
Cellulase Production by Pink Pigmented Facultative Methylotrophic Strains (PPFMs
Upload
independentCategory
view
0download
0
Exp Brain Res (1982) 48:228-237 Ex imental Brain Research �9 Springer-Verlag 1982
Projections of the Medial Terminal Nucleus of the Accessory Optic System Upon Pretectal Nuclei in the Pigmented Rat
R. H. I. Blanks 1'2, R.A. Giolli 1'3'4, and S.V. Pham 1
1 Dept. of Anatomy, California College of Medicine, University of California, Irvine, CA 92717, USA 2 Dept. of Surgery (Otolaryngology), California College of Medicine, University of California, Irvine, CA 92717, USA 4 Dept. of Psychobiology, University of California, Irvine, CA 92717, USA
Summary. The projections of the medial terminal nucleus (MTN) of the accessory optic system (AOS) upon pretectal nuclei have been studied in pigmented rats by means of (i) the anterograde transport of 3H- leucine with the use of light autoradiography and (ii) the retrograde transport of horseradish peroxidase (HRP). Injections of 3H-leucine largely restricted to the MTN and minimally involving adjacent ventral midbrain structures, produced heavy terminal axonal labeling within the ipsilateral nucleus of the optic tract (NOT) and the dorsal terminal nucleus (DTN) of the AOS. Terminal labeling was observed in all superficial portions of the NOT, except for a small ventromedial segment in the rostral two thirds and a larger medial segment in the caudal one third of this nucleus. Thus the MTN-NOT projections we describe entirely overlap the retinal-NOT projection and partially overlap the visual cortical-NOT, as reported by others. Within the DTN, the dense terminal fields covered the entire nucleus.
After postinjection survival times of 3-7 days, the pattern of axonal labeling showed that the MTN- NOT projection consisted of three bundles: (i) a superficial mesencephalic bundle coursing within the superior fasciculus, posterior fibers of the AOS which enters the caudal portions of the NOT and the DTN; (ii) a deep mesencephalic bundle that tra- versed the midbrain tegmentum dorsolaterally, also reaching the caudal one-half of the NOT and all of the DTN; and (iii) a mesodiencephalic bundle that passed first laterally through midbrain tegmentum and then dorsally through lateral thalamus to enter the rostral one-half of the NOT.
2 Supported by USPHA research grant EY03018 and Research Career Development Award EY000160 from the National Eye Institute
3 Supported by USPHA research grant EY03642 from the National Eye Institute
Offprint requests to: Dr. Robert H.I. Blanks (address see above)
Pretectal injections of HRP that invade the NOT and DTN produced retrograde labeling of most (ca. 75%) of the neurons of the ipsilateral MTN, without labeling the adjacent substantia nigra or ventral tegmental area. This finding confirms our autoradio- graphic data by showing that the MTN provides the major, ventral tegmental projection to the NOT and DTN. The present finding of a MTN-NOT projec- tion, combined with available anatomical and phy- siological data, suggests that the MTN may play a more significant role in visual-vestibular aspects of oculomotor control than formerly thought.
Key words: Mammalian accessory optic system - Medial terminal nucleus - Efferent projections - HRP and light autoradiographic techniques - Visual- oculomotor pathways
Introduction
An accessory optic system (AOS) has been described in virtually all vertebrate species in which it has been studied (Marg 1973). Its organization and degree of development varies between species of different vertebrate classes as well as between species of the same order, but, aside from members of the order Primates, one feature characterizes all vertebrates thus far studied: the chief terminus for accessory optic axons is the medial terminal nucleus (MTN) of the AOS defined by Hayhow and coworkers (see Hayhow 1959, 1966; Hayhow et al. 1960). In addi- tion, there is now anatomical evidence for non- retinal projections to the MTN (as well as to the DTN and LTN) from the substantia nigra, posterior thalamus, nucleus of the optic tract (NOT) and mesencephalic reticular formation (Giolli et al. 1968; Carpenter and Pierson 1973; Itoh 1977; Simpson and
0014-4819/82/0048/0228/$ 2.00

R. H. I. Blanks et al. : Projections of the Medial Terminal Nucleus 229
S o o d a k 1978; T e r a s a w a et al. 1979; H o l s t e g e and Col lewi jn 1980; B e r s o n and G r a y b i e l 1980). M o r e o v e r , in the cat and m o n k e y t h e r e a re neocor t i - cal p ro j ec t i ons f rom the visual co r tex ( G a r e y et al. 1968; C a m p o s - O r t e g a et al, 1970; B e r s o n and G r a y - biel 1980), a l though in r abb i t no visual cor t ica l p ro j ec t i on to the M T N could be f o u n d using e i ther f iber d e g e n e r a t i o n m e t h o d s (Gio l l i and G u th r i e 1971) or 3H-amino acid l ight a u t o r a d i o g r a p h y (Gio l l i et al. 1978; Hol l f inder e t al. 1979). These da ta , c o m b i n e d with the ava i l ab le e l ec t rophys io log ica l and les ion s tudies (see Discuss ion) , suggest tha t the M T N is involved in s o m e aspec t of v i s u o m o t o r cont ro l . H o w e v e r , m o r e i n f o r m a t i o n on the e f fe ren t p ro jec - t ions of this nucleus is r e q u i r e d b e f o r e the func t iona l role of the M T N can be d e t e r m i n e d .
A de ta i l ed account of the p r o j e c t i o n s of the p igeon M T N (ca l led nucleus of the basa l opt ic roo t ) has been given by B r e c h a et al. (1980), bu t da t a in m a m m a l s r ega rd ing M T N ef feren ts is m e a g e r and con t rad ic to ry . The p r e s e n t r e p o r t concerns wha t we have found to be one of the m a j o r p r o j e c t i o n s of the M T N in the ra t i . e . , a la rge , h ighly specif ic , to ta l ly uncrossed p r o j e c t i o n ar is ing f rom b o t h ven t ra l and dorsa l divisions of the M T N and t e rmina t i ng wi thin the nucleus of the op t ic t rac t ( N O T ) and the dorsa l t e rmina l nucleus ( D T N ) of the A O S . Pa r t i a l resul ts were pub l i shed as an abs t rac t (Gio l l i e t al. 1981).
Methods
In a first series of experiments, 13 pigmented rats (Long-Evans strain) were anesthetized with 35% chloral hydrate (0.1 ml/100 g b.wt.) and a small craniotomy was performed. The MTN was localized using the stereotaxic coordinates of Fifkovgt and MarSala (1967) and injected with 0.05-0.1 ul of tritiated leucine (4-538 L- leucine, specific activity 58.5 Ci/mmol, New England Nuclear) through a glass micropipette (tip diameter = 4-6 ~m) connected to a pressure injection system. Before injection, the isotope was dried and reconstituted in sterile 0.9% NaC1 to give a final radioactivity of 50 ~tCi/ul. Thus, from 2.5-5.0 uCi of isotope was injected into each animal. Following postinjection survival periods of as short as 2 days, to allow for terminal fields, or as long as 8 days, to show axonal trajectories, the rats were killed by transcar- dial perfusion with 10% formalin; blocks of their brains were then dehydrated, embedded in paraffin and sectioned in the transverse plane at 15 ~tm. Every section was subsequently mounted onto slides and processed according to the autoradiographic (ARG) technique of Cowan et al. (1972) which includes coating with Kodak NTB-2 emulsion, storage in the dark for the two weeks at 5 ~ C, development in freshly prepared D-19, and counterstaining with 0.12% thionin.
In a second series of experiments, 3 rats were surgically operated on as described above for the ARG experiments. Finely drawn glass micropipettes (tip diameter = 1 ~tm) filled with a 5% solution of HRP (Boehringer-Mannheim, 10809-0) in 0.5% KC1 were inserted stereotaxically into the pretectum. Iontophoretic injections of 1 ~tA, DC current for 2-5 rain were generally sufficient to produce injection sites less than 0.75 mm in dia.
Following a 24-30 h post-injection survival period, the rats were again anesthetized and transcardially perfused with 1 1 of cold saline followed by 2 1 of a fixative consisting of 1.25% paraform- aldehyde and 1.0% glutaraldehyde in 0.1 M phosphate buffer. The brains were stored overnight at 4 ~ C in 0.1 M phosphate buffer containing 30% sucrose and then sectioned at 40 ~tm. Free floating sections were reacted in tetramethyl benzidine/hydrogen peroxide according to the protocol of deOlmos et al. (1978). The sections were subsequently mounte d from 0.9% saline at 4 ~ C onto gelatin- chrome alum subbed slides, counterstained lightly with 0.12% thionin at 5 ~ C, and coverslipped.
To demonstrate the precise locations of 3H-leucine label within brain nuclei and pathways, appropriate sections were drawn using a Trisimplex Bausch and Lomb projector. Labeled axons and terminal fields, as seen in the autoradiographically prepared sections, were examined under darkfield illumination and plotted onto the drawings. The position and numbers of HRP-labeled neurons were similarly plotted using a Zeiss microscope equipped with a drawing tube.
Results
Tritiated Amino Acid Injections into M T N
The M T N was successful ly i n j ec t ed in 7 animals . The mos t conspicuous f inding in these cases was the p resence of h e a v y t e r m i n a l l abe l ing wi th in two com- ponen ts of the p re t ec t a l complex ; the N O T , as de f ined in the ra t by Scal ia (1972), and wi th in the D T N , as de l i ne a t e d in the ra t by H a y h o w et al. (1960) 1. T h e r e was no ind ica t ion of an M T N pro jec - t ion to the an te r io r , pos t e r i o r or o l ivary p re t ec t a l nuclei o r to the m e d i a l p r e t ec t a l a r ea de l im i t ed by Rose (1942). Case 128-81, was typica l of the 7 cases s tud ied (Fig. 1 and 3A) . The axona l t e rmina l f ields covered superf ic ia l po r t i ons of mos t of the ros t ra l t h ree -qua r t e r s of the N O T (Figs. 1 A - C ; 2 A - C ) , thus over l app ing the r e t i n a l - N O T p r o j e c t i o n zone as desc r ibed in the ra t (Scal ia 1972; Scal ia and A r a n g o 1979). In Figs. 1C and 2C, the f ie ld of l abe l ed te rmina ls a re seen to con t inue into the ex tens ion of the N O T , desc r ibed by Scal ia (1972) wi th in the s t ra tum op t i cum of the s u p e r i o r coll iculus. H o w e v e r , at the mos t cauda l level of the N O T , the m e d i a l quadran t s , which c o r r e s p o n d to the p r i m e focus of the visual cor t ica l p r o j e c t i o n to the N O T as r e p o r t e d in o p o s s u m (L inden and R o c h a - M i r a n d a 1981), rab- bi t (Giol l i and G u t h r i e 1971) and cat ( U p d y k e 1977), were only l ight ly l a b e l e d or u n l a b e l e d (Figs. 1C, D;
1 The terminology regarding the components of the pretectal complex and the AOS is often confusing. The DTN is located adjacent to the posterior pole of the NOT and for this reason is considered here as part of the pretectum. The physiological receptive field properties of DTN neurons are similar to those of the NOT and different from the other AOS nuclei (Simpson et al. 1979) which further supports this association. However, the DTN is also a component of the AOS given the defination that the terminal nuclei of the AOS receive a differential retinal input via fibers of the accessory optic tracts (Hayhow et al. 1960)

Fig. l A D (legend see p 231)

R. H.I. Blanks et al.: Projections of the Medial Terminal Nucleus 231
Fig. 1. A-E Serial transverse sections through the pretectal complex, arranged from rostral to caudal (A to E), show the distribution of terminal axonal labeling within the NOT and DTN following injection of 3H-leucine into the ipsilateral MTN (rat 128). The corresponding brightfield (left) and darkfield (right) photomicrographs are from five representative levels of the pretectum. The cytoarchitectural boundaries of the NOT (A-D) and DTN (E) are indicated by the dashed line in the darkfield photomicrographs. Arrows a and b (C-E) point to two of the three axonal bundles shown to contribute to the terminal axonal fields in the NOT and DTN
2D). The DTN is uniformly covered by a highly dense zone of silver grains (Figs. 1E; 2E).
We could distinguish three bundles that arose from the MTN and terminated in the NOT and DTN. The axons of the superficial mesencephalic bundle coursed through the superior fasciculus, posterior- fibers of the AOS (AOS-SFp) to reach the pretectal region at the level of the DTN (Fig. 2D, E). Enroute, a few of these axons gave rise to a sparse terminal field within the lateral terminal nucleus (LTN) (Fig. 2D). A second, deep mesencephalic bundle origi- nated chiefly from the ventral division of the MTN (MTNv), ran through the dorsal division (MTNo), and coursed dorsolaterally through the midbrain tegmentum to enter the caudal half of the NOT and the DTN (Fig. 2D, E). This is the largest of the three bundles and was of particular interest because some of its axons, perhaps collateral branches, gave rise to a dense terminal field within the MTNa (Fig. 2E) before they proceeded to the pretectum. Finally, a mesodiencephalic bundle arose from the MTNv, coursed laterally through the midbrain tegmentum and continued into the lateral thalamus (Fig. 2C, D). After joining the optic tract fibers, this bundle coursed through the lateral geniculate nuclei and pulvinar and then entered the rostral half of the NOT (Fig. 2C).
Although we could trace three distinct fiber bundles from the MTN to the pretectum, the blend- ing of their terminal fields within the NOT and DTN made it difficult to determine whether there was a differential pretectal input for each bundle.
The MTN-pretectal projections described above were observed only when the injection sites were confined to the MTN (5 experimental cases, see Fig. 3A for an example), or partially included the MTN (2 experimental cases). Punctate, injections into adja- cent nuclei i.e., the substantia nigra (2 cases), subcuneiform nucleus (2 cases) and the mammillary nuclei (2 cases), failed to provide evidence for nigral, reticular or hypothalamic afferents to either NOT or DTN. These findings strongly suggest that in the rat the MTN is the primary, if not the sole, source of axonal projections from ventral tegmentum to pre- tectal nuclei.
H R P In jec t ions into P r e t e c t u m
Because of the proximity of the MTN to the substan- tia nigra and to other ventral tegmental nuclei, it was unavoidable that our injections of 3H-leucine margi- nally included these areas and thus may have contri- buted to pretectal labeling. In order to confirm the ARG results, and to provide additional details on the MTN-pretectal projection, we used the method of retrograde transport of HRP.
Following injections of HRP into the pretectum in three experimental cases, all of which included large portions of NOT and the DTN, numerous labeled neurons were found within the MTN (Figs. 3B, C and 4). Cellular labeling within the MTN was entirely ipsilateral and, remarkably involved an over- whelmingly large proportion of neurons in both MTN v and MTNo. The distribution of HRP-labeled

232 R.H.I. Blanks et al.: Projections of the Medial Terminal Nucleus
A
SubtN
v
B 128-23.6
SGS Aq
SGS
C 128-23.2
fw ZI , ,
SNr
MB IMTN. - HITr
D 128-22.4
MTN v
SNr
E 128-21.8
[! l I ,J
q i l l I i I
MGN
AOS-SFp
Fig. 2. A-E Serial transverse sections in rostral to caudal sequence (A-E) illustrating the axonal (dashed lines) and terminal labeling (dots) in the lateral thalamus, midbrain tegmentum and pretectum following injection of 3H-leucine into the ipsilateral MTN. The extent of the injection site in this experimental case (rat 128) is shown by black area in C-E. Note that the arrows in D and E point to labeled axons within the AOS-SFp
neurons within the ventral t egmentum is illustrated in Fig. 4A-C. In this experimental case (118-81), the boundaries of the injection site which includes the "dense core" plus the flair encompassed the MTN projection zone within the pre tec tum and also involved portions of other pretectal nuclei, the medial part of the pulvinar, and the rostral edge of the superior colliculus (Fig. 4D). Neuronal counts indicated that approximately 75% of the neurons in the MTN d (94/127) and MTNv (811/905) project to
pretectum. However , because of the highly cellular composition of the MTNv, 8 of 9 of the MTN-NO T / DTN neurons were of MTNv origin. The pretectum- projecting neurons of both divisions were heterogeneous in size and shape, involving large and small triangular, fusiform and round neurons of the nucleus similar to these identified by Giolli et al. (1968) in Golgi sections of rabbit brain. Comparable results were obtained in the other two experimental cases analyzed.
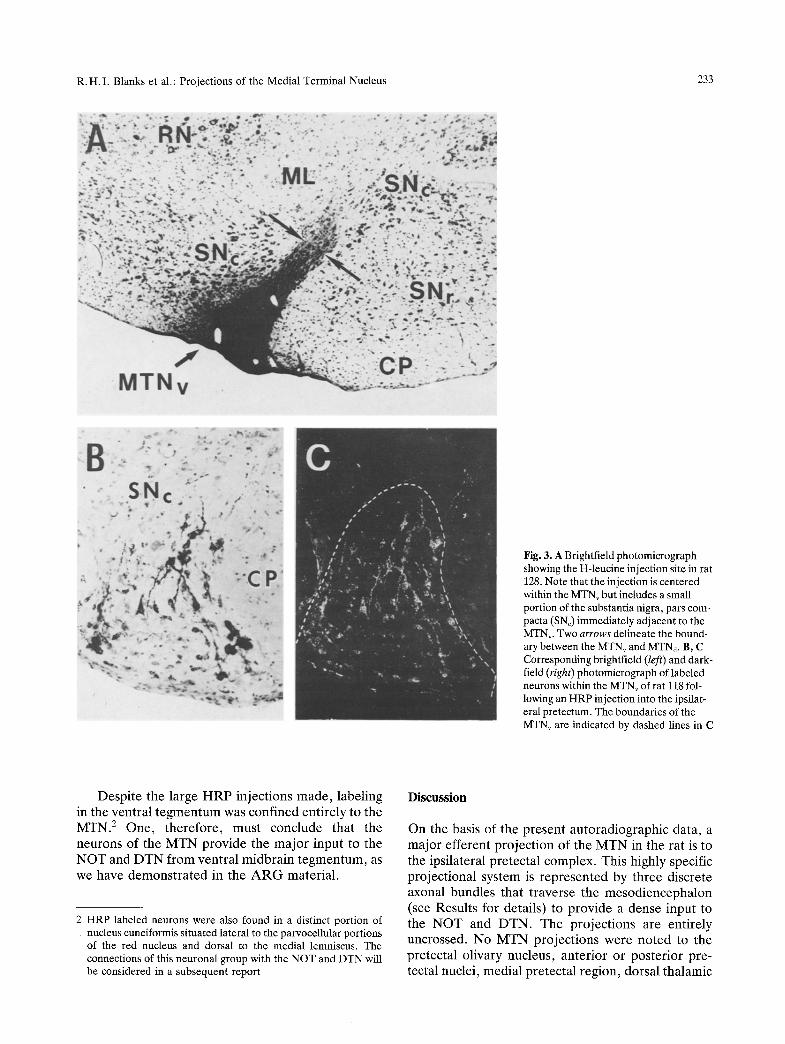
R.H.I. Blanks et al.: Projections of the Medial Terminal Nucleus 233
Fig. 3. A Brightfield photomicrograph showing the H-leucine injection site in rat 128. Note that the injection is centered within the MTN v but includes a small portion of the substantia nigra, pars com- pacta (SNc) immediately adjacent to the MTNv. Two arrows delineate the bound- ary between the MTNv and MTNd. B, C Corresponding brightfield (left) and dark- field (right) photomicrograph of labeled neurons within the MTNv of rat 118 fol- lowing an HRP injection into the ipsilat- eral pretectum. The boundaries of the MTNv are indicated by dashed lines in C
Desp i t e the la rge H R P in jec t ions m a d e , labe l ing in the ven t ra l t e g m e n t u m was conf ined en t i re ly to the M T N . 2 One , t he r e fo r e , mus t conc lude tha t the neurons of the M T N p r o v i d e the m a j o r inpu t to the N O T and D T N f rom ven t ra l m i d b r a i n t e g m e n t u m , as we have d e m o n s t r a t e d in the A R G mate r i a l .
2 HRP labeled neurons were also found in a distinct portion of nucleus cuneiformis situated lateral to the parvocellular portions of the red nucleus and dorsal to the medial lemniscus. The connections of this neuronal group with the NOT and DTN will be considered in a subsequent report
Discussion
On the basis of the p r e s e n t a u t o r a d i o g r a p h i c da ta , a m a j o r e f ferent p r o j e c t i o n of the M T N in the ra t is to the ips i la tera l p r e t ec t a l complex . This h ighly specific p ro j ec t i ona l sys tem is r e p r e s e n t e d by th ree d iscre te axona l bundles tha t t r ave r se the m e s o d i e n c e p h a l o n (see Resul t s for deta i l s ) to p r o v i d e a dense input to the N O T and D T N . T h e p r o j e c t i o n s are en t i r e ly uncrossed . No M T N p r o j e c t i o n s were n o t e d to the p re t ec t a l o l ivary nucleus , an t e r io r or pos t e r i o r pre- tec ta l nuclei , med ia l p r e t ec t a l r eg ion , dorsa l tha lamic

234 R.H.I. Blanks et al.: Projections of the Medial Terminal Nucleus
.&
ML M.TN d
D OT
O T LGNd
LGN d
Pc OT
MTNv 4 ~ k ~ )>/,~c:~,~ ~8-,4.s ML MTNd , I MP ~ \ ~ _
�9 ~ SN
Fig. 4. A-D Drawings of labeled neurons within the MTN following an injection of HRP into the ipsilateral pretectum in rat 118. In A, B, and C, the MTN is depicted in black, and extended draw- ings of the MTN, as seen at each of the three levels, show the neuronal distribu- tion, size and shape. In these extended figures, HRP-labeled neurons of the MTN v and MTNd are shown as filled profiles; unlabeled neurons are indi- cated by open profiles. Small arrows in B demarcate MTNv from MTNd. The extent of the HRP-injection site is depicted by cross-hatching in D
nuclei, or the superior colliculus, except for that portion of the NOT described by Scalia (1972) which extends into the stratum opticum. These findings confirm a previous description by Brecha et al. (1980) of a homologous MTN-pretectal projection in pigeon. Also, an ipsilateral MTN-NOT projection has been briefly described in the rabbit (Simpson and Soodak 1978), and there are suggestions that the MTN in the cat may project to dorsal parts of the ipsilateral NOT (Berson and Graybiel 1980).
The autoradiographic evidence for an MTN-NOT projection in the rat have been confirmed by retro- grade transport experiments utilizing HRP. These experiments have revealed that projections to the NOT and DTN arise from both MTNv and MTNd, but that no other ventral tegmentum nucleus sends
detectable numbers of axons to these pretectal com- ponents. In particular, it is important to emphasize that the present MTN-pretectal projections are sepa- rate and distinct from the nigral-tectal pathway that arises primarily from the adjacent substantia nigra, pars reticulata, and that terminates within the inter- mediate and deep layers of the superior colliculus (Hopkins and Niessen 1976; Rinvik et al. 1976; Jayaraman et al. 1977; Anderson and Yoshida 1980).
The MTN-pretectal terminal fields were distrib- uted to the superficial portions of the N O T through- out most of its rostral-caudal extent. From the detailed retinotopic maps of the rat N O T provided by Scalia and Arango (1979), it can be inferred that the MTN projects to both the temporal (rostral NOT) and the nasal regions (caudal NOT) of the contralat-

R. H. I. Blanks et al.: Projections of the Medial Terminal Nucleus 235
eral visual field. Additionally, the MTN-NOT pro- jection overlaps, in part, the visual cortical projec- tion as demonstrated in non-rodent species (Garey and Powell 1968; Pierson and Carpenter 1974; Updyke 1977; Benevento et al. 1977; Kanaseki and Sprague 1974; Lent et al. 1976; Berman 1977; Kawamura et al. 1974; Linden and Rocha-Miranda 1981). Such a projection confirms the general finding that the superficial pretectum, like superficial por- tions of tectum in mammals, is the prime recipient of visual projections, whereas deeper portions of pre- tectal nuclei and tectum receive input from somatosensory and auditory pathways (Garey et al. 1968; Berkley and Mash 1978; Kuypers and Law- rence 1967; Linden and Rocha-Miranda 1981).
The rodent MTN has been subdivided into dorsal (MTNd) and ventral (MTNv) portions on the basis of cytoarchitecture (Wallenberg 1903; Kosaka and Hiraiwa 1915; Gillilan 1941; Hayhow et al. 1960) and differential retinal input via superior and inferior accessory optic fasciculi (Hayhow et al. 1960). One observation made in the present study further sup- ports this subdivision, viz., that both nuclear subdivi- sions project upon the NOT and DTN, but the MTN v also provides an axonal input to the MTNd, most probably via the abundant Golgi II neurons of the MTNv described from Golgi preparations of rabbit brain (Giolli et al. 1968) and/or through collateral branches of MTN~ axons projecting to pretectum. Functionally, the axonal projection system arising in the MTNv and terminating in the MTNd may play a role in regulating the high level of spontaneous activity reported in the rabbit MTN d by Hill and Marg (1963).
The strong MTN-NOT projection we have found confirms other evidence that the AOS and pretectum are extensively interconnected and that the MTN and NOT are functionally related to oculomotor events, particularly optokinetic nystagmus and visual-ves- tibular interaction. Several anatomical studies have shown connections between terminal accessory optic nuclei and between the NOT and these terminal nuclei (see Carpenter and Pierson 1973, rhesus monkey; Itoh 1977; Berson and Graybiel 1980, cat; Simpson and Soodak 1978, rabbit; Terasawa et al. 1979, rat; Brecha et al. 1980, pigeon; Holstege and Collewijn 1980, rabbit). Maekawa and Simpson have demonstrated this interaction electrophysiologicalty by showing that the AOS and lateral pretectum (NOT included) represent the first relay stations connecting the contralateral retina with the dorsal cap of Kooy (Maekawa and Simpson 1972, 1973), the portion of the inferior olive conveying a crossed climbing fiber input to the cerebellar flocculus (Alley et al. 1975; Hoddevik and Brodal 1977). The
descending dorsal cap (of Kooy) pathway, recently shown to arise from the DTN and NOT (Takeda and Maekawa 1976, 1980), is strongly inhibited by fibers that apparently arise from the MTN and ascend through the AOS-SFp to terminate in the DTN and NOT (Maekawa and Simpson 1972, 1973). That the NOT represents an essential relay station for hori- zontal optokinetic nystagmus (OKN) is evidenced by the fact that in the rabbit, electrical stimulation of the NOT evokes OKN (slow phase directed ipsilat- erally), whereas lesion of-the NOT abolishes OKN (Collewijn 1975, 1977). The hypothesis proposed by Oyster et al. (1972, 1980), i.e., that a class of direction- and speed-selective retinal ganglion cells in the visual streak of the rabbit project to the MTN and control OKN, has received support by several recent anatomical studies (Karten et al. 1977; Kimm et al. 1979; Oyster et al. 1980). The MTN may be involved in horizontal OKN, but it does not appear to be an essential link, for it persists, but is reduced in high stimulus-velocity gain following bilateral MTN lesions (Fite et al. 1979; Kimm et al. 1979).
Any consideration of the functional interaction of the MTN and NOT, must account for the apparent discrepancies in receptive field properties between these two nuclei. MTN neurons in the rabbit prefer vertical stimuli moving upwards and post riorly (Walley 1967; Simpson et al. 1979) and would be expected to control vertical and not horizontal OKN. In this regard many NOT (Collewijn 1975; Hoffmann and Schoppmann 1975; Cazin et al. 1980) and DTN neurons (Simpson et al. 1979) are excited by slowly moving, whole-field horizontal movement in a tem- poral-to-nasal direction. There is another class of cells in the cat NOT which shows a preferred orientation for vertical motion in an antero-ventral direction (Hoffman and Schoppmann 1975), 180 ~ opposite to the upward-posterior direction selective responses of MTN neurons (Simpson et al. 1979). This class of NOT responses might be accounted for on the basis of an inhibitory MTN-NOT interaction, for which electrophysiological evidence exists (Maekawa and Simpson 1972, 1973). However, further investigation is required before conclusions can be drawn regarding the function of the MTN- NOT connection.
Abbreviations to Figures
AOS-SFp - Accessory optic tract-superior fasciculus, posterior fibers; APN - Anterior pretectal nucleus; Aq - Cerebral aqueduct; BSC - Brachium of superior colliculus; CP - Cerebral peduncle; DTN - Dorsal terminal nucleus, accessory optic system; HITr - Habenulointerpeduncular tract; IpN - Interpeduncular nucleus; L - Lateral thalamic nuclear complex; LGNd - Lateral geniculate nucleus, pars dorsalis; LTN - Lateral terminal nucleus, accessory

236 R.H.I . Blanks et al.: Projections of the Medial Terminal Nucleus
optic system; LGNv - Lateral geniculate nucleus, pars ventralis; MB - Mammillary body; MGN - Medial geniculate nucleus; MGNp -Medial geulculate nucleus, pars principalis; ML - Medial lemniscus; MP - Mammillary peduncle; MTNd - Medial terminal nucleus, dorsal division, accessory optic system; MTNv - Medial terminal nucleus, ventral division, accessory optic system; NOT - Nucleus of the optic tract; OT - Optic tract; Pag - Periaqueductal grey; P c - Posterior commissure; P O N - Pretectal olivary nucleus; PPN- Posterior pretectal nucleus; Pr - Pretectal nuclear complex; Pu l - Pulvinar; RN - Red nucleus; S C - Superior coUiculus; SCc - Commissure of superior colliculus; SGS - Superior colliculus, stratum gresium superficiale; SNc - Substantia nigra, pars com- pacta; SNr - Substantia nigra, pars reticulata; so - Superior colliculus, stratum opticum; Subt N. - Subthalamic nucleus; V - Ventral thalamic nuclear complex; ZI - Zona incerta; III N - Ocnlomotor nerve
Acknowledgements. The authors gratefully acknowledge the assist- ance provided by Ms. Carol M. Williams and Ms. Stephanie A. Dresie in preparing the illustrations and Mrs. Natalie M. Sepion for the excellent effort in typing the manuscript. We would also like to thank Dr. Jerry Simpson for his helpful discussion of the manuscript.
References
Alley K, Baker R, Simpson JI (1975) Afferents to the vestibulo- cerebellum and the origin of the visual climbing fibers in the rabbit. Brain Res 98:582-589
Anderson ME, Yoshida M (1980) Axonal branching patterns and location of nigrothalamic and nigrocollicular neurons in the cat. J Neurophysiol (Lond) 43:883-895
Benevento LA, Rezak M, Santos-Anderson R (1977) An auto- radiographic study of the projections of the pretectum in the rhesus monkey (Macaca mulatta): evidence for sensorimotor links to the thalamus and oculomotor nuclei. Brain Res 127: 197-218
Berkley KJ, Mash DC (1978) Somatic sensory projections to the pretectum in the cat. Brain Res 158:445-449
Berman N (1977) Connections of the pretectum in the cat. J Comp Neurol 174:227-254
Berson DM, Graybiel AM (1980) Some cortical and subcortical fiber projections to the accessory optic nuclei in the cat. Neuroscience 5:2203-2217
Brecha N, Karten HJ, Hunt SP (1980) Projections of the nucleus of the basal optic root in the pigeon: An autoradiographic and horseradish peroxidase study. J Comp Neurol 189:615-670
Campos-Ortega JA, Hayhow WR, Cluver PF de V (1970) The descending projections from the cortical visual fields of Macaca mulatta with particular reference to the question of a cortico-lateral geniculate-pathway. Brain Behav Evol 3: 368-414
Carpenter MB, Pierson RJ (1973) Pretectal region and the pupillary light reflex. An anatomical analysis in the monkey. J Comp Neurol 149:271-300
Cazin L, Precht W, Lannou J (1980) Firing characteristics of neurons mediating optokinetic responses to rats vestibular neurons. Pfluegers Arch Physiol 386:221-230
Collewijn H (1975) Oculomotor areas in the rabbit's midbrain and pretectum. J Neurobiol 6:3-22
Collewijn H (1977) Optokinetic and vestibulo-ocular reflexes in dark-reared rabbits. Exp Brain Res 27:287-300
Cowan WM, Gottlieb DI, Hendrickson AE, Price JL, Woolsey TA (1972) The autoradiographic demonstration of axonal connections in the central nervous system. Brain Res 37: 21-51
deOlmos J, Hardy H, Heimer L (1978) The afferent connections of the main and the accessory olfactory bulb formations in the rat: an experimental HRP-study. J Comp Neurol 181:213-244
Fifkovfi E, Margala J (1967) Stereotaxic atlas for the cat, rabbit and rat. In: Bures J, Petran M, Zachar J (eds) Electrophy- siological methods in biological research. Academic Press, New York, pp 653-711
Fite KV, Reiner A, Hunt SP (1979) The accessory optic system and optokinetic nystagmus. Brain Behav Evol 16:192-202
Garey LJ, Jones EG, PoweU TPS (1968) Interrelationships of striate and extrastriate cortex with the primary relay sites of the visual pathway. J Neurol Neurosurg Psychiatry 31: 135-157
Garey LJ, Powell TPS (1968) The projection of the retina in the cat. J Anat 102:189-222
Gillilan LA (1941) The connections of the basal optic root (posterior accessory optic tract) and its nucleus in various mammals. J Comp Neurol 74:367-408
Giolli RA (1961) An experimental study of the accessory optic tracts (transpeduncular tracts and anterior accessory optic tracts) in the rabbit. J Comp Neurol 117:77-95
Giolli RA, Blanks RItI, Pham S (1981) Projections of medial terminal nucleus of accessory optic system to pretectum in pigmented rat. Neurosci Abstr 7:461
Giolli RA, Braithwaite JR, Streeter TF (1968) Golgi study of the nucleus of the transpeduncular tract in the rabbit. J Comp Neurol 133:309-328
Giolli RA, Guthrie MD (1971) Organization of subcortical projec- tions of visual areas I and II in the rabbit. An experimental degeneration study. J Comp Neurol 142:351-376
Giolli RA, Towns LC, Takahashi TT, Karamanlidis AN, Williams DD (1978) An autoradiographic study of the projections of visual cortical area I to the thalamus, pretectum and superior colliculus of the rabbit. J Comp Neurol 180:743-752
Hayhow WR (1959) An experimental study of the accessory optic fiber system in the cat. J Comp Neurol 113:281-314
Hayhow WR (1966) The accessory optic system in the marsupial phalanger, Trichosurus vulpecula. An experimental degenera- tion study. J Comp Neurol 126:653-672
Hayhow WR, Webb C, Jervie A (1960) The accessory optic fiber system in the rat. J Comp Neurol 115:187-216
Hill RM, Marg E (1963) Single cell responses of the nucleus of the transpeduncular tract in rabbit to monochromatic light on the retina. J Neurophysiol 26:249-257
Hoddevik GH, Brodal A (1977) The olivocerebellar projection studied with the method of retrograde axonal transport of horseradish peroxidase. V. The projections to the floc- culonodular lobe and the paraflocculus in the rabbit. J Comp Neurol 176:269-280
Hoffmann K-P, Schoppmann A (1975) Retinal input to direction selective cells in the nucleus tractus opticus of the cat. Brain Res 99:359-366
Holl~inder H, Tietze J, Distel H (1979) An autoradiographic study of the subcortical projections of the rabbit striate cortex in the adult and during postnatal development. J Comp Neurol 184: 783-794
Holstege G, Collewijn H (1980) The efferent connections of the nucleus of optic tract of the rabbit. Neurosci Lett [Suppl] 5: $183
Hopkins DA, Niessen LW (1976) Substantia nigra projections to the reticular formation, superior colliculus and central gray in the rat, cat and monkey. Neurosci Lett 2:253-259
Itoh K (1977) Efferent projections of the pretectum in the cat. Exp Brain Res 30:89-105
Jayaraman A, Batton R, Carpenter MB (1977) Nigrotectal projec- tions in the monkey: An autoradiographic study. Brain Res 135:147-152

R. H. I. Blanks et al.: Projections of the Medial Terminal Nucleus 237
Kanaseki T, Sprague JM (1974) Anatomical organization of pretectal nuclei and tectal laminae in the cat. J Comp Neurol 158:319-338
Karten HJ, Fite KV, Brecha N (1977) Specific projection of displaced retinal ganglion cells upon the accessory optic system in the pigeon (Columbia livia). Proc Natl Acad Sci USA 74:1753-1756
Kawamura S, Sprague JM, Niimi K (1974) Corticofugal projec- tions from the visual cortices to the thalamus, pretectum and superior coUiculus in the cat. J Comp Neurol 158:339-362
Kimm J, Winfield JA, Hendrickson AE (1979) Visual-vestibular interaction and the role of the flocculus in the vestibulo-ocular reflex. In: Granit R, Pompeiano O (eds) Reflex control of posture and movement. Elsevier/North Holland, Amsterdam, pp 703-714
Kosaka K, Hiraiwa K (1915) Zur Anatomie der Sehnervenbahnen und ihrer Zentren. Folia Neurobiol 9:367-389
Kuypers HGJM, Lawrence DG (1967) Cortical projections to the red nucleus and the brain stem in the rhesus monkey. Brain Res 4:151-188
Lent R, Cavalcante LA, Rocha-Miranda CE (1976) Retinofugal projections in the opossum. An anterograde degeneration and radioautographic study. Brain Res 107:9-26
Linden R, Rocha-Miranda CE (1981) The pretectal complex in the opossum: Projections from the striate cortex and correlation with retinal terminal fields. Brain Res 207:267-277
Maekawa K, Simpson JI (1972) Climbing fiber activation of Purkinje cells in the flocculus by impulses transferred through the visual pathway. Brain Res 39:245-251
Maekawa K, Simpson JI (1973) Climbing fiber responses evoked in vestibulocerebellum of rabbit from visual system. J Neurophysiol 36:649-666
Marg E (1973) Neurophysiology of the accessary optic system. In: Jung R (ed) Handbook of sensory physiology Springer, Berlin Heidelberg New York, pp 103-111
Oyster CW, Takahashi E, Collewijn H (1972) Direction-selective retinal ganglion cells and control of optokinetic nystagmus in the rabbit. Vision Res 12:183-193
Oyster CW, Simpson JI, Takahashi ES, Soodak RE (1980) Retinal ganglion cells proiecting to the rabbit accessory optic system. J Comp NeuroI 190:49-61
Pierson R J, Carpenter MB (1974) Anatomical analysis of pupillary reflex pathways in the rhesus monkey. J Comp Neurol 158: 121-144
Rinvik E, Grofova I, Ottersen OP (1976) Demonstration of nigrotectal and nigroreticular projections in the cat by axonal transport of proteins. Brain Res 112:388-394
Rose JE (1942) The thalamus of the sheep: Cellular and fibrous structure and comparison with pig, rabbit and cat. J Comp Neurol 77:469-523
Scalia F (1972) The termination of retinal axons in the pretectal region of mammals. J Comp Neurol 145:223-258
Scalia F, Arango V (1979) Topographic organization of the projections of the retina to the pretectal region in the rat. J Comp Neurol 186:271-292
Simpson JI, Soodak RE (1978) The accessory optic system: A visual system in vestibular coordinates. Neurosci Abstr 4:645
Simpson JI, Soodak RE, Hess R (1979) The accessory optic system and its relation to the vestibulocerebellum. In: Granit R, Pompeiano O (eds) Reflex control of posture and movement. Elsevier/North Holland, Amsterdam, pp 715-724
Takeda T, Maekawa K (1976) The origin of the pretecto-olivary tract. A study using the horseradish peroxidase method. Brain Res 117:319-325
Takeda T, Maekawa K (1980) Bilateral visual inputs to the dorsal cap of inferior olive: differential localization and inhibitory interactions. Exp Brain Res 39:461-471
Terasawa K, Otani K, Yamada J (1979) Descending pathways of the nucleus of the optic tract in the rat. Brain Res 173: 405-417
Updyke BV (1977) Topographic organization of the projections from cortical areas 17, 18, and 19 onto the thalamus, pretectum and superior colliculus in the cat. J Comp Neurol 173:81-122
Wallenberg A (1903) Notiz zur Anatomie des Tractus pedun- cularis transversus beim Meerschweinchen. Anat Anz 24: 119-200
Walley RE (1967) Receptive fields in the accessory optic system of the rabbit. Exp Neurol 17:27-43
Accepted May 18, 1982
Copyright © 2022 FDOKUMEN




















