
Quinolizidine-Based Variations and Antifungal Activity of Eight ...
17
Citation: Cely-Veloza, W.; Quiroga, D.; Coy-Barrera, E. Quinolizidine-Based Variations and Antifungal Activity of Eight Lupinus Species Grown under Greenhouse Conditions. Molecules 2022, 27, 305. https://doi.org/10.3390/ molecules27010305 Academic Editors: Asta Judžentien ˙ e and Kristina Ložien ˙ e Received: 25 November 2021 Accepted: 30 December 2021 Published: 4 January 2022 Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affil- iations. Copyright: © 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https:// creativecommons.org/licenses/by/ 4.0/). molecules Article Quinolizidine-Based Variations and Antifungal Activity of Eight Lupinus Species Grown under Greenhouse Conditions Willy Cely-Veloza *, Diego Quiroga and Ericsson Coy-Barrera * Bioorganic Chemistry Laboratory, Facultad de Ciencias Básicas y Aplicadas, Universidad Militar Nueva Granada, Cajica 250247, Colombia; [email protected] * Correspondence: [email protected] (W.C.-V.); [email protected] (E.C.-B.) Abstract: Fusarium oxysporum is an aggressive phytopathogen that affects various plant species, resulting in extensive local and global economic losses. Therefore, the search for competent alter- natives is a constant pursuit. Quinolizidine alkaloids (QA) are naturally occurring compounds with diverse biological activities. The structural diversity of quinolizidines is mainly contributed by species of the family Fabaceae, particularly the genus Lupinus. This quinolizidine-based chemo diversity can be explored to find antifungals and even mixtures to address concomitant effects on F. oxysporum. Thus, the antifungal activity of quinolizidine-rich extracts (QREs) from the leaves of eight greenhouse-propagated Lupinus species was evaluated to outline promising QA mixtures against F. oxysporum. Thirteen main compounds were identified and quantified using an external standard. Quantitative analysis revealed different contents per quinolizidine depending on the Lupinus plant, ranging from 0.003 to 32.8 mg/g fresh leaves. Bioautography showed that all extracts were active at the maximum concentration (5 μg/μL). They also exhibited >50% mycelium growth inhibition. All QREs were fungistatic except for the fungicidal QRE of L. polyphyllus Lindl. Angustifoline, matrine, 13α-hydroxylupanine, and 17-oxolupanine were ranked to act jointly against the phytopathogen. Our findings constitute reference information to better understand the antifungal activity of naturally afforded QA mixtures from these globally important plants. Keywords: Fabaceae; Lupinus; quinolizidines; alkaloids; Fusarium oxysporum; antifungals 1. Introduction Fabaceae is the second largest family of flowering plants, containing approximately 490 species that have been used for medicinal purposes [1]. These plants are distributed in more than 65 genera of Fabaceae and can produce specialized metabolites such as quinolizidine alkaloid (QAs) (specially Genisteae Tribe) and non-nitrogenous compounds [2,3]. One of these genera is Lupinus, which is distributed in the Mediterranean, North and East Africa, and North and South America, and hosts the largest number of species [4]. About 170 Lupinus species can be found worldwide, including wild and domestic plants, widely called lupins [5]. In Colombia, 47 Lupinus species have been reported in the central and eastern mountain ranges in the Colombian Massif and Sierra Nevada de Santa Marta [6]. Lupinus-based products, particularly lupin seed proteins, are becoming good alternatives for multiple foods, including beverages, bakery, imitation meat, and dairy products, which have promoted intensive lupin cultivation in several parts of the world to produce grain protein [7]. However, the presence of QAs in lupin seed-based products provides a bitter taste. Therefore, debittering procedures should be performed on lupin seeds containing high QA contents for human and animal consumption [8]. Lupinus is one of the genera of Fabaceae that shows more considerable variability in QAs, which are biosynthesized by the L-lysine pathway in aerial green tissues, particularly leaves, and translocated to all plant parts whose accumulation is mainly in seeds (4– 8% dry weight) [9]. Above 170 QAs have been detected, identified, and quantified in Molecules 2022, 27, 305. https://doi.org/10.3390/molecules27010305 https://www.mdpi.com/journal/molecules
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Quinolizidine-Based Variations and Antifungal Activity of Eight ...

�����������������
Citation: Cely-Veloza, W.; Quiroga,
D.; Coy-Barrera, E.
Quinolizidine-Based Variations and
Antifungal Activity of Eight Lupinus
Species Grown under Greenhouse
Conditions. Molecules 2022, 27, 305.
https://doi.org/10.3390/
molecules27010305
Academic Editors: Asta Judžentiene
and Kristina Ložiene
Received: 25 November 2021
Accepted: 30 December 2021
Published: 4 January 2022
Publisher’s Note: MDPI stays neutral
with regard to jurisdictional claims in
published maps and institutional affil-
iations.
Copyright: © 2022 by the authors.
Licensee MDPI, Basel, Switzerland.
This article is an open access article
distributed under the terms and
conditions of the Creative Commons
Attribution (CC BY) license (https://
creativecommons.org/licenses/by/
4.0/).
molecules
Article
Quinolizidine-Based Variations and Antifungal Activity ofEight Lupinus Species Grown under Greenhouse ConditionsWilly Cely-Veloza *, Diego Quiroga and Ericsson Coy-Barrera *
Bioorganic Chemistry Laboratory, Facultad de Ciencias Básicas y Aplicadas, Universidad Militar Nueva Granada,Cajica 250247, Colombia; [email protected]* Correspondence: [email protected] (W.C.-V.); [email protected] (E.C.-B.)
Abstract: Fusarium oxysporum is an aggressive phytopathogen that affects various plant species,resulting in extensive local and global economic losses. Therefore, the search for competent alter-natives is a constant pursuit. Quinolizidine alkaloids (QA) are naturally occurring compoundswith diverse biological activities. The structural diversity of quinolizidines is mainly contributedby species of the family Fabaceae, particularly the genus Lupinus. This quinolizidine-based chemodiversity can be explored to find antifungals and even mixtures to address concomitant effects on F.oxysporum. Thus, the antifungal activity of quinolizidine-rich extracts (QREs) from the leaves of eightgreenhouse-propagated Lupinus species was evaluated to outline promising QA mixtures againstF. oxysporum. Thirteen main compounds were identified and quantified using an external standard.Quantitative analysis revealed different contents per quinolizidine depending on the Lupinus plant,ranging from 0.003 to 32.8 mg/g fresh leaves. Bioautography showed that all extracts were active atthe maximum concentration (5 µg/µL). They also exhibited >50% mycelium growth inhibition. AllQREs were fungistatic except for the fungicidal QRE of L. polyphyllus Lindl. Angustifoline, matrine,13α-hydroxylupanine, and 17-oxolupanine were ranked to act jointly against the phytopathogen.Our findings constitute reference information to better understand the antifungal activity of naturallyafforded QA mixtures from these globally important plants.
Keywords: Fabaceae; Lupinus; quinolizidines; alkaloids; Fusarium oxysporum; antifungals
1. Introduction
Fabaceae is the second largest family of flowering plants, containing approximately 490species that have been used for medicinal purposes [1]. These plants are distributed in morethan 65 genera of Fabaceae and can produce specialized metabolites such as quinolizidinealkaloid (QAs) (specially Genisteae Tribe) and non-nitrogenous compounds [2,3]. One ofthese genera is Lupinus, which is distributed in the Mediterranean, North and East Africa,and North and South America, and hosts the largest number of species [4]. About 170Lupinus species can be found worldwide, including wild and domestic plants, widelycalled lupins [5]. In Colombia, 47 Lupinus species have been reported in the central andeastern mountain ranges in the Colombian Massif and Sierra Nevada de Santa Marta [6].Lupinus-based products, particularly lupin seed proteins, are becoming good alternativesfor multiple foods, including beverages, bakery, imitation meat, and dairy products, whichhave promoted intensive lupin cultivation in several parts of the world to produce grainprotein [7]. However, the presence of QAs in lupin seed-based products provides a bittertaste. Therefore, debittering procedures should be performed on lupin seeds containinghigh QA contents for human and animal consumption [8].
Lupinus is one of the genera of Fabaceae that shows more considerable variability inQAs, which are biosynthesized by the L-lysine pathway in aerial green tissues, particularlyleaves, and translocated to all plant parts whose accumulation is mainly in seeds (4–8% dry weight) [9]. Above 170 QAs have been detected, identified, and quantified in
Molecules 2022, 27, 305. https://doi.org/10.3390/molecules27010305 https://www.mdpi.com/journal/molecules

Molecules 2022, 27, 305 2 of 17
different Lupinus species from which bicyclic-, tricyclic-, and tetracyclic-type QAs canbe obtained [10–13] and whose structural pattern is highly variable depending on thespecies [14]. The QA biosynthesis is highly regulated, influenced by genetic (e.g., genotype),biotic (e.g., presence of phytophagous insects and/or microbial pathogens), and abiotic(e.g., environmental and soil conditions) factors [13,14].
Lupin QAs constitute a natural defensive chemical barrier facing biotic pressures [15].Thus, despite their well-known acute anticholinergic toxicity, they can be explored asalternatives (or even compound templates) against pests and pathogens. In this regard,some QAs isolated from Lupinus, such as sparteine, lupanine, and lupinine, have beenused to control intestinal ectoparasites in cattle [16], and farmers occasionally use a QA-rich infusion from seeds for pest control in plants [17]. In addition, QA-rich Lupinusextracts (QREs) have shown antimicrobial activity against different microorganisms. Forinstance, the extracts of L. mutabilis seeds have inhibited 95% of the growth of Staphylococcusaureus [18], while Candida fungi have shown >60% inhibition in the presence of QREs of L.arboreus leaves [19], L. albescens [20], L. angustifolius [21], and L. densiflorus [22]. The seed-derived QREs from L. exaltatus and L. mexicanus have also been reported, showing inhibition>72% against Sclerotium rolfsii, Alternaria solani, Rhizoctonia solani, and F. oxysporum [23]. Inthe case of phytopathogens of the genus Fusarium, F. oxysporum and F. verticillioides showedsusceptibility to QREs of L. albescens, with IC50 values < 30 µg/mL [20].
The current phytosanitary problems associated with the attack of F. oxysporum remainto be solved. Crops infected with this phytopathogen experience fusariosis, a plant diseasethat causes damage from leaf chlorosis to vascular necrosis [24]. These adverse effectsproduce significant economic losses in the agricultural sector [25], specifically in cropssuch as cape gooseberrys [26], bananas [27], carnations [28], lulos, and strawberries [29].Fungicides such as dithane-M45, manzate 200WP, fitoraz, rovral-Flo, folicur EW, strobySC, azoxystrobin, benomyl, carbendazim, and fludioxonil are currently used for the man-agement of fusariosis [30,31]. However, the excessive use of these products led to adverseeffects such as the production of resistant strains that seriously affect some important crops,such as bananas [32].
Similarly, other problems related to the inappropriate use of fungicides have affectedthe environment, microbiota, and soil fertility due to non-biodegradable, persistent, andbioaccumulative chemicals [33,34]. Therefore, there is a continual need to search for compe-tent alternatives to synthetic pesticides to control this phytopathogen that can eventuallybe incorporated into rigorously controlled integrated pest management (IPM) programs.The exploration to find antifungals might be advantageous from plant sources with favor-able propagation, such as species of the genus Lupinus. In addition, since the particularproduction mainly contributes to the structural diversity in Lupinus leaves and consideringthat QAs are biologically active specialized metabolites and plant defense products [2],naturally-supplied QA mixtures could be examined as a promising approach for exertingconcomitant effects on F. oxysporum.
Therefore, the present study evaluated the antifungal activity against F. oxysporum ofQREs obtained from eight Lupinus species cultivated under greenhouse conditions. Thus,the quinolizidine variations in the test Lupinus species were employed to recognize patternsand identify the most active QAs and their mixtures as plausible antifungals from foliarmaterials of cultivated lupin plants.
2. Results and Discussion2.1. Characteristics of Propagated Plants
The leaf collection stage of native species was defined when plants showed their firstflower buds and reached ca. 50 cm in height. Only average heights around 20 cm werereached in exotic species, except L. albus specimens that exceeded 50 cm in height. In thecase of L. polyphyllus and L. perennis, no floral buds were observed. The flowering could beaffected by the unusual temperature conditions of the greenhouse (mainly cold periods

Molecules 2022, 27, 305 3 of 17
and irradiation) for these exotic plants, since vernalization and photoperiods have beendescribed to affect the flowering periods of some lupins seriously, e.g., L. luteus [35].
Additionally, high temperatures can alter the phenological stages of Lupinus [36] insuch a way that lower production of flowers and seeds is observed, while in more extremecases, the flowering stage is not reached [37]. However, exotic lupin plants grew underthe employed propagation conditions, indicating good acclimatization of Lupinus speciesto such a growing environment, which is beneficial when mass cultivating for harvestingpurposes. Thus, all lupin plants reached satisfactory growth to collect enough leaf biomassfor the subsequent chemical analysis.
2.2. Chemical Analysis
The young leaves were collected once the lupin plants reached physiological maturity(i.e., 70 ± 19 days after transplantation). Hence, QREs from fresh leaves (5 g) of the eightLupinus species were prepared to afford the yields shown in Table S1 (supplementarymaterial), following an acid-base extraction method. The highest yields were achievedfor L. bogotensis, L. mirabilis, and L. mutabilis (yields > 5%). Contrarily, the exotic speciesyielded lower extract amounts, but the best yields between them resulted for L. polyphyllus,L. perennis, and L. albus (yields > 4%), while L. argenteus and L. arboreus showed the lowestyields (<3%). The yield differences of native lupine plants compared to exotic ones can beexplained by biomass and specialized metabolite variations due to those intrinsic (genetic)factors favored by extrinsic (environmental) factors related to the propagation conditionsmanaged in the present study [38].
GC-MS analysis subsequently recorded the QA composition of these QREs obtainedfrom the eight Lupinus plants. With the diagnostic analysis of the mass spectra combinedwith the retention indices compared to the available information in literature, QAs 1–13,which are listed in Table 1 and whose structures are shown in Figure S1, were identified.
Table 1. Identified quinolizidines (1–13) in eight lupin species propagated under greenhouse condi-tions.
# a Rt b (min) Name RI c RI d Reference d
1 20.5 sparteine 1785 1805 [39]2 21.6 11,12-dehydrosparteine 1841 1841 [40]3 24.4 N-methylcytisine 1918 1924 [41]4 25.8 angustifoline 2079 2073 [39]5 26.3 5,6-dehydrolupanine 2104 2092 [40]6 27.1 α-isolupanine 2107 2123 [39]7 27.7 lupanine 2170 2146 [42]8 29.0 nuttalline 2338 2348 [18]9 30.9 matrine 2366 2365 [43]
10 31.1 anagyrine 2390 2377 [40]11 31.4 13α-hydroxylupanine 2405 2400 [39]12 35.7 multiflorine 2440 2469 [18]13 36.0 17-oxolupanine 2482 2473 [18]
a Compound numbers according to the chromatographic elution presented in Figure 1; b Rt = retention time (min);c Calculated retention index (RI) according to the Equation (S1); and d Reported RI according to the respectivereference.
The identified QAs have already been reported as phytocomponents in Lupinus species.Some of them are commonly found as the most abundant QAs, being part of certainmixtures at differentiated proportions between species or even due to the effects causedby biotic and abiotic factors [13,44]. Qualitative and quantitative comparisons were madebetween the eight lupin species propagated under the same conditions in the present study.Accordingly, the stacked chromatographic profiles are shown in Figure 1 and the measuredcontents per QA of these test plants, expressed as mg/g fresh leaves (fl), are presented inTable 2. Some compound signals (i.e., 6) are not entirely visualized in Figure 1 due to therelative abundance, despite being quantified according to the limit of quantification (see

Molecules 2022, 27, 305 4 of 17
Section 3.3), since the most abundant compounds per GC-MS chromatogram reduced thevisible intensity of those signals related to the minor compounds.
Molecules 2022, 27, x FOR PEER REVIEW 4 of 18
study. Accordingly, the stacked chromatographic profiles are shown in Figure 1 and the measured contents per QA of these test plants, expressed as mg/g fresh leaves (fl), are presented in Table 2. Some compound signals (i.e., 6) are not entirely visualized in Figure 1 due to the relative abundance, despite being quantified according to the limit of quanti-fication (see Section 3.3), since the most abundant compounds per GC-MS chromatogram reduced the visible intensity of those signals related to the minor compounds.
Figure 1. Stacked GC-MS chromatograms of quinolizidine-rich extracts (QREs) were obtained from the eight species of Lupinus. The box-enclosed numbers represent each identified quinolizidine listed in Table 1.
The QRE profiles changed in the presence and abundance of QAs, whose contents varied within a 0.031–32.8 mg/g fl range. Some common QAs were found in all test spe-cies, as in the case of 6 and 7, with contents between 0.046–3.12 and 5.62–25.7 mg/g fl, respectively. In contrast, other QAs were not commonly found in the sample set, such as 9 (0.043–3.54 mg/g fl), as most reported in the genus Sophora [43]. In the case of the species L. argenteus and L. arboreus, QAs containing the 2-pyridone moiety, such as 3 and 10, were the most common compounds, comprising contents between 4.61 and 6.08 mg/g fl. A pre-vious study reported that L. argenteus ecotypes exhibited the presence of seventeen QAs, including lupanine derivatives such as 5,6-dehydrolupanine, α-isolupanine, 11,12-dehy-drolupanine, 3-hydroxylupanine, and thermopsine [45], but the abundance and presence of lupanine derivatives were found to be different in L. argenteus under study. QAs 7 and 11 have been previously reported for L. arboreus, but 11 mainly accumulates in L. arboreus seeds; therefore, it was not detected in this study [46]. In the case of L. albus, QAs 4, 7, 8, and 12 were identified and quantified (between 0.91 and 13.8 mg/g fl). However, the pres-ence of albine was reported in previous studies [47], but it was not detected in the L. albus-derived extracts under investigation. Finally, in native species such as L. mutabilis and L. bogotensis, compounds 1, 2, 4–8, and 12 have previously been reported [8,48], agreeing with the results of the present study involving the highly-varied contents of such QAs (between 0.033 and 32.8 mg/g fl). The presence of 1, 7, 9, and 10 has been generally re-ported in the genera Lupinus, Sophora, and Genista species.
Figure 1. Stacked GC-MS chromatograms of quinolizidine-rich extracts (QREs) were obtained fromthe eight species of Lupinus. The box-enclosed numbers represent each identified quinolizidine listedin Table 1.
The QRE profiles changed in the presence and abundance of QAs, whose contentsvaried within a 0.031–32.8 mg/g fl range. Some common QAs were found in all testspecies, as in the case of 6 and 7, with contents between 0.046–3.12 and 5.62–25.7 mg/g fl,respectively. In contrast, other QAs were not commonly found in the sample set, such as 9(0.043–3.54 mg/g fl), as most reported in the genus Sophora [43]. In the case of the species L.argenteus and L. arboreus, QAs containing the 2-pyridone moiety, such as 3 and 10, were themost common compounds, comprising contents between 4.61 and 6.08 mg/g fl. A previousstudy reported that L. argenteus ecotypes exhibited the presence of seventeen QAs, includinglupanine derivatives such as 5,6-dehydrolupanine, α-isolupanine, 11,12-dehydrolupanine,3-hydroxylupanine, and thermopsine [45], but the abundance and presence of lupaninederivatives were found to be different in L. argenteus under study. QAs 7 and 11 havebeen previously reported for L. arboreus, but 11 mainly accumulates in L. arboreus seeds;therefore, it was not detected in this study [46]. In the case of L. albus, QAs 4, 7, 8, and 12were identified and quantified (between 0.91 and 13.8 mg/g fl). However, the presence ofalbine was reported in previous studies [47], but it was not detected in the L. albus-derivedextracts under investigation. Finally, in native species such as L. mutabilis and L. bogotensis,compounds 1, 2, 4–8, and 12 have previously been reported [8,48], agreeing with the resultsof the present study involving the highly-varied contents of such QAs (between 0.033 and32.8 mg/g fl). The presence of 1, 7, 9, and 10 has been generally reported in the generaLupinus, Sophora, and Genista species.

Molecules 2022, 27, 305 5 of 17
Table 2. Contents of quinolizidines 1–13 in leaves of eight Lupinus species.
PlantsQA Content (mg LE/g FL) a
1 2 3 4 5 6 7
L.po. 1.43 ± 0.12 C n.d. n.d. 5.39 ± 0.18 A 0.391 ± 0.008 B 0.169 ± 0.009 E 17.1 ± 0.9 C
L.pe. n.d. 0.031 ± 0.001 D n.d. 3.12 ± 0.12 B n.d. 1.607 ± 0.032 B 20.0 ± 0.2 B
L.b. 22.4 ± 0.7 B 0.033 ± 0.002 D n.d. n.d. n.d. 0.083 ± 0.002 F 19.3 ± 0.8 B
L.mu. n.d. n.d. n.d. 0.87 ± 0.09 C 1.42 ± 0.16 A 0.452 ± 0.009 C 25.7 ± 0.6 A
L.mi. 32.8 ± 0.6 A 0.102 ± 0.004 C n.d. 0.102 ± 0.008 D n.d. 0.151 ± 0.006 E 20.1 ± 0.31 B
L.arg. n.d. 1.38 ± 0.19 B 4.98 ± 0.19 B 0.051 ± 0.003 E n.d. 0.201 ± 0.016 D 5.62 ± 0.14 F
L.arb n.d. 2.07 ± 0.16 A 5.69 ± 0.21 A 0.046 ± 0.002 E n.d. 0.046 ± 0.003 G 6.56 ± 0.20 G
L.al. n.d. n.d. n.d. 3.35 ± 0.14 B n.d. 3.12 ± 0.06 A 13.8 ± 0.9 D
PlantsQA Content (mg LE/g FL) a
8 9 10 11 12 13
L.po. 1.08 ± 0.09 C 3.54 ± 0.08 A n.d. 2.62 ± 0.09 A 0.328 ± 0.013 C 9.40 ± 0.75 A
L.pe. 0.735 ± 0.059 D 0.043 ± 0.003 C 0.048 ± 0.001 C 0.068 ± 0.002 D 0.154 ± 0.005 E 4.09 ± 0.11 C
L.b. 5.41 ± 0.16 B n.d. n.d. n.d. 0.209 ± 0.017 D 0.051 ± 0.005 F
L.mu. n.d. 0.729 ± 0.008 B n.d. 0.727 ± 0.011 B 0.812 ± 0.012 B 4.98 ± 0.23 B
L.mi. 0.176 ± 0.011 E n.d. n.d. n.d. 0.045 ± 0.002 F 0.063 ± 0.006 F
L.arg. n.d. n.d. 6.08 ± 0.07 A n.d. 4.88 ± 0.34 A n.d.L.arb n.d. n.d. 4.61 ± 0.13 B n.d. n.d. 0.104 ± 0.008 E
L.al. 6.15 ± 0.08 A n.d. n.d. 0.377 ± 0.011 C 0.910 ± 0.017 B 4.97 ± 0.21 B
a Contents expressed as mg of lupanine equivalents per gram of fresh leaves (mg LE/g FL); the presented valuescomprise means ± standard deviation (n = 10). L.po. = L. polyphyllus, L.pe. = L. perennis, L.b. = L. bogotensis, L.mu.= L. mutabilis, L.mi. = L. mirabilis, L.arg = L. argenteus, L.arb. = L. arboreus, L.al. = L. albus. Different uppercasecapital letters indicate statistically significant differences according to the post hoc Tukey test (p < 0.05). n.d. = notdetected.
All Lupinus species were characterized by the high production and accumulation oflupanine (7), exploited as a useful chemotaxonomic marker to recognize Lupinus species [2].In native species, close lupanine contents were found, although L. mutabilis exhibited thehighest and significantly different content. In the case of exotic species, some differenceswere found and could be gathered into two groups as follows: In the first group, L. argenteusand L. arboreus were characterized by the low content of 7 (<7 mg/g fl) and the presence ofQAs 3 and 10, which are not commonly reported in Lupinus species but highly occurredin Genista species [49,50]. In the second group, the species L. polyphyllus, L. albus, and L.perennis showed important contents of 7 but the highest abundance of 4 (5.39, 3.35, and3.12 mg/g fl, respectively) and other characteristic compounds of these species such as 8and 11 (within content ranges of 0.735–6.15 and 0.068–2.62 mg/g fl, respectively).
2.3. Direct Bioautography Assay
Bioautography allowed qualitatively observing the sensitivity of the phytopathogenspores to the test QREs (Figure 2A). In this way, the negative control (D3, distilled wa-ter) exhibited a completely dark surface, representing the phytopathogen growth in thatsquare [51]. In contrast, positive controls Dithane and Rovral (squares B1 and B3) revealedclear zones, indicating no fungal growth due to the inhibition of spore germination [52].Similarly, QRE-based treatments (squares A1–A3, B2, C1–C3, and D1–D2) produced similarinhibition halos to positive controls. Thus, although some distinctive intensities were evi-denced, these results qualitatively indicated that all test extracts exhibit antifungal activityon F. oxysporum spores.

Molecules 2022, 27, 305 6 of 17Molecules 2022, 27, x FOR PEER REVIEW 6 of 18
Figure 2. Antifungal activity against F. oxysporum QREs from eight lupin species. (A) Bioautography results using 50 µg of QREs; (B) Mycelial growth inhibition percentages of eight QREs at three con-centrations.
2.4. Mycelial Growth Inhibition Assay The antifungal activity evaluation of the Lupinus-derived QREs against F. oxysporum
was quantitatively extended by assessing the mycelium growth inhibition through the amended-medium (poisoned food) technique. Thus, QREs were evaluated at three doses (i.e., 5.0, 1.0, and 0.1 µg/µL) and, subsequently, the respective inhibition percentages were measured, obtaining values >50% at the maximum concentration (Figure 2B). The L. poly-phyllus-derived QRE showed the best antifungal activity at the three test concentrations (inhibition > 85%), and such inhibitions were very similar to those of positive controls. However, the observed antifungal activity variations depended on the treatment and QAs composition involved significant differences (p < 0.05). Thus, using the post hoc Tukey test, the statistically significant differences based on variations in composition and con-centration were obtained. Consequently, twenty different mean groups were achieved (Figure S2, supplementary material). The first significantly different groups (i.e., A–F) were related to the best inhibition values (>80%), comprising positive controls and extracts from L. polyphyllus, L. bogotensis, L. mirabilis, and L. mutabilis at the highest test concentra-tions (1 and 5 µg/µL). The other groups represented a lesser inhibition (<75%) showed by the other extracts, mainly dependent on the concentration of each QRE. The measured inhibition values agree with the values reported for QREs obtained from L. exaltatus [53]. L. argenteus, L. arboreus, L. albus, and L. perennis exhibited the lowest inhibitory activity on mycelial growth (inhibition between 20 and 60%, depending on the concentration). How-ever, L. albus and L. perennis exhibited clear zones of inhibition in the bioautography, even similar to other QREs treatments (Figure 2A), suggesting a plausible selective effect de-pending on the fungal structure (spore > mycelium), which might be clarified in further studies.
The test QREs showed differences in their chemical composition and antifungal ac-tivity against F. oxysporum. In general, the eight lupin species showed the best antifungal activity at the maximum test concentration (5 µg/µL), comprising inhibition percentages between 50–95%. As mentioned above, the most active QREs were those obtained from L. polyphyllus and L. bogotensis. These QREs showed dose-dependent differences, as shown in Figure 3A. For example, C1 (treatment at 5 µg/µL) highly inhibited the mycelial growth of the phytopathogen (inhibition > 95%), while C2 (treatment at 1 µg/µL) showed lower mycelial growth (inhibition > 80%). In the case of C3 (treatment at 0.1 µg/µL), moderate mycelial growth was observed (inhibition > 70%). On the other hand, the intermediate doses (C2) of the QREs obtained from L. polyphyllus, L. bogotensis, and L. mirabilis showed the best results (inhibition > 80%), while a significant inhibitory reduction was found for the other five species (inhibition < 60%). Finally, the least active concentration was C3. At this dose, L. polyphyllus, L. bogotensis, and L. mirabilis afforded the best results (inhibition > 70%), while QREs from the other species were found to be below 60%. In this sense,
Figure 2. Antifungal activity against F. oxysporum QREs from eight lupin species. (A) Bioautographyresults using 50 µg of QREs; (B) Mycelial growth inhibition percentages of eight QREs at threeconcentrations.
2.4. Mycelial Growth Inhibition Assay
The antifungal activity evaluation of the Lupinus-derived QREs against F. oxysporumwas quantitatively extended by assessing the mycelium growth inhibition through theamended-medium (poisoned food) technique. Thus, QREs were evaluated at three doses(i.e., 5.0, 1.0, and 0.1 µg/µL) and, subsequently, the respective inhibition percentageswere measured, obtaining values >50% at the maximum concentration (Figure 2B). The L.polyphyllus-derived QRE showed the best antifungal activity at the three test concentrations(inhibition > 85%), and such inhibitions were very similar to those of positive controls.However, the observed antifungal activity variations depended on the treatment and QAscomposition involved significant differences (p < 0.05). Thus, using the post hoc Tukey test,the statistically significant differences based on variations in composition and concentrationwere obtained. Consequently, twenty different mean groups were achieved (Figure S2,supplementary material). The first significantly different groups (i.e., A–F) were relatedto the best inhibition values (>80%), comprising positive controls and extracts from L.polyphyllus, L. bogotensis, L. mirabilis, and L. mutabilis at the highest test concentrations (1and 5 µg/µL). The other groups represented a lesser inhibition (<75%) showed by the otherextracts, mainly dependent on the concentration of each QRE. The measured inhibitionvalues agree with the values reported for QREs obtained from L. exaltatus [53]. L. argenteus,L. arboreus, L. albus, and L. perennis exhibited the lowest inhibitory activity on mycelialgrowth (inhibition between 20 and 60%, depending on the concentration). However, L.albus and L. perennis exhibited clear zones of inhibition in the bioautography, even similarto other QREs treatments (Figure 2A), suggesting a plausible selective effect depending onthe fungal structure (spore > mycelium), which might be clarified in further studies.
The test QREs showed differences in their chemical composition and antifungal ac-tivity against F. oxysporum. In general, the eight lupin species showed the best antifungalactivity at the maximum test concentration (5 µg/µL), comprising inhibition percentagesbetween 50–95%. As mentioned above, the most active QREs were those obtained from L.polyphyllus and L. bogotensis. These QREs showed dose-dependent differences, as shown inFigure 3A. For example, C1 (treatment at 5 µg/µL) highly inhibited the mycelial growthof the phytopathogen (inhibition > 95%), while C2 (treatment at 1 µg/µL) showed lowermycelial growth (inhibition > 80%). In the case of C3 (treatment at 0.1 µg/µL), moderatemycelial growth was observed (inhibition > 70%). On the other hand, the intermediatedoses (C2) of the QREs obtained from L. polyphyllus, L. bogotensis, and L. mirabilis showedthe best results (inhibition > 80%), while a significant inhibitory reduction was found forthe other five species (inhibition < 60%). Finally, the least active concentration was C3. Atthis dose, L. polyphyllus, L. bogotensis, and L. mirabilis afforded the best results (inhibition >70%), while QREs from the other species were found to be below 60%. In this sense, these

Molecules 2022, 27, 305 7 of 17
results suggest that the optimal threshold for an effective dose to inhibit the growth of F.oxysporum mycelium by these QREs is 5 µg/µL (0.5% w/v).
Molecules 2022, 27, x FOR PEER REVIEW 7 of 18
these results suggest that the optimal threshold for an effective dose to inhibit the growth of F. oxysporum mycelium by these QREs is 5 µg/µL (0.5% w/v).
Figure 3. (A) Antifungal activity (mycelial growth inhibition) evaluated for the most-active QREs from L. polyphyllus (left) and L. bogotensis (right) against F. oxysporum. (B) (PDA 0.5%), C1 (5 µg/µL), C2 (1 µg/µL), and C3 (0.1 µg/µL). (B) Fungicide vs. Fungistatic of the most-active QREs of L. poly-phyllus and L. bogotensis.
Once the mycelial growth inhibition assay time elapsed, a subsequent test was per-formed to classify the extracts as fungicidal or fungistatic. This test was accomplished by further monitoring the fungal growth in fresh PDA after completion of QRE treatment at 5 µg/µL. This test suggested that seven lupin-derived extracts were fungistatic (Figure 3B). In contrast, there was no mycelium growth for the L. polyphyllus-derived QRE, indi-cating a fungicidal behavior. In addition, there are some studies related to QAs and bisquinolizidine alkaloids, such as spirocytisine, 3,5-dibromocytisine, bromo-N-boccytis-ine, N-boccytisine, 3-bromobenzyl cytisine, 4-bromo-benzylcytisine, 3-iodobenzylcytis-ine, and 4-iodobenzylcytisine, evaluated against Aspergillus niger van Tieghem ATCC 6275, and all of them showed fungistatic activity [54].
The detailed scrutiny of the chemical and antifungal activity data integration of test QREs led us to recognize meaningful patterns. Initially, intuitive visualization of the com-parison of autoscaled QA contents revealed plausible contributions by some QAs to the antifungal activity (Figure 4A). In this regard, QREs containing particular combinations of specific QAs were generally the most active extracts. Thus, the most active QRE, de-rived from L. polyphyllus, exhibited the abundant presence of 4, 9, 11, whereas QREs of L. bogotensis (the second most active extract against F. oxysporum) contained 1 and 8 in high abundance and L. mirabilis exhibited the highest content of 1. Remarkably, compound 9 was only detected in the most active QREs from L. mutabilis and L. polyphyllus, whose interesting inhibitory properties on conidia germination of F. oxysporum, Sphaeropsis spainea, Valsa pini (IC50 < 600 µg/mL) were previously evaluated [55]. The other lupin spe-cies that did not contain 1, 4, 9, and/or 11, showed the lowest activity, with inhibition < 70% in the three treatments. This in-depth exploration was extended by an unpaired fold-change (FC) analysis performed on QA content data of the test QREs, involving two anti-fungal activity-related datasets, i.e., more active QREs with >80% inhibition and less active QREs with <75% inhibition. The resulting plot (Figure 4B) showed the important QAs se-lected by this FC analysis (threshold = 2), and the red and blue dots represent QAs above and below such a threshold, respectively. This logarithmic relative change between the two activity-related datasets can be statistically interpreted as a positive and negative con-tribution to the antifungal activity by a QA due to its particular presence in a more active or less active QRE, respectively. Thus, QAs 2, 3, 6, 10, and 12 contributed negatively to the antifungal activity, whereas 5, 7, and 8 exhibited no contribution since the FC threshold was not exceeded. In contrast, the significance of 1, 4, 9, 11, and 13 was statistically evi-denced by their positive contribution to the antifungal activity. They can be considered essential constituents of a bioactive QRE and a plausible, promising mixture against F. oxysporum. The particular combination of QAs in the L. polyphyllus-derived QRE might
Figure 3. (A) Antifungal activity (mycelial growth inhibition) evaluated for the most-active QREsfrom L. polyphyllus (left) and L. bogotensis (right) against F. oxysporum. (B) (PDA 0.5%), C1 (5 µg/µL),C2 (1 µg/µL), and C3 (0.1 µg/µL). (B) Fungicide vs. Fungistatic of the most-active QREs of L.polyphyllus and L. bogotensis.
Once the mycelial growth inhibition assay time elapsed, a subsequent test was per-formed to classify the extracts as fungicidal or fungistatic. This test was accomplishedby further monitoring the fungal growth in fresh PDA after completion of QRE treat-ment at 5 µg/µL. This test suggested that seven lupin-derived extracts were fungistatic(Figure 3B). In contrast, there was no mycelium growth for the L. polyphyllus-derived QRE,indicating a fungicidal behavior. In addition, there are some studies related to QAs andbisquinolizidine alkaloids, such as spirocytisine, 3,5-dibromocytisine, bromo-N-boccytisine,N-boccytisine, 3-bromobenzyl cytisine, 4-bromo-benzylcytisine, 3-iodobenzylcytisine, and4-iodobenzylcytisine, evaluated against Aspergillus niger van Tieghem ATCC 6275, and allof them showed fungistatic activity [54].
The detailed scrutiny of the chemical and antifungal activity data integration of testQREs led us to recognize meaningful patterns. Initially, intuitive visualization of thecomparison of autoscaled QA contents revealed plausible contributions by some QAs tothe antifungal activity (Figure 4A). In this regard, QREs containing particular combinationsof specific QAs were generally the most active extracts. Thus, the most active QRE, derivedfrom L. polyphyllus, exhibited the abundant presence of 4, 9, 11, whereas QREs of L. bogotensis(the second most active extract against F. oxysporum) contained 1 and 8 in high abundanceand L. mirabilis exhibited the highest content of 1. Remarkably, compound 9 was onlydetected in the most active QREs from L. mutabilis and L. polyphyllus, whose interestinginhibitory properties on conidia germination of F. oxysporum, Sphaeropsis spainea, Valsapini (IC50 < 600 µg/mL) were previously evaluated [55]. The other lupin species that didnot contain 1, 4, 9, and/or 11, showed the lowest activity, with inhibition < 70% in thethree treatments. This in-depth exploration was extended by an unpaired fold-change(FC) analysis performed on QA content data of the test QREs, involving two antifungalactivity-related datasets, i.e., more active QREs with >80% inhibition and less active QREswith <75% inhibition. The resulting plot (Figure 4B) showed the important QAs selected bythis FC analysis (threshold = 2), and the red and blue dots represent QAs above and belowsuch a threshold, respectively. This logarithmic relative change between the two activity-related datasets can be statistically interpreted as a positive and negative contribution tothe antifungal activity by a QA due to its particular presence in a more active or less activeQRE, respectively. Thus, QAs 2, 3, 6, 10, and 12 contributed negatively to the antifungalactivity, whereas 5, 7, and 8 exhibited no contribution since the FC threshold was notexceeded. In contrast, the significance of 1, 4, 9, 11, and 13 was statistically evidencedby their positive contribution to the antifungal activity. They can be considered essentialconstituents of a bioactive QRE and a plausible, promising mixture against F. oxysporum.The particular combination of QAs in the L. polyphyllus-derived QRE might rationalize its

Molecules 2022, 27, 305 8 of 17
observed fungicidal activity. In this regard, such a QA-based combination could be dueto the growing behavior at the test greenhouse conditions since this introduced plant didnot reach the flowering stage. This fact may be a critical factor influencing the compositionand, consequently, the antifungal activity of this most active lupin plant.
Molecules 2022, 27, x FOR PEER REVIEW 8 of 18
rationalize its observed fungicidal activity. In this regard, such a QA-based combination could be due to the growing behavior at the test greenhouse conditions since this intro-duced plant did not reach the flowering stage. This fact may be a critical factor influencing the composition and, consequently, the antifungal activity of this most active lupin plant.
Figure 4. Comparison of QA contents (mg/g fresh leaves) among QREs obtained from leaves of eight lupin species. (A) Heatmap visualization of the normalized content distribution (unit of variance scaling) of QAs 1–13 from test lupins. Autoscaled QA contents are related to the color scale: 2 = high content; −2 = low content. (B) Unpaired fold-change (FC) analysis plot for selecting important QAs with threshold 2, by integrating the chemical (QA contents) and biological (mycelial growth inhibi-tion) data. Antifungal activity was used as a categorical variable, subdividing the antifungal activity of QREs into two datasets: more active (>80% inhibition) and less active (<75% inhibition), according to the Tukey test, to supervise the QA selection. According to the antifungal activity, positive and negative contributions to the FC-based QA selection are depicted as red and blue numbers and dots.
On the other hand, this integrated examination also suggests that very high contents of 7 are not a determinant for antifungal activity against F. oxysporum as the five selected QAs. This observation agrees with a previous study on a QRE obtained from L. exaltatus seeds. Such an extract did not show antifungal activity against F. oxysporum and its QA composition was based on a high relative abundance of 7 (53.2%), along with other deriv-atives and aphylline and sparteine in lower relative abundance (<10%) [53]. Therefore, a QRE whose composition mainly contains compound 7 and some of its analogs, such as L. albus-derived QRE, cannot be considered effective in inhibiting the F. oxysporum growth (<50%). A similar result was found in a previous study with a QRE obtained from L. mex-icanus seeds, whose practically insignificant antifungal activity against F. oxysporum was observed. The test extract was mainly composed of high contents of 7 (content = 21.2 mg/g, relative abundance = 76.2%), along with other QAs with lower content (<3 mg/g) and rel-ative abundance (<10%) [56]. Finally, the activity of QREs having good contents of 3 and 10 (4.61–6.08 mg/g fl) was found to be moderate to low. In this regard, other QAs contain-ing the 2-pyridone moiety (e.g., cytisine) have exhibited bactericidal and antifungal activ-ities [54,57]. However, the antifungal activity of 3 and 10 could not be statistically recog-nized, possibly by low inhibition by structural reasons that should be deepened in further structure-activity relationship studies.
Figure 4. Comparison of QA contents (mg/g fresh leaves) among QREs obtained from leaves of eightlupin species. (A) Heatmap visualization of the normalized content distribution (unit of variancescaling) of QAs 1–13 from test lupins. Autoscaled QA contents are related to the color scale: 2 =high content; −2 = low content. (B) Unpaired fold-change (FC) analysis plot for selecting importantQAs with threshold 2, by integrating the chemical (QA contents) and biological (mycelial growthinhibition) data. Antifungal activity was used as a categorical variable, subdividing the antifungalactivity of QREs into two datasets: more active (>80% inhibition) and less active (<75% inhibition),according to the Tukey test, to supervise the QA selection. According to the antifungal activity,positive and negative contributions to the FC-based QA selection are depicted as red and bluenumbers and dots.
On the other hand, this integrated examination also suggests that very high contentsof 7 are not a determinant for antifungal activity against F. oxysporum as the five selectedQAs. This observation agrees with a previous study on a QRE obtained from L. exaltatusseeds. Such an extract did not show antifungal activity against F. oxysporum and itsQA composition was based on a high relative abundance of 7 (53.2%), along with otherderivatives and aphylline and sparteine in lower relative abundance (<10%) [53]. Therefore,a QRE whose composition mainly contains compound 7 and some of its analogs, such as L.albus-derived QRE, cannot be considered effective in inhibiting the F. oxysporum growth(<50%). A similar result was found in a previous study with a QRE obtained from L.mexicanus seeds, whose practically insignificant antifungal activity against F. oxysporumwas observed. The test extract was mainly composed of high contents of 7 (content = 21.2mg/g, relative abundance = 76.2%), along with other QAs with lower content (<3 mg/g)and relative abundance (<10%) [56]. Finally, the activity of QREs having good contents of3 and 10 (4.61–6.08 mg/g fl) was found to be moderate to low. In this regard, other QAscontaining the 2-pyridone moiety (e.g., cytisine) have exhibited bactericidal and antifungalactivities [54,57]. However, the antifungal activity of 3 and 10 could not be statisticallyrecognized, possibly by low inhibition by structural reasons that should be deepened infurther structure-activity relationship studies.

Molecules 2022, 27, 305 9 of 17
2.5. Variation in the QA Profiles of L. polyphyllus Induced by Pruning Events
L. polyphyllus “Russell” was an exotic lupin species that quickly propagated underthese greenhouse conditions, although a flowering stage was not reached. However, theleaf production was abundant, so periodically (every two weeks), the young leaves werepruned to obtain 5 g of fresh plant material. This pruning process was performed twelvetimes, and consequently, twelve QREs were obtained which were further analyzed byGC-MS. This pruning-based exploration on this most active QRE-producing lupin planthelped refine the chemical and biological data integration to select bioactive QAs throughinherent variations.
The resulting chromatograms are shown in Figure 5. The QREs of L. polyphyllus werequalitatively and quantitatively compared, and some changes associated with the presenceand abundance of detected alkaloids in each resulting QRE were observed. Hence, tenQAs were detected and quantified (contents between 0.003 and 1.86 mg/g fl), along withpruning events (PEs) (Table 3).
Molecules 2022, 27, x FOR PEER REVIEW 9 of 18
2.5. Variation in the QA Profiles of L. polyphyllus Induced by Pruning Events L. polyphyllus “Russell” was an exotic lupin species that quickly propagated under
these greenhouse conditions, although a flowering stage was not reached. However, the leaf production was abundant, so periodically (every two weeks), the young leaves were pruned to obtain 5 g of fresh plant material. This pruning process was performed twelve times, and consequently, twelve QREs were obtained which were further analyzed by GC-MS. This pruning-based exploration on this most active QRE-producing lupin plant helped refine the chemical and biological data integration to select bioactive QAs through inherent variations.
The resulting chromatograms are shown in Figure 5. The QREs of L. polyphyllus were qualitatively and quantitatively compared, and some changes associated with the pres-ence and abundance of detected alkaloids in each resulting QRE were observed. Hence, ten QAs were detected and quantified (contents between 0.003 and 1.86 mg/g fl), along with pruning events (PEs) (Table 3).
In general, all QREs contained 4, 5, 7, 13, while 1, 6, 8, 9, 11, 12 were evidenced in selected PEs. Notably, compound 1 showed content variations for each PE (0.009–0.375 mg/g fl), with PE7-8 showing significantly higher abundance. Compound 1 has been pre-viously reported in Lupinus genotypes, and its antimicrobial history against K. pneumoniae and P. aeruginosa has been highlighted [18]. A content increase for 9 was observed from PE3-7, but no longer detected from PE9. This compound is vital due to its antimicrobial [55,58], antiparasitic [59], cytotoxic [60], anticarcinogenic [61], antiviral, [62] and antima-larial properties [63,64].
Figure 5. Stacked GC-MS chromatograms of quinolizidine-rich extract (QRE), obtained after prun-ing events (PE1–12) on L. polyphyllus. The box-enclosed numbers represent each identified quinoliz-idines, listed in Table 1. PE = pruning event.
Figure 5. Stacked GC-MS chromatograms of quinolizidine-rich extract (QRE), obtained after pruningevents (PE1–12) on L. polyphyllus. The box-enclosed numbers represent each identified quinolizidines,listed in Table 1. PE = pruning event.
In general, all QREs contained 4, 5, 7, 13, while 1, 6, 8, 9, 11, 12 were evidencedin selected PEs. Notably, compound 1 showed content variations for each PE (0.009–0.375 mg/g fl), with PE7-8 showing significantly higher abundance. Compound 1 hasbeen previously reported in Lupinus genotypes, and its antimicrobial history against K.pneumoniae and P. aeruginosa has been highlighted [18]. A content increase for 9 wasobserved from PE3-7, but no longer detected from PE9. This compound is vital due to itsantimicrobial [55,58], antiparasitic [59], cytotoxic [60], anticarcinogenic [61], antiviral, [62]and antimalarial properties [63,64].
Molecules 2022, 27, 305 10 of 17
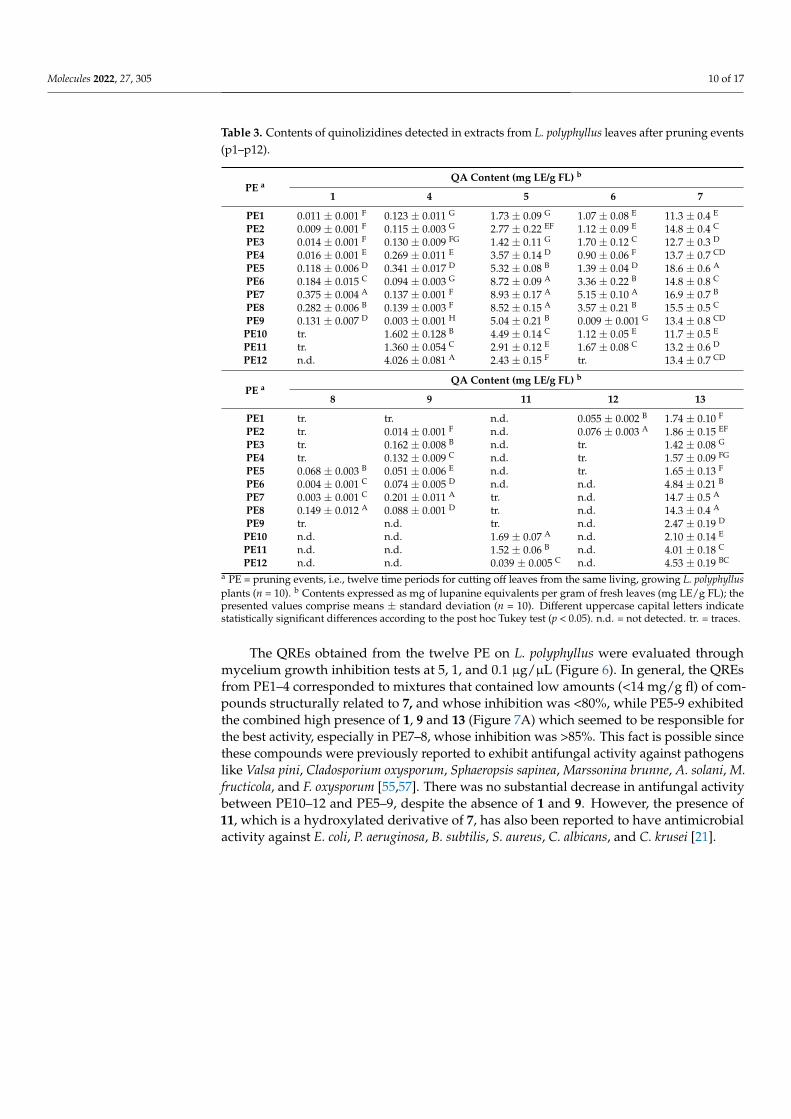
Table 3. Contents of quinolizidines detected in extracts from L. polyphyllus leaves after pruning events(p1–p12).
PE aQA Content (mg LE/g FL) b
1 4 5 6 7
PE1 0.011 ± 0.001 F 0.123 ± 0.011 G 1.73 ± 0.09 G 1.07 ± 0.08 E 11.3 ± 0.4 E
PE2 0.009 ± 0.001 F 0.115 ± 0.003 G 2.77 ± 0.22 EF 1.12 ± 0.09 E 14.8 ± 0.4 C
PE3 0.014 ± 0.001 F 0.130 ± 0.009 FG 1.42 ± 0.11 G 1.70 ± 0.12 C 12.7 ± 0.3 D
PE4 0.016 ± 0.001 E 0.269 ± 0.011 E 3.57 ± 0.14 D 0.90 ± 0.06 F 13.7 ± 0.7 CD
PE5 0.118 ± 0.006 D 0.341 ± 0.017 D 5.32 ± 0.08 B 1.39 ± 0.04 D 18.6 ± 0.6 A
PE6 0.184 ± 0.015 C 0.094 ± 0.003 G 8.72 ± 0.09 A 3.36 ± 0.22 B 14.8 ± 0.8 C
PE7 0.375 ± 0.004 A 0.137 ± 0.001 F 8.93 ± 0.17 A 5.15 ± 0.10 A 16.9 ± 0.7 B
PE8 0.282 ± 0.006 B 0.139 ± 0.003 F 8.52 ± 0.15 A 3.57 ± 0.21 B 15.5 ± 0.5 C
PE9 0.131 ± 0.007 D 0.003 ± 0.001 H 5.04 ± 0.21 B 0.009 ± 0.001 G 13.4 ± 0.8 CD
PE10 tr. 1.602 ± 0.128 B 4.49 ± 0.14 C 1.12 ± 0.05 E 11.7 ± 0.5 E
PE11 tr. 1.360 ± 0.054 C 2.91 ± 0.12 E 1.67 ± 0.08 C 13.2 ± 0.6 D
PE12 n.d. 4.026 ± 0.081 A 2.43 ± 0.15 F tr. 13.4 ± 0.7 CD
PE aQA Content (mg LE/g FL) b
8 9 11 12 13
PE1 tr. tr. n.d. 0.055 ± 0.002 B 1.74 ± 0.10 F
PE2 tr. 0.014 ± 0.001 F n.d. 0.076 ± 0.003 A 1.86 ± 0.15 EF
PE3 tr. 0.162 ± 0.008 B n.d. tr. 1.42 ± 0.08 G
PE4 tr. 0.132 ± 0.009 C n.d. tr. 1.57 ± 0.09 FG
PE5 0.068 ± 0.003 B 0.051 ± 0.006 E n.d. tr. 1.65 ± 0.13 F
PE6 0.004 ± 0.001 C 0.074 ± 0.005 D n.d. n.d. 4.84 ± 0.21 B
PE7 0.003 ± 0.001 C 0.201 ± 0.011 A tr. n.d. 14.7 ± 0.5 A
PE8 0.149 ± 0.012 A 0.088 ± 0.001 D tr. n.d. 14.3 ± 0.4 A
PE9 tr. n.d. tr. n.d. 2.47 ± 0.19 D
PE10 n.d. n.d. 1.69 ± 0.07 A n.d. 2.10 ± 0.14 E
PE11 n.d. n.d. 1.52 ± 0.06 B n.d. 4.01 ± 0.18 C
PE12 n.d. n.d. 0.039 ± 0.005 C n.d. 4.53 ± 0.19 BC
a PE = pruning events, i.e., twelve time periods for cutting off leaves from the same living, growing L. polyphyllusplants (n = 10). b Contents expressed as mg of lupanine equivalents per gram of fresh leaves (mg LE/g FL); thepresented values comprise means ± standard deviation (n = 10). Different uppercase capital letters indicatestatistically significant differences according to the post hoc Tukey test (p < 0.05). n.d. = not detected. tr. = traces.
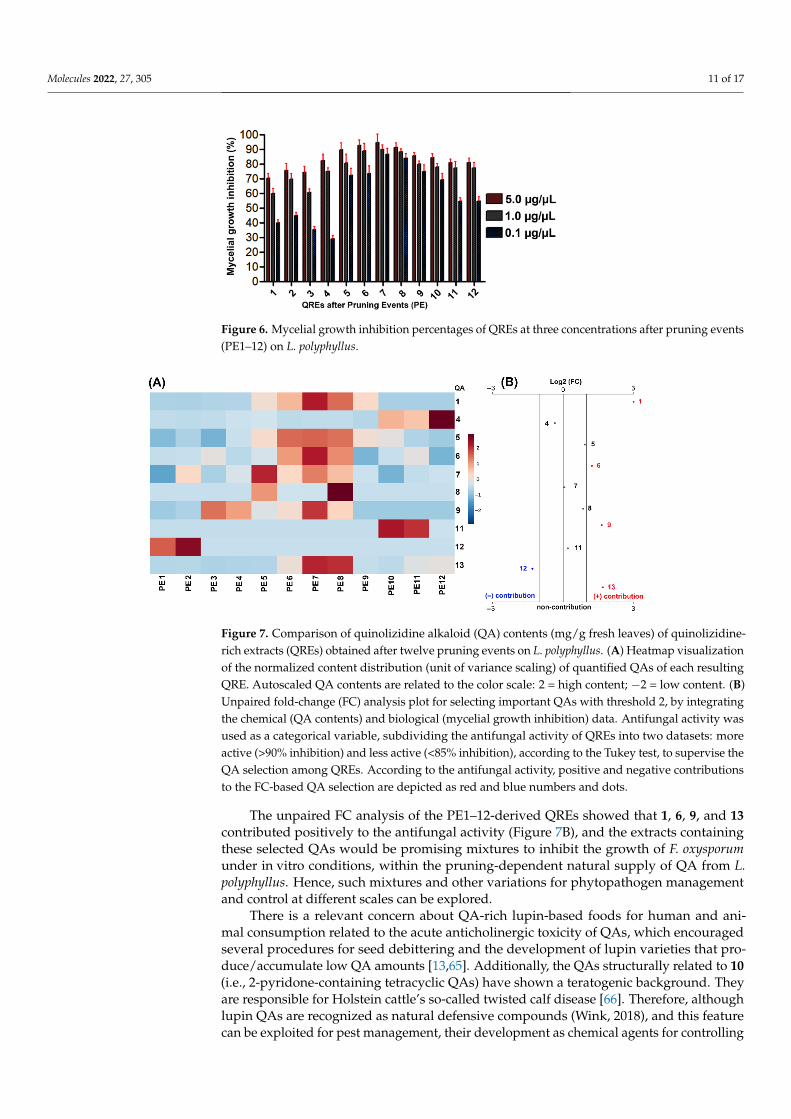
The QREs obtained from the twelve PE on L. polyphyllus were evaluated throughmycelium growth inhibition tests at 5, 1, and 0.1 µg/µL (Figure 6). In general, the QREsfrom PE1–4 corresponded to mixtures that contained low amounts (<14 mg/g fl) of com-pounds structurally related to 7, and whose inhibition was <80%, while PE5-9 exhibitedthe combined high presence of 1, 9 and 13 (Figure 7A) which seemed to be responsible forthe best activity, especially in PE7–8, whose inhibition was >85%. This fact is possible sincethese compounds were previously reported to exhibit antifungal activity against pathogenslike Valsa pini, Cladosporium oxysporum, Sphaeropsis sapinea, Marssonina brunne, A. solani, M.fructicola, and F. oxysporum [55,57]. There was no substantial decrease in antifungal activitybetween PE10–12 and PE5–9, despite the absence of 1 and 9. However, the presence of11, which is a hydroxylated derivative of 7, has also been reported to have antimicrobialactivity against E. coli, P. aeruginosa, B. subtilis, S. aureus, C. albicans, and C. krusei [21].
Molecules 2022, 27, 305 11 of 17Molecules 2022, 27, x FOR PEER REVIEW 11 of 18
Figure 6. Mycelial growth inhibition percentages of QREs at three concentrations after pruning events (PE1-12) on L. polyphyllus.
The unpaired FC analysis of the PE1–12-derived QREs showed that 1, 6, 9, and 13 contributed positively to the antifungal activity (Figure 7B), and the extracts containing these selected QAs would be promising mixtures to inhibit the growth of F. oxysporum under in vitro conditions, within the pruning-dependent natural supply of QA from L. polyphyllus. Hence, such mixtures and other variations for phytopathogen management and control at different scales can be explored.
Figure 7. Comparison of quinolizidine alkaloid (QA) contents (mg/g fresh leaves) of quinoliz-idine-rich extracts (QREs) obtained after twelve pruning events on L. polyphyllus. (A) Heatmap visualization of the normalized content distribution (unit of variance scaling) of quantified QAs of each resulting QRE. Autoscaled QA contents are related to the color scale: 2 = high content; –2 = low content. (B) Unpaired fold-change (FC) analysis plot for selecting important QAs with threshold 2, by integrating the chemical (QA contents) and biological (mycelial growth inhibi-tion) data. Antifungal activity was used as a categorical variable, subdividing the antifungal activity of QREs into two datasets: more active (>90% inhibition) and less active (<85% inhibi-tion), according to the Tukey test, to supervise the QA selection among QREs. According to the antifungal activity, positive and negative contributions to the FC-based QA selection are de-picted as red and blue numbers and dots.
There is a relevant concern about QA-rich lupin-based foods for human and animal consumption related to the acute anticholinergic toxicity of QAs, which encouraged sev-eral procedures for seed debittering and the development of lupin varieties that pro-duce/accumulate low QA amounts [13,65]. Additionally, the QAs structurally related to 10 (i.e., 2-pyridone-containing tetracyclic QAs) have shown a teratogenic background. They are responsible for Holstein cattle’s so-called twisted calf disease [66]. Therefore, although lupin QAs are recognized as natural defensive compounds (Wink, 2018), and this feature can be exploited for pest management, their development as chemical agents
Figure 6. Mycelial growth inhibition percentages of QREs at three concentrations after pruning events(PE1–12) on L. polyphyllus.
Molecules 2022, 27, x FOR PEER REVIEW 11 of 18
Figure 6. Mycelial growth inhibition percentages of QREs at three concentrations after pruning events (PE1-12) on L. polyphyllus.
The unpaired FC analysis of the PE1–12-derived QREs showed that 1, 6, 9, and 13 contributed positively to the antifungal activity (Figure 7B), and the extracts containing these selected QAs would be promising mixtures to inhibit the growth of F. oxysporum under in vitro conditions, within the pruning-dependent natural supply of QA from L. polyphyllus. Hence, such mixtures and other variations for phytopathogen management and control at different scales can be explored.
Figure 7. Comparison of quinolizidine alkaloid (QA) contents (mg/g fresh leaves) of quinoliz-idine-rich extracts (QREs) obtained after twelve pruning events on L. polyphyllus. (A) Heatmap visualization of the normalized content distribution (unit of variance scaling) of quantified QAs of each resulting QRE. Autoscaled QA contents are related to the color scale: 2 = high content; –2 = low content. (B) Unpaired fold-change (FC) analysis plot for selecting important QAs with threshold 2, by integrating the chemical (QA contents) and biological (mycelial growth inhibi-tion) data. Antifungal activity was used as a categorical variable, subdividing the antifungal activity of QREs into two datasets: more active (>90% inhibition) and less active (<85% inhibi-tion), according to the Tukey test, to supervise the QA selection among QREs. According to the antifungal activity, positive and negative contributions to the FC-based QA selection are de-picted as red and blue numbers and dots.
There is a relevant concern about QA-rich lupin-based foods for human and animal consumption related to the acute anticholinergic toxicity of QAs, which encouraged sev-eral procedures for seed debittering and the development of lupin varieties that pro-duce/accumulate low QA amounts [13,65]. Additionally, the QAs structurally related to 10 (i.e., 2-pyridone-containing tetracyclic QAs) have shown a teratogenic background. They are responsible for Holstein cattle’s so-called twisted calf disease [66]. Therefore, although lupin QAs are recognized as natural defensive compounds (Wink, 2018), and this feature can be exploited for pest management, their development as chemical agents
Figure 7. Comparison of quinolizidine alkaloid (QA) contents (mg/g fresh leaves) of quinolizidine-rich extracts (QREs) obtained after twelve pruning events on L. polyphyllus. (A) Heatmap visualizationof the normalized content distribution (unit of variance scaling) of quantified QAs of each resultingQRE. Autoscaled QA contents are related to the color scale: 2 = high content; −2 = low content. (B)Unpaired fold-change (FC) analysis plot for selecting important QAs with threshold 2, by integratingthe chemical (QA contents) and biological (mycelial growth inhibition) data. Antifungal activity wasused as a categorical variable, subdividing the antifungal activity of QREs into two datasets: moreactive (>90% inhibition) and less active (<85% inhibition), according to the Tukey test, to supervise theQA selection among QREs. According to the antifungal activity, positive and negative contributionsto the FC-based QA selection are depicted as red and blue numbers and dots.
The unpaired FC analysis of the PE1–12-derived QREs showed that 1, 6, 9, and 13contributed positively to the antifungal activity (Figure 7B), and the extracts containingthese selected QAs would be promising mixtures to inhibit the growth of F. oxysporumunder in vitro conditions, within the pruning-dependent natural supply of QA from L.polyphyllus. Hence, such mixtures and other variations for phytopathogen managementand control at different scales can be explored.
There is a relevant concern about QA-rich lupin-based foods for human and ani-mal consumption related to the acute anticholinergic toxicity of QAs, which encouragedseveral procedures for seed debittering and the development of lupin varieties that pro-duce/accumulate low QA amounts [13,65]. Additionally, the QAs structurally related to 10(i.e., 2-pyridone-containing tetracyclic QAs) have shown a teratogenic background. Theyare responsible for Holstein cattle’s so-called twisted calf disease [66]. Therefore, althoughlupin QAs are recognized as natural defensive compounds (Wink, 2018), and this featurecan be exploited for pest management, their development as chemical agents for controlling

Molecules 2022, 27, 305 12 of 17
phytopathogens should be rationally and carefully conducted. In this regard, these findingscan serve as a reference in future studies to explore the composition-activity (and evenstructure-activity) relationship of QAs to optimize such an antifungal potential and reducepossible side effects due to the toxicity.
3. Materials and Methods3.1. Propagation of Lupinus Plants under Greenhouse Conditions
Seeds of the three native species, i.e., L. bogotensis Benth., L. mutabilis Sweet, and L.mirabilis C.P. Sm., were collected on the campus at Military University Nueva Granada(UMNG) (4◦56′ N, 74◦00′ W and 2562 masl). Seeds of five exotic species, i.e., L. albus L., L.argenteus Pursh, L. polyphyllus Lindl. “Russell”, L. perennis L., and L. arboreus Sims, werecommercially purchased from Sow Right Seeds (https://sowrightseeds.com, accessed on30 November 2021).
All the Lupinus seeds were superficially disinfected with 70% ethanol (2 min) andthen with 3% sodium hypochlorite (5 min), subsequently washed with sterile distilledwater (3 × 3 min), and cleaned with a drop of Tween 20. The seeds were planted in 72-cellseedbeds that contained a substrate of loamy-silty soil (LSS) with rice husk (RiH) at a 3:1ratio and were maintained under greenhouse conditions (temperature = 21 ± 4 ◦C; relativehumidity (RH) = 65 ± 15%, altitude = 2562 masl, total light transmission = 85 ± 5%, totallight diffusion = 55 ± 5%, and UV transmission between 290–340 nm = 5%) during thewhole propagation experiment (30 days). At 8–10 days after seed planting (dasp), the seedsstarted the germination process (i.e., seed coat breakdown) and subsequent embryonic root(radicle) growing. Cotyledons appeared between 12–15 dasp on average.
After 20–30 dasp, the first and second pairs of leaves emerged, and the resultingseedlings (n = 20) were transplanted in 2 L bags on a substrate containing a mixture ofLSS/RiH 3:1 and maintained under the same greenhouse conditions. After transplanting,the seedlings were watered with water (500 mL) every two days, and once the elongation ofthe central axis began to be observed, we proceeded to apply commercial triple-15 fertilizer(5%). After 70 ± 19 days of development under these conditions (i.e., after transplanting)to reach the flowering stage, fresh leaves were collected for QA extraction.
In addition, a new set of plants of L. polyphyllus were propagated at the same green-house conditions. After 50 days of transplanting, leaves were removed every two weeksfrom the same living, growing plants (n = 10) and subsequently prepared for QA extraction.Each pruning event (PE) was performed at the same time of the day (i.e., 10 a.m.). Thispruning process was repeated twelve times (PE1–PE12).
3.2. Preparation of Quinolizidine-Rich Extracts (QREs)
Fresh young leaves (5 g) of each biological replicate (n = 10) were extracted with 0.5M HCl (20 mL) under stirring for 24 h at 130 rpm in an orbital shaker. Subsequently, theacidic solution was filtered and alkalinized to pH = 10 with a 15% aqueous NH3 solution.Subsequently, liquid-liquid extraction was performed using chloroform to obtain an organicphase enriched in QAs. Finally, the solvent was removed by distillation under reducedpressure at 375 mbar for 5 min, and Lupinus QREs were then obtained.
3.3. Gas Chromatography Coupled to Mass Spectrometry (GC-MS)
The chromatographic profiles and mass spectra were obtained with a Thermo Trace1300 equipped with a flame ionization detector (FID) and coupled to an ISQ LT massspectrometer with a single quadrupole analyzer. For the analysis, an Rxi® 5Sil MS column(5% diphenyl/95% dimethylpolysiloxane, 60 m, 0.25 mm ID, and 0.25 µm) was used. Atemperature program was implemented; the starting temperature was 120 ◦C, maintainedfor 2 min, and then a 6 ◦C/min program was applied until 300 ◦C and kept for 10 min. Thetest QREs were prepared at 1 µg/µL in CH2Cl2 (GC-MS grade, SupraSolv®). The injectionvolume was 1 µL in split mode (split ratio = 30). The transfer line temperature was 250 ◦C,and the carrier gas was grade-5 helium (flow = 1 mL/min). The ionization mode was the

Molecules 2022, 27, 305 13 of 17
electronic impact (EI) at 70 eV. The retention indices (RI) were calculated using a series ofC10-C24 n-alkanes [67], according to Equation (S1) (supplementary material). Compounds1–13 were identified by diagnostic analysis of their mass spectra and comparison of RIswith available literature. On the other hand, the QAs quantification was performed usingthe external standard method by GC-FID under the same chromatographic conditions. Thestandard curve was built by injecting eight solutions (ranging from 1 to 500 µg/mL) of(+)-lupanine (100 µg/mL, Sigma-Aldrich, St. Louis, MO, USA). The instrument responsewas verified by adding caffeine (100 µg/mL, Sigma-Aldrich, St. Louis, MO, USA) as aninternal standard. Owing to limitations related to commercial availability or purity degreeof reference compounds of identified QAs compounds, 1–13 were quantified as lupanineequivalents and expressed as mg lupanine equivalents per gram of fresh leaves (mg LE/g fl).Relative response factors were used to correct the peak areas of detected QAs. Quantitativeanalysis was performed in triplicate. The intra and inter-day analyzes of (+)-lupanine wereused to evaluate the method precision, whose relative standard deviations (RSD %) were2.4 and 3.7%, respectively. The limit of detection (LOD) and limit of quantification (LOQ)of (+)-lupanine was 1 and 2 µg/mL, respectively. Finally, the GC-MS-derived data wereprocessed in MZmine 2 software to compare the resulting chromatographic profiles ofQREs. Thus, the baseline correction was performed and exported as total ionic current(TIC) to a CSV file. Data were normalized and autoscaled. The resulting autoscaled profileswere stacked in OriginPro 8.5 for intuitive visualization of the QA variations between eachQRE.
3.4. Direct Bioautography Assay
A spore suspension of the fungus F. oxysporum was prepared at a concentration of 1 ×106 CFU/mL on a nutrient broth containing KH2PO4 (7 g), Na2HPO4.2H2O (3 g), KNO3 (4g), MgSO4.7H2O (1 g), and NaCl (1 g) per liter of water. The culture broth was sterilized for1 h in an autoclave at 120 ◦C. Then the spore suspension was prepared, adding 30% glucose(10 mL) for every 60 mL standard solution. Respective QRE solutions were prepared inCHCl3 at 1 mg/mL. Subsequently, the QRE solutions (50 µL) were seeded on silica gel60 F254 thin-layer chromatography (TLC) plates (20 × 20 cm, 0.20 mm layer thickness,subdivided into squares), and the solvent was removed in an extraction chamber. Then, thespore solution of F. oxysporum was sprayed on the plate surface and placed into a humidand dark chamber at 25 ◦C for 72 h. Dithane (mancozeb) and Rovral (iprodione) were usedas positive controls at the same doses. In this test, the antifungal activity was determined bylight areas on the squares that indicated no fungal growth by inhibiting spore germination,contrasted with dark areas that revealed fungal growth [68].
3.5. Mycelial Growth Inhibition Assay
Antifungal activity evaluation of QREs was performed by measuring the growthhalo of the phytopathogen F. oxysporum with the presence of the extracts at differentconcentrations compared to that of a blank (0.5% PDA), using the amended-mediumprocedure [69]. The culture medium contained 2.4% PDB and 1.5% bacteriological agar in100 mL of distilled water. The medium was homogenized for 2 min in a microwave ovenand then sterilized in an autoclave for 1 h at 120 ◦C. The culture medium (20 mL) was thenplaced into a previously sterilized Petri dish to propagate the fungus. Once it cooled andsolidified, a 2-mm plug from a previously prepared monosporic culture was placed ontothe central part of the Petri dish and left to grow at 28 ◦C for 8 days.
Three treatments per QRE were prepared for the antifungal assays according to threefinal QRE concentrations (i.e., 5, 1, and 0.1 µg/µL). Thus, the required amount of therespective QRE was dispersed in 0.5% PDA to afford the QRE-amended medium pertreatment. Subsequently, each treatment was randomly placed in a 12-well glass plate (79× 63 × 4 mm). Finally, a 1.1-mm plug (equivalent to the internal diameter of a 1.1-mmborosilicate capillary tube) was taken from an 8-day phytopathogen culture and placedonto the center of each well (QRE-amended and non-amended). This plate was placed

Molecules 2022, 27, 305 14 of 17
into a humid chamber for 72 h at 25 ◦C. The evaluation of each concentration per QREtreatment was performed in triplicate. Dithane (mancozeb) and Rovral (iprodione) wereused as positive controls at the same doses. After the incubation time, a photograph of the12-well plate was taken and analyzed in ImageJ software, whose growth areas of control(non-amended) and QRE-amended wells were measured. The comparison of these areasled to the determination of the inhibition percentage using Equation (1).
Inhibition Percentage =areacontrol − areaQRE
areacontrol× 100% (1)
3.6. Fungicidal (FC) and Fungistatic (FS) Activity
For the fungicidal or fungistatic activity classification procedure, the central plug ofthe phytopathogen used in the prior 5-µg/µL-amended treatment was retrieved and placedonto fresh, non-amended PDA medium for 72 h. After this time, mycelial growth wasadditionally monitored. The QRE was classified as fungistatic or fungicidal if mycelialgrowth or no mycelial growth, respectively, was observed [70].
3.7. Data Analysis
A Shapiro-Wilks normality test was accomplished to examine the normal distributionof the quantitative data (p > 0.05). Once the normal distribution of the data was verified, ananalysis of variance (ANOVA) was subsequently performed, followed by a post hoc Tukeytest to establish significant differences between samples (p < 0.05). These analyses wereperformed in Infostat statistical software [71]. In addition, a heatmap was constructed usingnormalized QA contents through autoscaling. The antifungal activity was subsequentlydivided into two datasets (i.e., more and less active) depending on the significantly differentmean groups after the Tukey test. An unpaired fold-change analysis (threshold = 2) wasfinally performed on chemical and antifungal activity datasets to select important QAs thatpositively and negatively contribute to mycelial growth inhibition.
4. Conclusions
The present study attempts to combine the information of QA profiles and the anti-fungal activities against F. oxysporum of lupin derived QREs. This chemical and antifungaldataset integration provided valuable information of naturally afforded QA mixtures tounderstand the promising concomitant effects against F. oxysporum. In summary, we foundthat the test QREs showed important antifungal activity since all extracts showed inhibition> 50% at the maximum test concentration (5 µg/µL). The best antifungal results wereobtained for the QREs of L. mirabilis, L. polyphyllus “Russell”, and L. bogotensis, reachinginhibitions ≥ 90% and similar to positive controls. In addition, all QREs were classified asfungistatic except for the L. polyphyllus-derived QRE ranked as a fungicide. QREs obtainedafter twelve pruning events on L. polyphyllus exhibited antifungal activity variations, withthe QREs derived from PE7–8 being the most active extracts. Each test extract in the presentstudy showed differential QA profiles. In-depth qualitative and quantitative analysis ofsuch QA differences indicated that QREs containing particular combinations of angusti-foline (4), α-isolupanine (6), matrine (9), 13α-hydroxylupanine (11), and 17-oxolupanine(13) best inhibit the growth of F. oxysporum. Contrarily, lupanine (7) seems to have a lowcontribution to the antifungal activity despite its high abundance in test QREs. A betterunderstanding of such bioactivity of QA mixtures would promote further studies to deeplyunderstand and exploit their antifungal potential at different levels (i.e., greenhouse andfield conditions) for managing and controlling F. oxysporum within IPM programs.
Supplementary Materials: The following supporting information can be downloaded online. Ta-ble S1: Yields of QREs obtained from propagated Lupinus species under greenhouse, Table S2:Replicates of the mycelial growth inhibition percentages of each lupin-based treatment, Figure S1:structures of QA identified in the eight species of Lupinus, Figure S2: Antifungal activity against F.oxysporum of eight Lupinus species, and Equation (S1): Retention index calculation.

Molecules 2022, 27, 305 15 of 17
Author Contributions: Conceptualization, E.C.-B.; methodology, W.C.-V.; software, W.C.-V. andE.C.-B.; validation, W.C.-V., D.Q. and E.C.-B.; formal analysis, W.C.-V. and E.C.-B.; resources, D.Q.and E.C.-B.; data curation, W.C.-V. and E.C.-B.; writing—original draft preparation, W.C.-V.; writing—review and editing, W.C.-V., D.Q. and E.C.-B.; supervision, E.C.-B.; project administration, E.C.-B.;funding acquisition, D.Q. and E.C.-B. All authors have read and agreed to the published version ofthe manuscript.
Funding: This study was funded by the Vicerrectoría de Investigaciones at Universidad MilitarNueva Granada through the project IMP-CIAS-2924, validity 2019.
Institutional Review Board Statement: Not applicable.
Informed Consent Statement: Not applicable.
Data Availability Statement: The data that support the findings of this study are available from thecorresponding author upon reasonable request.
Acknowledgments: Authors thank the Universidad Militar Nueva Granada for the financial support.
Conflicts of Interest: The authors declare no conflict of interest.
Sample Availability: Samples of the substances are available from the corresponding author uponreasonable request.
References1. Krishna, P.M.; Rao, K.N.V.; Sandhya, S.; Banji, D. A review on phytochemical, ethnomedical and pharmacological studies on
genus Sophora, Fabaceae. Braz. J. Pharmacogn. 2012, 22, 1145–1154. [CrossRef]2. Wink, M. Evolution of secondary metabolites in legumes (Fabaceae). S. Afr. J. Bot. 2013, 89, 164–175. [CrossRef]3. Confortin, T.C.; Todero, I.; Soares, J.F.; Brun, T.; Luft, L.; Ugalde, G.A.; Prá, V.D.; Mazutti, M.A.; Zabot, G.L.; Tres, M.V. Extraction
and composition of extracts obtained from Lupinus albescens using supercritical carbon dioxide and compressed liquefiedpetroleum gas. J. Supercrit. Fluids 2017, 128, 395–403. [CrossRef]
4. Kole, C. Wild Crop Relatives: Genomic and Breeding Resources. Legume Crops and Forages; Springer: Berlin/Heidelberg, Germany,2011; ISBN 9783642143878.
5. Gresta, F.; Wink, M.; Prins, U.; Abberton, M.; Capraro, J.; Scarafoni, A.; Hill, G. Lupins in European Cropping Systems. In Legumesin Cropping Systems; Murphy-Bokern, D., Stoddard, F., Watson, C., Eds.; CABI Publishing: Wallingford, CT, USA, 2017; pp. 88–108.
6. Contreras-Ortiz, N.; Jara-Muñoz, O.A.; Hughes, C.E. The acaulescent rosette species of Lupinus L. (Fabaceae) of Colombia andEcuador including a new species from Colombia. Phytotaxa 2018, 364, 61–70. [CrossRef]
7. Lucas, M.M.; Stoddard, F.; Annicchiarico, P.; Frias, J.; Martinez-Villaluenga, C.; Sussmann, D.; Duranti, M.; Seger, A.; Zander, P.;Pueyo, J. The future of lupin as a protein crop in Europe. Front. Plant Sci. 2015, 6, 705. [CrossRef]
8. Cortés-Avendaño, P.; Tarvainen, M.; Suomela, J.P.; Glorio-Paulet, P.; Yang, B.; Repo-Carrasco-Valencia, R. Profile and content ofresidual alkaloids in ten ecotypes of Lupinus mutabilis Sweet after aqueous debittering process. Plant Foods Hum. Nutr. 2020, 75,184–191. [CrossRef] [PubMed]
9. Wink, M. Introduction: Biochemistry, Physiology and Ecological Functions of Secondary Metabolites. In Biochemistry of PlantSecondary Metabolism; Wiley-Blackwell: Oxford, UK, 2010; Volume 40, pp. 1–19. ISBN 9781444320503.
10. Hernández, E.M.; Rangel, M.L.C.; Corona, A.E.; del Angel, J.A.C.; López, J.A.S.; Sporer, F.; Wink, M.; Torres, K.B. Quinolizidinealkaloid composition in different organs of Lupinus aschenbornii. Rev. Bras. Farmacogn. 2011, 21, 824–828. [CrossRef]
11. Neto, A.T.; Oliveira, C.Q.; Ilha, V.; Pedroso, M.; Burrow, R.A.; Dalcol, I.I.; Morel, A.F. Quinolizidine alkaloids from Lupinus lanatus.J. Mol. Struct. 2011, 1004, 174–177. [CrossRef]
12. Aisyah, S.; Vincken, J.P.; Andini, S.; Mardiah, Z.; Gruppen, H. Compositional changes in (iso)flavonoids and estrogenic activity ofthree edible Lupinus species by germination and Rhizopus-elicitation. Phytochemistry 2016, 122, 65–75. [CrossRef]
13. Frick, K.M.; Kamphuis, L.G.; Siddique, K.H.M.; Singh, K.B.; Foley, R.C. Quinolizidine alkaloid biosynthesis in Lupins andprospects for grain quality improvement. Front. Plant Sci. 2017, 8, 87. [CrossRef]
14. Boschin, G.; Resta, D. Alkaloids Derived from Lysine: Quinolizidine (a focus on Lupin alkaloids) BT—Natural products:Phytochemistry, botany and metabolism of alkaloids, phenolics and terpenes. In Natural Products. Phytochemistry, Botany andMetabolism of Alkaloids, Phenolics and Terpenes; Ramawat, K.G., Mérillon, J.-M., Eds.; Springer: Berlin/Heidelberg, Germany, 2013;pp. 381–403. ISBN 978-3-642-22144-6.
15. Wink, M. Plant secondary metabolites modulate insect behavior-steps toward addiction? Front. Physiol. 2018, 9, 364. [CrossRef]16. Arias Alemán, L.S.E.; Ulloa Ramones, L.A.; Rojas Oviedo, L.A.; Noboa Abdo, T.E. Effect of alkaloids of Lupinus mutabilis Sweet on
gastrointestinal parasites in guinea pigs. Cienc. Digit. 2019, 3, 221–228. [CrossRef]17. Wink, M. Chemical defense of Lupins. Mollusc-repellent properties of Quinolizidine Alkaloids. Z. Naturforsch. C 1984, 39,
553–558. [CrossRef]

Molecules 2022, 27, 305 16 of 17
18. Romeo, F.; Fabroni, S.; Ballistreri, G.; Muccilli, S.; Spina, A.; Rapisarda, P. Characterization and antimicrobial activity of alkaloidextracts from seeds of different genotypes of Lupinus spp. Sustainability 2018, 10, 788. [CrossRef]
19. Ohadoma, S.C.; Nnatuanya, I.; Amazu, L.U.; Okolo, C.E. Antimicrobial activity of the leaf extract and fractions of Lupinus arboreus.J. Med. Plants Res. 2014, 8, 386–391. [CrossRef]
20. Confortin, T.C.; Todero, I.; Soares, J.F.; Luft, L.; Brun, T.; Rabuske, J.E.; Nogueira, C.U.; Mazutti, M.A.; Zabot, G.L.; Tres, M.V.Extracts from Lupinus albescens: Antioxidant power and antifungal activity in vitro against phytopathogenic fungi. Environ.Technol. 2019, 40, 1668–1675. [CrossRef] [PubMed]
21. Erdemoglu, N.; Ozkan, S.; Tosun, F. Alkaloid profile and antimicrobial activity of Lupinus angustifolius L. alkaloid extract.Phytochem. Rev. 2007, 6, 197–201. [CrossRef]
22. El-Shazly, A.; Ateya, A.M.; Wink, M. Quinolizidine alkaloids profiles of Lupinus varius orientalis, L. albus albus, L. hartweguii and L.desinflorus. Z. Naturforsch. C 2001, 56, 21–30. [CrossRef] [PubMed]
23. Bernal-Alcocer, A.; Zamora-Natera, J.F.; Virgen-calleros, G.; Nuño-romero, R. In vitro biological activity of extracts of Lupinus spp.on phytopathogenic fungi. Rev. Mex. Fitopatol. 2005, 23, 140–146.
24. Pegg, K.G.; Coates, L.M.; O’Neill, W.T.; Turner, D.W. The epidemiology of Fusarium wilt of banana. Front. Plant Sci. 2019, 10,1395. [CrossRef]
25. Edel-Hermann, V.; Lecomte, C. Current status of Fusarium oxysporum formae speciales and races. Phytopathology 2019, 109, 512–530.[CrossRef] [PubMed]
26. Giraldo-Betancourt, C.; Velandia-Sánchez, E.A.; Fischer, G.; Gómez-Caro, S.; Martínez, L.J. Hyperspectral response of capegooseberry (Physalis peruviana L.) plants inoculated with Fusarium oxysporum f. sp. physali for vascular wilt detection. Rev. Colomb.Cienc. Hortic. 2020, 14, 301–313. [CrossRef]
27. García-Bastidas, F.A.; Quintero-Vargas, J.C.; Ayala-Vasquez, M.; Schermer, T.; Seidl, M.F.; Santos-Paiva, M.; Noguera, A.M.;Aguilera-Galvez, C.; Wittenberg, A.; Hofstede, R.; et al. First report of Fusarium wilt tropical race 4 in cavendish bananas causedby Fusarium odoratissimum in Colombia. Plant Dis. 2020, 104, 994. [CrossRef]
28. Basallote-Ureba, M.J.; Vela-Delgado, M.D.; Capote, N.; Melero-Vara, J.M.; López-Herrera, C.J.; Prados-Ligero, A.M.; Talavera-Rubia, M.F. Control of Fusarium wilt of carnation using organic amendments combined with soil solarization, and report ofassociated Fusarium species in southern Spain. Crop Prot. 2016, 89, 184–192. [CrossRef]
29. Jangir, M.; Pathak, R.; Sharma, S.; Sharma, S. Biocontrol mechanisms of Bacillus sp., isolated from tomato rhizosphere, againstFusarium oxysporum f. sp. lycopersici. Biol. Control 2018, 123, 60–70. [CrossRef]
30. Amini, J.; Dzhalilov, F.S. The effects of fungicides on Fusarium oxysporum f. sp. lycopersici associated with Fusarium wilt of tomato.J. Plant Prot. Res. 2010, 50, 172–178. [CrossRef]
31. Ajilogba, C.F.; Babalola, O.O. Integrated management strategies for tomato Fusarium Wilt. Biocontrol Sci. 2013, 18, 117–127.[CrossRef]
32. Hernández Martínez, R.; López Benítez, A.; Borrego Escalante, F.; Espinoza Velázquez, J.; Sánchez Aspeytia, D.; MaldonadoMendoza, I.E.; López Ochoa, L.A. Races of Fusarium oxysporum f. sp. lycopersici in tomato farmlands in San Luis Potosí. Rev. Mex.Cienc. Agrícolas 2018, 5, 1169. [CrossRef]
33. Petz, W.; Foissner, W. The effects of mancozeb and lindane on the soil microfauna of a spruce forest: A field study using acompletely randomized block design. Biol. Fertil. Soils 1989, 7, 225–231. [CrossRef]
34. Walia, A.; Mehta, P.; Guleria, S.; Chauhan, A.; Shirkot, C.K. Impact of fungicide mancozeb at different application rates on soilmicrobial populations, soil biological processes, and enzyme activities in soil. Sci. World J. 2014, 2014, 702909. [CrossRef]
35. Adhikari, K.N.; Buirchell, B.J.; Sweetingham, M.W. Length of vernalization period affects flowering time in three lupin species.Plant Breed. 2012, 131, 631–636. [CrossRef]
36. Clapham, W.L.; Willcott, J.B. A Thermosensitivity index for Lupinus albus L. Crop Sci. 1999, 39, 578–580. [CrossRef]37. Podlesny, J.; Podlesna, A. The effect of high temperature during flowering on growth, development and yielding of blue
lupine-barley mixture. J. Food Agric. Environ. 2012, 10, 500–504.38. Li, Y.; Kong, D.; Fu, Y.; Sussman, M.R.; Wu, H. The effect of developmental and environmental factors on secondary metabolites
in medicinal plants. Plant Physiol. Biochem. 2020, 148, 80–89. [CrossRef]39. Przybylak, J.K.; Ciesiołka, D.; Wysocka, W.; García-López, P.M.; Ruiz-López, M.A.; Wysocki, W.; Gulewicz, K. Alkaloid profiles of
Mexican wild lupin and an effect of alkaloid preparation from Lupinus exaltatus seeds on growth and yield of paprika (Capsicumannuum L.). Ind. Crop. Prod. 2005, 21, 1–7. [CrossRef]
40. Cook, D.; Lee, S.T.; Gardner, D.R.; Pfister, J.A.; Welch, K.D.; Green, B.T.; Davis, T.Z.; Panter, K.E. The alkaloid profiles of Lupinussulphureus. J. Agric. Food Chem. 2009, 57, 1646–1653. [CrossRef]
41. McDougal, O.M.; Heenan, P.B.; Jaksons, P.; Sansom, C.E.; Smallfield, B.M.; Perry, N.B.; van Klink, J.W. Alkaloid variation in NewZealand kowhai, Sophora species. Phytochemistry 2015, 118, 9–16. [CrossRef]
42. Martínez-Herrera, J.; Robledo-Quintos, N.; Mora-Escobedo, R.; Dávila-Ortíz, G. Alkaloid composition of Lupinus campestris fromMexico. J. Food Biochem. 2001, 25, 117–125. [CrossRef]
43. Kianbakht, S.; Dabaghian, F.H. Sophora alopecuroides L. var. alopecuroides alleviates morphine withdrawal syndrome in mice:Involvement of alkaloid fraction and matrine. Iran. J. Basic Med. 2016, 19, 1090–1095.

Molecules 2022, 27, 305 17 of 17
44. Basilico, N.; Parapini, S.; Sparatore, A.; Romeo, S.; Misiano, P.; Vivas, L.; Yardley, V.; Croft, S.L.; Habluetzel, A.; Lucantoni, L.; et al.In vivo and in vitro activities and ADME-tox profile of a quinolizidine-modified 4-aminoquinoline: A potent anti-P. falciparumand Anti-P. vivax blood-stage antimalarial. Molecules 2017, 22, 2102. [CrossRef]
45. Wink, M.; Carey, D.B. Variability of quinolizidine alkaloid profiles of Lupinus argentous (Fabaceae) from North America. Biochem.Syst. Ecol. 1994, 22, 663–669. [CrossRef]
46. Adler, L.S.; Kittelson, P.M. Variation in Lupinus arboreus alkaloid profiles and relationships with multiple herbivores. Biochem.Syst. Ecol. 2004, 32, 371–390. [CrossRef]
47. Kroc, M.; Rybinski, W.; Wilczura, P.; Kamel, K.; Kaczmarek, Z.; Barzyk, P.; Swiecicki, W. Quantitative and qualitative analysis ofalkaloids composition in the seeds of a white lupin (Lupinus albus L.) collection. Genet. Resour. Crop Evol. 2017, 64, 1853–1860.[CrossRef]
48. Vargas-Medina, L.; Yamaguchi, L.F.; Coy-Barrera, E. LC-ESI-HRMS-based Chemical characterization of Lupinus bogotensis roots.Rev. Fac. Cienc. Basic. 2016, 12, 200–211. [CrossRef]
49. Wink, M.; Witte, L. Quinolizidine alkaloids in Genista acanthoclada and its holoparasite, Cuscuta palaestina. J. Chem. Ecol. 1993, 19,441–448. [CrossRef]
50. Kacem, N.; Goossens, J.-F.; Duhal, N.; Roumy, V.; Hennebelle, T.; Christen, P.; Hostettmann, K.; Rhouati, S. Determination ofalkaloids in endemic Genista quadriflora Munby (Fabaceae). Biochem. Syst. Ecol. 2014, 56, 83–87. [CrossRef]
51. Choma, I.; Jesionek, W. Effects-directed biological detection. In Instrumental Thin-Layer Chromatography; Elsevier: Amsterdam, TheNetherlands, 2015; pp. 279–312. ISBN 9780124172845.
52. Dewanjee, S.; Gangopadhyay, M.; Bhattacharya, N.; Khanra, R.; Dua, T.K. Bioautography and its scope in the field of naturalproduct chemistry. J. Pharm. Anal. 2015, 5, 75–84. [CrossRef] [PubMed]
53. Zamora-Natera, J.F.; Bernal-alcocer, A.; Ruiz-lópez, M.; Soto-hernández, M.; Vibrans-lindemann, H. Alkaloid profile of seeds fromLupinus exaltatus Zucc. (Fabaceae) and the antifungal evaluation of alkaloid extract and lupanin against phytopathogens. Rev.Mex. Fitopatol. 2005, 23, 124–129.
54. Kwasniewska, P.W.; Cofta, G.; Mazela, B.; Gobakken, L.R.; Przybył, A.K. Fungistatic activity of quinolizidine and bisquinolizidinealkaloids against A. niger. Int. Res. Gr. Wood Prot. 2016, 47, 1–9.
55. Yang, X.; Zhao, B. Antifungal activities of matrine and oxymatrine and their synergetic effects with chlorthalonil. J. For. Res. 2006,17, 323–325. [CrossRef]
56. Zamora-Natera, F.; García-López, P.; Ruiz-López, M.; Salcedo-Pérez, E. Composition of alkaloids in seeds of Lupinus mexicanus(Fabaceae) and antifungal and allelopathic evaluation of the alkaloid extract. Agrociencia 2008, 42, 185–192.
57. Pérez-Laínez, D.; García-Mateos, R.; San Miguel-Chávez, R.; Soto-Hernández, M.; Rodríguez-Pérez, E.; Kite, G. Bactericidal andfungicidal activities of Calia secundiflora (Ort.) Yakovlev. Z. Naturforsch. C 2008, 63, 653–657. [CrossRef]
58. Shao, J.; Wang, T.; Yan, Y.; Shi, G.; Cheng, H.; Wu, D.; Wang, C. Matrine reduces yeast-to-hypha transition and resistance of afluconazole-resistant strain of Candida albicans. J. Appl. Microbiol. 2014, 117, 618–626. [CrossRef]
59. Goto, T.; Hirazawa, N.; Takaishi, Y.; Kashiwada, Y. Antiparasitic effect of matrine and oxymatrine (quinolizidine alkaloids) on theciliate Cryptocaryon irritans in the red sea bream Pagrus major. Aquaculture 2015, 437, 339–343. [CrossRef]
60. He, H.; Qin, X.; Dong, F.; Ye, J.; Xu, C.; Zhang, H.; Liu, Z.; Lv, X.; Wu, Y.; Jiang, X.; et al. Synthesis, characterization of two matrinederivatives and their cytotoxic effect on Sf9 cell of Spodoptera frugiperda. Sci. Rep. 2020, 10, 17999. [CrossRef]
61. ur Rashid, H.; Xu, Y.; Muhammad, Y.; Wang, L.; Jiang, J. Research advances on anticancer activities of matrine and its derivatives:An updated overview. Eur. J. Med. Chem. 2019, 161, 205–238. [CrossRef]
62. Yang, Y.; Xiu, J.; Zhang, X.; Zhang, L.; Yan, K.; Qin, C.; Liu, J. Antiviral effect of matrine against human enterovirus 71. Molecules2012, 17, 10370–10376. [CrossRef] [PubMed]
63. Lin, Z.; Huang, C.F.; Liu, X.S.; Jiang, J. In vitro anti-tumour activities of quinolizidine alkaloids derived from Sophora flavescensAit. Basic Clin. Pharmacol. Toxicol. 2011, 108, 304–309. [CrossRef]
64. Liu, W.; Shi, J.; Zhu, L.; Dong, L.; Luo, F.; Zhao, M.; Wang, Y.; Hu, M.; Lu, L.; Liu, Z. Reductive metabolism of oxymatrine iscatalyzed by microsomal CYP3A4. Drug Des. Devel. Ther. 2015, 9, 5771–5783. [CrossRef] [PubMed]
65. Schrenk, D.; Bodin, L.; Chipman, J.K.; Del Mazo, J.; Grasl-Kraupp, B.; Hogstrand, C.; Hoogenboom, L.R.; Leblanc, J.-C.; Nebbia,C.S.; Nielsen, E.; et al. Scientific opinion on the risks for animal and human health related to the presence of quinolizidinealkaloids in feed and food, in particular in lupins and lupin-derived products. Eur. Food Saf. Auth. J. 2019, 17, e05860.
66. Green, B.T.; Lee, S.T.; Welch, K.D.; Cook, D. Anagyrine desensitization of peripheral nicotinic acetylcholine receptors. A potentialbiomarker of quinolizidine alkaloid teratogenesis in cattle. Res. Vet. Sci. 2017, 115, 195–200. [CrossRef] [PubMed]
67. Babushok, V.I. Chromatographic retention indices in identification of chemical compounds. TrAC—Trends Anal. Chem. 2015, 69,98–104. [CrossRef]
68. Marques, J.V.; Kitamura, R.O.S.; Lago, J.H.G.; Young, M.C.M.; Guimarães, E.F.; Kato, M.J. Antifungal amides from Piper scutifoliumand Piper hoffmanseggianum. J. Nat. Prod. 2007, 70, 2036–2039. [CrossRef] [PubMed]
69. Marentes-Culma, R.; Orduz-Díaz, L.L.; Coy-Barrera, E. Targeted metabolite profiling-based identification of antifungal 5-n-alkylresorcinols occurring in different cereals against Fusarium oxysporum. Molecules 2019, 24, 770. [CrossRef] [PubMed]
70. Cole, M.D. Key antifungal, antibacterial and anti-insect assays—A critical review. Biochem. Syst. Ecol. 1994, 22, 837–856. [CrossRef]71. Di Rienzo, J.A.; Casanoves, F.; Balzarini, M.G.; Gonzalez, L.; Tablada, M.; Robledo, C.W. InfoStat Version 24. Available online:
http://www.infostat.com.ar/ (accessed on 12 November 2021).




















