
Do Salivary Proline-Rich Proteins Counteract Dietary Hydrolyzable Tannin in Laboratory Rats?
Upload
independentCategory
view
0download
0
Comparative Biochemistry and Physiology Part B 132(2002)739–750
1096-4959/02/$ - see front matter� 2002 Elsevier Science Inc. All rights reserved.PII: S1096-4959Ž02.00091-X
Proline biosynthesis genes and their regulation under salinity stressin the euryhaline copepodTigriopus californicus
Christopher S. Willett*, Ronald S. Burton
Marine Biology Research Division, Scripps Institution of Oceanography, University of California, San Diego, La Jolla,CA 92093-0202, USA
Received 4 February 2002; received in revised form 10 May 2002; accepted 14 May 2002
Abstract
Diverse organisms regulate concentrations of intracellular organic osmolytes in response to changes in environmentalsalinity or desiccation. In marine crustaceans, accumulation of high concentrations of proline is a dominant componentof response to hyperosmotic stress. In the euryhaline copepodTigriopus californicus, synthesis of proline from itsmetabolic precursor glutamate is tightly regulated by changes in environmental salinity. Here, for the first time in amarine invertebrate, the genes responsible for this pathway have been cloned and characterized. The two proteins displaythe sequence features of homologous enzymes identified from other eukaryotes. One of the cloned genes,d -pyrroline-1
5-carboxylase reductase(P5CR), is demonstrated to have the reductase enzyme activity when expressed in proline-auxotroph bacteria, while the second,d -pyrroline-5-carboxylase synthase(P5CS), does not rescue proline-auxotroph1
bacteria. In contrast to results from higher plants, neither levels of P5CS nor P5CR mRNAs increase in response tosalinity stress inT. californicus. Hence, regulation of proline synthesis during osmotic stress inT. californicus is likelymediated by some form of post-transcriptional regulation of either P5CS or P5CR. Understanding the regulation thispathway may elucidate the mechanisms limiting the salinity ranges of marine taxa.� 2002 Elsevier Science Inc. Allrights reserved.
Keywords: Crustacean; Free amino acid; Hyperosmotic stress; Organic osmolytes; Osmotic response; Amino acid synthesis;d -1
pyrroline-5-carboxylase synthase;d -pyrroline-5-carboxylase reductase1
1. Introduction
The ability of organisms to maintain a stableintracellular environment in the face of changes inthe external environment is crucial for survivalacross a wide range of habitats. Maintaining ionicbalance and cell volume are key parts of this stableintracellular environment. In response to hyperos-motic stress, diverse organisms exhibit a commoncellular response: the accumulation of intracellularorganic osmolytes(Gilles, 1979; Yancey et al.,
*Corresponding author. Tel.:q1-858-534-7827; fax:q1-858-534-7313.
E-mail address: [email protected](C.S. Willett).
1982; Burg et al., 1996; Hare et al., 1998; Kempfand Bremer, 1998). In osmo-conforming marineinvertebrates, body fluids change dramatically inosmotic concentration with salinity changes in theexternal environment; in response to increases inhemolymph osmolarity, intracellular organic com-pounds accumulate to comprise over 50% of theintracellular osmolyte pool in many invertebrates(Yancey, 1994).
Despite wide interest in the role of intracellularorganic osmolytes during hyperosmotic stress, themolecular mechanisms leading to osmolyte accu-mulation are not understood in marine animals. Ineuryhaline crustaceans, free amino acids(FAA)comprise the bulk of the organic osmolyte pool.

740 C.S. Willett, R.S. Burton / Comparative Biochemistry and Physiology Part B 132 (2002) 739–750
Fig. 1. Pathway for synthesis of proline from glutamate. Stepsin pathway are the same in both eukaryotes and prokaryotes,but the initial two reactions are performed by the bifunctionalP5CS enzyme in higher eukaryotes.
FAA accumulation is evident with increasing salin-ities in harpacticoid copepods in the genusTigrio-pus. These copepods experience rapid changes inenvironmental salinities when the splash pools theyinhabit in the rocky, high intertidal dessicate,accumulate fresh rainwater, or wash out withseawater. A British species,Tigriopus brevicornis,is an osmoconformer over the range of salinitiesof 15–330% seawater(McAllen et al., 1998).Intracellular FAA accumulation in its congener,T.californicus, has been found primarily to involveproline, alanine and glycine(Burton and Feldman,1982). Of the three FAA, proline shows the largestincrease in concentration under hyperosmoticstress(Burton and Feldman, 1982; Goolish andBurton, 1989). When copepods are transferredfrom 50 to 100% seawater proline concentrationsincrease nearly 7-fold within a period of 3 h(Goolish and Burton, 1989). Studies of incorpo-ration of C-labeled precursors have shown that14
the synthesis of proline(but not alanine) is tightlyregulated; little or no new proline is synthesizedin a constant salinity environment, but high levelsof label were incorporated into proline in responseto hyperosmotic stress(Burton, 1986). The meta-bolic precursor for proline synthesis, glutamate, issynthesized under conditions of constant salinity,implying that the important regulatory point inproline synthesis occurs in the steps leading toproline synthesis subsequent to glutamate(Burton,1986, 1991a). Burton showed that protein synthe-sis was required for the upregulation of prolinesynthesis (Burton, 1991b). The T. californicusregulatory checkpoint for proline accumulationdoes not appear to be unique; studies on othercrustacean euryhaline osmoconformers(blue crabjuveniles,Callinectes sapidus, and penaeid shrimppost-larvae,Penaeus aztecus) that accumulate pro-line in response to hyperosmotic stress show asimilar pattern of proline synthesis with tightregulation of the steps from glutamate to proline(Burton, 1992; Bishop and Burton, 1993).
The synthesis of proline from glutamate is afour-step process catalyzed by two enzymes inhigher eukaryotes, the bifunctional enzymed -1
pyrroline-5-carboxylase synthase(P5CS), andd -pyrroline-5-carboxylase reductase(P5CR)1
(Delauney and Verma, 1990; Hu et al., 1992,1999). P5CS catalyzes the first two steps of thepathway, the third occurs spontaneously, and thefourth is catalyzed by P5CR(Fig. 1). Plantsaccumulate proline in response to increasing salin-
ities (and desiccation)and the mRNA levels forboth P5CS and P5CR increase when subjected tothese stresses(Delauney and Verma, 1990; Hu etal., 1992). P5CS rather than P5CR appears to bethe rate-limiting enzyme for proline accumulationin plants; levels and activity of P5CR protein donot increase with increasing proline productionwhile increased expression of P5CS causesincreased proline production(Kishor et al., 1995;Nanjo et al., 1999; Hua et al., 2000). Analogousto its regulation in bacteria, P5CS is feedbackinhibited in plants by proline, but the inhibitionmay lessen under salinity stress conditions(Bog-gess et al., 1976; Hong et al., 2000). Combined,these results suggest that regulation of P5CS activ-ity, largely through increased enzyme synthesis, iscrucial for the regulation of stress-induced prolinesynthesis in plants.
Given the importance of the enzymes P5CS andP5CR for the accumulation of proline inT. cali-fornicus and other marine arthropods in responseto salinity stress, we sought to characterize theseenzymes and their regulation. Our previousattempts to directly measure enzyme activities forP5CS have been unsuccessful; here we employ

741C.S. Willett, R.S. Burton / Comparative Biochemistry and Physiology Part B 132 (2002) 739–750
molecular cloning to characterize for the first timein a marine invertebrate the genes encoding P5CSand P5CR. Based upon sequence identity, thecopepod genes encode proteins that are clearlyhomologous to their counterparts in other eukary-otes. T. californicus P5CR protein but not P5CSprotein is able to complement bacteria which aremutants for the homologous enzyme. Regulationof this pathway in copepods is different than thatfor plants because our analysis of gene expressionin T. californicus exposed to salinity stress failedto detect upregulation of either P5CS or P5CRmRNA. We suggest that proline synthesis inT.californicus may be under either translational orpost-translational control.
2. Materials and methods
2.1. P5CS and P5CR gene characterization
We used a 39 rapid amplification of cDNA endsprocedure(39 RACE) to PCR amplify a fragmentof P5CS fromT. californicus. T. californicus werecollected from tidepools in San Diego, CA andacclimated to 50% seawater for a period of 3 days.Animals were then subjected to hyperosmoticstress by transferring them from 50 to 100%seawater for a period of 90 min. RNA was isolatedfrom hyperosmotically stressed copepods usingTRI reagent following suppliers protocols(SigmaChemical, St. Louis, MO). Complementary DNA(cDNA) was synthesized from total RNA usingthe Superscript II reverse transcriptase enzyme(Gibco BRL, Gaithersburg, MD) and supplier’sprotocols. For the 39 RACE, we designed degen-erate primers to match conserved regions of analignment of P5CS fromDrosophila melanogaster,Homo sapiens, Arabidopsis thaliana, andCaenor-habditis elegans. A T7ypoly-T primer was usedfor cDNA first strand synthesis(59-TAATAC-GACTCACTATAGGGTTTTTTTTTTTTTTTTTT-V-39) so that a single degenerate forward primer(P5CS-F2.5, 59-GANTAYCCNGCNGCNTGYAA-YGC-39) could be used with a T7 primer toamplify a product of 850 bp in the subsequentPCR reaction. This fragment of P5CS was gelpurified (using QIAQUICK kit, QIAGEN, Chat-sworth, CA), and directly sequenced. A 59RACEprocedure, Generacer(Invitrogen, Carlsbad, CA)was used to obtain the remainder of the cDNA forP5CS. Except for the large first intron(onlysequenced from San Diego animals), the genomic
sequence of P5C was obtained from three Califor-nia T. californicus populationswSan Diego(SD),Abalone Cove(AB, Los Angeles County), andSanta Cruz(SC)x by PCR amplification and directsequencing.
We used a similar strategy to identify P5CRfrom T. californicus. We designed degenerateprimers from an alignment of P5CR of distantlyrelated taxa. For P5CR the Generacer kit wasused for both 39 RACE and 59 RACE. In theinitial 39 RACE the degenerate primer P5CR.F2 (59-GTNTGYTCNCCNGGNGGNGCNACNA-T-39) was used in conjunction with a kit 39RACEprimer to amplify a 600 bp region of P5CR.Subsequent 59 RACE with species–specific prim-ers yielded the remainder of the cDNA. Againsequences were obtained for the genomic P5CRfrom the SD, AB and SCT. californicus popula-tions using PCR amplification with species–spe-cific primers followed by direct sequencing.Linkage of P5CR and P5CS was examined byusing population diagnostic PCR to look forrecombinant multi-locus genotypes in backcrossindividuals as described in Willett and Burton(2001). Primer sequences and PCR profiles forP5CS and P5CR population-diagnostic PCR areavailable from authors upon request.
Editing of sequences was performed with theprogram Sequencer v4.1(Genecodes, Ann ArborMI). Alignments were made using Clustal X. Theprogram PAUP* version 4.06b(Swofford, 2001)was used to generate tables of ammo acid similar-ity and to construct phylogenetic trees. Trees weremade with the parsimony criterion using a branch-and-bound search algorithm.
2.2. P5CS and P5CR protein expression
We expressedT. californicus P5CS and P5CRproteins inEscherichia coli strains that were dele-tion mutants for these enzyme functions.E. colistrains used were the following: G9(proA- ),y
G13 (proB ), and CSH26(proAB ) (strains pro-y y
vided by L.N. Csonka). The genesproB andproAencodeg-glutamyl kinase andg-glutamyl phos-phate reductase enzymes, respectively,wboth func-tions of P5CS in higher eukaryotes(Fig. 1)x.x strain is aproC mutant (obtained fromE.342 y
coli stock center). The geneproC encodes theE.coli P5CR enzyme homolog(Fig. 1). All strainswere capable of growth on minimal media M63only in presence of supplemental proline(also

742 C.S. Willett, R.S. Burton / Comparative Biochemistry and Physiology Part B 132 (2002) 739–750
supplemented with 200mM threonine, 200mMleucine, 50mM thiamine-HCl, 40mgyml methio-nine, 2 mM MgSO , and 0.2% glycerol or glu-4
cose). Chemically competent cells were made fromall strains and electro-competent cells from the G9and CSH26 strains.
PCR products corresponding to the codingregions for both P5CS and P5CR were insertedinto pBAD-TOPO expression vectors(pBAD-TOPO) TA cloning kit, Invitrogen, Carlsbad, CA).For P5CS, four constructs were generated corre-sponding to combinations of two potential startcodons and inclusion or exclusion of vector C-terminal protein sequence(encoding V5 epitopetag). Vectors were transformed into mutant strainsusing either heat shock for chemically competentor electroporation for electro-competent cells.Transformed cells were plated on both selectivemedia(minimal media with ampicillin, no proline,and the plasmid inducerL-arabinose) or on LByampicillin plates. Transformant colonies from LByampicillin were tested for enzyme activity bystreaking onto selective media and incubating at37 8C for 2 days or longer.
The identity and direction of inserts werechecked by PCR assay of transformant coloniesusing kit pBAD.fwd primer and a gene-specificreverse primer. Protein expression was verified byWestern blotting using the antibody, anti-V5-HRP(Invitrogen, Carlsbad, CA) that recognizes the V5epitope tag added to both the P5CR and P5CS bythe pBAD-TOPO vector. Lysates of bacteria cul-tures grown with either 2% glucose or 2% arabi-nose for 4 h were run on 10% SDS-PAGE gels(Garfin, 1990). Blotting with anti-V5-HRP wasperformed using manufacturer’s protocols andbands were detected by chemiluminesence usingthe reagent Supersignal West Dura Extended(Pierce Chemical Co., Rockford, IL).
2.3. Quantification of mRNA expression
The quantification of mRNA levels before andafter hyperosmotic stress were performed usingreal-time reverse transcriptase PCR(RT-PCR) inthe Lightcycler (Roche, Indianapolis, IN). Twoseparate experiments with different RNA isolationprotocols were conducted to quantify P5CS mRNAlevels; only the second experimental RNA set wasthen used to quantify P5CR mRNA levels. Thefirst RNA isolation protocol used San Diego cope-pods that had moderate levels of lipid reserves
wlipid levels have been shown to be an importantfactor in the amounts of proline synthesized(Gool-ish and Burton, 1989)x. Approximately 1000 cope-pods were placed in each of the five 500 mlbeakers with filtered 50% seawater and allowed toacclimate for a period of three days(a smallamount of fish food was added to beakers). Theanimals from four beakers were transferred tofiltered 100% seawater. RNA was isolated fromcultures using the TRI reagent(Sigma Chemical,St. Louis, MO) immediately for the unstressed andat timepoints of 15, 45, 90 and 240 min aftertransfer to 100% seawater. DNA contaminationwas removed with a DNAse I treatment using theDNA-free kit (Ambion, Austin, TX). Total RNAwas measured by a flourometric assay using Ribo-green RNA quantification kit(Molecular probes,Eugene, OR) in a plate-reader spectroflourometer.
The Lightcycler assay quantifies the initialamount of template by measuring at what pointthe exponential phase(crossing point) of the PCRreaction begins in each reaction. The relativeamount of each template is determined by com-parison of crossing-points with a standard curve.We used the RNA amplification kit, SYBR GreenI (Roche) following manufacturer’s protocols. Toassay P5CS we amplified a fragment of 600 bp ofmRNA with two gene-specific primers. In the firstP5CS assay we made a series of dilutions of thecontrol (no stress) RNA to make a standard curve(0.2, 0.07, 0.02, 0.007ml of RNA) for comparisonwith each of the hyperosmotic stress treatmentRNA’s. Three replicates of 2ml of 1:100 dilutedRNA were run for each of the stress treatmentswhile two replicates were run for each of the fourcontrolled dilutions.
A second set of P5CS assays were performedon RNA isolations from two replicate transfersfrom 50 to 100% seawater of copepods for thetime periods of 0, 5, 15, 30 and 90 min(only asingle replicate for 90 min). Ten male and tenfemale copepods were used per RNA isolation inthis experiment(again, only animals with at leastmoderate lipid reserves were included). RNA wasisolated with the kit RNAqueous-4PCR(Ambion,Austin, TX) followed by DNAse I treatment usingDNA-free (Ambion, Austin, TX). The transcriptlevels were measured in the Lightcycler asdescribed previously with the following differenc-es: The DNA amplification kit, SYBR Green I(Roche) was used to create a standard curve fromfive serial dilutions of 10 to 10 of a fragmenty3 y7

743C.S. Willett, R.S. Burton / Comparative Biochemistry and Physiology Part B 132 (2002) 739–750
of cloned P5CS PCR product. Two replicates ofeach mRNA quantification were performed using2 ml of 1:10 diluted total RNA (approx. 0.06copepodsyreaction).
The P5CR mRNA levels were determined forthe second set of RNA isolations described above.Once again the RNA amplification kit, SYBRGreen I, was used with the Lightcycler to quantifymessage levels. Two P5CR-specific primers gen-erating a product of 650 bp were used in thequantitative PCR reactions. Quantification wasdone against a standard of serial dilutions of apurified P5CR PCR product. Again two replicatemeasurements were performed for each RNA prep,now using 5ml of undiluted mRNA for the PCRreaction(approx. 1.5 copepodsyreaction).
3. Results
3.1. Identification of P5CS and P5CR genes
Degenerate primers were successfully employedfor 39 RACE to isolate partial sequences of P5CSand P5CR from T. californicus. These partialsequences allowed us to designT. californicus-specific primers that were used to amplify theremainder of the cDNAs using 59 RACE. ForP5CS a cDNA of 3028 bp was sequenced contain-ing an open reading frame encoding 857 aminoacids. GenBank searches with this protein recoveronly P5CS genes from other organisms(or theirbacterial constituent homologs). Fig. 2 presents analignment with human P5CSwthis homolog hasbeen demonstrated to have P5CS enzyme functions(Hu et al., 1999)x. The N-termini of these twoproteins display no significant similarity, possiblyreflecting the lower conservation of the putativemitochondrial signaling peptides. Potential cleav-age sites for this signal peptide are shown in Fig.2 (using the program mitoprot II 1.0a4). Theremainder of the protein contains large regions ofidentity or similarity. Alignments ofT. californicusP5CS with other homologs have between 61%(D.melanogaster) and 33%(E. coli proB) amino acididentity (Table 1). Like other higher eukaryotes,T. californicus P5CS appears to be bifunctional,having domains homologous toproA and proBfrom E. coli. When phylogenetic trees are con-structed using the amino acid sequences and rootedusing theE. coli sequences, the expected organis-mal pattern of relationships are recovered:T. cali-fornicus and D. melanogaster group first, then
both group with human, thenC. elegans, andfinally A. thaliana, consistent with an orthologousrelationship of these genes(tree not shown). Thesequence data clearly suggest that the cloned geneencodes a P5CS homolog inT. californicus.
For P5CR, the cDNA was 1130 bp and encodeda protein of 270 amino acids. In Fig. 3 thisT.californicus protein is aligned with aD. melano-gaster protein that has been shown to have P5CRenzyme activity(Misener and Walker, 2001)andanother uncharacterized homolog fromD. melan-ogaster. GenBank searches recover three homologsin humans as well as homologs in other species.The T. californicus P5CR ranges from 33 to 45%identical in amino acid sequence to other P5CRsand homologs(Table 2). If a phylogenetic tree isconstructed from these homologous genes androoted with E. coli proC, the only apparent rela-tionship groups the first and second human P5CRhomologs together. All other relationships areunresolved or change dramatically depending onwhich of the taxa are included(trees not shown).The sequence evidence for theT. californicusP5CR gene indicates that this protein is in thesame gene family as other P5CRs, but the exactorthology with homologs from other species can-not be determined with the current taxon sampling.
3.2. Gene structure, sequence variation, and link-age relationships of P5CR and P5CS
Previous work has demonstrated extensivegenetic divergence amongT. californicus popula-tions (e.g. Burton, 1998; Rawson et al., 2000);therefore, we sequenced portions of both P5CRand P5CS genomic DNA for two individuals fromeach of three California populations(SC, SD, AB).Genomic structure is conserved acrossT. califor-nicus populations and consists of 7 introns inP5CS and 2 in P5CR(Fig. 2 and Fig. 3); althoughmost of the introns are small(69–190 bp), thefirst intron in P5CS is 1841 bp. WithinT. califor-nicus, numerous polymorphic nucleotide positionsand insertionydeletion variation(in introns anduntranslated regions) distinguish populations forboth P5CR and P5CS(Table 3). Four amino acidsubstitutions between populations are found inP5CS but none in P5CR. Taking advantage of thepopulation-specific nucleotide variation, we scoredalleles in backcross individuals and determinedthat P5CR and P5CS are not linked to one another.

744C
.S.W
illett,R
.S.B
urton/
Com
parativeB
iochemistry
andP
hysiologyP
artB
132(2002)
739–750
Fig. 2. Amino acid alignment ofT. californicus P5CS with human P5CS. Downward arrow indicates a predicted mitochondrial signal peptide cleavage site in copepod protein.Upward arrows indicate potential cleavage sites in human protein(Hu et al., 1999). T. californicus intron positions are numbered 1 to 7 followed by nucleotide position after whichintron occurs in codon. Amino acid differences between populations are indicated by the variant amino acid and the population in which it occurs(SC-Santa Cruz, AB-AbaloneCove). Amino acid sequence shown is the San Diego sequence.

745C.S. Willett, R.S. Burton / Comparative Biochemistry and Physiology Part B 132 (2002) 739–750
Table 1Percent amino acid identities forT. californicus P5CS protein in comparison to other P5CS homologs and overlapping regions of theproA andproB genes ofE. coli
D. mel Human C. elegans A. thaliana E. coli proB E. coli proA
T. cal. 61.0 53.5 51.3 47.4 32.5 36.2D. mel. 59.3 57.1 46.2 35.3 36.4Human 56.0 47.7 36.1 35.5C. elegans 46.1 33.7 35.9A. thaliana 36.3 39.0
Identities are calculated from a ClustalX alignment of genes beginning after the putative human P5CS mitochondrial targetingsequence cleavage site in the N-terminuswcorresponding to amino acid 130 ofT. californicus sequence(Hu et al., 1999)x. Distanceswere calculated using the program PAUP* and corrected for gaps(Swofford, 2001). Accession numbers are; human-U76542,D.melanogaster-AAF51799,C. elegans-P54889,A. thaliana-X87330,proA-P07004, andproB-P07005.
3.3. Functional expression in E. coli
To further characterize the putative P5CR andP5CS genes fromT. californicus, we tested forfunction via expression inE. coli. Four prolineauxotrophic strains were used, three mutant forproA and proB and one mutant forproC. Whenthe full translated region for P5CR was expressedin proC mutant bacteria, complementation wasobserved and the bacteria were able to growwithout added proline(Fig. 4). This complemen-tation occurs in the presence of an inducer of thepBAD plasmid (L-arabinose), but does not occurin the presence of a repressor(glucose) indicatingthat rescue is from the protein expressed from theplasmid and not from reversion of theproC muta-tion. Therefore, T. californicus P5CR has theenzyme activity to rescue theproC mutation(bac-terial P5C reductase mutant) despite only modestamino acid similarity between the two proteins(Table 2).
In contrast to the results seen for P5CR, wewere not able to complement theproA andyorproB mutations using the putativeT. californicusP5CS gene. Transformants with the P5CS constructin proper orientation (verified by PCR andsequencing) were recovered from medium contain-ing proline. We have also been able to verify thatthese transformant strains express the cloned pro-tein encoding gene(transformants with eitherP5CR or P5CS plasmid produce a recombinantprotein of the proper size identified with a V5epitope tag and anti-V5 antibody in Western blots).To test if the presence of mitochondrial signalpeptide from the P5CS protein was preventingcomplementation, we constructed a plasmid thatencodes a protein lacking this N-terminalsequence; transformants expressing this protein
(beginning at Met residue 106 of the P5CS pro-tein) also fails to complement theE. coli auxo-trophs. Finally, the exclusion of theplasmid-specific amino acids added for the V5epitope tag again failed to facilitatecomplementation.
3.4. Levels of mRNA for proline synthetic enzymes
We measured the levels of P5CR and P5CSmRNA by using real-time RT-PCR assays in theLightcycler. In the first set of experiments withP5CS we examined mRNA levels at four time-points after hyperosmotic stress and comparedthese levels relative to the unstressed control(Table 4). The level of P5CS mRNA increasedslightly at the 15 and 45 min timepoints but notconsistently across replicates and measures. Dotblot experiments with P5CS probes and the RNA’sused in this experiment gave equivalent results(unpublished results). A second experiment con-ducted with additional replicate timepoints showedno significant increase in mRNA levels relative tocontrols(Table 4). Similarly, we see no evidenceof an increase in mRNA levels for P5CR afterhyperosmotic stress(Table 5).
4. Discussion
The regulation of proline synthesis inT. califor-nicus (and other crustaceans) plays a significantrole in their response to changes in environmentalsalinity. Organic osmolytes like proline have beenshown to be key effectors of cell volume regulationacross a wide range of organisms. We have isolatedgenes encoding the structural enzymes P5CR andP5CS that catalyze the key steps of the synthesisof proline from glutamate in these copepods. When

746C
.S.W
illett,R
.S.B
urton/
Com
parativeB
iochemistry
andP
hysiologyP
artB
132(2002)
739–750
Fig. 3. Amino acid alignment ofT. californicus P5CR withD. melanogaster homologs. FirstD. melanogaster protein is currently called unidentified gene product in Genbank whilesecond is P5CR with demonstrated enzyme function(Misener and Walker, 2001). Intron positions inT. californicus are numbered 1 and 2 followed by nucleotide position in codonafter which intron occurs.
747C.S. Willett, R.S. Burton / Comparative Biochemistry and Physiology Part B 132 (2002) 739–750
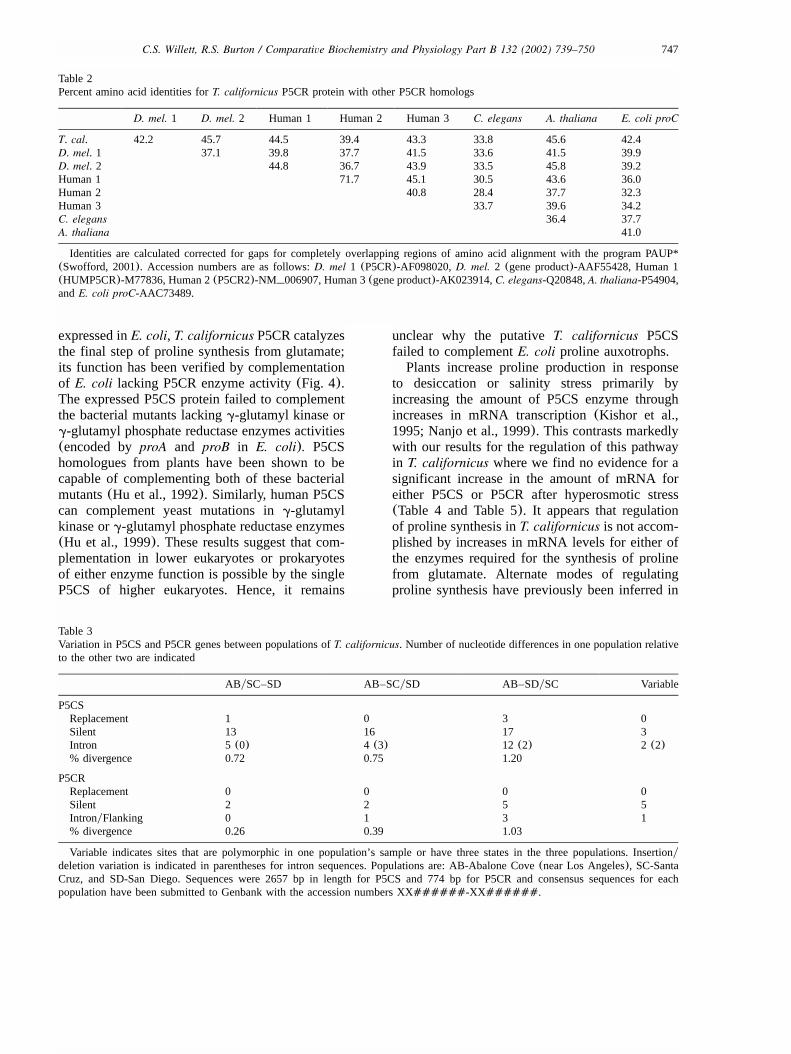
Table 2Percent amino acid identities forT. californicus P5CR protein with other P5CR homologs
D. mel. 1 D. mel. 2 Human 1 Human 2 Human 3 C. elegans A. thaliana E. coli proC
T. cal. 42.2 45.7 44.5 39.4 43.3 33.8 45.6 42.4D. mel. 1 37.1 39.8 37.7 41.5 33.6 41.5 39.9D. mel. 2 44.8 36.7 43.9 33.5 45.8 39.2Human 1 71.7 45.1 30.5 43.6 36.0Human 2 40.8 28.4 37.7 32.3Human 3 33.7 39.6 34.2C. elegans 36.4 37.7A. thaliana 41.0
Identities are calculated corrected for gaps for completely overlapping regions of amino acid alignment with the program PAUP*(Swofford, 2001). Accession numbers are as follows:D. mel 1 (P5CR)-AF098020,D. mel. 2 (gene product)-AAF55428, Human 1(HUMP5CR)-M77836, Human 2(P5CR2)-NM 006907, Human 3(gene product)-AK023914,C. elegans-Q20848,A. thaliana-P54904,andE. coli proC-AAC73489.
Table 3Variation in P5CS and P5CR genes between populations ofT. californicus. Number of nucleotide differences in one population relativeto the other two are indicated
ABySC–SD AB–SCySD AB–SDySC Variable
P5CSReplacement 1 0 3 0Silent 13 16 17 3Intron 5 (0) 4 (3) 12 (2) 2 (2)% divergence 0.72 0.75 1.20
P5CRReplacement 0 0 0 0Silent 2 2 5 5IntronyFlanking 0 1 3 1% divergence 0.26 0.39 1.03
Variable indicates sites that are polymorphic in one population’s sample or have three states in the three populations. Insertionydeletion variation is indicated in parentheses for intron sequences. Populations are: AB-Abalone Cove(near Los Angeles), SC-SantaCruz, and SD-San Diego. Sequences were 2657 bp in length for P5CS and 774 bp for P5CR and consensus sequences for eachpopulation have been submitted to Genbank with the accession numbers XX������-XX������.
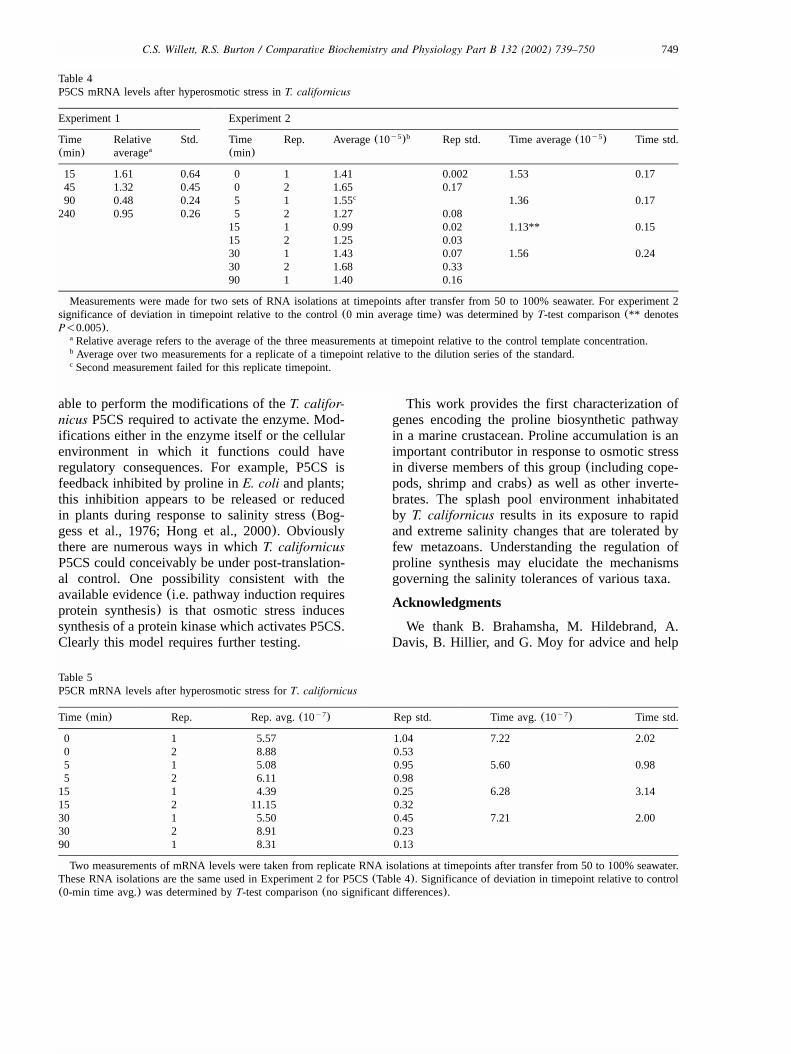
expressed inE. coli, T. californicus P5CR catalyzesthe final step of proline synthesis from glutamate;its function has been verified by complementationof E. coli lacking P5CR enzyme activity(Fig. 4).The expressed P5CS protein failed to complementthe bacterial mutants lackingg-glutamyl kinase org-glutamyl phosphate reductase enzymes activities(encoded byproA and proB in E. coli). P5CShomologues from plants have been shown to becapable of complementing both of these bacterialmutants(Hu et al., 1992). Similarly, human P5CScan complement yeast mutations ing-glutamylkinase org-glutamyl phosphate reductase enzymes(Hu et al., 1999). These results suggest that com-plementation in lower eukaryotes or prokaryotesof either enzyme function is possible by the singleP5CS of higher eukaryotes. Hence, it remains
unclear why the putativeT. californicus P5CSfailed to complementE. coli proline auxotrophs.
Plants increase proline production in responseto desiccation or salinity stress primarily byincreasing the amount of P5CS enzyme throughincreases in mRNA transcription(Kishor et al.,1995; Nanjo et al., 1999). This contrasts markedlywith our results for the regulation of this pathwayin T. californicus where we find no evidence for asignificant increase in the amount of mRNA foreither P5CS or P5CR after hyperosmotic stress(Table 4 and Table 5). It appears that regulationof proline synthesis inT. californicus is not accom-plished by increases in mRNA levels for either ofthe enzymes required for the synthesis of prolinefrom glutamate. Alternate modes of regulatingproline synthesis have previously been inferred in

748 C.S. Willett, R.S. Burton / Comparative Biochemistry and Physiology Part B 132 (2002) 739–750
Fig. 4. Complementation ability ofT. californicus P5CR forE. coli proline auxotrophs. Plate in(A) is minimal media with inducerL-arabinose while plate in(B) is minimal media plus repressor glucose. Strain G13(proB mutant) was transformed with plasmid isolatedfrom x -6 �3 bacterial strain and plated on non-selective media. Six of these colonies are streaked in the first row of each plate. Six342
colonies from non-selective transformation ofx with T. californicus P5CR containing plasmid are plated in the second row of each342
plate.
other systems; e.g. certain plant tissues that accu-mulate high levels of proline independent of stress(e.g. mature grapes) do not have increases in theamount of P5CS mRNA or protein coincident withincreases in proline concentration(Stines et al.,1999).
What are some possible models for prolinesynthesis regulation under hyperosmotic stress inT. californicus? One possibility is translationalcontrol. Under this scenario mRNA for the rate-limiting enzyme is always present but translationoccurs only under conditions of hyperosmoticstress. This model would still be consistent withprevious results demonstrating that protein synthe-sis is essential for proline accumulation(Burton,1991a). The prediction of this model is an increase
in the protein levels of either P5CS or P5CR underhyperosmotic stress leading to increased prolinesynthesis. The generation of specific antibodies tothese proteins will facilitate the testing of thishypothesis.
A second possible mode of regulation would bemodification of either the P5CS or P5CR proteinsafter translation. Under this mechanism we wouldpredict that the amount of protein for the rate-limiting enzyme would stay the same but theactivity would change upon hyperosmotic stress.The inability of theT. californicus P5CS to com-plement theg-glutamyl kinase org-glutamyl phos-phate reductase mutations inE. coli might beexplained by the need for post-translational acti-vation of P5CS. PresumablyE. coli would not be
749C.S. Willett, R.S. Burton / Comparative Biochemistry and Physiology Part B 132 (2002) 739–750
Table 4P5CS mRNA levels after hyperosmotic stress inT. californicus
Experiment 1 Experiment 2
Time Relative Std. Time Rep. Average(10 )y5 b Rep std. Time average(10 )y5 Time std.(min) averagea (min)
15 1.61 0.64 0 1 1.41 0.002 1.53 0.1745 1.32 0.45 0 2 1.65 0.1790 0.48 0.24 5 1 1.55c 1.36 0.17
240 0.95 0.26 5 2 1.27 0.0815 1 0.99 0.02 1.13** 0.1515 2 1.25 0.0330 1 1.43 0.07 1.56 0.2430 2 1.68 0.3390 1 1.40 0.16
Measurements were made for two sets of RNA isolations at timepoints after transfer from 50 to 100% seawater. For experiment 2significance of deviation in timepoint relative to the control(0 min average time) was determined byT-test comparison(** denotesP-0.005).
Relative average refers to the average of the three measurements at timepoint relative to the control template concentration.a
Average over two measurements for a replicate of a timepoint relative to the dilution series of the standard.b
Second measurement failed for this replicate timepoint.c
Table 5P5CR mRNA levels after hyperosmotic stress forT. californicus
Time (min) Rep. Rep. avg.(10 )y7 Rep std. Time avg.(10 )y7 Time std.
0 1 5.57 1.04 7.22 2.020 2 8.88 0.535 1 5.08 0.95 5.60 0.985 2 6.11 0.98
15 1 4.39 0.25 6.28 3.1415 2 11.15 0.3230 1 5.50 0.45 7.21 2.0030 2 8.91 0.2390 1 8.31 0.13
Two measurements of mRNA levels were taken from replicate RNA isolations at timepoints after transfer from 50 to 100% seawater.These RNA isolations are the same used in Experiment 2 for P5CS(Table 4). Significance of deviation in timepoint relative to control(0-min time avg.) was determined byT-test comparison(no significant differences).
able to perform the modifications of theT. califor-nicus P5CS required to activate the enzyme. Mod-ifications either in the enzyme itself or the cellularenvironment in which it functions could haveregulatory consequences. For example, P5CS isfeedback inhibited by proline inE. coli and plants;this inhibition appears to be released or reducedin plants during response to salinity stress(Bog-gess et al., 1976; Hong et al., 2000). Obviouslythere are numerous ways in whichT. californicusP5CS could conceivably be under post-translation-al control. One possibility consistent with theavailable evidence(i.e. pathway induction requiresprotein synthesis)is that osmotic stress inducessynthesis of a protein kinase which activates P5CS.Clearly this model requires further testing.
This work provides the first characterization ofgenes encoding the proline biosynthetic pathwayin a marine crustacean. Proline accumulation is animportant contributor in response to osmotic stressin diverse members of this group(including cope-pods, shrimp and crabs) as well as other inverte-brates. The splash pool environment inhabitatedby T. californicus results in its exposure to rapidand extreme salinity changes that are tolerated byfew metazoans. Understanding the regulation ofproline synthesis may elucidate the mechanismsgoverning the salinity tolerances of various taxa.
Acknowledgments
We thank B. Brahamsha, M. Hildebrand, A.Davis, B. Hillier, and G. Moy for advice and help

750 C.S. Willett, R.S. Burton / Comparative Biochemistry and Physiology Part B 132 (2002) 739–750
with microbiology and molecular biology tech-niques. We also thank L. Csonka for providingE.coli strains. M. Servedio and J. Flowers providedhelpful comments on the manuscript. Work sup-ported by National Science Foundation grant(DEB98-15424)to RSB.
References
Bishop, J.S., Burton, R.S., 1993. Amino acid synthesis duringhyperosmotic stress inPenaeus aztecus post-larvae. Comp.Biochem. Physiol. 106A, 49–56.
Boggess, S.F, Aspinall, D., Paleg, L.G., 1976. Stress metabo-lism IX. The significance of end-product inhibition ofproline biosynthesis and of compartmentation in relation tostess-induced proline accumulation. Aust. J. Plant Physiol.3, 513–525.
Burg, M.B., Kwon, E.D., Kultz, D., 1996. Osmotic regulation¨of gene expression. FASEB J. 10, 1598–1606.
Burton, R.S., 1986. Incorporation of C-bicarbonate into the14
free amino acid pool during hyperosmotic stress in anintertidal copepod. J. Exper. Zool. 238, 55–61.
Burton, R.S., 1991a. Regulation of proline synthesis duringosmotic stress in the copepodTigriopus californicus. J.Exper. Zool. 259, 166–173.
Burton, R.S., 1991b. Regulation of proline synthesis in osmoticresponse: effects of protein synthesis inhibitors. J. Exper.Zool. 259, 272–277.
Burton, R.S., 1992. Proline synthesis during osmotic stress inmegalopa stage larval of the blue crabCallinectes sapidus.Biol. Bull. 182, 409–415.
Burton, R.S., 1998. Intraspecific phylogeography across thePoint Conception biogeographic boundary. Evolution 52,734–745.
Burton, R.S., Feldman, M.W, 1982. Changes in free aminoacid concentrations during osmotic response in the intertidalcopepodTigriopus californicus. Comp. Biochem. Physiol.73A, 441–445.
Delauney, A.J., Verma, D.P.S., 1990. A soybean gene encodingd -pyrroline-5-carboxylate reductase was isolated by func-1
tional complementation inEscherichia coli and is found tobe osmoregulated. Mol. Gen. Genet. 221, 299–305.
Garfin, D.E., 1990. One-dimensional gel electrophoresis.. In:Deutscher, M.P.(Ed.), Guide to Protein Purification. Aca-demic Press, San Diego, CA, pp. 425–441.
Gilles, R., 1979. Intracellular organic osmotic effectors. In:Gilles, R(Ed.), Mechanisms of Osmoregulation in Animals.John Wiley and Sons, New York, NY, pp. 111–156.
Goolish, E.M., Burton, R.S., 1989. Energetics of osmoregula-tion in the intertidal copepodTigriopus californicus. Funct.Ecol. 3, 81–89.
Hare, P.D., Cress, W.A., Van Staden, J., 1998. Dissecting theroles of osmolyte accumulation during stress. Plant CellEnviron. 21, 535–553.
Hong, Z., Lakkineni, K., Zhang, Z., Verma, D.P.S., 2000.Removal of feedback inhibition ofd -pyrroline-5-carboxy-1
late synthase results in increased proline accumulation andprotection of plants from osmotic stress. Plant Physiol. 122,1136–1229.
Hu, C.-A.A., Delauney, A.J., Verma, D.P.S., 1992. A bifunc-tional enzymed -pyrroline-5-carboxylate synthase) catalyz-1
es the first two steps in proline biosynthesis in plants. Proc.Nad. Acad. Sci. USA 89, 9354–9358.
Hu, C.-A.A., Lin, W.-W., Obie, C., Valle, D., 1999. Molecularenzymology of mammaliand -pyrroline-5-carboxylate syn-1
thase. J. Biol. Chem. 274, 6754–6762.Hua, X.J., van de Cotte, B., Van Montagu, M., Verbruggen,
N., 2000. The 59 untranslated region of theAt-P5CR geneis involved in both transcriptional and post-transcriptionalregulation. Plant J. 26, 157–169.
Kempf, B., Bremer, E., 1998. Upttake and synthesis of com-patible solutes as microbial stress responses to high-osmo-lality environments. Arch. Microbiol. 170, 319–330.
Kishor, P.B.K., Hong, Z., Miao, G.-H., Hu, C.-A.A., Verma,D.P.S., 1995. Overexpression of -pyrroline-5-carboxylate1
synthase increases proline production and confers osmoto-lerance in transgenic plants. Plant Physiol. 108, 1387–1394.
McAllen, R.J., Taylor, A.C, Davenport, J., 1998. Osmotic andbody density response in the harpacticoid copepodTigriopusbrevicornis in supralittoral rock pools. J. Mar. Biol. Ass.UK 78, 1143–1153.
Misener, S.R., Walker, V.K., 2001. Complementation, cloning,and characterization of the pyrroline 5-carboxylate reductasegene fromDrosophila melanogaster. Biochem. Genet. 39,15–31.
Nanjo, T., Kobayashi, M., Yoshiba, Y., et al., 1999. Biologicalfunctions of proline in morphogenesis and osmotolerancerevealed in antisense transgenicArabidopsis thaliana. PlantJ. 18, 185–193.
Rawson, P.D., Brazeau, D.A., Burton, R.S., 2000. Isolationand characterization of cytochromec from the marinecopepodTigriopus californicus. Gene 248, 15–22.
Stines, A.P., Naylor, D.J., Høj, P.B., van Heeswijck, R., 1999.Proline accumulation in developing grapevine fruit occursindependently of changes in the levels -pyrroline-5-carbox-1
ylate syntheses mRNA or protein. Plant Physiol. 120,923–931.
Swofford, D.L., 2001. PAUP*: Phylogenetic Analysis UsingParsimony(*and other Methods) (Sinauer Associates, Sun-derland, MA), Version 4.
Willett, C.S., Burton, R.S., 2001. Viability of cytochromecdepends on cytoplasmic background inTigriopus californi-cus. Evolution 55, 1592–1599.
Yancey, P.H., 1994. Compatible and counteracting solutes. In:Strange, K.(Ed.), Cellular and Molecular Physiology ofCell Volume Regulation. CRC Press, Ann Arbor, MI, pp.81–110.
Yancey, P.H., Clark, M.E, Hand, S.C., Bowlus, R.D., Somero,G.N., 1982. Living with water stress: evolution of osmolytesystem. Science 217, 1214–1222.
Copyright © 2022 FDOKUMEN




















