
Rehabilitation of Swamp Paperbark ( Melaleuca ericifolia ) wetlands in south-eastern Australia:...
14
ORIGINAL PAPER Rehabilitation of Swamp Paperbark (Melaleuca ericifolia) wetlands in south-eastern Australia: effects of hydrology, microtopography, plant age and planting technique on the success of community-based revegetation trials Elisa J. Raulings Paul I. Boon Paul C. Bailey Michael C. Roache Kay Morris Randall Robinson Received: 8 December 2005 / Accepted: 24 October 2006 / Published online: 13 February 2007 Ó Springer Science+Business Media B.V. 2007 Abstract Wetlands dominated by Swamp Paper- barks (Melaleuca spp., Myrtaceae) are common in coastal regions across Australia. Many of these wetlands have been filled in for coastal develop- ment or otherwise degraded as a consequence of altered water regimes and increased salinity. Substantial resources, often involving community groups, are now being allocated to revegetating the remaining wetland sites, yet only rarely is the effectiveness of the rehabilitation strategies or on-ground procedures robustly assessed. As part of a larger project investigating the condition and rehabilitation of brackish-water wetlands of the Gippsland Lakes, we overlaid a scientifically informed experimental design on a set of community-based planting trials to test the effects of water depth, microtopography, plant age and planting method on the survival and growth of seedlings of Melaleuca ericifolia Sm. in Dowd Morass, a degraded, Ramsar-listed wetland in south-eastern Australia. Although previous laboratory and greenhouse studies have shown M. ericifolia seedlings to be salt tolerant, the strongly interactive effects of waterlogging and salinity resulted in high seedling mortality (>90%) in the field-based revegetation trials. Seedlings survived best if planted on naturally raised hummocks vegetated with Paspalum disti- chum L. (Gramineae), but their height was reduced compared with seedlings planted in shallowly flooded environments. Age of plants and depth of water were important factors in the survival and growth of M. ericifolia seedlings, whereas planting method seemed to have little effect on survival. Improved testing of revegeta- tion methods and reporting of success or other- wise of revegetation trials will improve the effectiveness and accountability of projects aim- ing to rehabilitate degraded coastal wetlands. Keywords Gippsland Lakes Á Hummock Á Microtopography Á Water regime Introduction Hydrology is the most powerful element control- ling the structure and function of wetland ecosys- tems. Bedford (1996) identified three core hydrological attributes affecting wetlands: the source of the water, the quality of the water, and the spatial and temporal characteristics of a E. J. Raulings (&) Á P. C. Bailey Á M. C. Roache Á K. Morris School of Biological Sciences and Australian Centre for Biodiversity: Analysis, Policy and Management, Monash University, Clayton 3800, Australia e-mail: [email protected] P. I. Boon Á R. Robinson Institute for Sustainability and Innovation, Victoria University, St. Albans 8001, Australia 123 Wetlands Ecol Manage (2007) 15:175–188 DOI 10.1007/s11273-006-9022-6
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Rehabilitation of Swamp Paperbark ( Melaleuca ericifolia ) wetlands in south-eastern Australia:...

ORIGINAL PAPER
Rehabilitation of Swamp Paperbark (Melaleuca ericifolia)wetlands in south-eastern Australia: effects of hydrology,microtopography, plant age and planting techniqueon the success of community-based revegetation trials
Elisa J. Raulings Æ Paul I. Boon Æ Paul C. Bailey Æ Michael C. Roache ÆKay Morris Æ Randall Robinson
Received: 8 December 2005 / Accepted: 24 October 2006 / Published online: 13 February 2007� Springer Science+Business Media B.V. 2007
Abstract Wetlands dominated by Swamp Paper-
barks (Melaleuca spp., Myrtaceae) are common in
coastal regions across Australia. Many of these
wetlands have been filled in for coastal develop-
ment or otherwise degraded as a consequence of
altered water regimes and increased salinity.
Substantial resources, often involving community
groups, are now being allocated to revegetating
the remaining wetland sites, yet only rarely is the
effectiveness of the rehabilitation strategies or
on-ground procedures robustly assessed. As part
of a larger project investigating the condition and
rehabilitation of brackish-water wetlands of the
Gippsland Lakes, we overlaid a scientifically
informed experimental design on a set of
community-based planting trials to test the effects
of water depth, microtopography, plant age and
planting method on the survival and growth of
seedlings of Melaleuca ericifolia Sm. in Dowd
Morass, a degraded, Ramsar-listed wetland
in south-eastern Australia. Although previous
laboratory and greenhouse studies have shown
M. ericifolia seedlings to be salt tolerant, the
strongly interactive effects of waterlogging and
salinity resulted in high seedling mortality
(>90%) in the field-based revegetation trials.
Seedlings survived best if planted on naturally
raised hummocks vegetated with Paspalum disti-
chum L. (Gramineae), but their height was
reduced compared with seedlings planted in
shallowly flooded environments. Age of plants
and depth of water were important factors in the
survival and growth of M. ericifolia seedlings,
whereas planting method seemed to have little
effect on survival. Improved testing of revegeta-
tion methods and reporting of success or other-
wise of revegetation trials will improve the
effectiveness and accountability of projects aim-
ing to rehabilitate degraded coastal wetlands.
Keywords Gippsland Lakes � Hummock �Microtopography � Water regime
Introduction
Hydrology is the most powerful element control-
ling the structure and function of wetland ecosys-
tems. Bedford (1996) identified three core
hydrological attributes affecting wetlands: the
source of the water, the quality of the water, and
the spatial and temporal characteristics of a
E. J. Raulings (&) � P. C. Bailey � M. C. Roache �K. MorrisSchool of Biological Sciences and Australian Centrefor Biodiversity: Analysis, Policy and Management,Monash University, Clayton 3800, Australiae-mail: [email protected]
P. I. Boon � R. RobinsonInstitute for Sustainability and Innovation, VictoriaUniversity, St. Albans 8001, Australia
123
Wetlands Ecol Manage (2007) 15:175–188
DOI 10.1007/s11273-006-9022-6

wetland’s wetting and drying cycle. The hydrological
attributes of wetlands across the world has been
altered, often with devastating impacts on wetland
distribution and integrity. In Australia for exam-
ple, altered water regimes affect or potentially
threaten 27% of the country’s highest-value wet-
lands, that is those described as Wetlands of
National and International Significance (Davis
et al. 2001). Not only have patterns of wetting
and drying been modified globally, but water
quality also is often very poor in the remaining
wetlands, commonly as a result of changed inter-
actions between wetland bathymetry, surface-
water flows and ground-water inputs (Middleton
1999). As an example, many Australian wetlands
are currently subject to secondary salinisation due
to the rise of saline water tables and increasing
river-water salinity (Hart et al. 2003).
Dowd Morass State Game Reserve is a large,
brackish-water wetland in the Gippsland Lakes
Ramsar site (Victoria, south-eastern Australia)
that exemplifies these issues of wetland degrada-
tion. In order to provide a water regime condu-
cive to waterfowl hunting and to limit seawater
intrusions from the neighboring Lake Wellington,
the wetland has been more-or-less permanently
inundated for the past three decades. Despite the
maintenance of high water levels, salinities in
Dowd Morass range from <0.2 to over 19 g l–1
(5–25 dS m–1; Sinclair Knight Merz 2001), due
mostly to the continuing inputs of saline water
from Lake Wellington. These changes to hydro-
logical attributes have had marked effects on the
wetland vegetation. The dominant woody species
in Dowd Morass is Melaleuca ericifolia Sm.
(Swamp Paperbark). Melaleuca ericifolia is a
small clonal tree in the Family Myrtaceae that
grows in coastal (freshwater and estuarine)
swamps across southern and eastern Australia,
from southern Tasmania through to northern
New South Wales. Constant flooding, combined
with high salinity, has resulted in death of adult
plants and very poor recruitment of juveniles into
the population at Dowd Morass.
Since the distribution and abundance of
M. ericifolia has decreased markedly with the
clearing and draining of wetlands along the entire
coast of Australia (Bowkett and Kirkpatrick
2003), a priority of natural-resource management
agencies and non-government organisations
throughout Australia is the rehabilitation of
the wetlands that contain, or did contain, M.
ericifolia and other Melaleuca species (de Jong
1997; Turner and Lewis 1997; de Jong 2000).
Substantial resources are now being allocated
to rehabilitating degraded coastal wetlands in
south-eastern Australia. The rehabilitation strat-
egy is based largely on revegetation efforts,
usually commissioned by management agencies
and then devolved to contractors or community
groups, who in turn frequently transfer revegeta-
tion methods to wetlands that have been devel-
oped in terrestrial or riparian environments. With
few exceptions, the effectiveness of wetland
revegetation efforts have not been experimentally
tested and there is little accountability of the
success of the projects that have been undertaken
(de Jong 2000; Close and Davidson 2002).
In this paper, we report on a 2-year study in
which we collaborated with community groups
working in Dowd Morass by providing an exper-
imental design to a set of revegetation trials that
tested the effectiveness of a range of techniques
commonly used in wetland rehabilitation in
south-eastern Australia. Specifically, we tested
the effectiveness of planting at different water
depths, with different planting methods, using
seedlings of different age classes, and planting
according to wetland microtopography. The
results of the study will be useful not only in
identifying the optimal methods for revegetating
degraded coastal wetlands with M. ericifolia, but
also in more general terms in reporting on
the effectiveness of projects aiming at wetland
rehabilitation.
Methods
Species description
Ten species of Melaleuca dominate coastal and
inland wetlands throughout Australia (Specht,
1990). The Swamp Paperbark, M. ericifolia, forms
a vegetation type commonly known as Swamp
Scrub: Melaleuca-dominated Swamp Scrub
extends throughout southern and eastern Australia
176 Wetlands Ecol Manage (2007) 15:175–188
123

and is especially extensive in the coastal districts of
Gippsland, Victoria (Bird 1962; Ladd et al. 1976).
Melaleuca ericifolia is a shrub or small tree,
reaching a maximum height of 8 m. It grows
mostly at low altitude in poorly drained soils,
swamps and stream flats (Bird 1962; Gibson et al.
1987; Costermans 1998). Although M. ericifolia is
typically a wetland or wetland-margin dominant
species (Bird 1961, 1962) it can grow also in well-
drained soil (Bowkett and Kirkpatrick 2003).
While many Melaleuca species are reliant on seed
to produce new individual plants, M. ericifolia is
unusual in that it is extensively clonal. Recruit-
ment from seed is comparatively rare and the
seed bank in sediments seems quite depauperate;
seed is stored in the canopy and released follow-
ing desiccation or fire (Gibson et al. 1987).
Although adult trees produce great numbers of
seeds, seed viability at Dowd Morass is typically
low. Moreover, the germination of M. ericifolia
seeds is delayed by submergence in water
(Ladiges et al. 1981), and seeds that do germinate
often float on the water surface and are unlikely
to take root under flooded conditions. Seedlings
do persist, however, on small, sometimes vegetated
mounds (‘hummocks’) of other clonal species,
such as Paspalum distichum L., Juncus ingens
N.A. Wakefield, and Phragmites australis (Cav.)
Steud across Dowd Morass.
Study site
Dowd Morass is a large (1,500 ha), deep (40–
60 cm), brackish-water wetland on the south-
western shore of Lake Wellington in the Gippsland
Lakes, Victoria, south-eastern Australia (Fig. 1).
The Gippsland Lakes are connected to the
Southern Ocean via an artificial opening cut to
the sea in 1889 at Lakes Entrance to improve
navigation access. Under the wetland inventory
system used in Victoria, Dowd Morass is classified
as a Deep Permanent Freshwater Marsh, despite
the salinity regime noted in the ‘‘Introduction’’
section.
The Morass has a catchment area of about
65 km2 and receives fresh water as overbank
flows from the Latrobe River on its northern
boundary, nutrient-enriched overland flow from
surrounding agricultural land, and brackish water
from Lake Wellington. The electrical conductivity
Fig. 1 Dowd MorassState Game Reserve,located between theLatrobe River to thenorth and LakeWellington to the east,in the Gippsland LakesRamsar site, south-eastern Australia
Wetlands Ecol Manage (2007) 15:175–188 177
123

of surface water in the wetland has fluctuated
over the past decade from about 5 - 25 dS m–1
(Sinclair Knight Merz 2001). It is thought that
Dowd Morass would have dried every 5 years or
so prior to European settlement; as noted in the
Introduction, the wetland has been almost
permanently inundated since 1975.
Revegetation trials
Melaleuca ericifolia tubestock was planted at two
sites in Dowd Morass (Fig. 1). These two sites
were chosen on the basis of internal homogeneity,
ease of access, presence of a suitable range of
water depths and hummocks, and the presence of
juvenile M. ericifolia in the surrounding areas.
Seed for the tubestock was collected from
M. ericifolia stands at Dowd Morass to ensure
local provenance, and germinated in composted
pine in a growing medium. Tubestock were
established from seed by a commercial nursery
(Gippsland Indigenous Plants Pty. Ltd., Valencia
Creek) and grown in this medium in 5-cm
diameter forestry tubes. Previous revegetation
projects using similar tubestock have over 90%
survival in terrestrial sites (John Topp, Gippsland
Indigenous Plants Pty. Ltd., personal communi-
cation). To assist in acclimatisation, tubestock
were placed in the Morass for 2–3 days before
each revegetation trial commenced.
The revegetation trials were undertaken in
collaboration with members of three community
groups interested in the ecology and manage-
ment of the Lake Wellington wetlands: Field and
Game Victoria (Sale and District), the Field
Naturalists Club of Victoria (Sale branch), and
Waterwatch Victoria. A small number of volun-
teers from other groups, for example, local
schools, also attended. Up to 60 volunteers
attended each planting session. Volunteers were
trained in planting techniques before each trial
to minimise differences in planting experience
among participants. Three experimental vegeta-
tion trials were undertaken. Of necessity, each
experimental trial developed iteratively on the
basis of knowledge generated in prior trials and
on the number of community volunteers avail-
able. Due to these factors, methods varied
slightly from trial to trial.
Experiment 1: effect of water depth and age class
Experiment 1 used a split-plot design to test the
effects of water depth and plant age on the
success of seedling establishment. This experi-
ment was undertaken at Site 1 in March 2004
(Fig. 1). Seed of M. ericifolia would naturally
germinate and grow during spring and summer,
so seedlings growing naturally in the Morass
would be 4 to 6-months-old by March. Four-
month-old and 6-month-old seedlings were
therefore selected to replicate the natural age
classes of seedlings in the wetland. The seed-
lings varied not only in age but also in height:
4-month-old seedlings were typically 16 cm high
whereas 6-month-old seedlings were typically
54 cm high.
Four replicate plots (each 18 m · 10 m) were
placed parallel to the water line at three water
depths at Site 1: (i) ‘exposed’ (saturated sediment;
mean surface water depth = 0 cm); (ii) ‘water-
logged’ (mean water depth = 1 cm); and (iii)
‘flooded’ (mean water depth = 9 cm). Three
water depths were chosen because community-
based replanting activities may have little control
over the depth of water in a wetland targeted for
revegetation (de Jong 2000). Within each plot,
4-month-old plants (n = 20) and 6-month-old
plants (n = 25) were planted 2 m apart using a
mattock to create a hole in the sediment into
which the tubestock was inserted. After planting,
volunteers measured the height of the tallest
green foliage (‘plant height’) and water depth for
each plant. Plant survival, plant height, and water
depth were recorded monthly for the following
8 months. A datalogger, located approximately
200 m from Site 1, collected data hourly for
electrical conductivity and water depth.
On the basis of the site’s long term meteoro-
logical records, it was anticipated that the water
level at Site 1 would initially fall, and gradually
increase with the onset of winter rains. However,
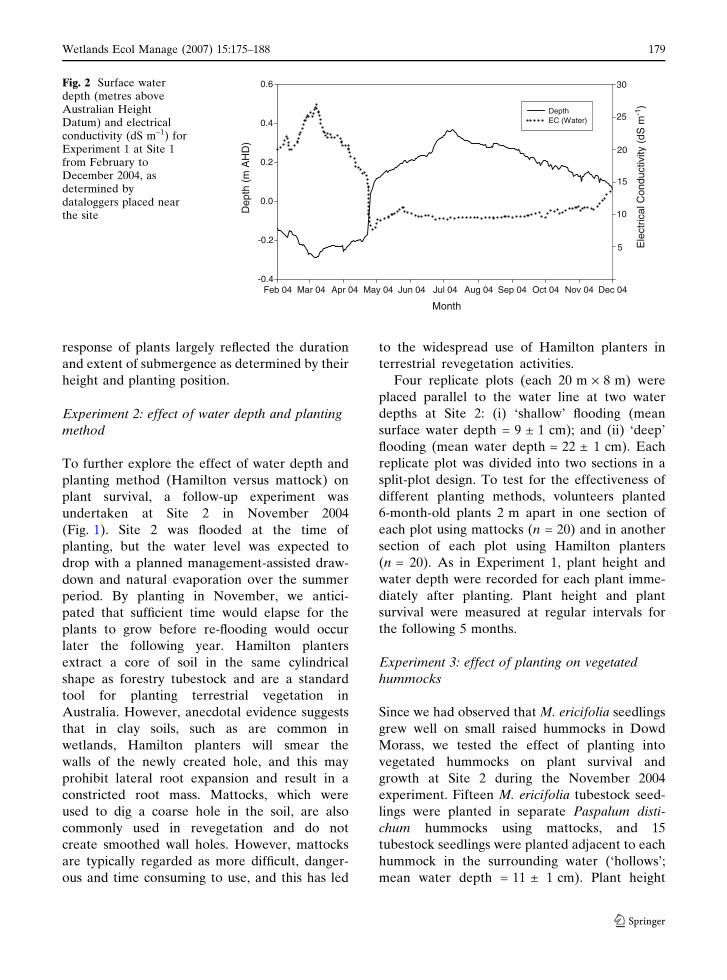
a storm in late April 2004 increased the water
depth at Site 1 by more than 30 cm (Fig. 2). Due
to ongoing wet weather, water levels continued to
increase until July 2004 and fell slowly thereafter.
Due to the unexpected flood event, the planned
water depth treatments became obscured within
1 month of planting. After the flood event, the
178 Wetlands Ecol Manage (2007) 15:175–188
123
response of plants largely reflected the duration
and extent of submergence as determined by their
height and planting position.
Experiment 2: effect of water depth and planting
method
To further explore the effect of water depth and
planting method (Hamilton versus mattock) on
plant survival, a follow-up experiment was
undertaken at Site 2 in November 2004
(Fig. 1). Site 2 was flooded at the time of
planting, but the water level was expected to
drop with a planned management-assisted draw-
down and natural evaporation over the summer
period. By planting in November, we antici-
pated that sufficient time would elapse for the
plants to grow before re-flooding would occur
later the following year. Hamilton planters
extract a core of soil in the same cylindrical
shape as forestry tubestock and are a standard
tool for planting terrestrial vegetation in
Australia. However, anecdotal evidence suggests
that in clay soils, such as are common in
wetlands, Hamilton planters will smear the
walls of the newly created hole, and this may
prohibit lateral root expansion and result in a
constricted root mass. Mattocks, which were
used to dig a coarse hole in the soil, are also
commonly used in revegetation and do not
create smoothed wall holes. However, mattocks
are typically regarded as more difficult, danger-
ous and time consuming to use, and this has led
to the widespread use of Hamilton planters in
terrestrial revegetation activities.
Four replicate plots (each 20 m · 8 m) were
placed parallel to the water line at two water
depths at Site 2: (i) ‘shallow’ flooding (mean
surface water depth = 9 ± 1 cm); and (ii) ‘deep’
flooding (mean water depth = 22 ± 1 cm). Each
replicate plot was divided into two sections in a
split-plot design. To test for the effectiveness of
different planting methods, volunteers planted
6-month-old plants 2 m apart in one section of
each plot using mattocks (n = 20) and in another
section of each plot using Hamilton planters
(n = 20). As in Experiment 1, plant height and
water depth were recorded for each plant imme-
diately after planting. Plant height and plant
survival were measured at regular intervals for
the following 5 months.
Experiment 3: effect of planting on vegetated
hummocks
Since we had observed that M. ericifolia seedlings
grew well on small raised hummocks in Dowd
Morass, we tested the effect of planting into
vegetated hummocks on plant survival and
growth at Site 2 during the November 2004
experiment. Fifteen M. ericifolia tubestock seed-
lings were planted in separate Paspalum disti-
chum hummocks using mattocks, and 15
tubestock seedlings were planted adjacent to each
hummock in the surrounding water (‘hollows’;
mean water depth = 11 ± 1 cm). Plant height
Month
Feb 04 Mar 04 Apr 04 May 04 Jun 04 Jul 04 Aug 04 Sep 04 Oct 04 Nov 04 Dec 04
Dep
th (
m A
HD
)
-0.4
-0.2
0.0
0.2
0.4
0.6
Ele
ctric
al C
ondu
ctiv
ity (
dS m
-1)
DepthEC (Water)
5
10
15
20
25
30Fig. 2 Surface waterdepth (metres aboveAustralian HeightDatum) and electricalconductivity (dS m–1) forExperiment 1 at Site 1from February toDecember 2004, asdetermined bydataloggers placed nearthe site
Wetlands Ecol Manage (2007) 15:175–188 179
123

and water depth were measured for each plant at
the time of planting and at regular intervals for
the following 5 months.
Soil chemistry
A single soil core was taken from each plot at the
two sites and separated into two depths: 0–10 cm
and 10–20 cm. At Site 1 cores were taken only at
the time of planting in March 2004. At Site 2,
cores were taken at the time of planting (Novem-
ber 2004) and again at the end of the experiment
when the site had dried (April 2005). Soil cores
were weighed before and after drying at 105�C to
determine soil moisture content. Soil electrical
conductivity (EC) and pH of 1:5 soil:water
extracts were measured using the procedures of
Rayment and Higginson (1992). Electrical
conductivity and pH were measured using a
calibrated TPSTM WP-81 probe and meter
(Springwood, Queensland, Australia). The in situ
soil salinity was calculated using the soil moisture
content and the EC of the 1:5 soil extracts.
Statistical analyses
Data were analysed using SYSTATTM version 11,
and screened for missing values, outliers and
normality, then transformed where required prior
to analysis. Experiments 1 and 2 were unreplicat-
ed, split-plot designs, so analysis followed the
procedures outlined by Underwood (1997) and
Quinn and Keough (2002). Water depth (Exper-
iments 1 and 2), age class (Experiment 1), and
planting method (Experiment 2) were treated as
fixed factors and plot location a random factor.
Age class (Experiment 1) and planting method
(Experiment 2) were treated as within-plot fac-
tors. Soil chemistry data were analysed by treat-
ing soil depth as a fixed within-plots factor.
Variables measured over time were analysed with
time as a fixed within-plot factor.
Logistic regression was used to determine
which factors contributed most to plant survival.
Logistic regression predicts the likelihood of
plants surviving in a particular treatment relative
to the likelihood that plants will die. McFadden’s
q, which is conceptually similar to the r2 value in
linear regression, measures the strength of the
association between the outcome (survival or
mortality) and the predictors (e.g. age class).
Values in the 0.2–0.4 range are considered to be
highly satisfactory (Tabachnick and Fidell 2001).
The odds ratio measures the increase (or
decrease, if the ratio is < 1) in the probability of
a plant surviving when the value of the predictor
unit (e.g. water depth) increases by one step
(Tabachnick and Fidell 2001). It is calculated in
relation to a reference group; the reference group
for this model is 6-month-old plants in the
exposed treatment (Experiment 1) and plants in
shallow water (Experiment 2). A chi-square
analysis, which compares the constant-only model
with the full model, was used to determine
whether the predictor variables reliably predicted
plant survival. A useful guide to logistic regres-
sion can be found at http://www.statpages.org/
logistic.htm.
Results
Experiment 1: effect of water depth and age
class
Sediment and water characteristics
At the time of planting a piezometer located at
the perimeter of the wetland indicated that the
groundwater was likely to be <50 cm below the
sediment surface in the wetland. The conductivity
of the groundwater and surface water was
11 dS m–1 and 25 dS m–1, respectively. At the
start of the experiment, the mean water level in
the exposed, waterlogged and flooded treatments
were 0, 1, and 9 cm, respectively (Table 1).
Depending on wind strength and direction, the
water surface could fluctuate by about 10 cm. The
storm of late April 2004 increased the water
depth by more than 30 cm at Site 1 and lowered
the salinity of the surface water to <10 dS m–1
(Fig. 2). Water levels continued to rise until July
2004, then fell steadily thereafter.
The pH of the soil at Site 1 ranged from 3.7 to
5.6, indicating mildly to strongly acidic soils. Soil
cores 0–10 cm deep (mean pH 4.6) were slightly
more alkaline than soil from the 10–20 cm depth
(mean pH 4.2: F1,18 = 7.757, P = 0.012; Table 2).
180 Wetlands Ecol Manage (2007) 15:175–188
123

There was no significant difference in soil pH
across the three water depth treatments
(P = 0.118). There were, however, significant
differences in soil moisture across water depth
treatments (F2, 18 = 9.059, P = 0.002) and soil core
depths (F1, 18 = 17.388, P = 0.001); the interaction
term was not significant. Soil moisture increased
with increasing water depth at Site 1 and, as
expected, was significantly higher in flooded plots
than in exposed plots (Tukey HSD, P = 0.014).
Soil from the upper 0–10 cm layer had signifi-
cantly less soil moisture than the deeper 10–20 cm
layer (Table 2).
Soil salinity, adjusted for in situ soil moisture,
ranged from ~ 16–36 dS m–1 at Site 1 (Table 2).
There was a significant effect of both water depth
treatment (F2, 18 = 4.631, P = 0.024) and soil
core depth (F1, 18 = 15.091, P = 0.001) on soil
conductivity, but there was no interactive effect.
Soil electrical conductivity was significantly greater
in exposed and waterlogged sites than flooded
sites, and soil cores 0–10 cm deep were signifi-
cantly more saline than cores 10–20 cm deep.
Note that the electrical conductivity of surface
water in the wetland has fluctuated over the past
decade from 5 to 25 dS m–1 (Sinclair Knight Merz
Table 1 Percentage survival, plant height and water depthfor Experiment 1 over a 9-month period for 4-month-oldand 6-month-old seedlings planted at three water depth
treatments at Site 1 in Dowd Morass. All the 4-month-oldplants died in May 2004
Date Age class (months) Water depth treatment Water depth (cm) % Survival Plant height (cm)
March 2004 4 Exposed 0 100 ± 0.0 15 ± 3Waterlogged 1 ± 1 100 ± 0.0 14 ± 4Flooded 9 ± 1 100 ± 0.0 18 ± 7
6 Exposed 0 ± 0 100 ± 0.0 58 ± 31Waterlogged 2 ± 1 100 ± 0.0 53 ± 19Flooded 9 ± 0 100 ± 0.0 50 ± 20
April 2004 4 Exposed 1 ± 1 29 ± 19 17 ± 21Waterlogged 3 ± 0 25 ± 7 13 ± 5Flooded 11 ± 0 26 ± 9 16 ± 6
6 Exposed 2 ± 1 76 ± 9 58 ± 27Waterlogged 3 ± 1 52 ± 10 50 ± 6Flooded 11 ± 1 59 ± 10 47 ± 15
May 2004 4 Exposed Not recorded 0 –Waterlogged Not recorded 0 –Flooded Not recorded 0 –
6 Exposed 36 ± 1 54 ± 5 58 ± 27Waterlogged 40 ± 1 37 ± 13 52 ± 31Flooded 47 ± 1 33 ± 10 50 ± 12
June 2004 6 Exposed 43 ± 1 42 ± 4 56 ± 18Waterlogged 48 ± 2 34 ± 12 49 ± 25Flooded 52 ± 0 29 ± 9 50 ± 8
July 2004 6 Exposed 54 ± 1 42 ± 9 56 ± 27Waterlogged 61 ± 2 8 ± 2 56 ± 19Flooded 64 ± 2 18 ± 6 53 ± 54
August 2004 6 Exposed 47 ± 1 43 ± 5 51 ± 12Waterlogged 53 ± 2 25 ± 10 52 ± 25Flooded 59 24 57
September 2004 6 Exposed 40 ± 1 42 ± 1 43 ± 12Waterlogged 46 ± 1 32 ± 13 46 ± 15Flooded 51 ± 0 29 ± 10 48 ± 19
November 2004 6 Exposed 38 ± 1 3 ± 2 –Waterlogged 35 ± 1 8 ± 2 –Flooded 44 ± 0 2 ± 1 –
Data represents mean ± standard error (n = 20 for 4-month-old plants; n = 25 for 6-month-old plants)
Wetlands Ecol Manage (2007) 15:175–188 181
123
2001), indicating that the soil salinities in soil pore
water were likely to be considerably higher than
were surface-water salinities.
Plant responses
Due to the unexpected flood in late April 2004, the
effects of the planned water depth treatments
could be evaluated using only data collected in
early April 2004. After this time plant responses
also reflected the duration and extent of submer-
gence. In April, 1 month after planting and prior to
the flood event, there was very high mortality of
plants (Table 1). Both the age of plants and water
depth were significant factors in seedling survival,
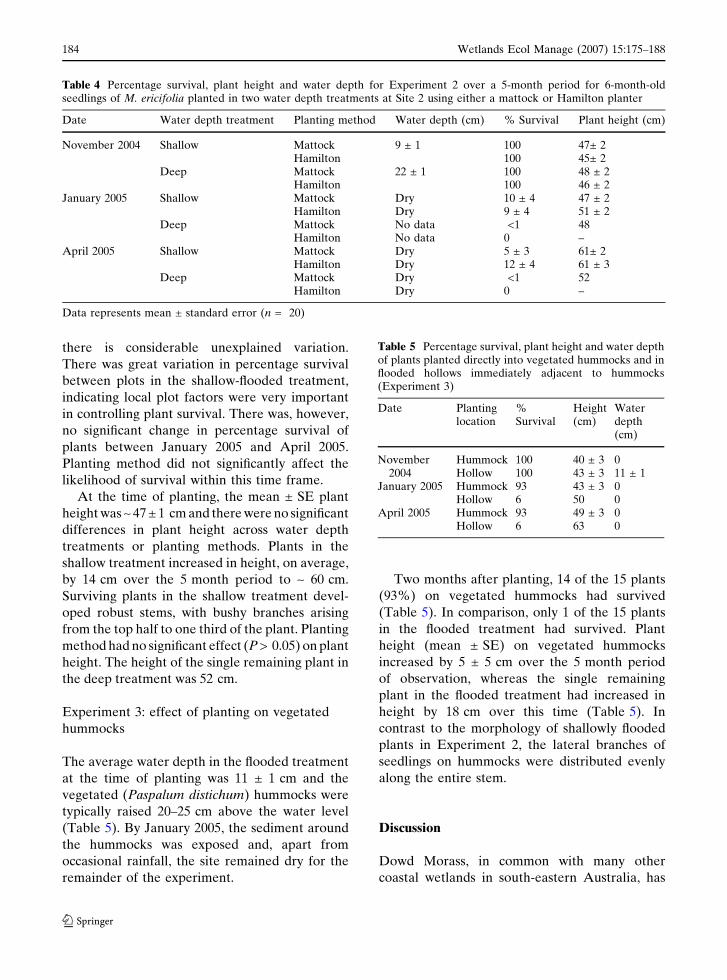
but there was no interaction. As indicated by the
odds ratio (Table 3), seedlings were three times
more likely to survive in the exposed treatment
than in the waterlogged treatment, although the
strength of this effect is somewhat equivocal given
the variance associated with the estimate (Wald
statistic = 3.5, P < 0.001; Table 3). Survival was
greater in older plants, and only 27% of the
4-month-old plants survived the first month
whereas 65% of the 6-month-old plants survived
for this time (Table 1). Statistical analysis demon-
strated that 4-month-old plants were significantly
more likely to die than survive (Wald statistic =
-5.7, P < 0.001; Table 3). The chi-square test for
the difference between the constant-only model
and the model specified was significant (P < 0.001),
although McFadden’s q value of 0.1 was not
completely satisfactory. Much of the unexplained
variation may be accounted for by differences in
survival across plots.
The height of many plants decreased over the
first month as a result of die-back of the apical
meristem (Table 1). Water depth was a significant
factor in determining changes in plant height
(F2,238 = 4.346, P = 0.048); plants in the exposed
treatment were significantly taller than those in
the flooded treatment (Tukey’s HSD, P = 0.039).
In contrast, age class was not a significant factor
in predicting plant height; this may be related to
the high mortality of 4-month-old plants and the
loss of statistical power. There was, however, a
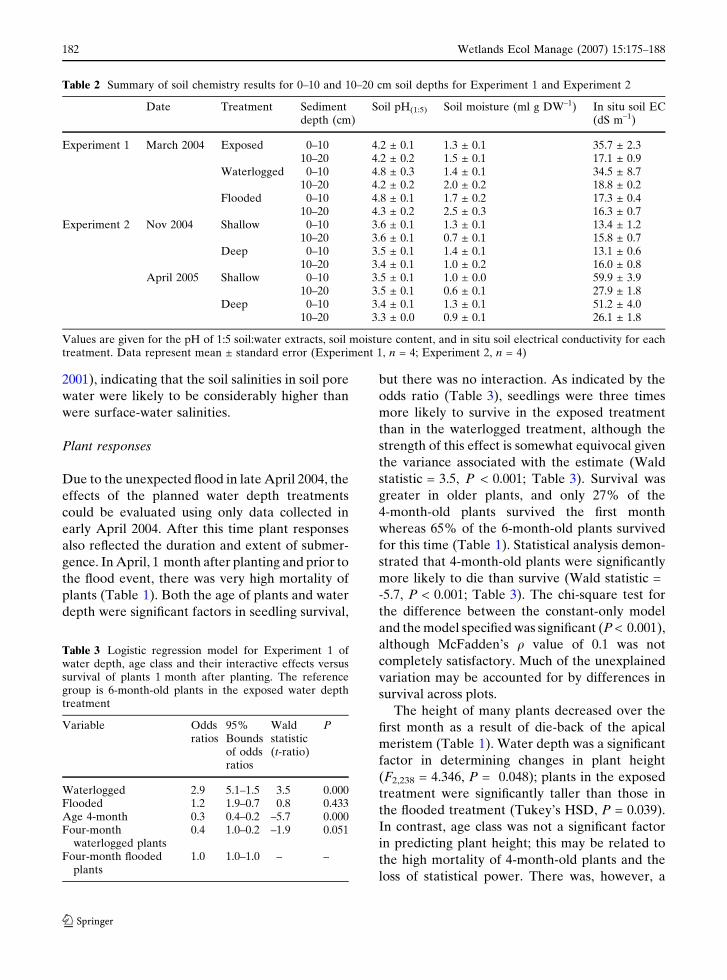
Table 2 Summary of soil chemistry results for 0–10 and 10–20 cm soil depths for Experiment 1 and Experiment 2
Date Treatment Sedimentdepth (cm)
Soil pH(1:5) Soil moisture (ml g DW–1) In situ soil EC(dS m–1)
Experiment 1 March 2004 Exposed 0–10 4.2 ± 0.1 1.3 ± 0.1 35.7 ± 2.310–20 4.2 ± 0.2 1.5 ± 0.1 17.1 ± 0.9
Waterlogged 0–10 4.8 ± 0.3 1.4 ± 0.1 34.5 ± 8.710–20 4.2 ± 0.2 2.0 ± 0.2 18.8 ± 0.2
Flooded 0–10 4.8 ± 0.1 1.7 ± 0.2 17.3 ± 0.410–20 4.3 ± 0.2 2.5 ± 0.3 16.3 ± 0.7
Experiment 2 Nov 2004 Shallow 0–10 3.6 ± 0.1 1.3 ± 0.1 13.4 ± 1.210–20 3.6 ± 0.1 0.7 ± 0.1 15.8 ± 0.7
Deep 0–10 3.5 ± 0.1 1.4 ± 0.1 13.1 ± 0.610–20 3.4 ± 0.1 1.0 ± 0.2 16.0 ± 0.8
April 2005 Shallow 0–10 3.5 ± 0.1 1.0 ± 0.0 59.9 ± 3.910–20 3.5 ± 0.1 0.6 ± 0.1 27.9 ± 1.8
Deep 0–10 3.4 ± 0.1 1.3 ± 0.1 51.2 ± 4.010–20 3.3 ± 0.0 0.9 ± 0.1 26.1 ± 1.8
Values are given for the pH of 1:5 soil:water extracts, soil moisture content, and in situ soil electrical conductivity for eachtreatment. Data represent mean ± standard error (Experiment 1, n = 4; Experiment 2, n = 4)
Table 3 Logistic regression model for Experiment 1 ofwater depth, age class and their interactive effects versussurvival of plants 1 month after planting. The referencegroup is 6-month-old plants in the exposed water depthtreatment
Variable Oddsratios
95%Boundsof oddsratios
Waldstatistic(t-ratio)
P
Waterlogged 2.9 5.1–1.5 3.5 0.000Flooded 1.2 1.9–0.7 0.8 0.433Age 4-month 0.3 0.4–0.2 –5.7 0.000Four-month
waterlogged plants0.4 1.0–0.2 –1.9 0.051
Four-month floodedplants
1.0 1.0–1.0 – –
182 Wetlands Ecol Manage (2007) 15:175–188
123

significant interactive effect of water depth with
age class (F2,238 = 7.88, P < 0.001); the height of
4-month-old plants in April is less in the water-
logged treatment than either the exposed or
flooded treatment, but for 6-month-old seedlings
the height is lowest in both the waterlogged and
flooded treatments (Table 1). As before, among-
plot effects were a significant factor in determin-
ing plant height.
As a result of the flood in April, by May 2004
all the 4-month-old seedlings were deeply sub-
merged (shoot tips >20 cm below the water
surface) and had died (Table 1). Although the
height of some 6-month-old plants exceeded the
water depth, the plant tips did not emerge from
the water but instead lay flat on the surface. The
extent and duration of submergence was consid-
erably less for these taller, older plants than for
the younger tubestock and this was correlated
with their improved survival: more than 20% of
6-month-old plants survived 3–4 months of com-
plete submergence, and survival was highest
(40%) in exposed plants. Nevertheless, prolonged
inundation of 6-month-old plants caused the
growing tip to die, emergent leaves to be shed,
and the bark of the main stem to become thick
and spongy where submerged.
In August and September 2004, when water
levels started to drop again, survival of 6-month-
old plants appeared to increase in the water-
logged and flooded treatments (Table 1), possibly
as new shoots were produced on plants that had
previously been scored as ‘dead’ or because the
green shoots were easier to see when they were
not immersed in turbid water. The increase in
survival, however, proved to be transient and
more plants died within 1 month as water levels
fell further. Of the total number of seedlings
planted originally (540), <10% survived over the
8 months of observation.
Experiment 2: effect of water depth
and planting method
Sediment and water characteristics
Water levels fell over the experimental period
and, by January 2005, the surface water had
evaporated from the shallow plots to leave the
sediments fully exposed (Table 4). Water in the
deep plots at this time was 3–5 cm deep. By April
2005, all the shallow and the deep flooded sites
were dry. Apart from occasional rainfall, Site 2
remained dry for the remainder of the observa-
tion period.
The pH of soil at Site 2 ranged from 3.3 to
3.8, again indicating mildly to strongly acidic
conditions (Table 2). Shallow-flooded soils had
significantly higher pH than deep-flooded soils
(F1,12 = 12.002, P = 0.005), but there was no
significant change in soil pH over the duration
of the experiment. Soil from 0–10 cm depth had
significantly higher soil moisture than soil
10–20 cm deep (F1, 12 = 62.049, P < 0.001),
and soil cores taken from deep flooded sites had
significantly greater soil moisture than cores
in shallow flooded sites (F1, 12 = 21.694,
P = 0.001; Table 2).
When seedlings were planted in November 2004,
soil salinities were ~ 13 dS m–1 in the 0–10 cm
profile and ~ 16 dS m–1 in the 10–20 cm profile
(Table 2). There were no significant differences in
soil salinity across soil depths or water depth
treatments at this time. As the wetland dried out
however, the electrical conductivity of soil from the
0–10 cm depth increased to ~ 60 ± 4 dS m–1 in the
shallow flooded site and 51 ± 4 dS m–1 in the deeply
flooded site. These soil salinities are very much
higher than the range of water-column salinities
observed in the wetland. Salinities in the 10–20 cm
deep profile were around half the values observed
for the upper soil layers. By the end of April 2005,
salt encrusted the soil surface in both shallow and
deep flooded sites.
Plant responses
The logit regression model for seedling survival
2 months after planting at Site 2 was significant
(P < 0.001), and increasing water depth clearly
affected the likelihood of seedling survival. Two
months after planting, only 9 ± 3% of plants in
the shallow water treatment had survived
(Table 4). Deeply flooded plants were even more
likely to die (Wald statistic = –11.4, P < 0.001),
and only one plant had survived after 2 months
under these conditions. However, the low McF-
adden’s q2 value (0.01) for the model suggests
Wetlands Ecol Manage (2007) 15:175–188 183
123
there is considerable unexplained variation.
There was great variation in percentage survival
between plots in the shallow-flooded treatment,
indicating local plot factors were very important
in controlling plant survival. There was, however,
no significant change in percentage survival of
plants between January 2005 and April 2005.
Planting method did not significantly affect the
likelihood of survival within this time frame.
At the time of planting, the mean ± SE plant
height was ~ 47 ± 1 cm and there were no significant
differences in plant height across water depth
treatments or planting methods. Plants in the
shallow treatment increased in height, on average,
by 14 cm over the 5 month period to ~ 60 cm.
Surviving plants in the shallow treatment devel-
oped robust stems, with bushy branches arising
from the top half to one third of the plant. Planting
method had no significant effect (P > 0.05) on plant
height. The height of the single remaining plant in
the deep treatment was 52 cm.
Experiment 3: effect of planting on vegetated
hummocks
The average water depth in the flooded treatment
at the time of planting was 11 ± 1 cm and the
vegetated (Paspalum distichum) hummocks were
typically raised 20–25 cm above the water level
(Table 5). By January 2005, the sediment around
the hummocks was exposed and, apart from
occasional rainfall, the site remained dry for the
remainder of the experiment.
Two months after planting, 14 of the 15 plants
(93%) on vegetated hummocks had survived
(Table 5). In comparison, only 1 of the 15 plants
in the flooded treatment had survived. Plant
height (mean ± SE) on vegetated hummocks
increased by 5 ± 5 cm over the 5 month period
of observation, whereas the single remaining
plant in the flooded treatment had increased in
height by 18 cm over this time (Table 5). In
contrast to the morphology of shallowly flooded
plants in Experiment 2, the lateral branches of
seedlings on hummocks were distributed evenly
along the entire stem.
Discussion
Dowd Morass, in common with many other
coastal wetlands in south-eastern Australia, has
Table 4 Percentage survival, plant height and water depth for Experiment 2 over a 5-month period for 6-month-oldseedlings of M. ericifolia planted in two water depth treatments at Site 2 using either a mattock or Hamilton planter
Date Water depth treatment Planting method Water depth (cm) % Survival Plant height (cm)
November 2004 Shallow Mattock 9 ± 1 100 47± 2Hamilton 100 45± 2
Deep Mattock 22 ± 1 100 48 ± 2Hamilton 100 46 ± 2
January 2005 Shallow Mattock Dry 10 ± 4 47 ± 2Hamilton Dry 9 ± 4 51 ± 2
Deep Mattock No data <1 48Hamilton No data 0 –
April 2005 Shallow Mattock Dry 5 ± 3 61± 2Hamilton Dry 12 ± 4 61 ± 3
Deep Mattock Dry <1 52Hamilton Dry 0 –
Data represents mean ± standard error (n = 20)
Table 5 Percentage survival, plant height and water depthof plants planted directly into vegetated hummocks and inflooded hollows immediately adjacent to hummocks(Experiment 3)
Date Plantinglocation
%Survival
Height(cm)
Waterdepth(cm)
November2004
Hummock 100 40 ± 3 0Hollow 100 43 ± 3 11 ± 1
January 2005 Hummock 93 43 ± 3 0Hollow 6 50 0
April 2005 Hummock 93 49 ± 3 0Hollow 6 63 0
184 Wetlands Ecol Manage (2007) 15:175–188
123

been degraded by near-permanent flooding, low
soil pH and highly saline soil and surface water (de
Jong 1997; Turner and Lewis 1997; de Jong 2000;
Davis et al. 2001). The very poor survival of
tubestock seedlings observed in our study is
undoubtedly related to this highly stressful
combination of adverse environmental factors,
and demonstrates the difficulties associated with
rehabilitating degraded brackish-water wetlands.
The study clearly demonstrated that age class,
water depth and topography were critical factors
in the survival and growth of M. ericifolia seed-
lings. Due to the low survival, it was more difficult
to assess the importance of planting method, but
preliminary results suggest little difference between
Hamilton planters and the use of a mattock.
Our experience at Dowd Morass shows how
water regime and water quality interact to control
seedling establishment and thus affect the likely
success of revegetation trials. Location from the
waterline, up a strong water depth gradient,
strongly influenced soil salinities throughout the
surface (0–20 cm) soil profile in Dowd Morass. In
sediments that had dried out, the in situ soil
salinities increased dramatically, due to both the
concentration and accumulation of salts. The soil
salinity increased fourfold in the surface (0–10 cm)
layers and doubled in the deeper (10–20 cm)
layers. Moreover, the salinity of the soil pore
water (up to 60 dS m–1) almost always exceeded
the salinity of surface water in the wetland
(10–30 dS m–1). In locations where the sediments
had been exposed and then re-flooded, the soil
salinities may greatly exceed the salinity of the
overlying water because of the accumulation in
surface profiles of salts from deeper in the soil
profile. In contrast, where the sediments rarely
become exposed, the soil salinities may better
reflect those of the surface water or ground water.
Measurement of the water column salinity when
assessing environmental conditions is therefore
not a good indicator of salinities that rooted plants
are likely to experience in revegetation trials and
rehabilitation strategies.
Effect of seedling age
The survival of M. ericifolia seedlings at Dowd
Morass depended strongly on the age of the
tubestock plants. The mortality of 4-month-old
plants 1 month after planting was at least 20%
greater than that of 6-month-old seedlings at
the same time. Four-month-old plants had a
small root system that did not bind soil, and it
was difficult to plant these young seedlings in
the wetland without damaging their roots. In
contrast, 6-month-old plants had a larger root
system and planting was comparatively easy
across all water depth treatments. Damage to
the roots of younger seedlings may also have
increased salt uptake by the plants: the smaller
root system of 4-month-old plants may have
restricted them to the more saline surface layers
of the soil existing in the exposed and water-
logged sites, whereas the better developed roots
of 6-month-old plants may have been able to
penetrate to less saline zones deeper in the soil
profile.
Effect of water depth and soil salinity
Seedlings and young plants of many Melaleuca
species, including M. ericifolia, are known to be
highly tolerant of waterlogging (Ladiges et al.
1981; de Jong 2000; Salter et al. 2006), and are
capable of active root growth in waterlogged soils
through the production of aerenchyma in the
roots (Ladiges et al. 1981). It is noteworthy,
however, that the survival of 6-month-old seed-
lings of M. ericifolia in the exposed (i.e., dry to
semi-damp) plots was 15–25% higher than for
seedlings planted in the waterlogged or fully
flooded plots 1 month after planting. In a reveg-
etation experiment in New South Wales, M. erici-
folia had high survivorship under extremes of
inundation and drought (de Jong 2000), but the
soil was markedly less saline than in our trials at
Dowd Morass.
The depth to which plants are flooded imme-
diately after planting may be an important factor
in the survival of M. ericifolia seedlings during
revegetation efforts. Once submerged, all the
4-month-old seedlings had died within 1 month;
in contrast, some 6-month-old plants survived up
to 8 months fully submerged, and survival was
highest in exposed plots. Similarly, Denton and
Ganf (1994) reported that 4-month seedlings of
M. halmaturorum did not survive complete
Wetlands Ecol Manage (2007) 15:175–188 185
123

inundation over a 6-week period in a wetland in
South Australia. In a laboratory experiment, 1-
year-old M. halmaturorum plants covered by
water to 50% of their height had > 86% survi-
vorship after 14 weeks, whereas plants com-
pletely submerged had only 29% survival
(Denton and Ganf 1994). In Experiment 2,
seedlings of M. ericifolia covered by 20% of their
height (i.e., in the shallow flooded treatment) had
9% survival after 2 months, whereas plants cov-
ered by 44% (i.e., the deep flooded treatment)
had only one individual remaining by the time
observations had ceased.
There is likely to be a strong interaction in
field-based revegetation trials between age of
plant and depth of water on the survival of
seedling tubestock. Clearly, the taller, older
seedlings should be better able to withstand
inundation since they can maintain a greater
proportion of their foliage above the water
surface. This prediction was vindicated by the
results of Experiment 1; where on average
only 18% of the shoot of the 6-month-old
plants was submerged, compared with ~ 50%
of the shoot in 4-month-old seedlings. This
difference was reflected in the markedly better
survival of the older tubestock under flooded
conditions.
Seedlings of M. ericifolia are reportedly
tolerant also of high salinities (van der Moezel
et al. 1991; Salter et al. 2006). In a mesocosm
experiment, 90% of M. ericifolia seedlings
survived 10 weeks in exposed sediments where
soil salinities exceeded 66 dS m–1; however,
when these sediments were waterlogged or
flooded, there was 100% mortality within
9 weeks (Salter et al. 2006). Similar results were
observed in Experiment 2, since seedlings in the
shallow flooded treatment had high mortality in
the first month when the sediments were
flooded but, once the sediments became
exposed, the surviving plants persisted in soil
salinities ~ 60 dS m–1 for at least 5 months.
Nevertheless, the interaction of high soil salin-
ities with water depth is likely to have contrib-
uted to the very high mortality of seedlings at
Dowd Morass. The extremely low pH of the
soil at the two revegetation sites probably also
contributed substantially to the poor revegeta-
tion success.
Importance of hummocks
In contrast to the high mortality of seedlings
planted directly in water, planting tubestock into
raised hummocks vegetated with a dense growth
of Paspalum distichum increased markedly the
survival of M. ericifolia tubestock at Dowd
Morass. This finding is consistent with our field
observations that M. ericifolia seedlings and juve-
niles predominantly occur on hummocks that are
raised, even slightly, out of the surrounding water,
and can be explained in terms of hummocks
offering a refuge from the stressful combination of
waterlogging, salinity and soil acidity occurring in
the surrounding sediments.
Vegetated hummocks are composed predomi-
nantly of decomposing organic matter with large air
spaces, and may enhance the survival of M. ericifolia
by elevating the shoots out of the water column and
allowing improved root aeration. Vegetated hum-
mocks may also be less affected by salinity than
lower lying, waterlogged areas; the vegetation and
organic matter may facilitate the collection of
rainfall, favouring percolation and leaching of salts
from the root zone of seedlings.
There may however be an adverse effect of
hummocks on plant growth, since young plants on
hummocks may become water stressed during
periods of low rainfall if roots do not rapidly
locate deeper water sources (Rothwell et al.
1993). Although plants on vegetated hummocks
had the best survivorship, growth of M. ericifolia
was almost three times better in shallow flooded
sites that dried out over time compared with that
of plants in vegetated hummocks.
Implications for revegetation trials
at the landscape scale
Despite the reported tolerance of M. ericifolia
seedlings to inundation and salinity, our research
demonstrated that, in wetlands that experience
high salinities, planting tubestock directly into
waterlogged soils, irrespective of water depth, is
likely to be almost completely unsuccessful as a
186 Wetlands Ecol Manage (2007) 15:175–188
123

revegetation strategy. This result is most likely a
consequence of the interactive, and detrimental,
effects of waterlogging with soil salinity and/or
low pH on seedling survival. The combined
impacts of waterlogging and salinity, however,
may be minimized by selecting elevated planting
spots, after consideration of microrelief, soil
characteristics, species, water depth and local
climate (Roy et al. 1999). In a site such as Dowd
Morass, it is preferable to use the natural topog-
raphy of the wetland for revegetation, using
raised sites such as existing vegetated hummocks,
Phragmites beds, and localised mounds formed
through tree fall.
Where advantage cannot be taken of the
natural microtopography, areas may have to be
artificially raised to improve the chances of
seedling survival. Planting onto hummocks is
similar to a site preparation technique known as
mounding, which has been used since the 18th
century for reestablishing tree species on wet sites
(Londo and Mroz 2001). This technique has been
successfully applied in some Australian systems
and has been recommended for revegetation
because of its good soil moisture retention and
high plant survival (Close and Davidson 2002).
Unless intrinsic edaphic (e.g., salinity, pH) and
hydrological (e.g., water quality and water regime)
factors are fully understood, community-based
revegetation trials in coastal Melaleuca wetlands
are likely to be unsuccessful. The effectiveness of
community-based revegetation efforts needs to be
rigorously and quantitatively assessed, so that the
process is more accountable and transparent. The
mere act of planting large number of seedlings is
insufficient as a rehabilitation strategy if the young
plants do not mature into a self-sustaining popu-
lation. This is the result we observed in our
collaboration with community groups, as <10%
(and in some trials <1%) of tubestock seedlings
survived over the period of observation.
Acknowledgements We thank Parks Victoria forallowing us to conduct the experiments at Dowd Morassand for their support of the project. The Rangers, AndrewSchulz, Chris Holmes and Peter Kambouris, are thankedespecially. We are very grateful to John Topp of GippslandIndigenous Plants for growing the seedlings, and to theGippsland community for their assistance with planting,particularly Gary Howard and members of Field and
Game Australia (Sale and District), Field Naturalists andWaterwatch. We thank also Ni Luh Watiniasih, JacquiSalter and Matthew Hatton for their assistance in the field.This research was funded under Land and Water GrantsUTV2 and MU041, with additional support from WestGippsland Catchment Management Authority,Department of Sustainability and Environment,Department of Primary Industries, Parks Victoria andthe Gippsland Coastal Board.
References
Bedford BL (1996) The need to define hydrologicalequivalence at the landscape scale for freshwaterwetland mitigation. Ecol Appl 6:57–68
Bird ECF (1961) Reed growth in the Gippsland Lakes. VicNat 77:262–268
Bird ECF (1962) The Swamp paper-bark. Vic Nat 79:72–81
Bowkett LA, Kirkpatrick JB (2003) Ecology and conser-vation of remnant Melaleuca ericifolia stands in theTamar Valley, Tasmania. Aust J Bot 51:405–413
Close DC, Davidson NJ (2002) Revegetation to combattree decline in the midlands and Derwent Valleylowlands of Tasmania: practices for improved plantestablishment. Ecol Mgmt Rest 4:29–36
Costermans LF (1998) Native trees and shrubs of south-eastern Australia. Toppan Printing Company Pty.Ltd., Singapore
Davis J, Froend RH, Hamilton D et al (2001) Environ-mental water requirements to maintain wetlands ofnational and international importance. EnvironmentAustralia, Canberra
de Jong NH (2000) Woody plant restoration and naturalregeneration in wet meadow at Coomonderry swampon the south coast of New South Wales. MarFreshwater Res 51:81–89
de Jong TJ (1997) Register of wetland restoration projectsin Australia and New Zealand. Department ofEnvironment and Natural Resources, Adelaide
Denton M, Ganf GG (1994) Response of juvenile Melal-euca halmaturorum to flooding: management implica-tions for a seasonal wetland, Bool Lagoon, SouthAustralia. Aust J Mar Freshwater Res 45:1395–1408
Gibson N, Williams K, Marsden-Smedley J et al (1987)Regeneration characteristics of a swamp forest innorthwestern Tasmania. Proc Royal Soc Tasmania121:93–99
Hart BT, Lake PS, Webb JA et al (2003) Ecological risk toaquatic systems from salinity increases. Aust J Bot51:689–702
Ladd PG, Ladiges P, Calder DM (1976) Vegetationsurrounding the Gippsland Lakes and Lake Tyers:final report. Prepared for the Ministry for Conserva-tion, Victoria
Ladiges P, Foord PC, Willis RJ (1981) Salinity andwaterlogging tolerance of some populations of Melal-euca ericifolia Smith. Aust J Ecol 6:203–215
Wetlands Ecol Manage (2007) 15:175–188 187
123

Londo AJ, Mroz GD (2001) Bucket mounding as amechanical site preparation technique in wetlands.North J App For 18:7–13
Middleton B (1999) Wetland restoration, flood pulsing anddisturbance dynamics. Wiley, USA
Quinn G, Keough M (2002) Experimental design and dataanalysis for biologists. Cambridge University Press,Cambridge
Rayment GE, Higginson FR (1992) Australian laboratoryhandbook of soil and water chemical methods. InkataPress, Melbourne
Rothwell RL, Woodard PM, Rivard PG (1993) The effectof peatland drainage and planting position on thegrowth of White Spruce seedlings. North J Appl For10:154–160
Roy V, Bernier PY, Plamondon AP et al (1999) Effect ofdrainage and microtopography in forested wetlandson the microenvironment and growth of plantedBlack Spruce seedlings. Can J For Res 29:563–574
Salter J, Morris K, Bailey PCE et al (2006) Interactiveeffects of salinity and water depth on the performanceof Melaleuca ericifolia Smith (Swamp Paperbark)
seedlings. Aquatic Botany Available online 21 Octo-ber 2006
Sinclair Knight Merz (2001) Lake Wellington catchmentsalinity management plan wetlands monitoring pro-ject. Part A. Analysis and interpretation of wetlandmonitoring data. Department of Natural Resourcesand Environment, Victoria
Specht RL (1990) Forested wetlands in Australia. In: LugoAE, Brinson M, Brown S (eds) Forested wetlands.Ecosystem of the world. Elsevier, Amsterdam, 15:387–405
Tabachnick BG, Fidell LS (2001) Using multivariatestatistics. Allyn and Bacon, USA
Turner RE, Lewis III RR (1997) Hydrologic restoration ofcoastal wetlands. Wetl Ecol Manage 4:65–72
Underwood AJ (1997) Experiments in ecology: theirlogical design and interpretation using analysis ofvariance. University Press, Cambridge
van der Moezel PG, Pearce-Pinto GVN, Bell DT (1991)Screening for salt and waterlogging tolerance inEucalyptus and Melaleuca species. For Ecol Manage40:27–37
188 Wetlands Ecol Manage (2007) 15:175–188
123




















