
Macaque monkeys learn by observation in the ghost display ...
Upload
independentCategory
view
1download
0
Znt. J . Cancer: 41, 60-608 (1988) Publication of the International Union Against Cancer Publication de I’Union Internationale Contre le Cancer @ 1988 Alan R. Liss, Inc.
PREVALENCE OF ANTIBODIES TO 3 RETROVIRUSES IN A CAPTIVE COLONY OF MACAQUE MONKEYS Muthiah D. DANIEL, Norman L. LETVM, Prabhat K. SEHGAL, Diane K. SCHMIDT, Daniel P. SILVA, Keith R. SOLOMON, F. Stephen HODI, JR., Douglas J. RINGLER, Ronald D. HUNT, Norval W. KING and Ronald C. DESROSIERS New England Regional Primate Research Center, Harvard Medical School, Southborough, MA 01 772.
The prevalence of antibodies to 3 retroviruses in the ma- caque colony of the New England Regional Primate Research Center (NERPRC) was determined using enzyme-linked im- munosorbent assay procedures as well as radioimmunoprecip- itation-SDS polyacrylamide gel electrophoresis and indirect immunofluorescence tests. Out of 848 macaques, 3 (0.35%) had antibodies to simian immunodeficiency virus (SIV), 27 (3.2%) had antibodies to simian T-lymphotropic virus type I (STLV-I) and approximately 285 (34%) had antibodies to type D retrovirus. Of 3 macaques infected with SlV, 2 were rhesus monkeys (Macaca mulatta) and I was a cynomolgus monkey (Macaca fwcicularis). STLV-I and D retrovirus infection oc- curred in all 4 macaque species examined. SlV, STLV-I and D retroviruses were isolated from sero-positive macaques. The low prevalence of SIV infection suggests that SIV is not being readily transmitted among macaques at NERPRC; this con- trasts markedly with the high SIV prevalence in some captive mangabey colonies. In contrast to African green monkeys from eastern Africa, 160 Caribbean green monkeys examined showed no sign of SIV infection. These results provide a framework for monitoring spontaneous disease associated with infection by these 3 retroviruses and will help in further definition of STLV-I and SIV infection of non-human primates as animal models for human disease.
Four groups of retroviruses are now known to infect ma- caques (genus Macaca), primates of Asian origin: foamy vi- rus, type-D retrovirus, simian T-lymphotropic virus type I (STLV-I), and simian immunodeficiency virus (SIV). Exten- sive natural infection of macaque monkeys with foamy virus has been known for some time (Rustigian et al . , 1955; Hooks and Gibbs, 1975). However, natural transmission of the other 3 groups of retroviruses among macaques has only become recognized in the last few years. Although Mason-Pfizer mon- key virus, the prototype of the D-type retroviruses, was origi- nally isolated from a macaque by Chopra and Mason (1970), it remained the only macaque type-D retrovirus available for study until 1984 (Daniel et al., 1984; Marx et al., 1984). Since that time, the potential for extensive transmission of D-type retroviruses in macaque colonies has become generally recog- nized. STLV-I and SIV are monkey counterparts of the human viruses HTLV-I and HIV. The prevalence of STLV-I infection has been reported previously for macaques in the wild and in captive colonies in Japan (Miyoshi et al., 1982; Hayami et al., 1984). STLV-I has about 90% sequence identity with HTLV-I (Watanabe et al . , 1985, 1986). SIV was first isolated from captive rhesus macaques at NERPRC (Daniel et al., 1985); SIV is also related in sequence to its human counterpart HIV (Kanki et al., 198%; Hirsch et al., 1986). Between 30% and 50% of African green monkeys (AGMs) in eastern Africa appear to be SIV-seropositive (Kanki et al., 1985a) and SIV has now also been isolated from the mangabey, another Afri- can primate (Fultz et al . , 1986; Murphey-Corb et al., 1986). However, other data on SIV prevalence are only beginning to emerge.
It is important to monitor the extent of infection with these 3 groups of retroviruses in captive macaque colonies for at least 3 reasons. First, these retroviruses can be responsible for naturally occurring disease in this precious resource. Only through such monitoring can infections be traced and disease
associations drawn with a view to taking appropriate preven- tive actions. Second, 2 of these viruses (STLV-I and SIV) provide excellent models for retroviral diseases of humans, adult T-cell leukemia (ATL) and acquired immunodeficiency syndrome (AIDS). More detailed descriptions of modes of transmission, the extents of infection and the effects of these infections are needed to provide a baseline for continued study of these primate models. Third, macaques are often used in a variety of research studies; it is important to know that infec- tion with one or more of these viruses has not confounded the interpretation of research results.
In this study, we report the extents of infection with SIV, STLV-I and D-type retrovirus in the captive macaque colony at the NERPRC and the isolation of these viruses from sero- positive animals.
MATERIAL AND METHODS
Animals Four species of macaque are cared for at NERPRC: Macaca
mulatta (rhesus monkey); Macaca fascicularis (cynomolgus monkey); Macaca cyclopis (Taiwanese rock macaque); and Macaca arctoides (stump-tailed macaque). These macaques are housed in individual cages, in pairs for breeding purposes or in groups containing 10-30 animals according to DHSS specifications (NIH, 1985). Most of these macaques were born in captivity at NERPRC, some were obtained from other captive colonies and some, particularly M. fascicularis, were captured in the wild. Serum collection
Macaques were anesthetized by intramuscular inoculation of Ketamine hydrochloride (Ketaset, Bristol Myers, Syracuse, NY) at 10-20 mg per kg. Five to 10 ml of blood were drawn into vacutainer SST tubes (Becton Dickinson, Mountain View, CA). Serum was transferred from these tubes following cen- trifugation into screw-cap glass vials and aliquots were stored at -70°C and 4°C. Macaque serum was collected between November 14, 1985 and February 6, 1986.
Sera from AGMs were obtained from a supplier who re- cently imported these animals from eastern Africa (Kenya and Ethiopia). Sera from Caribbean AGMs were obtained from a pharmaceutical company which purchased animals recently captured in Barbados. Cells and viruses
Strain 225 of retrovirus D/New England (D/NE) was pro- duced in Raji cells (Daniel et al . , 1984; Desrosiers et a l . , 1985). The M10P16 cell line from a Macaca radiata, kindly provided by Dr. G. Hunsmann and Dr. M. Hayami, was used as a source of macaque STLV-I (Tsujimoto et al . , 1985; Jurkiewicz et a l . , 1986). The 251 and 239 strains of SIV originally isolated from macaques at NERPRC were produced in HUT-78 cells (Daniel et al., 1985). Uninfected Raji and HUT-78 cells were added to the DINE and SIV cultures
Received: July 14, 1987 and in revised form Septembcr 26, 1987.

602 DANIEL ET AL.
TABLE I - INITIAL SCREENING OF 848 MACAQUE SERA BY ELISA FOR ANTIBODIES TO 3 RETROVIRUSES
Number of sera producing absorbance values of 0.0-0.1 0.1-0.2 0.2-0.3 0.3-0.4 0.4-0.5 0.5-0.6 0.6-0.7 0.7-0.8 0.8-0.9 0.9-1.0 >1.0
D/NE 392 94 42 18 17 17 21 25 21 21 1 80 SIV 764 53 19 2 1 0 3 2 1 I 2 STLV-I 682 97 14 8 7 3 4 4 2 6 21
respectively every 7-14 days in approximately equal numbers to maintain maximal reverse transcriptase activity in the cell- free supernatant. These lymphoid cell lines, as well as other lymphoid cell lines, were grown in RPMI 1640 containing 10% fetal calf serum, penicillin and streptomycin in stationary suspension cultures. Other cell lines that were used include the HTLV-I-producing human cell lines MT-2 and TCL-KAN; the ROD isolate of LAV-2 (Clavel et al., 1986) was produced in CEM cells. Virus pur$cation
Cells for virus production were grown in 600 ml T-150 plastic cell culture flasks in volumes of 200 ml per flask. Cells were harvested 48-72 hr after splitting by centrifugation at 1,500 g for 15 min and the supernatants were filtered through disposable 0.45 p filters to remove all traces of cells and cell debris. Virus contained in the filtered cell-free supernatant was pelleted by centrifugation in a Beckman type-19 rotor at 48,000 g at 4°C for 3 hr. Following removal of the supernatant and complete drainage, virus pellets from 1.5 1 contained in 6 type-19 bottles were resuspended in 1.0 ml of TNE (0.02M Tris HCl, 0 . 1 ~ NaCl, 0 . 0 0 1 ~ EDTA, PH 7.5) and stored at 4°C overnight. Virus was then further purified by Sepharose 4B column chromatography in a biosafety cabinet using TNE buffer (McGrath et al., 1978). Virus contained in the void volume was pelleted at 4°C by micro-centrifugation using 1.5- ml screw-cap Sarstedt tubes. Virus from 1.5 1 of starting culture was finally resuspended in 150 pI of phosphate-buff- ered saline (PBS calcium- and magnesium-free) containing 0.5% Triton X-100 to yield 10,OOOX virus. Purified 10,OOOX virus was stored at -70°C until use. Coating of plates with antigen
To coat plates with virus antigen, purified 10,OOO X virus in Triton X-100-PBS was diluted 1:1,000 with PBS, and 100 p1 were added to each well of 96-well Nunc (Roskbilde, Den- mark) immunoplates. After overnight incubation at room tem- perature, liquid was removed from wells by inversion, the wells were rinsed with distilled water by immersion and the water was removed by inversion. Unbound sites were blocked by adding 100 p1 of 0.3 % BSA (Kirkegaard and Perry, Gaith- ersburg, MD) in PBS to each well and incubating approxi- mately 20 min. Wells were washed again as above and stored at 4°C in a humidified sealed bag until use.
To coat plates with uninfected cell antigens for preadsorp- tion, 400 ml of uninfected cells were first pelleted and then resuspended in 5 ml PBS. Following 2 repeated freeze-thaws in a dry-ice-ethanol bath, the cell lysate was clarified by centrifugation at 1,OOO g for 2 min. The supernatant was diluted to 150 ml with PBS and used to coat wells of Nunc immunoplates as described above.
Plates for HTLV-I ELISA were obtained commercially from Biotech, Dupont, Billerica, MA. Enzyme-linked irnrnunosorbent assay (ELISA)
For initial ELISA determination of binding antibodies to virus antigens by ELISA, 200 pl of serum diluent (PBS con- taining 0.05% Tween 20, 5% fetal calf serum and 5% goat serum) was added to each virus antigen well followed by 10 pl
monkey test serum. Monkey test serum was omitted from wells used to blank the ELISA plate reader. Following a 1-hr incubation at room temperature, the fluid was discarded and the wells were rinsed with PBS containing 0.05% Tween 20. The rinsing was repeated twice and the plates were tapped dry. Binding antibodies were reacted with affinity-purified goat anti-human IgG conjugated with alkaline phosphatase and the color was developed using P-nitrophenylphosphate according to the manufacturer’s recommendations. Absorbance at 410 nm was recorded with a Dynatech (Cambridge, MA) ELISA reader.
Additional testing of selected sera by ELISA employed a serum pre-adsorption step. Serum diluent and monkey test serum were first added to plates coated with appropriate unin- fected cell antigens (Raji cells for D virus determinations and HUT-78 cells for SIV and STLV-I determinations). Following a 1-hr incubation at room temperature, 200 pi were then transferred to the appropriate virus antigen plates and pro- cessed as described above.
Radioimmunoprecipitation assay-SDS polyacrylamide gel electrophoresis (RIPA)
H9 or SIV-infected H9 cells were radiolabelled with 35S methionine (150 uCi/ml, NEN. Boston, MA) for 4 hr. The labelled cells were resuspended in lysis buffer (1 % Triton X- 100, 0.05 M Tris-HC1 buffer, PH 7.2, 0.15 M NaCl, h M EDTA, 1% aprotinin, and 1 mM phenylmethylsulfonyl fluo- ride) and cleared by centrifugation for 1 hr at 100,000 g. Macaque plasma, 10-20 pI, was incubated with protein-A- sepharose CL4B (Sigma, St. Louis, MO) and, after washing, antibody-coated beads were incubated with the cell lysate. After further washing, immunoprecipitates were eluted from the protein-A-sepharose beads with sample buffer [5% 2- mercaptoethanol, 2% sodium dodecyl sulfate (SDS), 0.08 M Tris-HC1, PH 6.8, 10% glycerol, and 0.001% bromophenol blue] by boiling at 100°C for 3 min. Samples were analyzed by electrophoresis in 10% SDS-polyacrylamide gels with a 3.5% stacking gel according to the discontinuous buffer sys- tem of Laemmli (1970). Indirect imrnunofluorescence
Infected lymphoid cells were smeared on a glass slide (Carl- son Scientific, Peotone, IL), within 24 hr of splitting, air-dried at room temperature for 1-2 hr and fixed in acetone for 7 min. The fixed slides were used immediately or stored at -20°C for future use. Fixed cells on the slides were incubated with 1 : 10 dilutions of sera for 18 hr at 4°C or 30 min at 37°C and then rinsed 3 times with PBS. Fluorescein isothiocyanate (F1TC)-conjugated goat anti-human IgG (Cappel, Cochran- ville, PA) was applied for 30 min at 37°C and then the slides were rinsed again as above. Cells were counterstained for 5 min with 0.06% Evans’ blue and examined with a Leitz flu- orescent microscope. Virus isolations
Type-D virus isolations were performed by co-cultivation of macaque peripheral blood mononuclear cells (PBMC) with Raji cells (Daniel et al., 1984; Desrosiers et al., 1985). SIV isolations were performed by co-cultivation of macaque PBMC
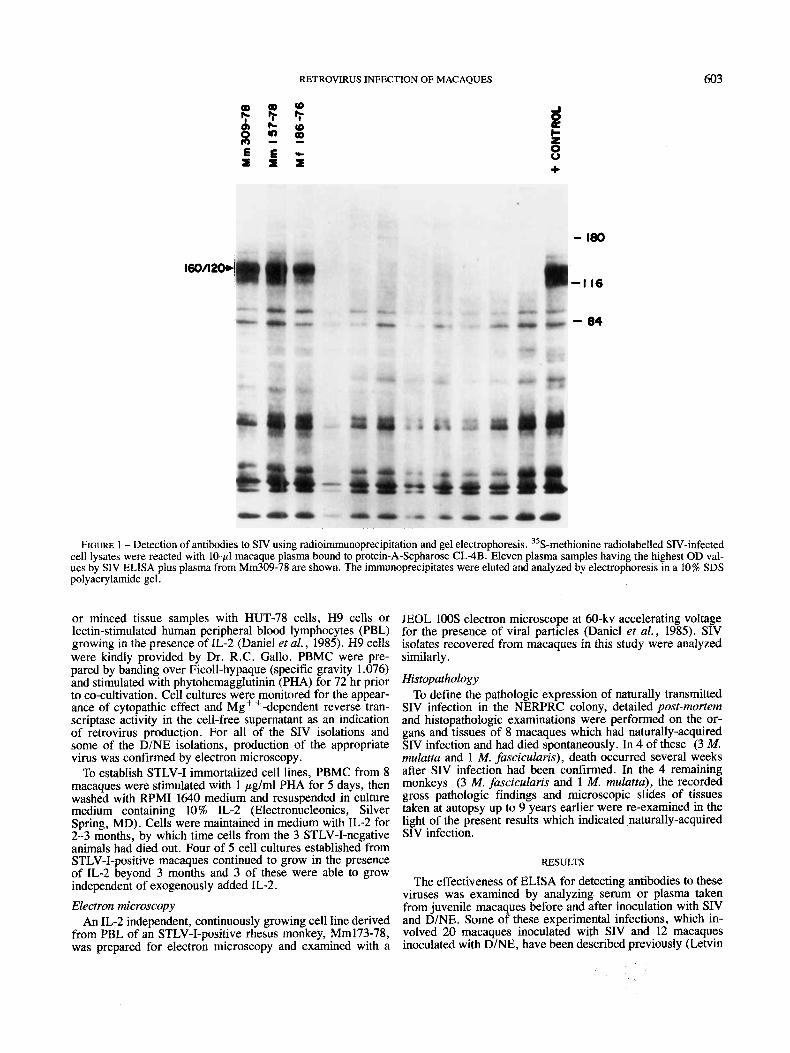
RETROVIRUS INFECTION OF MACAQUES 603
+
FIGURE 1 - Detection of antibodies to SIV using radioimmunoprecipitation and gel electrophoresis. 35S-methionine radiolabelled SIV-infected cell lysates were reacted with 10-pl macaque plasma bound to protein-A-Sepharose CL-4B. Eleven plasma samples having the highest OD val- ues by SIV ELISA plus plasma from Mm309-78 are shown. The immunoprecipitates were eluted and analyzed by electrophoresis in a 10% SDS polyacrylamide gel.
or minced tissue samples with HUT-78 cells, H9 cells or lectin-stimulated human peripheral blood lymphocytes (PBL) growing in the presence of IL-2 (Daniel et al., 1985). H9 cells were kindly provided by Dr. R.C. Gallo. PBMC were pre- pared by banding over Ficoll-hypaque (specific gravity 1.076) and stimulated with phytohemagglutinin (PHA) for 72 hr prior to co-cultivation. Cell cultures were monitored for the appear- ance of cytopathic effect and Mg + +-dependent reverse tran- scriptase activity in the cell-free supernatant as an indication of retrovirus production. For all of the SIV isolations and some of the D/NE isolations, production of the appropriate virus was confirmed by electron microscopy.
To establish STLV-I immortalized cell lines, PBMC from 8 macaques were stimulated with 1 pg/ml PHA for 5 days, then washed with RPMI 1640 medium and resuspended in culture medium containing 10 % IL-2 (Electronucleonics, Silver Spring, MD). Cells were maintained in medium with IL-2 for 2-3 months, by which time cells from the 3 STLV-I-negative animals had died out. Four of 5 cell cultures established from STLV-I-positive macaques continued to grow in the presence of IL-2 beyond 3 months and 3 of these were able to grow independent of exogenously added IL-2. Electron microscopy
An IL-2 independent, continuously growing cell line derived from PBL of an STLV-I-positive rhesus monkey, Mm173-78, was prepared for electron microscopy and examined with a
JEOL 100s electron microscope at 60-kv accelerating voltage for the presence of viral particles (Daniel et al . , 1985). SIV isolates recovered from macaques in this study were analyzed similarly.
Histopathology To define the pathologic expression of naturally transmitted
SIV infection in the NERPRC colony, detailed post-mortem and histopathologic examinations were performed on the or- gans and tissues of 8 macaques which had naturally-acquired SIV infection and had died spontaneously. In 4 of these (3 M. mulatta and 1 M. fascicularis), death occurred several weeks after SIV infection had been confirmed. In the 4 remaining monkeys (3 M . fascicularis and 1 M. mulatta), the recorded gross pathologic findings and microscopic slides of tissues taken at autopsy up to 9 years earlier were re-examined in the light of the present results which indicated naturally-acquired SIV infection.
RESULTS
The effectiveness of ELISA for detecting antibodies to these viruses was examined by analyzing serum or plasma taken from juvenile macaques before and after inoculation with SIV and D/NE. Some of these experimental infections, which in- volved 20 macaques inoculated with SIV and 12 macaques inoculated with D/NE, have been described previously (Letvin
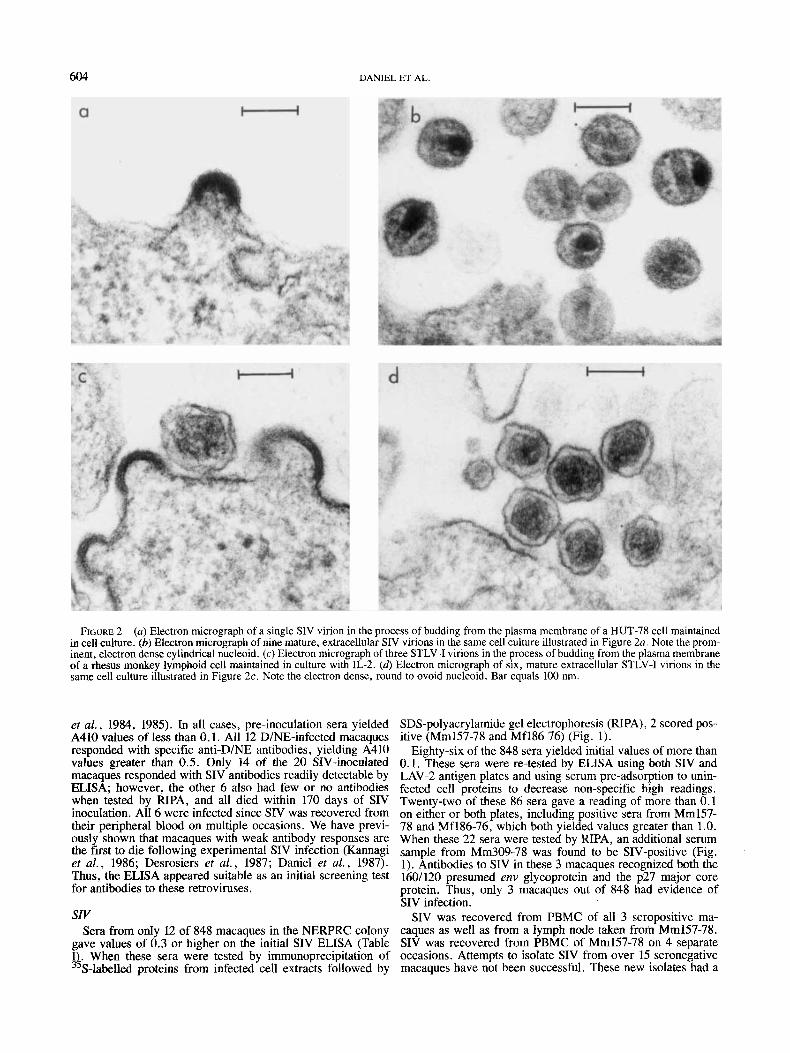
604 DANIEL ET AL.
FIGURE 2 - (a) Electron micrograph of a single SIV virion in the process of budding from the plasma membrane of a HUT-78 cell maintained in cell culture. (b) Electron micrograph of nine mature, extracellular SIV virions in the same cell culture illustrated in Figure 2a. Note the prom- inent, electron dense cylindrical nucleoid. (c) Electron micrograph of three STLV-I virions in the process of budding from the plasma membrane of a rhesus monkey lymphoid cell maintained in culture with IL-2. (4 Electron micrograph of six, mature extracellular STLV-I virions in the same cell culture illustrated in Figure 2c. Note the electron dense, round to ovoid nucleoid. Bar equals 100 nm.
et al., 1984, 1985). In all cases, pre-inoculation sera yielded SDS-polyacrylamide gel electrophoresis (RIPA), 2 scored pos- A410 values of less than 0.1. All 12 D/NE-infected macaques itive (Mm157-78 and Mf186-76) (Fig. 1). responded with specific anti-D/NE antibodies, yielding A410 Eighty-six of the 848 Sera yielded initial values of more than values greater than 0.5. Only 14 of the 20 SIV-inoculated 0.1. These sera were re-tested by ELISA using both SIV and macaques responded with SIV antibodies readily detectable by LAV-2 antigen plates and using serum pre-adsorption to unin- ELISA; however, the other 6 also had few or no antibodies fected cell proteins to decrease non-specific high readings. when tested by RIPA, and all died within 170 days of SIV Twenty-two of these 86 sera gave a reading of more than 0.1 inoculation. All 6 were infected since SIV was recovered from on either or both plates, including positive sera from Mm157- their peripheral blood on multiple occasions. We have previ- 78 and Mf186-76, which both yielded values greater than 1.0. ously shown that macaques with weak antibody responses are When these 22 sera were tested by RIPA, an additional serum the first to die following experimental SIV infection (Kannagi sample from Mm309-78 was found to be SIV-positive (Fig. et al., 1986; Desrosiers et d. , 1987; Daniel et a[. , 1987). 1). Antibodies to SIV in these 3 macaques recognized both the Thus, the ELISA appeared suitable as an initial screening test 160/120 presumed env glycoprotein and the p27 major core for antibodies to these retroviruses. protein. Thus, only 3 macaques out of 848 had evidence of
SIV infection. szv SIV was recovered from PBMC of all 3 seropositive ma-
Sera from only 12 of 848 macaques in the NERPRC colony caques as well as from a lymph node taken from Mm157-78. gave values of 0.3 or higher on the initial SIV ELISA (Table SIV was recovered from PBMC of Mm157-78 on 4 separate I& When these sera were tested by immunoprecipitation of occasions. Attempts to isolate SIV from over 15 seronegative
S-labelled proteins from infected cell extracts followed by macaques have not been successful. These new isolates had a

RETROVIRUS INFEC
Mm 91-78 Mm44-84
200 -
+ 160/120 I 0 6 -
71 -
.c 5 5
44 -
23 -
18- . . du-, -
4 2 7
FIGURE 3 - Demonstration of anti-SIV seroconversion of 2 ma- caques using radioimmunoprecipition and gel electrophoresis. Paired lasma samples shown were taken from 2 macaques 17 months apart.
h-methionine-radiolabelled SIV-infected cell lysates were reacted with 10 pl macaque plasma bound to protein A-sepharose CL4B. The immunoprecipitates were eluted and analyzed by electrophoresis in a 10% SDS polyacrylamide gel.
morphogenesis and morphology typical of lentiviruses, includ- ing the human AIDS virus HIV (Gonda et al., 1985) and its monkey counterpart SIV (Daniel et al., 1985); i .e . , a crescent shape upon budding in the manner of type-C retroviruses and a cylindrical or rod-shaped nucleoid in mature particles (Fig. 2a and b). Cells infected with these new SIV isolates reacted positively in indirect immunofluorescence tests with SIV-pos- itive sera but not SIV-negative sera.
Of the 3 SIV-positive macaques, one, Mm157-78, was pur- chased from a primate importer in April 1978; one, Mf186- 76, was born at NERPRC in May 1976 and one, Mm309-78, was obtained in July 1978 from another primate colony in the US. Stored serum samples from Mm157-78 and Mf186-76 taken in January 1979 were negative for SIV antibodies and the mother of Mf186-76 was SIV-negative. Serum from Mm309-78 taken late in 1978 was negative for SIV antibodies;
TION OF MACAQUES 605
serum taken in November 1985 was only weakly positive for SIV antibodies; however, by November 1986, Mm309-78 had become strongly positive. These results indicate that all 3 macaques had become infected while at NERPRC.
Mm157-78 (a female) gave birth in 1983 to Mm142-83, a male rhesus that yielded 1 of our initial 4 SIV isolates in early 1985 (Daniel et al., 1985). Inspection of records and further analyses indicated that Mm142-83 must have been infected in utero. This animal was delivered by cesarean section and reared in our nursery away from the mother. He had constant health problems until his death in 1985. All serum samples from Mm142-83, including those taken 1 !h months after birth, were positive for SIV antibodies.
Mm157-78, Mf186-76 and more recently Mm309-78 have been removed from general contact with the rest of the ma- caques. Mm157-78 and Mf186-76 were placed in small breed- ing groups isolated from the rest of the colony in an effort to continue the study of natural transmission; however, both animals died during the next few months. Mm309-78 also died within 2 months of transfer to this isolated breeding group. We are continuing to monitor the remaining isolated animals for signs of infection as well as the rest of the macaques in order to maintain an SIV-free colony.
The histopathologic findings at autopsy in Mm157-78 in- cluded: generalized cytomegalovirus (CMV) infection, multi- ple bacterial abscesses of the lung, severe lymphocytic depletion of peripheral and visceral lymph nodes and scattered nodular infiltrates of lymphocytes (lymphoproliferative disor- der) in the bone marrow. Mm142-83 (the cesarean-derived offspring of Mm157-78), which was infected in utero, died with a massive, polyclonal proliferation of B lymphocytes (a severe lymphoproliferative disorder) previously described in detail by Chalifoux et al. (1986). Mm142-83 was also infected with type-D retrovirus. Mm309-78 had hepatobiliary crypto- sporidiosis, splenic amyloidosis, purulent necrotizing gas- troenteritis and necrotizing ulcerative gingivitis. This animal was probably also infected with type-D retrovirus as well. In Mf186-76, the post mortem findings consisted of enteric sal- monellosis, granulomatous hepatitis, generalized lymphoid de- pletion and mononuclear myocarditis of undetermined etiology.
Among approximately 300 previously stored NERPRC ma- caque sera tested, we found 4 additional SIV seropositives; all 4 of the animal donors were dead at the time of testing. Of these, 3 were M. fascicularis and 1 a M. mulatta. None had been inoculated with any fluids or tissue materials as part of any study and thus must have been infected naturally. A retrospective examination of microscopic slides prepared from the tissues taken at autopsy of these animals revealed the following pathologic findings. Mt222-73 died of generalized malignant lymphoma, CMV infection and an unusual giant- cell lymphadenopathy. Mfll4-73, a juvenile, had profound thymic atrophy and cystic Hassal’s corpuscles, giant-cell lymphadenopathy involving the visceral lymph nodes similar to that observed in Mf222-73 and lymphocytic depletion of peripheral lymph nodes. Mf21-77 died of a diffuse bacterial pneumonia, the etiology of which was not identified, and also had marked lymphocytic depletion of lymph nodes and spleen. The only rhesus in this group, Mm425-75, had a localized necrotizing gingivitis and intestinal cryptosporidiosis.
In early 1987, 17 months after obtaining serum samples from NERPRC macaques for the present survey, we again obtained sera from the NERPRC macaques. Sera from 2 female ma- caques showed antibodies to SIV (Fig. 3). Comparison with sera from these same 2 macaques taken 17 months earlier revealed that both animals had seroconverted during the year.
We have also analyzed sera from 160 AGMs of Caribbean origin and 50 AGMs from eastern Africa. In both groups, serum samples were obtained during the process of importa-

606 DANIEL ET AL.
TABLE I1 - PREVALENCE AND SPECIES PREDILECTION FOR SIV, STLV-I AND D/NE INFECTIONS
Number of antibody-positive sera (%) SIV' STLV-I~ DINE' Species
M . arctoides 0/28 (0%) 1/28 (3.6%) 14/28 (50%) M. cyclopis 0148 (0%) 1/48 (2.1%) 25/48 (52%) M. fascicularis 1/333 (0.3%) 15/333 (4.5%) 1431333 (43%) M. mulatta 2/439 (0.45%) 101439 (2.3%) 1031439 (24%) Total 3/848 (0.35%) 21/848 (3.2%) 2851848 (34%) 'SIV-positive sera are those confirmed by RIPA-SDS. See text.-*STLV-I-positive sera are those confirmed positive by
HTLV-I ELISA and indirect immunofluorescence tests. See text.-'Sera with ELISA A410 greater than 0.5 were considered positive.
tion. ELISA tests using SIV and LAV-2 antigen plates with pre-adsorption were performed with all AGM serum samples and RIPA was performed on selected samples. Approximately 30% of the sera from eastern African AGMs were SIV-sero- positive, confirming an earlier report of high SIV seropositiv- ity in AGMs from this region (Kanki et al., 1985~). None of the 160 Caribbean AGMs was SIV-seropositive. sizv-I
Of 848 sera tested without pre-adsorption, 69 gave readings greater than 0.2 in the initial STLV-I ELISA (Table I). These 69 sera were re-tested by an indirect immunofluorescence (IF) test and by ELISA using a commercial HTLV-I plate. Twenty- seven of these 69 sera were positive for STLV-I antibodies by IF using TCL-KAN and MT-2, HTLV-I-positive indicator cell lines, and 173, an STLV-I-positive cell line established from an NERPRC macaque (see below). All of these 27 sera yielded A410 values greater than 0.2 with a median of 0.641 when tested in the HTLV-I ELISA. These 27 also yielded A410 values greater than 0.2 when retested by STLV-I ELISA, with or without pre-adsorption (medians of 1.540 and 1.690). Only 3 sera that yielded A410 values greater than 0.2 (0.475,0.516, 0.545) in the HTLV-I ELISA scored negative in the IF test. The remaining 39 sera yielded values of less than 0.2 in the HTLV-I ELISA (median 0.023) and all 39 scored negative in the IF test. Most of these 39 also yielded values of less than 0.2 by STLV-I ELISA when preadsorption was used. Fifty- three sera which gave A410 values of less than 0.1 on the original STLV-I ELISA were retested with the commercial HTLV-I ELISA plate; all 53 yielded A410 values of less than 0.1. We conclude from these analyses that 27 of 848 sera (3.2 %) were positive for STLV-I antibodies.
Antibodies to STLV-I were found in all 4 macaque species tested and were most prevalent in M. fascicularis (Table 11). However, 4 of the 15 STLV-I positive M. fascicularis were part of a single shipment of 6 from another US laboratory to NERPRC in 1985.
PBL were obtained from 5 of the 27 STLV-I-positive ma- caques in an attempt to establish STLV-I-immortalized cell lines. Four of these yielded continuously growing cell lines. PBL were also obtained at the same time from 3 STLV-I- negative macaques; none of these 3 yielded continuously growing cell lines.
Of the 4 continuously growing cell lines, 3 were able to grow independently of exogenously added IL-2; these 3 also produced significant reverse transcriptase activity and were stained positively by STLV-I-positive sera in indirect immu- nofluorescence tests. DNA from these 3 cell lines, but not from an uninfected cell line, hybridized specifically to a cloned HTLV-I DNA probe upon Southern blot hybridization (data not shown). Electron microscopy showed that virus produced by the newly established cell line from Mm173-78 had the morphogenesis and morphology expected for STLV-I, i. e . , that of a type-C retrovirus (Fig. 2c and 6).
Four of the 27 STLV-I-positive animals died within the year following sampling for the serological survey. Unfortunately, all 4 animals had been previously assigned to other investiga- tors at the time of their death and, as a consequence, no tissues were available from them for retrospective pathologic evaluation. D-type retrovirus
Taking all DlNE ELISA values over 0.5 as positive, we estimate that approximately 34% of NERPRC macaques are D-positive (Table I); as with STLV-I, type-D retrovirus was more prevalent in M. fascicularis than in M. mulatta (Table 11). This large percentage of D-positive macaques is consistent with our isolation of type-D retrovirus from over 40 NERPRC macaques since 1984 (Desrosiers et al., 1985) and knowledge of its mode of transmission (Lerche et al., 1984).
In the 12 months following this serological survey, type-D retrovirus was isolated from 10 macaques with clinical signs of illness that included diarrhea, lymphadenopathy, necrotiz- ing gingivitis and weight loss. Most of these macaques were negative for IgG antibodies to type-D retrovirus at the time of the original sample survey and at the time of D-retrovirus isolation, which indicates primary type-D retrovirus infection.
DISCUSSION
Our results indicate that 3 of 848 NERPRC macaques (0.35%) are SIV-positive, 27 of 848 (3.2%) are STLV-I posi- tive and approximately 34 % are type-D retrovirus-positive (Table 11).
The low prevalence of SIV infection in the NERPRC ma- caque colony suggests that SIV is not readily transmitted among macaques. We know that Mm157-78 had been infected at least since April 1983 when she gave birth to Mm142-83; since that time she was in contact with numerous other ma- caques, including 5 different males with which she was bred on numerous occasions. The apparently inefficient transmis- sion of SIV among these animals contrasts markedly with the situation in captive mangabey colonies in which more than 80% of the animals are SIV-infected (Fultz et al., 1986; Murphey-Corb et al., 1986).
The disease associations in SIV-infected macaques in this study and in a previous report (Daniel et a l . , 1985) indicate that SIV is often pathogenic during natural transmission among macaques. The constellation of specific and non-specific path- ologic findings in the 8 macaques dying with naturally-ac- quired SIV infection is similar in many respects to the original descriptions of the naturally occurring acquired immunodefi- ciency syndrome at NERPRC (King et al . , 1983). The findings of King et al. (1983), based upon a study of only 16 macaques, were published prior to the isoiation of either the type-D retrovirus or SIV from macaques at NERPRC . Since a number of these pathologic findings are also found in animals dying from type-D retrovirus infection, SIV and/or type-D retrovirus

may have contributed to the spontaneous disease described by King et al. (1983). Giant-cell lymphadenopathy and giant-cell pneumonia were encountered in the 8 SIV-infected animals described here but these were not described in King et al. (1983). Interestingly, giant-cell lymphadenopathy and giant- cell pneumonia are now being recognized in macaques infected experimentally with SIV at the NERPRC (data not shown). A striking difference, however, between the naturally and exper- imentally SIV-infected macaques at NERPRC is the lack of any SIV-induced encephalopathy in the naturally infected animals.
The natural habitat of green monkeys (Cercopithecus ae- thiops) is obviously Africa. However, in the 17th and 18th centuries, sailors involved in the slave trade often brought African green monkeys with them to the New World; since that time, free-roaming troops of African green monkeys have become established on 3 islands in the Caribbean: Barbados, St. Kitts and Nevis (Denham, 1981). Caribbean AGMs have been shown to be STLV-I seronegative (Blakeslee et al., 1985). In this report, we found no antibodies to SIV in any of the 160 Caribbean AGMs from Barbados examined. We have also confirmed the high SIV seropositivity of AGMs from eastern Africa. It is tempting to conclude from these studies that SIV was introduced into the African AGM population at some time since the 17th century. However, such a conclusion is not justified for at least 2 reasons. First, it is not known how complex the genetic stock was that gave rise to the free- roaming AGMs present in the Caribbean area today. It is possible that all Caribbean AGMs are descendants of only a few animals, and these could all have been SIV-negative. Second, it is not known from what region of Africa the AGMs that populated these Caribbean islands came. There possibly could be a great variation in SIV prevalence among AGMs in different regions of Africa.
It is interesting that some macaque colonies, such as that at NERPRC , have extensive D-retrovirus infections while other colonies, such as those at the Delta Regional Primate Research Center in Louisiana and the Yerkes Regional Primate Research
Center in Georgia, apparently have little or no D-retrovirus infection (McClure, Fultz and Marx, personal communication; Murphey-Corb and Marx, personal communication). These latter colonies contain several thousand macaques and no spe- cific measures have been taken to keep them free of D-type retrovirus. These results suggest that type-D retrovirus may have been introduced into captive M. mulatta from rare sero- positive animals or perhaps from some other species. Previ- ously we have noted the absence of restriction fragment polymorphisms in different isolates of D-retrovirus from NERPRC macaques (Desrosiers et al., 1985); this finding is consistent with the above hypothesis.
There has been only a single documented case of a sponta- neous adult T-cell leukemia (ATL)-like disease in a monkey naturally infected with STLV-I (Sakakibara et al., 1986). This may not be surprising since the lifetime chance of contracting ATL in HTLV-I-positive humans has been estimated at 1 in 10,OOO. Also, attempts to induce ATL experimentally in pri- mates have not been successful (Nakamura et al., 1986). Knowledge of which macaques in our colony are STLV-I positive will allow us to look carefully for diseases resembling ATL or spastic paraparesis (Gessain et al., 1985) in these animals. STLV-I-immortalized cell lines can also be used in attempts to induce leukemia in macaques as models for the human ATL disease.
ACKNOWLEDGEMENTS
This work was supported by the Public Health Service and by the Division of Research Resources. Research was also sponsored in part by the National Cancer Institute, DHHS, under contract with Program Resources, Inc. and by the Mas- sachusetts Department of Public Health. The contents of this publication do not necessarily reflect the views or policies of the DHHS, nor does mention of trade names, commercial products or organizations imply endorsement by the US Gov- ernment. We thank Ms. J. Newton for preparation of the manuscript and Ms. B. Blake for editorial assistance.
RETROVIRUS INFECTION OF MACAQUES 607
REFERENCES
BLAKESLEE, J.R., JR., SOWDER, W.G., and BAULU, J., Wild African green monkeys of Barbados are HTLV negative. Lancer, I, 525 (1985). CHALIFOUX, L.V., KING, N.W., DANIEL, M.D., KANNAGI, M., DESRO- SIERS, R E . , SEHGAL, P.K., WALDRON, L.M., HUNT, R.D., and LETVIN, N.L., Lymphoproliferative syndrome in an immunodeficient monkey nat- urally infected with an HTLV-111-like virus (STLV-III). Lab. Invest., 55,
CHOPRA, H.C., and MASON, M.M., A new virus in a spontaneous mam- mary tumor of a rhesus monkey. Cancer Res., 30,2081-2086 (1970). CLAVEL, F., GUBTARD, D., BRUN-VEZINET, F., CHAMARET, S., REY, M.- A , , SANTOS-FERREIRA, M.O., LAURENT, A.G., DAUGUET, C., KATLAMA, C., ROUZIOUX, C., KLATZMANN, D., CHAMPALIMAUD, J.L., and MONTAG- NIER, L., Isolation of a new human retrovirus from West African patients with AIDS. Science, 223,343-346 (1986). DANIEL, M.D., KING, N.W., LETVIN, N.L., HUNT, R.D., SEHGAL, P.K., and DESROSIERS, R.C., A new type D retrovirus isolated from macaques with an immunodeficiency syndrome. Science, 223, 602-605 (1984). DANIEL, M.D., LETVIN, N.L., KING, N. W., KANNAGI, M., SEHGAL, P.K., HUNT, R.D., KANKI, P.J., ESSEX, M., and DESROSIERS, R.C., Isolation of a T-cell tropic HTLV-111-like retrovirus from macaques. Science, 228,
DANIEL, M.D., LETVIN, N.L., SEHGAL, P.K., HUNSMANN, G., SCHMIDT, D.K., KING, N.W., and DESROSIERS, R.C., Long-term persistent infection of macaque monkeys with the simian immunodeficiency virus. J. gen. Virol., 68, 3183-3189 (1987). DENHAM, W.W., History of green monkeys in the West Indies. Part I. Migration from Africa. Part II. Population dynamics of Barbadian mon- keys. J. Barbados Mus. Hist. Soc., 36,211-228, 353-371 (1981).
43-50 (1986).
1201-1204 (1985).
DESROSIERS, R.C., DANIEL, M.D., BUTLER, C.V., SCHMIDT, D.K., LET- VIN, N.L., HUNT, R.D., KING, N.W., BARKER, C.S., and HUNTER, E., Retrovirus D/New England and its relation to Mason-Pfizer monkey virus. J. Virol., 54, 552-560 (1985). DESROSIERS, R.C., LETVIN, N.L., KING, N.W., HUNT, R.D., BLAKE, B.J., ARTHUR, L.O., and DANIEL, M.D., Three retroviruses infecting macaques at the New England Regional Primate Research Center. In: R.C. Gallo, W. Haseltine, G. Klein and H. zur Hausen (eds.), Viruses and human cancer, UCLA Symposia on molecular and cellular biology, New Series, Vol. 43, pp. 451-466, A.R. Liss, New York (1987). FULTZ, P.N., MCCLURE, H.M., ANDERSON, D.C., SWENSON, R.B., AN- AND, R., and SRINIVASAN, A., Isolation of a T-lymphotropic retrovirus from naturally infected sooty mangabey monkeys (Cercocebus atys). Proc. nut. Acad. Sci. (Wash.), 83,5286-5290 (1986). GESSAIN, A., BARIN, F., VERNANT, J.C., GOUT, O., MAURS, L., CALEN- DER, A., and DE THE, G., Antibodies to human T-lymphotropic virus type-I in patients with tropical spastic paraparesis. Lancet, II, 407-410 (1985). GONDA, M.A., WONG-STAAL, F., GALLO, R.C., CLEMENTS, J.E., NA- RAYAN, O., and GILDEN, R.V., Sequence homology and morphologic similarity of HTLV-111 and visna virus, a pathogenic lentivirus. Science,
HAYAMI, M., KOMURO, A., NOZAWA, K., SHOTAKE, T., ISHIKAWA, K., YAMAMOTO, K., ISHIDA, T., HONJO, S., and HINUMA, Y., Prevalence of antibody to adult T-cell leukemia virus-associated antigens (ATLA) in Japanese monkeys and other non-human primates. In?. J. Cancer, 33,
HIRSCH, V., RIEDEL, N., KORNFELD, H., KANKI, P.J., ESSEX, M., and MULLINS, J.I., Cross-reactivity to human T-lymphotropic virus type III/
227, 173-177 (1985).
179-183 (1984).

608 DANIEL ET AL
lymphadenopathy-associated virus and molecular cloning of simian T-cell lymphotropic virus type 111 from African green monkeys. Proc. mr. Acud. Sci. wash.), 83, 9754-9758 (1986). HOOKS, J.J., and GIBBS, C.J., JR., The foamy viruses. Buct. Rev., 39, 169-185 (1975). JURKIEWICZ, E., NAKAMURA, H., SCHNEIDER, J., YAMAMOTO, N., HAY- AMI, M., and HUNSMANN, G., Structural analysis of p19 and p24 core polypeptides of primate lymphotropic retroviruses (PLRV) . Virology, 150,
KANKI, P.J., KURTH, R., BECKER, W., DREESMAN, G., MCLANE, M.F., and ESSEX, M., Antibodies to simian T-lymphotropic retrovirus type 111 in African green monkeys and recognition of STLV-IU viral proteins by AIDS and related sera. Lancet, I, 1330-1332 (1985~). KANKI, P.J., MCLANE, M.F., KING, N.W., JR., LETVIN, N.L., HUNT, R.D., SEHGAL, P., DANIEL, M.D., DESROSIERS, R.C., and ESSEX, M., Serologic identification and characterization of a macaque T-lymphotropic retrovirus closely related to HTLV-III. Science, 228, 1199-1201 (1985b). KANNAGI, M., KIYOTAKI, M., DESROSIERS, R.C., REIMANN, K.A., KING, N.W., WALDRON, L.M., and LETVIN, N.L., Humoral immune responses to T cell tropic retrovirus simian T lymphotropic virus type ILI in monkeys with experimentally induced acquired immune deficiency-like syndrome. J. din. Invest., 78, 1229-1236 (1986). KING, N.W., HUNT, R.D., and LETVIN, N.L., Histopathologic changes in macaques with an acquired immunodeficiency syndrome (AIDS). Amer. J . Parhol., 113, 382-388 (1983). LAEMMLI, U.K., Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature (Lond.), 227,680-685 (1970). LERCHE, N.W., HENRICKSON, R.V., MAUL, D.H., and GARDNER, M.B., Epidemiologic aspects of an outbreak of acquired immunodeficiency in rhesus monkeys (Macaca mulatfu). Lab. Anim. Sci., 34, 146-150 (1984). LETVIN, N.L., DANIEL, M.D., SEHGAL, P.K., CHALIFOUX, L.V., KING, N.W., HUNT, R.D., ALDRICH, W.R., HOLLEY, K., SCHMIDT, D.K., and DESROSIERS, R.C., Experimental infection of rhesus monkeys with type D retrovirus. J. Virol., 52, 683-686 (1984). LETVIN, N.L., DANIEL, M.D., SEHGAL, P.K., DESROSIERS, R.C., HUNT, R.D., WALDRON, L.M., MACKEY, J.J., SCHMIDT, D.K., CHALIFOUX, L.V., and KING, N.W., Induction of AIDS-like disease in macaque mon- keys with T-cell tropic retrovirus STLV-111. Science, 230, 71-73 (1985). MARX, P.A., MAUL, D.H., OSBORN, K.G., LERCHE, N.W., MOODY, P.,
291-298 (1986).
LOWENSTINE, L.J., HENRICKSON, R.V., ARTHUR, L.O., GILDEN, R.V., GRAVEL, M., LONDON, W.T., SEVER, J.L., LEVY, J.A., MUNN, R.J., and GARDNER, M.B., Simian AIDS: isolation of a type D retrovirus and transmission of the disease. Science, 223, 1083-1086 (1984). MCGRATH, M., WITTE, O., PINCUS, T., and WEISSMAN, I.L., Retrovirus purification: method that conserves envelope glycoprotein and maximizes infectivity. J. Virol., 25, 923-927 (1978). MIYOSHI, I., YOSHIMOTO, S., FUJISHITA, M., TAGUCHI, H., KUBONISHI, I., NIIYA, K., and MINEZAWA, M., Natural adult T-cell leukaemia virus infection in Japanese monkeys. Lancet, 11, 658 (1982). MURPHEY-CORB, M., MARTIN, L.N., RANGAN, S.R.S., BASKIN, G.B., GORMUS, B.J., WOLF, R.H., ANDES, W.A., WEST, M., and MONTELARO, R.C., Isolation of an HTLV-111-related retrovirus from macaques with simian AIDS and its possible origin in asymptomatic mangabeys. Nature
NAKAMURA, H., TANAKA, Y., KOMURO-TSUJIMOTO, A, , ISHIKAWA, K., TAKADAYA, K.I., TOZAWA, H., TSUJIMOTO, H., HONJO, S., and HAYAMI, M., Experimental inoculation of monkeys with autologous lymphoid cell lines immortalized by and producing human T-cell leukemia virus type-I. Int. J . Cancer, 38, 867-875 (1986). NIH. Guide for the care and use of laboratory animals. DHHS 85-23 Bethesda, MD (1985). RUSTIGIAN, R., JOHNSTON, P., and REIHART, H., Infection of monkey kidney tissue cultures with virus like agents. Proc. Soc. exp. Bid. (N. Y ) ,
SAKAKIBARA, I., SUGIMOTO, Y., SASAGAWA, A., HONJO, S., TSUJIMOTO, H., NAKAMURA, H., and HAYAMI, M., Spontaneous malignant lymphoma in an African green monkey naturally infected with simian T-lymphotropic virus (STLV). J. med. Primatol, 15, 311-318 (1986). TSUJIMOTO, H., KOMURO, A, , IIJIMA, K., MIYAMOTO, J., ISHIKAWA, K.-I., and HAYAMI, M., Isolation of simian retroviruses closely related to human T-cell leukemia virus by establishment of lymphoid cell lines from various non-human primates. Inr. J. Cancer, 35, 377-384 (1985). WATANABE, T., SEIKI, M., HIRAYAMA, Y., and YOSHIDA, M., Human T- cell leukemia virus type I is a member of the African subtype of simian viruses (STLV). Virology, 148, 385-388 (1986). WATANABE, T., SEIKI, M., TSUJIMOTO, H., MIYOSHI, I., HAYAMI, M. , and YOSHIDA, M., Sequence homology of the simian retrovirus genome with human T-cell leukemia virus type 1. Virology, 144, 59-65 (1985).
(Lond.), 321,435-437 (1986).
88, 8-16 (1955).
Copyright © 2022 FDOKUMEN




















