
Proteome analysis of brown spider venom: Identification of loxnecrogin isoforms inLoxosceles gaucho...
12
135 Journal of Protein Chemistry, Vol. 22, No. 2, February 2003 (© 2003) 0277-8033/03/0200-0135/0 © 2003 Plenum Publishing Corporation Purification and Characterization of Loxnecrogin, a Dermonecrotic Toxin from Loxosceles gaucho Brown Spider Venom Ricardo Bastos Cunha, 1,2 Katia Cristina Barbaro, 3,5 Daniela Muramatsu, 3 Fernanda Calheta Vieira Portaro, 4 Wagner Fontes, 1 and Marcelo Valle de Sousa 1 Received November 4, 2002 The most common manifestation of Loxosceles spider envenoming is a dermonecrotic lesion at the bite site. Dermonecrotic toxins from Loxosceles gaucho venom were purified and characterized by mass spectrometry (capillary liquid chromatography followed by mass spectrometry detection). Two components were purified: a major one of 31,444 Da, called loxnecrogin A, and a minor one of 31,626 Da, called loxnecrogin B, being probably two isoforms of the toxin. The N-terminal sequence of loxnecrogin A showed similarity with N termini of other sphingomyelinolytic dermonecrotic toxins isolated from venoms of different Loxosceles species. The internal sequences did not present any statistically significant hits in sequence databases searches. However, loxnecrogin A partial sequence showed high similarity to regions of L. intermedia LiD1 recombinant protein sequence, recently described in the literature but not yet deposited in databanks. KEY WORDS: Loxosceles gaucho; dermonecrotic toxin; brown spider venom; purification; mass spectrometry. 1. INTRODUCTION Loxosceles spiders are found in both South and North America, as well as in Middle Eastern regions. In Brazil, it is widely distributed with seven species ranging from northern to southern states. In the last years, an increase in notified cases (2.200 cases/year), probably due to intradomiciliar infestation, was reported (Ministry of Health, 1998), turning loxoscelism the most important form of arachnidism in Brazil. In the southern and south- eastern regions of Brazil, three species of Loxosceles are most commonly found: L. gaucho, L. laeta, and L. inter- media. In the state of São Paulo, L. gaucho is responsible for the large majority of bites. The most common clinical picture presentation is a dermonecrotic lesion at the bite site. Most patients (approximately 90%) present only this local injury, which is classified as cutaneous loxoscelism. Some pa- tients (3%–16%) present with a more severe systemic re- sponse to Loxosceles envenomation (viscerocutaneous loxoscelism), characterized by intravascular hemolysis, disseminated intravascular coagulation, and acute renal 1 Brazilian Center for Protein Research, Department of Cell Biology, Institute of Biology, University of Brasília, Brasília, DF, 70910-900, Brazil. 2 Analytical Chemistry Area, Institute of Chemistry, University of Brasília, Brasília, DF, 70910-900, Brazil. 3 Laboratory of Immunopathology, Institute Butantan, São Paulo, SP, 05503-900, Brazil. 4 Center for Applied Toxinology (CAT/CEPID), Institute Butantan, São Paulo, SP, 05503-900, Brazil. 5 To whom correspondence should be addressed at Laboratory of Immunopathology, Butantan Institute, Av. Vital Brazil, 1500, São Paulo, SP, 05503-900 Brazil. E-mail: [email protected] 6 Abbreviations: BLAST, basic local alignment search tool; CID, collision-induced dissociation; cLC-MS, capillary liquid chromatog- raphy followed by mass spectrometry detection; DTT, dithiothreitol; ESI-MS, electrospray ionization–mass spectrometry; FPLC, fast pro- tein liquid chromatography; HPLC, high-performance liquid chro- matography; LD 50 , lethal dose (dose that killed 50% of the animals); LiD1, Loxosceles intermedia dermonecrotic protein 1; MS, mass spectrometry; NCBI, National Center for Biotechnology Information; recLiD1, recombinant LiD1; RP, reversed-phase; SDS-PAGE, sodium dodecyl sulfate–polyacrylamide gel electrophoresis; TFA, trifluoroacetic acid.
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Proteome analysis of brown spider venom: Identification of loxnecrogin isoforms inLoxosceles gaucho...

135
Journal of Protein Chemistry, Vol. 22, No. 2, February 2003 (© 2003)
0277-8033/03/0200-0135/0 © 2003 Plenum Publishing Corporation
Purification and Characterization of Loxnecrogin,a Dermonecrotic Toxin from Loxosceles gauchoBrown Spider Venom
Ricardo Bastos Cunha,1,2 Katia Cristina Barbaro, 3,5 Daniela Muramatsu,3
Fernanda Calheta Vieira Portaro,4 Wagner Fontes,1 and Marcelo Valle de Sousa1
Received November 4, 2002
The most common manifestation of Loxoscelesspider envenoming is a dermonecrotic lesion at thebite site. Dermonecrotic toxins from Loxosceles gauchovenom were purified and characterized bymass spectrometry (capillary liquid chromatography followed by mass spectrometry detection).Two components were purified: a major one of 31,444 Da, called loxnecrogin A, and a minor oneof 31,626 Da, called loxnecrogin B, being probably two isoforms of the toxin. The N-terminalsequence of loxnecrogin A showed similarity with N termini of other sphingomyelinolyticdermonecrotic toxins isolated from venoms of different Loxoscelesspecies. The internal sequencesdid not present any statistically significant hits in sequence databases searches. However, loxnecroginA partial sequence showed high similarity to regions of L. intermediaLiD1 recombinant proteinsequence, recently described in the literature but not yet deposited in databanks.
KEY WORDS: Loxosceles gaucho;dermonecrotic toxin; brown spider venom; purification; mass spectrometry.
1. INTRODUCTION
Loxoscelesspiders are found in both South and NorthAmerica, as well as in Middle Eastern regions. In Brazil,it is widely distributed with seven species ranging fromnorthern to southern states. In the last years, an increasein notified cases (2.200 cases/year), probably due tointradomiciliar infestation, was reported (Ministry ofHealth, 1998), turning loxoscelism the most importantform of arachnidism in Brazil. In the southern and south-eastern regions of Brazil, three species of Loxoscelesare
most commonly found: L. gaucho, L. laeta,and L. inter-media.In the state of São Paulo, L. gauchois responsiblefor the large majority of bites.
The most common clinical picture presentation isa dermonecrotic lesion at the bite site. Most patients(approximately 90%) present only this local injury,which is classified as cutaneous loxoscelism. Some pa-tients (3%–16%) present with a more severe systemic re-sponse to Loxoscelesenvenomation (viscerocutaneousloxoscelism), characterized by intravascular hemolysis,disseminated intravascular coagulation, and acute renal
1 Brazilian Center for Protein Research, Department of Cell Biology,Institute of Biology, University of Brasília, Brasília, DF, 70910-900,Brazil.
2 Analytical Chemistry Area, Institute of Chemistry, University ofBrasília, Brasília, DF, 70910-900, Brazil.
3 Laboratory of Immunopathology, Institute Butantan, São Paulo, SP,05503-900, Brazil.
4 Center for Applied Toxinology (CAT/CEPID), Institute Butantan,São Paulo, SP, 05503-900, Brazil.
5 To whom correspondence should be addressed at Laboratory ofImmunopathology, Butantan Institute, Av. Vital Brazil, 1500, SãoPaulo, SP, 05503-900 Brazil. E-mail: [email protected]
6 Abbreviations: BLAST, basic local alignment search tool; CID,collision-induced dissociation; cLC-MS, capillary liquid chromatog-raphy followed by mass spectrometry detection; DTT, dithiothreitol;ESI-MS, electrospray ionization–mass spectrometry; FPLC, fast pro-tein liquid chromatography; HPLC, high-performance liquid chro-matography; LD50, lethal dose (dose that killed 50% of the animals);LiD1, Loxosceles intermediadermonecrotic protein 1; MS, massspectrometry; NCBI, National Center for Biotechnology Information;recLiD1, recombinant LiD1; RP, reversed-phase; SDS-PAGE,sodium dodecyl sulfate–polyacrylamide gel electrophoresis; TFA,trifluoroacetic acid.

1998). The presence of calcium-dependent sphin-gomyelinases in the L. reclusa(Kurpiewski et al.,1981)and L. intermedia(Tambourgi et al.,1998) venoms sug-gests that this class of toxic enzymes might also bindmetals. Portaro et al. (personal communication) showedthat EDTA and 1,10-phenanthroline inhibit the sphin-gomyelinase activity of the dermonecrotic fraction of theL. gauchovenom, indicating that the sphingomyelinaseactivity could actually be metal dependent.
Barbaro and co-workers (1996) showed that thevenoms of L. gaucho, L. laeta,and L. intermediacouldbe separated over Sephadex G-100 into three fractions.Only fraction A, containing components with molecularmass above 20 kDa, showed the dermonecrotic andlethal activities in all venoms. The electrophoretic profileof Brazilian Loxoscelesspp. venoms revealed majorcomponents, of 30–35 kDa, responsible for der-monecrotic activity. These components are probablyhomologous proteins, because they have high similar-ity in the N-terminal sequences (Barbaro et al., 1996),and show intense cross-reactivity when species-specificanti-sera were used (Barbaro et al., 1994). However,monoclonal antibodies produced against the 35-kDacomponent of L. gaucho venom cross-reacted onlyweakly with the 35-kDa component of L. intermediavenom and did not recognize the 32-kDa component ofL. laetavenom (Guilherme et al.,2001). So, despite theirhomology, the dermonecrotic components of the venomsof different Loxoscelesspecies are distinct.
The aim of the present study was to purify and tocharacterize the dermonecrotic proteins from L. gauchovenom. For the characterization, electrospray mass spec-trometry (ESI-MS)6 was used for both to determine iso-forms of the dermonecrotic toxin and to generate internalamino acid sequences.
2. MATERIALS AND METHODS
2.1. Chemicals and Reagents
Methanol and acetonitrile (sequenal grade) werepurchased from Applied Biosystems. NH4HCO3 (analyt-ical grade) was obtained from Riedel-Dehaënag Seelze.The water used was purified by a Millipore Milli-Q sys-tem. Unless otherwise indicated, all other chemicals andreagents were purchased from Sigma Chemical Co.
2.2. Animals
Male Swiss mice weighing 18–20 g and adult whiterabbits weighing 3–4 kg were provided by the InstitutoButantan animal house.
136 Cunha, Barbaro, Muramatsu, Portaro, Fontes, and Valle de Sousa
failure, which is the main cause of death from lox-oscelism (Cardoso et al., 1988; Sezerino et al., 1998).The bite is initially painless. Within 2–6 hr, the bittensite becomes painful, accompanied by erythema, itching,swelling, and tenderness. From 1–7 days later, an escharis formed, leaving an ulcer that may take months to heal,usually associated with scar formation. Histologically,the injury appears to be a cutaneous Arthus reaction, pre-senting with edema, polymorphonuclear infiltration, fib-rinoid necrosis, vacuolization of the arterial walls, andintense intradermal hemorrhage. The systemic effects aremuch less common: fever, malaise, chills, weakness,nausea, and vomiting can occur up to 24 hr after the bite.The hematological disturbances may include hemolyticanemia, thrombocytopenia, and consumption coagulo-pathy (Futrell, 1992). Differences in the severity of thecases may be due to spider species, spider sex (Oliveiraet al.,1999), developmental stage of the spider (Andradeet al.,1999), amount of injected venom, and patient age(Sezerino et al.,1998) or genetics (Barreto et al.,1985).The definite therapy for the treatment of the lesioncaused by Loxoscelesspider bites has not yet been es-tablished. Usually, treatment with antiserum is generallyindicated in cases when the patient comes early to a hos-pital. Currently, specific treatment with serum therapywith horse polyvalent anti-arachnid or anti-loxoscelicvenom sera is the most widely used, sometimes associ-ated with corticoids (Ministry of Health, 1998).
A number of different enzymes have been identifiedas constituents of Loxoscelesspp. venom (reviewed inRash and Hodgson, 2002), including alkaline phos-phatases (Heitz and Norment, 1974), hyaluronidases(Kaiser, 1956; Wright, 1973; Elgert et al., 1974; Gerenet al., 1976; Schenone and Suarez, 1978; Young andPincus, 2001), hydrolases, lipases (Norment et al.,1979),and proteases (Wright, 1973; Jong et al.,1979; Barbaroet al.,1996; Feitosa et al.,1998; Young and Pincus, 2001).
Sphyngomyelinase D, a major component of theLoxoscelesvenom, was early recognized as related to thedermonecrosis (Forrester et al.,1978; Kurpiewski et al.,1981). More recent reports confirm this finding (Futrell,1992; Tambourgi et al.,1998), although it is clear that aset of enzymes must work in a concerted manner in theformation of the necrotic injury. The enzyme sphyngo-myelinase interacts with the cellular membrane, deregu-lating the complement system, activating platelets andpolymorphonuclear leukocytes, and thereby causing thedermonecrotic lesion (Futrell, 1992). Sphyngomyeli-nases purified from L. reclusaand L. intermediahas beenshown to induce dermonecrosis similar to that producedby the whole venom when injected intradermally intorabbits (Kurpiewski et al., 1981; Tambourgi et al.,

2.3. Venom
Specimens of L. gaucho were collected in thestate of São Paulo, Brazil. Venom was obtained byelectrostimulation, as described by Barbaro et al.(1992). A pool of venoms from approximately 4000spiders was used. The protein content of the venomand of the chromatographic fractions was determinedby the method of Bradford (1976).
2.4. Chromatographic Separation of the VenomFractions
L. gauchovenom was fractionated by cation ex-change FPLC (Amersham Biosciences) with a Mono-SHR 5/5 column, equilibrated with a 50 mmol/L sodiumacetate buffer, pH 5.0. One milliliter of dialyzed samples(1 mg/ml) was applied to the column and eluted with alinear gradient of NaCl (0–1 mol/L) in the same buffer.The flow rate was 1 ml/min, and the absorbance wasmonitored at 280 nm. Fractions of 1.0 ml correspondingto each protein peak were then pooled, lyophilized, andstored at 220°C.
The active fraction obtained from cation-exchangechromatography was submitted to reversed-phase HPLC(Perkin Elmer series LC 200 with an Applied Biosystems783A UV absorbance detector) using a wide-pore butylC4 Vydac column (4.6 3 150 mm, 5-mm particle size). Thefractions were eluted with a linear gradient of 0%–70%acetonitrile in 0.1% aqueous trifluoroacetic acid, at aflow rate of 1 ml/min. The elutate was monitored by itsabsorbance at 216 nm. Manually collected fractions weredried in a vacuum centrifuge.
2.5. Polyacrylamide Gel Electrophoresis in Presenceof SDS (SDS-PAGE)
Fractions were analyzed by SDS-PAGE (12% poly-acrylamide gels) under nonreducing conditions using themethod of Laemmli (1970). The L. gauchovenom (3 mg)and the fractions (0.5 mg) were diluted in sample bufferand boiled for 1 min before electrophoresis. Phosphory-lase B (94 kDa), BSA (67 kDa), ovalbumin (43 kDa),carbonic anhydrase (30 kDa), trypsin (20.1 kDa), anda-lactalbumin (14.4 kDa) were used as molecular weightmarkers. The gels were silver stained, as described byBlum et al. (1987), and interpreted with the aid of theImageMaster 2D Elite program, version 3.01, fromAmersham Biosciences.
2.6. Local Reaction and Dermonecrotic Activity
Different samples of L. gauchovenom or its purifiedfractions were diluted in 200 mL of 0.15 mol/L NaCl,
and injected intradermally into the shaved backs of adultrabbits. Control sites were injected with an equal volumeof 0.15 mol/L NaCl. The size of the affected areas wasmeasured after 48 hr.
2.7. Lethal Activity
Groups of four Swiss mice were injected intra-dermally with 200 ml of 0.15 mol/L NaCl containing dif-ferent concentrations (0.25–2.50 mg/kg body weight) ofL. gauchovenom. The LD50 observed 48 hr after injec-tion was determined by Probit analysis (Finney, 1971).The assays were carried out in duplicate. The LD50 de-termined in mice 48 hr after venom injection was 0.97mg/kg to L. gauchovenom. To determine the lethal ac-tivity of the purified fractions, male Swiss mice were in-jected intradermally with 200 ml of 0.15 mol/L NaClcontaining 0.485 mg (1/2LD50) of each fraction perkilogram of the animal’s weight.
2.8. Edman Sequencing
The protein was reduced with DTT and alkylatedwith 4-vinylpyridine, cleaned up by reversed-phase HPLC(C4 column), and dried in a vacuum centrifuge. It wasthen dissolved in acetonitrile/0.1% v/v trifluoroacetic acid(2:8) and applied directly into the reaction chamber of anApplied Biosystems 477A automated protein sequencercoupled to a 120A analyzer. The equipment was program-med according to the manufacturer’s instructions, withalterations as described in the literature (Fontes et al.,1998). The similarity searches were performed using theBLAST program at NCBI (Altschul et al.,1990).
2.9. Trypsin Digestion
The purified protein was dissolved in 100 ml of guani-dine 6 mol/L–Tris 0.6 mol/L (pH 8.6) containing 1.5 mg/mlof DTT and incubated under N2 for 1 hr 30 min at 37°C.After the addiction of 8 ml of 4-vynilpyridine, the samplewas incubated again for an additional 1 hr 30 min under N2
at 37°C. The lyophilized alkylated protein was then dis-solved in 0.1 mol/L NH4HCO3, buffer pH 8.1, containing2% of trypsin (w/w trypsin/protein) and incubated underN2 for 3 hr at 37°C. The digestion was stopped by freezingand lyophilizing the sample. Prior to MS, the digest wasdesalinized by using a Poros R2 microcolumn.
2.10. Electrospray Mass Spectrometry
The samples were dissolved in 50% acetonitrile(v/v) in 5% acetic acid (v/v), at a final concentration of5 pmol/ml, and injected into a Perkin Elmer-Sciex electro-spray triple quadrupole model API 300 mass spectrometer
Purification and Characterization of Loxnecrogin 137

equipped with a Protana nanoelectrospray ionizationprobe, using a 52-mm coated capillary with an 0.8-mmtip length, 2 mm ID, 1.2 mm OD, at a flow rate of 10–40 nl/min. The spectra were collected at the range of200–2000 Da with 0.3-Da step size, 0.5-ms dwell time,10-ms pause time in typically 30 scans. The data werecollected at the positive mode and interpreted with theaid of the software BioMultiView version 1.2 fromPerkin Elmer-Sciex.
2.11. Capillary Liquid Chromatography with MassSpectrometry Detection (cLC-MS)
For the cLC-MS experiments, the mass spectro-meter was coupled to an Applied Biosystems 140C micro-gradient system coupled to a 785A programmable ab-sorbance detector and an 112A oven/injector unit. Threemicrograms of the sample was dissolved in 40 ml of0.1% formic acid, from which 2 ml was applied directlyinto a homemade capillary column (150 mm ID 3 25 cmlength) packed with Vydac C18 resin at 5-mm particle dia-meter. The fractions were eluted by a 0%–100% aceto-nitrile gradient (containing 0.1% formic acid), at a flowrate of 40 ml/min regulated by a splitter, which alloweda flow rate of 30 ml/hr into the column. The solvent flowwas directed straight from the column to the microion-spray probe of the mass spectrometer described above.
2.12. Tandem Mass Spectrometry (MS/MS)
The samples, previously digested with trypsin,were injected into the electrospray probe as describedabove. The peptides were selected at the first quadru-pole and fragmented by collision-induced dissociation(CID) with nitrogen gas at the second quadrupole. Thespectra were collected with 30-Da start mass (the stopmass depended on the peptide being fragmented), 0.2-Dastep size, 1-ms dwell time, 2-ms pause time, and 50scans. The fragments obtained were analyzed at the thirdquadrupole. The sequence data were deduced both man-ually and with the aid of the package BioMultiViewversion 1.2 from Perkin Elmer-Sciex instruments. TheMS/MS searches were performed via the programsMascot from Matrix Science Ltd. (http://www.matrixscience.com/search_form_select.html) (Perkins et al.,1999) and PepSea from Protana (http://195.41.108.38/PepSeaIntro.html).
3. RESULTS
The proteins of L. gauchovenom were fractionatedby cation-exchange chromatography into 15 fractions
(Fig. 1). Fraction 8, which contains proteins of approxi-mately 30 kDa by SDS-PAGE (Fig. 2), was the majorfraction in terms of both protein content and activity.Dermonecrotic and lethal activities were detected only infraction 8 (Table 1), which also produced more intenselocal reactions (edema, erythema, hemorrhagic and paleareas). Other fractions induced only minor local reactions.
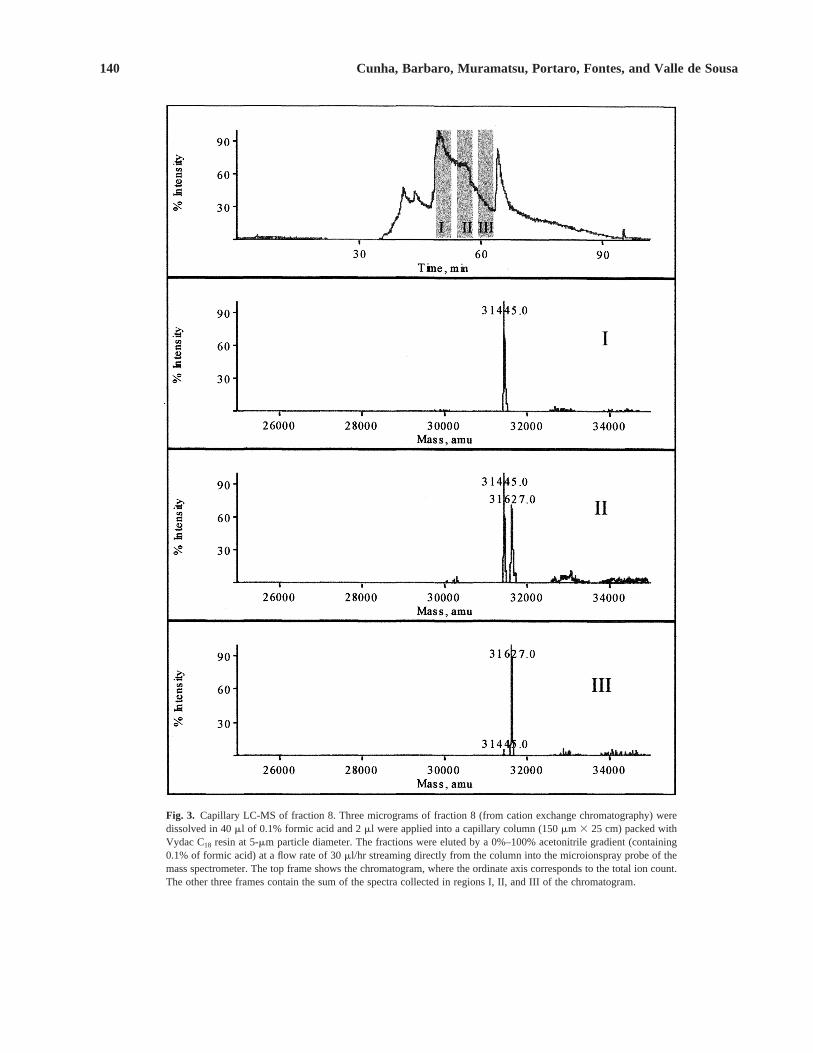
The dermonecrotic fraction 8 was then submitted toa cLC-MS experiment to characterize its protein content(Fig. 3). Region I of the chromatogram showed only amass of 31,445 6 3 Da, whereas region III containedmainly a peak of 31,627 6 4 Da. Region II showed amixture of the two peaks presented in regions I and III.The two peaks before region I on the chromatogram car-ried smaller proteins and peptides, and the peak after re-gion III was from the solvent. This result suggested theexistence of two isoforms of dermonecrotic protein,which was called loxnecrogin here.
Fraction 8 was further resolved by reversed-phasechromatography, resulting in two peaks: a major peak8A and a minor peak 8B (Fig. 4). ESI-MS of fractions8A and 8B showed the presence of two molecules:loxnecrogin A of 31,444 6 3 Da, more intense in frac-tion 8A, and loxnecrogin B of 31,626 6 4 Da, more in-tense in fraction 8B, which is notably contaminated byloxnecrogin A (Fig. 5). An injection peak was the firstone to be seen on the chromatogram, and showed no pro-tein masses associated to it by ESI-MS (data not shown).
The dermonecrotic activities of fractions 8A and 8Bwere tested in rabbits, and it was found that both frac-tions 8A and 8B induced necrosis, but the lesion areaswere smaller than for fraction 8 (Table 1). Neither 8Anor 8B alone could induce death in mice, in contrast tofraction 8. The small peak eluting immediately after 8Bon the RP-HPLC chromatogram (Fig. 4) was negativefor dermonecrotic and lethal activities. To check if thelethal activity was related to the combination of the two
138 Cunha, Barbaro, Muramatsu, Portaro, Fontes, and Valle de Sousa
Fig. 1. Chromatographic profile of L. gauchovenom fractionated bycation exchange FPLC. Proteins (samples of 1 mg) bound to thecolumn (MonoS HR 5/5) were eluted with a linear gradient of NaCl(0–1 mol/L) in 50 mmol/L sodium acetate buffer, pH 5.0. Samples of1.0 ml each were collected and their protein content was monitored byabsorbance at 280 nm.

sequencing. The N-terminal sequence ofloxnecrogin A(ADNRRPIWVMGHMVNSLAQIDEFVNLGANSIETDV) showed a high degree of similarity with N ter-mini of other dermonecrotic toxins isolated from venomsof Loxoscelesspp. (see Fig. 8). Only the N-terminal re-gions of these toxins were sequenced and deposited inthe databanks, but no internal sequence data were avail-able for them. Such a lack of information led us to pro-duce some internal sequence for loxnecrogin A by tan-dem MS (MS/MS) technique.
The reduced and vinylpyridylethylated loxnecro-gin A was cleaved with trypsin. The resulting peptidemap is shown in Fig. 6. Many peptides from the trypsindigest were subjected to MS/MS generating spectrafrom which some internal sequences could be assigned(Fig. 7).
Neither the peptide fingerprint nor the sequencestags searches showed any statistically significant hit, pos-sibly because there are no similar sequences depositedin the available databanks. However, a recent report(Kalapothakis et al., 2002) showed the complete se-quence (obtained by cDNA sequencing) of a 31,253-Dadermonecrotic toxin (LiD1) isolated from the venom ofL. intermedia. Figure 8 shows the deduced LiD1 proteinsequence aligned with fragment sequences of loxnecro-gin (MS/MS, Fig. 7, and Edman sequencing) and withother loxoscelic dermonecrotic toxins. Since isobaricamino acid cannot be distinguished by MS/MS sequenc-ing, the amino acids at the positions 87, 190, 254, and262 in loxnecrogin sequence may be Gln or Lys. Simi-larly, the amino acids at the positions 88, 93, 194, and256 may be Leu or Ile. In Figs. 7 and 8, we assigned theamino acid residues of loxnecrogin partial sequence ac-cording to their similarity with LiD1 sequence, and con-sidering the fact that Lys (and not Gln) is probably theC-terminal residue of a tryptic peptide.
4. DISCUSSION
Although the entire mechanism of action of Lox-oscelesvenom is not fully understood, several studieshave demonstrated that the development of the der-monecrotic lesion is due to several factors involving thedirect action of the venom on the tissue and the organ-ism’s response to this aggression. The initial interactionbetween the venom and the tissue could trigger endoge-nous mechanisms such as release of proteolytic enzymesfrom polymorphonuclear leukocytes (Smith and Micks,1970), platelet aggregation (Kurpiewski et al.,1981) andactivation of the complement system (Tambourgi et al.,1995). The venom also degrades components of the
Purification and Characterization of Loxnecrogin 139
Fig. 2. SDS-PAGE of L. gauchovenom and its fractions obtained bycation exchange FPLC. Three micrograms of the venom (V) and 500 ngof the fractions were analyzed by SDS-PAGE (12% polyacrylamidegels) under nonreducing conditions. Gels were silver stained. Numbersat the left side correspond to the molecular masses (kDa) of markers.
Table 1. Local Reaction, Dermonecrotic, and Lethal Activity ofL. gauchoFractions
Dermonecrosisa Local reactiona LethalFractions (cm2) (cm2) activityb
1 ND ND 0/62 ND 0.5 6 0.1 0/63 ND ND 0/64 ND 1.1 6 0.2 0/65 ND 1.7 6 0.9 0/66 ND 0.3 6 0.1 0/67 ND 3.1 6 0.7 0/68 1.0 6 0.7 31.1 6 5.7 6/68A 0.5 6 0.3 25.5 6 5.8 0/68B 0.5 6 0.2 14.7 6 4.2 0/69 ND 3.7 6 2.6 0/6
10 ND 0.8 6 0.3 0/611 ND ND 0/612 ND 0.5 6 0.2 0/613 ND ND 0/614 ND ND 0/615 ND 1.5 6 0.7 0/6
Venom 2.1 6 0.8 20.9 6 6.7 6/6
a The dorsum of shaved rabbits were injected, intradermally, with200 m of 0.15 mol/L NaCl containing the venom (3.0 mg) or thefraction (0.5 mg) obtained by FPLC. Data are expressed as mean 6
SD. ND means “not detectable.” The size of the affected areas wasmeasured after 48 hr. Local reactions are characterized by edema,erythema, and hemorrhagic and pale areas.
b Swiss mice were injected, intradermally, with 200 m of 0.15 mol/L NaClcontaining 2 LD50 of venom or 1/2LD50 of each fraction. The mortalitywas verified 48 hr after the injections. The results are expressed as num-ber of deaths per number of injected mice.
fractions, we produced a mixture of 8A and 8B; how-ever, this combination was not lethal to mice.
Loxnecrogin A was reduced and vinylpyridylethy-lated, and then submitted to automated N-terminal Edman
140 Cunha, Barbaro, Muramatsu, Portaro, Fontes, and Valle de Sousa
Fig. 3. Capillary LC-MS of fraction 8. Three micrograms of fraction 8 (from cation exchange chromatography) weredissolved in 40 ml of 0.1% formic acid and 2 ml were applied into a capillary column (150 mm 3 25 cm) packed withVydac C18 resin at 5-mm particle diameter. The fractions were eluted by a 0%–100% acetonitrile gradient (containing0.1% of formic acid) at a flow rate of 30 ml/hr streaming directly from the column into the microionspray probe of themass spectrometer. The top frame shows the chromatogram, where the ordinate axis corresponds to the total ion count.The other three frames contain the sum of the spectra collected in regions I, II, and III of the chromatogram.

extracellular matrix (Feitosa et al., 1998, Souza et al.,1998; Veiga et al., 2001), causing the tissue injury totravel following gravity (Futrell, 1992). All of thesemechanisms would contribute to an increase of the lesionthrough damage of the microvascularization system, dis-turbances in blood flow, induction of edema and is-chemic action. These would lead to cellular degenerationand damage of the local tissue.
Barbaro et al.,(1996) showed that 35-kDa L. gauchovenom components were the main components responsi-ble for dermonecrotic and lethal venom activities. Herewe confirmed by gel image analysis of SDS-PAGE re-sults that L. gauchodermonecrotic toxin falls at approx-imately 30 kDa. By MS, we were able to more accuratelydetermine the mass of the toxin as 31,444 Da (loxnecro-gin A), accompanied by an isoform of 31,626 Da (lox-necrogin B). More accurate measurements could simi-larly be made for dermonecrotic toxins of other Loxoscelesspiders if higher accuracy methods for molecular massdetermination, such as MS, were used.
Mass spectrometric data demonstrated a molecularmicrodiversity of loxnecrogin, with the presence of twoisoforms, differing by 182 6 5 Da between each other.Further MS experiments carried out with more accurateequipment should be performed to determine the aminoacid residue substitutions between the isoforms.
The presence of isoforms of Loxoscelessphingo-myelinases had been described previously byKurpiewski et al. (1981), who separated four active iso-forms of 32-kDa protein from L. reclusavenom (pI of8.7, 8.4, 8.2, and 7.8) by isoelectric focusing. All theseisoforms presented sphyngomyelinase D activity, wereable to develop dermonecrosis in rabbits, and offeredcross-reactivity, suggesting structural homology. How-ever, it was not clear if all the four L. reclusasphyngo-myelinase D varieties were originally present in vivo orwere formed by chemical modification due to the am-pholines in the isoelectric focusing experiment. Tam-bourgi et al. (1998) purified two isoforms of the der-monecrotic toxin from L. intermediaand sequenced theN-terminal regions of them, showing three residue substi-tutions in the 29 first amino acid residues. A third partiallypurified isoform did not yield an unambiguous N-terminalsequence.
In a recent report, Kalapothakis et al.(2002) clonedand expressed a dermonecrotic toxin from L. intermediavenom (LiD1) and used the recombinant purified protein(recLiD1) to produce anti-recLiD1 antibodies. LiD1 wascloned based on the sequence of the native form of thedermonecrotic toxin from L. intermediatoxin. Then,the recombinant toxin recLiD1 was supposed to presentsimilar activity. However, recLiD1 did not present anyactivity. A tentative explanation for that is a possible mis-folding of the recombinant protein due to errors in disul-fide bond formation and the presence of b-galactosidasein the recombinant fusion protein (Kalapothakis et al.,2002). Also, the immunization power of the anti-recLiD1antibodies is less than the antibodies generated againstthe dermonecrotic fraction or the whole venom, possiblydue to the structural difference between native and re-combinant molecules or the presence of different isoformsof the dermonecrotic family in the whole venom and inthe purified fraction, which are absent in the antibodyproduction with recLiD1.
The decrease of dermonecrotic and lethal activitydetected on fraction 8A and 8B after HPLC separation(Table 1) suggested that more than one venom compo-nent might be involved in tissue damage as observed byGuilherme et al., (2001) using monoclonal antibodies.Alternatively, some authors (Sadler et al.,1984; Wu et al.,1986; Herold et al.,1991) have reported conformationalchanges of proteins under typical HPLC conditions,
Purification and Characterization of Loxnecrogin 141
Fig. 4. Reversed-phase chromatographic profile of fraction 8 fromL. gaucho venom. Fraction 8, obtained from cation-exchangechromatography, was submitted to reversed-phase HPLC in a wide-pore butyl C4 Vydac column (4.6 3 150 mm, 5-mm particle size) undera linear gradient of 0%–70% acetonitrile in 0.1% TFA at a flow rate of1 ml/min. Protein content was monitored by absorbance at 216 nm.

determined for loxnecrogin A because the former prepa-ration was just in a partially pure state while the latterwas obtained in a high degree of purity.
The presence of sphingomyelinase activity in Lox-oscelesvenom has been known (Forrester et al., 1978).This enzyme acts through a mechanism that involves thehydrolysis of sphingomyelin with the respective libera-tion of ceramide, which is a known mediator of apopto-sis. Recent results obtained by our group showed thatloxnecrogin was able to hydrolyze sphingomyelin. Acomplete report on loxnecrogin9s enzymatic characteris-tics will be published elsewhere (Portaro et al.,personalcommunication).
To further characterize loxnecrogin, we generated apeptide mass map (Fig. 6) followed by MS/MS spectra(Fig. 7) from peptides of a tryptic digest of the isoformA. Internal sequences were deduced by de novose-quencing (Fig. 7). No relevant matches were revealed byeither peptide fingerprint or sequence searching strate-
142 Cunha, Barbaro, Muramatsu, Portaro, Fontes, and Valle de Sousa
which could also explain the decrease of activities. Theloxnecrogin partially lost its activity upon HPLC, proba-bly due to change in conformation by the contact withacidic and organic solvent. This suggests that the main-tenance of the correct toxin folding is important forpreservation of its activity. Use of MS combined withcircular dichroism and other physical methods to probepossible conformational changes of the loxnecroginscould shed light on the causes of the loss of activities.
The N-terminal sequence of loxnecrogin A showeda high degree of similarity with other dermonecrotic tox-ins, some of them possessing described sphingomyeli-nase activity, from venoms of some spiders of the genusLoxosceles(Fig. 8). Unfortunately, only the N-terminalregion of these proteins had been chemically sequencedand deposited in the databanks. The L. gauchotoxin se-quence showed in Fig. 8 (AAB50794) may actually bethat of one of the loxnecrogin isoforms. However, its se-quence might not be as accurate as the present sequence
Fig. 5. ESI-MS of fractions 8A and 8B. Fractions 8A and 8B (from reversed-phase HPLC) were collected, lyophilized, dissolved in 50% acetonitrile(v/v) in 5% acetic acid (v/v), at a final concentration of 5 pmol/ml, and injected using the nanoelectrospray ionization probe at a flow rate of 10–40nL/min. The data were collected at the positive mode. The figure shows the reconstructed spectra.

gies. Perhaps loxnecrogin and the dermonecrotic toxinsfrom other Loxoscelesspecies will constitute a new toxinfamily.
The partial sequences of loxnecrogin, obtained byboth chemical sequencing and MS/MS approaches, pre-sented many sequence similarities with the deduced se-quence of L. intermediaLiD1 toxin (Fig. 8). The de-duced molecular mass of the mature LiD1 toxin (31,253Da) and the MS-determined masses of loxnecrogin A(31,444 Da) and B (31,626 Da) are also very similar toeach other.
Despite the overall similarities, some specific dif-ferences were found between the LiD1 and loxnecroginA sequences. So far, loxnecrogin presented the substitu-tions G2D, I9V, A16S, I17L, G18A, L86Q, I188R,S191F, D192G, and D254K in relation to LiD1. The twosequences have 86% of identity and 90% of similarityusing the BLOSUM62 matrix (Henikoff and Henikoff,1992). According to Henikoff and Henikoff (1992) crite-ria, the substitutions I9V, A16S, and I17L are conserva-
tive, the substitutions G2D, G18A, D192G, and D254Kare semiconservative, and the substitutions L86Q,I188R, and S191F are nonconservative. Differences be-tween the sequences agree with the hypothesis that im-portant changes can be found among the epitopes pres-ent in the dermonecrotic proteins of Loxoscelesvenomsas described previously by Guilherme et al.(2001) usingmonoclonal antibodies.
All these structural and conformational differencesas well as the presence of some active isoforms amongthe venoms of different Loxoscelesspecies should beconsidered to improve the loxoscelism serum therapy.
ACKNOWLEDGMENTS
We are grateful to Dr. Dário E. Kalume for his con-tribution in interpretation the MS/MS data, Mr. ElvioDias Botelho for his technical assistance, and Dr. RogerKautz, Barnett Institute, Boston, MA, for critical reading
Purification and Characterization of Loxnecrogin 143
Fig. 6. Peptide mass fingerprint of loxnecrogin A. The tryptic digest was submitted to electrospray mass spectrometry. The peaks marked by anasterisk (*) produced good informative MS/MS spectra, which are shown in Fig. 7.

144 Cunha, Barbaro, Muramatsu, Portaro, Fontes, and Valle de Sousa
Fig
. 7.
MS
/MS
dat
a ob
tain
ed b
y el
ectr
ospr
ay t
ande
m m
ass
spec
trom
etry
und
er n
itrog
en c
ollis
ion-
indu
ced
diss
ocia
tion
(CID
). T
he p
eaks
wer
e se
lect
ed b
ased
on
the
pept
ide
mas
sfin
gerp
rint
of lo
xnec
rogi
n A
(F
ig.
6).

Purification and Characterization of Loxnecrogin 145
Fig
. 8.
Alig
nmen
t of
the
seq
uenc
e fr
agm
ents
of
loxn
ecro
gin
A a
nd o
ther
der
mon
ecro
tic t
oxin
seq
uenc
es.
LiD
1 re
fers
to
the
com
plet
e pr
ote
in s
eque
nce,
ded
uced
fro
m t
he c
DN
A s
eque
nce,
of a
der
mon
ecro
tic p
rote
in i
sola
ted
from
the
ven
om o
f L.
int
erm
edia
(Kal
apot
haki
s et
al.,
2002
). A
3810
7 is
the
PIR
acc
essi
on n
umbe
r of
a d
erm
onec
rotic
tox
in p
artia
l se
quen
ce f
rom
L.r
ecl
usa
(Cis
ar e
t a
l.,19
89).
AA
B50
794,
AA
B50
795,
and
AA
B50
796
are
the
Gen
Ban
k ac
cess
ion
num
bers
for
der
mon
ecro
tic t
oxin
N-t
erm
inal
seq
uenc
es f
rom
L
. g
au
cho
, L
. in
term
ed
ia,
and
L.
lae
ta,r
espe
ctiv
ely
(Bar
baro
et a
l.,19
96).
P83
045
and
P83
046
are
the
Sw
issP
rot
acce
ssio
n nu
mbe
rs f
or s
phin
gom
yelin
ases
P1
and
P2
sequ
ence
s, r
espe
ctiv
ely,
bot
h fr
omL
.in
term
ed
ia(T
ambo
urgi
et
al.,
1998
). T
he u
nder
lined
seq
uenc
e of
lox
necr
ogin
A r
epre
sent
s th
e N
-ter
min
al s
eque
nce
obta
ined
che
mic
ally
, ev
en t
houg
h pa
rt o
f th
at
sequ
ence
was
confi
rmed
by
MS
/MS
seq
uenc
ing.
Pai
rs o
f am
ino
acid
s Le
u/Ile
and
Gln
/Lys
can
not
be d
istin
guis
hed
by M
S/M
S s
eque
ncin
g fo
r be
ing
isob
aric
and
wer
e as
sign
ed i
n lo
xnec
rogi
n pa
rtia
lse
quen
ces
acco
rdin
g to
the
ir si
mila
rity
with
LiD
1 se
quen
ce,
and
cons
ider
ing
the
fact
tha
t Ly
s (a
nd n
ot G
ln)
is p
roba
bly
the
C-t
erm
inal
res
idue
of
a tr
yptic
pep
tide.

of the manuscript. This work was funded in part byFAPESP (São Paulo, Brazil, grants 97/14187-0 and97/09681-6), CNPq-PADCT (Brasília, Brazil, grant62.0545/98-4), and Universidade de Brasília, Brazil.
REFERENCES
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J.(1990). J. Mol. Biol.215: 403–410.
Andrade, R. M. G., Oliveira, K. C., Giusti, A. L., Silva, W. D., andTambourgi, D. V. (1999). Toxicon37: 627–632.
Barbaro, K. C., Cardoso, J. L. C., Eickstedt, V. R. D., and Mota, I.(1992). Toxicon30: 331–338.
Barbaro, K. C., Eickstedt, V. R. D., and Mota, I. (1994). Toxicon32:113–120.
Barbaro, K. C., Sousa, M. V., Morhy, L., Eickstedt, V. R. D., andMota, I. (1996). J. Protein Chem.15: 337–343.
Barreto, O. C. O., Cardoso, J. L. C., and De Cillo, D. (1985). Rev. Inst.Med. Trop. São Paulo27: 264–267.
Blum, H., Beier, H., and Gross, H. J. (1987). Electrophoresis8: 93–95.
Bradford, M. M. (1976). Anal. Biochem.72: 248–254.Cardoso, J. L. C., França, F. O. S., Eickstedt, V. R. D., Borges, I., and
Nogueira, M. T. (1988). Rev. Soc. Bras. Toxicol.1: 58–60.Cisar, C. R., Fox, J. W., and Geren, C. R. (1989). Toxicon27(37).Elgert, K. D., Ross, M. A., Campbell, B. J., and Barrett, J. T. (1974).
Infect. Immun.10: 1412–1419.Feitosa, L., Gremski, W., Veiga, S. S., Elias, M. C. Q. B., Graner, E.,
Mangili, O. C., et al. (1998). Toxicon36: 1039–1051.Finney, D. J. (1971). Probit Analysis, 3rd Ed. Cambridge University
Press, Cambridge.Fontes, W., Cunha, R. B., Sousa, M. V., and Morhy, L. (1998). Anal.
Biochem.258: 259–267.Forrester, L. J., Barrett, J. T., and Campbell, B. J. (1978). Arch.
Biochem. Biophys.187: 355–365.Futrell, J. M. (1992). Loxoscelism. Am. J. Med. Sci.304: 261–267.Geren, C. R., Chan, T. K., Howell, D. E., and Odell, G. V. (1976).
Arch. Biochem. Biophys.174: 90–99.Guilherme, P., Fernandes, I., and Barbaro, K. C. (2001). Toxicon39:
1333–1342.Heitz, J. R., and Norment, B. R. (1974). Toxicon12: 181–187.Henikoff, S., and Henikoff, J. G. (1992). Proc. Natl. Acad. Sci. USA
89: 10915–10919.
Herold, M., Rozing, G. P., and Curtis, J. L. (1991). Biotechniques10:656–662.
Jong, Y. S., Norment, B. R., and Heitz, J. R. (1979). Toxicon17: 529–537.
Kaiser, E. (1956). Enzymatic activity of spider venoms. In: Buckley,E. E., and Porges, N. (eds.), Venoms, American Association forthe Advancement of Science, Washington, pp. 91–93.
Kalapothakis, E., Araújo, S. C., Castro, C. S., Mendes, T. M., Gomez,M. V., Mangili, O. C., et al. (2002). Toxicon40: 1691–1699.
Kurpiewski, G., Forrester, L. J., Barret, J. T., and Campbell, J. (1981).Biochim. Biophys. Acta678: 467–476.
Laemmli, U. K. (1970). Nature227: 680–685.Ministry of Health (1998). Brazil: Manual de Diagnóstico e Trata-
mento de Acidentes por Animais Peçonhentos, CENEPI, Fun-dação Nacional de Saúde/Coordenação de Controle de Zoonosese Animais Peçonhentos, Brasília, pp. 57–61.
Norment, B. R., Jong, Y. S., and Heitz, J. R. (1979). Toxicon17: 539–548.
Oliveira, K. C., Andrade, R. M. G., Giusti, A. L., Silva, W. D., andTambourgi, D. V. (1999). Toxicon37: 217–221.
Perkins, D. N., Pappin, D. J., Creasy, D. M., and Cottrell, J. S. (1999).Electrophoresis20: 3551–3567.
Rash, L. D., and Hodgson, W. C. (2002). Toxicon40: 225–254.Sadler, A. J., Micanovic, R., Katzenstein, G. E., Lewis, R. V., and
Middaugh, C. R. (1984). J. Chromatogr.317: 93–101.Schenone, H., and Suarez, G. (1978). Venoms of Scytodidae. In: Bet-
tini, S. (ed.), Handbook of Experimental Pharmacology, Vol. 48,Springer-Verlag, New York, pp. 247–275.
Sezerino, U. M., Zannin, M., Coelho, L. K., Gonçalves, J. R. J.,Grando, M., Mattosinho, S. G., et al.(1998). Trans. R. Soc. Trop.Med. Hyg.92: 546–548.
Smith, C. W., and Micks, D. W. (1970). Lab. Invest.22: 90–93.Souza, G. A., Ribeiro, A. S., Santos, V. L. P., Veiga, S. S., Mangili,
O. C., and Gremski, W. (1998). Acta Biol. Par.27: 97–109.Tambourgi, D. V., Magnoli, F. C., van den Berg, C. W., Morgan, B. P.,
Araujo, P. S., Alves, E. W., et al. (1998). Biochem. Biophys. Res.Commun.251: 366–373.
Tambourgi, D. V., Mangoli, F. C., von Eickstedt, V. R., Benedetti,Z. C., Petricevich, V. L., and da Silva, W. D. (1995). J. Immunol.155: 4459–4466.
Veiga, S. S., Zanetti, V. C., Braz, A., Mangili, O. C., and Gremski, W.(2001). Braz. J. Med. Biol. Res.34: 843–850.
Wright, R. P. (1973). Bull. Mo. Acad. Sci.(Suppl. 2): 1–94.Wu, S. L., Figueroa, A., and Karger, B. L. (1986). J. Chromatogr.371:
3–27.Young, A. R., and Pincus, S. J. (2001). Toxicon39: 391–400.
146 Cunha, Barbaro, Muramatsu, Portaro, Fontes, and Valle de Sousa




















