
Monitoring cerebral oxygenation during balloon occlusion with multichannel NIRS
Upload
independentCategory
view
0download
0
ORIGINAL ARTICLE
Protective Effects of Cannabidiol Against Hippocampal CellDeath and Cognitive Impairment Induced by Bilateral CommonCarotid Artery Occlusion in Mice
Angelica Pupin Schiavon • Lıgia Mendes Soares • Jessica Mendes Bonato •
Humberto Milani • Francisco Silveira Guimaraes • Rubia Maria Weffort de Oliveira
Received: 29 October 2013 / Revised: 13 January 2014 / Accepted: 2 February 2014
� Springer Science+Business Media New York 2014
Abstract The present study investigated whether canna-
bidiol (CBD), a major non-psychoactive constituent of
marijuana, protects against hippocampal neurodegenera-
tion and cognitive deficits induced by brain ischemia in
adult mice. Male Swiss mice were subjected to a 17 min of
bilateral common carotid artery occlusion (BCCAO) and
tested in the Morris water maze 7 days later. CBD (3, 10,
and 30 mg/kg) was administered 30 min before and 3, 24,
and 48 h after BCCAO. After behavioral testing, the brains
were removed and processed to evaluate hippocampal cell
survival and degeneration using Nissl staining and Flu-
oroJade C histochemistry, respectively. Astroglial response
was examined using immunohistochemistry for glial
fibrillary acidic protein (GFAP). CBD (3–30 mg/kg)
improved spatial learning performance in BCCAO mice.
The Nissl and FJC staining results showed a decrease in
hippocampal neurodegeneration after CBD (10 and 30 mg/
kg) treatment. GFAP immunoreactivity was also decreased
in ischemic mice treated with CBD (30 mg/kg). These
findings suggest a protective effect of CBD on neuronal
death induced by ischemia and indicate that CBD might
exert beneficial therapeutic effects in brain ischemia. The
mechanisms that underlie the neuroprotective effects of
CBD in BCCAO mice might involve the inhibition of
reactive astrogliosis.
Keywords Cannabidiol � Morris water maze �Hippocampus � Bilateral common carotid artery occlusion �Mice
Introduction
Ischemic injury that results from global circulatory arrest in
the brain is one of the major causes of death and disability
in the adult population (Nikonenko et al. 2009; Neumann
et al. 2013). Prolonged circulatory deficits induce irre-
versible changes in brain tissue. The resulting neuronal
damage and death lead to a broad range of neurological and
behavioral dysfunctions (Anderson and Arciniegas 2010;
Peskine et al. 2010). Cognitive impairments are among the
better characterized sequelae following brain ischemia,
with memory being the most affected domain, followed by
attention and executive function (Zola-Morgan et al. 1986;
Kartsounis et al. 1995; Moulaert et al. 2010; Mateen et al.
2011). Post mortem studies have suggested that hippo-
campal damage may be a key factor associated with cog-
nitive impairment after ischemic events (Zola-Morgan
et al. 1986; Bachevalier and Meunier 1996; Blum et al.
2012). Despite intense efforts, few pharmacological treat-
ments have effectively minimized the functional impair-
ments following cerebral ischemia (Auriel and Bornstein
2010). Therefore, studies that seek to identify safe and
effective strategies for the treatment of ischemic brain
disease are needed.
A considerable number of preclinical studies have
demonstrated that cannabidiol (CBD), a major non-psy-
choactive constituent of marijuana, may provide neuro-
protection against acute or chronic brain damage
(Mechoulam and Shohami 2007; Garcia et al. 2011; Sag-
redo et al. 2011; Fernandez-Ruiz et al. 2013; Harvey et al.
A. P. Schiavon � L. M. Soares � J. M. Bonato � H. Milani �R. M. Weffort de Oliveira (&)
Department of Pharmacology and Therapeutics, State University
of Maringa, Av. Colombo, 5790, Maringa, PR 87020-900, Brazil
e-mail: [email protected]
F. S. Guimaraes
Department of Pharmacology, School of Medicine, USP, Av.
Bandeirantes, Ribeirao Preto, SP 14015-000, Brazil
123
Neurotox Res
DOI 10.1007/s12640-014-9457-0

2012; Pazos et al. 2012, 2013; Valdeolivas et al. 2012).
In vitro, CBD produces a significant reduction of b-amy-
loid-induced neuronal death because of its ability to scav-
enge reactive oxygen species, reduce lipid peroxidation
(Iuvone et al. 2004), and decrease the expression of the
inducible form of nitric oxide synthase (iNOS) and inter-
leukin-1b (IL-1b) expression (Castillo et al. 2010). CBD
also reduces necrotic and apoptotic damage in forebrain
slices in newborn mice exposed to oxygen-glucose depri-
vation (Castillo et al. 2010). Moreover, CBD has been
shown to protect the immature brain from hypoxic-ische-
mic (HI) injury. The administration of CBD 30 min after
HI insult in newborn piglets (Lafuente et al. 2011), mice
(Castillo et al. 2010), and rats (Pazos et al. 2012) resulted
in histological, functional, and biochemical improvements.
Repeated treatment with CBD (3 mg/kg) also decreased
glial activation and improved survival rates after middle
cerebral artery occlusion (MCAO) in mice (Hayakawa
et al. 2008, 2009). From a functional perspective, Pazos
et al. (2012) showed that CBD improved the neurological
score (i.e., cycling, rolling, and leaning to the ipsilateral
side of the lesion) and motor coordination in MCAO mice.
Moreover, CBD (5 mg/kg) administered 5 min after bilateral
common carotid artery occlusion (BCCAO) antagonized
electroencephalographic flattening and hyperlocomotion in
gerbils (Braida et al. 2003).
To date, only one study has evaluated the effects of
CBD on cognitive deficits induced by experimental brain
ischemia. Pazos et al. (2012) showed that CBD (1 mg/kg)
administration in newborn rats after HI injury led to long-
lasting neuroprotective effects, reflected by improved
neurobehavioral performance in both sensorimotor tests
and the novel object recognition test 30 days after the
insult. The latter is considered a non-spatial memory test.
Remaining to be determined, however, the effects of CBD
on spatial memory deficits induced by brain ischemia in
adult animals. Therefore, the aim of this study was to
investigate the effects of CBD on the ability to perform a
spatial learning and memory task (the Morris water maze)
in adult mice after BCCAO. Because astrocytes play a
fundamental role in the pathogenesis of neuronal death
(Takuma et al. 2004; Szydlowska et al. 2010; Duan et al.
2011), we also examined the effects of CBD on hippo-
campal cell survival and astroglial response to experi-
mental brain ischemia.
Materials and Methods
Animals
Male Swiss albino mice (30–40 g, 35–45 days old) were
obtained from the central vivarium of the State University
of Maringa, Maringa, Brazil. Prior to and throughout the
experiments, the animals were maintained under conditions
of controlled temperature (22 ± 1 �C) with a 12/12 h light/
dark cycle (lights on at 7:00 AM). The animals were
housed in groups (n = 3–5) and given standard commer-
cial chow and tap water ad libitum. A total of 81 mice were
included in the experiments. Of these, 15 mice were
assigned to sham operation, and 66 were subjected to brain
ischemia. They were then, randomly distributed into the
independent experimental groups. The experimental pro-
cedures were approved by the Ethics Committee on Animal
Experimentation of the State University of Maringa (CEEA
004/2011) and are in accordance with the guidelines of
NIH and Brazilian College for Animal Experimentation.
Surgery
Transient global cerebral ischemia was induced by
BCCAO as previously described (Soares et al. 2013). The
mice were anesthetized with a mixture of isoflurane
(Isoforine, Cristalia, SP, Brazil) and oxygen delivered
through a universal vaporizer (Oxigel, SP, Brazil) con-
nected to a plastic mask adapted to the animal’s nose. The
vaporizer was regulated to release the minimal burble flow
(2.0 l/min). The mixture delivered to the animal was
monitored to maintain the minimal isoflurane concentration
required for an efficient anesthesia (evaluated by pinching
the animal’s tail). Under these conditions, the animal was
fixed in a stereotaxic frame and the anesthesia maintained
with 1.3-1.5 % isoflurane in 100 % oxygen during
*6 min, time necessary to make an incision in the ventral
neck to expose the common carotid arteries. Rectal tem-
perature was carefully monitored during surgery and
maintained at *37.5 �C using a heating blanket. Brain
ischemia was induced by 17 min of BCCAO using aneu-
rysm clips (ADCA, MG, Brazil). Throughout the occlusion
procedure, the mice were maintained in a warming box
(inner temperature, 30 ± 1 �C) to avoid ischemia-induced
brain hypothermia (Seif el Nasr et al. 1992). At the end of
each occlusion, the aneurism clips were removed, and the
carotid arteries were visually inspected for reperfusion.
Then, each animal was again anesthetized for 2 min and
the incision closed with sutures. For 3 h after reperfusion,
the mice were maintained in a warming box at 30 �C.
Sham-operated animals were subjected to the same anes-
thetic and surgical interventions, with the exception that
the carotid arteries remained intact.
Treatment
Vehicle (0.9 % NaCl with 1 % Tween 80) or CBD (THC
Pharma, Germany) was prepared immediately before use
and injected intraperitoneally (i.p.) in a 1 ml/kg volume.
Neurotox Res
123

The animals were randomly assigned to receive vehicle or
CBD (3, 10, or 30 mg/kg) 30 min before and 3, 24, and
48 h after surgery. The treatment regimen and doses of
CBD were based on Hayakawa et al. (2008) and Campos
et al. (2012).
Morris Water Maze
The behavioral testing occurred during the light phase
between 8:00 AM and 2:00 PM under identical conditions.
The experiments were video-recorded, and the behavioral
scores were later analyzed using ANY-maze image ana-
lyzer software (Stoelting, Wood Dale, IL, USA). All of the
behavioral tests were conducted 7 days after BCCAO
because the neuropathological and neurobehavioral con-
sequences of BCCAO in Swiss mice are well established
over that time interval so the neuroprotective effect of a
given strategy can be properly assessed (Soares et al.
2013).
The Morris water maze apparatus consisted of a swim-
ming pool made of black painted fiberglass (90 cm diam-
eter, 35 cm height). For the tests, the tank was filled with
water maintained at 24 ± 1 �C. The target platform
(10 cm2) was made of transparent acrylic and submerged
1 cm beneath the surface of the water. The starting points
for the animals were marked on the outside of the pool as
north (N), south (S), east (E), and west (W). Four distant
visual cues were placed on the walls of the experimental
room.
The mice were subjected to the Morris water maze using
a procedure that was similar to one described previously
(Prediger et al. 2008; Soares et al. 2013). In brief, the
training session consisted of ten consecutive trials, during
which the animals were left in the tank facing the wall, and
then allowed to swim freely to the submerged platform.
The platform was located in a constant position (middle of
the southwest quadrant), equidistant from the center and
wall of the pool. If the animal did not find the platform
during a 60 s trial period, then it was gently guided to it.
The animal was allowed to remain on the platform for 10 s.
This procedure was repeated ten times, with the starting
point (i.e., the axis of one imaginary quadrant) varying in a
pseudo-random manner. The latency to find the platform
was recorded for each animal. 24 h after the last training
trial (i.e., acquisition phase), the probe trial test (i.e.,
retention phase) began and consisted of a single trial in
which the platform was removed from the pool. The mouse
was allowed to swim for 60 s, and the time spent in the
correct quadrant (i.e., where the platform was located
during the training session) was recorded as an index of
memory strength.
Histology
Two histological groups were generated by random group
assignment. One group (n = 6–15 subjects/experimental
group) underwent histological assessment of hippocampal
cell survival using Nissl staining. A second group (n = 5–8
subjects/experimental group) underwent Fluorojade-C
(FJC) staining and immunohistochemical assays. Coronal
brain sections were obtained at a stereotaxic level between
-1.70 and -2.70 mm posterior to bregma (Franklin and
Paxinos 1997). The histological analysis was performed
blind to the treatment groups.
Nissl Staining
The mice were deeply anesthetized with 50 mg/kg sodium
thiopental (Thiopentax; Cristalia, SP, Brazil) and tran-
scardially perfused with 0.9 % saline followed by Bouin’s
fixative. Following decapitation, the head was immersed in
crushed ice (1–2 �C) for 2 h to avoid the appearance of
dark neurons, which could confound the actual extent of
neurodegeneration. The brain was carefully removed and
postfixed in Bouin’s solution for 3 days. Using a rotating
microtome (RM2445, Leica, Goettingen, Germany), 7 lm
paraffin-embedded coronal sections were cut and distrib-
uted into four sets of slides that contained four coronal
sections, each 28 lm apart. After standard dehydration and
diaphanization procedures, slides that contained adjacent
sections were immersed in distilled water and submerged
in 0.2 % Cresyl violet solution (Nissl staining) for 5 min.
The slides were then rinsed in distilled water, dehydrated in
a graded series of ethanol (70, 80, 90, and 100 %), cleared
in xylene, and coverslipped with xylene using Permount
(Fisher Scientific, Sao Paulo, Brazil). In each hemisphere,
the number of cells that presented a well delimited,
spherical form with a distinct nucleus and nucleolus was
counted throughout the CA1, CA2, CA3, and CA4 sub-
fields of the hippocampus (4009 magnification; Olympus
BX-41 microscope). Neurons that had shrunken cell bodies
or surrounding empty spaces were considered destined to
die and excluded from the counting. The number of intact-
appearing cells is expressed as the mean ± SEM.
Fluorojade-C and Immunohistochemistry
The animals were deeply anesthetized and transcardially
perfused with saline followed by 4 % paraformaldehyde in
0.2 M phosphate buffer (PB). The brains were removed,
postfixed in the same fixative for 2 h, and cryoprotected by
immersion in 30 % sucrose. Frozen tissue was serially
sectioned on a cryostat (Criocut 1800, Reichert-Jung,
Heidelberg, Germany) into 30 lm coronal sections that
Neurotox Res
123

were collected in quadruplicate in Eppendorf tubes that
contained antifreeze solution.
FJC is an anionic fluorochrome that detects neurons that
are undergoing the death process (Schmued et al. 2005).
Slides containing frozen sections were incubated in citrate
buffer for 30 min at 95 �C. The slides were then rinsed in
distilled water for 1 min and incubated in a 0.06 %
potassium permanganate solution for 15 min. The slides
were washed again in distilled water and incubated in
0.0001 % FJC (Histo-Chem, Jefferson, AR, USA) dis-
solved in a solution that contained 0.1 % acetic acid for
30 min in a dark room at 25 �C. They were then washed in
water, dried at room temperature, dehydrated, cleared in
xylene, and coverslipped after the addition of Permount
media. Using a fluorescence microscope (4009 magnifi-
cation, Zeiss microscope, Axiostar Plus, Jena, Germany),
FJC-positive cells were counted throughout the CA1-CA4
subfields in four coronal sections per animal from both
brain hemispheres. The number of FJC-positive cells is
expressed as the mean ± SEM.
Immunohistochemistry was performed for glial fibrillary
acidic protein (GFAP). The free-floating sections were
quenched in 1 % H2O2 for 30 min, and then blocked with
2 % bovine serum albumin in 0.1 M phosphate-buffered
saline (PBS) for 60 min. The sections were incubated
overnight with rabbit polyclonal anti-GFAP antibody
(1:500, Santa Cruz Biotechnology, Santa Cruz, CA, USA)
in PBS that contained 0.3 % Triton X-100 at room tem-
perature. The sections were then incubated with the
respective biotinylated secondary antibodies (1:500; Santa
Cruz Biotechnology, Santa Cruz, CA, USA) for 2 h and
further incubated in ABC solution (Vectastain Elite ABC
Kit, Vector Laboratories, Burlingame, CA, USA) for 2 h.
The peroxidase reaction was performed by incubating the
sections with 3-30-diaminobenzidine (DAB; Sigma) and
0.05 % H2O2. The sections were mounted on gelatin-
coated slides, air dried, cleared with xylene, and covers-
lipped with Permount.
GFAP immunoreactivity was evaluated by measuring
the integrated optical density (IOD) using ImageJ software
(National Institutes of Health, Bethesda, MD, USA). Color
images of the CA1 region from both brain hemispheres
were captured at 409 objective using a camera (QColor,
Olympus, America Inc.) and an Olympus BX 41 research
microscope. The region of interest was selected and a
previously defined area located between the CA1 pyrami-
dal cell layer and lacunosum molecular lay of the hippo-
campus (Franklin and Paxinos 1997) in both brain
hemispheres. The image was then converted to gray scale,
the background subtracted and, the IOD obtained. Results
were presented as the mean ± SEM of three sections/
animal.
Statistical Analysis
The SAS package (version 9.3) was used. Both behavioral and
histological data were examined for the assumptions of nor-
mality, homocedasticity, and sphericity (repeated measures).
In the Morris water maze test, learning performance was
measured across ten consecutive trials. The values obtained
for each mouse across two consecutive trials were aver-
aged, yielding five blocks with two trials per block. These
individual values (in blocks) were used to compute the
mean ± SEM for each group. A two-way repeated-mea-
sures analysis of variance (ANOVA), with experimental
group as the independent factor and test block (five blocks)
as the repeated factor, was performed. The sham- and
CBD-treated animals were compared with vehicle-treated
ischemic mice using a simple-contrast analysis. One-way
ANOVA was used to quantify the total latency (summed
over the 10 trials) and time spent in the correct quadrant.
The histological data were normalized to the mean
values of the sham- (Nissl and GFAP) or BCCAO-operated
(FJC) groups. The data were analyzed using the general-
ized linear model using the Poisson distribution for mod-
eling the count data (number of cells; Nissl and FJC) and
the Gamma distribution for the continuing measurements
(IOD; GFAP), respectively.
Differences with a probability value of p \ 0.05 were
considered statistically significant.
Results
Survival
Overall, 81 animals entered the experiment. Fifteen ani-
mals underwent sham surgery and were namely vehi-
cle ? sham group (veh sham; n = 15). Of the 66 mice
subjected to BCCAO, seven (10.6 %) died after complete
recovery from anesthesia, likely reflecting a severe and
fatal effect of brain ischemia. The remaining 59 ischemic
animals were randomly distributed into the following
experimental groups: vehicle ? ischemia (veh isch;
n = 23), CBD 3 mg/kg ? ischemia (CBD3 isch; n = 11),
CBD 10 mg/kg ? ischemia (CBD10 isch; n = 14), and
CBD 30 mg/kg ? ischemia (CBD30 isch; n = 11). 7 days
after sham or BCCAO surgery the animals were tested in
the Morris water maze task.
Morris Water Maze Task
Learning and memory performance in the Morris water maze
are shown in Fig. 1. The latency to find the platform was
measured in a single session (day) across 10 consecutive
Neurotox Res
123

trials and is expressed as the learning curve during five
blocks with two trials each (latency in blocks) and total
latency (summed across the 10 trials). The latency to find the
platform decreased regularly across the trials for all of the
experimental groups (F4,276 = 20.5, p \ 0.001). A signifi-
cant effect of treatment was found (F4,73 = 16.7,
p \ 0.001). The sham and ischemic animals treated with
CBD (3–30 mg/kg) exhibited a significant decrease in the
latency to find the platform compared with vehicle-treated
ischemic mice (Fig. 1a; p \ 0.05). No significant difference
was found in the time spent in the correct quadrant (Fig. 1c).
Nissl Staining
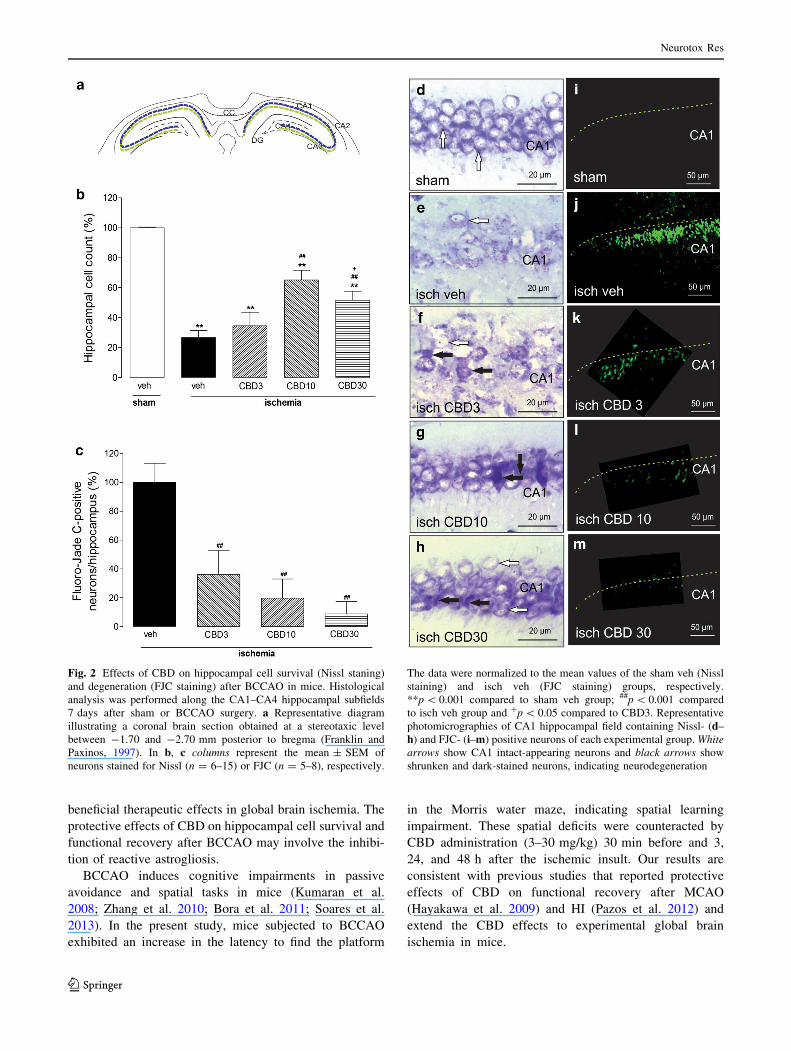
Representative photomicrographs of Nissl staining for the
hippocampus are shown in Fig. 2. BCCAO resulted in
severe neurodegeneration in the hippocampus as seen by
the presence of shrunken and dark-stained neurons (Fig. 2).
Compared with sham surgery, BCCAO caused significant
cell loss over the entire CA1-CA4 hippocampal fields
(v2 = 6,337.54, p \ 0.0001). Compared with vehicle,
CBD treatment at 10 and 30 mg/kg significantly increased
the number of intact hippocampal cells compared to veh
isch group (p \ 0.0001). There was also a significant dif-
ference between ischemic animals treated with CBD 10
and 30 mg/kg (p \ 0.05), indicating that the dose of
10 mg/kg was more effective in attenuating the neurode-
generative effect of brain ischemia.
Fluorojade-C Staining
FJC mainly stained cell bodies of degenerating neurons and
occasionally their processes (Schmued et al. 2005). The
majority of FJC-positive neurons were detected in the CA1
pyramidal layer of the hippocampus (Fig. 2c, e), which
paralleled the reduction of Nissl-stained neurons (Fig. 2b,
d). No FJC-positive cells were observed in sham-operated
animals. The statistical analysis revealed a significant
decrease in the number of FJC-positive cells in the hip-
pocampus in ischemic animals treated with CBD (3, 10 and
30 mg/kg) compared with vehicle-treated ischemic animals
(v2 = 1,656.90, p \ 0.0001).
Immunohistochemistry
GFAP-immunoreactive cells are shown in Fig. 3. BCCAO
significantly increased the IOD in ischemic animals treated
with vehicle or CBD 3 mg/kg compared with the sham
group (v2 = 17.63, p = 0.001). Compared with vehicle,
CBD 30 mg/kg significantly decreased the IOD of GFAP-
positive cells in vehicle (p = 0.0003) and CBD 3 mg/kg
(p = 0.0013) treated animals.
Discussion
We found that CBD reduced behavioral impairment and
hippocampal neurodegeneration in response to brain
ischemia in mice. Treatment with CBD (3-30 mg/kg)
30 min before and 3, 24, and 48 h after BCCAO improved
spatial learning performance in mice tested in the Morris
water maze 7 days after the ischemic insult. The Nissl
staining results showed an increase in the hippocampal cell
survival in ischemic animals after CBD treatment, sug-
gesting a protective effect of CBD on neuronal death
induced by ischemia. On the other hand, the number of
cells undergoing degeneration (FJC-positive) was reduced
by CBD treatment. GFAP immunoreactivity was also
decreased in ischemic mice treated with CBD (10 and
30 mg/kg). These findings indicate that CBD might exert
Fig. 1 Effects of CBD on learning deficits induced by BCCAO in
mice. The animals received vehicle or CBD (3, 10 or 30 mg/kg)
30 min before and 3, 24 and 48 h after surgery. Learning performance
was assessed 7 days after surgery and was expressed by the latency to
find the platform along 5 blocks of 2 trials each, i.e., ‘‘latency in
blocks’’ (a) and ‘‘total latency’’ (b). Memory retention was given by
the ‘‘time spent in the correct quadrant’’ during the probe trial (c).
Values are mean ± SEM of the groups: sham mice treated with
vehicle (sham veh, n = 15), ischemic mice treated with vehicle (isch
veh, n = 23), ischemic mice treated with CBD 3 (isch CBD3,
n = 11), 10 (isch CBD10, n = 14), or 30 (isch CBD 30, n = 11) mg/
kg. *p \ 0.05, **p \ 0.001 in comparison with isch veh group
Neurotox Res
123
beneficial therapeutic effects in global brain ischemia. The
protective effects of CBD on hippocampal cell survival and
functional recovery after BCCAO may involve the inhibi-
tion of reactive astrogliosis.
BCCAO induces cognitive impairments in passive
avoidance and spatial tasks in mice (Kumaran et al.
2008; Zhang et al. 2010; Bora et al. 2011; Soares et al.
2013). In the present study, mice subjected to BCCAO
exhibited an increase in the latency to find the platform
in the Morris water maze, indicating spatial learning
impairment. These spatial deficits were counteracted by
CBD administration (3–30 mg/kg) 30 min before and 3,
24, and 48 h after the ischemic insult. Our results are
consistent with previous studies that reported protective
effects of CBD on functional recovery after MCAO
(Hayakawa et al. 2009) and HI (Pazos et al. 2012) and
extend the CBD effects to experimental global brain
ischemia in mice.
Fig. 2 Effects of CBD on hippocampal cell survival (Nissl staning)
and degeneration (FJC staining) after BCCAO in mice. Histological
analysis was performed along the CA1–CA4 hippocampal subfields
7 days after sham or BCCAO surgery. a Representative diagram
illustrating a coronal brain section obtained at a stereotaxic level
between -1.70 and -2.70 mm posterior to bregma (Franklin and
Paxinos, 1997). In b, c columns represent the mean ± SEM of
neurons stained for Nissl (n = 6–15) or FJC (n = 5–8), respectively.
The data were normalized to the mean values of the sham veh (Nissl
staining) and isch veh (FJC staining) groups, respectively.
**p \ 0.001 compared to sham veh group; ##p \ 0.001 compared
to isch veh group and ?p \ 0.05 compared to CBD3. Representative
photomicrographies of CA1 hippocampal field containing Nissl- (d–
h) and FJC- (i–m) positive neurons of each experimental group. White
arrows show CA1 intact-appearing neurons and black arrows show
shrunken and dark-stained neurons, indicating neurodegeneration
Neurotox Res
123

CBD 10 and 30 mg/kg significantly increased the
number of intact-appearing, Nissl-stained cells and
decreased the number of FJC-positive cells in the hippo-
campus of ischemic animals. These results indicate that
CBD not only promoted functional recovery but also
decreased the delayed hippocampal cell loss and mitigated
the undergoing neurodegenerative processes induced by
global brain ischemia in mice. Despite the hippocampal
damage observed, the animals treated with CBD 3 mg/kg
were able to learn the task and perform at level similar to
controls. This finding suggests that the effect of CBD in
cognition was distinct from the structural damage in the
hippocampus. Therefore, one issue is how CBD 3 mg/kg
could have facilitated learning despite the hippocampal
lesion. Notably, dysfunction of complex behaviors and
recovery of function may reflect alterations at the subcel-
lular, synaptic, or electrophysiologic levels, or even
widespread morphological changes that would not be
restricted to a given brain structure, such as the hippo-
campus (de la Tremblaye and Plamondon 2011). In fact,
existing studies rarely assess neuronal damage outside the
hippocampus in relationship to behavioral impairments
following global ischemia. Only few authors have dem-
onstrated ischemia-induced alterations of cellular activity
in brain regions outside hippocampus, which could have a
significant impact on behavioral recovery following
ischemia. For example, Caruana et al. (2008), reported
epileptiform EEG activity in the CA1/subcular region,
perirhinal cortex and prefrontal cortex following 15 min of
global ischemia in rats. Moreover, significant losses of
CRH-positive neurons have been demonstrated in amyg-
dala 6 weeks following hypoxia–ischemia (Carty et al.
2010) a phenomenon associated with hyperactivity in
response to novel open-field exposure in ischemic rats. In
humans, reduced amygdalar volume has been correlated in
stroke and cardiac arrest survivors with cognitive
Fig. 3 Effects of CBD on
GFAP expression after BCCAO
in mice. a Representative
diagram illustrating a coronal
brain section containing the
hippocampus with a selected
area where the IOD of GFAP
immunostaining was obtained.
In b columns represent the
mean ± SEM of IOD of GFAP-
immunoreactive cells in the
different experimental groups
(n = 5–8). The data were
normalized to the mean values
of the isch veh group.
**p \ 0.001 and *p \ 0.05
compared to sham veh group;##p \ 0.001 compared to isch
veh group; ?p \ 0.05 compared
to CBD3 group. c–
g Representative
photomicrographies of GFAP
immunostaining showing
morphological changes in
GFAP expression of ischemic
compared to sham animals.
Details in a, c–g showing the
selected area where GFAP
immunolabeling was evaluated.
Py pyramidal cell lay, Lmol
lacunosum moleculare layer,
DG dentate gyrus
Neurotox Res
123

impairments (Sachdev et al. 2007). These findings support
the notion that factors other than hippocampal lesion might
influence behavioral output after brain ischemia. However,
whether or not the CBD treatment would provide transient
or enduring protective effects on neurohistological out-
comes of BCCAO in mice, is still unclear. The degree of
neuroprotection afforded by other drugs, such as the a-
amino-3-hydroxy-5-methyl-4-isoxazolepropionic acid
receptor antagonist NBQX, N-channel calcium blocker
SNX, and noncompetitive N-methyl-D-aspartate antagonist
MK801, has been shown to decline when the postischemic
time of the analysis is extended from the first 7–30 days
overall (Corbett and Crooks 1997; Corbett and Nurse
1998).
The protective effect of CBD was not limited to neurons
but extended to astrocytes. The administration of 30 mg/kg
CBD decreased reactive astrogliosis induced by BCCAO,
suggesting that astrocytes might be involved in the
observed neuroprotective effect of CBD. Astrocytes are the
most numerous cell type in the brain and dynamically
involved in synaptic transmission, metabolic and ionic
homeostasis, the anti-inflammatory response, and antioxi-
dant defense (Robel et al. 2011). An increase in astrocyte
expression, characterized by increased cell proliferation,
cell hypertrophy, and upregulation of the intermediate fil-
aments GFAP and vimentin (Robel et al. 2011), has been
demonstrated in nearly every type of central nervous sys-
tem injury, including focal (Cheung et al. 1999; Mao et al.
2011) and transient global (Kindy et al. 1992; Norenberg
1996; Ridet et al. 1997) brain ischemia. Our data are
consistent with other reports. However, astrocytes can exert
both beneficial and detrimental effects during repair pro-
cesses in the injured brain (Stoll et al. 1998; Robel et al.
2011). Astrocytes secrete neurotrophins and may provide a
permissive substratum to support axonal regrowth during
injury, but sustained reactive astrogliosis may be a major
impediment to axonal regeneration and protection (Lee
et al. 1996; Ridet et al. 1997). For example, when GFAP
filaments are genetically ablated in mice, they develop less
dense scars and show improved posttraumatic regeneration
after hemisection of the spinal cord and improved survival
and differentiation of transplanted stem cells (Pekny and
Pkna 2004; Pekny and Nilsson 2005). Moreover, increased
GFAP immunoreactivity in the CA1 hippocampal region
after brain ischemia has been associated with the extent and
maturation of neuronal necrosis (Petito and Halaby 1993;
Stoll et al. 1998). Therefore, decreased astrogliosis in
CBD-treated mice might have influenced neuronal death
induced by the ischemic insult. However, the inhibition of
astrogliosis by CBD may be a consequence, not the cause,
of the reduced neuronal damage provoked by CBD.
Additional experimentation addressing the signaling pro-
tective mechanism of CBD need be conducted.
A limitation of the present study is that the behavior
improvements observed in the Morris test were not dose-
dependent and did not correlate directly with the histo-
logical findings. The reason for this discrepancy is
unknown. However, CBD usually produces bell-shaped
dose–response curves, an effect not completely understood
but could involve the different pharmacological effects
triggered by this drug (for review, see Campos et al. 2012).
Although CBD has been reported to increase hippocampal
cell proliferation, this effect disappears at high concentra-
tions of the drug (Campos et al. 2013). CBD-mediated
neuroprotection after experimental brain ischemia has
generally been associated with the modulation of neuro-
toxicity, inflammation, and oxidative stress (Castillo et al.
2010; Pazos et al. 2012). Pharmacological studies have
shown that serotonin-1A, cannabinoid CB2, and adenosine
receptors are also involved in the neuroprotective effect of
CBD in the immature ischemic brain (Castillo et al. 2010;
Lafuente et al. 2011; Pazos et al. 2012). Recently, it was
proposed that CBD exerts its effects on brain injury by
inhibiting the astroglial differentiation of neural stem pre-
cursor cells (Shinjyo and Di Marzo 2013), an effect that
could be detrimental to neurogenesis (Robel et al. 2011;
Colangelo et al. 2012). CBD markedly downregulated
reactive astrogliosis by activating peroxisome proliferator-
activated receptors following b-amyloid neurotoxicity,
which in turn resulted in neuroprotection (Esposito et al.
2011). Therefore, the protective effects of CBD on hip-
pocampal cell death and functional recovery after BCCAO
may involve the inhibition of reactive astrogliosis.
Remaining to be investigated is whether similar mecha-
nisms are involved in CBD-induced neuroprotection after
global brain ischemia.
Acknowledgments The authors thank Marco Alberto Trombelli for
his technical support. This study was supported by Conselho Nacional
de Desenvolvimento Cientıfico e Tecnologico (CNPq), FAPESP,
State University of Maringa and Fundacao Araucaria.
Conflict of interest The authors have no conflict of interest.
References
Anderson CA, Arciniegas DB (2010) Cognitive sequelae of hypoxic-
ischemic brain injury: a review. Neurorehabilitation 26(1):47–63
Auriel E, Bornstein NM (2010) Neuroprotection in acute ischemic
stroke-current status. J Cell Mol Med 4(9):2200–2202
Bachevalier J, Meunier M (1996) Cerebral ischemia: are the memory
deficits associated with hippocampal cell loss? Hippocampus
6(5):553–560
Blum S, Luchsinger JA, Manly JJ, Schupf N, Stern Y, Brown TR,
DeCarli C, Small SA, Mayeux R, Brickman AM (2012) Memory
after silent stroke: hippocampus and infarcts both matter.
Neurology 78(1):38–46
Bora KS, Arora S, Shri R (2011) Role of Ocimum basilicum L. in
prevention of ischemia and reperfusion-induced cerebral
Neurotox Res
123

damage, and motor dysfunctions in mice brain. J Ethnopharmacol
137(3):1360–1365
Braida D, Pegorini S, Arcidiacono MV, Consalez GG, Croci L, Sala
M (2003) Post-ischemic treatment with cannabidiol prevents
electroencephalographic flattening, hyperlocomotion and neuro-
nal injury in gerbils. Neurosci Lett 346(1–2):61–64
Campos AC, Moreira FA, Gomes FV, Del Bel EA, Guimaraes FS
(2012) Multiple mechanisms involved in the large-spectrum
therapeutic potential of cannabidiol in psychiatric disorders.
Philos Trans R Soc Lond B 367(1607):3364–3378
Campos AC, Ortega Z, Palazuelos J, Fogaca MV, Aguiar DC, Dıaz-
Alonso J, Ortega-Gutierrez S, Vazquez-Villa H, Moreira FA,
Guzman M, Galve-Roperh I, Guimaraes FS (2013) The anxio-
lytic effect of cannabidiol on chronically stressed mice depends
on hippocampal neurogenesis: involvement of the endocannab-
inoid system. Int J Neuropsychopharmacol 16(6):1407–1419.
doi:10.1017/S1461145712001502
Carty ML, Wixey JA, Kesby J, Reinebrant HE, Colditz PB, Gobe G
(2010) Long-term losses of amygdala corticotropin-releasing
factor neurons are associated with behavioural outcomes follow-
ing neonatal hypoxia–ischemia. Behav Brain Res 208:609–618
Caruana DA, Nesbitt C, Mumby DG, Chapman CA (2008) Seizure
activity in the rat hippocampus, perirhinal and prefrontal cortex
associated with transient global cerebral ischemia. J Neural
Transm 115(3):401–411
Castillo A, Tolon MR, Fernandez-Ruiz J, Romero J, Martinez-Orgado
J (2010) The neuroprotective effect of cannabidiol in an in vitro
model of newborn hypoxic-ischemic brain damage in mice is
mediated by CB(2) and adenosine receptors. Neurobiol Dis
37(2):434–440
Cheung WM, Wang CK, Kuo JS, Lin TN (1999) Changes in the level
of glial fibrillary acidic protein (GFAP) after mild and severe
focal cerebral ischemia. Chin J Physiol 42(4):227–235
Colangelo AM, Cirillo G, Lavitrano ML, Alberghina L, Papa M
(2012) Targeting reactive astrogliosis by novel biotechnological
strategies. Biotechnol Adv 30(1):261–271
Corbett D, Crooks P (1997) Ischemic preconditioning: a long term
survival study using behavioural and histological endpoints.
Brain Res. 760(1–2):129–136
Corbett D, Nurse S (1998) The problem of assessing effective
neuroprotection in experimental cerebral ischemia. Prog Neuro-
biol. 54(5):531–548
de la Tremblaye PB, Plamondon H (2011) Impaired conditioned
emotional response and object recognition are concomitant to
neuronal damage in the amygdala and perirhinal cortex in
middle-aged ischemic rats. Behav Brain Res 219(2):227–233
Duan YL, Wang SY, Zeng QW, Su DS, Li W, Wang XR, Zhao Z
(2011) Astroglial reaction to delta opioid peptide [D-Ala2,
D-Leu5] enkephalin confers neuroprotection against global
ischemia in the adult rat hippocampus. Neuroscience 192:81–90
Esposito G, Scuderi C, Valenza M, Togna GI, Latina V, De Filippis
D, Cipriano M, Carratu MR, Iuvone T, Steardo L (2011)
Cannabidiol reduces Ab-induced neuroinflammation and pro-
motes hippocampal neurogenesis through PPARc involvement.
PLoS One 6(12):e28668
Fernandez-Ruiz J, Sagredo O, Pazos MR, Garcıa C, Pertwee R,
Mechoulam R, Martınez-Orgado J (2013) Cannabidiol for
neurodegenerative disorders: important new clinical applications
for this phytocannabinoid? Br J Clin Pharmacol 75(2):323–333
Franklin KBJ, Paxinos G (1997) The mouse brain in stereotaxic
coordinates, 2nd edn. Editora Academic Press, San Diego
Garcia C, Palomo-Garo C, Garcia-Arencibia M, Ramos J, Pertwee R,
Fernandez-Ruiz J (2011) Symptom-relieving and neuroprotective
effects of the phytocannabinoid D9-THCV in animal modeks of
Parkinson’s disease. Br J Pharmacol 164(7):1495–1506
Harvey BS, Ohlsson KS, Maag JL, Musgrave IF, Smid SD (2012)
Contrasting protective effects of cannabinoids against oxidative
stress and amyloid-b evoked neurotoxicity in vitro. Neurotox-
icology 33(1):138–146
Hayakawa K, Mishima K, Irie K, Hazekawa M, Mishima S, Fujioka
M, Orito K, Egashira N, Katsurabayashi S, Takasaki K, Iwasaki
K, Fujiwara M (2008) Cannabidiol prevents a post-ischemic
injury progressively induced by cerebral ischemia via a high-
mobility group box1-inhibiting mechanism. Neuropharmacology
55(8):1280–1286
Hayakawa K, Irie K, Sano K, Watanabe T, Higuchi S, Enoki M,
Nakano T, Harada K, Ishikane S, Ikeda T, Fujioka M, Orito K,
Iwasaki K, Mishima K, Fujiwara M (2009) Therapeutic time
window of cannabidiol treatment on delayed ischemic damage
via high-mobility group box1-inhibiting mechanism. Biol Pharm
Bull 32(9):1538–1544
Iuvone T, Esposito G, Esposito R, Santamaria R, Di Rosa M, Izzo AA
(2004) Neuroprotective effect of cannabidiol, a non-psychoac-
tive component from Cannabis sativa, on beta-amyloid-induced
toxicity in PC12 cells. J Neurochem 89(1):134–141
Kartsounis LD, Rudge P, Stevens JM (1995) Bilateral lesions of CA1
and CA2 fields of the hippocampus are sufficient to cause a
severe amnesic syndrome in humans. J Neurol Neurosurg
Psychiatry 59(1):95–98
Kindy MS, Bhat AN, Bhat NR (1992) Transient ischemia stimulates
glial fibrillary acid protein and vimentin gene expression in the
gerbil neocortex, striatum and hippocampus. Brain Res Mol
Brain Res 13(3):199–206
Kumaran D, Udayabanu M, Kumar M, Aneja R, Katyal A (2008)
Involvement of angiotensin converting enzyme in cerebral
hypoperfusion induced anterograde memory impairment and
cholinergic dysfunction in rats. Neuroscience 155(3):626–639
Lafuente H, Alvarez FJ, Pazos MR, Alvarez A, Rey-Santano MC,
Mielgo V, Murgia-Esteve X, Hilario E, Martinez-Orgado J
(2011) Cannabidiol reduces brain damage and improves func-
tional recovery after acute hypoxia–ischemia in newborn pigs.
Pediatr Res 70(3):272–277
Lee TH, Kato H, Kogure K, Itoyama Y (1996) Temporal profile of
nerve growth factor like immunoreactivity after transient focal
cerebral ischemia in rats (1996). Brain Res 713:199–210
Mao X, Yin W, Liu M, Ye M, Liu P, Liu J, Xu S, Pi R (2011) Osthole,
a natural coumarin, improves neurobehavioral functions and
reduces infarct volume and matrix metalloproteinase-9 activity
after transient focal cerebral ischemia in rats. Brain Res
1385:275–280
Mateen FJ, Josephs KA, Trenerry MR, Felmlee-Devine MD, Weaver
AL, Carone M, White RD (2011) Long-term cognitive outcomes
following out-of-hospital cardiac arrest: a population-based
study. Neurology 77(15):1438–1445
Mechoulam R, Shohami E (2007) Endocannabinoids and traumatic
brain injury. Mol Neurobiol 36(1):68–74
Moulaert VR, Wachelder EM, Verbunt JA, Wade DT, van Heugten
CM (2010) Determinants of quality of life in survivors of cardiac
arrest. J Rehabil Med 42(6):553–558
Neumann JT, Cohan CH, Dave KR, Wright CB, Perez-Pinzon MA
(2013) Global cerebral ischemia: synaptic and cognitive dys-
function. Curr Drug Targets 14(1):20–35
Nikonenko AG, Radenovic L, Andjus PR, Skibo GG (2009)
Structural features of ischemic damage in the hippocampus.
Anat Rec (Hoboken) 292(12):1914–1921
Norenberg MD (1996) Astrocytic-ammonia interactions in hepatic
encephalopathy. Semin Liver Dis 16(3):245–253
Pazos MR, Cinquina V, Gomez A, Layunta R, Santos M, Fernandez-
Ruiz J, Martınez-Orgado J (2012) Cannabidiol administration
after hypoxia–ischemia to newborn rats reduces long-term brain
Neurotox Res
123

injury and restores neurobehavioral function. Neuropharmacol-
ogy 63(5):776–783
Pazos MR, Mohammed N, Lafuente H, Santos M, Martınez-Pinilla E,
Moreno E, Valdizan E, Romero J, Pazos A, Franco R, Hillard CJ,
Alvarez FJ, Martınez-Orgado J (2013) Mechanisms of cannabi-
diol neuroprotection in hypoxic newborn pigs: role of 5HT1A
and CB2 receptors. Neuropharmacology 71:282–291
Pekny M, Pekna M (2004) Astrocyte intermediate filaments in CNS
pathologies and regeneration. J Pathol. 204(4):428–437
Pekny M, Nilsson M (2005) Astrocyte activation and reactive gliosis.
Glia 50(4):427–434
Peskine A, Picq C, Pradat-diehl P (2010) Neurological sequelae after
cerebral anoxia. Brain Inj 24(5):755–761
Petito CK, Halaby IA (1993) Relationship between ischemia and
ischemic neuronal necrosis to astrocyte expression of glial
fibrillary acidic protein. Int J Dev Neurosci 11(2):239–247
Prediger RD, Fernandes MS, Rial D, Wopereis S, Pereira VS, Bosse
TS, Da Silva CB, Carradore RS, Machado MS, Cechinel-Filho
V, Costa-Campos L (2008) Effects of acute administration of the
hydroalcoholic extract of mate tea leaves (Ilex paraguariensis)
in animal models of learning and memory. J Ethnopharmacol
120(3):465–473
Ridet JL, Malhotra SK, Privat A, Gage FH (1997) Reactive
astrocytes: cellular and molecular cues to biological function.
Trends Neurosci 20(12):570–577
Robel S, Berninger B, Gotz M (2011) The stem cell potential of glia:
lessons from reactive gliosis. Nat Rev Neurosci 12(2):88–104
Sachdev PS, Chen X, Joscelyne A, Wen W, Brodaty H (2007)
Amygdala in stroke/transient ischemic attack patients and its
relationship to cognitive impairment and psychopathology: the
Sydney stroke study. Am J Geriatr Psychiatry 15:487–496
Sagredo O, Pazos MR, Satta V, Ramos JA, Pertwee RG, Fernandez-
Ruiz J (2011) Neuroprotective effects of phytocannabinoid-
based medicines in experimental models of Huntington’s
disease. J Neurosci Res 89(9):1509–1518
Schmued LC, Stowers CC, Scallet AC, Xu L (2005) Fluoro-Jade C
results in ultra high resolution and contrast labeling of degen-
erating neurons. Brain Res 1035(1):24–31
Seif el Nasr M, Nuglisch J, Krieglstein J (1992) Prevention of ischemia-
induced cerebral hypothermia by controlling the environmental
temperature. J Pharmacol Toxicol Methods 27(1):23–26
Shinjyo N, Di Marzo V (2013) The effect cannabichromene on adult
neural stem/progenitor cells. Neurochem 63(5):432–437
Soares LM, Schiavon AP, Milani H, de Oliveira RM (2013) Cognitive
impairment and persistent anxiety-related responses following
bilateral common carotid artery occlusion in mice. Behav Brain
Res 249:28–37
Stoll G, Jander S, Schroeter M (1998) Inflammation and glial
responses in ischemic brain lesions. Prog Neurobiol
56(2):149–171
Szydlowska K, Gozdz A, Dabrowski M, Zawadzka M, Kaminska B
(2010) Prolonged activation of ERK triggers glutamate-induced
apoptosis of astrocytes: neuroprotective effect of FK506. J Neu-
rochem 113(4):904–918
Takuma K, Baba A, Matsuda T (2004) Astrocyte apoptosis:
implications for neuroprotection. Prog Neurobiol 72(2):111–127
Valdeolivas S, Satta V, Pertwee RG, Fernandez-Ruiz J, Sagredo O
(2012) Sativex-like combination of phytocannabinoids is neuro-
protective in malonate-lesioned rats, an inflammatory model of
Huntington’s disease: role of CB1 and CB2 receptors. ACS Chem
Neurosci 3(5):400–406
Zhang L, Fu F, Zhang X, Zhu M, Wang T, Fan H (2010) Escin
attenuates cognitive deficits and hippocampal injury after
transient global cerebral ischemia in mice via regulating certain
inflammatory genes. Neurochem Int 57(2):119–127
Zola-Morgan S, Squire LR, Amaral DG (1986) Human amnesia and
the medial temporal region: enduring memory impairment
following a bilateral lesion limited to field CA1 of the
Hippocampus. J Neurosci 6(10):2950–2967
Neurotox Res
123
Copyright © 2022 FDOKUMEN




















