
Control of Nitrification by Tree Species in a Common-Garden Experiment
Upload
independentCategory
view
3download
0
Rn
Ca
b
c
d
e
a
ARRA
KAEFNPR
1
acboggrftot
0d
Enzyme and Microbial Technology 46 (2010) 229–236
Contents lists available at ScienceDirect
Enzyme and Microbial Technology
journa l homepage: www.e lsev ier .com/ locate /emt
elationship between respirometric activity and community of entrappeditrifying bacteria: Implications for partial nitrification
haiwat Rongsayamanonta,b, Tawan Limpiyakornc, Benedict Lawd, Eakalak Khane,∗
International Postgraduate Programs in Environmental Management, Graduate School, Chulalongkorn University, Bangkok 10330, ThailandNational Center of Excellence for Environmental and Hazardous Waste Management, Chulalongkorn University, Bangkok 10330, ThailandDepartment of Environmental Engineering, Faculty of Engineering, Chulalongkorn University, Bangkok 10330, ThailandDepartment of Pharmaceutical Sciences, College of Pharmacy, Nursing, and Allied Sciences, North Dakota State University, Fargo, ND 58108, USADepartment of Civil Engineering, College of Engineering and Architecture, North Dakota State University, Fargo, ND 58108, USA
r t i c l e i n f o
rticle history:eceived 24 July 2009eceived in revised form 27 October 2009ccepted 28 October 2009
eywords:mmonia oxidizing bacteria (AOB)ntrapped cellsluorescence in situ hybridization (FISH)itrite oxidizing bacteria (NOB)artial nitrificationespirometry
a b s t r a c t
Activated sludge obtained from two municipal wastewater treatment facilities (WWTF) was used as seedsludge for enriched nitrifiers, which were later entrapped in polyvinyl alcohol. Seed sludge from oneWWTF was acclimated to high ammonia level (1813 mg NH3-N l−1) through the return of sludge digestersupernatant back to primary clarifier while seed sludge from the other WWTF was un-acclimated. To elu-cidate on how to control partial nitrification by entrapped cells, which could be different from suspendedcells, kinetics of entrapped enriched nitrifiers were studied using a respirometric assay. The community ofnitrifiers within the entrapment matrix, which was observed by fluorescence in situ hybridization (FISH)technique, was related to the nitritation and nitratation kinetics based on oxygen uptake rate. Maximumoxygen uptake rate, and substrate and oxygen affinities of both ammonia oxidizing bacteria (AOB) fornitritation and nitrite oxidizing bacteria (NOB) for nitratation in entrapped cells were lower than those ofcorresponding suspended cells. Under dissolved oxygen (DO) limiting conditions, nitratation was moresuppressed than nitritation for suspended cells, while for the entrapped cells, the results were the con-
trary. A free ammonia (FA) inhibition affected only the un-acclimated sludge. Either FA inhibition or DOlimitation might not be a sole effective control parameter to achieve partial nitrification by entrappedcells. FISH results revealed that Nitrosomonas europaea was the dominant AOB while Nitrobacter specieswas the dominant NOB in all cases. Heterotrophs were also present in the entrapment at 22.8 ± 18.6%and 41.5 ± 4.3% of total bacteria for acclimated and un-acclimated originated sludge. The availability ofsubstrate and oxygen governed the distributions of AOB, NOB and heterotrophs within the entrapmentf entr
and nitritation kinetics o. Introduction
Partial nitrification of ammonium rich wastewater is nowcommon practice. To generate and maintain partial nitrifi-
ation, the ammonia oxidizing bacteria (AOB) activity has toe maintained while simultaneously the activity of the nitritexidizing bacteria (NOB) needs to be inhibited. For suspendedrowth systems, at high ammonium or limited dissolved oxy-en (DO) concentrations, the activities of NOB or the nitratationate are suppressed due to ammonia inhibition or lower affinity
or oxygen than AOB and thus, nitrite accumulates in the sys-ems. The effectiveness of these two common strategies dependsn what specific species of nitrifying bacteria are in the reac-or and how they interact with environmental conditions such as∗ Corresponding author. Tel.: +1 701 231 7717; fax: +1 701 231 6185.E-mail address: [email protected] (E. Khan).
141-0229/$ – see front matter © 2009 Elsevier Inc. All rights reserved.oi:10.1016/j.enzmictec.2009.10.014
apped nitrifiers.© 2009 Elsevier Inc. All rights reserved.
their affinity to substrate and ability to tolerate inhibiting condi-tions.
For attached growth systems, which have less substrate trans-fer and space for growth compared to suspended growth systems,nitrite accumulation depends on other factors such as cell localiza-tion [1]. Recently, there have been studies suggesting that microbiallocalization within biofilm has significant effects on its substratediffusion resistance or substrate utilization [2–4]. For example,Okabe et al. [5] reported that heterotrophs outcompeted nitrifiersfor oxygen and space. As a result, hetetorophs dominated the outer-most biofilm while nitrifying bacteria were found only in the innerlayer causing reduced nitrification.
Cell entrapment within polymeric gels such as polyvinyl alcohol
(PVA) and carrageenan has been extensively applied for nitrifi-cation of wastewater [6–8]. Nitrification in full scale wastewatertreatment plants by using polyethylene glycol (PEG) entrappedcells has been practiced in Japan and the United States [9].Moreover, advantages of entrapped nitrifying bacteria over corre-
2 d Mic
swob
or[s([twcsdactd
dicawfss
anrmiopnns
2
2
neNFarI
2
FsMda(oswpaaw
30 C. Rongsayamanont et al. / Enzyme an
ponding suspended cells including effective protection against cellash-out, higher conversion rates, higher cell concentrations, ease
f cell separation, and protection from unfavorable conditions haveeen reported [10,11].
Although there have been a number of studies and applicationsf entrapped cells for nitrification, there have been limited peer-eviewed articles that address partial nitrification by entrapped cell10,12–14]. Isaka et al. [14] reported nitrite accumulation in theirtudy which used PEG-entrapped cells to treat landfill leachate150–200 mg l−1 of NH3-N) at low temperatures. Hill and Khan13] found that by using calcium alginate-entrapped cells, morehan 70% of ammonia in anaerobic sludge digester supernatantas converted to nitrite even under an unlimited dissolved oxygen
ondition in bulk liquid. Although these studies showed the pos-ibility for applying entrapped cells for partial nitrification, theyid not provide any understandings on the kinetics of nitritationnd nitratation by entrapped cells and in situ nitrifying bacterialommunity within the entrapment matrix to elucidate on howo control partial nitrification by entrapped cells, which could beifferent from suspended cells.
Nitritation and nitratation can become the rate limiting step atifferent environmental conditions. An understanding of the kinet-
cs of these two steps is needed for setting up design criteria andontrol strategy for partial nitrification. Although there have beennumber of kinetic studies of nitritation and nitratation processesith suspended cells [15,16], such work has never been conducted
or entrapped cells. The knowledge on the kinetics of these twoteps for entrapped cells, particularly associated with the controltrategy, could be used to achieve effective partial nitrification.
The main objectives of this study were to examine nitritationnd nitratation kinetics based on oxygen uptake rate of enricheditrifying sludge entrapped in PVA and to observe how they areelated to bacterial community and population. A batch respiro-etric assay coupled with nitritation and nitratation chemical
nhibitors was used to determine the bacterial activities. A flu-rescence in situ hybridization (FISH) technique was chosen forrobing nitrifying bacteria within the entrapment matrix since theitritation and nitratation kinetics are related to the make up ofitrifying bacterial community. This study was also conducted withuspended cells in parallel.
. Materials and methods
.1. Enrichment medium and synthetic wastewater
An enrichment medium, which was synthetic wastewater with a total ammoniaitrogen (TAN) level of 150 mg N l−1 (0.72 g of (NH4)2SO4 per liter), was used fornriching nitrifying bacteria. The synthetic wastewater contained NaHCO3 (1.5 g),a2HPO4 (4.05 g), K2HPO4 (2.1 g), MgSO4·7H2O (0.05 g), CaCl2·2H2O (0.01 g), andeSO4·7H2O (0.09 g) in one liter of distilled water. The synthetic wastewater plusmmonia ((NH4)2SO4) and/or nitrite (NaNO2) at various concentrations was used forespirometric experiments. All chemicals were purchased from VWR international,nc. (West Chester, PA, USA).
.2. Enrichment and entrapment of nitrifying sludge
Activated sludge obtained from the Moorhead (MH) Wastewater Treatmentacility (WWTF) in Moorhead, Minnesota, USA (High purity oxygen activated sludgeystem, solid retention times of 3 days) and Fergus Falls (FF) WWTF in Fergus Falls,innesota, USA (Complete-mix activated sludge system, solid retention times of 5
ays) were enriched separately for nitrifying bacteria. The Moorhead WWTF hadreturn flow of sludge digester supernatant, which was very high in ammonia
average NH3-N concentration of 1813 mg N l−1 and average NH3-N pulse loadingf 222 kg N d−1) while the Fergus Falls WWTF did not practice the return of digester
upernatant. Two 17 l laboratory-scale enrichment reactors, one for each sludge,ere aerated and operated in a two day cycle fill and draw mode at room tem-erature (22–23 ◦C). The reactors were fed with the enrichment medium with a pHdjustment between 7.5 and 8.2. After 10 weeks, more than 95% of TAN was removednd no nitrite accumulation was observed in both reactors, then the enriched sludgeas harvested.robial Technology 46 (2010) 229–236
The harvested biomass was entrapped in phosphorylated PVA gel beads at acell-to-matrix ratio of 4% (w/v) (g of centrifuged mixed liquor volatile suspendedsolids/ml of media) according to the method by Chen and Lin [11]. In brief, a PVAaqueous solution (20%, w/v) was mixed thoroughly with an equal volume of con-centrated enriched sludge with volatile suspended solid (VSS) concentration of70 g l−1. The mixture was dropped into a saturated boric acid solution at a rate of0.83 ml min−1 resulting in a droplet diameter of 3–4 mm. About one hour after thedropping ended, the formed beads were then transferred to a 1 M sodium orthophos-phate solution (pH 7) for 2 h for hardening. After that, each batch of the entrappedcells was returned to the enrichment reactor for maintenance.
2.3. Respirometric assay
A 300 ml completely sealed glass vessel with a port at the top for the insertion ofa DO probe was employed for respirometry. The DO probe was connected througha RS-232 port to a personnel computer, which was used for storing and monitoringall data transmitted by the probe. For each test, the suspended cells or entrappedcell beads previously washed with the synthetic wastewater without nitrogen sup-plement were added to the vessel, which was later filled up with the syntheticwastewater previously aerated for 1–2 min. The pH of the synthetic wastewater wasadjusted between 7.9 and 8.2 and was maintained by the phosphate buffer (part ofthe synthetic wastewater). In the vessel, the concentration of suspended cells was0.47 ± 0.03 g VSS l−1 while around 6.0–9.0 g wet entrapped cell beads (30 beads)were used. The vessel was magnetically stirred and operated at room temperature(22–23 ◦C). DO depletion in the vessel due to substrate utilization was monitoredover time and the specific oxygen uptake rate (SOUR) was determined. The SOURwas determined twice for every substrate concentration tested.
The kinetic of nitrification process is dependent mainly on substrate and DOconcentrations. It is normally described by a Double Monod expression (Eq. (1)). Foreach experiment, the data were fitted to Eq. (1) using a nonlinear regression modulein SIGMAPLOT® 10.0 to determine maximum SOUR and half saturation coefficientsfor nitritation and nitratation.
SOUR = SOURMAX
([S]
KS + [S]
)([DO]
KO + [DO]
)(1)
where SOUR is specific oxygen uptake rate for nitritation or nitratation (mgO2 [mg VSS h]−1), SOURMAX is maximum SOUR for nitritation or nitratation(SOURMAX,AOB, SOURMAX,NOB) (mg O2 [mg VSS h]−1), [S] is substrate concentration(TAN or NO2
−) (mg N l−1), [DO] is DO concentration (mg O2 l−1), KS is half satura-tion coefficient for substrate (KS,AOB, KS,NOB) (mg N l−1), and KO is half saturationcoefficient for oxygen (KO,AOB, KO,NOB) (mg O2 l−1).
2.3.1. Effect of substrate (ammonia and nitrite) concentrationsTo investigate the effect of substrate concentrations, two tests, nitritation and
nitratation, were conducted. For the nitritation test, the synthetic wastewater whichhad initial ammonia concentrations of 0, 3, 5, 10, 20, 40, 80, 150, 300 mg N l−1 wasused to study the influence of ammonia on nitritation rate by both suspended andentrapped cells. To inhibit nitrite oxidation in the nitritation test, sodium azide wasadded at a concentration of 50 �M. For the nitratation test, the initial nitrite concen-trations of synthetic wastewater were 0, 5, 10, 20, 40, 80, 120, 150 mg N l−1 to studythe influence of nitrite on nitratation rate by both types of cells. Allylthiourea (ATU)was added simultaneously at a concentration of 86 �M to inhibit ammonia oxida-tion [17]. To avoid the occurrence of DO limitation in the vessel, the experimentwas terminated at 4.0 mg O2 l−1 for suspended cells and 6.0 mg O2 l−1 for entrappedcells. The initial DO was at 7.5 mg O2 l−1.
2.3.2. Effect of bulk DO concentrationExperiments to examine the effect of bulk DO concentration were conducted in
the same manner as described in Section 2.3.1. An initial ammonia concentration of150 mg N l−1 and an initial nitrite concentration of 120 mg N l−1 were chosen for thenitritation and nitratation tests, respectively.
2.3.3. Inhibitory effect of free ammoniaA free ammonia (FA) inhibition on ammonia oxidation was tested using syn-
thetic wastewater at the initial FA concentrations of 0, 0.6, 1.2, 2.4, 4.9, 9.8, 18.3,36.6, 61.1, 91.7, 122.2 mg N l−1 with 50 �M sodium azide. For the nitratation test,the initial FA concentrations of synthetic wastewater were 0, 0.6, 1.2, 2.4, 4.9, 9.8,18.3, 36.6, 61.1 mg N l−1 while the nitrite concentration was 120 mg N l−1. Ammoniaoxidation was inhibited by adding ATU in the same manner as described in Section2.3.1.
2.4. Chemical analyses
DO (Thermo Orion 850A meter, Thermo Orion DO 083005D probe), pH (ThermoOrion 250A + pH Electrode), and VSS were determined according to Standard Meth-ods [18]. Ammonia and nitrate nitrogen were analyzed using ion selective electrodes(Thermo Orion 250A+, VWR SympHony Ammonia Combination Electrode, VWRSympHony Nitrate Ion Selective Electrode, VWR SympHony Double Junction Ref-erence Half-Cell) in accordance with Standard Methods [18]. Nitrite was analyzed
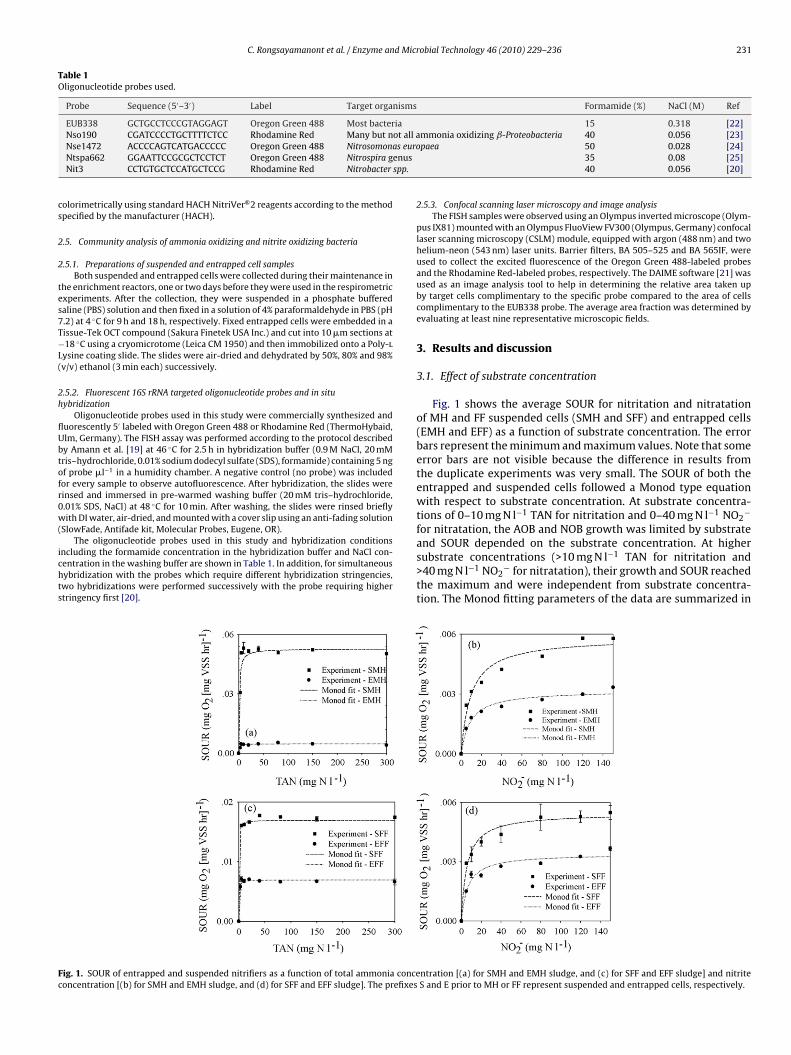
C. Rongsayamanont et al. / Enzyme and Microbial Technology 46 (2010) 229–236 231
Table 1Oligonucleotide probes used.
Probe Sequence (5′–3′) Label Target organisms Formamide (%) NaCl (M) Ref
EUB338 GCTGCCTCCCGTAGGAGT Oregon Green 488 Most bacteria 15 0.318 [22]ot all aas euroenusspp.
cs
2
2
tes7T−L(
2h
flUbtofr0w(
ichts
Fc
Nso190 CGATCCCCTGCTTTTCTCC Rhodamine Red Many but nNse1472 ACCCCAGTCATGACCCCC Oregon Green 488 NitrosomonNtspa662 GGAATTCCGCGCTCCTCT Oregon Green 488 Nitrospira gNit3 CCTGTGCTCCATGCTCCG Rhodamine Red Nitrobacter
olorimetrically using standard HACH NitriVer®2 reagents according to the methodpecified by the manufacturer (HACH).
.5. Community analysis of ammonia oxidizing and nitrite oxidizing bacteria
.5.1. Preparations of suspended and entrapped cell samplesBoth suspended and entrapped cells were collected during their maintenance in
he enrichment reactors, one or two days before they were used in the respirometricxperiments. After the collection, they were suspended in a phosphate bufferedaline (PBS) solution and then fixed in a solution of 4% paraformaldehyde in PBS (pH.2) at 4 ◦C for 9 h and 18 h, respectively. Fixed entrapped cells were embedded in aissue-Tek OCT compound (Sakura Finetek USA Inc.) and cut into 10 �m sections at18 ◦C using a cryomicrotome (Leica CM 1950) and then immobilized onto a Poly-lysine coating slide. The slides were air-dried and dehydrated by 50%, 80% and 98%v/v) ethanol (3 min each) successively.
.5.2. Fluorescent 16S rRNA targeted oligonucleotide probes and in situybridization
Oligonucleotide probes used in this study were commercially synthesized anduorescently 5′ labeled with Oregon Green 488 or Rhodamine Red (ThermoHybaid,lm, Germany). The FISH assay was performed according to the protocol describedy Amann et al. [19] at 46 ◦C for 2.5 h in hybridization buffer (0.9 M NaCl, 20 mMris–hydrochloride, 0.01% sodium dodecyl sulfate (SDS), formamide) containing 5 ngf probe �l−1 in a humidity chamber. A negative control (no probe) was includedor every sample to observe autofluorescence. After hybridization, the slides wereinsed and immersed in pre-warmed washing buffer (20 mM tris–hydrochloride,.01% SDS, NaCl) at 48 ◦C for 10 min. After washing, the slides were rinsed brieflyith DI water, air-dried, and mounted with a cover slip using an anti-fading solution
SlowFade, Antifade kit, Molecular Probes, Eugene, OR).
The oligonucleotide probes used in this study and hybridization conditionsncluding the formamide concentration in the hybridization buffer and NaCl con-entration in the washing buffer are shown in Table 1. In addition, for simultaneousybridization with the probes which require different hybridization stringencies,wo hybridizations were performed successively with the probe requiring highertringency first [20].
ig. 1. SOUR of entrapped and suspended nitrifiers as a function of total ammonia conceoncentration [(b) for SMH and EMH sludge, and (d) for SFF and EFF sludge]. The prefixes
mmonia oxidizing ˇ-Proteobacteria 40 0.056 [23]paea 50 0.028 [24]
35 0.08 [25]40 0.056 [20]
2.5.3. Confocal scanning laser microscopy and image analysisThe FISH samples were observed using an Olympus inverted microscope (Olym-
pus IX81) mounted with an Olympus FluoView FV300 (Olympus, Germany) confocallaser scanning microscopy (CSLM) module, equipped with argon (488 nm) and twohelium-neon (543 nm) laser units. Barrier filters, BA 505–525 and BA 565IF, wereused to collect the excited fluorescence of the Oregon Green 488-labeled probesand the Rhodamine Red-labeled probes, respectively. The DAIME software [21] wasused as an image analysis tool to help in determining the relative area taken upby target cells complimentary to the specific probe compared to the area of cellscomplimentary to the EUB338 probe. The average area fraction was determined byevaluating at least nine representative microscopic fields.
3. Results and discussion
3.1. Effect of substrate concentration
Fig. 1 shows the average SOUR for nitritation and nitratationof MH and FF suspended cells (SMH and SFF) and entrapped cells(EMH and EFF) as a function of substrate concentration. The errorbars represent the minimum and maximum values. Note that someerror bars are not visible because the difference in results fromthe duplicate experiments was very small. The SOUR of both theentrapped and suspended cells followed a Monod type equationwith respect to substrate concentration. At substrate concentra-tions of 0–10 mg N l−1 TAN for nitritation and 0–40 mg N l−1 NO2
−
for nitratation, the AOB and NOB growth was limited by substrate
and SOUR depended on the substrate concentration. At highersubstrate concentrations (>10 mg N l−1 TAN for nitritation and>40 mg N l−1 NO2− for nitratation), their growth and SOUR reachedthe maximum and were independent from substrate concentra-tion. The Monod fitting parameters of the data are summarized in
ntration [(a) for SMH and EMH sludge, and (c) for SFF and EFF sludge] and nitriteS and E prior to MH or FF represent suspended and entrapped cells, respectively.
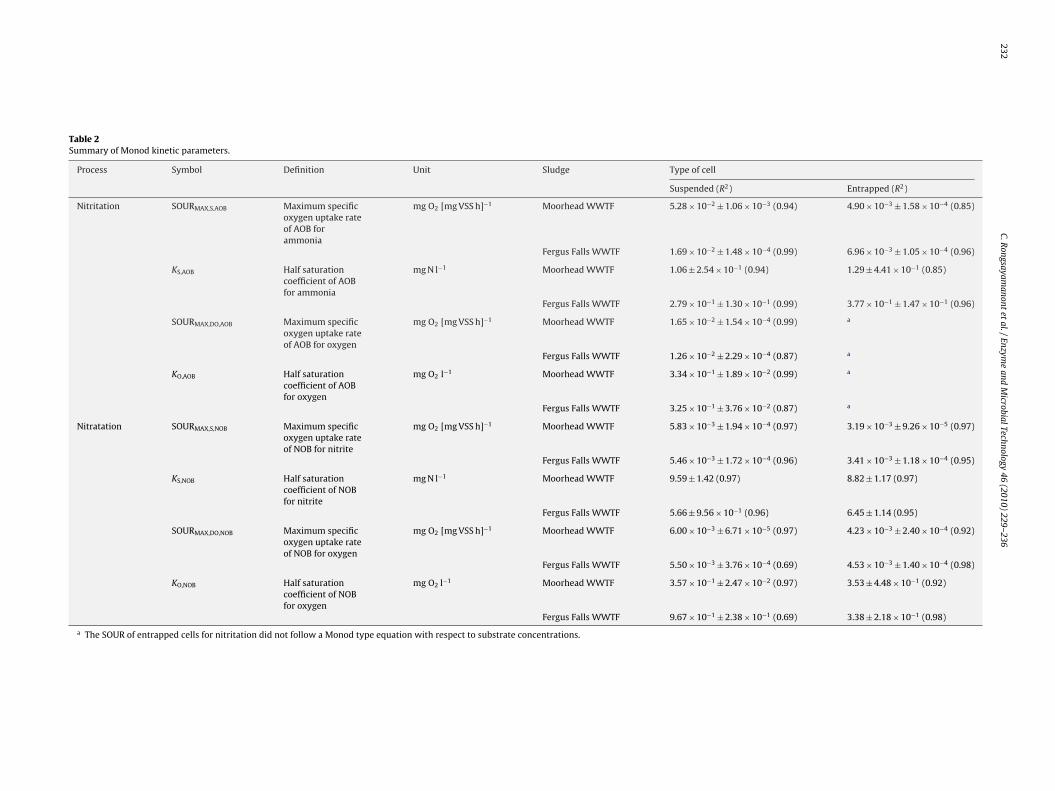
232C.R
ongsayamanont
etal./Enzym
eand
MicrobialTechnology
46 (2010) 229–236
Table 2Summary of Monod kinetic parameters.
Process Symbol Definition Unit Sludge Type of cell
Suspended (R2) Entrapped (R2)
Nitritation SOURMAX,S,AOB Maximum specificoxygen uptake rateof AOB forammonia
mg O2 [mg VSS h]−1 Moorhead WWTF 5.28 × 10−2 ± 1.06 × 10−3 (0.94) 4.90 × 10−3 ± 1.58 × 10−4 (0.85)
Fergus Falls WWTF 1.69 × 10−2 ± 1.48 × 10−4 (0.99) 6.96 × 10−3 ± 1.05 × 10−4 (0.96)
KS,AOB Half saturationcoefficient of AOBfor ammonia
mg N l−1 Moorhead WWTF 1.06 ± 2.54 × 10−1 (0.94) 1.29 ± 4.41 × 10−1 (0.85)
Fergus Falls WWTF 2.79 × 10−1 ± 1.30 × 10−1 (0.99) 3.77 × 10−1 ± 1.47 × 10−1 (0.96)
SOURMAX,DO,AOB Maximum specificoxygen uptake rateof AOB for oxygen
mg O2 [mg VSS h]−1 Moorhead WWTF 1.65 × 10−2 ± 1.54 × 10−4 (0.99) a
Fergus Falls WWTF 1.26 × 10−2 ± 2.29 × 10−4 (0.87) a
KO,AOB Half saturationcoefficient of AOBfor oxygen
mg O2 l−1 Moorhead WWTF 3.34 × 10−1 ± 1.89 × 10−2 (0.99) a
Fergus Falls WWTF 3.25 × 10−1 ± 3.76 × 10−2 (0.87) a
Nitratation SOURMAX,S,NOB Maximum specificoxygen uptake rateof NOB for nitrite
mg O2 [mg VSS h]−1 Moorhead WWTF 5.83 × 10−3 ± 1.94 × 10−4 (0.97) 3.19 × 10−3 ± 9.26 × 10−5 (0.97)
Fergus Falls WWTF 5.46 × 10−3 ± 1.72 × 10−4 (0.96) 3.41 × 10−3 ± 1.18 × 10−4 (0.95)
KS,NOB Half saturationcoefficient of NOBfor nitrite
mg N l−1 Moorhead WWTF 9.59 ± 1.42 (0.97) 8.82 ± 1.17 (0.97)
Fergus Falls WWTF 5.66 ± 9.56 × 10−1 (0.96) 6.45 ± 1.14 (0.95)
SOURMAX,DO,NOB Maximum specificoxygen uptake rateof NOB for oxygen
mg O2 [mg VSS h]−1 Moorhead WWTF 6.00 × 10−3 ± 6.71 × 10−5 (0.97) 4.23 × 10−3 ± 2.40 × 10−4 (0.92)
Fergus Falls WWTF 5.50 × 10−3 ± 3.76 × 10−4 (0.69) 4.53 × 10−3 ± 1.40 × 10−4 (0.98)
KO,NOB Half saturationcoefficient of NOBfor oxygen
mg O2 l−1 Moorhead WWTF 3.57 × 10−1 ± 2.47 × 10−2 (0.97) 3.53 ± 4.48 × 10−1 (0.92)
Fergus Falls WWTF 9.67 × 10−1 ± 2.38 × 10−1 (0.69) 3.38 ± 2.18 × 10−1 (0.98)
a The SOUR of entrapped cells for nitritation did not follow a Monod type equation with respect to substrate concentrations.

C. Rongsayamanont et al. / Enzyme and Microbial Technology 46 (2010) 229–236 233
F tion inn es S a
Ttcbetto
f1wWf1oStStafsn(d
t6oEahsbmE
ig. 2. SOUR of entrapped and suspended nitrifiers as a function of DO concentraitratation [(b) for SMH and EMH sludge, and (d) for SFF and EFF sludge]. The prefix
able 2. The SOURMAX of suspended cells were clearly higher thanhose of corresponding entrapped cells while the half saturationoefficients for substrate of entrapped cells were higher proba-ly because of the internal substrate transfer limitation within thentrapment gel [26]. There was only one case that did not follow therend; the half saturation coefficient for nitrite of SMH was higherhan that of EMH which could be due to uncontrollable variabilityf the respirometric assay.
For suspended cells, the values of half saturation coefficientsor total ammonia nitrogen (Ks, AOB) between 2.79 × 10−1 and.06 mg N l−1 and nitrite (Ks, NOB) between 5.66 and 9.59 mg N l−1
ere agreeable with those reported in previous studies [26–29].hen comparing between nitritation and nitratation, the SOURMAX
or nitritation (5.28 × 10−2 mg O2 mg VSS−1 h−1 for SMH and.69 × 10−2 mg O2 mg VSS−1 h−1 for SFF) were higher than thosef corresponding nitratation (5.83 × 10−3 mg O2 mg VSS−1 h−1 forMH and 5.46 × 10−3 mg O2 mg VSS−1 h−1 for SFF). This is becausehe amount of AOB was higher than that of NOB as discussed later inection 3.4. The magnitudes of SOURMAX for nitritation and nitrata-ion of SMH were higher than those of SFF. This observation isgreeable with the FISH results (Section 3.4) which show higherractions of AOB and NOB in SMH than those in SFF. The seedludge from the Moorhead WWTF was acclimated to high ammo-ia and should be abundant in high growth rate nitrifier speciesr-strategist). As a result, after the enrichment under the same con-ition, there were more AOB and NOB in SMH than those in SFF.
For entrapped cells, the SOURMAX for nitrita-ion (4.90 × 10−3 mg O2 mg VSS−1 h−1 for EMH and.96 × 10−3 mg O2 mg VSS−1 h−1 for EFF) were higher than thosef corresponding nitratation (3.19 × 10−3 mg O2 mg VSS−1 h−1 forMH and 3.41 × 10−3 mg O2 mg VSS−1 h−1 for EFF) agreeing withn observation that the amounts of AOB in both EMH and EFF were
igher than that of NOB (see Section 3.4). Although the source ofludge affected the activity of suspended cells (SMH and SFF), thisehavior was less evident for entrapped cells (EMH and EFF). Theagnitudes of SOURMAX of EMH and EFF were closer. However,MH had lower SOURMAX than EFF while higher fractions of AOB
nitritation [(a) for SMH and EMH sludge, and (c) for SFF and EFF sludge] and innd E prior to MH or FF represent suspended and entrapped cells, respectively.
and NOB in EMH than those in EFF were observed (see Section3.4). This suggests that there was no relationship between thechange in the activity of entrapped nitrifiers and the change inthe amount of nitrifiers in the entrapment. It is possible that cellprotection against adverse effects provided by entrapment matrix[7] controlled the nitrifier activity through substrate and/or oxygendiffusion.
3.2. Effect of bulk DO concentration
Effects of DO concentration on SOUR for nitritation and nitrata-tion are presented in Fig. 2 and summarized in Table 2. Note thatthere are no error bars in Fig. 2 because the measurement of DO con-centration could not be controlled to be at the same values betweenthe duplicate experiments. As shown in Fig. 2, the nitrifier activitywas affected differently by DO between suspended and entrappedcells.
For suspended cells, at <2 mg l−1 DO, the SOUR for nitri-tation and nitratation were limited by the DO concentration.When DO > 2 mg l−1, the SOUR were constant and became max-imum. The SOURMAX values of SMH were higher than those ofSFF. The explanations for these observations were the same asthose given in Section 3.1. The half saturation coefficient for oxy-gen in nitritation (KO,AOB = 3.34 × 10−1 and 3.25 × 10−1 mg O2 l−1)and nitratation (KO,NOB = 3.57 × 10−1 and 9.67 × 10−1 mg O2 l−1) forSMH and SFF sludge were comparable to the published val-ues (KO,AOB = 0.22–0.56 mg O2 l−1 and KO,NOB = 0.17–4.32 mg O2 l−1)[29]. The half saturation coefficients for oxygen in nitritation werelower than those of nitratation suggesting that under oxygen-limited environment (as in partial nitrifying condition), AOB in SMHand SFF sludge would outcompete NOB for oxygen and NOB wouldeventually be washed out of the reactor.
For entrapped cells, the SOUR for nitratation followed theMonod kinetics with respect to DO concentration. NOB in both EMHand EFF needed DO around 4 mg O2 l−1 to achieve their maximumutilization while the SOUR of entrapped AOB for both EMH and EFFwere related to DO in the first order manner and they needed more
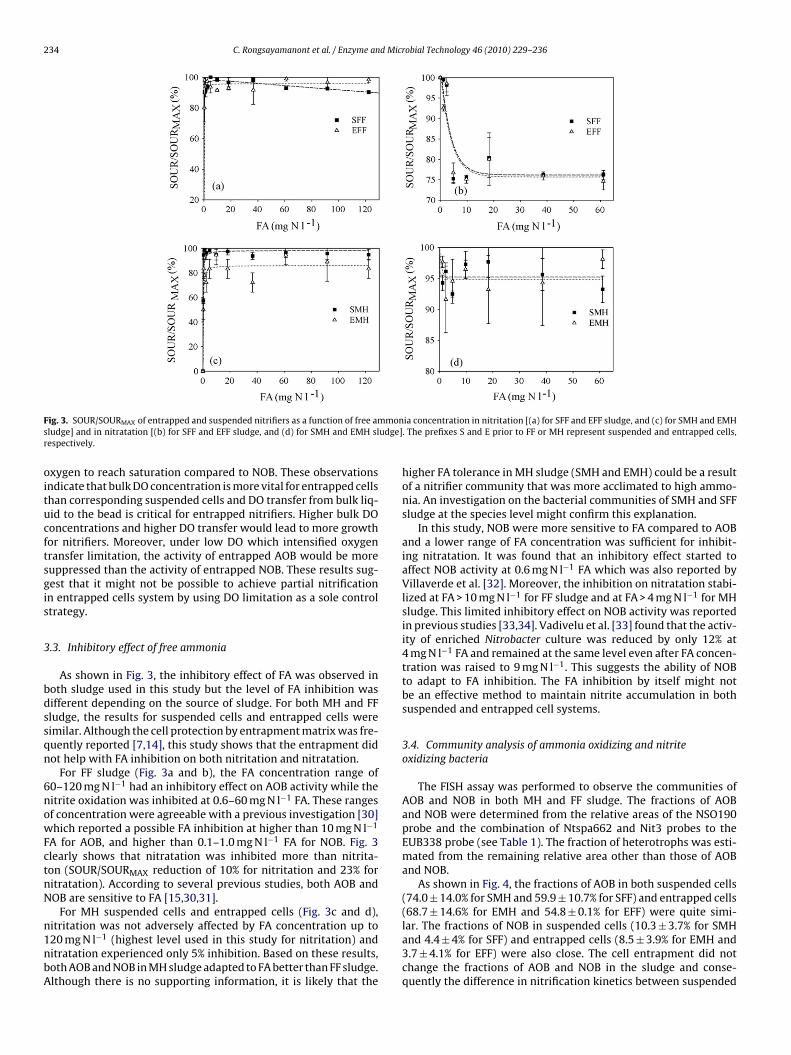
234 C. Rongsayamanont et al. / Enzyme and Microbial Technology 46 (2010) 229–236
F mons udge]r
oitucftsgis
3
bdssqn
6nowFctnN
n1nbA
ig. 3. SOUR/SOURMAX of entrapped and suspended nitrifiers as a function of free amludge] and in nitratation [(b) for SFF and EFF sludge, and (d) for SMH and EMH slespectively.
xygen to reach saturation compared to NOB. These observationsndicate that bulk DO concentration is more vital for entrapped cellshan corresponding suspended cells and DO transfer from bulk liq-id to the bead is critical for entrapped nitrifiers. Higher bulk DOoncentrations and higher DO transfer would lead to more growthor nitrifiers. Moreover, under low DO which intensified oxygenransfer limitation, the activity of entrapped AOB would be moreuppressed than the activity of entrapped NOB. These results sug-est that it might not be possible to achieve partial nitrificationn entrapped cells system by using DO limitation as a sole controltrategy.
.3. Inhibitory effect of free ammonia
As shown in Fig. 3, the inhibitory effect of FA was observed inoth sludge used in this study but the level of FA inhibition wasifferent depending on the source of sludge. For both MH and FFludge, the results for suspended cells and entrapped cells wereimilar. Although the cell protection by entrapment matrix was fre-uently reported [7,14], this study shows that the entrapment didot help with FA inhibition on both nitritation and nitratation.
For FF sludge (Fig. 3a and b), the FA concentration range of0–120 mg N l−1 had an inhibitory effect on AOB activity while theitrite oxidation was inhibited at 0.6–60 mg N l−1 FA. These rangesf concentration were agreeable with a previous investigation [30]hich reported a possible FA inhibition at higher than 10 mg N l−1
A for AOB, and higher than 0.1–1.0 mg N l−1 FA for NOB. Fig. 3learly shows that nitratation was inhibited more than nitrita-on (SOUR/SOURMAX reduction of 10% for nitritation and 23% foritratation). According to several previous studies, both AOB andOB are sensitive to FA [15,30,31].
For MH suspended cells and entrapped cells (Fig. 3c and d),
itritation was not adversely affected by FA concentration up to20 mg N l−1 (highest level used in this study for nitritation) anditratation experienced only 5% inhibition. Based on these results,oth AOB and NOB in MH sludge adapted to FA better than FF sludge.lthough there is no supporting information, it is likely that theia concentration in nitritation [(a) for SFF and EFF sludge, and (c) for SMH and EMH. The prefixes S and E prior to FF or MH represent suspended and entrapped cells,
higher FA tolerance in MH sludge (SMH and EMH) could be a resultof a nitrifier community that was more acclimated to high ammo-nia. An investigation on the bacterial communities of SMH and SFFsludge at the species level might confirm this explanation.
In this study, NOB were more sensitive to FA compared to AOBand a lower range of FA concentration was sufficient for inhibit-ing nitratation. It was found that an inhibitory effect started toaffect NOB activity at 0.6 mg N l−1 FA which was also reported byVillaverde et al. [32]. Moreover, the inhibition on nitratation stabi-lized at FA > 10 mg N l−1 for FF sludge and at FA > 4 mg N l−1 for MHsludge. This limited inhibitory effect on NOB activity was reportedin previous studies [33,34]. Vadivelu et al. [33] found that the activ-ity of enriched Nitrobacter culture was reduced by only 12% at4 mg N l−1 FA and remained at the same level even after FA concen-tration was raised to 9 mg N l−1. This suggests the ability of NOBto adapt to FA inhibition. The FA inhibition by itself might notbe an effective method to maintain nitrite accumulation in bothsuspended and entrapped cell systems.
3.4. Community analysis of ammonia oxidizing and nitriteoxidizing bacteria
The FISH assay was performed to observe the communities ofAOB and NOB in both MH and FF sludge. The fractions of AOBand NOB were determined from the relative areas of the NSO190probe and the combination of Ntspa662 and Nit3 probes to theEUB338 probe (see Table 1). The fraction of heterotrophs was esti-mated from the remaining relative area other than those of AOBand NOB.
As shown in Fig. 4, the fractions of AOB in both suspended cells(74.0 ± 14.0% for SMH and 59.9 ± 10.7% for SFF) and entrapped cells(68.7 ± 14.6% for EMH and 54.8 ± 0.1% for EFF) were quite simi-
lar. The fractions of NOB in suspended cells (10.3 ± 3.7% for SMHand 4.4 ± 4% for SFF) and entrapped cells (8.5 ± 3.9% for EMH and3.7 ± 4.1% for EFF) were also close. The cell entrapment did notchange the fractions of AOB and NOB in the sludge and conse-quently the difference in nitrification kinetics between suspended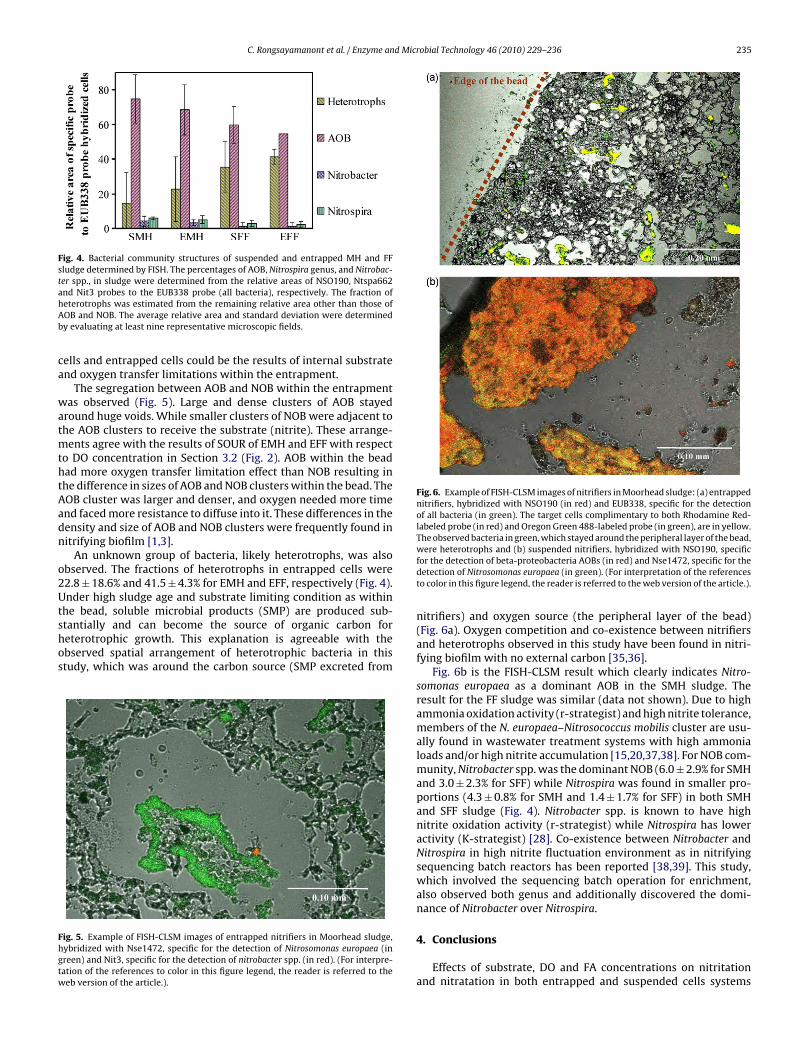
C. Rongsayamanont et al. / Enzyme and Microbial Technology 46 (2010) 229–236 235
Fig. 4. Bacterial community structures of suspended and entrapped MH and FFsludge determined by FISH. The percentages of AOB, Nitrospira genus, and Nitrobac-ter spp., in sludge were determined from the relative areas of NSO190, Ntspa662ahAb
ca
watmthtAadn
o2Utshos
Fhgtw
Fig. 6. Example of FISH-CLSM images of nitrifiers in Moorhead sludge: (a) entrappednitrifiers, hybridized with NSO190 (in red) and EUB338, specific for the detectionof all bacteria (in green). The target cells complimentary to both Rhodamine Red-labeled probe (in red) and Oregon Green 488-labeled probe (in green), are in yellow.The observed bacteria in green, which stayed around the peripheral layer of the bead,
nd Nit3 probes to the EUB338 probe (all bacteria), respectively. The fraction ofeterotrophs was estimated from the remaining relative area other than those ofOB and NOB. The average relative area and standard deviation were determinedy evaluating at least nine representative microscopic fields.
ells and entrapped cells could be the results of internal substratend oxygen transfer limitations within the entrapment.
The segregation between AOB and NOB within the entrapmentas observed (Fig. 5). Large and dense clusters of AOB stayed
round huge voids. While smaller clusters of NOB were adjacent tohe AOB clusters to receive the substrate (nitrite). These arrange-
ents agree with the results of SOUR of EMH and EFF with respecto DO concentration in Section 3.2 (Fig. 2). AOB within the beadad more oxygen transfer limitation effect than NOB resulting inhe difference in sizes of AOB and NOB clusters within the bead. TheOB cluster was larger and denser, and oxygen needed more timend faced more resistance to diffuse into it. These differences in theensity and size of AOB and NOB clusters were frequently found initrifying biofilm [1,3].
An unknown group of bacteria, likely heterotrophs, was alsobserved. The fractions of heterotrophs in entrapped cells were2.8 ± 18.6% and 41.5 ± 4.3% for EMH and EFF, respectively (Fig. 4).nder high sludge age and substrate limiting condition as within
he bead, soluble microbial products (SMP) are produced sub-
tantially and can become the source of organic carbon foreterotrophic growth. This explanation is agreeable with thebserved spatial arrangement of heterotrophic bacteria in thistudy, which was around the carbon source (SMP excreted fromig. 5. Example of FISH-CLSM images of entrapped nitrifiers in Moorhead sludge,ybridized with Nse1472, specific for the detection of Nitrosomonas europaea (inreen) and Nit3, specific for the detection of nitrobacter spp. (in red). (For interpre-ation of the references to color in this figure legend, the reader is referred to theeb version of the article.).
were heterotrophs and (b) suspended nitrifiers, hybridized with NSO190, specificfor the detection of beta-proteobacteria AOBs (in red) and Nse1472, specific for thedetection of Nitrosomonas europaea (in green). (For interpretation of the referencesto color in this figure legend, the reader is referred to the web version of the article.).
nitrifiers) and oxygen source (the peripheral layer of the bead)(Fig. 6a). Oxygen competition and co-existence between nitrifiersand heterotrophs observed in this study have been found in nitri-fying biofilm with no external carbon [35,36].
Fig. 6b is the FISH-CLSM result which clearly indicates Nitro-somonas europaea as a dominant AOB in the SMH sludge. Theresult for the FF sludge was similar (data not shown). Due to highammonia oxidation activity (r-strategist) and high nitrite tolerance,members of the N. europaea–Nitrosococcus mobilis cluster are usu-ally found in wastewater treatment systems with high ammonialoads and/or high nitrite accumulation [15,20,37,38]. For NOB com-munity, Nitrobacter spp. was the dominant NOB (6.0 ± 2.9% for SMHand 3.0 ± 2.3% for SFF) while Nitrospira was found in smaller pro-portions (4.3 ± 0.8% for SMH and 1.4 ± 1.7% for SFF) in both SMHand SFF sludge (Fig. 4). Nitrobacter spp. is known to have highnitrite oxidation activity (r-strategist) while Nitrospira has loweractivity (K-strategist) [28]. Co-existence between Nitrobacter andNitrospira in high nitrite fluctuation environment as in nitrifyingsequencing batch reactors has been reported [38,39]. This study,which involved the sequencing batch operation for enrichment,also observed both genus and additionally discovered the domi-nance of Nitrobacter over Nitrospira.
4. Conclusions
Effects of substrate, DO and FA concentrations on nitritationand nitratation in both entrapped and suspended cells systems

2 d Mic
westpcrocoatmniaisgtemstsfs
A
iUfifAs
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
36 C. Rongsayamanont et al. / Enzyme an
ere determined by a respirometric assay. Two nitrifying sludgenriched from high ammonia acclimated and un-acclimated seedludge, were used in this study. For both nitritation and nitratation,he SOURMAX and the affinity for substrate and oxygen of sus-ended cells were higher than those of corresponding entrappedells. Under DO limiting conditions, the SOUR for nitratation waseduced more than that for nitritation for suspended cells becausef higher oxygen affinity of AOB than NOB, while for entrappedells, the results were the opposite. The FA inhibitory effect wasbserved in the un-acclimated sludge at significant levels but thecclimated sludge was not inhibited by high FA. These results leado a conclusion that FA inhibition or DO limitation can be used to
aintain nitrite accumulation in entrapped cell systems but mightot be effective strategies, which need a detailed investigation to
dentify one. The fractions of AOB and NOB in both entrapped cellsnd suspended cells were comparable. N. europaea was the dom-nant AOB while Nitrobacter spp. was the dominant NOB for bothuspended and entrapped cells. The competition for space and oxy-en led to segregations among AOB, NOB and heterotrophs withinhe entrapment. Heterotrophic bacteria stayed within the periph-ral layer of the bead to take oxygen from bulk liquid and solubleicrobial products excreted from nitrifiers. Large clusters of AOB
tayed in huge voids and smaller clusters of NOB were adjacent tohe AOB clusters to receive nitrite. This study suggests that internalubstrate and oxygen transfers would be two of the most importantactors in controlling nitritation kinetics of entrapped nitrifyingludge.
cknowledgments
The authors would like to thank Departments of Civil Engineer-ng and Pharmaceutical Sciences, North Dakota State University,SA for providing research facilities. The authors acknowledgenancial support received from the National Center of Excellence
or Environmental and Hazardous Waste Management and the 90thnniversary of Chulalongkorn University Fund (Ratchadaphisek-omphot Endowment Fund), Chulalongkorn University, Thailand.
eferences
[1] Okabe S, Satoh H, Ito T, Watanabe Y. Analysis of microbial community struc-ture and in situ activity of nitrifying biofilms. J Water Environ Technol2004;2:65–74.
[2] Kim DJ, Chang JS, Lee DI, Han DW, Yoo IK, Cha GC. Nitrification of high strengthammonia wastewater and nitrite accumulation characteristics. Water Sci Tech-nol 2003;47:45–51.
[3] Schramm A, Larsen LH, Revsbech NP, Ramsing NB, Amann RI, Schleifer KH.Structure and function of a nitrifying biofilm as determined by in situ hybridiza-tion and the use of microelectrodes. Appl Environ Microbiol 1996;62:4641–7.
[4] Tanaka H, Dunn IJ. Kinetics of biofilm nitrification. Biotechnol Bioeng1992;24:669–89.
[5] Okabe S, Hiratia K, Ozawa Y, Watanabe Y. Spatial microbial distributions ofnitrifiers and heterotrophs in mixed-population biofilms. Biotechnol Bioeng1996;50:24–35.
[6] Cao G, Zhao Q, Sun X, Zhang T. Characterization of nitrifying and denitrify-ing bacteria coimmobilized in PVA and kinetics model of biological nitrogenremoval by coimmobilized cells. Enzyme Microb Technol 2002;30:49–55.
[7] Wijffels RH, Tramper J. Nitrification by immobilized cells. Enzyme Microb Tech-nol 1995;17:482–92.
[8] Hunik JH, van den Hoogen MP, de Boer W, Smit M, Tramper J. Quanti-tative determination of the spatial distribution of Nitrosomonas europaeaand Nitrobacter agilis cells immobilized in K-carrageenan gel beads by aspecific fluorescent antibody labelling technique. Appl Environ Microbiol1993;59:1951–4.
[9] Emori H, Mikawa K, Hamaya M, Yamaguchi T, Tanaka K, Takeshima T. PEGASUSInnovative biological nitrogen removal process using entrapped nitrifiers. In:Wijffels RH, Buitelaar RM, Bucke C, Tramper J, editors. Immobilized cells: basics
and applications. Amsterdam: Elsevier; 1996. p. 546–55.10] Rostron WM, Stuckey DC, Young AA. Nitrification of high strength ammo-nia wastewater: comparative study of immobilization media. Water Res2001;35:1169–78.
11] Chen KC, Lin YF. Immobilization of microorganisms with phosphorylatedpolyvinyl alcohol (PVA) gel. Enzyme Microb Technol 1994;16:79–83.
[
[
robial Technology 46 (2010) 229–236
12] Yan J, Hu YY. Partial nitrification to nitrite for treating ammonium-richorganic wastewater by immobilized biomass system. Bioresour Technol2009;100:2341–7.
13] Hill BC, Khan E. A comparative study of immobilized nitrifying and co-immobilized nitrifying and denitrifying bacteria for ammonia removal fromsludge digester supernatant. Water Air Soil Pollut 2008;195:23–33.
14] Isaka K, Yoshie S, Sumino T, Inamori Y, Tsuneda S. Nitrification of landfillleachate using immobilized nitrifying bacteria at low temperatures. BiochemEng J 2007;37:49–55.
15] Ahn JH, Yu R, Chandran K. Distinctive microbial ecology and biokinetics ofautotrophic ammonia and nitrite oxidation in a partial nitrification bioreactor.Biotechnol Bioeng 2008;100:1078–87.
16] Chandran K, Smets BF. Single-step nitrification models erroneously describebatch ammonia oxidation profiles when nitrite oxidation becomes rate limit-ing. Biotechnol Bioeng 2000;68:396–406.
17] Ginestet P, Audic JM, Urbain V, Block JC. Estimation of nitrifying bacterial activ-ities by measuring oxygen uptake in the presence of the metabolic inhibitorsallylthiourea and azide. Appl Environ Microbiol 1998;64:2266–8.
18] American Public Health Association. Standard methods for the examinationof water and wastewater. 20th ed. Washington, DC: American Public HealthAssociation; 1998.
19] Amann R, Ludwig W, Schleifer KH. Phylogenetic identification and in situdetection of individual microbial cells without cultivation. Microbiol Rev1995;59:143–69.
20] Wagner M, Rath G, Koops HP, Flood J, Amann RI. In situ analysis of nitrifyingbacteria in sewage treatment plants. Water Sci Technol 1996;34:237–44.
21] Daims H, Lücker S, Wagner M. Daime, a novel image analysis program formicrobial ecology and biofilm research. Environ Microbiol 2005;8:200–13.
22] Amann RI, Binder BJ, Olson RJ, Chisholm SW, Devereux R, Stahl DA. Acombination of 16S rRNA targeted oligonucleotide probes with flow cytom-etry for analyzing mixed microbial populations. Appl Environ Microbiol1990;56:1919–25.
23] Mobarry BK, Wagner M, Urbain V, Rittmann BE, Stahl DA. Phylo-genetic probes for analyzing abundance and spatial organizationof nitrifying bacteria. Appl Environ Microbiol 1996;62:2156–62.
24] Juretschko S, Timmermann G, Schmid M, Schleifer K-H, Pommerening-RöserA, Koops HP, et al. Combined molecular and conventional analysis of nitri-fying bacterium diversity in activated sludge: Nitrosococcus mobilis andNitrospira-like bacteria as dominant populations. Appl Environ Microbiol1998;64:3042–51.
25] Daims H, Nielsen JL, Nielsen PH, Schleifer KH, Wagner M. In situ characteriza-tion of Nitrospira-like nitrite-oxidizing bacteria active in wastewater treatmentplants. Appl Environ Microbiol 2001;67:5273–84.
26] Carrera J, Jubany I, Carvallo L, Chamy R, Lafuente J. Kinetic models for nitrifica-tion inhibition by ammonium and nitrite in a suspended and an immobilisedbiomass systems. Process Biochem 2004;39:1159–65.
27] Sanchez O, Marti MC, Aspe E, Roeckel M. Nitrification rates in a saline medium atdifferent dissolved oxygen concentrations. Biotechnol Lett 2001;23:1597–602.
28] Schramm A, de Beer D, van den Heuvel JC, Ottengraf S, Amann R. Microscaledistribution of populations and activities of Nitrosospira and Nitrospira spp.along a macroscale gradient in a nitrifying bioreactor: quantification byin situ hybridization and the use of microsensors. Appl Environ Microbiol1999;65:3690–6.
29] Laanbroek HJ, Bodelier PLE, Gerards S. Oxygen consumption kinetics of Nitro-somonas europaea and Nitrobacter hamburgensis grown in mixed continuouscultures at different oxygen concentrations. Arch Microbiol 1994;161:156–62.
30] Anthonisen AC, Loehr RC, Prakasam TBS, Srinath EG. Inhibition of nitrificationby ammonia and nitrous acid. J Wat Pollut Control Fed 1976;48:835–52.
31] Sinha B, Annachhatre AP. Partial nitrification-operational parameters andmicroorganisms involved. Rev Environ Sci Biotechnol 2007;6:285–313.
32] Villaverde S, Fdz-Polanco F, Garcia-Encina PA. Nitrifying biofilms acclima-tion to free ammonia in submerged biofilters. Start-up influence. Water Res2000;34:602–10.
33] Vadivelu VM, Keller J, Yuan Z. Effect of free ammonia on the respira-tion and growth processes of an enriched Nitrobacter culture. Water Res2007;41:826–34.
34] Turk O, Mavinic DS. Maintaining nitrite build-up in a system acclimated to freeammonia. Water Res 1989;23:1383–8.
35] Okabe S, Kindaichi T, Ito T. Fate of 14C-labeled microbial products derived fromnitrifying bacteria in autotrophic nitrifying biofilms. Appl Environ Microbiol2005;71:3987–94.
36] Kindaichi T, Ito T, Okabe S. Ecophysiological interaction between nitrify-ing bacteria and heterotrophic bacteria in autotrophic nitrifying biofilms asdetermined by microautoradiography-fluorescence in situ hybridization. ApplEnviron Microbiol 2004;70:1641–50.
37] Limpiyakorn T, Sakamoto Y, Kurisu F, Yagi O. Effect of ammonia andnitrite concentrations on communities and populations of ammonia-oxidizingbacteria in laboratory-scale continuous-flow reactors. FEMS Microbiol Ecol2007;60:501–12.
38] Dytczak MA, Londry KL, Oleszkiewicz JA. Activated sludge operational regimehas significant impact on the type of nitrifying community and its nitrificationrates. Water Res 2008;42:2320–8.
39] Kim D-J, Kim S-H. Effect of nitrite concentration on the distribution and com-petition of nitrite-oxidizing bacteria in nitratation reactor systems and theirkinetic characteristics. Water Res 2006;40:887–94.
Copyright © 2022 FDOKUMEN




















