
Primary responses of root and leaf elongation to water deficits ...
15
Journal of Experimental Botany Journal of Experimental Botany, Vol. 48, No. 310, pp. 985-999, May 1997 OPINION PAPER Primary responses of root and leaf elongation to water deficits in the atmosphere and soil solution Jiirgen Frensch 1 Lehrstuhl fOr Pflanzendkologie, Universita't Bayreuth, D-95440 Bayreuth, Germany Received 5 August 1996; Accepted 16 January 1997 Abstract Plants experience drought by a limitation of water supply and by enhanced transpiration. Both processes tend to decrease the plant's water potential, but affect growth responses in the root and leaf differently. The evaluation of the underlying mechanisms leads to a discussion of recent studies on biophysical aspects of cell expansion at a cellular, tissue and organ level. Two processes enable roots to compensate rapidly effects of water deficits originating in the medium: (i) adjustment of the minimum pressure in cells required for expansion (yield threshold), and (ii) solute transport within the elongation zone. Limitations of root growth are discussed with respect to hydraulic, mechanical, and solute relations in the root elongation zone. It is argued that the variable nature of both the yield threshold and solute transport challenges the applic- ability of the Lockhart concept to determine growth- related parameters from steady conditions of turgor and growth. On a whole organ level, the attenuation of xylem pressure along the root is important for the differential response of root and leaf growth. Experimental evidence is presented for the hydraulic separation of the elongation zones, which is closely related to root development and functioning. The data obtained over the past few years have been used to extend mathematical models of growth and water transport in roots. Key words: Extension growth, hydraulic conductivity, root development (xylem, endodermis), transport (water and solute), turgor pressure, water stress, xylem pressure, Zea mays. Introduction At least during short periods of time, terrestrial plants are exposed to water limiting conditions in almost every natural environment. Cell expansion is among the most sensitive processes in plants affected by drought which is the result of an imbalance in the plant between water uptake and water loss (Hsiao, 1973; Boyer, 1985). Water loss to the atmosphere is largely controlled by stomatal movement. Stomata respond to drought in the atmos- phere and in the soil, and much information on the underlying mechanisms has been accumulated (Schulze, 1986; Tardieu and Davies, 1993; Slovik et al., 1995). By contrast, the complexity between root performance and low plant water potentials is much less understood. To maintain uptake of water and nutrients under water limiting conditions, continuous proliferation of roots into new soil layers is important and requires adaptive mechan- isms on the cell and tissue level (Tomos et al., 1989; Carpita and Gibeaut, 1993; Cosgrove, 1993). This review aims to evaluate limitations of root and leaf growth effected by drought in the atmosphere and in the soil. In particular, short-term responses of hydraulic conductivity, cell wall yielding, and solute transport in the root elongation zone are analysed. Furthermore, the significance of root development for the differential response of roots and shoots to drought stress is empha- sized and demonstrated. Mechanisms of long-distance volume flow are discussed with respect to new experi- mental results on pressure propagation in the xylem. Current mathematical models of cell expansion and water uptake are extended on the basis of experimental data. Differential response of the root and the leaf to water stress: Lockhart's model For many plants, root growth is more resistant to water stress than shoot growth (Hsiao and Jing, 1987; Kramer and Boyer, 1995). The different responses may be exempli- fied by a simple experiment. Root and leaf elongation were measured continuously on the same intact plant 1 Fax: +49 921 552564. E-mail: jurgen.frenschQunl-bayreuth.de C Oxford University Press 1997 Downloaded from https://academic.oup.com/jxb/article/48/5/985/542707 by guest on 10 January 2022
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Primary responses of root and leaf elongation to water deficits ...

Journal ofExperimentalBotany
Journal of Experimental Botany, Vol. 48, No. 310, pp. 985-999, May 1997
OPINION PAPER
Primary responses of root and leaf elongation to waterdeficits in the atmosphere and soil solution
Jiirgen Frensch1
Lehrstuhl fOr Pflanzendkologie, Universita't Bayreuth, D-95440 Bayreuth, Germany
Received 5 August 1996; Accepted 16 January 1997
Abstract
Plants experience drought by a limitation of watersupply and by enhanced transpiration. Both processestend to decrease the plant's water potential, but affectgrowth responses in the root and leaf differently. Theevaluation of the underlying mechanisms leads to adiscussion of recent studies on biophysical aspects ofcell expansion at a cellular, tissue and organ level.Two processes enable roots to compensate rapidlyeffects of water deficits originating in the medium: (i)adjustment of the minimum pressure in cells requiredfor expansion (yield threshold), and (ii) solute transportwithin the elongation zone. Limitations of root growthare discussed with respect to hydraulic, mechanical,and solute relations in the root elongation zone. It isargued that the variable nature of both the yieldthreshold and solute transport challenges the applic-ability of the Lockhart concept to determine growth-related parameters from steady conditions of turgorand growth. On a whole organ level, the attenuationof xylem pressure along the root is important forthe differential response of root and leaf growth.Experimental evidence is presented for the hydraulicseparation of the elongation zones, which is closelyrelated to root development and functioning. The dataobtained over the past few years have been used toextend mathematical models of growth and watertransport in roots.
Key words: Extension growth, hydraulic conductivity, rootdevelopment (xylem, endodermis), transport (water andsolute), turgor pressure, water stress, xylem pressure,Zea mays.
Introduction
At least during short periods of time, terrestrial plantsare exposed to water limiting conditions in almost every
natural environment. Cell expansion is among the mostsensitive processes in plants affected by drought which isthe result of an imbalance in the plant between wateruptake and water loss (Hsiao, 1973; Boyer, 1985). Waterloss to the atmosphere is largely controlled by stomatalmovement. Stomata respond to drought in the atmos-phere and in the soil, and much information on theunderlying mechanisms has been accumulated (Schulze,1986; Tardieu and Davies, 1993; Slovik et al., 1995). Bycontrast, the complexity between root performance andlow plant water potentials is much less understood. Tomaintain uptake of water and nutrients under waterlimiting conditions, continuous proliferation of roots intonew soil layers is important and requires adaptive mechan-isms on the cell and tissue level (Tomos et al., 1989;Carpita and Gibeaut, 1993; Cosgrove, 1993).
This review aims to evaluate limitations of root andleaf growth effected by drought in the atmosphere and inthe soil. In particular, short-term responses of hydraulicconductivity, cell wall yielding, and solute transport inthe root elongation zone are analysed. Furthermore, thesignificance of root development for the differentialresponse of roots and shoots to drought stress is empha-sized and demonstrated. Mechanisms of long-distancevolume flow are discussed with respect to new experi-mental results on pressure propagation in the xylem.Current mathematical models of cell expansion and wateruptake are extended on the basis of experimental data.
Differential response of the root and the leaf towater stress: Lockhart's model
For many plants, root growth is more resistant to waterstress than shoot growth (Hsiao and Jing, 1987; Kramerand Boyer, 1995). The different responses may be exempli-fied by a simple experiment. Root and leaf elongationwere measured continuously on the same intact plant
1 Fax: +49 921 552564. E-mail: jurgen.frenschQunl-bayreuth.de
C Oxford University Press 1997
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/5/985/542707 by guest on 10 January 2022
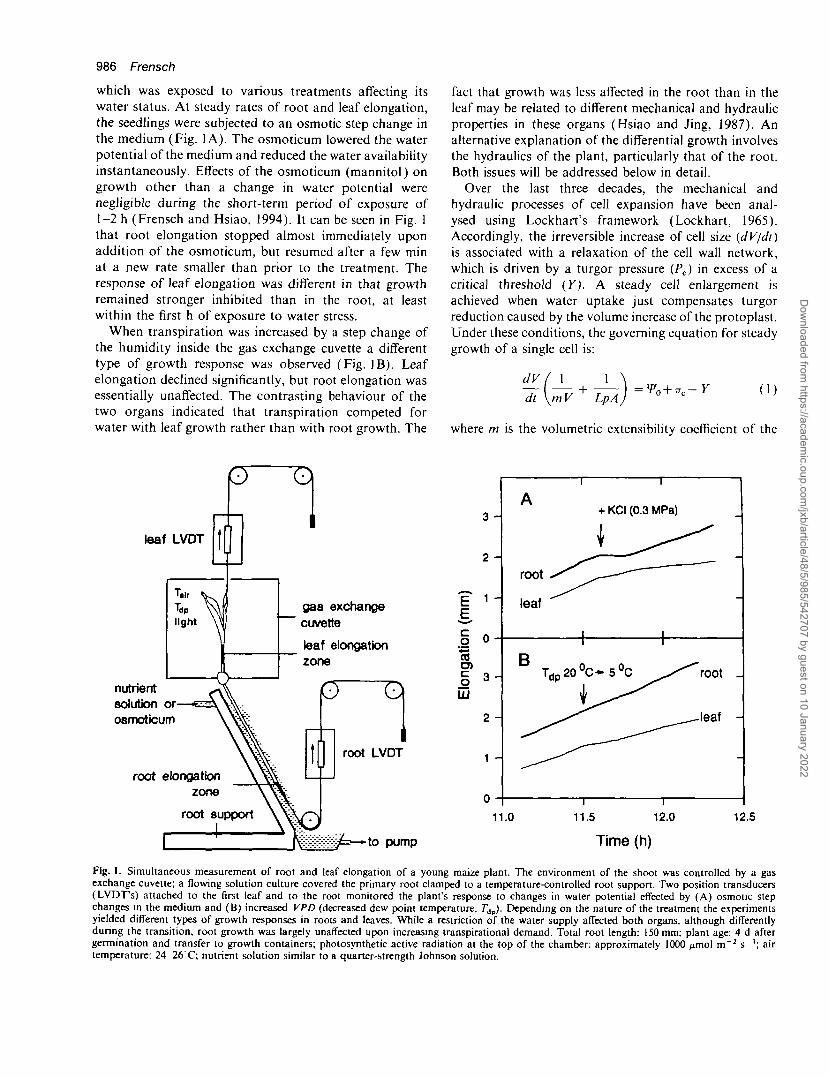
986 Frensch
which was exposed to various treatments affecting itswater status. At steady rates of root and leaf elongation,the seedlings were subjected to an osmotic step change inthe medium (Fig. 1A). The osmoticum lowered the waterpotential of the medium and reduced the water availabilityinstantaneously. Effects of the osmoticum (mannitol) ongrowth other than a change in water potential werenegligible during the short-term period of exposure of1-2 h (Frensch and Hsiao, 1994). It can be seen in Fig. 1that root elongation stopped almost immediately uponaddition of the osmoticum, but resumed after a few minat a new rate smaller than prior to the treatment. Theresponse of leaf elongation was different in that growthremained stronger inhibited than in the root, at leastwithin the first h of exposure to water stress.
When transpiration was increased by a step change ofthe humidity inside the gas exchange cuvette a differenttype of growth response was observed (Fig. IB). Leafelongation declined significantly, but root elongation wasessentially unaffected. The contrasting behaviour of thetwo organs indicated that transpiration competed forwater with leaf growth rather than with root growth. The
fact that growth was less affected in the root than in theleaf may be related to different mechanical and hydraulicproperties in these organs (Hsiao and Jing, 1987). Analternative explanation of the differential growth involvesthe hydraulics of the plant, particularly that of the root.Both issues will be addressed below in detail.
Over the last three decades, the mechanical andhydraulic processes of cell expansion have been anal-ysed using Lockhart's framework (Lockhart, 1965).Accordingly, the irreversible increase of cell size (dVjdt)is associated with a relaxation of the cell wall network,which is driven by a turgor pressure (Pc) in excess of acritical threshold (Y). A steady cell enlargement isachieved when water uptake just compensates turgorreduction caused by the volume increase of the protoplast.Under these conditions, the governing equation for steadygrowth of a single cell is:
( +dt \mV LpA
( i )
where m is the volumetric extensibility coefficient of the
leaf LVDT
nutrientsolution orosmoticum
root elongationzone
gas exchangecuvette
leaf elongationzone
root LVDT
root support
to pump
3 -
11.0 11.5 12.0 12.5
Time (h)
Fig. 1. Simultaneous measurement of root and leaf elongation of a young maize plant. Trie environment of the shoot was controlled by a gasexchange cuvette; a flowing solution culture covered the primary root clamped to a temperature-controlled root support. Two position transducers(LVDTs) attached to the first leaf and to the root monitored the plant's response to changes in water potential effected by (A) osmotic stepchanges in the medium and (B) increased VPD (decreased dew point temperature, 7"dp). Depending on the nature of the treatment the experimentsyielded different types of growth responses in roots and leaves. While a restriction of the water supply affected both organs, although differentlyduring the transition, root growth was largely unaffected upon increasing transpirational demand. Total root length: 150 mm; plant age: 4 d aftergermination and transfer to growth containers; photosynthetic active radiation at the top of the chamber: approximately 1000 ^unol m~2 s"1; airtemperature: 24 26 C; nutrient solution similar to a quarter-strength Johnson solution.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/5/985/542707 by guest on 10 January 2022
Roof elongation and drought 987
wall. Lp is the hydraulic conductivity of the cell, A andFare the surface area and volume of the cell, respectively,nc is the osmotic pressure of the cell sap, and y0 is thewater potential of the source. The two terms in bracketsrepresent mechanical and hydraulic 'growth resistances'which limit cell expansion according to their contributionto the overall resistance.
Provided that the mechanical resistance is much largerthan the hydraulic, Equation (1) reduces to
(2)dt \mV] P' Y
where (Pc-Y) denotes the 'growth-effective turgor'.Equation (2) rather than equation (1) is usually used toquantify cell expansion. When solute transport into thecell is not unlimited, however, the simplified equation canbe misleading. Because TTC is proportional to the ratio ofvolume and solute flows (Jy/J,), the criteria for thesimplification are not justified solely by showing that1/fmx V)»\/(LpxA) and, thus, Wos Wm (fm: mediumwater potential). Rather the relationship between -nc andJ, needs further consideration, which is largely ignoredin studies relating to cell and tissue growth. It is expectedthat a separation of solute and water flows would helpto elucidate the controversial debate on the significanceof hydraulic conductivities for growth.
Conceptually, the differential response of growth inroots and shoots to drought may be summarized in termsof different control loops, linking cell expansion to bio-physical and biochemical processes in growing tissues(Bradford and Hsiao, 1982). Turgor pressure (Pc) haslong been recognized a driving force for cell enlargement,although its meaning for growth control and adjustmentto water stress is debated. Doubts about Pc as a limitingfactor are based on results from experiments in the fieldand laboratory (Matsuda and Riazi, 1981; Michelena andBoyer, 1982; Shackel et al, 1987; Hsiao and Jing, 1987;Munns, 1988). However, growth may be sensitive tochanges in Pc only where expansion is limited or co-limitedby low turgor. This hypothesis was recently investigatedby exposing maize plants to changes in water potentialseffected in the root medium (osmotic and hydrostaticpressure steps) and in the atmosphere (humidity steps)(Frensch, Hsiao, Rochas-Lara, unpublished results).
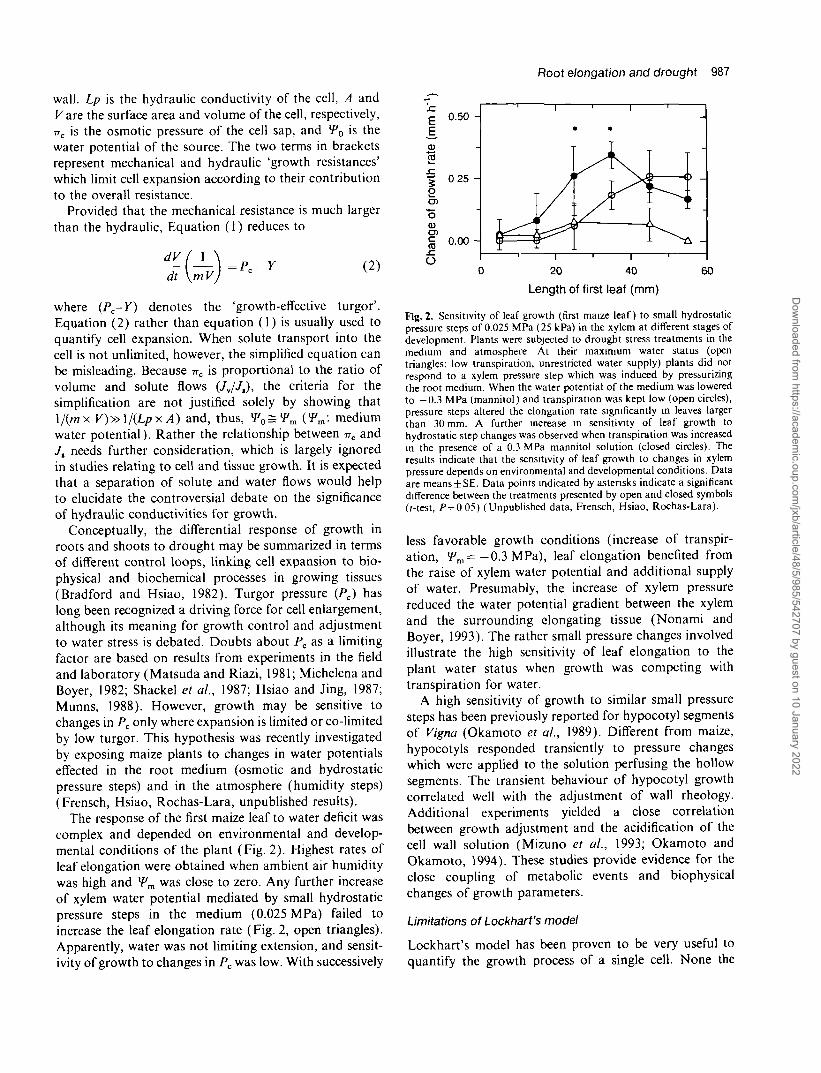
The response of the first maize leaf to water deficit wascomplex and depended on environmental and develop-mental conditions of the plant (Fig. 2). Highest rates ofleaf elongation were obtained when ambient air humiditywas high and Wm was close to zero. Any further increaseof xylem water potential mediated by small hydrostaticpressure steps in the medium (0.025 MPa) failed toincrease the leaf elongation rate (Fig. 2, open triangles).Apparently, water was not limiting extension, and sensit-ivity of growth to changes in Pc was low. With successively
0.50 -
0 25 -
0 .00-
1 1
T /
1 1
1 1 '-
> *
• 1 •
CD
2
g
~oCDO)CO
60 20 40 60
Length of first leaf (mm)
Fig. 2. Sensitivity of leaf growth (first maize leaf) to small hydrostaticpressure steps of 0.025 MPa (25 kPa) in the xylem at different stages ofdevelopment. Plants were subjected to drought stress treatments in themedium and atmosphere At their maximum water status (opentriangles; low transpiration, unrestricted water supply) plants did notrespond to a xylem pressure step which was induced by pressurizingthe root medium. When the water potential of the medium was loweredto - 0 . 3 MPa (mannitol) and transpiration was kept low (open circles),pressure steps altered the elongation rate significantly in leaves largerthan 30 mm. A further increase in sensitivity of leaf growth tohydrostatic step changes was observed when transpiration was increasedin the presence of a 0.3 MPa mannitol solution (closed circles). Theresults indicate that the sensitivity of leaf growth to changes in xylempressure depends on environmental and developmental conditions. Dataare means ±SE. Data points indicated by asterisks indicate a significantdifference between the treatments presented by open and closed symbols(f-test, /> = 0 05) (Unpublished data, Frensch, Hsiao, Rochas-Lara).
less favorable growth conditions (increase of transpir-ation, *Fm=-0.3MPa), leaf elongation benefited fromthe raise of xylem water potential and additional supplyof water. Presumably, the increase of xylem pressurereduced the water potential gradient between the xylemand the surrounding elongating tissue (Nonami andBoyer, 1993). The rather small pressure changes involvedillustrate the high sensitivity of leaf elongation to theplant water status when growth was competing withtranspiration for water.
A high sensitivity of growth to similar small pressuresteps has been previously reported for hypocotyl segmentsof Vigna (Okamoto et al, 1989). Different from maize,hypocotyls responded transiently to pressure changeswhich were applied to the solution perfusing the hollowsegments. The transient behaviour of hypocotyl growthcorrelated well with the adjustment of wall rheology.Additional experiments yielded a close correlationbetween growth adjustment and the acidification of thecell wall solution (Mizuno et al, 1993; Okamoto andOkamoto, 1994). These studies provide evidence for theclose coupling of metabolic events and biophysicalchanges of growth parameters.
Limitations of Lockhart's model
Lockhart's model has been proven to be very useful toquantify the growth process of a single cell. None the
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/5/985/542707 by guest on 10 January 2022

988 Frensch
less, there are obvious limitations of the model whenapplied to a growing tissue, which have not yet beenincorporated in the theoretical framework. Tissue growthdiffers from cell growth in that:(a) growth rates vary along the elongation zones in theplant, corresponding to (i) a spatial heterogeneity in thedemand for water and solutes, (ii) variable growth-relatedparameters with respect to position and time, and(b) water and solutes are supplied by different compart-ments (e.g. xylem, phloem, soil solution) and transportinvolves parallel pathways (apoplast and symplast), eachwith distinct physical and chemical properties.
Measurements at a high spatial and temporal resolutionare required to unravel causes and effects within thecomplex process of tissue growth, and to evaluate tissueresponse to environmental factors properly.
It is important to recall that equation (2) would predicta linear relationship between Pc and growth rate. Changesof Pc are commonly affected by altering the rate of wateruptake by means of changes in Wo, nc, and LpxA.Following perturbations of Pc, linearity would justify thatY and m can be determined from steady growth ratesand steady turgor pressures. Despite early concernsregarding the linearity (Acevedo et al., 1971; Green et al.,1971) this approach has been extensively used to deter-mine Y and m. Alternatively, Y has been determined fromturgor relaxation after water uptake into the cell wasinhibited (Ortega, 1985; Cosgrove, 1987). However, con-tinuous measurements of Pc during transient responses ofgrowth rates indicate that cell wall parameters are variableand adjust in a matter of min to new environmentalconditions (Shackel et al., 1987; Serpe and Matthews,1992; Frensch and Hsiao, 1994). Without adequate con-siderations of the variability, previous methods may leadto erroneous values of Y and m. Until now, Lockhart'smodel does not account for the variable nature of anyparameter in equation (1). A significant step toward thisdirection is the modelling of responses of microfibrils inthe cell wall to changes in turgor (Passioura and Fry,1992). Further measurements are required to relate meta-bolic events with physical restrictions or enhancements ofcell expansion.
The role of solute accumulation for growth
Water uptake and volume increase of the growing cellconstantly dilute the cell sap which would lead to turgorrelaxation until growth ceases at Pc= Y. Thus, cell expan-sion is intimately related to solute uptake and/or synthesis.To some extent, equation (1) accounts for this relationshipby the parameter nc. Introduced as part of the drivingforce for water flow, nc is also a measure of the soluteconcentration in the cell. It is evident, however, thatequation (1) does not govern the coupling of water andsolute flows during cell enlargement. This may be possible
by the introduction of a third 'growth resistance' thatwould couple solute flow to an appropriate force. Forexample, active solute uptake may be related to themetabolic energy of the cell, whereas passive solutemovement into the cell may be driven by a concen-tration gradient between adjacent cells connected byplasmodesmata.
Depending on the location, the various elongationzones of a plant gain solutes from different compartments.For the root elongation zone, solute accumulation isprobably sustained by two compartments, i.e. the soilsolution (nutrient salts) and the phloem (carbohydrates,nutrient salts). For above-ground organs, phloem andxylem supply growing cells. To measure rates of soluteaccumulation in a single cell, consider the followingexperiment. Turgor is lowered to a level well below Y,and cell expansion stops. As long as Pc < Y, the irrevers-ible volume increase is zero and changes in Pc would beproportional to changes in water and solute content ofthat cell. Provided that changes in solute concentrationwithin the protoplast would be the only process thatgenerates a driving force for water uptake, a change inturgor (APC) is a measure of the amount of soluteaccumulated by this cell (/l/is), and APC is given by(Steudle, 1989, 1992):
1
Cc+Cv,( A",X{RT~V
(3)
It can be seen from equation (3) that APC correspondsto the increase in Anc (Anc = RTxAnJV) and AVUstuc.A Vtiuuc denotes the net volume change of the entire cell(protoplast and cell wall). Changes of nc do not directlytranslate into APC. Rather, elastic and plastic deformationof the cell (Fxe/(e + nc), e is the elastic coefficient of acell) and the storage capacities of the wall (Cw) and theprotoplast (Cc) dampen effects of An, on APC. The factorF, which is approximately (1 + trjn xAt)'1 for <r»7rc,accounts for the plastic extension (mAt) of a cell. Incontrast to elastic extension (1/e), plastic extension istime-dependent; At denotes the time interval of relativevolume increase (AV/V). F approaches 1 for m->0 (i.e.in a non-growing cell) and for At—>0. If water gain forAPC is entirely from the apoplast of the cell, AVtiuue = 0and equation (3) becomes
e C w An.-T7- (4)
C c + w VBecause of the smaller storage capacity of the wallcompared to the protoplast, the ratio CW/(CC + CW) issignificantly smaller than unity and changes of APC willbe small. This relationship may illustrate the situation forshoot growth, when cell expansion strongly competeswith transpiration for water. Under these conditions,
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/5/985/542707 by guest on 10 January 2022

osmotic adjustment of the growing cell is rather inefficientto maintain Pc > Y.
With sufficient water supply, i.e. water loss in thewall due to uptake into the protoplast is immediatelyreplaced by an external water source, AVuauc equalsCc x RTxAnJV(Steud\e, 1992). Thus, effects of compart-mentation between apoplast and symplast cancel out andequation (3) reduces to
APC =An.
V(5)
Here, An, translates almost entirely into turgor changesprovided that e»7rc. Equation (5) represents the corres-ponding turgor change to an increase in solute contentof an expanding cell in the root elongation zone whenwater availability is unlimited. The net deposition rate ofsolutes into the elongation zone would then be given by:
An,
At
APC
At RT'
In the present study, AnjAt was calculated from previousdata of APJAt (Frensch and Hsiao, 1995). The calcula-tions assumed F=\, because APJAt was measured atzero root elongation and, hence, mAt = 0.
A different approach to obtain data on solute accumu-lation in tissues is to study growth in terms of fluid flow(Silk, 1984). This model yields quantitative informationon the magnitude and the spatial distribution of growthalong the elongation zone of an organ. Accordingly, netdeposition rates of matter for a small root segment arecalculated from concentrations of the molecule of interestwithin this segment and from the local growth rate ofthis segment. For maize roots, deposition rates of waterand several major osmotic compounds have been deter-mined for segments of 0.5 mm length (Sharp et al., 1990).Local deposition rates calculated by this model may becompared with measurements of AnJAt in single cells.
Mechanisms of adaptation to water deficits inroots
It has long been recognized that the expansion rate alongthe root elongation zone follows a bell-shaped curve. Fora maize root, the acceleration and deceleration phasesextent over a length of approximately 10 mm (Ericksonand Sax, 1956). More recently, elementary growth rateshave been combined with turgor measurements. Thesestudies show that turgor is rather uniform throughoutthe cortex of the elongation zone in spite of large differ-ences in local growth rates in wheat (Pritchard et al.,1990, 1991) and maize (Spollen and Sharp, 1991).Longitudinal gradients in wall rheology are thereforelikely the reason for the observed variability in localgrowth rates (Tomos and Pritchard, 1994). Despite the
Root elongation and drought 989
similarities in both species, a notable difference wasobserved in these studies when roots were exposed towater stress over medium long periods (hours to days).While maize roots seemed to increase the wall yieldingability, thus maintaining root growth at lower turgor,wheat roots appeared to do just the opposite, i.e. theyreduced wall yielding ability. The discrepancy may indi-cate a fundamental difference between the species duringthe adjustment to limited water supply.
Further investigations aimed to unravel primaryresponses in the root elongation zone, including wallrheology and water and solute transport properties, atshorter time intervals (Frensch and Hsiao, 1994, 1995).The results of turgor recordings during steady anddynamic conditions of root growth can be summarizedas follows:(i) Upon exposure of roots to hyperosmotic solutions, Pc
declined rapidly and growth decelerated or stopped almostimmediately. However, the inhibition of root growth wascompensated partially or completely within min due toboth a decline of the yield threshold (Y) and an osmoticresponse GdwJ that led to a recovery of Pc. A minimumpressure of 0.3 MPa was required for irreversible cellexpansion. During the short period of adjustment(1-30 min) the wall extensibility coefficient (m) remainedrather constant.(ii) The osmotic response of each cell was presumablythe result of carbohydrate uptake. Radial solute transportin the elongation zone was sensitive to turgor changes.With successively lower Pc in the cortex, the rate of solutetransport declined drastically.(iii) Consistent with earlier results on root growth (Sharpet al., 1988), the length of the elongation zone in maizewas reduced from originally 10 to approximately 7 mmat i/(m< —0.5 MPa. The permanent inhibition of growthin the proximal part of the elongation zone was likelydue to the lack of turgor recovery (solute transport).Therefore, Pc remained smaller than 0.3 MPa, the min-imum Y. Towards the apex growth resumed, becauseradial solute flow continued and exceeded Y eventually,(iv) The hydraulic conductivity of the tissue remainedhigh at all levels of Pc. Based on the hydraulic conductivitycoefficient of the tissue (K= 1.3 x 10"10 m2 s" 'MPa"')gradients of water potential across the cortex were calcu-lated. At maximum expansion rates in the middle of theelongation zone, gradients of only 0.03 MPa would beexpected.
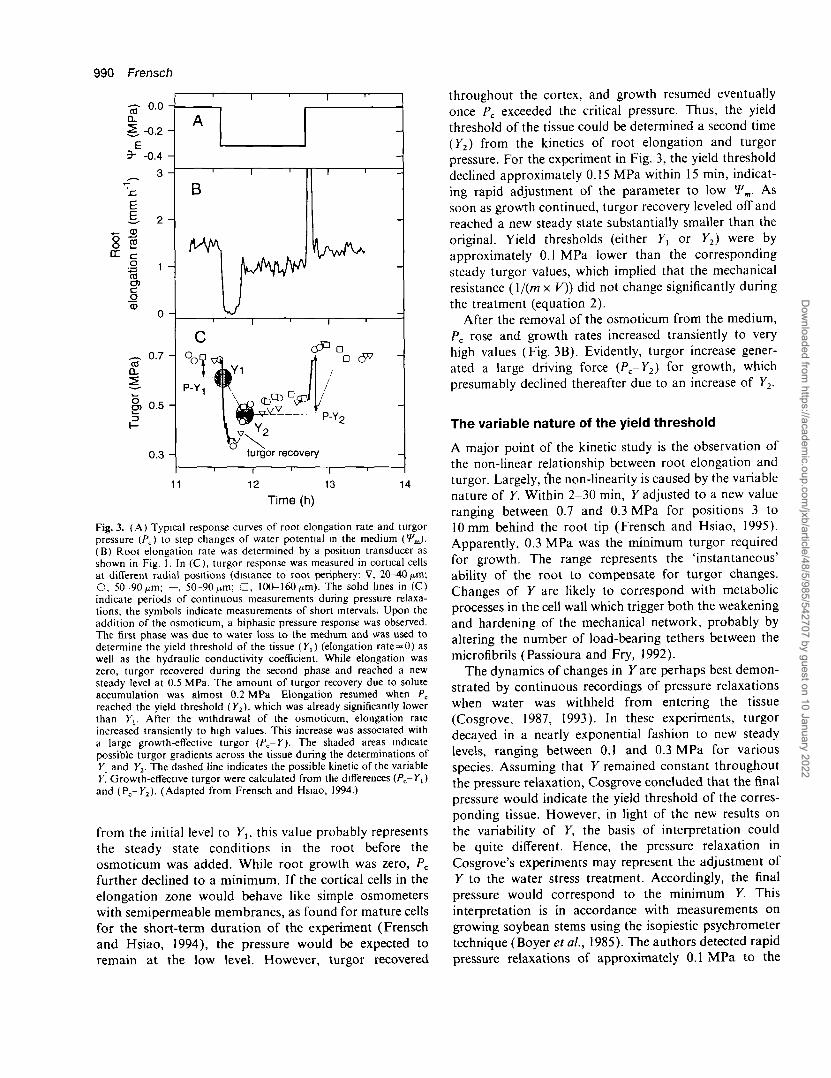
On the premise that root elongation stops at Pc = Y,and recalling that Pc was rather uniform across the cortex,the yield coefficient of the tissue (y,) was determinedfrom the decline of turgor following osmotic step changesin the medium (Fig. 3). The cross-hatched area indicatesthe approximate pressure gradient in the cortex at whichthe root elongation rate became zero. Because of thesmall time interval (few seconds) it took for Pc to drop
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/5/985/542707 by guest on 10 January 2022
990 Frensch
— 00 -CO
Q_
2 , -0.2 -
5* -0.4 -
_ 3 -'.cE•§- 2 -
* - CD
8 2•S 1 -COO)coCD
0 -
_ 0.7 -COa.~^8, 0.5-
Tur
0.3 -
1
A
1
B
AAM.
co g r t 3
1 ' 1 •
—
-
1 '
1 A L a
-
_
/
/
i ' i •
dP DA n rW
i /
1 j^zv____, p ^
turgor recovery
11I
12 13Time (h)
14
Fig. 3. (A) Typical response curves of root elongation rate and turgorpressure (/>c) to step changes of water potential in the medium (V.J.(B) Root elongation rate was determined by a position transducer asshown in Fig. 1. In (C), turgor response was measured in cortical cellsat different radial positions (distance to root periphery: V, 20-40 ̂ m;O, 50-90fxm; —, 50-90^m; D, 100-160 ^m). The solid lines in (C)indicate periods of continuous measurements during pressure relaxa-tions; the symbols indicate measurements of short intervals. Upon theaddition of the osmoticum, a biphasic pressure response was observed.The first phase was due to water loss to the medium and was used todetermine the yield threshold of the tissue (K,) (elongation rate = 0) aswell as the hydraulic conductivity coefficient. While elongation waszero, turgor recovered during the second phase and reached a newsteady level at 0.5 MPa. The amount of turgor recovery due to soluteaccumulation was almost 0.2 MPa Elongation resumed when Pcreached the yield threshold (Y2), which was already significantly lowerthan K,. After the withdrawal of the osmoticum, elongation rateincreased transiently to high values. This increase was associated witha large growth-effective turgor (Pc-Y). The shaded areas indicatepossible turgor gradients across the tissue during the determinations ofK, and Y2- The dashed line indicates the possible kinetic of the variableY. Growth-effective turgor were calculated from the differences (Pc-Yt)and (Pc-K2). (Adapted from Frensch and Hsiao, 1994.)
from the initial level to Yu this value probably representsthe steady state conditions in the root before theosmoticum was added. While root growth was zero, PQ
further declined to a minimum. If the cortical cells in theelongation zone would behave like simple osmometerswith semipermeable membranes, as found for mature cellsfor the short-term duration of the experiment (Frenschand Hsiao, 1994), the pressure would be expected toremain at the low level. However, turgor recovered
throughout the cortex, and growth resumed eventuallyonce Pc exceeded the critical pressure. Thus, the yieldthreshold of the tissue could be determined a second time(Y2) from the kinetics of root elongation and turgorpressure. For the experiment in Fig. 3, the yield thresholddeclined approximately 0.15 MPa within 15 min, indicat-ing rapid adjustment of the parameter to low Wm. Assoon as growth continued, turgor recovery leveled off andreached a new steady state substantially smaller than theoriginal. Yield thresholds (either Yt or Y2) were byapproximately 0.1 MPa lower than the correspondingsteady turgor values, which implied that the mechanicalresistance (\/(m x V)) did not change significantly duringthe treatment (equation 2).
After the removal of the osmoticum from the medium,Pc rose and growth rates increased transiently to veryhigh values (Fig. 3B). Evidently, turgor increase gener-ated a large driving force (Pc-Y2) f° r growth, whichpresumably declined thereafter due to an increase of Y2.
The variable nature of the yield threshold
A major point of the kinetic study is the observation ofthe non-linear relationship between root elongation andturgor. Largely, the non-linearity is caused by the variablenature of Y. Within 2-30 min, Y adjusted to a new valueranging between 0.7 and 0.3 MPa for positions 3 to10mm behind the root tip (Frensch and Hsiao, 1995).Apparently, 0.3 MPa was the minimum turgor requiredfor growth. The range represents the 'instantaneous'ability of the root to compensate for turgor changes.Changes of Y are likely to correspond with metabolicprocesses in the cell wall which trigger both the weakeningand hardening of the mechanical network, probably byaltering the number of load-bearing tethers between themicrofibrils (Passioura and Fry, 1992).
The dynamics of changes in Y are perhaps best demon-strated by continuous recordings of pressure relaxationswhen water was withheld from entering the tissue(Cosgrove, 1987, 1993). In these experiments, turgordecayed in a nearly exponential fashion to new steadylevels, ranging between 0.1 and 0.3 MPa for variousspecies. Assuming that Y remained constant throughoutthe pressure relaxation, Cosgrove concluded that the finalpressure would indicate the yield threshold of the corres-ponding tissue. However, in light of the new results onthe variability of Y, the basis of interpretation couldbe quite different. Hence, the pressure relaxation inCosgrove's experiments may represent the adjustment ofY to the water stress treatment. Accordingly, the finalpressure would correspond to the minimum Y. Thisinterpretation is in accordance with measurements ongrowing soybean stems using the isopiestic psychrometertechnique (Boyer et al., 1985). The authors detected rapidpressure relaxations of approximately 0.1 MPa to the
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/5/985/542707 by guest on 10 January 2022
Roof elongation and drought 991
presumed Y, followed by a slower pressure reductionafterwards.
Consistent with the results on leaf growth and resultsof other studies, (Pc-Y) for roots was small and cellexpansion responded to slight changes in Pc (Matysseket al., 1988; Okamoto et al, 1989). Compared with theconventional method, i.e. when Y is calculated fromsteady state relationships between growth rate and Pc,the direct approach yielded much higher values of Y and,thus, smaller values of (Pc-Y) (Frensch and Hsiao, 1994).The discrepancy may caution to evaluate Y and m apply-ing the linear relationship of the Lockhart equation toexperiments where the water status of the root is altered.Earlier conclusions drawn on the assumption that Ybehaves like a constant may be premature.
Is the determination of Y confounded by elasticresponses?
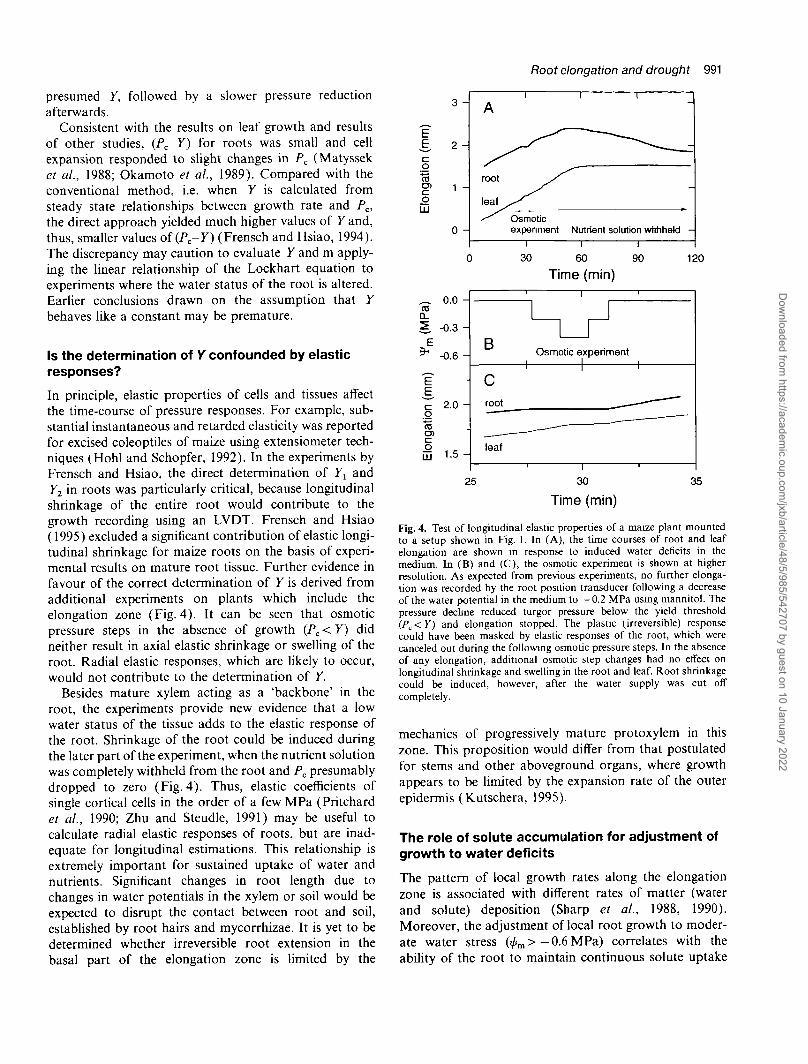
In principle, elastic properties of cells and tissues affectthe time-course of pressure responses. For example, sub-stantial instantaneous and retarded elasticity was reportedfor excised coleoptiles of maize using extensiometer tech-niques (Hohl and Schopfer, 1992). In the experiments byFrensch and Hsiao, the direct determination of Yx andY2 in roots was particularly critical, because longitudinalshrinkage of the entire root would contribute to thegrowth recording using an LVDT. Frensch and Hsiao(1995) excluded a significant contribution of elastic longi-tudinal shrinkage for maize roots on the basis of experi-mental results on mature root tissue. Further evidence infavour of the correct determination of Y is derived fromadditional experiments on plants which include theelongation zone (Fig. 4). It can be seen that osmoticpressure steps in the absence of growth (Pc < Y) didneither result in axial elastic shrinkage or swelling of theroot. Radial elastic responses, which are likely to occur,would not contribute to the determination of Y.
Besides mature xylem acting as a 'backbone' in theroot, the experiments provide new evidence that a lowwater status of the tissue adds to the elastic response ofthe root. Shrinkage of the root could be induced duringthe later part of the experiment, when the nutrient solutionwas completely withheld from the root and Pc presumablydropped to zero (Fig. 4). Thus, elastic coefficients ofsingle cortical cells in the order of a few MPa (Pritchardet al., 1990; Zhu and Steudle, 1991) may be useful tocalculate radial elastic responses of roots, but are inad-equate for longitudinal estimations. This relationship isextremely important for sustained uptake of water andnutrients. Significant changes in root length due tochanges in water potentials in the xylem or soil would beexpected to disrupt the contact between root and soil,established by root hairs and mycorrhizae. It is yet to bedetermined whether irreversible root extension in thebasal part of the elongation zone is limited by the
COO)
LLJ
3 -
2 -
1 -
0 -
A
root
leaf
I
Osmoticexperiment
i
i i
Nutrient solution withheld -
i i30 60 90
Time (min)120
25
Time (min)
Fig. 4. Test of longitudinal elastic properties of a maize plant mountedto a setup shown in Fig. 1. In (A), the time courses of root and leafelongation are shown in response to induced water deficits in themedium. In (B) and (C), the osmotic experiment is shown at higherresolution. As expected from previous experiments, no further elonga-tion was recorded by the root position transducer following a decreaseof the water potential in the medium to - 0 . 2 MPa using mannitol. Thepressure decline reduced turgor pressure below the yield threshold(PC<Y) and elongation stopped. The plastic (irreversible) responsecould have been masked by elastic responses of the root, which werecanceled out during the following osmotic pressure steps. In the absenceof any elongation, additional osmotic step changes had no effect onlongitudinal shrinkage and swelling in the root and leaf. Root shrinkagecould be induced, however, after the water supply was cut offcompletely.
mechanics of progressively mature protoxylem in thiszone. This proposition would differ from that postulatedfor stems and other aboveground organs, where growthappears to be limited by the expansion rate of the outerepidermis (Kutschera, 1995).
The role of solute accumulation for adjustment ofgrowth to water deficits
The pattern of local growth rates along the elongationzone is associated with different rates of matter (waterand solute) deposition (Sharp et al., 1988, 1990).Moreover, the adjustment of local root growth to moder-ate water stress (4>m> — 0.6 MPa) correlates with theability of the root to maintain continuous solute uptake
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/5/985/542707 by guest on 10 January 2022
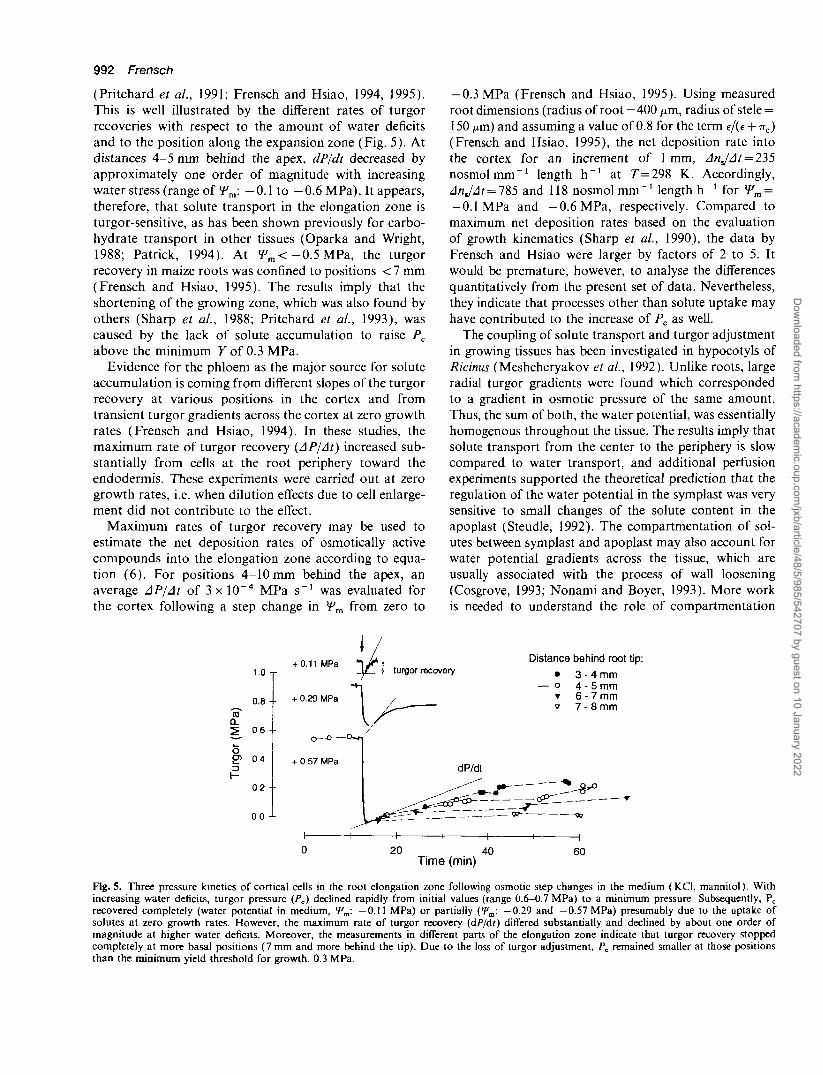
992 Frensch
(Pritchard et al., 1991; Frensch and Hsiao, 1994, 1995).This is well illustrated by the different rates of turgorrecoveries with respect to the amount of water deficitsand to the position along the expansion zone (Fig. 5). Atdistances 4-5 mm behind the apex, dP/dt decreased byapproximately one order of magnitude with increasingwater stress (range of *Fm: —0.1 to —0.6 MPa). It appears,therefore, that solute transport in the elongation zone isturgor-sensitive, as has been shown previously for carbo-hydrate transport in other tissues (Oparka and Wright,1988; Patrick, 1994). At f m < - 0 . 5 MPa, the turgorrecovery in maize roots was confined to positions < 7 mm(Frensch and Hsiao, 1995). The results imply that theshortening of the growing zone, which was also found byothers (Sharp et al., 1988; Pritchard et al., 1993), wascaused by the lack of solute accumulation to raise Pc
above the minimum Y of 0.3 MPa.Evidence for the phloem as the major source for solute
accumulation is coming from different slopes of the turgorrecovery at various positions in the cortex and fromtransient turgor gradients across the cortex at zero growthrates (Frensch and Hsiao, 1994). In these studies, themaximum rate of turgor recovery (AP/At) increased sub-stantially from cells at the root periphery toward theendodermis. These experiments were carried out at zerogrowth rates, i.e. when dilution effects due to cell enlarge-ment did not contribute to the effect.
Maximum rates of turgor recovery may be used toestimate the net deposition rates of osmotically activecompounds into the elongation zone according to equa-tion (6). For positions 4-10 mm behind the apex, anaverage AP/At of 3 x 10~4 MPa s"1 was evaluated forthe cortex following a step change in x¥m from zero to
—0.3 MPa (Frensch and Hsiao, 1995). Using measuredroot dimensions (radius of root=400 ^m, radius of stele =150 ̂ m) and assuming a value of 0.8 for the term e/(e + 77c)(Frensch and Hsiao, 1995), the net deposition rate intothe cortex for an increment of 1 mm, Anj'At = 235nosmolmm"1 length h"1 at 7=298 K. Accordingly,AnJAt = l&5 and 118 nosmolmm"1 length h"1 for ¥/
m =— 0.1 MPa and —0.6 MPa, respectively. Compared tomaximum net deposition rates based on the evaluationof growth kinematics (Sharp et al., 1990), the data byFrensch and Hsiao were larger by factors of 2 to 5. Itwould be premature, however, to analyse the differencesquantitatively from the present set of data. Nevertheless,they indicate that processes other than solute uptake mayhave contributed to the increase of Pc as well.
The coupling of solute transport and turgor adjustmentin growing tissues has been investigated in hypocotyls ofRicinus (Meshcheryakov et al, 1992). Unlike roots, largeradial turgor gradients were found which correspondedto a gradient in osmotic pressure of the same amount.Thus, the sum of both, the water potential, was essentiallyhomogenous throughout the tissue. The results imply thatsolute transport from the center to the periphery is slowcompared to water transport, and additional perfusionexperiments supported the theoretical prediction that theregulation of the water potential in the symplast was verysensitive to small changes of the solute content in theapoplast (Steudle, 1992). The compartmentation of sol-utes between symplast and apoplast may also account forwater potential gradients across the tissue, which areusually associated with the process of wall loosening(Cosgrove, 1993; Nonami and Boyer, 1993). More workis needed to understand the role of compartmentation
CO
a.
1.0 -r
0.8 --
06 --
OO) 04 --
02 -•
0.0 -1-
+ 0.11 MPa Distance behind root tip:
• 3 - 4 mm— o 4 - 5 mm
6 - 7 mm7 - 8 mm
20 40Time (min)
60
Fig. 5. Three pressure kinetics of cortical cells in the root elongationincreasing water deficits, turgor pressure (Pc) declined rapidly fromrecovered completely (water potential in medium, fm: —0.11 MPa)solutes at zero growth rates. However, the maximum rate of turgormagnitude at higher water deficits. Moreover, the measurements incompletely at more basal positions (7 mm and more behind the tip),than the minimum yield threshold for growth, 0.3 MPa.
zone following osmotic step changes in the medium (KCl, mannitol). Withinitial values (range 0.6-0.7 MPa) to a minimum pressure Subsequently, Pcor partially (>?„: -0.29 and -0.57 MPa) presumably due to the uptake ofrecovery (dP/dt) differed substantially and declined by about one order of
different parts of the elongation zone indicate that turgor recovery stoppedDue to the loss of turgor adjustment, Pc remained smaller at those positions
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/5/985/542707 by guest on 10 January 2022

and, especially, that of the apoplast within the complexityof tissue growth (Steudle and Frensch, 1996).
What limits root growth?
As already stated, limitations of growth are usuallydiscussed in terms of hydraulic and mechanical resist-ances. The relative magnitude of the resistances can beestimated from the ratio (Boyer, 1985)
(7)
Roof elongation and drought 993
Unstressed root
LpA Pc-Y
according to the Lockhart theory. Values significantlysmaller than 1 indicate a growth-limitation by the wallextensibility. Using measured values of (Pc — Y) and calcu-lated water potential gradients in growing maize roots(Silk and Wagner, 1980; Frensch and Hsiao, 1995), theratio of mechanical versus hydraulic resistance rangesbetween 0.1 and 0.5 for unstressed and moderatelystressed roots, depending on the axial and radial positionof the expanding cell in the tissue. Cells at their maximumlocal growth rate, which make up most of the over-allgrowth rate, have values closer to 0.5, which suggeststhat both (or neither) of the parameters is rate-limiting.
It is tempting to argue that solute accumulation couldhave become the rate-limiting process for growth underwater stress. In favor of this hypothesis are the possiblepressure-sensitive distribution of carbohydrates and theclose relationship between the reduced length of theelongation zone and the loss of turgor recovery (Frenschand Hsiao, 1995). Carbohydrates account for approxi-mately 50% of the net solute deposition rate in maizeroots (Sharp et ai, 1990). They arrive from the shootand are distributed radially into the growth zone alongthe symplastic route (Fig. 6). The flow rate of sugarsdecreases progressively with farther distance from thephloem until it eventually drops to zero in the epidermis.Simultaneously, water enters the tissue largely across theepidermis, driven by water potential gradients induced byeither wall relaxation or solute gradients along the radialpath. Under steady conditions, a 'functional equilibrium'is expected for this position, where the local growth rateis determined by cells limited in either carbohydrate orwater supply. The ratio between the rate constants ofwater and solute flow would then determine whether theoverall growth rate is closer associated with cell layers atthe periphery or with those in the center of the root.With increasing water stress and lower levels of Pc, theresults in Fig. 5 suggest that the epidermis could becomethe growth-limiting cell layer in the root, provided that(Pc-Y) can adjust to positive values. On the other hand?turgor recovered rapidly in cortical cells following smallperturbations in the medium ( f m > = —0.1 MPa). Thisindicates that there is no obvious limiting factor or tissueduring steady growth conditions.
Carbohydrate flow » Water flow
Low medium water potential
Fig. 6. Schematical drawing of a cross section of the maize rootillustrating radial water and carbohydrate flows in the elongation zonefor unstressed and water stressed roots. The location of the phloem isrepresented by the cross-hatched ring. The arrows indicate the directionsof flow, and the thickness approximates the radial flow rates within thegrowing tissue.
There is growing evidence that plasmodesmata repres-ent the major resistance for symplastic solute flow (Bret-Harte and Silk, 1994). Dye-injection studies suggest thatsymplastic unloading from the phloem ceases when rootgrowth is reduced (Oparka et al., 1995). Although thereduced permeability would explain the results presentedin the present study, there is also evidence for the contrary.Using electron microscopy, Schulz (1995) reports awidening of plasmodesmata in pea roots in response toosmotic step changes in the medium, which wouldenhance symplastic solute flow at constant concentrationgradients across the tissue. Perhaps, the lack of turgorrecovery correlates with a smaller driving force for phloemunloading at low fm, which requires further attention infuture experiments. In either case, solute transport intothe elongation zone appears critical for continuous rootgrowth in water-stressed and unstressed roots.
Growth limitations: scaling up from cell toorgan level
So far, the discussion on interactions between cellenlargement and water shortage has been confined to the
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/5/985/542707 by guest on 10 January 2022

994 Frensch
elongation zones of the root and leaf. Different plantorgans, however, do not experience the same amount ofwater stress which relates to the observed differentialgrowth responses of the two organs (Fig. 1). The gradi-ents in water potential between the elongation zones inroot and shoot largely depend on the hydraulics of theroot system. Therefore, growth limitations as related toaspects of water transport in roots shall now be discussed.
Water loss of the plant to the atmosphere generates atension in the xylem which, according to the cohesiontheory, propagates into the root and reduces the waterpotential of the xylem below that of the soil. Limited byhydraulic resistances in series and parallel, water movesalong the gradient from the soil to the shoot. It is useful,therefore, to separate the volume flow into a radial andan axial component. The longitudinal flow is solely drivenby hydrostatic pressure gradients in the xylem. Withinthe radial pathway, water potential gradients may behydrostatic, osmotic, or matric in nature. In the absenceof transpiration, the osmotic gradient dominates, whichleads to guttation and xylem exudation in many plants.
Radial water flow in roots is commonly analysedapplying a two-compartment model in which the xylemis separated from the soil by a complex barrier (Passioura,1988; Steudle, 1994; Steudle and Frensch, 1996). Theapplication of the root pressure probe technique to rootsof herbaceous and woody species has yielded the remark-able result that the radial hydraulic conductivity of theroot (Lpr) strongly depended on the nature of the drivingforce. In tree roots, for example, Lpr(hydrostatic) waslarger than L/;r(osmotic) by one to three orders of magni-tude, whereas no difference between L/?r(hydrostatic) andL/?r(osmotic) was found in Phaseolus and barley (Steudle,1994). Radial volume flow rates in maize are larger byapproximately one order of magnitude in the presence ofhydrostatic pressure gradients compared to osmotic(Fig. 7). The experiments on plants of different ageshow that discrepancies between LpT(hydrostatic) andL/>r(osmotic) were found consistently in young roots aswell as in three-weeks old root systems, when the lengthof the main root axis exceeded 600 mm.
The data in Fig. 7 further illustrate a decreasing effici-ency of roots in water uptake when the main axis exceededa length of approximately 200 mm. This result is notsurprising if one considers the developmental gradientsalong the root axis. From the apex toward the base, thefunction of the root shifts from a tissue more specializedin material uptake to one more specialized in materialtranslocation. To some extent this process may be com-pensated by the development of laterals, although thiseffect appeared to be small in the present experiments onmaize roots. For roots exposed to uniform environmentalconditions, water uptake characteristics in hydrostaticexperiments remained rather constant along the apical150 mm. The decline of L/?r(hydrostatic) by one order of
,-'nj
,r\nE
30 -
20 -
i
oo
1
1 1
o•
o
o
1 1
1 1
Lp, (hydrostatic) _
Lp, (osmotic)
-
1 I
10 -
0 -
0 200 400 600
Length of main root axis (mm)
Fig. 7. Radial hydraulic conductivity (Lp,) of maize roots (age 2-24 d)grown in solution culture The primary root of a maize plant wasexcised and attached to the root pressure probe (Steudle and Frensch,1989). Hydrostatic pressure steps were induced by means of the probe;osmotic step changes were applied by replacing the root medium witha solution of higher osmotic pressure. Both experiments resultedin a radial flow of water which caused a pressure relaxation in thexylem. The two types of pressure relaxation experiments yieldedL/>r(hydrostatic) and L/>r(osmotic). Each data point is the mean of2-14 measurements on a single root. The data on maize roots showtwo important results: L/>r(hydrostatic)>Lpr(osmotic), and Lp, declineswith increasing root size The dashed lines indicate Lp,(hydrostatic)before the emergence of laterals (25 x 10~8m s"1 MPa"1) and afterlaterals amounted for more than 90% of the total root surface area(2.2x IO"8m s"
magnitude coincided with the growth of root laterals,emerging from the cortex approximately 150 mm behindthe apex. After 14 d in nutrient solution, when the mainaxis had reached lengths of approximately 500 mm,laterals accounted for more than 90% of the total rootsurface area (Frensch, 1990). Hence, small LpT of olderroots largely represents the hydraulic conductivity of thelaterals. It may be concluded that the laterals of maizeroots grown in solution culture are less efficient in termsof water uptake and transport than the main axis.
Radial water transport in roots is composite in nature(Steudle, 1994). The corresponding model involves twoparallel pathways: an apoplastic and a cell-to-cell path,each exhibiting different hydraulic and osmotic properties.It has been supported by pressure clamp experiments onPrunus using a different experimental and theoreticalapproach (Magnani et a!., 1996). Despite its advantages,it is worth noting that this model simplifies the processof water transport in roots considerably. Root develop-ment and function depends much on the physical andchemical conditions of the immediate environment, whichis largely unexplored for most plants under natural condi-tions (Waisel et al., 1991; McCully, 1995). Both axialand radial resistances as well as water potential gradientsvary with time and space along the root. The implicationsfor the hydraulic architecture of the root have beenstudied theoretically and experimentally on excised andintact roots using pressure probe techniques (Frensch and
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/5/985/542707 by guest on 10 January 2022

Steudle, 1989; Frensch and Hsiao, 1993; Frensch et al.,1996), and will be subject of the following chapter.
Pressure propagation across the root andimplications for leaf and root growth
Information on soil water content is transmitted to theshoot by hydraulic and metabolic 'signals' originating inthe root. They modulate stomatal movement and shootgrowth within complex control loops (Tardieu andDavies, 1993). Over the past decade, the action of thephytohormone abscisic acid has been investigated extens-ively, whereas the meaning of the hydraulic componentwas largely neglected. To some extent, the negligence mayresult from the lack of appropriate techniques to measureand to modify xylem pressures in plants. Numerousmeasurements of xylem water potential by indirectmethods (e.g. psychrometry, Scholander's pressure-chamber technique) have predicted substantial tensionsin the xylem of a transpiring plant. Recent studies haveyielded satisfactory results on the reliability of the pressurechamber technique, i.e. to correlate applied (positive)pressures with tensions in excised plant material(Holbrook et al., 1995; Pockman et al., 1995). Data ofnegative pressures in these studies are in accordance withthe cohesion theory, which has prevailed the literature onwater movement in plants over the last 100 years.However, the dogmas of this theory have been attackedon the basis of direct measurements in the xylem using amodified cell pressure probe (Balling and Zimmermann,1990; Zimmermann et al., 1993). The authors reporthigher (less negative) values in the xylem of herbs andtree species compared to Px obtained by the pressurechamber technique. Direct xylem measurements in treesindicated that longitudinal pressure gradients were toosmall to move water from the roots to the leaf by a purelyhydraulic mechanism (Benkert et al., 1995). If true, theresults of Zimmermann and co-workers would changebasic concepts of the mechanism of long-distance trans-port in the xylem. However, the results have to be takencautiously. Due to technical limitations of the cell pressureprobe in the negative pressure range, which have not beenaddressed satisfactorily in the studies by Zimmermannand co-workers, the critique against the cohesion theoryhas been refused (Steudle, 1995).
The existence of longitudinal pressure gradients is akey feature of the cohesion theory and also relevant forplant growth. How effectively do pressure changes in theshoot propagate into the root and further into the soil?Recently, the propagation of hydrostatic pressures wasmeasured in the cortex and in the xylem of maize rootsystems (Frensch and Hsiao, 1993; Frensch et al., 1996).These experiments provided novel results on the inter-actions between root structure and function. When theshoot was replaced by a modified root pressure probe,
Roof elongation and drought 995
volume flows could be induced either into or out of thecut end of the excised root (Fig. 8). The cell pressureprobe was used to scan the kinetics of the pressurepropagation along mature and immature xylem (AP^) ofroot systems up to 500 mm in length. As expected, APipp
was rapidly transmitted (fractions of a second) alongconductive xylem. Longitudinal pressure propagation wasdetermined from constant values of APX and APhpp.Ratios of APJAPBpp ranged from values close to 1 at thebase of the root to almost zero at the root tip (Frenschet al., 1996). Based on measured APJAPnpp, a hypothet-ical pressure profile was calculated for the boundarycondition Px= - 0 . 5 MPa atz = 500 mm and Wm =0 MPa(closed symbols in Fig. 8). The data show that a tensionin the shoot is maintained for a long distance in the rootxylem, but steeply attenuates toward zero in the apical200 mm.
The attenuation of xylem pressure is important for thedifferential growth of roots and shoots in the presence ofwater deficits in the atmosphere and in the soil. In ourexample, the leaf elongation zone would experience atension in the xylem (apoplast) of —0.5 MPa or less,while Px in the root elongation zone would be almostzero. Frensch and Hsiao (1993) concluded that rootelongation was rather hydraulically isolated from changesin water potential that originate in the shoot. On thecontrary, water potential gradients along the root aredifferent when water stress is imposed through osmoticchanges in the medium. In the absence of significanttranspiration, the osmoticum reduces P% more evenly overthe entire length of the root. Due to reflection coefficientsof the root smaller than unity, values of APX are smallerthan the corresponding A-nm (Steudle and Frensch, 1989).Hence, root growth is probably more affected than shootgrowth when Wm is altered.
Assuming that transpiration is the major driving forcefor water transport, the experiments on pressure propaga-tion along the xylem resemble the situation in the intactplant. Further steps toward an understanding of dynamicpressure changes in the xylem under transpiring condi-tions require direct measurements in the xylem. Theseexperiments were performed on plants kept in eithersolution culture or in soil (Frensch, Dieffenbach andGottlein, unpublished results). Reliable tests of the cellpressure probe along with further control experimentsdemonstrated convincingly that the measurements wereperformed in conductive xylem. Negative xylem pressuresof down to —0.5 MPa absolute were recorded in rootsof maize plants. They depended not only on transpir-ational water loss, but also on the availability of waterin the medium and soil. Essentially, the results verifiedthat transpiration generates a rapid and effective drivingforce for water uptake in the xylem. Furthermore, thestudies explain the apparent lack between transpirationand xylem pressure, which was used by Zimmermann and
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/5/985/542707 by guest on 10 January 2022
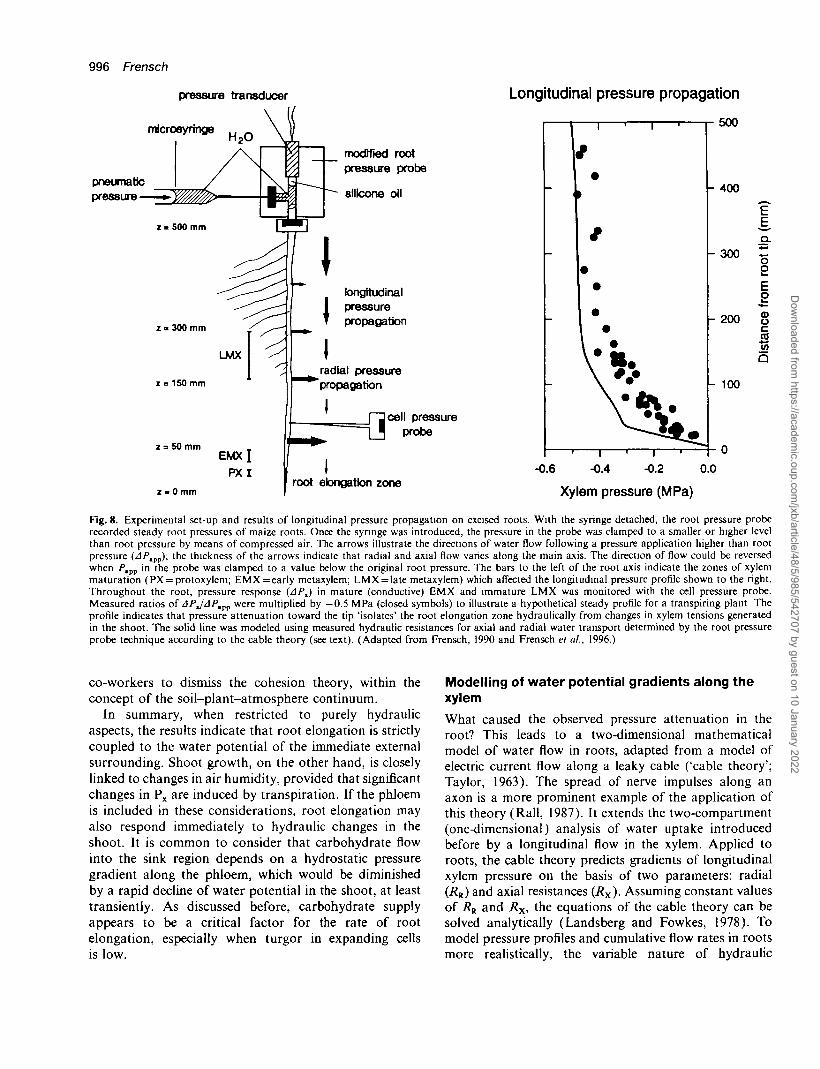
996 Frensch
pressure transducer Longitudinal pressure propagation
rrdcroeyringe
pneumaticpressure —
500
modified rootpressure probe
8lllcone oil
longitudinalI pressure* propagation
z = 150 mm
z = 50 mm
z « 0 mm
radial pressurepropagation
cell pressureprobe
Ioot elongation zone
-0.6 -0.4 -0.2 0.0
Xylem pressure (MPa)
Fig. 8. Experimental set-up and results of longitudinal pressure propagation on excised roots. With the syringe detached, the root pressure proberecorded steady root pressures of maize roots. Once the syringe was introduced, the pressure in the probe was clamped to a smaller or higher levelthan root pressure by means of compressed air. The arrows illustrate the directions of water flow following a pressure application higher than rootpressure (APtvp); the thickness of the arrows indicate that radial and axial flow varies along the main axis. The direction of flow could be reversedwhen P,m in the probe was clamped to a value below the original root pressure. The bars to the left of the root axis indicate the zones of xylemmaturation (PX = protoxylem; EMX = early metaxylem; LMX = late metaxylem) which affected the longitudinal pressure profile shown to the right.Throughout the root, pressure response (APJ in mature (conductive) EMX and immature LMX was monitored with the cell pressure probe.Measured ratios of APJAPtpp were multiplied by —0.5 MPa (closed symbols) to illustrate a hypothetical steady profile for a transpiring plant Theprofile indicates that pressure attenuation toward the tip 'isolates' the root elongation zone hydraulically from changes in xylem tensions generatedin the shoot. The solid line was modeled using measured hydraulic resistances for axial and radial water transport determined by the root pressureprobe technique according to the cable theory (see text). (Adapted from Frensch, 1990 and Frensch et al., 1996.)
co-workers to dismiss the cohesion theory, within theconcept of the soil-plant-atmosphere continuum.
In summary, when restricted to purely hydraulicaspects, the results indicate that root elongation is strictlycoupled to the water potential of the immediate externalsurrounding. Shoot growth, on the other hand, is closelylinked to changes in air humidity, provided that significantchanges in Px are induced by transpiration. If the phJoemis included in these considerations, root elongation mayalso respond immediately to hydraulic changes in theshoot. It is common to consider that carbohydrate flowinto the sink region depends on a hydrostatic pressuregradient along the phloem, which would be diminishedby a rapid decline of water potential in the shoot, at leasttransiently. As discussed before, carbohydrate supplyappears to be a critical factor for the rate of rootelongation, especially when turgor in expanding cellsis low.
Modelling of water potential gradients along thexylem
What caused the observed pressure attenuation in theroot? This leads to a two-dimensional mathematicalmodel of water flow in roots, adapted from a model ofelectric current flow along a leaky cable ('cable theory';Taylor, 1963). The spread of nerve impulses along anaxon is a more prominent example of the application ofthis theory (Rail, 1987). It extends the two-compartment(one-dimensional) analysis of water uptake introducedbefore by a longitudinal flow in the xylem. Applied toroots, the cable theory predicts gradients of longitudinalxylem pressure on the basis of two parameters: radial(RR) and axial resistances (Rx)- Assuming constant valuesof 7?R and Rx, the equations of the cable theory can besolved analytically (Landsberg and Fowkes, 1978). Tomodel pressure profiles and cumulative flow rates in rootsmore realistically, the variable nature of hydraulic
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/5/985/542707 by guest on 10 January 2022

resistances was incorporated in the theory (Frensch andSteudle, 1989; Frensch, 1990). Using the root pressureprobe technique to evaluate RR and Rx (Frensch andSteudle, 1989; Frensch, 1990), the pressure attenuationalong the xylem was modeled for the maize root system(Fig. 8, solid line). Similar pressure profiles were calcu-lated for roots of Agave and Opuntia (Aim et al., 1992).It can be seen in Fig. 8 that the two independentlydetermined profiles correspond reasonably well. Dis-crepancies between the profiles are probably due to thefact that the model (i) neglects the distribution of resist-ances within the laterals (largely unknown), and (ii)overestimates the contribution of axial resistances in thetip region (0-20 mm), which is difficult to assess with theroot pressure probe.
According to the cable theory, the ratio RfJRx is afundamental parameter for the longitudinal pressuregradient in the xylem and, hence, for the distribution ofwater potential differences between the xylem and theroot surface. An increase of Rx would be associated witha higher proportion of immature xylem. Because resist-ances decrease with the forth power of the vessel radius,changes of Rx due to xylem maturation are huge. Withthe differentiation of the early and late metaxylem, ^?x
decreased by 5 orders of magnitude in maize roots(Frensch, 1990); changes in Rx by 3 orders of magnitudewere reported for onion roots (Melchior and Steudle,1993). Both studies emphasize to measure Rx rather thanto estimate this parameter from Poiseuille's law becauseof significant differences between the two approaches.Changes in RR along the main root axis are affected bychanges in both the apoplastic and cell-to-cell path. Inolder parts of grass roots, the endodermis is a likelycandidate for the radial hydraulic barrier (Sanderson,1983; McCully and Canny, 1988). The dominating roleof the endodermis for radial as well as axial pressureattenuation was demonstrated for maize roots (Frenschet al., 1996). The results are consistent with measurementsof water uptake in various root zones of barley andspruce (Sanderson, 1983; Haussling et al., 1988). Withinthe radial pathway, the lateral walls of mature vesselsmay also pose a significant resistance to water uptake(Peterson and Steudle, 1993), whereas the contributionof developmental changes in the hypodermis appears tobe rather small (Clarkson et al., 1987).
Taken together, experimental data on pressure propaga-tion in the xylem and cortex validate predictions of thecable theory. The model provides a sound basis to studythe relationship between root structure and function at amechanistic level. Future work should aim to include rootgrowth and the responses of growth to water stress as well.
Conclusions
It has been known for a long time that cell expansion isextremely sensitive to changes in plant water potential,
Root elongation and drought 997
and that growth recovers rapidly from mild and moderatewater stress. Recent progress toward a better understand-ing of the underlying processes is highlighted by thisreview. Measurements ranging from the cell to the organlevel emphasize to study growth responses of plants atboth high spatial and temporal resolution. The resultsconfirm the fundamental relationship between turgor andcell expansion, which is usually masked when conven-tional approaches are applied, i.e. when steady conditionsof turgor and growth are compared. In the past, this hasled to confusion about the meaning of turgor for cellenlargement.
Evidence is presented that rapid modifications of theyield threshold as well as turgor-dependent solute trans-port into the growth zone are important features of rootsduring the process of adjustment. From these data itshould be clear that growth and its recovery from transi-ent inhibition is closely linked to solute transport. Anadequate model of tissue expansion, which is still missing,should include pathways, mechanisms, and amounts ofmatter supplied by different compartments.
The differential responses of root and shoot growth arerelated to longitudinal water potential gradients in theplant. The attenuation of xylem pressure in roots isreadily explained by the 'cable theory'. It combines fea-tures of root structure (hydraulic resistances) and rootfunction (water transport) and demonstrates that theelongation zones in the root and shoot of maize arehydraulically separated, consistent with previous experi-mental results. The data emphasize the need to studyinteractions between growth and water relations on awhole plant level.
Acknowledgements
The research was supported by the Award of a Feodor Lynen-Fellowship from the Alexander von Humboldt-Foundation(Germany) and by a Fellowship from the DeutscheForschungsgemeinschaft to JF. I gratefully acknowledge thesupport of Drs Theodore C Hsiao, University of California,Davis, and Ernst Steudle, University of Bayreuth.
References
Acevedo E, Hsiao TC, Henderson DW. 1971. Immediate andsubsequent growth responses of maize leaves to changes inwater status. Plant Physiology 48, 631-6.
Aim DM, Cavelier J, Nobel PS. 1992. A finite-element modelof radial and axial conductivities for individual roots:development and validation for two desert succulents. Annalsof Botany 69, 87-92.
Balling A, Zinunennann U. 1990. Comparative measurementsof the xylem pressure of Nicotiana plants by means of thepressure bomb and probe. Planta 182, 325-45.
Benkert R, Zhu JJ, Zimmennann G, TOrk R, Bentrup F-W,Zinunennann U. 1995. Long-term xylem pressure measure-ments in the liana Tetrastigma voinierianum by means of thexylem pressure probe. Planta 196, 804-13.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/5/985/542707 by guest on 10 January 2022

998 Frensch
Boyer JS. 1985. Water transport. Annual Review of Plan!Physiology 36, 473-516.
Boyer JS, Cavalieri AJ, Schulze E-D. 1985. Control of the rateof cell enlargement: excision, wall relaxation, and growth-induced water potentials. Planta 163, 527-43.
Bradford KJ, Hsiao TC. 1982. Physiological responses tomoderate water stress. In: Lange OL, Nobel PS, OsmondCB, Ziegler H, eds. Encyclopedia of plant physiology Vol. 12B.Physiological Plant Ecology If. Water relations and carbonassimilation. Berlin: Springer-Verlag, 263-324.
Bret-Harte MS, Silk WK. 1994. Nonvascular, symplasmicdiffusion of sucrose cannot satisfy the carbon demands ofgrowth in the primary root tip of Zea mays L. PlantPhysiology 105, 19-33.
Carpita NC, Gibeaut DM. 1993. Structural models of primarycell walls in flowering plants: consistency of molecularstructure with the physical properties of the walls duringgrowth. The Plant Journal 3, 1-30.
Clarkson DT, Robards AW, Stephens JE, Stark M. 1987.Suberin lamellae in the hypodermis of maize {Zea mays)roots: development and factors affecting the permeability ofhypodermal layers. Plant, Cell and Environment 10, 83-93.
Cosgrove DJ. 1987. Wall relaxations in growing stems: compar-ison of four species and assessment of measurement tech-niques. Planta 171, 266-78.
Cosgrove DJ. 1993. Wall extensibility: its nature, measurementand relationship to plant cell growth. New Phvtologist124, 1-23.
Erickson RO, Sax KB. 1956. Elemental growth rate of theprimary root of Zea mays. Proceedings of the AmericanPhilosophical Society 100, 487-98.
Frenscb J. 1990. Wasser- und Teilchentransport in Maiswurzeln.PhD thesis, Bayreuth, Germany.
Frensch J, Hsiao TC. 1993. Hydraulic propagation of pressurealong immature and mature xylem vessels of roots of Zeamays measured by pressure probe techniques. Planta 190,263-70.
Frensch J, Hsiao TC. 1994. Transient responses of cell turgorand growth of maize roots as affected by changes in waterpotential. Plant Physiology 104, 247-54.
Frensch J, Hsiao TC. 1995. Rapid response of the yieldthreshold and turgor regulation during adjustment of rootgrowth to water stress in Zea mays. Plant Physiology108, 303-12.
Frensch J, Hsiao TC, Steudle E. 1996. Changes of water andsolute transport along developing maize roots. Planta 198,348-55.
Frensch J, Steudle E. 1989. Axial and radial hydraulic resistanceto roots of maize. Plant Physiology 91, 719-26.
Green PB, Erikson RO, Buggy J. 1971. Metabolic and physicalcontrol of cell elongation rate—in vivo studies in Nitella.Plant Physiology 47, 423-30.
HSussling M, Joins CA, Lehmbecker G, Hecht-Buchholz C,Marschner H. 1988. Ion and water uptake in relation to rootdevelopment in Norway spruce (Picea abies (L.) Karst.).Journal of Plant Physiology 133, 486-91.
Hohl M, Schopfer P. 1992. Physical extensibility of maizecoleoptile cell walls: apparent plastic extensibility is due toelastic hysteresis. Planta 92, 498-504.
Holbrook NM, Burns MJ, Field CB. 1995. Negative xylempressure in plants: a test of the balancing pressure technique.Science 270, 1193-4.
Hsiao TC. 1973. Plant responses to water stress. Annual Reviewof Plant Physiology 24, 519-70.
Hsiao TC, Jing J. 1987. Leaf and root expansive growth inresponse to water deficits. In: Cosgrove DJ, Knievel DP, eds.
Physiology of cell expansion during plant growth. TheAmerican Society of Plant Physiologists, 180-92.
Kramer PJ, Boyer JS. 1995. Water relations of plants and soils.San Diego: Academic Press.
Kutscbera U. 1995. Tissue pressure and cell turgor in axial plantorgans: Implications for the organismal theory of multicellul-arity. Journal of Plant Physiology 146, 126-32.
Landsberg JJ, Fowkes ND. 1978. Water movement throughplant roots. Annals of Botany 42, 493-508.
Lockhart JA. 1965. An analysis of irreversible plant cellelongation. Journal of Theoretical Biology 8, 264-76.
Magnani F, Centritto M, Grace J. 1996. Measurement ofapoplasmic and cell-to-cell components of root hydraulicconductance by a pressure-clamp technique. Planta 199,296-306.
Matsuda K, Riazi A. 1981. Stress-induced osmotic adjustmentin growing regions of barley leaves. Plant Physiology68,571-6.
Matyssek R, Mamyama S, Boyer JS. 1988. Rapid wallrelaxation in elongating tissues. Plant Physiology 86, 1163-7.
McCully ME. 1995. How do real roots work? Some new viewsof root structure. Plant Physiology 109, 1-6.
McCully ME, Canny MJ. 1988 Pathways and processes ofwater and nutrient movement in roots. Plant and Soil111, 159-70.
Melchior W, Steudle E. 1993. Water transport in onion (Alliumcepa L.) roots. Changes of axial and radial hydraulicconductivities during root development. Plant Physiology101, 1305-15.
Mesbcheryakov A, Steudle E, Komor E. 1992. Gradients ofturgor, osmotic pressure, and water potential in the cortex ofthe hypocotyl of growing Ricinus seedlings. Effects of thesupply of water from the xylem and of solutes fromthe phloem. Plant Physiology 98, 840-52.
Michelena VA, Boyer JS. 1982. Complete turgor maintenanceat low water potentials in the elongating region of maizeleaves. Plant Physiology 69, 1145-9.
Mizuno A, Nakahori K, Katou K. 1993. Acid-induced changesin in vivo wall yielding properties of hypocotyl sections ofVigna unguiculata. Physiologia Plantarum 89, 693-8.
Munns R. 1988. Why measure osmotic adjustment? AustralianJournal of Plant Physiology 15, 717-26.
Nonami H, Boyer JS. 1993. Direct demonstration of a growth-induced water potential gradient. Plant Physiology 102, 13-19.
Okamoto H, Liu Q, Nakahori K, Katou K. 1989. A pressure-jump method as a new tool in growth physiology formonitoring physiological wall extensibility and effectiveturgor. Plant and Cell Physiology 30, 979-85.
Okamoto H, Okamoto A. 1994. The pH-dependent yieldthreshold of the cell wall in a glycerinated hollow cylinder (invitro system) of cowpea hypocotyl. Plant, Cell andEnvironment 17, 979-83.
Oparka KJ, Prior DAM, Wright KM. 1995. Symplasticcommunication between primary and developing lateral rootsof Arabidopsis thaliana. Journal of Experimental Botany46, 187-97.
Oparka KJ, Wright KM. 1988. Influence of cell turgor onsucrose partitioning in potato tuber storage tissues. Planta175, 520-6.
Ortega JKE. 1985. Augmented growth equation for cell wallexpansion. Plant Physiology 79, 318-20.
Passioura JB. 1988. Water transport in and to roots. AnnualReview of Plant Physiology 39, 245-65.
Passioura JB, Fry SC. 1992. Turgor and cell expansion: beyondthe Lockhart equation. Australian Journal of Plant Physiology19, 565-76.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/5/985/542707 by guest on 10 January 2022

Patrick JW. 1994. Turgor-dependent unloading of photo-synthates from coats of developing seed of Phaseolus vulgarisand Viciafaba. Turgor homeostasis and set points. PhvswlogiaPlantarum 90, 367-77.
Peterson CA, Steudle E. 1993. Lateral hydraulic conductivity ofearly metaxylem vessels in Zea mays L. roots. Planta189, 288-97.
Pockman WT, Sperry JS, O'Leary JW. 1995. Sustained andsignificant negative water pressure in xylem. Nature 378,715-16.
Pritchard J, Barlow PW, Adam JS, Tomos AD. 1990. Biophysicsof the inhibition of the growth of maize roots by loweredtemperature. Plant Physiology 93, 222-30.
Pritchard J, Hetherington PR, Fry SC, Tomos AD. 1993.Xyloglucan endotransglycosylase activity, microfibril orienta-tion and the profiles of cell wall properties along growingregions of maize roots. Journal of Experimental Botany44, 1281-9.
Pritchard J, Wyn Jones RG, Tomos AD. 1991. Turgor, growthand rheological gradients of wheat roots following osmoticstress. Journal of Experimental Botany 42, 1043-9.
Rail W. 1987. Neuron, cable properties. In: Adelman G, ed.Encyclopedia of Neuroscience. Boston: BirkhSuser Inc.,816-20.
Sanderson J. 1983. Water uptake by different regions of thebarley root. Pathways of radial flow in relation to developmentof the endodermis. Journal of Experimental Botany 34, 240-53.
Schulz A. 1995. Plasmodesmal widening accompanies the shortterm increase in symplasmic phloem unloading in pea roottips under osmotic stress. Protoplasma 188, 22-37.
Schulze E-D. 1986. Carbon dioxide and water vapor exchangein response to drought in the atmosphere and in the soil.Annual Review of Plant Physiology 37, 247-74.
Serpe MD, Matthews MA. 1992. Rapid changes in cell wallyielding of elongating Begonia argenteo-guttata L. leaves inresponse to changes in plant water status. Plant Phvsiologv100, 1852-7.
Shackel KA, Matthews MA, Morrison JC. 1987. Dynamicrelation between expansion and cellular turgor in growinggrape (Vitis vinifera L.) leaves. Plant Physiology 84, 1166-71.
Sharp RE, Hsiao TC, Silk WK. 1990. Growth of the maizeprimary root at low water potentials. II. The role of growthand deposition of hexose and potassium in osmotic adjust-ment. Plant Physiology 93, 1337^6.
Sharp RE, Silk WK, Hsiao TC. 1988. Growth of the maizeprimary root at low water potentials. I. Spatial distributionof expansive growth. Plant Physiology 87, 50-7.
Silk WK. 1984. Quantitative descriptions of development.Annual Review of Plant Physiology 35, 479-518.
Silk WK, Wagner KK. 1980. Growth-sustaining water potentialdistributions in the primary corn root. A noncompartmentedcontinuum model. Plant Physiology 66, 859-63.
Root elongation and drought 999
Slovik S, Daeter W, Hartung W. 1995. Compartmental redis-tribution and long-distance transport of abscisic acid (ABA)in plants as influenced by environmental changes in therhizosphere—a biomathematical model. Journal ofExperimental Botany 46, 881-94.
Spollen WG, Sharp RE. 1991. Spatial distribution of turgor androot growth at low water potentials. Plant Physiology96, 438-43.
Steudle E. 1989. Water flow in plants and its coupling to otherprocesses: an overview. In: Fleischer S, Fleischer R, eds.Methods in enzymology, No. 174. Biomembranes, Part U:Cellular and subcellular transport: Eukaryotic (non-epithelial)cells. London: Academic Press, 183-225.
Steudle E. 1992. The biophysics of plant water: compart -mentation, coupling with metabolic processes, and flow ofwater in plant roots. In: Somero GN, Osmond CB, Bolis CL,eds. Water and life: comparative analysis of water relationshipsat the organismic, cellular, and molecular levels. Heidelberg,Germany: Springer-Verlag, 173-204.
Steudle E. 1994. Water transport across roots. Plant and Soil167, 79-90.
Steudle E. 1995. Trees under tension. Nature 378, 663-4.Steudle E, Frensch J. 1989. Osmotic responses of maize roots:
water and solute relations. Planta 177, 281-95.Steudle E, Frensch J. 1996. Water transport in plants: role of
the apoplast. Plant and Soil, 187, 67-79.Tardieu F, Davies WJ. 1993. Integration of hydraulic and
chemical signalling in the control of stomatal conductanceand water status of droughted plants. Plant, Cell andEnvironment 16, 341-9.
Taylor RE. 1963. Cable theory. In: Nastuk WL, ed. Physicaltechniques in biological research. IV. Electrophysiologicalmethods, Part B. New York: Academic Press, 219-62.
Tomos AD, Malone M, Pritchard J. 1989. The biophysics ofdifferential growth. Environmental and Experimental Botany29, 7-23.
Tomos AD, Pritchard J. 1994. Biophysical and biochemicalcontrol of cell expamsion in roots and leaves. Journal ofExperimental Botany 45, 1721-31.
Waisel Y, Eshel A, Kalkafi U. 1991. Plant roots: the hidden half.New York: Marcel Decker.
Zhu GL, Steudle E. 1991. Water transport across maize roots.Simultaneous measurement of flows at the cell and root levelby double pressure probe technique. Plant Physiology 95,305-15.
Zimmcrmann U, Haase A, Langbein D, Meinzer F. 1993.Mechanisms of long-distance water transport in plants: are-examination of some paradigms in the light of newevidence. Philosophical Transaction of the Royal Society ofLondon, Series B 341, 19-31.
Dow
nloaded from https://academ
ic.oup.com/jxb/article/48/5/985/542707 by guest on 10 January 2022




















