
Interhemispheric transfer of visual perceptual learning in callosal agenesis
Upload
independentCategory
view
3download
0
BRAIN AND COGNITION 7, 39-53 (1988)
Processing from LVF, RVF and BILATERAL Presentations: Examinations of Metacontrol and Interhemispheric
Interaction
JOSEPH B. HELLIGE, JON E. JONSSON, AND CHIKASHI MICHIMATA
Observers indicated whether two successively presented drawings of facts were identical or differed in one feature (hair. eyes. mouth, jaw). The first face of each pair was presented at the fixation point and the second was presented to the left visual field-right hemisphere (LVF-RH). right visual field-left hemisphere (RVF-LH). or to both visual fields simultaneously (BILATERAL). On DIF- FERENT trials the RT of correct responses depended on which feature differed and the pattern of feature location effects was significantly different on LVF- RH and RVF-LH trials. On BILATERAL trials the feature location effect was identical to that obtained on RVF-LH trials and significantly different from that obtained on LVF-RH trials. In addition. the percentage of errors and RT of correct responses were both higher on BILATERAL trials than on unilateral trials. Implications of these results are considered for the concept of “metacontrol” in neurologically normal humans and for models of interhemispheric interaction. G 19% Academic Prc\\. Inc.
While a great deal of research has been devoted to the study of in- formation-processing differences between the left and right cerebral hem- ispheres, much less has been devoted to the manner in which the two hemispheres interact to produce integrated behavioral responses. As hu- mans look around their world, hear sounds, etc., most information is presented to both hemispheres and it is important to discover the range of interhemispheric interactions in such situations. With this in mind, the present experiment was designed to shed light on the nature of interhemispheric interaction.
The research reported here was supported in part by National Science Foundation Grant BNS-8608893 to Joseph B. Hellige. The authors are grateful to Justine Sergent for providing copies of the facial features used in her earlier studies, to Michael Bloch and Annette Kujawski for discussion of the results, and to Jerre Levy for comments on an earlier draft. Reprint requests should be addressed to Joseph B. Hellige. Department of Psychology SGM 502. University of Southern California, Los Angeles, CA 90089-1061.
39 0278-2626188 $3.00
Cownaht @ IY88 by Academic Press. Inc.

40 HELLIGE, JON&SON, AND MICHIMATA
Of particular interest for the present experiment is a concept referred to by Levy and Trevarthen (1976) as “metacontrol.” They define me- tacontrol as the neural mechanisms that determine which hemisphere will attempt to control cognitive operations. From their investigations of four commissurotomy patients, they conclude that when both hemi- spheres are presented with information to process, one hemisphere fre- quently asserts control. While the controlling hemisphere was often the one most effective for the task being performed, this was not always the case. That is, there was some dissociation of relative hemispheric com- petence and metacontrol. Furthermore, the pattern of metacontrol was not the same for all four subjects (see also Levy, 1985).
A major problem in extending the investigation of metacontrol to normal subjects is to determine which hemisphere in some sense “dominates” the processing when both have access to relevant stimulus input. One way to deal with this problem is to use tachistoscopic presentation tasks for which both cerebral hemispheres have some competence, but for which processing differs qualitatively depending on which visual field (hemisphere) is stimulated. By including “bilateral” presentation of the same stimulus simultaneously to both visual fields, it is possible to compare qualitative aspects of processing when both hemispheres are stimulated with the processing found on each of the unilateral trials. In this way, information is obtained about which of the two bilateral stimuli (LVF or RVF) dominates performance and, consequently, information is obtained about how the hemispheres act either together or independently to produce a correct response. The operational definition of a qualitative difference in processing is that there is an interaction between visual field (hemisphere) and some manipulated task variable, that is, what Hellige (1983) has referred to as a Hemisphere by Task interaction and what Zaidel (1983) has termed a processing dissociation.
The task used in the present experiment was based on findings reported by Sergent (1982). She required subjects to indicate whether a probe face presented to the left (LVF-RH) or right (RVF-LH) visual field was identical to a target face presented previously in the center of the viewing screen. The two faces were either identical or differed in only one or two features. The relevant features were hair, eyes, mouth, and jaw. The most important trials are those on which the two faces differ by only one feature. Sergent reported a Visual Field by Location of Different Feature interaction with RT as the dependent variable. On RVF-LH trials RT was significantly faster if the feature difference occurred in the upper rather than lower part of the face, with this pattern reversed on LVF-RH trials. Such results suggest that both hemispheres have some competence to perform this face comparison task, but do so in qualitatively different ways (see also Sergent, 1984).
The present experiment used the stimuli and comparison task employed

LVF, RVF, AND BILATERAL PRESENTATION 41
by Sergent (1982), with the addition of trials on which the probe face was presented simultaneously to both the LVF and RVF locations (referred to hereafter as BILATERAL presentation). On the unilateral trials in this experiement, one would hope to find a Visual Field by Location of Different Feature interaction similar to that reported by Sergent (1982) for RT of correct responses. The interesting question would then be what RT pattern emerges on the BILATERAL trials. There are many possibilities and it is instructive to consider a few here. Suppose that the two hem- ispheres operate in the normal brain as two completely independent processing systems, proceding in parallel with no funtional crosstalk on BILATERAL trials. In this case, the RT for each feature-location condition using BILATERAL presentation should be equal to the faster of the two unilateral RTs (with the error rate equal to the error rate associated with the faster visual field). That is, whichever hemisphere reaches a decision first would generate the response first on BILATERAL trials. As one alternative, suppose that one hemisphere consistently asserts metacontrol over this task. In this case the feature-location pattern on BILATERAL trials should be identical to the pattern obtained from the controlling hemisphere, for both response accuracy and RT.
METHODS Subjects. Five male and 10 female volunteers from introductory psychology courses at
the University of Southern California served as subjects. All subjects were right-handed native speakers of English with normal or corrected-to-normal vision in both eyes.’
Apparatus. Each subject sat at a table facing a 44 by 48cm screen approximately 60 cm away. A black posterboard covered the screen. with a rectangular window cut out approximately at eye level and centered in front of the subject. The window measured 6.5 cm vertically and 12.5 cm horizontally. A chinrest apparatus with forehead restraint bar was attached to the edge of the table, centered with respect to the viewing screen. Centered directly in front of the subject on the table was a 17 by 355cm response console. On top of the response console were two pairs of buttons, with the centermost button of each pair 7.5 cm from the center of the console. The two buttons within each pair were 3.5 cm apart. Appearing above each of the two innermost buttons was a small card with the same label (SAME or DIFFERENT) and above each of the two outermost buttons was a small card with the other label (DIFFERENT or SAME). Face stimuli and a fixation dot were rear-projected onto the viewing screen at the appropriate times by a Gerbrands three- channel projection tachistoscope (Model G 1176) equipped with three Kodak Carousel 850 slide projectors with Kodak Ektanar 12.8, 4 in lenses. Stimulus duration was controlled by a Gerbrands six-channel timer (Model 300-6T). Summary statistics were computed by an Apple II microprocessor.
Stimuhs materials. Stimuli for the present experiment were prepared from the drawings of features used by Sergent (1982). The I6 faces were prepared by using all combinations of the four binary-valued features, with the features being hair, eyes, mouth, and jaw (the nose was identical for all faces). The two versions of each feature and a sample face are
’ An additional I4 males and 8 females were run through all or part of the experiment, but were unable to detect certain of the feature differences at all. Thus, they would have had 0% correct and missing RT for one or more trial types.

42 HELLIGE, JONSSON, AND MICHIMATA
HAIR
JAW
1
FIG. I. Features used to construct the stimulus set and a sample face. Sergent (1982).
shown in Figure I. The 16 faces were photographed as high contrast negatives using Kodak Kodalith Ortho film so that they appeared on the viewing screen as white drawings on an opaque black background. When projected, each face spanned approximately 3.2” of visual angle horizontally and 4.3” of visual angle vertically. Faces presented centrally were centered around the fixation dot directly in front of the subject. Faces presented to the LVF or RVF were projected with the edge of the face nearest the center of the screen approximately 2.1” of visual angle from the center. The average luminance of each face stimulus was approximately 5.6 cd/m’. A fixation dot presented to the center of the viewing screen at appropriate times spanned approximately 0.2” of visual angle and had a luminance of approximately 4.0 cd/m’.
Each of the faces was photographed for presentation to the center of the viewing screen, for presentation to the LVF, RVF and for presentation to both LVF and RVF locations simultaneously. These displays were used to create a sequence of 192 trials consisting of the following trial types. Each trial consisted of two successively presented stimuli. The first stimulus was always 1 of the I6 faces presented in the center location and the second stimulus was I of the 16 faces presented to either the LVF alone (64 trials), the RVF alone (64 trials), or to both the LVF and the RVF simultaneously (BILATERAL trials, 64 trials). The trials within each visual field condition consisted of the following types. On 32 trials the second face was identical to the first (centered) face (SAME trials) and on 32 trials the second face differed from the first in one of the four features (DIFFERENT trials): hair (8 trials), eyes (8 trials), mouth (8 trials), and jaw (8 trials). The various trial types were ordered randomly with the constraints that there were no more than four successive occurrences of SAME or DIFFERENT stimuli or of the same visual field condition. Across all I92 trials each of the three visual field conditions consisted of exactly the same stimuli. The sequence of 192 trials was divided into successive sets of 48 trials each and the order of presentation of these four 48-trial sets was determined randomly for each subject.
Procedure. Each subject participated in three experimental sessions of approximately I hr each and separated by approximately 48 hr. At the beginning of the initial experimental session, subjects were told that their task would be to decide whether two successively presented faces were SAME or DIFFERENT. They were told that each trial would begin with presentation of a small fixation dot to the center of the viewing screen for 1 set and that the fixation dot would be followed immediately by a drawing of a face presented for I set to the center of the screen. They were told further that this centered face would be

LVF, RVF, AND BILATERAL PRESENTATION 43
followed immediately by a second face flashed briefly either to the LVF, RVF or to both visual fields simultaneously. Throughout the experiment it was emphasized repeatedly that on the BILATERAL trials the same face would always be presented to both visual fields. Subjects were told that their task would be to indicate as quickly and as accurately as possible whether the two successively presented faces were SAME or DIFFERENT, with each type of trial occurring about half the time. Once subjects understood the basic nature of the task, the importance of keeping their chin and forehead on the restraint apparatus was emphasized as was the importance of fixating their gaze on the fixation dot when it appeared and maintaining that eye position until after a response was made. Subjects were instructed to make their responses in the following manner. At the beginning of each trial block, subjects placed the index and middle fingers of the left and right hands on the two buttons within the left and right button-pairs, respectively-with the index fingers on the innermost buttons and the middle fingers on the outermost buttons. Half of the subjects were instructed to make SAME responses by pressing both of the inner buttons (index fingers) as quickly as possible and DIFFERENT responses by pressing both of the outer buttons (middle fingers) as quickly as possible. The remaining subjects had the opposite response-to-button mapping. Subjects were asked to respond as quickly and as accurately as possible.
After subjects demonstrated their understanding of the response procedure. the nature of the face stimuli was described completely and the subjects were given approximately 2 min to study a sheet containing the 16 possible faces. Subjects were told explicitly that on each trial the two successive faces would be either identical or differ on only one of the four variable features.
After studying the faces, subjects were given 15 practice trials to ensure that they understood the instructions to fixate and the response procedures. Following this. subjects received an additional block of 192 trials (the four 48-trial sets in a random order) which, unknown to the subjects, had been designated as additional practice and discarded. During a second and third experimental session subjects received two blocks of 192 experimental trials (two replications of the l92-trial sequence) each and it is the results from these latter two experimental sessions (768 trials) that are reported here. During each of these latter two sessions, subjects were given 2 min to restudy the faces prior to each block of 192 trials.
Each trial of both practice and experimental trials began with a I-set presentation of the fixation dot followed immediately by a I-set presentation of a centered face followed by a 250.msec presentation of a face in one of the three visual field conditions (LVF, RVF, BILATERAL). Within each set of 48 trials the intertrial interval was 5 set and between sets there was a pause of approximately 30 sec.
RESULTS
For each subject the percentage of errors and the median RT of correct responses were obtained for the 60 trial types defined by the following variables: four trial blocks, three visual field conditions (LVF, RVF and BILATERAL), and SAME/DIFFERENT stimulus type (with DIFFER- ENT trials further divided into the location of feature difference-hair, eyes, mouth, and jaw). Preliminary analyses of variance indicated no interactions of the within-subjects variables with sex of the subject and no interactions of the other variables with trial block. Therefore, the results presented here are collapsed across these two variables. An overall analysis of SAME and DIFFERENT trials (collapsing across the specific

44 HELLIGE, JONSSON, AND MICHIMATA
a
p BILATERAL
/
n=i5
2 970
‘1 - 960 w I + z 950 Cl F ” 2 940 n
SAME DIFFERENT STIMULUS TYPE
FIG. 2. Percentage of errors (a) and mean of median RT (b) for SAME and DIFFERENT trials. The parameters identify the three visual field conditions: LVF, left visual field: RVF, right visual field; BILATERAL, both visual fields.
feature difference) is presented first followed by examination of the effects of feature location on performance on DIFFERENT trials.
SAME/DIFFERENT comparison. Figure 2 shows the percentage of errors (Figure 2a) and mean of median RT (Figure 2b) for SAME and DIFFERENT trials. The parameter in each panel identifies the three visual field conditions (LVF, RVF and BILATERAL). For DIFFERENT trials the results are collapsed across the four feature locations which could differ. For both percentage of errors and RT, the results were subjected to an analysis of variance with SAME/DIFFERENT and Visual Field as within-subjects variables.
As suggested by Figure 2a, there was a significant main effect of visual field, F(2, 28) = 4.38, p < .05, and a significant SAME/DIFFERENT by Visual Field interaction, F(2, 28) = 6.58, p < .005. These effects occur because on SAME trials there were no differences among the visual field conditions, whereas on DIFFERENT trials errors increased slightly from RVF to LVF trials and increased more noticeably with BILATERAL presentation.
The high error rates in the present experiment make it necessary to be cautious in an evaluation of RT. Nevertheless, the RT of correct responses reinforces the finding that performance is worse in the BI- LATERAL condition than in the unilateral conditions. In the analysis of variance for RT of correct responses, the only effect to approach statistical significance was the main effect of visual field, F(2, 28) =

LVF, RVF, AND BILATERAL PRESENTATION 45
4.86, p < .025. As suggested by Figure 2b, RT was nonsignificantly longer on LVF trials than on RVF trials and significantly longer on BILATERAL trials than on unilateral trials.
Feature location effects. As noted in the introduction, it is of particular importance to examine performance on DIFFERENT trials as a function of the location of feature difference. Although in Sergent’s (1982) study an interaction between visual field and feature location was found only for RT of correct responses, the high error rates in the present experiment make it necessary to examine the feature location effects for both dependent variables. Accordingly, analyses of variance were conducted for each dependent variable with feature location and visual field as within-subjects variables. In order to be more precise about any feature location effects, the feature location variable was analyzed into linear, quadratic, and cubic trend components. The error rate results will be considered first, followed by the RT results.
Averaged across the three visual field conditions, the percentages of errors for the hair, eyes, mouth and jaw feature differences were 35.2, 26.0, 40.4, and 12.8, respectively. As these results suggest, there was a highly significant main effect of feature location, F(3, 42) = 8.38, p < .OOl, with each of the trend components showing a significant main effect: linear, F(1, 14) = 6.83, p < .025; quadratic, F(1, 14) = 6.51, p < .025; cubic, F(1, 14) = 10.62, p < .Ol. The linear trend reflects the finding that, in general, the percentage of errors is lower for features lower in the face as compared with features higher in the face, the quadratic trend reflects the tendency for the percentage of errors to be lower for the outer features (hair and jaw) than for the inner features (eyes and mouth), and the cubic trend reflects the fact that for the outer features there are fewer errors to the lower feature (jaw) than to the upper feature (hair) but for the inner features there are fewer errors to the upper feature (eyes) than to the lower feature (mouth).
There was also a significant main effect of visual field, F(2, 28) = 11.62, p < .OOl, with the RVF showing the least (26.4%) and the BI- LATERAL condition showing the greatest (34.1%) percentage of errors, with the LVF in between (28.1%). The feature location effect was the same for all three visual field conditions as reflected in a nonsignificant Feature Location by Visual Field interaction, F < 1.0.
In contrast to her response accuracy results, Sergent (1982, Experiment 1) did find a Feature Location by Visual Field interaction for RT of correct responses. It is instructive to consider the effects on RT of feature location and an interaction with visual field that is only partly similar to that reported by Sergent.
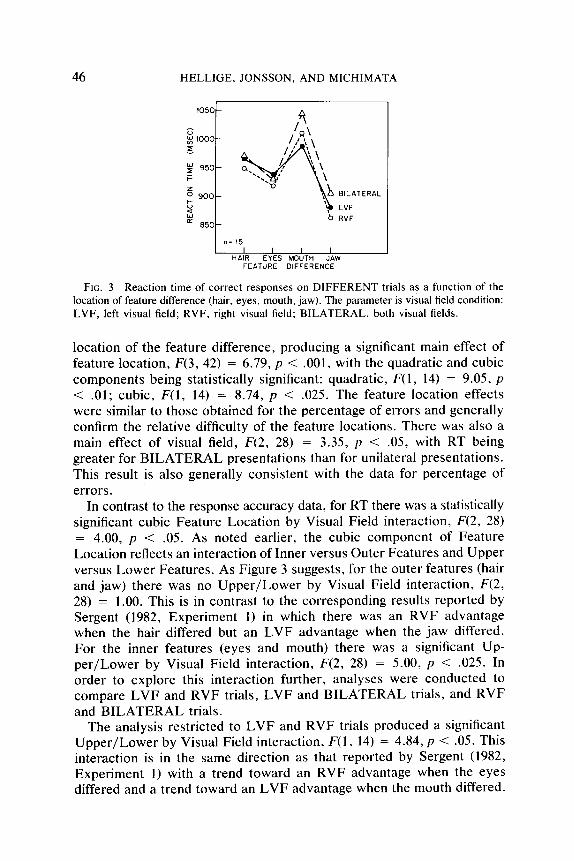
Figure 3 shows the RT of correct rsponses on DIFFERENT trials as a function of the location of feature difference. The parameter is visual field condition. As inspection of Figure 3 suggests, RT varied with the
46 HELLIGE, JONSSON, AND MICHIMATA
n= 15 I I I I
HAIR EYES MOUTH JAW FEATURE DIFFERENCE
FIG. 3 Reaction time of correct responses on DIFFERENT trials as a function of the location of feature difference (hair, eyes, mouth, jaw). The parameter is visual field condition: LVF, left visual field; RVF, right visual field; BILATERAL, both visual fields.
location of the feature difference, producing a significant main effect of feature location, F(3, 42) = 6.79, p < .OOl, with the quadratic and cubic components being statistically significant: quadratic, F(1, 14) = 9.05, p < .Ol; cubic, F(1, 14) = 8.74, p < .025. The feature location effects were similar to those obtained for the percentage of errors and generally confirm the relative difficulty of the feature locations. There was also a main effect of visual field, F(2, 28) = 3.35, p < .05, with RT being greater for BILATERAL presentations than for unilateral presentations. This result is also generally consistent with the data for percentage of errors.
In contrast to the response accuracy data, for RT there was a statistically significant cubic Feature Location by Visual Field interaction, F(2, 28) = 4.00, p < .05. As noted earlier, the cubic component of Feature Location reflects an interaction of Inner versus Outer Features and Upper versus Lower Features. As Figure 3 suggests, for the outer features (hair and jaw) there was no Upper/Lower by Visual Field interaction, F(2, 28) = 1.00. This is in contrast to the corresponding results reported by Sergent (1982, Experiment 1) in which there was an RVF advantage when the hair differed but an LVF advantage when the jaw differed. For the inner features (eyes and mouth) there was a significant Up- per/Lower by Visual Field interaction, F(2, 28) = 5.00, p < .025. In order to explore this interaction further, analyses were conducted to compare LVF and RVF trials, LVF and BILATERAL trials, and RVF and BILATERAL trials.
The analysis restricted to LVF and RVF trials produced a significant Upper/Lower by Visual Field interaction, F(1, 14) = 4.84, p < .05. This interaction is in the same direction as that reported by Sergent (1982, Experiment 1) with a trend toward an RVF advantage when the eyes differed and a trend toward an LVF advantage when the mouth differed.

LVF, RVF, AND BILATERAL PRESENTATION 47
Although the interaction appears less dramatic than that reported by Sergent, 11 of 15 subjects showed an interaction pattern in the direction reported by Sergent. Given that for inner features the feature location effect differs on LVF and RVF trials, it is instructive to examine whether the pattern of feature location effects on BILATERAL trials is more similar to one unilateral condition or the other.
Examination of Figure 3 (especially for the inner features) suggests that the pattern on BILATERAL trials is virtually identical to the pattern on RVF trials, but is increased by a constant amount, whereas the BILATERAL pattern is different from the LVF pattern. Specifically, the increase in RT from the eyes-different to the mouth-different condition is smaller for LVF presentations than for either RVF or BILATERAL presentations. These impressions are corroborated by analyses of variance comparing each unilateral field with the BILATERAL condition. Both analyses confirmed the Upper/Lower feature main effect for inner features. In the analysis comparing LVF and BILATERAL conditions there was a significant Upper/Lower Feature Location by Visual Field interaction, F(1, 14) = 9.50, p < .Ol, indicating that the pattern of feature location effects is not the same on BILATERAL and LVF trials. In contrast, in the analysis comparing RVF and BILATERAL conditions, an RVF ad- vantage averaging about 24 msec was constant across feature locations so that there was no Upper/Lower Feature Location by Visual Field interaction, F(1, 14) = 1.50.’
Anulysis of individual subjects. In order to examine the consistency of the pattern of results just described, the data from individual subjects were examined to determine whether the pattern of feature location effects obtained on BILATERAL trials were more similar to the pattern obtained on LVF-RH trials (LVF-RH model), the pattern obtained on RVF-LH trials (RVF-LH model), or the pattern predicted by the average of LVF-RH and RVF-LH trials (averaging model). If there were no tendency for one hemisphere or the other to exert metacontrol over the present task, one would expect the LVF-RH and RVF-LH models to provide the relatively best description of the data for about the same number of subjects or to find the averaging model providing a better fit than the other two because each prediction is based on two unilateral scores rather than one.3
’ Similar analyses using all four feature locations produced the same pattern of results. In the comparison of LVF and BILATERAL conditions there was a significant Feature Location by Visual Field interaction, F(3, 42) = 2.86, p < .05, with the interaction being significant for both the linear and the cubic components of feature location: linear, F( I, 14) = 4.62, p < .05; cubic, F(1, 14) = 6.44, p i ,025. In the comparison of RVF and BILATERAL conditions, there was no Feature Location by Visual Field interaction, F(3, 42) = 0.81, and no trend toward an interaction for linear, quadratic, or cubic components of feature location.
3 Specifically, the models used for both RT and percentage of errors are (I) BILATERAL

48 HELLIGE, JONSSON, AND MICHIMATA
For RT of correct responses, the RVF-LH model provides the best fit to the data from 10 of 15 subjects, the LVF-RH model provides the best fit to the data from 2 of 15 subjects, and the averaging model provides the best fit to the data from 3 of 15 subjects. Thus, the individual subject data suggest that on BILATERAL trials the pattern of feature location effects for RT is more similar to the RVF-LH pattern than to either the LVF-RH pattern or the average of LVF-RH and RVF-LH patterns for most subjects, consistent with the group data. For percentage of errors, the RVF-LH model provides the best fit to the data from 9 of 15 subjects, while each of the other two models provide the best fit to the data from 3 of 15 subjects, a pattern of results similar to that found for RT of correct responses. The RVF-LH model provides the best fit to the BI- LATERAL data for both percentage of errors and RT for 7 subjects, while the LVF-RH model provides the best fit for both dependent variables for only one subject.
DISCUSSION
For convenience of exposition, several aspects of the present results will be discussed separately. We will consider, in order, the results from the unilateral trials of the present experiment in comparison to results reported by Sergent (1982, 1984), the results from the BILATERAL trials compared with the unilateral trials, and possible reasons for individual differences in the pattern of effects on BILATERAL trials.
Unilateral trials and Set-gent’s results. For reasons that are unclear, the present experiment was more difficult for our subjects than Sergent’s
score = LVF-RH score + Kl, (2) BILATERAL score = RVF-LH score + K2, and (3) BILATERAL score = .S(LVF-RH score + RVF-LH score) + K3. The values of Kl, K2, and K3 equal the scores on BILATERAL trials (averaged across all four feature locations) minus the scores on LVF-RH trials, RVF-LH trials, and the average of LVF- RH and RVF-LH trials, respectively. Thus, the K values are scaling constants for each subject that allow feature location patterns to be predicted regardless of any constant difference between BILATERAL and unilateral conditions. The models were used to generate a predicted BILATERAL score for each of the four feature locations for each subject using the LVF-RH and/or RVF-LH scores for the corresponding feature location and the constant (K), which was the same for all four feature locations. For each model, the difference was computed between each of the four predicted values and the corresponding obtained BILATERAL values. These differences were then squared and summed and this sum of squared deviations was used to assess the relative goodness of fit of each model to the data from each subject. In order to determine whether this family of models provides a reasonable description of the group data for BILATERAL trials, the predictions of the best-fitting model for each subject were used to compute a predicted group average for each of the four feature locations. For percentage of errors the predicted (and obtained) values were as follows for the hair, eyes, mouth, and jaw conditions, respectively: 41.1 (39.2), 31.5 (31.0), 46.6 (47.8), and 17.6 (17.5). For RT of correct responses the corresponding predicted (and obtained) values in milliseconds were 985 (974), 944 (932), 1027 (1040), and 899 (905).

LVF, RVF, AND BILATERAL PRESENTATION 49
(1982) experiments were for her subjects. Despite this difference in overall performance level, there are important similarites in the pattern of results. For example, in neither case was there an overall advantage for either the LVF-RH or the RVF-LH, suggesting that both hemispheres have about equal competence for the present task. Furthermore, in both cases there was a LVF-RH/RVF-LH by Feature Location interaction for RT of correct responses, suggesting some qualitative differences in the pro- cessing of these stimuli from the two visual fields.
Despite these similarities, the specific pattern of feature location effects was not the same in the present experiment as in Sergent’s (1982) ex- periment. Sergent (1984) used multidimensional scaling techniques to determine that stimuli from this pool are processed in terms of their gestalt (rather than analytically, feature by feature) regardless of which visual field is stimulated. On this view, location of the DIFFERENT feature influences RT because the features do not contribute equally to the gestalt and interaction of Visual Field and Feature Location suggests that the relative contribution of the features to the gestalt is not the same for the two cerebral hemispheres (see Sergent, 1984, for discussion). With this in mind, it is instructive to consider why the relative contributions of the various features might be different in the present experiment than in Sergent’s (1982) experiments.
For both outer (hair and jaw) and inner (eyes and mouth) features, Sergent (1982) found faster RTs to the upper than to the lower feature on RVF-LH trials and exactly the opposite on LVF-RH trials. In contrast, as Figure 3 shows, for outer features we found faster RTs to the jaw than to the hair for both visual fields, with there being no LVF-RH versus RVF-LH by Feature Location interaction. For the inner features we found faster RT to the eyes than to the mouth for both visual fields, although in this case the effect was significantly smaller on LVF-RH trials than on RVF-LH trials, an interaction that is in the direction shown by Sergent. In Sergent’s experiments, the spatial location of the face on LVF-RH and RVF-LH trials overlapped somewhat the spatial location of the centrally presented face. Although central and lateralized faces were never presented simultaneously, they were immediately successive as in the present experiment. Thus, there may have been some forward and backward masking that influenced the relative contribution of the various features to the gestalt. Because the faces are not left-right sym- metrical, such masking could even contribute to the form of the Visual Field byeFeature Location interaction, especially for the outer features for which the overlap would be greater. In the present experiment, stimuli were located so that the central and lateralized stimuli would not overlap at all in their spatial locations so that any effects of forward and backward masking would be eliminated. While we can only speculate that this difference in the stimulus arrangements caused a difference in the feature

50 HELLIGE, JONSSON. AND MICHIMATA
location patterns, it seems the most reasonable possibility in view of what Sergent (1984) has learned about the processing of these stimuli.
Bilateral versus unilateral presentation. Two aspects of the results on BILATERAL trials will be discussed in order. The first is that BILATERAL presentation of the stimulus material led to worse performance than unilateral presentation. The second is that the pattern of feature location effects on DIFFERENT trials is identical to the pattern obtained on RVF-LH trials but different from the pattern obtained on LVF-RH trials.
As noted in the introduction, some plausible models of how the two hemispheres might function on BILATERAL trials predict that the RT on BILATERAL trials should always equal the faster of the two unilateral RTs. While this may prove true for other tasks, it was clearly not the case for the present task. Instead, for both percentage of errors and RT of correct responses performance was worse on BILATERAL trials than on unilateral trials. The only exception to this was for percentage of errors on SAME trials (see Figure 2a) where there were no visual field differences, but this may well be caused by a bias for subjects to respond “same” when they were not sufficiently certain that they had detected a difference.
In considering possible reasons for the difficulty produced by BILAT- ERAL presentation, it is useful to keep in mind that BILATERAL pres- entation occurred on only one-third of the trials and was unpredictable. It is possible that the relative infrequency of BILATERAL (compared with unilateral) trials resulted in a delay in stimulus processing which might increase both RT and error rates. One way to test this notion would be to block the presentation of BILATERAL trials, thereby making such presentation more predictable.
Another possibility is that each hemisphere began processing the stimulus with which it was stimulated and at some point in the information processing sequence the two hemispheres compared the results of their processing to that point (cf. Liederman, Merola, & Martinez, 1985). Such a comparison might be particularly encouraged by the requirement to make a bilateral response on each trial. Such a comparison would occur only on BI- LATERAL trials, would presumably take time, and would occur on both SAME and DIFFERENT trials. Furthermore, on DIFFERENT trials the comparison time would be independent of which feature was different. If it were the case that subjects most often continued processing only the RVF-LH stimulus after the comparison, then this notion is quite consistent with the finding that the RT on BILATERAL trials is well described as being equal to the RT on RVF-LH trials plus a constant of approximately 24 msec for both SAME and DIFFERENT trials and for all locations of the different feature.
It is also possible that the increase in RT and error rate on BILATERAL trials is related to attentional factors. Based on observations of a split-

LVF, RVF, AND BILATERAL PRESENTATION 51
brain patient, Holtzman and Gazzaniga (1982) concluded that the two cerebral hemispheres have access to a common pool of processing resources that is distributed between the two hemispheres when the subject is faced with demanding bilateral stimulation. There is no doubt that when the stimulus information is presented bilaterally, at least some processing of both stimuli is obligatory. If some of the obligatory processing is resource demanding, then on BILATERAL trials the resource pool must be appropriately divided between the two stimuli. In contrast, on unilateral trials all of the resources can be allocated to the single stimulus. Perhaps it is the need to divide resources between the hemispheres that causes the performance decrement on BILATERAL trials. If such attentional factors are operating prior to the SAME/DIFFERENT decision, then such an explanation is consistent with a performance decrement on BI- LATERAL trials that is constant across trial types.
The attentional factors just considered are consistent with physiological evidence, indicating that the increase in arousal in the hemisphere con- tralateral to a stimulus is typically accompanied by an inhibition of arousal in the opposite hemisphere, mediated at least in part by reciprocal inhibitory connections through the brainstem commissures. For example, animal studies suggest that unilateral stimulation not only produces facilitation of dopamine release in the contralateral substantia nigra (and ipsilateral caudate nucleus), but also inhibition of dopamine release in the ipsilateral substantia nigra (and contralateral caudate nucleus) (Leviel, Chesselet, Glowinski, & Cheramy, 1981). Thus, under conditions of bilateral stim- ulation, the contralateral arousal produced by a lateralized stimulus would be partially offset by the ipsilateral inhibition produced by the other lateralized stimulus. As a result, neither hemisphere may be as aroused as when it was presented with a unilateral stimulus.
Whatever the reason for the performance decrement on BILATERAL trials in the present experiment, it is unlikely that such a decrement will characterize all instances of BILATERAL presentation of the same stimulus to both visual fields. For example, in the only other study of hemispheric asymmetry we know of that presented the same stimuli simultaneously to both LVF and RVF locations, Jones (1982) found better performance on BILATERAL than on unilateral trials, even when unilateral and bilateral trials were randomly mixed. His task required subjects to name aloud the letter presented tachistoscopically, a task for which the left hemisphere must be involved to generate the overt verbal response. Perhaps per- formance increments are most likely for very easy tasks or for tasks whose response components are strongly lateralized, reducing the potential for confusability caused by having each hemisphere generate its own response. Performance decrements may be more likely in situations like the present one where the task is difficult and both hemispheres have competence to analyze the stimuli and generate the response.

52 HELLIGE, JONSSON, AND MICHIMATA
For reasons reviewed earlier, special importance is attached to a com- parison of the feature location effect on BILATERAL trials with the feature location effects on RVF-LH and LVF-RH trials. For RT of correct responses, where the feature location effects are reliably different on RVF-LH and LVF-RH trials, it is the case that the feature location effect on BILATERAL trials is identical to the feature location effect on RVF-LH trials and significantly different from the feature location effect on LVF-RH trials. Thus, for most subjects in the present task the RVF-LH stimulus dominates performance on BILATERAL trials. While important questions remain about exactly where this domination occurs in the sequence of processing stages, absence of a reliable RVF-LH advantage on unilateral trials rules out explanations in terms of such overt attentional factors as a bias in eye fixation toward the RVF.
The RVF-LH stimulus dominates performance on BILATERAL trials even when it is not advantageous for it to do so. For example, note from Figure 3 that the increase in RT when the feature difference is moved from the eyes to the mouth is significantly larger for RVF-LH trials than for LVF-RH trials. The corresponding increase on BILATERAL trials matches the increase on RVF-LH trials, even though on BILATERAL trials the LVF-RH stimulus is present and could potentially lead to more efficient responding. That is, there is some dissociation of the relative efficiency of LVF-RH and RVF-LH performance and the visual field to which subjects are most attentive when given a choice. This is reminiscent of the dissociation found by Levy and Trevarthen (1976; Levy, 1985) between relative hemispheric competence and metacontrol for split-brain patients. Such results suggest that the concept of metacontrol extends to the intact brain and that the use of BILATERAL presentation of the same stimulus may provide a useful way to study the direction and extent of metacontrol for a variety of tasks and subject populations and to separate metacontrol from hemispheric specialization.
Individual differences. In their investigation of split-brain patients, Levy and Trevarthen (1976) reported individual differences among their four patients in metacontrol for some tasks. An examination of the data from individual subjects in the present experiment provides at least a preliminary indication that the same is true of neurologically normal subjects. For both percentage of errors and RT of correct responses the pattern of feature location effects on BILATERAL trials was more similar to the pattern obtained on RVF-LH than on LVF-RH trials or on the average of LVF-RH and RVF-LH trials for most subjects. For RT this is hardly surprising in view of the group data shown in Figure 3 and one would hope that the error data not depart too much from the RT data. However, the analysis of individual subjects suggests that subjects are not uniform in the extent to how accurately BILATERAL performance is predicted by performance on one or the other visual field and it would

LVF, RVF, AND BILATERAL PRESENTATION 53
seem worthwhile to explore such individual differences in further studies. Such studies should examine the relationship between BILATERAL and unilateral performance across a variety of tasks for the same subjects or examine the relationship between individual differences such as those reported here and individual differences in other tasks thought to reveal patterns of metacontrol or arousal asymmetries between the left and the right hemispheres. In this way, stable individual differences in metacontrol can be separated from task-specific differences and from differences caused by random error.
REFERENCES
Bradshaw. J. L., & Nettleton. N. C. 1983. Hlr,nrr,t cer-ebrrrl asymmetry. New York: Academic Press.
Hellige. J. B. (1983) Hemisphere x task interaction and the study of laterality. In J. B. Hellige (Ed.), Cerebra/ hemisphere rrsymmefry: Method, rheory. and rrpplicution. New York: Praeger. Pp. 41 l-443.
Hellige, J. B.. & Sergent, J. (1986). Role of task factors in visual field asymmetries. Bruin and Cognition. 5, 200-222.
Holtzman. J. D., & Gazzaniga, M. S. (1982) Dual task interactions due exclusively to limits in processing resources. Science, 218, 1325-1327.
Jones, B. (1982) The integrative action of the cerebral hemispheres. Perception & Psy- chophysics, 32, 423-433.
Leviel, V., Chesselet, M. F., Glowinski, J., & Cheramy, A. (1981) Involvement of the thalamus in the asymmetric effects of unilateral sensory stimuli on the two nigrostriatal dopaminergic pathways of the cat. Brain Research, 223, 257-272.
Levy, J. (1985) Interhemispheric collaboration: Single-mindedness in the asymmetric brain. In C. T. Best (Ed.). Hemi.splleric,frrnction und collaborution in the child. New York: Academic Press. Pp. I l-32.
Levy, J., & Trevarthen. C. (1976) Metacontrol of hemispheric function in human split- brain patients. Jortrnul qfExperimenfu1 P.s~cholo~y: Human Perception and Perf<ormunc,e, 2, 299-3 12.
Liederman, J., Merola, J., & Martinez, S. (1985) Interhemispheric collaboration in response to simultaneous bilateral input. Neuropsychologiu. 23, 673-684.
Sergent, J. (1982) About face: Left-hemisphere involvement in processing physiognomies. Journal of Experimental Psychology: Human Perception und Performunce, 8, 253- 272.
Sergent. J. (1984) Conhgural processing of faces in the left and right cerebral hemispheres. Journul of Experimental Psychology: Humatz Perception and Perfkmrrnce, 10, 556 572.
Zaidel, E. (1983) Disconnection syndrome as a model for laterality effects in the normal brain. In J. B. Hellige (Ed.), Cerebral hemisphere asymmetry: Method, theory, and application. New York: Praeger. Pp. 95-151.
Copyright © 2022 FDOKUMEN




















