
Purification and characterisation of extracellular protease produced by Clostridium sp. from...
9
ELSEVIER Purification and characterization of an extracellular P-glucosidase from the filamentous fungus Acremonium persicinum and its probable role in P-glucan degradation Stuart M. Pitson, Robert J. Seviour, and Barbara M. McDougall Biotechnology Research Centre, TaTrobe University, Bend&o, Victoria, Australia A P-glucosidase from the culture filtrates of the filamentous fungus Acremonium persicinum has been purified by (NH4)J04 precipitation followed by anion-exchange and gel filtration chromatography. SDS-PAGE of the purified enzyme gave a single band with an apparent molecular mass of 128 kDa. The enzyme is a monomeric protein with an isoelectric point of 4.3 and a pH optimum of 5.5. Comparison of the N-terminal amino acid sequence revealed similarities between the A. persicinum enzyme and several other extracellular fungal P-glucosidases including those from Trichoderma reesei, Aspergillus aculeatus, Saccharomycopsis fibuligera, and Pichia anomala. In addition to the hydrolysis of p-nitrophenyl-/3-glucoside, the enzyme was also active against several other aryl-/3-glucosides as well as a range of P-linked oligoglucosides including laminaribiose, gentiobiose, cellobiose, and sophorose. D-Ghtcono-l,S-lactone and glucose are competitive inhibitors while the enzyme was also inhibited by r+bromosuccinimide, N-acetylimidazole, dicyclohexyl carbodiimide, Woodward’s Reagent K, 2-hydroxy-5-nitrobenzyl bromide, KMnO,, and some metal ions. Possible roles for this enzyme in the noncellulolytic fungus A. persicinum are discussed in light of the increase in the r-ate of reducing sugar release from P-glucans by (I + 3)- and (1 + 6)-P-glucanases when the P-glucosidase is also present in the reaction mixtures. 0 1997 Elsevier Science Inc. Keywords: P-Glucosidase; Acremonium persicinum: B-glucan hydrolysis Introduction P-Glucosidases (P-o-glucoside glucohydrolases, EC 3.2.1.21) can catalyze the hydrolysis of the P-glucosidic linkages of aryl and alkyl P-glucosides, P-linked oligoglu- cosides, and several other oligosaccharides with the release of glucose usually in the p configuration.’ The broad group of enzymes termed P-glucosidases display a diversity of substrate specificities apparently dependant on both enzyme source and perhaps more importantly, their role in the Address reprint requests to Dr. R. J. Seviour, Biotechnology Research Centre, La Trobe University, Bendigo, P.O. Box 199, Bendigo 3550, Victoria, Australia The present address of Dr. S. M. Pitson is the School of Biochemistry and Molecular Genetics, The University of New South Wales, Sydney 2052, Australia Received 16 May 1996; revised 21 November 1996; accepted 27 Novem- ber 1996 producing cell. 2,3 Some P-glucosidases are quite specific for aryl or alkyl j3-glucosides,4-6 while others show preference for short-chain P-linked oIigoglucosides;5*7 however, most commonly seen in fungi are the P-glucosidases with much broader specificity which act equally well on both p- oligoglucosides and aryl and alkyl P-glucosides.‘,3,8 Re- cently, a P-glucosidase isolated from Botrytis cinerae has even been reported to have the surprising ability to hydro- lyze both p- and cx-glucosides.’ While not strictly recognized as P-glucan hydrolytic enzymes, P-glucosidases have been implicated in P-glucan degradation through synergistic action with P-glucanases. For example, degradation of cellulose by the combined action of the various enzymes of the cellulase complex, including P-glucosidases, is well established;“-” however, surprisingly few studies have focused on their involvement in the degradation of other p-glucans. Stahmann et a1.‘4,‘5 did suggest that an extracellular (1 + 3)(1 -+ 6)-P-glucan from B. cinerea was degraded by this organism through Enzyme and Microbial Technology 21:182-190, 1997 0 1997 Elsevier Science Inc. All rights reserved. 655 Avenue of the Americas, New York, NY 10010 0141-0229/97/$17.00 PII SO141-0229(96)00263-3
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Purification and characterisation of extracellular protease produced by Clostridium sp. from...

ELSEVIER
Purification and characterization of an extracellular P-glucosidase from the filamentous fungus Acremonium persicinum and its probable role in P-glucan degradation Stuart M. Pitson, Robert J. Seviour, and Barbara M. McDougall
Biotechnology Research Centre, TaTrobe University, Bend&o, Victoria, Australia
A P-glucosidase from the culture filtrates of the filamentous fungus Acremonium persicinum has been purified by (NH4)J04 precipitation followed by anion-exchange and gel filtration chromatography. SDS-PAGE of the purified enzyme gave a single band with an apparent molecular mass of 128 kDa. The enzyme is a monomeric protein with an isoelectric point of 4.3 and a pH optimum of 5.5. Comparison of the N-terminal amino acid sequence revealed similarities between the A. persicinum enzyme and several other extracellular fungal P-glucosidases including those from Trichoderma reesei, Aspergillus aculeatus, Saccharomycopsis fibuligera, and Pichia anomala. In addition to the hydrolysis of p-nitrophenyl-/3-glucoside, the enzyme was also active against several other aryl-/3-glucosides as well as a range of P-linked oligoglucosides including laminaribiose, gentiobiose, cellobiose, and sophorose. D-Ghtcono-l,S-lactone and glucose are competitive inhibitors while the enzyme was also inhibited by r+bromosuccinimide, N-acetylimidazole, dicyclohexyl carbodiimide, Woodward’s Reagent K, 2-hydroxy-5-nitrobenzyl bromide, KMnO,, and some metal ions. Possible roles for this enzyme in the noncellulolytic fungus A. persicinum are discussed in light of the increase in the r-ate of reducing sugar release from P-glucans by (I + 3)- and (1 + 6)-P-glucanases when the P-glucosidase is also present in the reaction mixtures. 0 1997 Elsevier Science Inc.
Keywords: P-Glucosidase; Acremonium persicinum: B-glucan hydrolysis
Introduction
P-Glucosidases (P-o-glucoside glucohydrolases, EC 3.2.1.21) can catalyze the hydrolysis of the P-glucosidic linkages of aryl and alkyl P-glucosides, P-linked oligoglu- cosides, and several other oligosaccharides with the release of glucose usually in the p configuration.’ The broad group of enzymes termed P-glucosidases display a diversity of substrate specificities apparently dependant on both enzyme source and perhaps more importantly, their role in the
Address reprint requests to Dr. R. J. Seviour, Biotechnology Research Centre, La Trobe University, Bendigo, P.O. Box 199, Bendigo 3550, Victoria, Australia The present address of Dr. S. M. Pitson is the School of Biochemistry and Molecular Genetics, The University of New South Wales, Sydney 2052, Australia Received 16 May 1996; revised 21 November 1996; accepted 27 Novem- ber 1996
producing cell. 2,3 Some P-glucosidases are quite specific for aryl or alkyl j3-glucosides,4-6 while others show preference for short-chain P-linked oIigoglucosides;5*7 however, most commonly seen in fungi are the P-glucosidases with much broader specificity which act equally well on both p- oligoglucosides and aryl and alkyl P-glucosides.‘,3,8 Re- cently, a P-glucosidase isolated from Botrytis cinerae has even been reported to have the surprising ability to hydro- lyze both p- and cx-glucosides.’
While not strictly recognized as P-glucan hydrolytic enzymes, P-glucosidases have been implicated in P-glucan degradation through synergistic action with P-glucanases. For example, degradation of cellulose by the combined action of the various enzymes of the cellulase complex, including P-glucosidases, is well established;“-” however, surprisingly few studies have focused on their involvement in the degradation of other p-glucans. Stahmann et a1.‘4,‘5 did suggest that an extracellular (1 + 3)(1 -+ 6)-P-glucan from B. cinerea was degraded by this organism through
Enzyme and Microbial Technology 21:182-190, 1997 0 1997 Elsevier Science Inc. All rights reserved. 655 Avenue of the Americas, New York, NY 10010
0141-0229/97/$17.00 PII SO141-0229(96)00263-3
A. persicinum p-glucosidase: S. Pitson et al.
the combined action of a P-glucosidase and several exo-( 1 += 3)-P-glucanases. Similar results have been re- ported for CZavicepsfisiformis’6 and scleroglucan degrada- tion by Sclerotium glucanicum.‘7~‘8 Similar synergistic actions have been proposed to account for degradation of chemically complex fungal walls during cell autolysis and growth. ‘9,2o Endo-P-glucanases are considered responsible for partial wall hydrolysis. They release oligoglucosides from long-chain P-glucans which are then converted to glucose for cell metabolism by either P-glucosidases or exo-P-glucanases. *‘,** This interactive process has not been studied in detail, although Hiura et uL.*~-*~ proposed a mechanism whereby isolated Neurosporu crussu cell wall (1 + 3)( 1 + 6)-p-glucan is degraded by the cooperative involvement of a (1 -+ 6)-P-glucanase, exo-( 1 + 3)-p- glucanase, endo-( 1 + 3)-P-glucanase, and a P-glucosidase.
The filamentous fungus Acremonium persicinum se- cretes an extracellular P-glucan under nitrogen-limiting conditions. 26 Carbon source exhaustion results in the ap- pearance of (1 -+ 3)+glucanase, (1 + 6)-P-glucanase, and P-glucosidase activity in the culture filtrates of A. persici- num27.28 and a subsequent decrease in p-glucan levels. Two recent studies29*30 have described the purification and char- acterization of an endo-( 1 -+ 6)+Lglucanase and three extracellular (1 -+ 3)-P-glucanases from this fungus. This work reports the purification and characterization of a P-glucosidase secreted by A. persicinum and examines its probable role in the degradation of extracellular and cell wall polysaccharides.
Materials and methods
Organism and culture conditions
Growth of A. persicinum Nicot and Gams QM107a in liquid media and the collection of culture filtrates for enzyme purification were as described earlier.29
@Glucosidase assay
Routine P-glucosidase activities were determined in duplicate using 2 mM 4-nitrophenyl-P-n-glucopyranoside @NPG) as sub- strate. Assays were performed in 50 mM sodium acetate buffer pH 5.5 and incubated at 40°C for 30 min after which time NH,OH was added to a final concentration of 1 M. The amount of 4-nitrophenol (pNP) released was estimated by absorbance at 400 nm32 using a pNP standard curve. One unit (U) of B-glucosidase activity is defined as one pmol of pNP released min-‘. Activities against other 4-nitrophenyl derivatives were measured in the same way while activities on other substrates were determined by the release of reducing sugars.‘3.‘4
Purification of the P-glucosidase
Purification of the B-glucosidase from A. persicinum culture filtrates concentrated by ultrafiltration29 was achieved using (NH&SO, fractionation followed by FPLC chromatography with anion-exchange (Mono Q) and gel filtration (Superose 12) col- umns using the same separation protocols described earlier for purification of the extracellular (1 -+ 3)- and (1 -+ 6)-B-glucanases of A. persicinum.‘9~‘0
0 48 96 144 192 240
Time (h)
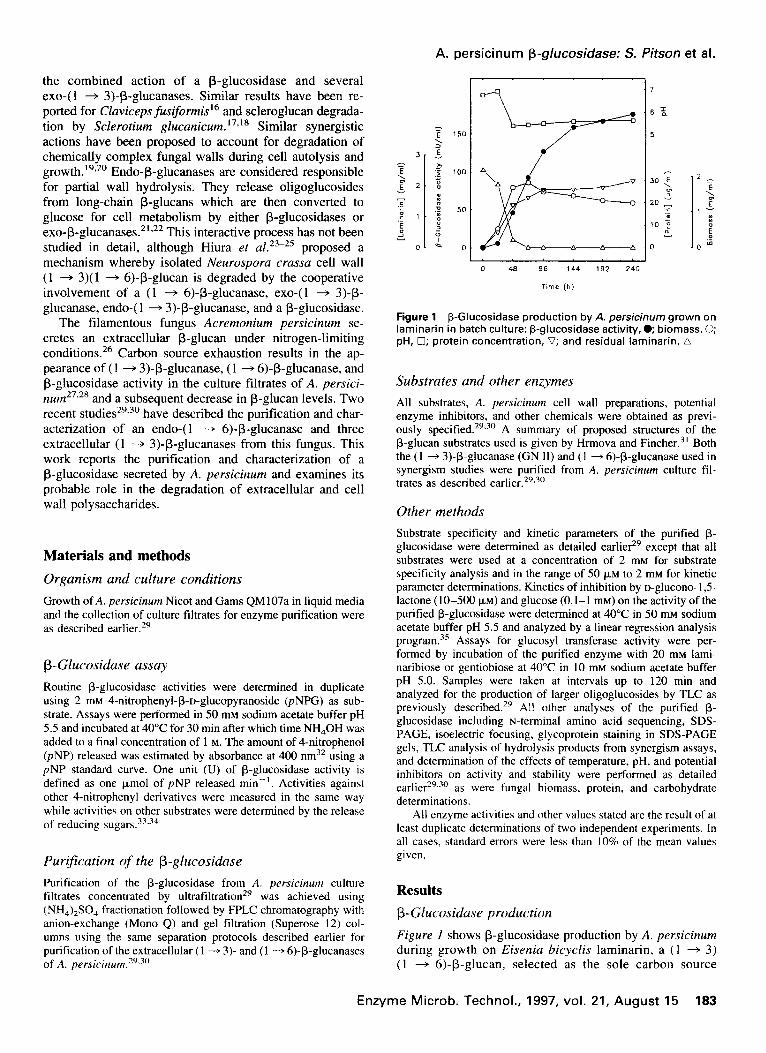
Figure 1 P-Glucosidase production by A. persicinum grown on laminarin in batch culture: p-glucosidase activity, e biomass, 0; pH, 0; protein concentration, V; and residual laminarin, A
Substrates and other enzymes
All substrates, A. persicinum cell wall preparations, potential enzyme inhibitors, and other chemicals were obtained as previ- ously specified. 29*30 A summary of proposed structures of the B-glucan substrates used is given by Hrmova and Fincher.3’ Both the (1 + 3)-B-glucanase (GN II) and ( 1 + 6)-B-glucanase used in synergism studies were purified from A. persicinum culture fil- trates as described earlier.29.3”
Other methods
Substrate specificity and kinetic parameters of the purified B- glucosidase were determined as detailed earliera except that all substrates were used at a concentration of 2 mM for substrate specificity analysis and in the range of 50 FM to 2 mM for kinetic parameter determinations. Kinetics of inhibition by D-glucono- I,5 lactone (lo-500 FM) and glucose (0. l-l mM) on the activity of the purified R-glucosidase were determined at 40°C in 50 ITIM sodium acetate buffer pH 5.5 and analyzed by a linear regression analysis program.‘s Assays for glucosyl transferase activity were per- formed by incubation of the purified enzyme with 20 mM lami- naribiose or gentiobiose at 40°C in 10 mM sodium acetate buffer pH 5.0. Samples were taken at intervals up to 120 min and analyzed for the production of larger oligoglucosides by TLC as previously described.29 All other analyses of the purified B- glucosidase including N-terminal amino acid sequencing, SDS- PAGE, isoelectric focusing, glycoprotein staining in SDS-PAGE gels, TLC analysis of hydrolysis products from synergism assays, and determination of the effects of temperature, pH, and potential inhibitors on activity and stability were performed as detailed earlier29.a0 as were fungal biomass, protein, and carbohydrate determinations.
All enzyme activities and other values stated are the result of at least duplicate determinations of two independent experiments. In all cases, standard errors were less than 10% of the mean values given.
Results
P-Glucosidase production
Figure I shows P-glucosidase production by A. persicinum during growth on Eisenia bicyclis laminarin, a ( 1 + 3) (1 + 6)-P-glucan, selected as the sole carbon source
Enzyme Microb. Technol., 1997, vol. 21, August 15 183
Papers
94 -
67 -
43 -
30 -
20.1 -
14.4 -
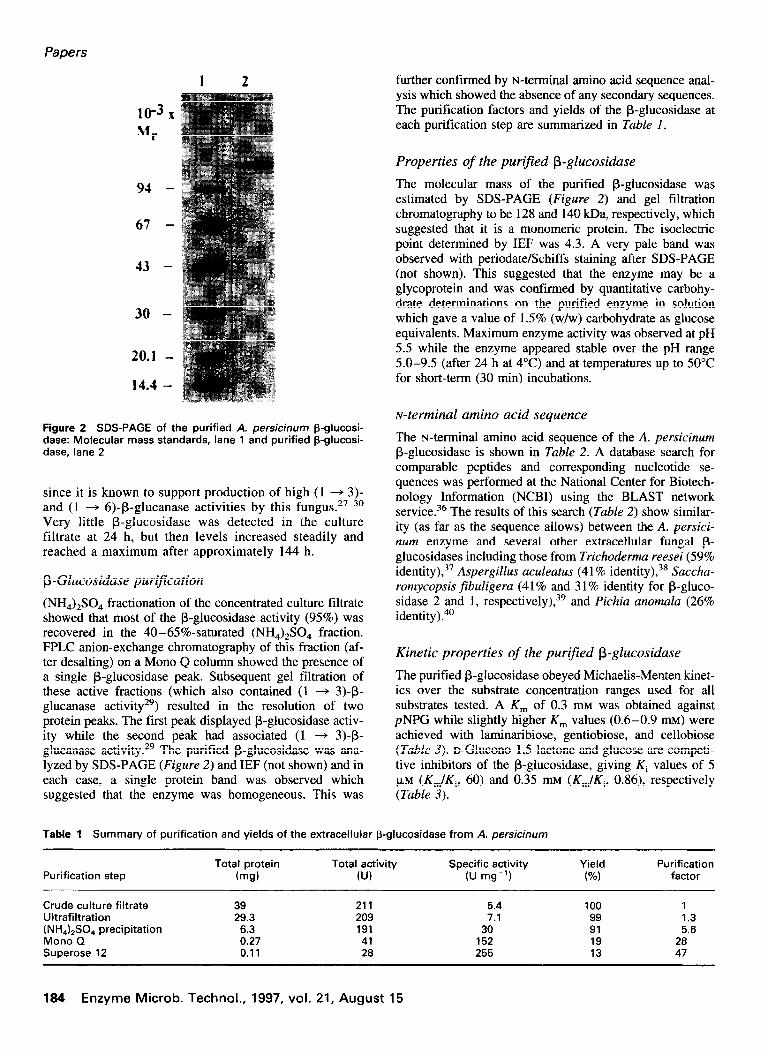
Figure 2 SDS-PAGE of the purified A. persicinum p-glucosi- dase: Molecular mass standards, lane 1 and purified p-glucosi- dase. lane 2
since it is known to support production of high (1 + 3)- and (1 + 6)-P-glucanase activities by this fungus.27-30 Very little B-glucosidase was detected in the culture filtrate at 24 h, but then levels increased steadily and reached a maximum after approximately 144 h.
@Glucosidase purification
(NH,),SO, fractionation of the concentrated culture filtrate showed that most of the P-glucosidase activity (95%) was recovered in the 40-65%-saturated (NH,),SO, fraction. FPLC anion-exchange chromatography of this fraction (af- ter desalting) on a Mono Q column showed the presence of a single &glucosidase peak. Subsequent gel filtration of these active fractions (which also contained (1 + 3)-B- glucanase activity29) resulted in the resolution of two protein peaks. The first peak displayed /3-glucosidase activ- ity while the second peak had associated (1 -+ 3)-p- glucanase activity. 29 The purified @glucosidase was ana- lyzed by SDS-PAGE (Figure 2) and IEF (not shown) and in each case, a single protein band was observed which suggested that the enzyme was homogeneous. This was
further confirmed by t+terminal amino acid sequence anal- ysis which showed the absence of any secondary sequences. The purification factors and yields of the @glucosidase at each purification step are summarized in Table 1.
Properties of the purified P-glucosidase
The molecular mass of the purified P-glucosidase was estimated by SDS-PAGE (Figure 2) and gel filtration chromatography to be 128 and 140 kDa, respectively, which suggested that it is a monomeric protein. The isoelectric point determined by IEF was 4.3. A very pale band was observed with periodate/Schiffs staining after SDS-PAGE (not shown). This suggested that the enzyme may be a glycoprotein and was confirmed by quantitative carbohy- drate determinations on the purified enzyme in solution which gave a value of 1.5% (w/w) carbohydrate as glucose equivalents. Maximum enzyme activity was observed at pH 5.5 while the enzyme appeared stable over the pH range 5.0-9.5 (after 24 h at 4°C) and at temperatures up to 50°C for short-term (30 min) incubations.
N-terminal amino acid sequence
The r+terminal amino acid sequence of the A. persicinum P-glucosidase is shown in Table 2. A database search for comparable peptides and corresponding nucleotide se- quences was performed at the National Center for Biotech- nology Information (NCBI) using the BLAST network service.36 The results of this search (Table 2) show similar- ity (as far as the sequence allows) between the A. persici- num enzyme and several other extracellular fungal p- glucosidases including those from Trichoderma reesei (59% identity), 37 Aspergillus aculeatus (4 1% identityJ3’ Saccha- romycopsis fibuligera (41% and 3 1% identity for B-gluco- sidase 2 and 1, respectively),39 and Pichia anomala (26% identity).40
Kinetic properties of the purified f3-glucosidase
The purified @-glucosidase obeyed Michaelis-Menten kinet- ics over the substrate concentration ranges used for all substrates tested. A K, of 0.3 mM was obtained against pNPG while slightly higher K, values (0.6-0.9 mu) were achieved with laminaribiose, gentiobiose, and cellobiose (Table 3). D-Glucono- 1,5-lactone and glucose are competi- tive inhibitors of the @glucosidase, giving Ki values of 5 pM (Km/K,, 60) and 0.35 mu (Km/K,, 0.86), respectively (Table 3).
Table 1 Summary of purification and yields of the extracellular @glucosidase from A. persicinum
Purification step Total protein Total activity
b-ng) KJ) Specific activity
(U mg-‘1
Crude culture filtrate 39 211 5.4 100 1 Ultrafiltration 29.3 209 7.1 99 1.3 (NH&SO, precipitation 6.3 191 30 91 5.6 Mono Q 0.27 41 152 19 28 Superose 12 0.11 28 255 13 47
Yield (%I
Purification factor
184 Enzyme Microb. Technol., 1997, vol. 21, August 15

A. persicinum p-glucosidase: S. Pitson et al.
Table 2 N-terminal amino acid sequence of the A. persicinum p-glucosidase and its similarity to other extracellular fungal 9-glucosidasesa
30 A. persicinum p- :(P)(P)WDEAYT1:AEAALSDLS2:ExKTx lVT(G)V
glucosidaseb (S) G D
6 T. reesei f3-glucosidase G’T ~GT~D~K~~AK~N;~~QD~VG[~S~
18 A. aculeatus f3-glucosidase QG E~Aj~jQR~V@IV~QMT~DE~VNLT~~4:
36 S. fibuligera 6-glucosidase 2 QG v~QD~TK~K@LV~QMTIVE[KJVNLTT~~:
36 S. fibuligera f3-glucosidase 1 QD V~QE~AR~K~IVGQMTIVE~VNLT&?
30 P. anomala @glucosidase DG K~QA@FYR~RELVDQMSIAE~VNLT+$
Waxed amino acids indicate those that are identical to the A. persicinum f3-glucosidase. Numbers indicate the amino acid position from the N-terminus of the purified extracellular enzymes as determined by N-terminal amino acid sequencing, and therefore excludes any signal peptides that may be present on the nascent protein “x’ indicates ‘residue not identifiable’. Amino acids in parentheses indicate tentative assignments, and where two amino acids are given, analysis did not permit confident discrimination
Substrate specificity of the purified P-glucosidase
The purified P-glucosidase hydrolyzed a range of oligo- and aryl P-glucosides (Table 4). Highest activity was observed against laminaribiose, although several other substrates including gentiobiose, laminaritriose, methylumbelliferyl P-glucoside, and pNPG were hydrolyzed almost as rapidly. In fact, the enzyme was active to differing extents against all the oligo+glucosides and aryl and alkyl P-glucosides tested; however, with the exception of 4-nitrophenyl p- xyloside, no activity could be detected against the non-p- glucosides or the long-chain P-glucans tested. Activities against (1 -+ 3)- and (1 + 4)-P-linked oligoglucosides decreased as the chain length increased with only low activi- ties obtained against laminariheptaose or celloheptaose.
Esfects of metal ions and potential inhibitors on the purified P-glucosidase
Several metal ions and potential inhibitors were examined for their effects on the activity of the purified P-glucosidase. Results are shown in Table 5. N-Bromosuccinimide, KMnO,, and Zn2+ were strong inhibitors while less severe
Table 3 Kinetic properties of the purified p-glucosidase from A. persicinuma
Kinetics with pNPG K,,, (mM) k,,, (s ‘1 k,,,/K, (1O-6 X s-’ Mm’)
K, with other substrates (mM) Laminaribiose Gentiobiose Cellobiose
Kib o-glucono-1,5-lactone Glucose
0.30 640 2.13
0.63 0.70 0.91
5 W 0.35 rnM
aAll kinetic parameters were determined at 40°C in 50 mM sodium acetate buffer pH 5.5 using 26 mU of 5-glucosidase ‘Determined with pNPG as substrate
inhibition was observed with 1 rnM Hg*+, Ct.?+, and pHMB, and slight stimulation of activity was noted with Sn’+ and Mg2+. The effects of a range of relatively group-specific potential inhibitors were also investigated with several showing marked inhibition of P-glucosidase activity; however, presence of substrate (pNPG at 2 tJIM)
decreased the inhibition observed with N-bromosuccini- mide, Woodward’s reagent K, and dicyclohexyl carbodi- imide (Table 5).
Glucosyl transferase activity of the purified P-glucosidase
TLC analysis of the hydrolysis products from the action of the purified P-glucosidase after a 60-min incubation with 20 mM laminaribiose or gentiobiose revealed the presence of a range of other, unidentified oligoglucosides (Figure 3) consistent with glucosyl transferase activity. Although no further attempts were made to identify or quantify these products, the results suggest that the enzyme acts with retention of configuration (e -+ e), releasing P-glucose4’ from these substrates.
Investigation of possible cooperation between the A. persicinum P-glucosidase and P-glucanases in P-glucan degradation
The effects of the P-glucosidase on hydrolysis of a range of /3-glucans by the previously purified (1 -+ 3)-p- and (1 + 6)-P-glucanases’9*30 was examined by determining the relative rates of reducing sugar formation by mixtures of the purified P-glucosidase and /3-glucanases. The measured activities do not necessarily reflect P-glucan degradation, but quantify the total reducing sugars present, the levels of which are clearly enhanced by P-glucosidase cleavage of oligoglucosides arising from an initial hydrolysis of the P-glucan by the P-glucanases.
A marked synergism was apparent between the p- glucosidase and (1 + 3)-P-glucanase, although the extent depended on the substrate (Table 6). For example, some
Enzyme Microb. Technol., 1997, vol. 21, August 15 185

Papers
Table 4 Relative rates of hydrolysis of oligoglucosides, aryl and alkyl glucosides, and p-glucans by the purified A. persici- num P-glucosidase
Substrate” Relative activity
(%)b
Laminaribiose Laminaritriose Laminaritetraose Laminaripentaose Laminarihexaose Laminariheptaose Cellobiose Cellotriose Cellotetraose Cellopentaose Cellohexaose Celloheptaose Gentiobiose Sophorose pNPG o-Nitrophenyl P-glucoside Methylumbelliferyl p-glucoside Methyl-p-glucoside Phenyl-p-glucoside Salicin p-Nitrophenyl P-xyloside p-Nitrophenyl P-cellobioside P,P’-Trehalose
100 81 63 41 24 11 48 42 31 20 12 9
86 41 73 20 84
24 31
9 3
aNo activity was detected against laminarins, curdlan, pachyman, CM-pachyman, schizophyllan, scleroglucan, (1 + 3) (1 --f 6)-p- glucan (Ssccharomyces cerevisiae), CM-(1 + 3)(1 + 6)-P-glucan (S. cereuisiae), yeast glucan, lichenin, barley P-glucan, pustulan, lutean, reduced pneumonococcal RSHI glucan, CM-cellulose, HE-cellulose, pullulan, p-nitrophenyl a-glucoside, p-nitrophenyl P-galactoside, p-nitrophenyl p-lactoside, or lactose bThe relative rates of hydrolysis of the P-glucosidase against laminaribiose was arbitrarily set at 100% and corresponds to 25 mU of enzyme activity
synergism was seen with the Laminaria laminarins and schizophyllan [( 1 + 3)-P-glucans with single (1 + 6)-p- linked glucose side chains], presumably through the p- glucosidase hydrolyzing the gentiobiose released by the action of (1 + 3)-P-glucanase29 since glucose was the sole product of the combined enzyme hydrolysis (data not shown). Even greater augmentation was noticed with sub- strates possessing more complex side branching such as the E. bicyclis laminarin and the yeast glucans; however, as expected, little increase in reducing sugar release was observed with the predominantly linear (1 -+ 3)-p-glucans curdlan and CM-pachyman since glucose is the only product of (1 + 3)-P-glucanase action on these substrates.29
Synergism was also observed with the hydrolysis of P-glucans by the P-glucosidase and (1 -+ 6)-P-glucanase (Table 7> especially with the linear (1 + 6)-p-glucans pustulan and lutean presumably through hydrolysis by the P-glucosidase of the (1 + 6)-P-linked oligoglucosides which were released earlier by (1 + 6)-P-glucanase ac- tion.30 Similar cooperative effects were noted with E. bicyclis laminarin and yeast glucans, and in all cases, glucose was the sole product of the combined enzyme hydrolysis (data not shown).
Although the A. persicinum (1 + 3)-P-glucanase de-
Table 5 Effects of metal ions and potential inhibitors on the activity of the purified A. persicinum (3-glucosidase
Compound addeda,b Residual activity
(%)
None 100
HgCl, 79 SnCI, 122 CUCI, 82
MgSO, 119 KMnO, 7.3 ZnSO, 44 pHMB 84 r+Bromosuccinimide (0.1 mM) 0.1 (17)d
(1 rnM) 0 (2) t+Acetylimidazole (10 mM) 36 (39)
(100 rnM) 23 (25) Woodward’s Reagent K (2 mM) 0.4 (12)
(20 rnM) 0 (2) Dicyclohexyl carbodiimide (10 mM) 0.2 (9)
(100 rnM) 0 (2.1) 2-Hydroxy-5-nitrobenzyl bromide (10 mM) 49 (56)
(100 rnM) 38 (43)
“Compounds added at 1 mM unless otherwise indicated bNo effect on activity was observed with the addition of 1 mM BaCI,, NaCI, LiCI, NiCI,, PbCI,, MgSO,, ZnSO,, MnCI,, CoCI,, KCI, AgCI, CaCI,, NH,MoO,, urea, EDTA, acetamide, iodoacetic acid, dithiothreitol, sodium diethyldithiocarbamate, or dieth- ylpyrocarbonate (100 mM) “Activities are expressed as relative percentages of the activity of the P-glucosidase against 2 mM pNPG with no potential inhibitors or metal ions added and corresponds to 25 mU dValues given in parentheses are from substrate protection experiments and are the relative residual activities after incuba- tion of the inhibitor and enzyme in the presence of 10 mM pNPG
grades the extracellular P-glucan from A. persicinum,‘9 and both the purified (1 -+ 3)- and (1 + 6)-P-glucanases from this organism can hydrolyze its cell wall fractions (CWF),28,30 exposure of both to a combination of these purified enzymes failed to increase the hydrolysis of either: however, the (1 -+ 3)+glucanase and P-glucosidase ap- peared able to act synergistically in the hydrolysis of the extracellular glucan, although the P-glucosidase alone showed no activity against this material (Table 4). Subse- quent addition of the (1 -+ 6)-p-glucanase to this reaction mixture failed to increase hydrolysis of this glucan. The P-glucosidase also seemed to slightly increase the rate of reducing sugar formation from the three susceptible A. persicinum cell wall fractions (particularly CWF 3) when combined with either the (1 + 3)-P-glucanase or the (1 + 6)-P-glucanase (Table 8). Further reducing sugar release was noticed when the cell wall fractions were exposed to all three enzymes (Table 8). No such synergism between the P-glucosidase and P-glucanases was noted with any p- glucan substrate which was resistant to (1 -+ 3)-P-glucanase or (1 + 6)+glucanase attack alone.
Discussion
Although multiple P-glucosidase activities are often se- creted by filamentous fungi,3z’ ‘,‘2,42-44 only a single p- glucosidase was isolated from the culture filtrates of A. persicinum grown on laminarin as the sole carbon source.
188 Enzyme Microb. Technol., 1997, vol. 21, August 15

;I. I .;imin:trihiosc h. (:cntiohiose
c; -
I.2 -
I., -
L4 -
I.< -
Figure 3 TLC analysis of possible glucosyl transferase activity of the purified A. persicinum (3-glucosidase with laminaribiose (a) and gentiobiose (b). Sugars present after 60 min incubation of 25 mU of P-glucosidase with 20 mM laminaribiose or gentio- biose. Standards (Std) used were glucose (G), laminaribiose (LJ, laminaritriose (LJ, laminaritetraose (LJ, and laminaripen- taose (LJ
Many fungal P-glucosidases have been isolated and char- acterized, and comparisons of their physicochemical prop- erties with those for the P-glucosidase described here reveals many similarities.2,8 N-terminal amino acid se- quence analysis showed the A. persicinum P-glucosidase to have a high similarity to an extracellular P-glucosidase from T. reesei3’ as well as lower similarities to four other extracellular fungal p-glucosidases (Table 2). Since all five ‘similar’ enzymes have been classified (based on amino acid sequence similarity) into Glycosyl Hydrolase Family 3 45,46 it would be tempting to speculate that the A. pkrsicinum P-glucosidase also belongs to this Family par-
A. persicinum p-glucosidase: S. Pitson et al.
titularly in light of the number of conserved amino acids that exist between all of these enzymes (Table 2). Clearly, however, further sequence analysis would be needed to confirm this.
The purified A. persicinum P-glucosidase has a substrate specificity similar to the P-glucosidases from Trichoderma koningii,’ ’ Aspergillus niger,47 Aspergillus phoenicis,’ Pyricularia oryzae,42 and Sclerotium rolfsii” since it can hydrolyze a range of short-chain (1 + 2)-, (1 -+ 3)-, (1 + 4)-, and (1 + 6)-P-oligoglucosides and aryl @-glucosides, but acts only slowly on methyl p-glucoside. Not all p-glucosi- dases act in this manner, and some are more specific for oligoglucosides with particular P-linkage types. For exam- ple, the P-glucosidases isolated from the bacterium Cyto- phaga arvensicola49 and an Acremonium SP.~O can hydro- lyze both sophorose and laminaribiose, but show no activity against cellobiose and gentiobiose. A similar enzyme from Orpinomyces sp. 5’ also showed no activity toward gentio- biose, but hydrolyzed sophorose, laminaribiose, and cello- biose. Other P-glucosidases have a specificity for aryl P-glucosides and do not hydrolyze P-oligoglucosides.4-‘~52 The A. persicinum P-glucosidase slowly hydrolyzes p- nitrophenyl j3-xyloside, and although rare for fungal p- glucosidases,3 this ability has been reported for the p- glucosidases isolated from A. phoenicis,‘” B. cinereu,” Lenzites trabea.‘” and Stachybotrys atra.54
Inhibition with dicyclohexyl carbodiitnide and Wood- ward’s Reagent K may suggest the importance of aspartyl and glutamyl residues55 possibly at or near the active site (catalytic or binding sites) since the presence of the sub- strate decreases this inhibition. N-Acetylimidazole and 2- hydroxy-5-nitrobenzyl bromide inhibition may also suggest a role for tyrosyl and tryptophanyl residues, respectively,55 although possibly not at the active site since the substrate provided little protection against the recorded loss of activ- ity (Table 5).
P-Glucosidases have been widely implicated in cellulose degradation through synergistic interactions with endo- (1 + 4)-P-glucanases and cellobiohydrolases;‘0-‘3 how- ever, their role in the hydrolysis of other P-glucans, al-
Table 6 Investigations of possible synergistic hydrolysis of (3-glucans by p-glucosidase and (1 + 3)-(3-glucanase from A. persicinuma
Substrate (1 + 3)-P-glucanase (1 + 3)-P-glucanase +
(mUI P-glucosidase (mu)
Increase in hydrolysis
(%)
Laminarin Laminaria digitata Laminaria hyperborea E. bicyclis
Curdlanb CM-pachyman (DS 0.23) Schizophyllan (1 + 3)(1 + 6)-P-Glucan (S. cerevisiae)b CM-(1 + 3111 + 6)-P-Glucan (S. cerevisiae) Yeast glucan
5.76 6.67 15 1.72 2.72 58 1.10 3.06 178 0.08 0.08 0 0.98 1.00 2 2.21 4.31 95 0.01 0.02 100 0.36 0.89 147
0.31 0.86 179
“Assay mixtures contained 5.8 mU of GN II, 25 mU of p-glucosidase, and 2 mg ml-’ substrate in 50 mM sodium acetate buffer pH 5.0. One unit (U) of activity is defined as one pmol of reducing sugar (as glucose equivalents) released min-’ blnsoluble substrates “The increase in hydrolysis was determined on the basis of the increase in reducing sugar release
Enzyme Microb. Technol., 1997, vol. 21, August 15 187
Papers
Table 7 investigations of possible synergistic hydrolysis of P-glucans by P-glucosidase and (1 + 6)-P-glucanase from A. persicinuma
Increase in (1 --f 6)-P-glucanase (1 ---f 6)-P-glucanase + hydrolysis
Substrate (mu) P-glucosidase (mu) (%)”
Pustulan Lutean f. bicyclis laminarin (1 + 3)(1 + 6)-p-Glucan (S. cerevisiae)b CM-(1 --f 3)(1 -+ Gf-P-Glucan ( S. cefevisiae) Yeast glucan
14.43 51.1 253 12.47 41.3 231 6.66 17.7 166 0.03 0.05 67 0.1 0.21 110
0.23 0.52 126
aAssay mixtures contained 14.5 mU of (1 + 6)-p-glucanase, 25 mU of P-glucosidase, and 2 mg ml-’ substrate in 50 mM sodium acetate buffer pH 5.0. One unit (U) of activity is defined as one pmol of reducing sugar (as glucose equivalents) released min-’ “Insoluble substrate “The increase in hydrolysis was determined on the basis of the increase in reducing sugar release
though reported, I4919 has not been as extensively studied. The P-glucosidase from A. persicinum, a noncellulolytic fungus, became of interest when it was noted that the measured (1 + 6)+glucanase activity detected in the crude culture medium decreased substantially (approxi- mately three-fold) upon purification,30 a decrease which can be explained by the removal of the cooperative action of the P-glucosidase and (1 + 6)-P-glucanase after their separa- tion. This type of interaction, although not previously reported for this combination of enzyme types, is not surprising since the substrate specificity of the p-glucosi- dase means it can readily degrade the products released from the endohydrolytic action of the (1 + 6)-P-glucanase on P-glucans, namely gentiobiose, gentiotriose, and larger (1 + 6)-P-oligoglucosides. 3o Similarly, the synergism ob- served with the exo-(1 + 3)-P-glucanases and p-glucosi- dase of A. persicinum on some substrates follows from the apparent ability of the (1 -+ 3)+glucanases to “bypass” (1 -+ 6)-P-linkages in (1 -+ 3)(1 -+ 6)-p-glucans, and thus liberate not only glucose, but also gentiobiose and larger P-linked oligoglucosides. 29 It would appear that all of these products can then be cleaved by the P-glucosidase, thereby
increasing the detectable levels of reducing sugars in the reaction mixtures.
The purified A. persicinum (1 -+ 3)+glucanase partially hydrolyzes the extracellular P-glucan synthesized by this fungus, 26,56 thereby releasing glucose as the only detectable hydrolysis product seen with TLC.29 Hydrolysis of this polymer would not be expected to be enhanced by the presence of P-glucosidase to the reaction mixture, since there are no obvious oligoglucoside products for it to degrade: however, an unexpected synergism was observed which possibly arose from P-glucosidase hydrolysis of long-chain oligoglucosides released by the action of the (1 -+ 3)-P-glucanases which were too large to be detected by the TLC conditions used. The hydrolysis of the A. persicinum CWF 3 by the (1 + 3)- and (1 + 6)-p- glucanase was enhanced six-fold and three-fold, respec- tively, by the presence of P-glucosidase again probably through the hydrolysis of oligoglucosides released from the cell wall P-glucans by P-glucanase action.** Since both the (1 + 3)- and (1 + 6)-P-glucanase release glucose as the major hydrolysis product from CWF 1 and 2,28 it is not surprising that little synergism was observed with these
Table 8 Investigations of possible synergistic action of P-glucosidase, (1 -+ 3)-P-glucanase and (1 + 6)-P-glucanase against the A. persicinum extracellular glucan and cell wall preparations
Activity ( mU)“,b
Enzyme(s) CWF 1 CWF 2 CWF 3 Extracellular
glucan
(1 -+ 3).p-glucanase (1 + 6).P-glucanase p-glucosidase (1 -+ 3).P-glucanase + P-glucosidase (1 + 6).p-glucanase + P-glucosidase (1 + 3).P-glucanase + (1 + 6)-P-glucanase (1 + 3).p-glucanase + (1 + 6)-P-glucanase
+ p-glucosidase
0.44 0.47 0.07 1.10 1.94 1.04 0.57 0.04 0 0 0 0 0.53 0.65 0.42 1.79 2.04 1.30 1.84 0.04 2.18 1.42 0.76 1.11 2.43 1.78 2.04 1.87
aAssay mixtures contained 5.8 mU of GN II, 14.5 mU of (1 -+ 6)-P-glucanase, 25 mU of P-glucosidase and 2 mg ml-’ substrate in 50 mM sodium aCetate buffer pH 5.0. One unit (U) of activity is defined as one kmol of reducing sugar (as glucose equivalents) released min-’ bNo activity was observed against A. persicinum CWF 4
188 Enzyme Microb. Technol., 1997, vol. 21, August 15

enzymes and the P-glucosidase with either of these wall fractions. All these data are consistent with a possible role for the P-glucosidase in the assimilation of extracellular or cell wall P-glucans as carbon sources through hydrolysis of oligoglucosides released by P-glucanase action in a manner analogous to cellobiose hydrolysis during cellulose degra- dation.57
Acknowledgments
We thank Prof. Bruce A. Stone and Ms. Rosemary Condron, both of the School of Biochemistry, LaTrobe University, for the N-terminal amino acid sequence.
References
I.
2.
3.
4.
5.
6.
7.
8.
9.
IO.
I I.
12.
13.
14.
15.
16.
17.
18.
19.
Webb. E. C. Enzyme Nomenclature: Recommendations of the Nomencluture Committee of the International Union of Biochemiq- TV und Molecular Biology. Academic Press, Orlando, 1992, 346- 365 Woodward, J. and Wiseman. A. Fungal and other @-D-glucosidases: Their properties and applications. Enzyme Microb. Technol. 1984.4, 73-79 Shewale. J. G. P-Glucosidase: Its role in cellulase synthesis and hydrolysis of cellulose. ht. J. Biochem. 1982, 14, 435-443 Jermyn. M. A. Fungal cellulases. VI. Substrate and inhibitor specificity of the P-glucosidase of Stachybotrys arm. Aust. .I. Biol. Sci. 1955. 8. 577-602 Jermyn. M. A. Glycosidases. Rev. Pure Appl. Chem. 1961. 11. 92-116 HGsel, W. and Conn, E. E. The aglycone specificity of plant glycosidases. Trends Biochem. Sri. 1982, 7, 219-221 Bucht. B. and Eriksson. K. E. Extracellular enzyme system utilized by the rot fungus Srereum sanguinoletum for the breakdown of cellulose. IV. Separation of cellobiase and ary-P-glucosidase ac- tivities. Arch. Biochem. Biophvs. 1969, 129, 416-420 Stone. B. A. and Clarke. A. E. Chemistry and Biology cg (I - _?J-/3-Glucans. LaTrobe University Press, Melbourne, 1992 Gueguen. Y., Chemardin, P., Arnaud, A., and Galzy, P. Purification and characterization of an intracellular P-glucosidase from Botrytis cinereu. Enzyme Microb. Tech&. 1995. 17, 900-906 Sternberg. D.. Vijayakumar, P., and Reese. E. T. P-Glucosidase: Microbial production and effect on enzymatic hydrolysis of cellu- lose. Con. J. Microbial. 1977, 23, 139-147 Wood. T. M. and McCrae, S. I. Purification and properties of the extracellular P-D-glucosidase of the cellulolytic fungus 7’ri- chodermu koningii. J. Gem Microbial. 1982, 128, 2973-2982 Sakamoto, R.. Arai, M.. and Murao, S. Enzymic properties of three B-glucosidases from Aspergillus acLdeatus No. F-50. Agric. Biol. Chem. 1985. 49, 1283-1290 Beldman, G., Voragen. A. G. J., Rombouts, F. M., and Pilnik, W. Synergism in cellulose hydrolysis by endoglucanases and exoglu- canases purified from Trichoderma viride. Biotechnol. Bioenx. 1988. 31, 173-178 Stahmann. K.-P., Pielken. P., Schimz, K.-L., and Sahm, H. Degra- dation of extracellular p-( I ,3)( 1,6)-o-glucan by Bottytis cinererr. Appl. Environ. Microbial. 1992, 58. 3347-3354 Stahmann, K.-P., Schimz, K.-L.. and Sahm, H. Purification and characterisation of four extracellular 1.3-p-glucanases of BotFtis cirrerecc. .I. Gen. Microbial. 1993, 139, 2833-2840 Dickerson, A. G.. Mantle, P. G., and Szczyrbak, C. A. Autolysis of extracellular glucans produced in vitro by a strain of Claviceps fusiformis. J. Gen. Microbial. 1970, 60, 403-415 Rapp, P. 1,3-P-Glucanase, 1.6.P-glucanase, and P-glucosidase ac- tivities of Sclerotium glucanirum: Synthesis and properties. J. Gen. Microbial. 1989, 135, 2847-2858 Rapp, P. Formation, separation, and characterisation of three 0-l .3- glucanases from Sclerotium &a2nicum. 1990, 1117.7-14
Biochim. Biophys: Acta
Copa-Patiiio, J. L.. Rodriqurz. J., Reyes, F.. and Perez-Leblic. M. 1.
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35. 36.
37.
38.
39.
40.
41.
42.
A. persicinum P-glucosidase: S. fitson et al.
Effect of P-glucanases on Penicillium oxulicum autolysates. FEMS Microbial. L.&t. 1990, 70, 233-240 Bielecki, S. and Galas, E. Microbial P-glucanases different from cellulases. Crit. Rev. Biorechnol. 1991. 10, 275--304 Santos, T., Sanchez. M., Villanueva. J. R., and Nombela, C. Regulation of the 1,3-P-glucanase system in Penicillium it&-urn: Localization of the various enzymes and correlation with cell wall glucan mobilization and autolysis. J. Bucteriol. 1978, 133,465-47 1 Del Rey, F., Garcia-Acha. I.. and Nombela, C. The regulation of P-glucanase synthesis in fungi and yeast. .I. Gen. Micmbinl. 1979. 110,83-89 Hiura, N.. Honjyo. I.. Nakajima, T., and Matsuda, K. Change of the structure of cell wall P-1.3~D-glucan with the growth of Neurosporu crussu cells. Agric. Biol. Chem. 1984. 48, lO4--1047 Hiura, N., Kobayashi, M., Nakajima. T.. and Matsuda, K. Purifica- tion and some properties of two wail-associated (pl.3)glucanases from Neurospom crussu cells. Agric. Biol. Chem. 1986. 50. 2461- 2467 Hiura. N.. Nakajima, T.. and Matsuda, K. Purification and some properties of an endo-@ I ,6-glucanase from Neuroyporu ~~russu. Agric. Biol. Chem. 1987, 51, 3315-3321 Stasinopoulos, S. J. and Seviour, R. J. Exopolysaccharide formation by isolates of Cephulosporium and A~~remonium. Myc 01. Re.r. 1989, 92, 55-60 Pitson, S., Seviour. R. J.. Bott, J.. and Stasinopoulos. S. J. Produc- tion and regulation of P-glucanases in Acremonium and Cephulos- porium isolates. ~~col. Res. 1991, 95, 352-356 Pitson. S. M. Punfication. characterisation. and regulation of syn- thesis of P-glucan degrading enzymes from the fungus Acremonium persicinum. Ph.D. thesis. LaTrobe University, Victoria. Australia, 199s Pitson. S. M.. Seviour. R. J.. McDougall. B. M.. Woodward, J. R., and Stone. B. A. Purification and characterization of three extracel- lular (I - 3).P-D-glucan glucanohydrolases from the filamentous fungus Acremonium persicinum. Biochem. J. 1995. 308. 733-741 Pitson, S. M.. Seviour, R. J., McDougall. B. M.. Stone. B. A., and Sadek, M. Purification and characterization of an extracellular ( I + 6). P-glucanase from the tilamentous fungus Acremonium petxicinum. Biochem. J. 1996, 316, 84 l-846 Hrmova, M. and Fincher, G. B. Purification and properties of three (I - 3).P-o-glucanase isoenzymes from young leaves of barley (Hordeurn vulgure). Biochem. .I. 1993. 289, 453-461 Berghem, L. E. R. and Petterson. L. G. The mechanism of enzymatic cellulose degradation. Characterization and enzymatic properties of a p- 1.4.glucan cellobiohydrolase from Trichodemwr viride. Eur. J. Biochem. 1973. 37, 21-30 Nelson, N. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem. 1944. 153, 375-380 Somogyi, M. Notes of sugar determination. .1. Ri~L Chem. 1952. 195, 19-23 Williams, P. A. Enzpuck. Elsevier-Biosoft, Cambridge, 1985 Altschul. S. F., Gish, W.. Miller, W.. Myers, E. W., and Lipman. D. J. Basic local alignment search tool. ./. Mol. Hiol. 1990. 215, 403-410 Barnett. C. C.. Berka. R. M., and Fowler. T. Cloning and amplifi- cation of the gene encoding an extracellular P-glucosidase from Trichoderma reesei: Evidence for improved rates of saccharification of cellulosic substrates. Biorechnologv I99 I. 9, 562-567 Kawaguchi. T.. Enoki, T.. Tsurumaki. S.. Sumitani, J., Ueda. M., Ooi, T., and Arai. M. Cloning and sequencing of the cDNA encoding P-glucosidase I from Aspeyqillus oculeutuv. Gene 1996. 173,x37-288 Machida, M., Ohtsuki. I.. Fukui, S.. and Yamashita. I. Nucleotide sequences of Succhurom~copsis ,fibuligeru genes for extracellular P-glucosidases as expressed in Succhuromyce.\ cerevisiue. Appl. Environ. Microbial. 1988. 54, 3 147-3 I S5 Kohchi. C. and Toh-e, A. Nucleotide sequence of Cundidu pellicu- losu P-glucosidase gene. Nucl. Acid Res. 1985, 13, 6273-6282 Sinnott, M.L. Catalytic mechanisms of enzymic glycosyl transfer. Chem. Rev. 1990, 90, 1 I7 I- I202 Hirayama. T.. Horie, S., Nagayama, H., and Matsuda, K. J. Studies on cellulases from a phytopathogenic fungus, Pyriculuriu or?/zue. Carvara. II. Purification and properties of a P-glucosidase. Biochem- htp 1978. 84, 27-37
Enzyme Microb. Technol., 1997, vol. 21, August 15 189

Papers
43.
44.
45.
46.
47.
48.
49.
50.
Bhat, K. M., Gaikwad, J. S., and Maheshwari, R. Purification and characterisation of an extracellular B-glucosidase from the thermo- phylic fungus Sporotrrchum fhermophile and its influence on cellulase activity. J. Gen. Microbial. 1993, 139,2825-2832 Himmel, M. E., Adney, W. S., Fox, J. W., Mitchell, D. J.. and Baker, J. 0. Isolation and characterization of two forms of P-D-
glucosidase from Aspergillus niger. Appl. Biochem. Biotechnol. 1993,39/40,213-225 Hen&at, B. A classification of glycosyl hydrolases based on amino acid sequence similarities. Biochem. J. 1991, 280, 309-316 Henrissat, B. and Bairoch, A. New families in the classification of glycosyl hydrolases based on amino acid sequence similarities. Biochem. J. 1993, 293, 781-788 Unno, T., Ide, K., Yazaki, T., Tanaka, Y., Nakakuki, T., and Okada, G. High recovery purification and some properties of a B-glucosi- dase from Aspergillus niger. Biosci. Biotechnol. Biochem. 1993.57, 2172-2173 Sadana, J. C., Shewale, J. G., and Patil, R. V. B-o-Glucosidases of Sclerotium rolfsii. Substrate specificity and mode of action. Carbo- hydr. Res. 1983, 118, 205-214 Mendoza, N. S. and Amemura, A. (1 + 2)-B-o-Glucan-hydrolysing enzymes in Cyzophaga arvensicola: Partial purification and some properties of endo-( 1 -+ 2)-B-o-glucanase and B-D-glucosidase specific for (1 4 2)- and (1 + 3)-linkages. J. Ferment. Technol. 1983, 61,473-481 Kitahata, S. and Edagawa, S. Cyclic (1 + 2)-B-o-glucan-hydrolys-
51.
52.
53.
54.
55.
56.
57.
ing enzymes from Acremonium sp. 15: Purification and some properties of endo-(1 + 2)-B-o-glucanase and B-D-glucosidase. Agric. Biol. Chem. 1987, 51, 2701-2708 Chen, H., Li, X., and Ljungdahl, L. G. Isolation and properties of an extracellular B-glucosidase from the polycentric rumen fungus Orpinomyces sp. strain PC-2. Appl. Environ. Microbial. 1994. 60, M-70 Meyer, H.-P. and Canevascini, G. Separation and some properties of two intracellular B-glucosidases of Sporotrichum (Chrysosporium) thermophile. Appl. Environ. Microbial. 1981, 41, 924-931 Herr, D., Baumer, F., and Dellweg, H. Purification and characteri- sation of an extracellular B-glucosidase from Lenzites rabea. Eur. J. Appl. Microbial. Biotechnol. 1978, 5, 29-36 Gussem, R. L. D., Aerts, G. M., Claeyssens, M., and Debruyne, C. K. Purification and properties of an induced B-glucosidase from Stachybotrys atra. Biochim. Biophys. Acta 1978, 525, 142-153 Freedman, R. B. Determination of essential residues in enzymes by chemical modification. Tech. Protein Enzyme Eiorechnol. 1978, B117, 229-235 Stasinopoulos, S. .I. Polysaccharide production by isolates of Ceph- alosporium and Acremonium. M. Appl. SC. thesis, College of Advanced Education, Bendigo, Victoria, Australia, 1990 Goyal, A.. Ghosh, B., and Eveleigh, D. Characteristics of fungal cellulases. &ores. Technol. 199 1, 36, 37-50
190 Enzyme Microb. Technol., 1997, vol. 21, August 15




















