
Functional monoclonal antibodies to p75 neurotrophin receptor raised in knockout mice
Upload
independentCategory
view
0download
0
This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Qualitative and quantitative analysis of anthraquinone derivatives inrhizomes of tissue culture-raised Rheum emodi Wall. plants
Sonia Malik, Nandini Sharma, Upendra K. Sharma, Narendra P. Singh, Shashi Bhushan, Madhu Sharma n,Arun K. Sinha n, Paramvir S. Ahuja
Institute of Himalayan Bioresource Technology, Palampur 176061 (H. P.), India
a r t i c l e i n f o
Article history:
Received 17 July 2009
Received in revised form
17 December 2009
Accepted 17 December 2009
Keywords:
Anthraquinone
Plant tissue culture
Polygonaceae
Rheum emodi
Secondary metabolite
a b s t r a c t
This paper presents quantification of five anthraquinone derivatives (emodin glycoside, chrysophanol
glycoside, emodin, chrysophanol and physcion) in rhizomes of hardened micro-propagated Rheum
emodi plants using high-performance liquid chromatography (HPLC). Aseptic shoot cultures were raised
using rhizome buds. Shoot multiplication occurred in both agar gelled and liquid Murashige and Skoog
(MS) medium supplemented with 10.0 mM 6-benzylaminopurine (BAP) and 5.0 mM indole-3-butyric
acid (IBA). Rooted plantlets obtained on plant growth regulator (PGR)-free medium were transferred to
soil with 92% survival. HPLC analysis revealed the presence of five anthraquinone derivatives: emodin
glycoside, chrysophanol glycoside, emodin, chrysophanol and physcion in rhizomes of tissue culture-
raised plants. Only emodin glycoside (1) and chrysophanol glycoside (2) were present in 6-month-old
hardened tissue cultured plants. In addition, the other three derivatives (emodin (3), chrysophanol (4)
and physcion (5)) were also detected after 9 months.
& 2010 Elsevier GmbH. All rights reserved.
Introduction
Rheum emodi (Polygonaceae), a plant found in the westernHimalayas, is a source of anthraquinone derivatives, includingemodin, aloe emodin, physcion, chrysophanol, rhein, emodinglycoside and chrysophanol glycoside (Verma et al., 2005).Anthraquinone derivatives exhibit antifungal, anti-microbial,cytotoxic and antioxidant activities (Agarwal et al., 2000; Matsudaet al., 2001; Babu et al., 2004). In addition, these metabolites arealso used as dye in textile and food industries. Due to presence ofanthraquinone derivatives in its rhizomes, R. emodi has beenoverexploited and has become endangered in its natural habitat.Therefore, plant tissue culture technology offers an importantalternative for rapid multiplication and germplasm conservationof this rare and endangered plant species. In vitro plant multi-plication has an advantage over naturally occurring populations,as it offers a defined production system that ensures continuoussupply with uniform quality and yield (Fay, 1992; Zhao et al.,2004).
Lal and Ahuja (1989) standardized a micropropagation proto-col in order to conserve this important and rare medicinal plantspecies. Walkey used meristem tips for regeneration to obtainvirus-free Rheum rhaponticum plants (Walkey, 1968). Laterreports for in vitro multiplication and regeneration in the samespecies were provided by Walkey and Matthews (1979),Roggemans and Claes (1979) and Malik et al. (2009). In Polygonum
multiflorum, higher emodin and physcion content was recorded in3-month-old in vitro shoots as well as 6-month-old-tissueculture-raised hardened plants compared with field-grown plants(Lin et al., 2003). However, the qualitative and quantitative fatesof anthraquinone derivatives in tissue culture-raised plantsof R. emodi are still unknown. The present study was thereforecarried out to perform qualitative and quantitative analysis ofanthraquinone derivatives in tissue culture-raised plants.
Materials and methods
Plant material and micropropagation
Rheum emodi plants were collected from Chamba (westernHimalayas, altitude 2200 m above mean sea level) and main-tained under polyhouse conditions with an annual averagetemperature of 20–30 1C and relative humidity of 60–70%.Rhizome buds were used as explants for initiation of asepticcultures. Explants were thoroughly washed in running tap water
ARTICLE IN PRESS
Contents lists available at ScienceDirect
journal homepage: www.elsevier.de/jplph
Journal of Plant Physiology
0176-1617/$ - see front matter & 2010 Elsevier GmbH. All rights reserved.
doi:10.1016/j.jplph.2009.12.007
Abbreviations: BAP, 6-benzylaminopurine; HPLC, high performance liquid
chromatography; IBA, indole-3-butyric acid; MS, Murashige and Skoog (1962);
PGR, plant growth regulator; PPFD, photosynthetic photon flux densityn Corresponding authors. Tel.: +91 1894 233337; fax: +91 1894 230433.
E-mail addresses: [email protected] (M. Sharma),
[email protected] (A.K. Sinha).
Journal of Plant Physiology 167 (2010) 749–756

Author's personal copyARTICLE IN PRESS
followed by distilled water containing 1–2 drops of Tween-20(HiMedia) using a sable hairbrush. Buds were then surfacesterilized with 0.5% (w/v) bavistin (BASF) and 0.5% (w/v)streptomycin sulphate (Sigma) for 8–10 min, followed by a dipin 70% ethanol. The explants were subsequently surface sterilizedwith 0.1% (w/v) HgCl2 (Qualigens) having 1–2 drops of Tween-20 as wetting agent for 9–10 min, followed by 4–5 washings withsterile distilled water. These were then inoculated vertically in a250 mL Erlenmeyer flask containing 100 mL Murashige and Skoog(1962) (MS) medium with sucrose (3%, w/v; Qualigens) and agar(0.8%, w/v; Qualigens). MS medium was supplemented with6-benzylaminopurine (BAP) (5.0 and 10.0 mM; Sigma) and indole-3-butyric acid (IBA) (0.5 and 5.0 mM; Sigma). Plant growthregulator (PGR)-free MS medium served as control. To avoid theproblem of phenolic exudation in the medium, ascorbic acid(100 mg/L) was added. The pH of the medium was adjusted to 5.8prior to autoclaving at 15 psi and 121 1C for 20 min.
We examined the effects of agar gelled, liquid static and liquidshake culture medium on shoot multiplication. For liquid staticand shake cultures, 20 and 30 mL liquid media, respectively, wereused in 250 mL Erlenmeyer flasks. For liquid shake cultures, flaskswere kept on a rotary shaker at 100 rpm. The multiple shootsobtained were transferred to PGR-free agar gelled medium forrooting. The cultures were maintained in bright light (photosyn-thetic photon flux density (PPFD), 70 mmol m�1 s�2) at 2572 1Cwith a 16 h photoperiod. Five replicates were used for eachexperiment and the experiment was repeated in triplicate. Rootedplantlets were transferred to pots containing a sand and soilmixture (1:1 ratio) under greenhouse conditions. After duehardening, these were shifted to pots containing soil.
Extraction and isolation of anthraquinone derivatives
Rhizomes collected from the natural habitat were used forisolation of standard anthraquinone derivatives. The dried andpowdered rhizomes (225 g) were extracted with three differentsolvents (3�500 mL each with hexane, chloroform and ethylacetate) using Soxhlet extractions. The extracts were filtered andsolvents were removed under reduced pressure. Concentratedextracts obtained from different solvents were subjected tocolumn chromatography over silica gel using the gradient elutionmethod. Ethyl acetate extracts upon column purification withmethanol–chloroform (20:80, v/v) yielded emodin glycoside(1) and chrysophanol glycoside (2). Chloroform extract uponcolumn purification with ethyl acetate-hexane (30:70, v/v)yielded emodin (3), whereas hexane extract upon columnpurification with ethyl acetate-hexane (5:95, v/v) yielded chry-sophanol (4) and physcion (5) (Singh et al., 2005). 1H- (300 MHz)and 13C NMR (75.4 MHz) data for compounds (1–5) were recordedon a Bruker Avance 300 spectrometer. The structures of all
Fig. 1. Shoot cultures of R. emodi in (a) agar gelled, (b) liquid static and (c) liquid shake flasks on MS medium supplemented with 10.0 mM BAP and 5.0 mM IBA.
Table 1Effect of status of medium on shoot multiplication and leaf size on MS medium
with BAP (10.0 mM) and IBA (5.0 mM).
Status of medium Fold multiplication Leaf area (cm2)
Agar gelled 6.6570.18 0.1–2.0
Liquid (static) 4.6670.13 2.0–3.0
Liquid (shake culture) 8.470.08 2.0–3.0
Data recorded after 4 weeks; no. of replicates: 5.
S. Malik et al. / Journal of Plant Physiology 167 (2010) 749–756750

Author's personal copyARTICLE IN PRESS
Fig. 2. Plantlets with a well-developed root system.
S. Malik et al. / Journal of Plant Physiology 167 (2010) 749–756 751
Author's personal copyARTICLE IN PRESS
Fig. 3. Hardened plants in greenhouse conditions.
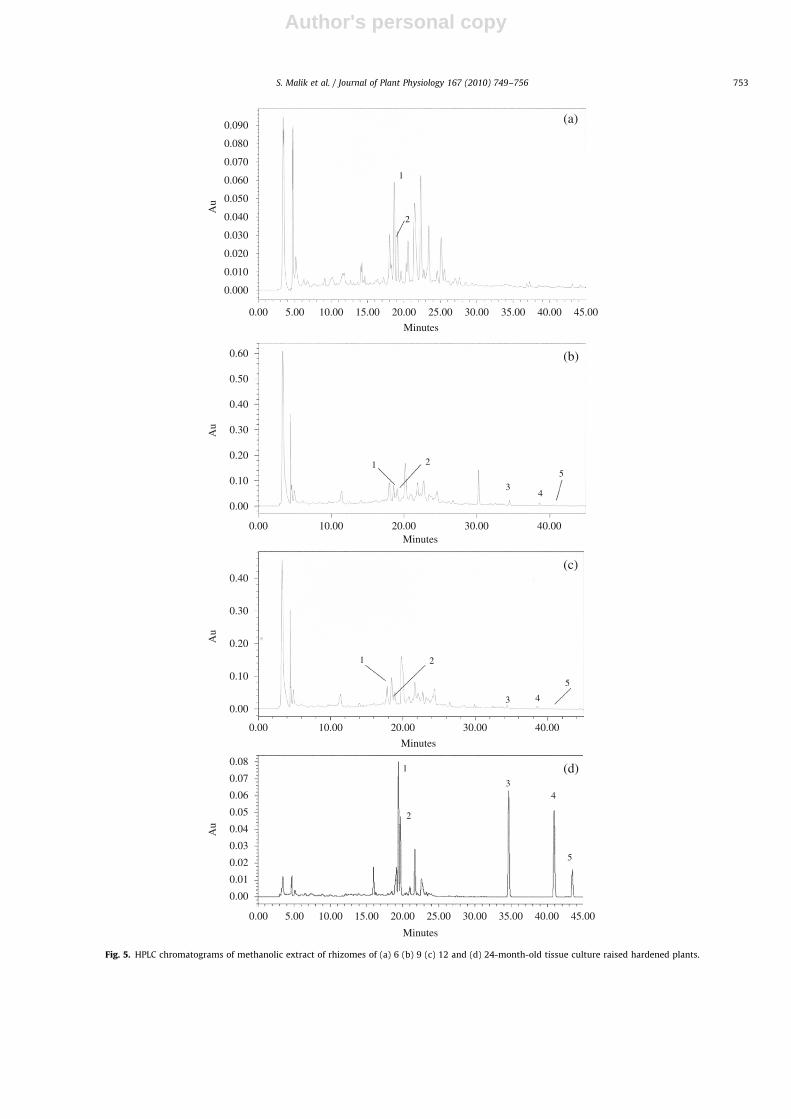
Fig. 4. HPLC chromatogram of standard emodin glycoside (1), chrysophanol glycoside (2), emodin (3), chrysophanol (4) and physcion (5).
S. Malik et al. / Journal of Plant Physiology 167 (2010) 749–756752
Author's personal copyARTICLE IN PRESS
0.090
0.080
0.070
0.060
0.050
0.040
0.030
0.020
0.010
0.000
0.60
0.50
0.40
0.30
0.20
0.10
0.00
Au
0.08
0.07
0.06
0.05
0.04
0.03
0.02
0.01
0.00
Au
Au
0.40
0.30
0.20
0.10
0.00
Au
0.00 5.00 10.00 15.00 20.00 25.00 30.00 35.00 40.00 45.00
1
2
34
5
Minutes
0.00 10.00 20.00 30.00 40.00
Minutes
1 2
3 4
5
0.00 10.00 20.00 30.00 40.00Minutes
1 2
34
5
0.00 5.00 10.00 15.00 20.00 25.00 30.00 35.00 40.00 45.00
Minutes
1
2
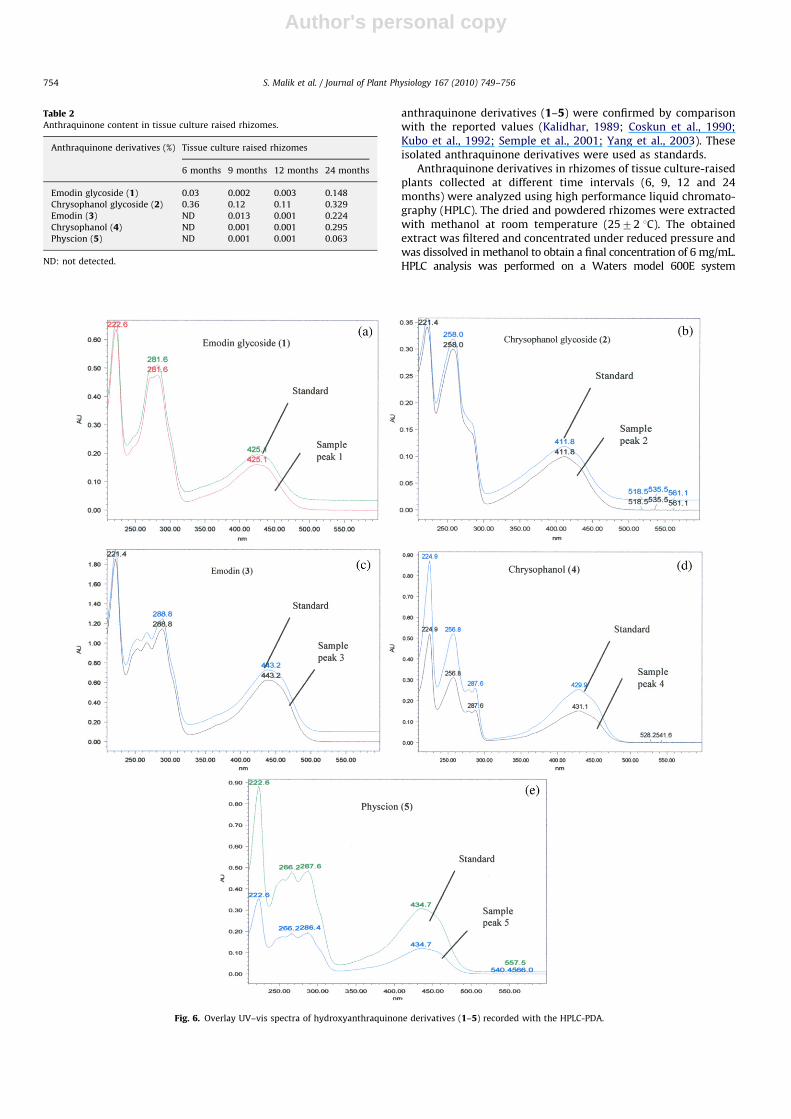
Fig. 5. HPLC chromatograms of methanolic extract of rhizomes of (a) 6 (b) 9 (c) 12 and (d) 24-month-old tissue culture raised hardened plants.
S. Malik et al. / Journal of Plant Physiology 167 (2010) 749–756 753
Author's personal copyARTICLE IN PRESS
anthraquinone derivatives (1–5) were confirmed by comparisonwith the reported values (Kalidhar, 1989; Coskun et al., 1990;Kubo et al., 1992; Semple et al., 2001; Yang et al., 2003). Theseisolated anthraquinone derivatives were used as standards.
Anthraquinone derivatives in rhizomes of tissue culture-raisedplants collected at different time intervals (6, 9, 12 and 24months) were analyzed using high performance liquid chromato-graphy (HPLC). The dried and powdered rhizomes were extractedwith methanol at room temperature (2572 1C). The obtainedextract was filtered and concentrated under reduced pressure andwas dissolved in methanol to obtain a final concentration of 6 mg/mL.HPLC analysis was performed on a Waters model 600E system
Table 2Anthraquinone content in tissue culture raised rhizomes.
Anthraquinone derivatives (%) Tissue culture raised rhizomes
6 months 9 months 12 months 24 months
Emodin glycoside (1) 0.03 0.002 0.003 0.148
Chrysophanol glycoside (2) 0.36 0.12 0.11 0.329
Emodin (3) ND 0.013 0.001 0.224
Chrysophanol (4) ND 0.001 0.001 0.295
Physcion (5) ND 0.001 0.001 0.063
ND: not detected.
Fig. 6. Overlay UV–vis spectra of hydroxyanthraquinone derivatives (1–5) recorded with the HPLC-PDA.
S. Malik et al. / Journal of Plant Physiology 167 (2010) 749–756754

Author's personal copyARTICLE IN PRESS
(Waters, Milford, MA, USA) equipped with a Purosphers-Star RP-18ecolumn (4.6 mm i.d.�250 mm, 5 mm, Merck, Darmstadt, Germany), aphoto-diode array detector (PDA) (Waters 2996), an inline-degasserAF (Waters), a 20 mL loop manual injector and Waters empowersoftware. We used acetonitrile: methanol (95:5, v/v) (solvent A) andwater: acetic acid (99.9:0.1, v/v, pH 3.5) (solvent B) as the mobilephase with a linear gradient elution as described by Verma et al.(2005). The detection wavelength was set at 290 nm. The stocksolution (1 mg/mL) of each standard anthraquinone derivative (1–5)was freshly prepared in a mixture of methanol: acetonitrile (80:20,v/v), and desired concentrations were obtained by serial dilution forstandard curve preparation. The calibration graphs were plotted afterlinear regression of the peak area versus concentration.
Results and discussion
Shoot multiplication
Rhizome buds sprouted after 20–25 d of inoculation in all themedia combinations used. These sprouted buds gave rise toshoots after 2 weeks, upon transfer to fresh medium. According toour previous results, Murashige and Skoog (MS) medium with10.0 mM 6-benzylaminopurine (BAP) and 5.0 mM indole-3-butyricacid (IBA) yields the maximum shoot multiplication rate (Maliket al., 2009). This, medium was used for further experiments forshoot proliferation.
The status of the medium varied the shoot proliferationresponse. There was an 8.4-fold increase in shoot number onmedium with BAP (10.0 mM) and IBA (5.0 mM) in liquid shakecultures, followed by agar gelled (6.65-fold) and liquid staticcultures (4.66-fold) (Fig. 1, Table 1). On the other hand, in plantgrowth regulator (PGR)-free medium (control), we observed onlyelongation of shoots with occasional formation of 1–2 new shoots.Multiple shoots obtained after 30 d on medium with BAP and IBAwere transferred to PGR-free medium (liquid static, shake flaskand agar gelled) for elongation. The shoots showed a maximumlength (6.2 cm) in shake cultures, followed by liquid staticcultures (4.5 cm) and agar gelled (3.4 cm) in PGR-free medium.Lal and Ahuja (1993) also observed high shoot multiplication inliquid shake cultures compared with liquid static and agar gelledcultures in R. emodi. Large leaves in liquid medium could beattributed to the fact that more surface area of the tissue is incontact with the medium for uptake of nutrients and PGRs,resulting in greater shoot growth and faster multiplication (Liuet al., 2001). Moreover, there is better exchange of atmosphericgases, such as O2 and CO2, in liquid shake cultures due to constantagitation of medium (Biondi and Thorpe, 1981). Low shootmultiplication and growth in liquid static cultures comparedwith liquid shake flasks could be due to asphyxia (Alvard et al.,1993; Mehrotra et al., 2007).
Rooting and hardening
Root initiation from in vitro shoots was observed after 30–35 dof culture on PGR-free medium, and plants with well developedroots were obtained after 40–45 d (Fig. 2a, b). Rooted plantletswere transferred to pots and hardened under greenhouseconditions (Fig. 3). Plants showed 92% survival and rhizomedevelopment started after 6 months.
Analysis of anthraquinone derivatives
All of the isolated anthraquinone derivatives (1–5) wereanalyzed under optimized high performance liquid chromatography
(HPLC) conditions (Verma et al., 2005) and were well separated fromeach other with retention times of 18.8 min (emodin glycoside),19.3 min (chrysophanol glycoside), 34.5 min (emodin), 38.8 min(chrysophanol) and 40.7 min (physcion) (Fig. 4).
Temporal analysis revealed qualitative as well as quantitativevariations in anthraquinone derivatives in the rhizomes of tissueculture-raised plants. HPLC analysis of extracts from rhizomes of6-month-old-plants showed the presence of only emodin glyco-side (1) and chrysophanol glycoside (2) (Fig. 5a), thus indicatingtheir early synthesis compared with the other derivatives. In9- and 12-month-old rhizomes, all five anthraquinone derivativeswere detected, but the amounts of emodin glycoside (1) andchrysophanol glycoside (2) were lower than in the 6-month-oldsample (Fig. 5b, c). In extracts of 24-month-old rhizomes, theamounts of emodin (3), chrysophanol (4) and physcion (5) weremuch higher than in the earlier stages (Fig. 5d, Table 2). Thestructures of all the anthraquinone derivatives (1–5) wereconfirmed by spiking and comparison of their UV–vis spectrawith reference standards (Fig. 6) to clarify the observeddiscrepancy in retention times (see Figs. 4 and 5d). From theseresults, we concluded that the production of emodin glycoside (1)and chrysophanol glycoside (2) started prior to other threeanthraquinone derivatives (3–5). It is well known thatsecondary compounds toxic to plant cells are stored in theirglycoside form (Wink, 1987; Veeresham, 2004). In R. emodi, theseprobably act as precursors to their conversion to active formsduring stress or adverse environmental conditions.
Conclusion
The present study provides insights into an optimized harvestschedule of different anthraquinone derivatives in R. emodi. Thedata show that there is a correlation between synthesis of thesesecondary compounds and age of the plant. Anthraquinoneglycosides, which are reported to have pharmacological impor-tance (cf. Singh et al., 2004), were detected in 6-month-oldrhizomes, while other derivatives were formed at later stages ofdevelopment. These latter forms are used in traditional Japaneseand Chinese systems of medicine. Therefore, tissue culture-raisedplants can be used for the extraction of these derivatives asrequired. Based on the results of the present study, an effectivestrategy can be developed not only for this endangered plantspecies, but also for the production of an important class ofmedicinally important anthraquinones.
Acknowledgement
Financial support from Department of Biotechnology, NewDelhi, India is gratefully acknowledged.
References
Agarwal SK, Singh SS, Verma S, Kumar S. Antifungal activity of anthraquinonederivatives from Rheum emodi. J Ethnopharmacol 2000;72:43–6.
Alvard D, Cote F, Teisson C. Comparison of methods of liquid medium culture forbanana micropropagation: effects of temporary immersion of explants. PlantCell Tissue Org Cult 1993;32:55–60.
Babu KS, Tiwari AK, Srinivas PV, Ali AZ, Raju BN, Rao M. Yeast and mammaliana-glucosidase inhibitory constituents from Himalayan Rheum emodi Wall. exMeisson. Bioorg Med Chem Lett 2004;14:3841–5.
Biondi S, Thorpe TA. Requirements for a tissue culture facility. In: Thorpe TA,editor. Plant tissue culture: methods and applications in agriculture. NewYork: Academic Press; 1981. p. 1–20.
Coskun M, Satake T, Hori K, Saiki Y, Tanker M. Anthraquinone glycosides fromRhamnus libanoticus. Phytochemistry 1990;29:2018–20.
Fay MF. Conservation of rare and endangered plants using in vitro methods.In Vitro Cell Dev Biol Pl 1992;28:1–4.
S. Malik et al. / Journal of Plant Physiology 167 (2010) 749–756 755

Author's personal copyARTICLE IN PRESS
Kalidhar SB. Location of glycosylation and alkylation sites in anthraquinones by1H NMR. Phytochemistry 1989;28:2455–8.
Kubo I, Murai Y, Soediro I, Soeetarno S, Sastrodihardjo S. Cytotoxic anthraquinonesfrom Rheum palmatum. Phytochemistry 1992;31:1063–5.
Lal N, Ahuja PS. Propagation of Indian Rhubarb (Rheum emodi Wall.) using shoot-tip and leaf explant culture. Plant Cell Rep 1989;8:493–6.
Lal N, Ahuja PS. Assessment of liquid culture procedures for in vitro propagation ofRheum emodi. Plant Cell Tissue Org Cult 1993;34:223–36.
Lin LC, Nalawade SM, Mulabagal V, Yeh MS, Tsay HS. Micropropagation ofPolygonum multiflorum THUNB and quantitative analysis of the anthraqui-nones emodin and physcion formed in in vitro propagated shoots and plants.Biol Pharm Bull 2003;26:1467–71.
Liu CZ, Moon KH, Honda H, Kobayashi T. In situ regeneration of rice (Oryza sativa L.)callus immobilized in polyurethane foam. J Biosci Bioeng 2001;91:76–80.
Malik S, Kumar R, Vats SK, Bhushan S, Sharma M, Ahuja PS. Regeneration in Rheumemodi Wall: a step towards conservation of an endangered medicinal plantspecies. Eng Life Sci 2009;9:130–4.
Matsuda H, Morikawa T, Toguchida I, Park JY, Harima S, Yoshikawa M. Antioxidantconstituents from Rhubarb. Structural requirements of stilbenes for theactivity and structures of two new anthraquinone glucosides. Bioorg MedChem 2001;9:41–50.
Mehrotra S, Goel MK, Kukreja AK, Mishra BN. Efficiency of liquid culture systemsover conventional micropropagation: a progress towards commercialization.Afr J Biotechnol 2007;6:1484–92.
Murashige T, Skoog F. A revised medium for rapid growth and bioassays withtobacco tissue cultures. Physiol Plant 1962;15:473–97.
Roggemans J, Claes MC. Rapid clonal propagation of Rhubarb by in vitro culture ofshoot tips. Sci Hortic 1979;11:241–6.
Semple SJ, Pyke SM, Reynolds GD. Flower RLP. In vitro antiviral activity of theanthraquinone chrysophanic acid against poliovirus. Antiviral Res 2001;49:169–78.
Singh R, Geetanjali, Chauhan SMS. 9,10-Anthraquinones and other biologicallyactive compounds from the genus Rubia. Chem Biodiversity 2004;1:1241–63.
Singh NP, Gupta AP, Sinha AK, Ahuja PS. High-performance thin layer chromato-graphy method for the quantitative determination of four major anthraqui-none derivatives in Rheum emodi. J Chromatogr A 2005;1077:202–6.
Veeresham C. Secondary metabolic products (phytopharmaceuticals) from plantcell cultures. Medicinal plant biotechnology. 1st ed.. New Delhi: CBS Publish-ers and Distributors; 2004. p. 49–120.
Verma SC, Singh NP, Sinha AK. Determination and locational variation in the quantityof hydroxyanthraquinones and their glycosides in rhizomes of Rheum emodi usinghigh-performance liquid chromatography. J Chromatogr A 2005;1097:59–65.
Walkey DGA. The production of virus free rhubarb by apical tip-culture. J Hortic Sci1968;43:283–7.
Walkey DGA. Matthews KAM. Rapid clonal propagation of rhubarb (Rheumrhaponticum) from meristem-tips in tissue culture. Plant Sci Lett 1979;14:287–9.
Wink M. Physiology of the accumulation of secondary metabolites with specialreference to alkaloids. In: Constable F, Vasil I, editors. Cell culture and somaticcell genetics of plants. San Diego: Academic Press; 1987. p. 255–66.
Yang YC, Lim MY, Lee HS. Emodin isolated from Cassia obtusifolia (Leguminosae)seed shows larvicidal activity against three mosquito species. J Agric FoodChem 2003;51:7629–31.
Zhao P, Iwamoto Y, Kouno I, Egami Y, Yamamoto H. Stimulating the production ofhomoisoflavonoids in cell suspension cultures of Caesalpinia pulcherrima usingcork tissue. Phytochemistry 2004;65:2455–61.
S. Malik et al. / Journal of Plant Physiology 167 (2010) 749–756756
Copyright © 2022 FDOKUMEN




















