
Preparation and structural characterization of salts of oxotetrachlorotechnetate(V)
Upload
khangminh22Category
view
1download
0
Preparation, structural analysis and prebiotic potential of arabinoxylo-oligosaccharides
Helena Pastell
ACADEMIC DISSERTATION
To be presented, with the permission of the Faculty of Agriculture and Forestry of the University of Helsinki, for public criticism in lecture hall B3, Viikki,
on January 29th 2010, at 12 o´clock noon.
University of Helsinki Department of Applied Chemistry and Microbiology
Chemistry and Biochemistry / Food Chemistry
Helsinki 2010
brought to you by COREView metadata, citation and similar papers at core.ac.uk
provided by Helsingin yliopiston digitaalinen arkisto

Custos: Professor Vieno Piironen Department of Applied Chemistry and Microbiology University of Helsinki Helsinki, Finland
Supervisors: Professor Maija Tenkanen Department of Applied Chemistry and Microbiology University of Helsinki
Helsinki, Finland Docent Päivi Tuomainen Department of Applied Chemistry and Microbiology University of Helsinki Helsinki, Finland
Reviewers: Professor Alphons G.J. Voragen
Laboratory of Food Chemistry Wageningen University Wageningen, The Netherlands
Docent Tarja Tamminen VTT Technical Research Centre Helsinki, Finland
Opponent: rofessor Luc Saulnier
Research Unit on Biopolymers - Interactions and Assemblies INRA, Nantes Research Centre
Nantes, France
ISBN 978-952-10-6054-0 (paperback) ISBN 978-952-10-6055-7 (pdf) ISSN 0355-1180 Cover picture: Tuuli Koivumäki Helsinki University Print Helsinki 2010
Associate P

”Whole meal of flour is recommended for its salutary effects on the bowels.”
Hippocrates, 4th century BC


Pastell, H. 2010. Preparation, structural analysis and prebiotic potential of arabinoxylo-oligosaccharides (dissertation). EKT-Series 1463. University of Helsinki. Department of Applied Chemistry and Microbiology. 112 + 48 pp.
ABSTRACT
Arabinoxylo-oligosaccharides (AXOS) can be prepared enzymatically from arabinoxylans (AX) and AXOS are known to possess prebiotic potential. Here the structural features of 10 cereal AX were examined. AX were hydrolysed by Shearzyme® to prepare AXOS, and their structures were fully analysed. The prebiotic potential of the purified AXOS was studied in the fermentation experiments with bifidobacteria and faecal microbiota. In AX extracted from flours and bran, high amounts of �-L-Araf units are attached to the �-D-Xylp main chain, whereas moderate or low degree of substitution was found from husks, cob and straw. Nuclear magnetic resonance (NMR) spectroscopy showed that flour and bran AX contain high amounts of �-L-Araf units bound to the O-3 of �-D-Xylp residues and doubly substituted �-D-Xylp units with �-L-Araf substituents at O-2 and O-3. Barley husk and corn cob AX contain high amounts of �-D-Xylp(1�2)-�-L-Araf(1�3) side chains, which can also be found in AX from oat spelts and rice husks, and in lesser amounts in wheat straw AX. Rye and wheat flour AX and oat spelt AX were hydrolysed by Shearzyme® (with Aspergillus aculeatus GH10 endo-1,4-�-D-xylanase as the main enzyme) for the production of AXOS on a milligram scale. The AXOS were purified and their structures fully analysed, using mass spectrometry (MS) and 1D and 2D NMR spectroscopy. Monosubstituted xylobiose and xylotriose with �-L-Araf attached to the O-3 or O-2 of the nonreducing end �-D-Xylp unit and disubstituted AXOS with two �-L-Araf units at the nonreducing end �-D-Xylp unit of xylobiose or xylotriose were produced. Xylobiose with �-D-Xylp(1�2)-�-L-Araf(1�3) side chain was also purified. These AXOS were used as standards in further identification and quantification of corresponding AXOS from the hydrolysates in high-performance anion-exchange chromatography with pulsed amperometric detection (HPAEC-PAD) analysis. The prebiotic potential of AXOS was tested in in vitro fermentation experiments. Bifidobacterium adolescentis ATCC 15703 and B. longum ATCC 15707 utilized AXOS from the AX hydrolysates. Both species released L-arabinose from AXOS, but B. adolescentis consumed the XOS formed, whereas B. longum fermented the L-arabinose released. The third species tested, B. breve ATCC 15700, grew poorly on these substrates. When cultivated on pure AXOS, the bifidobacterial mixture utilized pure singly substituted AXOS almost completely, but no growth was detected with pure doubly substituted AXOS as substrates. However, doubly substituted AXOS were utilized from the mixture of xylose, XOS and AXOS. Faecal microbiota utilized both pure singly and doubly substituted AXOS. Thus, a mixture of singly and doubly substituted AXOS could function as a suitable, slowly fermenting prebiotic substance. This thesis contributes to the structural information on cereal AX and preparation of mono and doubly substituted AXOS from AX. Understanding the utilization strategies is fundamental in evaluating the prebiotic potential of AXOS. Further research is still required before AXOS can be used in applications for human consumption.

Acknowledgements
This study was carried out at the Department of Applied Chemistry and Microbiology, Chemistry and Biochemistry Division, at the University of Helsinki during the years 2003-2009. Part of the study was carried out during the research visit at the Technical University of Denmark (DTU). The work was financially supported by The Raisio Research Foundation, The Danisco Foundation, The Academy of Finland and The Finnish Cultural Foundation, which are gratefully acknowledged. COST action 928 and NordForsk Network; Food and Bioresource Enzyme Technology are acknowledged for funding the research visit at DTU. I am deeply grateful to my supervisors, Professor Maija Tenkanen and Docent Päivi Tuomainen for making this thesis possible. You got me interested in research work, and you gave me numerous good advice, constructive criticism and patience. Thank you for supporting me throughout the work with your inspiring guidance. I thank Professor Vieno Piironen for valuable comments concerning this thesis and for encouragement during the work. I thank warmly Docent Liisa Virkki for your help in preparing the manuscripts and this thesis and your continuous interest in my work. I sincerely thank Professor Anne Mayer and Associate Professor Peter Westermann for providing the great working facilities at DTU and for introducing me into the research world of microbiology and molecular biology. I owe special thanks to Louise Vigsnæs, Gitte Hinz-Berg and many others who guided me in the laboratory at DTU and made my stay in Denmark unforgettable. I want to thank my co-authors Associate Professor Henk Schols and Dr. Mirjam Kabel for your help with the MALDI-TOF-MS experiments and for constructive comments in preparing the manuscript. I thank all my current and former colleagues in the Viikki D-building for creating a supportive working atmosphere and for the nice moments we have shared. Special thanks are owed to the hemicellulose research group: Leena Pitkänen, Ndegwa Maina, Sanna Koutaniemi, Sun-Li Chong, Susanna Heikkinen, Kirsi Mikkonen, Mari Heikkilä, Kirsti Parikka, Henna Pynnönen, Liisa Johansson, Paula Koivisto, Riku Talja, Saara Hamarila and Annemai Soovre. Thank you all for commenting my work, presenting your ideas and sharing great moments as well as listening and helping in difficult times. I am very grateful to Essi Harju and Jaakko Arpiainen for the help in the lab. Professor Alphons G.J. Voragen and Docent Tarja Tamminen are thanked for careful pre-examination of this thesis, their comments and suggestions for improvements.

My heartfelt thanks go to my parents Hanna and Tapio Rantanen and my little sister Riitta. You have always believed in me, helped and supported me in every way possible. I also wish to thank my relatives and friends for the interest you have shown towards my work, for happy moments and comfort when needed. Thank you for longlasting friendship through all these years and for helping me to forget science sometimes completely... Finally, my loving thanks belong to my husband Matti and our son Aleksanteri. Thank you, Matti, for loving me for what I am, believing in me and supporting me. You have made my life complete. Aleksanteri, you can always make mom laugh with your “keksijä-Kyösti” jokes and happy with a wet kiss. I appreciate your exquisite joy of life – the world is a sunny place with you! Helsinki, January 2010 Helena Pastell

Contents
ABSTRACT 5 ACKNOWLEDGEMENTS 6 LIST OF ORIGINAL PUBLICATIONS 10 LIST OF ABBREVIATIONS 12
1 INTRODUCTION 15
2 REVIEW OF THE LITERATURE 18
2.1 Cereal arabinoxylans 18 2.1.1 Grains 18 2.1.2 Occurrence of AX 18 2.1.3 Structure of AX 20 2.1.4 Distribution of substituents in cereal AX 23
2.2 Enzymes hydrolysing arabinoxylans 25 2.2.1 Enzymes hydrolysing the xylan main chain 25 2.2.2 Enzymes acting on �-L-Araf substituents 26 2.2.3 Enzymes acting on other substituents 27
2.3 Enzymatic preparation of arabinoxylo-oligosaccharides 29 2.3.1 AXOS from isolated AX 29 2.3.2 AXOS from lignocellulosic materials 33
2.4 Separation and detection methods in AXOS analytics 33 2.4.1 Retention of monosaccharides 34 2.4.2 Retention of oligosaccharides 35 2.4.3 Pulsed amperometric detection 35
2.5 Prebiotics and probiotics 37 2.5.1 Definition of pro- and prebiotics 37 2.5.2 Potential prebiotic (oligo)saccharides 38 2.5.3 Production of SCFA during fermentation of prebiotics 39 2.5.4 Intestinal microbiota 40 2.5.5 Bifidobacterial enzymes in AXOS utilization 40
3. AIMS OF THE STUDY 44
4. MATERIALS AND METHODS 45
4.1 Materials 45 4.1.1 Carbohydrates 45 4.1.2 Enzymes 46 4.1.3 Bacteria used in the fermentation experiments 47

4.2 Experimental 47 4.2.1 Extraction of AX (III) 47 4.2.2 Monosaccharide composition analysis of AX (I, III) 47 4.2.3 Enzymatic production and chromatographic purification of AXOS (I-III) 48 4.2.4 Preparative chromatography methods (I-III) 49 4.2.5 Quantification of AXOS (I-III) 50 4.2.6 Analytical chromatographic methods (I-IV) 51
4.2.6.1 TLC 51 4.2.6.2 HPAEC-PAD 51 4.2.6.3 GC 53
4.2.7 Structural elucidation of AXOS (I-III) 55 4.2.7.1 Mass spectrometry 56 4.2.7.2 NMR spectroscopy 57
4.2.8 Microbiological and molecular biological methods (IV) 58 4.2.8.1 Fermentation experiments 58 4.2.8.2 DNA extraction, purification, PCR and t-RFLP 58
5. RESULTS 61
5.1 Carbohydrate composition of starting materials (I-IV) 61 5.2. Structures of enzymatically prepared AXOS (I-III) 61 5.3 Structural features of different cereal AX (I-III) 66 5.4 Action of Shearzyme (I-III) 68 5.5 Behaviour of AXOS in chromatography (I-III) 68
5.5.1 Separation of AXOS on TLC 68 5.5.2 Elution order of AXOS and their responses in HPAEC-PAD 69
5.6 Fermentation experiments (IV) 71 5.6.1 Carbohydrate composition of XOS and AX hydrolysates 71 5.6.2. Growth of pure bifidobacterial cultures 72 5.6.3 Fermentations with faecal microbiota 74
6. DISCUSSION 77 6.1 Carbohydrate composition of starting materials 77 6.2 Structural features of different cereal AX 78 6.3 Production of AXOS on a preparative scale 82 6.4 Challenges in identification and quantification of AXOS 84 6.5 Behaviour of AXOS in HPAEC-PAD system 85
6.5.1 Elution order of AXOS 85 6.5.2 Responses of AXOS in PAD 86
6.6 Fermentation experiments 87 6.6.1 Utilization of substrates 87 6.6.2 Substrate utilization strategies of pure bifidobacteria 90 6.6.3 Specific features in utilization of purified AXOS 92
7. CONCLUSIONS 95
8. REFERENCES 97
ORIGINAL PUBLICATIONS

LIST OF ORIGINAL PUBLICATIONS
This thesis is based on the following original publications, which are referred to in the text by the Roman numerals: I Rantanen, H., Virkki, L., Tuomainen, P., Kabel, M., Schols, H., Tenkanen, M. 2007.
Preparation of arabinoxylobiose from rye xylan using family 10 Aspergillus aculeatus endo-1,4-�-D-xylanase. Carbohydrate Polymers, 68, 350-359.
II Pastell, H., Tuomainen, P., Virkki, L., Tenkanen, M. 2008. Step-wise enzymatic
preparation and structural characterization of singly and doubly substituted arabinoxylo-oligosaccharides with non-reducing end terminal branches. Carbohydrate Research, 343, 3049-3057.
III Pastell, H., Virkki, L., Harju, E., Tuomainen, P., Tenkanen, M. 2009. Presence of 2-
O-�-D-xylopyranosyl-�-L-arabinofuranosyl side chains in cereal arabinoxylans. Carbohydrate Research, 344, 2480-2488.
IV Pastell, H., Westermann, P., Meyer, A.S., Tuomainen, P., Tenkanen, M. 2009. In
vitro fermentation of arabinoxylan-derived carbohydrates by bifidobacteria and mixed fecal microbiota. Journal of Agricultural and Food Chemistry, 57, 8598-8606.
The papers are reproduced with kind permission from the copyright holders: Elsevier (I, II, III) and American Chemical Society (IV).
Contribution of the author to papers I to IV:
I Helena Pastell (neé Rantanen) planned the study together with Prof. M. Tenkanen and Docent P. Tuomainen. Docent L. Virkki planned the NMR analyses, interpreted the results and wrote the NMR section in the manuscript. Associate Prof. H. Schols and Ph.D. M. Kabel helped in planning and executing the MALDI-TOF-MS analysis. Other experimental work, except for enzyme activity measurements and acid methanolysis analyses, was carried out by Helena Pastell. She wrote the manuscript which was jointly edited with the other authors. Helena Pastell acted as the corresponding author of the paper.
II Helena Pastell planned the study together with Prof. M. Tenkanen and Docent P.
Tuomainen, who also carried out the MALDI-TOF-MS measurements. Docent L. Virkki planned the NMR analyses, interpreted the results and wrote the NMR section in the manuscript. Helena Pastell was responsible for the other experimental work. She wrote the manuscript which was jointly edited with the other authors, and she acted as the corresponding author of the paper.

III Helena Pastell planned the study together with Prof. M. Tenkanen and Docent P. Tuomainen. Docent L. Virkki planned the NMR analyses, interpreted the results and wrote the NMR section in the manuscript. B.Sc E. Harju isolated the arabinoxylans and oligosaccharide. Other experimental work, except the preceding and ESI-MS analyses, was carried out by Helena Pastell. She wrote the manuscript which was commented on by the other authors, and she acted as the corresponding author of the paper.
IV Helena Pastell planned the study together with Prof. M. Tenkanen, Assoc. Prof. P.
Westermann and Prof. A.S. Meyer. Helena Pastell was responsible for the experimental work, except for the t-RFLP analysis. She wrote the manuscript which was jointly edited with the other authors. Helena Pastell acted as the corresponding author of the paper.

LIST OF ABBREVIATIONS
�-L-Araf alpha-L-arabinofuranosyl AX arabinoxylan(s) AzX A; �-L-Araf + �-D-Xylp, z; linkage position between �-L-Araf
and �-D-Xylp, X; (�)-D-Xyl(p) AXH-d3 �-L-arabinofuranosidase (able to release only (1�3)-linked �- L-Araf units from disubstituted �-D-Xylp residues) AXH-m �-L-arabinofuranosidase (acting on (1�2)- and (1�3)-linked �-L-Araf units on monosubstituted �-D-Xylp residues) AXOS arabinoxylo-oligosaccharide(s) �-D-Xylp beta-D-xylopyranosyl BHAX barley husk arabinoxylan CCAX corncob arabinoxylan DP degree of polymerization DS degree of substitution DQF-COSY double-quantum filtered correlation spectroscopy D2,3X �-D-Xylp(1�2)-�-L-Araf(1�3)-�-D-Xylp-(1�4)-D-Xyl ESI-MS electrospray ionization mass spectrometry FOS fructo-oligosaccharides FOSHU foods for specified healthy use GC gas chromatography GH glycoside hydrolase GOS galacto-oligosaccharides GPC gel permeation chromatography HMQC heteronuclear multiple-quantum correlation HMBC heteronuclear multibond connectivity HPAEC-PAD high-performance anion-exchange chromatography with pulsed amperometric detection HSQC heteronuclear single-quantum correlation LC/MSD liquid chromatography/mass selective detection LDL low-density lipoprotein MALDI-TOF-MS matrix-assisted laser desorption/ionization time-of-flight mass spectrometry Mw weight-average molar mass 1D NMR one-dimensional nuclear magnetic resonance 2D NMR two-dimensional nuclear magnetic resonance OBAX oat bran arabinoxylan OSAX oat spelt arabinoxylan RAX rye arabinoxylan RBAX rye bran arabinoxylan RiHAX rice husk arabinoxylan Rf retention factor SCFA short-chain fatty acids TLC thin-layer chromatography

tR retention time t-RFLP terminal restriction fragment length polymorphism WAX wheat arabinoxylan WBAX wheat bran arabinoxylan WEFT water-eliminated Fourier transform WStAX wheat straw arabinoxylan XOS xylo-oligosaccharides


15
1 Introduction
In most human diets carbohydrates are the main source of energy. Carbohydrates can be
divided into two categories: 1) carbohydrates digested and absorbed in the human intestine
and 2) dietary fibres which are nondigestible carbohydrates passing to the large intestine.
Thus, dietary fibre can be defined as “edible plant parts or analogous carbohydrates that
are resistant to digestion and absorption in the human small intestine with complete or
partial fermentation in the large intestine” (American Association of Cereal Chemists
(AACC), 2001). According to the latest definition from the European Food Safety
Authority (EFSA), dietary fibre is simply “non-digestible carbohydrates plus lignin”. The
main constituents of dietary fibre are nonstarch polysaccharides (e.g. cellulose,
hemicelluloses, pectins and �-glucans), resistant oligosaccharides (e.g. fructo-
oligosaccharides (FOS) and galacto-oligosaccharides (GOS)), resistant starch and lignin.
Utilization of foods rich in dietary fibre is associated e.g. with reduced risk for type 2
diabetes, reduced total and low-density lipoprotein (LDL)-cholesterol concentrations and
possible reduction of the risk of colon cancer. To obtain these health effects, adequate
fibre intakes for adults are considered to be 25 g/d (EFSA, 2009).
Various nondigestable oligosaccharides, categorized as dietary fibre, or oligosaccharides
obtained from polysaccharides that are classified as dietary fibre, are included among new
food ingredients and may be used in functional foods. Oligosaccharides have many
beneficial physiological properties, such as growth promotion of beneficial intestinal
bacteria (Bifidobacterium and Lactobacillus) (= prebiotic effect) (Gibson and Roberfroid,
1995; Crittenden and Playne, 1996; Voragen, 1998) and protection against colon cancer by
producing short-chain fatty acids (SCFA) in the large intestine during fermentation
(Voragen, 1998; Scharlau et al., 2009). Interest in oligosaccharides as health-promoting
food incredients has increased in Europe, but only inulin, FOS and GOS are currently
allowed for human consumption in functional foods (van Loo et al., 1999; Pascal, 2008).
In Japan, foods for specified healthy use (FOSHU) were already legislated in 1991.
FOSHU products include fructo-, galacto-, soybean, palatinose, xylo- and isomalto-
oligosaccharides, lactulose and lactosucrose. The main health claim for these products is
that they are “foods designed to help maintain a good gastrointestinal environment, and act
to increase intestinal bifidobacteria” (Crittenden and Playne, 1996).

16
FOS, GOS, xylo-oligosaccharides (XOS) and lactulose stimulate specifically the growth
of Bifidobacterium (Imaizumi et al., 1991; Crittenden et al., 2002). However, linear
oligosaccharides may be fermented too rapidly by bifidobacteria and the present objective
is to find oligosaccharides that ferment slowly in the large intestine. Slow fermentation
produces abundant SCFA, of which especially butyric and propionic acids seem to play a
key role in preventing colon cancer (Scharlau et al., 2009). Potential slowly fermenting
oligosaccharides include arabinoxylo-oligosaccharides (AXOS), produced by degrading
polymeric arabinoxylans (AX). Branched AXOS are nondigestible oligosaccharides, but
are fermented in the large intestine by the intestinal microbiota (Gibson and Roberfroid,
1995; van Laere et al., 2000). AXOS are fermented by some health-promoting
bifidobacteria and by the predominant intestinal bacteria, Bacteroides spp., but harmful
Clostridium spp. showed low utilization of branched AXOS (van Laere et al., 2000; Kabel
et al., 2002b).
To obtain health effects mediated by health-promoting bacteria, one strategy is to add
bifidobacteria to food. However, orally taken bifidobacteria may not remain in the
intestine and as soon as intake is stopped, the enterobacterial flora returns to the previous
state (Suwa et al., 1999). Another strategy is to fortify food with SCFA, but it is unlikely
that SCFA added directly to food would reach the large bowel. Thus, orally taken
fermentable fibre or oligosaccharide that would reach the lower gut may promote
proliferation of bifidobacteria and provide the positive effects of SCFA (Campbell et al.,
1997b; Suwa et al., 1999).
For preparation of nondigestable oligosaccharides, several plant sources rich in
carbohydrates are available. In the cell walls, lignin, cellulose and hemicelluloses are
closely associated and together can be referred to as lignocellulosic material (Aspinall,
1980; Puls, 1993). Forestial, agricultural and industrial wastes or by-products contain
considerable amounts of lignocellulosic materials. Processing of residual biomass as raw
materials provides economic and ecological benefits due to its biorenewability and
abundance (Alonso et al., 2003). Xylans occur as the major constituents of hemicelluloses
and thus, after cellulose, are the second most abundant renewable plant resource with high
potential for utilization as useful products (Kulkarni et al., 1999). Xylans constitute about

17
20-30% of the dry weight of agricultural residues, such as cereal straws and grain hulls
(Aspinall, 1970).
This thesis reviews the literature on the occurrence and structures of arabinoxylans in
cereals, enzymatic production of AXOS, as well as their prebiotic potential. The
experimental part of the thesis is a summary of the data published in the attached papers I-
IV, in which the substitution patterns of several polymeric AX are studied, the production,
purification and structural analysis of AXOS are characterized, and the prebiotic potentials
of AXOS are evaluated in in vitro studies. The results obtained are evaluated in the
Discussion section.

18
2 Review of the literature
2.1 Cereal arabinoxylans
2.1.1 Grains
Cereal grains consist typically of bran, germ and starchy endosperm. In addition, some
common cereals also carry husks (hulls); wheat (Triticum aestivum L.) and rye (Secale
sereale L.) are dehulled during harvest, whereas barley (Hordeum vulgare L.) and oat
(Avena sativa L.) are not, but husks are removed in traditional ways before utilization in
the human diet. Brans are manufactured from grains, using methods adequate for
structurally different cereal species, resulting in brans containing diverse grain layers.
Wheat brans are fairly clean of other layers, barley brans contain some husk remnants and
oat brans often contain substantial quantities of starchy endosperm (Fulcher and Duke,
2002). Cereal plant cell walls consist mainly of polysaccharides, lignin and protein (Puls,
1993; Aspinall, 1980). The major polymers in the cell walls are cellulose (25-35%),
hemicelluloses (40-50%) and lignin (7-10%) (Ishii, 1997). Both cellulose and
hemicelluloses function as structural supporting materials in the cell walls; cellulose has a
high tensile strength and gives rigidity to the walls, whereas hemicelluloses impart
elasticity to the structure by cross-linking cellulose microfibrils (Sjöström, 1981; Carpita
and Gibeaut, 1993).
2.1.2 Occurrence of AX
Xylans occur as the most common hemicelluloses, and after cellulose they are the second
most abundant polysaccharide in the plant kingdom. Xylans can be divided into three
groups: 1) (glucurono-)arabinoxylans, which are present in softwoods, lignified tissues of
grasses and annual plants, 2) neutral arabinoxylans in cereal grains and 3)
glucuronoxylans found in hardwoods (Aspinall, 1959, 1970; Sjöström, 1981; Ebringerová
and Heinze, 2000). Cereal whole grains contain AX from 1.2 wt% (in rice (Oryza sativa
L.)) to 8.5 wt% (in rye), whereas xylans in more lignified tissues of cereals (leaves, straws,
husks) represent 25-35% of dry biomass (Aspinall, 1970; Henry, 1985; Härkönen et al.,
1997; Moure et al., 2006). Even though the proportion of AX is quite small in whole

19
grain, the relative proportion is considerable in cereal endosperm cell walls, where AX
comprise from 20% (w/w; in barley) to 72% (w/w; in wheat) (Fincher, 1975; Bacic and
Stone, 1980). Thus, AX are the main cell wall components of several cereals (Hoffmann et
al., 1992). However, endosperm cell walls contribute only 0.5-5% of the dry weight of
cereal whole grains and they also constitute nonstarch polymers other than AX, e.g.
cellulose, (1�3)(1�4)-�-D-glucans and glucomannans (Bacic and Stone, 1981; Fulcher
and Duke, 2002). An average of 2.1 wt% of AX in flour (mainly endosperm) was reported
(Andersson and Åman, 2001). Quantitatively, most of the AX are situated in the bran
layers, which contribute about 25% of the dry weight of the grain (Fulcher and Duke,
2002).
Arabinoxylans can be divided into water-extractable and water-unextractable AX. The
water-unextractability is due to a combination of noncovalent interactions and covalent
bonds with other cell wall components, such as proteins, cellulose and lignin (McNeil et
al., 1975; Markwalder and Neukom, 1976; Andrewartha et al. 1979; Jeffries, 1990). Many
of these linkages are alkaline-labile and thus some AX can be liberated and extracted by
alkaline treatment (Ishii, 1997; Courtin and Delcour, 2001). Gruppen et al. (1991) showed
that with barium hydroxide (Ba(OH)2) the major part of AX was selectively extracted
from different cereal samples. Other alkals, such as sodium hydroxide (NaOH) and
potassium hydroxide (KOH), have also been used to extract AX. KOH is preferred over
NaOH because mannans are more or less insoluble in KOH (Puls and Schuseil, 1993). In
addition (1�3, 1�4)-�-glucans are coextracted with KOH and NaOH, and thus they are
not as selective as Ba(OH)2 (Gruppen et al., 1991). The xylan group includes both alkali-
and water-extractable polysaccharides (Aspinall, 1970) but regardless of the extraction
method, AX have different solubilities in water, since water-solubility is dependent on the
structure of the polymer. Unsubstituted xylans are nearly water-insoluble, but with
increasing amounts of arabinose side groups, the polymers become more water-soluble
(Sternemalm et al., 2008; Pitkänen et al., 2009). The amount and structural arrangement of
the side chains of AX vary between cereal species and even among different parts of the
same plant.

20
2.1.3 Structure of AX
The linear main chain of arabinoxylans is composed of (1�4)-linked �-D-xylopyranosyl
(�-D-Xylp) residues. Cereal xylans are mainly substituted with (1�2)- and/or (1�3)-
linked �-L-arabinofuranosyl (�-L-Araf) residues, thus resulting in mono- and
disubstituted �-D-Xylp residues. The �-L-Araf side groups are usually (1�3)-linked with
�-D-Xylp residues in all cereals (Ebringerová et al., 1990; Gruppen et al., 1992; Viëtor et
al., 1992; Izydorczyk and Biliaderis, 1993). Monosubstituted �-D-Xylp units with (1�2)-
linked �-L-Araf groups are usually found in alkali-extractable barley flour AX (22%) but
they have been found in small amounts in all cereal AX. Water-extractable wheat bran AX
possesses the highest amounts of doubly (1�2)- and (1�3)-substituted �-D-Xylp units
(40% of substituted units) (Viëtor et al., 1992; Shiiba et al., 1993; Izydorczyk and
Biliaderis, 1995). In addition to �-L-Araf residues, the main chain �-D-Xylp units can also
carry �-D-glucopyranosyluronic acid (�-D-GlcA) or its 4-O-methyl ether (4-O-Me-�-D-
GlcA) and acetyl substituents (Aspinall, 1959, 1980; Vázquez et al., 2000). Ferulic and p-
coumaric acids may be ester-linked to the AX structure at the O-5 position of some of the
�-L-Araf units (Saulnier et al., 1995b; Ishii, 1997). Acetyl groups, ferulic and p-coumaric
acids are easily released in alkaline-extraction (Puls and Schuseil, 1993). The main
structural units of AX are presented in Figure 1 and the hypothetical structure of AX in
Figure 2.
O
OHH
OHOH
HOH2C
H
H
OH
HH
OHOH
H OH
H
OH
L-arabino-furanose
D-xylo-pyranose
OH
HH
OH
H OH
COOH
OH
H3CO
4-O-MeGlcA
HO
OCH3
CH CH COOH
ferulic acid
HO
CH CH COOH
p-coumaric acid
Figure 1. Main structural units of arabinoxylans.
Some cereal xylans may also carry side chains containing more than one sugar unit
(Aspinall, 1970). In these more extended side chains, the �-L-Araf residues attached to the
xylan main chain carry additional substituents, such as galactose or xylose units (Aspinall,
1980). These diverse side chains were isolated from more highly lignified tissues of the
cereal plants (Wilkie, 1979).

21
The disaccharide 2-O-�-D-Xylp-L-Araf side chain, attached to the O-3 of the main xylan
chain, was first reported in corncob AX and barley husk AX (Whistler and McGilvray,
1955; Aspinall and Ferrier, 1958; Kusakabe et al., 1983; Ebringerová et al., 1992). Later
on, this side chain structure was also identified in corn (Zea mays L.) hull and corn bran
AX (Hromádková and Ebringerová, 1995; Saulnier et al., 1995a). The existence of this
disaccharidic side chain in alkali-extracted barley husk AX was recently shown by Höije
et al. (2006). Wende and Fry (1997) reported that the disaccharide, usually esterified by
ferulic acid at the O-5 position of the Araf unit, is a widespread component of grass cell
walls. Different main substituent patterns in various cereal-based plant parts are shown in
Table 1.

22
Tabl
e 1.
The
mai
n su
bstit
uent
s of v
ario
us A
X fr
om c
erea
l pla
nts.
Subs
titue
nts
AX
sour
ce
��-L
-Ara
f (1
�3)
mon
o �
-L-A
raf
(1�
2)m
ono
�-L
-Ara
f (1
�2,
1�
3)di
(4
-O-M
e-)�
-D-G
lcA
(1
�2)
2-
O- �
-D-X
ylp-�
-L-A
raf
(1�
3)
Ref
eren
ce
Barle
y flo
ur
x x
x
V
iëto
r et a
l., 1
992;
Tr
ogh
et a
l., 2
004
Bar
ley
husk
x
x x
Asp
inal
l and
Fer
rier,
1957
; H
öije
et a
l., 2
006
Cor
ncob
x
x x
Ebrin
gero
vá e
t al.,
199
2
Cor
n hu
sk
x
x
x Eb
ringe
rová
and
Hro
mád
ková
, 199
7
Oat
end
ospe
rm
x
x
W
este
rlund
et a
l., 1
993
O
at b
ran
x
x
Wes
terlu
nd e
t al.,
199
3
Ric
e en
dosp
erm
x
Sh
ibuy
a an
d M
isaki
, 197
8
Ric
e br
an
x
x
Eb
ringe
rová
and
Hei
nze,
200
0
Rye
flou
r x
x x
Cyr
an et
al.,
200
3 V
erw
imp
et a
l., 2
007
Rye
bra
n
x
x
R
oubr
oeks
et a
l., 2
000
Whe
at fl
our
x
x
C
leem
put e
t al.,
199
7
Whe
at b
ran
x
x x
Br
illou
et e
t al.,
198
2;
Scho
onev
eld-
Ber
gman
s et
al.,
199
9 W
heat
stra
w
x
x
Su
n et
al.,
199
6

23
2.1.4 Distribution of substituents in cereal AX
Substituents are not always regularly distributed on the AX backbone. Particularly for AX
with high amounts of substituents, an alternation of highly branched and less branched
regions has been proposed (Ewald and Perlin, 1959; Gruppen et al., 1993). Generally,
cereal endosperm cell walls contain highly branched arabinoxylans, whereas more highly
lignified tissues, such as grasses, straw, husks and corncobs, usually contain less branched
AX with additional (4-O-methyl) glucuronic acid side groups (Aspinall, 1980). Rye,
wheat, rice and oat brans contain highly branched AX similar to endosperm AX from
wheat, rice and rye. AX with low degrees of branching have been reported earlier from
rice hull, wheat straw and oat spelts (Hromádková et al., 1987; Schooneveld-Bergmans et
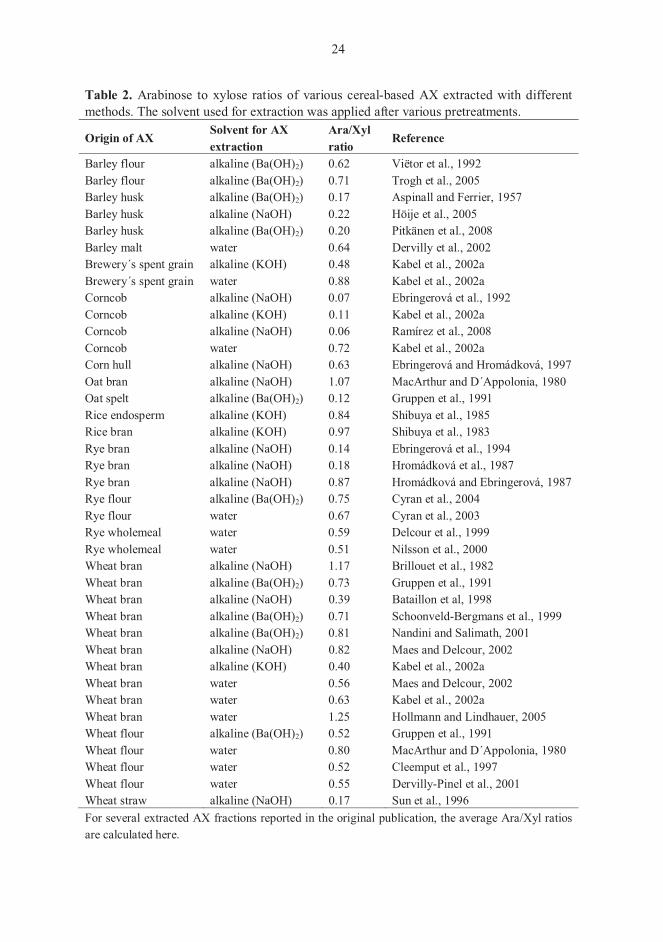
al., 1999; Puls et al., 2006). Arabinose-to-xylose (Ara/Xyl) ratios ranging from 0.5 to 0.8
in cereal endosperm and from 0.4 to 1.2 in bran commonly occur, but wide natural
variations are found. The Ara/Xyl ratios in the endosperm, aleurone layer, inner pericarp,
testa and outer pericarp are diverse, and the outer pericarp may possess the highest
Ara/Xyl ratios in several cereals (Izydorczyk and Biliaderis, 1995; Glitsø and Bach
Knudsen, 1999; Ordaz-Ortiz et al., 2005). Different manufacturing techniques for cereal
brans and difficulties in separation of cereal cell wall layers may cause alternation in
comparing Ara/Xyl ratios, because the various cell wall layers are represented in the
extracted materials.
The cell walls of wheat and rye endosperms are rich in AX. The Ara/Xyl ratio is about 0.5
in water-soluble wheat and rye flour AX from Megazyme International Ireland Ltd. (Bray,
Ireland), but the structure regarding the type of �-L-Araf substitution differs significantly.
In wheat flour AX, about one third of the �-L-Araf are (1�3)-linked with
monosubstituted �-D-Xylp residues and two thirds are (1�2)- and (1�3)-linked to
doubly substituted �-D-Xylp (Sørensen et al., 2006; Pitkänen et al., 2009). Rye flour AX
has significantly more singly substituted �-D-Xylp residues, since two thirds of the �-L-
Araf are (1�3)-linked to mono- and one third to doubly substituted �-D-Xylp,
respectively (Höije et al., 2008; Pitkänen et al., 2009). The alkaline-extracted wheat
endosperm AX structure corresponds to that of water-extracted wheat endosperm,
although small variations in the Ara/Xyl ratio and molecular weight were reported
(Gruppen et al., 1993). The Ara/Xyl ratios of various AX extracted from different cereal
materials are shown in Table 2.
24
Table 2. Arabinose to xylose ratios of various cereal-based AX extracted with different methods. The solvent used for extraction was applied after various pretreatments.
Origin of AX Solvent for AX extraction
Ara/Xyl ratio
Reference
Barley flour alkaline (Ba(OH)2) 0.62 Viëtor et al., 1992 Barley flour alkaline (Ba(OH)2) 0.71 Trogh et al., 2005 Barley husk alkaline (Ba(OH)2) 0.17 Aspinall and Ferrier, 1957 Barley husk alkaline (NaOH) 0.22 Höije et al., 2005 Barley husk alkaline (Ba(OH)2) 0.20 Pitkänen et al., 2008 Barley malt water 0.64 Dervilly et al., 2002 Brewery´s spent grain alkaline (KOH) 0.48 Kabel et al., 2002a Brewery´s spent grain water 0.88 Kabel et al., 2002a Corncob alkaline (NaOH) 0.07 Ebringerová et al., 1992 Corncob alkaline (KOH) 0.11 Kabel et al., 2002a Corncob alkaline (NaOH) 0.06 Ramírez et al., 2008 Corncob water 0.72 Kabel et al., 2002a Corn hull alkaline (NaOH) 0.63 Ebringerová and Hromádková, 1997 Oat bran alkaline (NaOH) 1.07 MacArthur and D´Appolonia, 1980 Oat spelt alkaline (Ba(OH)2) 0.12 Gruppen et al., 1991 Rice endosperm alkaline (KOH) 0.84 Shibuya et al., 1985 Rice bran alkaline (KOH) 0.97 Shibuya et al., 1983 Rye bran alkaline (NaOH) 0.14 Ebringerová et al., 1994 Rye bran alkaline (NaOH) 0.18 Hromádková et al., 1987 Rye bran alkaline (NaOH) 0.87 Hromádková and Ebringerová, 1987 Rye flour alkaline (Ba(OH)2) 0.75 Cyran et al., 2004 Rye flour water 0.67 Cyran et al., 2003 Rye wholemeal water 0.59 Delcour et al., 1999 Rye wholemeal water 0.51 Nilsson et al., 2000 Wheat bran alkaline (NaOH) 1.17 Brillouet et al., 1982 Wheat bran alkaline (Ba(OH)2) 0.73 Gruppen et al., 1991 Wheat bran alkaline (NaOH) 0.39 Bataillon et al, 1998 Wheat bran alkaline (Ba(OH)2) 0.71 Schoonveld-Bergmans et al., 1999 Wheat bran alkaline (Ba(OH)2) 0.81 Nandini and Salimath, 2001 Wheat bran alkaline (NaOH) 0.82 Maes and Delcour, 2002 Wheat bran alkaline (KOH) 0.40 Kabel et al., 2002a Wheat bran water 0.56 Maes and Delcour, 2002 Wheat bran water 0.63 Kabel et al., 2002a Wheat bran water 1.25 Hollmann and Lindhauer, 2005 Wheat flour alkaline (Ba(OH)2) 0.52 Gruppen et al., 1991 Wheat flour water 0.80 MacArthur and D´Appolonia, 1980 Wheat flour water 0.52 Cleemput et al., 1997 Wheat flour water 0.55 Dervilly-Pinel et al., 2001 Wheat straw alkaline (NaOH) 0.17 Sun et al., 1996 For several extracted AX fractions reported in the original publication, the average Ara/Xyl ratios are calculated here.

25
2.2 Enzymes hydrolysing arabinoxylans
2.2.1 Enzymes hydrolysing the xylan main chain
The arabinoxylan backbone is degraded by glycoside hydrolases (GH) that are able to
hydrolyse the glycosidic bond between (1�4)-linked �-D-Xylp units. Such enzymes are
produced e.g. by various bacteria and fungi. The plant AX backbone is randomly
hydrolysed, using endo-1,4-�-D-xylanases (EC 3.2.1.8), producing a mixture of various
oligosaccharides. Xylanases are currently classified based on the structural and sequence
similarities in GH families 5, 7, 8, 10, 11 and 43, but research has mainly focused on the
family GH10 and GH11 enzymes (CAZy - Carbohydrate Active enZymes; Collins et al.,
2005).
GH10 and GH11 xylanases possess distinct catalytic properties. Pell et al. (2004) and
Fujimoto et al. (2004) observed that the family GH10 contains enzymes with slightly
diverse catalytic sites compared with GH11 enzymes, resulting in small differences in
their hydrolytic end-products. The xylo-oligosaccharides resulting from the activity of
GH10 xylanases are shorter than those produced by GH11 xylanases. GH11 xylanases are
hindered by the presence of side groups, whereas GH10 xylanases are able to act near the
substitution, forming oligosaccharides that carry subsituents at the nonreducing terminal
of the �-D-Xylp residue (Biely et al., 1997; Vardakou et al., 2003; Beaugrand et al., 2004;
Maslen et al., 2007) (Figure 2). The shortest AXOS produced were �-L-Araf-(1�3)-�-D-
Xylp-(1�4)-D-Xylp (A3X) and �-D-Xylp-(1�4)[�-L-Araf-(1�3)]-�-D-Xylp-(1�4)-D-
Xylp (XA3X). The feruloyl substituents do not impede the action of xylanases, since
feruloyl arabinoxylobiose is the shortest feruloylated oligosaccharide liberated by GH10
xylanase (Vardakou et al., 2003). The naming system for oligosaccharides is adopted from
Fauré et al. (2009).
In addition to endo-1,4-�-D-xylanases, there are enzymes capable of hydrolysing the
xylose chain from the ends. Exo-1,4-�-D-xylosidases (EC 3.2.1.37) release xylose units
from the nonreducing end of XOS, but the effectivity decreases with increasing chain
length of XOS (Biely, 1985). Some of the exo-1,4-�-D-xylosidases can also slowly
hydrolyse polymeric xylan (Margolles-Clark et al., 1996), but terminal substituents, such

26
as (1�2)-linked 4-O-�-D-MeGlcA, at the nonreducing end of the xylose chain prevent
the action of the enzyme (Poutanen et al., 1990). Further, the penultimate (1�3)-linked
�-L-Araf residue prevents the exo-1,4-�-D-xylosidase from Trichoderma reesei from
hydrolysing the �-1,4-glycosidic linkage before the substituted xylose unit (Tenkanen et
al., 1996). In addition, the reducing end xylose-releasing exo-oligoxylanases (EC
3.2.1.156) are able to hydrolyse the (A)XOS further into even shorter oligosaccharides
(Tenkanen et al., 2003). The last-mentioned enzyme is not able to hydrolyse xylobiose, in
contrast to exo-1,4-�-D-xylosidases. The sites of activity of the xylan main chain-
hydrolysing enzymes are presented in Figure 2.
2.2.2 Enzymes acting on ��-L-Araf substituents
The �-L-arabinofuranosidases (EC 3.2.1.55) are the enzymes that cleave terminal �-L-
Araf residues from different polysaccharides and oligosaccharides. The �-L-
arabinofuranosidases acting on polymeric AX (arabinoxylan arabinofuranohydrolases,
AXH) are further divided according to their substrate specificities, since some act on
(1�2)- and (1�3)-linked �-L-Araf units on monosubstituted �-D-Xylp residues (AXH-
m), whereas others release only (1�3)-linked �-L-Araf units from disubstituted �-D-
Xylp residues (AXH-d3) (van Laere et al., 1997) (Figure 2). Most �-L-
arabinofuranosidases isolated, such as from Aspergillus awamori (Kormelink et al.,
1993c), Pseudomonas fluorescens (Kellett, 1990) and wheat (Beldman et al., 1996), are
active only on �-L-Araf residues on singly substituted �-D-Xylp residues (AXH-m).
AXH-d3-type �-arabinofuranosidases were isolated from Bifidobacterium adolescentis
(van Laere et al., 1997, 1999) and Humicola insolens (Sørensen et al., 2006). The first
reported �-arabinofuranosidase, able to release �-L-Araf from both singly and doubly
substituted �-D-Xylp, was isolated from barley malt (Ferré et al., 2000). This �-
arabinofuranosidase is able to act on linkages at the nonreducing terminal �-D-Xylp with
one or two �-L-Araf substituents, but shows no activity towards doubly substituted
internal �-D-Xylp. Highly specific AXH-m and AXH-d3, which act on polymers and
oligomers, are excellent tools to use in modifying the branching ratio of AX and AXOS.

27
2.2.3 Enzymes acting on other substituents
Xylan �-1,2-glucuronidases (EC 3.2.1.131) hydrolyse the linkage between the (4-O-
methyl)-�-D-glucopyranosyl substituent and the �-D-Xylp unit. �-1,2-Glucuronidase
activity is found in bacteria and fungi (Puls et al., 1987). Some glucuronidases are able to
act on polymeric xylan and some on oligosaccharides (Khandke et al., 1989; Tenkanen
and Siika-aho, 2000). Acetyl groups are removed by acetyl xylan esterases (EC 3.1.1.72),
which cleave the acetyl groups from positions O-2 and O-3 of �-D-Xylp units (Biely,
1985; Kormelink and Voragen, 1993). The enzyme is more important in modification of
water-extractable AX, since during alkaline-extraction of AX most acetyl groups are
removed. Furthermore, feruloyl esterases (EC 3.1.1.73) are able to cut the ester linkages
between ferulic or p-coumaric acids and �-L-Araf units (Smith and Hartley, 1983).

28
Figu
re. H
ypot
heti
cal
cere
al-b
ased
ara
bino
xyla
n st
ruct
ure
and
the
glyc
anas
es h
ydro
lysi
ng t
he x
ylan
bac
kbon
e (a
-c)
and
the
glyc
anas
es
rele
asin
g th
e su
bsti
tuen
ts p
rese
nted
(d-
f). T
he f
unct
ion
of r
educ
ing
end
xylo
se-r
elea
sing
exo
-oli
goxy
lana
se (
g) is
sho
wn
on A
XO
S.
OO
O
OO
OH H
OC
H 2O
H
OH
OO
H
OH
OO
OO
O
OO
OH
OH
HO
H 2C
OO
OH H
OC
H 2O
H
OH
O
HO
OH
OH
OO
HO
OH
OO
O
OO
OH
OH
HO
H 2C
OH
OO
HO
O
OC
H 3 HO OH
O
HO
OC
OO
OO
OO
O
HO
CH 2
OH
OH
OO
H
OH
OH
OH
O
OH
OH
OO
HO
OH
OH
OO
OH H
OC
H 2O
H
O
HO
OH
OO
HO H
OO
H a. e
ndo-
1,4--
D-x
ylan
ase
GH
10 (E
C 3
.2.1
.8)
b. e
ndo-
1,4--
D-x
ylan
ase
GH
11 (E
C 3
.2.1
.8)
c. e
xo-1
,4-
-D-x
ylos
idas
e (E
C 3
.2.1
.37)
d.
-L-a
rabi
nofu
rano
sida
seAX
H-m
(EC
3.2
.1.5
5)e.
-L
-ara
bino
fura
nosi
dase
AXH
-d3
(EC
3.2
.1.5
5)f.
-glu
curo
nida
se(E
C 3
.2.1
.131
)g.
redu
cing
end
xylo
se-r
elea
sing
exo-
olig
oxyl
anas
e(E
C 3
.2.1
.156
)
a, b
, ca
de a
aa,
ba
f
d
d
a OO
OO
O
OO
OH
OH
HO
H 2C
OH
O
HO
OH
OH
OH
HO
OH
OHO
OH
g
d
c
2

29
2.3 Enzymatic preparation of arabinoxylo-oligosaccharides
Arabinoxylo-oligosaccharides (AXOS) can be produced from polymeric arabinoxylans,
using restricted acid hydrolysis, hydrothermal processing (autohydrolysis), extensive dry
ball milling or enzymatic hydrolysis (Garrote and Parajó, 2002; Tenkanen, 2004; Van
Craeyveld et al., 2009). Only enzymatic hydrolysis allows specific regulation of the end-
products and it is possible to obtain AXOS having the desired degree of polymerization
(DP) and substitution patterns. Others of the above-mentioned methods produce more
indiscriminate oligosaccharide mixtures with varying polymerization and substitution
degrees. The enzymatic preparation of AXOS is described in detail in the following
sections.
Purified enzymes allow the hydrolysis end-products to be successfully regulated, while
commercial enzyme mixtures usually contain a combination of different enzymes that may
affect the hydrolysis pattern. The DP and degree of substitution (DS) can be effectively
reduced with different enzymes. The hydrolysis products are mainly xylose, unsubstituted
linear XOS and AXOS. However, if the xylose backbone is heavily substituted, there are
no sites on the backbone accessible for the enzymes. For structural analysis, AXOS must
be isolated from the other hydrolysis end-products. Gel permeation chromatography
(GPC) and anion-exchange chromatography (AEC) are widely used methods in
preparative separation of monosaccharides and various oligosaccharides, based on their
differences in size and charge (Kormelink et al., 1993b; Kabel et al., 2002c). AXOS,
formed in hydrolysis, provide information on the substrate specificity of the enzymes
used, and the data obtained from the hydrolysis products can be used in making structural
models for AX (Gruppen et al., 1993).
2.3.1 AXOS from isolated AX
AXOS, with DP of 3-11, were prepared and identified in the studies presented in Table 3.
The most comprehensive studies on isolation and structural characterization of AXOS
were carried out after the hydrolysis of wheat endosperm AX with Aspergillus awamori
xylanases I and III, which unfortunately have not been assigned into GH families but most
probably are members of GH10 and GH11, respectively (Gruppen et al., 1992; Kormelink
et al., 1993a; Viëtor et al., 1994). Kormelink et al. (1993b) reported that A. awamori endo-

30
1,4-�-D-xylanase I converted 72% of the wheat flour AX into mono-, di- and
oligosaccharides with DP 3-10, whereas xylanase III hydrolysed 50% of AX into
oligosaccharides with DP 5-10. Endo-1,4-�-D-xylanase I was able to produce singly and
doubly substituted AXOS with nonreducing terminal branches. AXOS with at least one
unsubstituted xylose unit in the nonreducing terminal before the branched xylose residue
were produced by endo-1,4-�-D-xylanase III. Similarly, shorter AXOS were also formed
by other GH10 endo-1,4-�-D-xylanases than by those from GH11 presented in Table 3.
The amounts of AXOS produced have seldom been determined. Viëtor et al. (1994)
reported a 24 wt% yield for all quantified mono- and oligosaccharides from barley AX
after xylanase treatment. In the study of Ordaz-Ortiz et al. (2004), 8 wt% of AXOS with
DP 4-9 were identified and quantified after wheat flour AX xylanase treatment. The main
AXOS (XA3XX) with the highest yield comprised 3% (77 mg) of all the AXOS
quantified.
A disaccharidic side chain fairly uncommon in cereals, namely 2-O-�-D-Xylp-�-L-Araf,
was reported in AXOS obtained by GH family 11 endo-1,4-�-D-xylanase hydrolysis of
barley husk AX. This hexasaccharide (�-D-Xylp-(1�4)-[�-D-Xylp-(1�2)-�-L-Araf-
(1�3)]-�-D-Xylp-(1�4)-�-D-Xylp-(1�4)-D-Xyl) (XD2,3XX), was isolated and the
structure determined by Höije et al. (2006).
The commercial preparation Ultraflo (Novo-Nordisk A/S, Bagsvaerd, Denmark) has also
been used to prepare AXOS. Ultraflo is a �-glucanase preparation produced by Humicola
insolens and it contains side activities such as cellulase, xylanase, arabinase and feruloyl
esterase (Faulds et al., 2002). Broberg et al. (2000) treated the extracted barley malt AX
with Ultraflo and identified six different AXOS (A2XX, A2+3XX, XA2+3XX, A2XXX,
A2+3XXX). The AXOS amounts in the fractions collected were 3-18 �g (4-26 nmol).

31
Table 3. Arabinoxylo-oligosaccharides obtained in hydrolysis of various AX extracted from cereal-based materials. The main AXOS produced are underlined. Enzyme, GH family (Production organism)
Substrate (AX) origin AXOS Reference
Endo-1,4-�-D-xylanase HC-II GH family not determined (Ceratocystis paradoxa)
Wheat endosperm
A3X A3XX A3XXX
Dekker and Richards, 1975
Endo-1,4-�-D-xylanase GH11* (Aspergillus)
Wheat flour XA3XX XA2+3XX XA3XXX XXA3XX XA2+3XXX XXA2+3XX
Hoffmann et al., 1991
Endo-1,4-�-D-xylanase I GH10* (Aspergillus awamori)
Wheat flour A3X XA3X A3A3X A2+3XX A2+3A3X XA3A3X XA2+3XX A3A2+3XX A2+3A2+3XX XA3A2+3XX XA2+3A2+3XX XA2+3XA2+3XX
Kormelink et al., 1993b; Gruppen et al., 1993
Endo-1,4-�-D-xylanase III GH11* (Aspergillus awamori)
Wheat flour XA3XX XA2+3XX XXA3XX XA3A3XX XXA2+3XX XA3A2+3XX XA2+3A2+3XX XA2+3A3XX XXA3A3XX XXA3A2+3XX XA3A2+3XXX XA3XA3XX XA2+3XA2+3XX XA3XXA3XX XXA3XA3XX
Kormelink et al., 1993b; Gruppen et al., 1993
Endo-1,4-�-D-xylanase I GH10* (Aspergillus awamori)
Barley cell wall A3X XA3X A2XX A2+3XX XA2+3XX A3A3X XA3A3X A2A3X A2+3A3X
Viëtor et al., 1994

32
Table 3. continued Enzyme, GH family (Production organism)
Substrate (AX) origin AXOS Reference
Endo-1,4-�-D-xylanase GH11 (Aspergillus tubingensis)
Wheat flour XA2XX A2+3XX XA2+3XX XA3A2+3XX X 2+3A2+3XX XA2+3XA2+3XX
van Laere et al., 2000
Endo-1,4-�-D-xylanase I GH10* (Aspergillus awamori)
Brewery´s spent grain
A3X XA3X A2+3XX XA2+3XX
Kabel et al., 2002a, Kabel et al., 2002b
Endo-1,4-�-D-xylanase I GH10* (Aspergillus awamori)
Corncob A3X A2XX XA3X
Kabel et al., 2002a
Endo-1,4-�-D-xylanase I GH10* (Aspergillus awamori)
Wheat bran A3X XA3X
Kabel et al., 2002a
Endo-1,4-�-D-xylanase GH11 (Trichoderma viride)
Wheat flour XA3X XA3XX XA2+3XX XA3A3XX XA3A2+3XX XA2+3A3XX XA2+3A2+3XX XA3XA3XX
Ordaz-Ortiz et al., 2004
Endo-1,4-�-D-xylanase M3 GH11 (Trichoderma longibrachiatum)
Barley husks XD2,3XX Höije et al., 2006
Endo-1,4-�-D-xylanase† GH11 (Thermomyces lanuginosus)
Wheat flour XA3X XA2+3X XA3XX XA2+3XX XA3A3XX XA2+3A3XX XA3A2+3XX XA3XA3XX XXA2+3XX XA3A2+3A3XX XA2+3XA2+3XX XA2+3XA3XX XA2+3XXA2+3XX XA3XA3XA3XX XA2+3XXA3XX XA2+3A2+3XX XA3XXA3XX
Puchart and Biely, 2008
*Belong most probably to the GH families indicated † AXOS obtained from the fermentation of AX by fungus T. lanuginosus, not purified enzyme

33
2.3.2 AXOS from lignocellulosic materials
In addition to preparing AXOS from extracted AX, AXOS can also be produced directly
from lignocellulosic material. In the study of Swennen et al. (2006), AXOS were produced
on a large scale from wheat bran (500 kg). Wheat bran was first enzymatically destarched
and deproteinized and later treated with Bacillus subtilis GH11 endo-1,4-�-D-xylanase.
The AXOS obtained were fractionated with a graded ethanol precipitation, in which the
ethanol concentration was increased stepwise until a final concentration of 90% (v/v) was
reached. The AXOS produced had an average DP of 15 and Ara/Xyl ratio of 0.27. The
yield of AXOS was 6 wt% with a purity of 72%. The �-D-Xylp units were generally
monosubstituted with �-L-Araf O-3 residues, but the AXOS obtained with 40-70%
ethanol precipitation contained nearly as much �-L-Araf doubly substituted �-D-Xylp
units. In the study of Maes et al. (2004), destarched and deproteinized wheat bran (1000 g)
was incubated with endo-1,4-�-D-xylanase from B. subtilis (GH11; Grindamyl H640,
Danisco A/S, Copenhagen, Denmark) and Aspergillus aculeatus (GH10; Shearzyme 500L,
Novozymes A/S, Bagsvaerd, Denmark). Endo-1,4-�-D-xylanase from B. subtilis was able
to hydrolyse 41 wt% of water-unextractable AX into AXOS with average DP of 15, while
A. aculeatus endo-1,4-�-D-xylanase hydrolysed only 18 wt% (average DP of 8). The same
study showed that, A. aculeatus endo-1,4-�-D-xylanase prefentially degrades water-
extractable AX.
2.4 Separation and detection methods in AXOS analytics
Carbohydrates have hardly any ultraviolet (UV) absorption, due to absence of �-bonding,
and therefore either refractive index (RI) or electrochemical (EC) detection are used in
their analysis. EC detection has become the preferred analytic method, especially in trace
analysis, due to its low detection limits (picomolar level) (Johnson and LaCourse, 1990;
Lee, 1990; Johnson et al., 1993). High-performance anion-exchange chromatography
coupled with pulsed amperometric detection (HPAEC-PAD) is currently routinely used
for oligosaccharide separation and analysis. HPAEC-PAD is capable of exceptional
resolution of neutral and charged oligosaccharide isomers (Lee, 1990, 1996; Kabel et al.,
2002c). Futhermore, the reproducibility of HPAEC-PAD is favourable (Thayer et al.,

34
1998). One of the greatest advantages of HPAEC-PAD is its capability for separating
underivatized oligosaccharides (Lee, 1990).
Carbohydrates are weak acids with pKa > 12, and in anion-exchange columns separation is
achieved at high pH, using strong alkaline solutions (0.1 M NaOH) (Lee, 1990; Johnson et
al., 1993). Dissociation of protons from hydroxyl groups at high pH creates oxyanions that
are retained by the HPAEC column (Thayer et al., 1998). The retention time (tR) is
inversely correlated with pKa value and increases with increasing molecular mass of the
carbohydrates. Retention times can be substantially decreased by addition of acetate ions
(OAc-) (Johnson et al., 1993).
2.4.1 Retention of monosaccharides
Predicting the elution order of monosaccharides is complicated, due to their complex
equilibrium between pyranoses and furanoses, and �- and �-anomers. Some principal
elution rules can be stated based on carbohydrate stereochemistry and functional groups.
The anomeric hydroxyl group of carbohydrates is more acidic than the others and affects
the most. The elution order largely corresponds to the pKa values of carbohydrates, and
among pentoses the monosaccharide with the higher pKa elutes first, e.g. arabinose (pKa
12.43) elutes before xylose (pKa 12.29). Furthermore, the axial 1-OH �-anomers are
expected to be stronger acids than the equatorial 1-OH �-anomers, and the amount of �-
anomers is expected to be much higher in mannose than in glucose, which may explain the
earlier elution of glucose. Regarding homomorphous hexose and pentose, the pentose
seems to elute earlier (L-arabinose < D-galactose). Yet, xylose which lacks 5-CH2OH
(Figure 3) but otherwise is similar to glucose, elutes later than the more acidic glucose
(pKa 12.35) (Lee, 1990).
D-gluco-pyranose
OH
HH
OHOH
H OH
H
OH
D-xylo-pyranose
O
HH
H
OHOH
H OH
OH
OHO
OHH
OHOH
HOH2C
H
H
L-arabino-furanose
O
HH
OH
HOH
H OH
OH
OH
D-galacto-pyranose
OHO
OHH
H
OHOH
H H
OH
D-manno-pyranose
Figure 3. Structures of L-arabinofuranose, D-galactopyranose, D-mannopyranose, D-glucopyranose and D-xylopyranose.

35
2.4.2 Retention of oligosaccharides
With the oligosaccharides, the position of the glycosidic linkages and the overall
conformational structures become important factors and elements other than acidity of the
1-OH groups have more influence on the elution order. In the study of Roberts et al.
(1971), different pKa values were found for each of the hydroxyl groups. They reported
the acidity of the hydroxyl groups in methyl �- or �-D-glucopyranoside to be 2-OH > 6-
OH > 3-OH > 4-OH. Thus, masking or transformation of some of the hydroxyl groups
influences the elution order of oligosaccharides. Generally the retention volume increases
as the DP increases. The linkage position influences the elution in the following order for
glucose oligosaccharides: �-1,3 > �-1,3 > �-1,4 > �-1,4 > �-1,6 > �-1,2 > �-1,6 (Koizumi
et al., 1991) and therefore the elution order is different for various D-glucose
disaccharides. Since the retention times for oligosaccharides from the Xyl�(1,4) series are
shorter than for the Glc�(1,4) series, and the only difference between the series is the
absence of the -CH2OH group from xylose, it has been concluded that 6-OH may well be
accessible by the resin surface. On the other hand, even with free 6-OH groups the
Man�(1,4) (Figure 3) series elutes even before the corresponding xylose series, and thus it
was suggested that the presence of axial 2-OH somehow hinders the interactions between
the oligosaccharide and the resin (Lee, 1990).
Information on the retention of branched oligosaccharides is limited. However, two factors
are operative in determining the elution order: 1) the relative acidities of the hydroxyl
groups and 2) the accessibility of the oligosaccharide oxyanions to the functional groups
of the stationary phase. Hardy and Townsend (1988) studied the separation of neutral
oligosaccharides from glycoproteins, but the results cannot straightforwardly be applied to
AXOS.
2.4.3 Pulsed amperometric detection
In pulsed amperometric detection, the carbohydrates are oxidized at the gold (Au)
electrode (Johnson and LaCourse, 1990). PAD, with an Au working electrode, was
developed to enable frequent renewal of the working electrode surface. High sensitivity
for anodic detection of carbohydrates at Au electrodes requires pH � 12 (Johnson et al.,

36
1993). Functioning of the electrode can be divided into three steps. During the first step
the carbohydrates are oxidized at the Au electrode, resulting in a specific oxidation
current, and within this step the signal is collected. This is followed by a large positive
potential to achieve anodic formation of surface oxides to clean the electrode. The last
phase is a reactivation step, since the oxidized electrode surface is reduced back to a
reactive Au surface. Oxidation of glucose in the acyclic form on PAD during the first step
is described as follows: a first very rapid two-electron oxidation of the aldehyde to the
corresponding carboxylic acid occurs, followed by relatively rapid steps resulting in
cleavage of the C1-C2 bond with production of CHO2H, after which C2 and C6 are
converted to the corresponding carboxylates. Further sequental anodic cleavage of the
remaining terminal carbons occurs slowly and the electrons released during oxidation are
detected (Johnson and LaCourse, 1990).
PAD does not provide a uniform response to the carbohydrates composed of the same
monosaccharide units but with different structural arrangement of the residues. Therefore
this technique requires a quantification standard for each oligosaccharide to be determined
(Lee, 1990). This is a serious disadvantage, since reference compounds for complicated
oligosaccharides are not available and thus must be self-prepared. Previous information on
responses for oligosaccharides, including AXOS, is limited. When quantitative analyses
are performed, the choice between measuring area or height is a compromise between
several factors. In integrating the peaks, either peak height or peak area can be used when
good resolution and peak shape are achieved. Sometimes peak height is more precise than
area in measuring the quantity, even though area is more widely used in chromatography
(Dyson, 1998). Snyder (1972) showed that height is less affected by overlap than area
with symmetric peaks, while Meyer (1995) demonstrated that height is a safer choice in
measuring small overlapped peaks. Furthermore, integration errors for height
measurements are smaller in almost every situation than the corresponding area
measurements. A shortcoming of using the peak height is that height is a function of peak
width, making this dependent on column efficiency, not the quantity of the measured
molecule. For integration, however, peak height can be recommended in several cases
(Bicking, 2006).

37
2.5 Prebiotics and probiotics
2.5.1 Definition of pro- and prebiotics
Research on prebiotic compounds and probiotic bacteria has been active for over a decade.
A probiotic is “a live microbial food ingredient that is beneficial to health” (Salminen et
al., 1998). The health effects associated with probiotic intake include: relieving of lactose
malabsorption symptoms, improvement of natural resistance to infectious diseases of the
gastrointestinal tract, suppression of cancer, serum cholesterol concentration reduction,
and improved digestion and stimulation of gastrointestinal immunity (Collins and Gibson,
1999).
Prebiotic compounds are targeted to stimulate the growth of the bifidobacteria and
lactobacilli that are recognized as health-promoting probiotics (Gibson and Roberfroid,
1995). Gibson et al. (2004) provided the latest definition for prebiotic compound: "A
prebiotic is a selectively fermented ingredient that allows specific changes, both in the
composition and/or activity in the gastrointestinal microflora that confers benefits upon
host well-being and health." Thus, to evaluate the prebiotic properties of different
compounds, three criteria were established by Gibson et al. (2004): 1) A prebiotic
compound should resist gastric acidity, hydrolysis by mammalian enzymes and
gastrointestinal absorption 2) the intestinal microflora is able to ferment the compound
and 3) a prebiotic compound can selectively stimulate the growth and/or activity of
intestinal bacteria associated with health and well-being.
The human gastrointestinal tract (GIT) is a microecosystem in which beneficial,
potentially pathogenic and harmful bacteria coexist. Normal physiological functions are
enabled, unless harmful and potentially pathogenic bacteria predominate. Thus, the
maintaining of proper equilibrium of the microbiota is important and may be aided, by
using probiotics, prebiotics or their combination, synbiotics, in the diet (Bielecka et al.,
2002).

38
2.5.2 Potential prebiotic (oligo)saccharides
Fructo-oligosaccharides, galacto-oligosaccharides and lactulose are selectively fermented
by probiotics and thus can be called prebiotics (Gibson et al., 2004). In addition, the
prebiotic potentials of several other oligosaccharides have been tested. For example, linear
xylo-oligosaccharides have shown promising results in in vitro and in vivo studies
(Okazaki et al., 1990; Crittenden and Playne, 1996; Vázquez et al., 2000). XOS are
reported to be stable over a wide range of pH values (2.5-8.0), and are indigestible in the
digestive tract (Vázquez et al., 2000). However, linear oligosaccharides may be fermented
rapidly in the colon and thus oligosaccharides with more complex carbohydrate structures
are currently studied to find slower-fermenting prebiotics (Voragen, 1998; Kabel et al.,
2002b). Furthermore, no evidence for a selective stimulation of bifidobacterial growth by
XOS has yet been demonstrated, since in addition to bifidobacteria, Bacteroides,
Clostridium and Lactobacillus are also able to metabolize XOS (Moura et al., 2007).
One potential group is arabinoxylan-derived arabinoxylo-oligosaccharides, which may
have varying chemical structures, depending on the xylan source and the degradation
method used (Grootaert et al., 2007). Both AXOS and AX have shown promising
prebiotic properties in pure culture in vitro studies (van Laere et al., 1997, 1999, 2000;
Palframan et al., 2003; Yuan et al., 2005; Grootaert et al., 2007). Some studies also
investigated the relationship between the structure of AX and the growth of mixed
intestinal bacteria cultures (Karppinen et al., 2001; Hopkins et al., 2003; Hughes et al.,
2007). Since AXOS differ in degree of polymerization and in degree of substitution, the
diversity of these compounds is higher than in other commercial prebiotic carbohydrates.
In evolving prebiotic compounds, the target is to increase the persistence of prebiotics in
the colon, since slow fermentation was related to decreasing the risk of colon cancer
(Grootaert et al., 2007). Until recently, little information has been available on which
AXOS would be the most beneficial. The challenge for these studies is to have enough
purified and well-defined AXOS preparations.
Grootaert et al. (2009) evaluated the prebiotic effect of an AXOS preparation with average
DP of 15 and Ara/Xyl ratio of 0.27. A model of the human intestinal microbial ecosystem
was used and a shift in part of the sugar fermentation toward the distal colon part was
observed with AXOS supplementation. AXOS also had a strong propionate-stimulating

39
effect. Additionally, xylanase-pretreated AX (not characterized) stimulated the colonic
bifidobacteria in the human gut model (Vardakou et al., 2007).
Only a few in vivo fermentation studies on AXOS can be found in the literature. Van
Crayeveld et al. (2008) evaluated the prebiotic potential of wheat bran-derived AXOS in
relation to their structure in rats. AXOS and XOS with DP � 3 resulted in increased
colonic acetate and butyric concentrations and increased amount of bifidobacteria. On the
other hand, AXOS with average DP of 61 effectively suppressed branched short-chain
fatty acid concentrations, thus shifting the balance away from protein fermentation, but the
long-chain AXOS did not increase the butyrate concentration or stimulate faecal
bifidobacteria development. AXOS with DP of 12, Ara/Xyl ratio of 0.69 and AXOS with
DP of 15, Ara/Xyl ratio of 0.27 affected similarly, suggesting that the influence of DP was
more pronounced. The best gut health effects were obtained with AXOS of DP 5 and
Ara/Xyl ratio of 0.27. Cloetens et al. (2008) studied the effect of AXOS preparation
produced from wheat on colonic metabolism in human volunteers. The results from the
study suggest that a minimal dose of 2.2 g of AXOS (DP 15, Ara/Xyl ratio 0.26)
favourably modulates colonic metabolism in healthy humans. However, long-term studies
are still required to confirm the results.
2.5.3 Production of SCFA during fermentation of prebiotics
The fermentation of short oligosaccharides (prebiotics) in the large intestine by colonic
bacteria leads, in addition to microbial growth, to the production of CO2, H2, SCFA and
lactate. SCFA and lactate can be metabolized further to provide energy to the host. In
addition, many health effects have been reported for SCFA, such as improvement in bowel
function, calcium absorption, lipid metabolism and reduction of the risk of colon cancer
(Scheppach et al., 2001; Scharlau et al., 2009). The production of unbranched SCFA
(acetate, propionate, and butyrate) is desired, since they are able to lower the pH in the
intestine and inhibit the growth of potentially harmful bacteria (Campbell et al., 1997a;
Wong et al., 2006). Butyrate was especially able to inhibit the growth of colonic
carcinoma cells in in vitro and in vivo experiments. Butyric acid formation during colonic
fermentation also appears to correlate with the cancer-suppressing properties of dietary
fibre (Scheppach et al., 1995; Wollowski et al., 2001). Some authors refer that butyrate

40
can also be produced from lactate by some intestinal bacteria (Duncan et al., 2004). In
contrast, the concentrations of branched SCFA (isobutyrate, isovalerate) should be
reduced, since they are formed during catabolism of proteins into potentially toxic
catabolites associated with bowel cancer (Visek, 1978; Van Craeyveld et al., 2008).
Probiotics may in some cases be able to suppress protein fermentation in the colon (De
Preter et al., 2004).
2.5.4 Intestinal microbiota
The intestinal microbiota play an important role in the maintainance of health and in
prevention of disease, which is well recognized (Holzapfel and Schillinger, 2002). In the
GIT there are more than 400 bacterial species, of which the lactic-acid bacteria constitute
approximately 10%. Gram-positive anaerobic Bacteroides, Eubacterium, and
Bifidobacterium species predominate in the large intestine, but there are also other
important groups such as clostridia and lactobacilli (Kontula et al., 2000; Holzapfel and
Schillinger, 2002). The presence of bifidobacteria in the GIT was associated with
beneficial health effects (Modler, 1994). The growth of these bacteria is linked with their
ability to produce proteins, such as extra- and intracellular hydrolytic enzymes, as well as
mono- and oligosaccharide transporters involved in the metabolism of nondigestive
carbohydrates (van den Broek et al., 2008).
2.5.5 Bifidobacterial enzymes in AXOS utilization
The genome sequences of B. longum NCC 2705 and B. adolescentis ATCC 15703 have
revealed high levels of proteins related to the utilization of nondigestive polysaccharides
and oligosaccharides, including putative �-D-xylosidases and �-L-arabinofuranosidases
for degradation of AXOS (Schell et al., 2002; van den Broek et al., 2008). Thus far,
bifidobacteria are not known at the protein level to produce endo-1,4-�-D-xylanase, but
one putative xylanase-encoding gene (GH family 8) was isolated (van den Broek et al.,
2005). Thus, other microorganisms in the GIT are needed for the hydrolysis of polymeric
nondigestive AX into oligosaccharides. The key enzymes for utilization of AXOS are the
�-L-arabinofuranosidases. Bifidobacterium adolescentis ATCC 15703 harbours genes for
two extracellular polymeric xylan-acting �-L-arabinofuranosidases: AXH-m (GH family

41
51) and AXH-d3 (GH family 43), which are able to release �-L-Araf from mono- and
doubly substituted �-D-Xylp residues, respectively (van Laere et al., 1999; van den Broek
et al., 2005, 2008). Surprisingly, the B. longum NCC 2705 genes annotated into familes
GH43 and GH51 have not revealed significant homology with currently known
extracellular �-L-arabinofuranosidases (van den Broek et al., 2008), although several B.
longum strains were able to grow on rye arabinoxylan, presumably by releasing and
metabolizing �-L-Araf substituents (Crittenden et al., 2002). Indeed, many other B.
longum strains evidently have GH family 51 �-L-arabinofuranosidase (Gueimonde et al.,
2007). The genome sequences of Clostridium and Lactobacillus strains have until now
revealed no enzymes in GH families 43 and 51 containing �-L-arabinofuranosidases and
�-xylosidases (van den Broek et al., 2008), which further indicates the potential use of
AXOS as prebiotic oligosaccharides with bifidobacterial selectivity.
Despite several studies suggesting that AXOS and, especially, various AX hydrolysates
are potential prebiotics (Grootaert et al., 2007; Vardakou et al., 2008), there is a lack of
information on the influence of the molecular structure of the putative prebiotic AXOS on
the growth of probiotic bifidobacteria. Table 4 summarizes the growth of several
bifidobacterial strains on L-arabinose, D-xylose, XOS, AXOS and (arabino)xylan
substrates. Usually the growth was determined by measuring the optical density (OD) and
detecting SCFA production in the fermentation samples. HPAEC-PAD was also used in
some studies to determine the utilization of carbohydrates (Crittenden et al., 2002; van
Laere et al., 2000; Moura et al., 2007; Gullón et al., 2008).

42
Tab
le 4
. G
row
th o
f va
rious
bifi
doba
cter
ial
stra
ins
on L
-ara
bino
se,
D-x
ylos
e, x
ylo-
olig
osac
char
ides
(X
OS)
, ar
abin
oxyl
o-ol
igos
acch
arid
es
(AX
OS)
and
(ara
bino
)xyl
an. +
= g
row
th, -
= n
o gr
owth
, n.a
. = n
ot a
naly
sed.
Org
anis
m
Subs
trat
e R
efer
ence
L
-ara
bino
se
D-x
ylos
e X
OS
AX
OS
(Ara
bino
)xyl
an
B. a
dole
scen
tis A
TCC
157
03
- -
n.a.
n.
a.
n.a.
R
oy a
nd W
ard,
199
0 B.
ado
lesc
entis
ATC
C 1
5703
n.
a.
n.a.
n.
a.
+ n.
a.
van
Laer
e et
al.,
199
7 B.
ado
lesc
entis
ATC
C 1
5703
-
+ +
+ -
van
Laer
e et
al.,
199
9 B.
ado
lesc
entis
ATC
C 1
5703
n.
a.
n.a.
+
+ -
van
Laer
e et
al.,
200
0 B.
ado
lesc
entis
ATC
C 1
5703
n.
a.
n.a.
+
n.a.
-
Crit
tend
en e
t al.,
200
2 B.
ado
lesc
entis
ATC
C 1
5703
n.
a.
+ +
n.a.
-
Palfr
aman
et a
l., 2
003
B. a
dole
scen
tis A
TCC
157
03
+ +
+ n.
a.
n.a.
M
oura
et a
l., 2
007
B. a
dole
scen
tis A
TCC
157
03
n.a.
n.
a.
+ n.
a.
n.a.
G
ulló
n et
al.,
200
8 B.
ado
lesc
entis
VTT
E-9
9143
6a +
+ +
n.a.
+
Crit
tend
en e
t al.,
200
2 B.
ang
ulat
um A
TCC
275
35
+ +
n.a.
n.
a.
n.a.
R
oy a
nd W
ard,
199
0 B.
ang
ulat
um A
TCC
275
35
n.a.
n.
a.
+ n.
a.
- C
ritte
nden
et a
l., 2
002
B. a
ngul
atum
ATC
C 2
7535
n.
a.
- -
n.a.
-
Palfr
aman
et a
l., 2
003
B. b
ifidu
m A
TCC
295
21
- -
n.a.
n.
a.
n.a.
R
oy a
nd W
ard,
199
0 B.
bifi
dum
ATC
C 2
9521
n.
a.
n.a.
+
n.a.
-
Crit
tend
en e
t al.,
200
2 B.
bifi
dum
ATC
C 2
9521
n.
a.
- +
n.a.
+
Palfr
aman
et a
l., 2
003
B. b
ifidu
m F
-35b
- -
n.a.
+
n.a.
Y
uan
et a
l., 2
005
B. b
reve
ATC
C 1
5700
-
- n.
a.
n.a.
n.
a.
Roy
and
War
d, 1
990
B. b
reve
ATC
C 1
5700
n.
a.
n.a.
n.
a.
- -
van
Laer
e et
al.,
200
0 B.
bre
ve A
TCC
157
00
n.a.
n.
a.
+ n.
a.
- C
ritte
nden
et a
l., 2
002
B. b
reve
ATC
C 1
5700
n.
a.
- -
n.a.
+
Palfr
aman
et a
l., 2
003
B. b
reve
ATC
C 1
5700
n.
a.
n.a.
+
n.a.
n.
a.
Gul
lón
et a
l., 2
008
B. b
reve
CIP
64.
68c
n.a.
n.
a.
(+)
n.a.
-
Crit
tend
en e
t al.,
200
2 B.
cat
enul
atum
ATC
C 2
7539
-
- n.
a.
n.a.
n.
a.
Roy
and
War
d, 1
990
B. c
aten
ulat
um A
TCC
275
39
n.a.
+
+ n.
a.
+ Pa
lfram
an e
t al.,
200
3
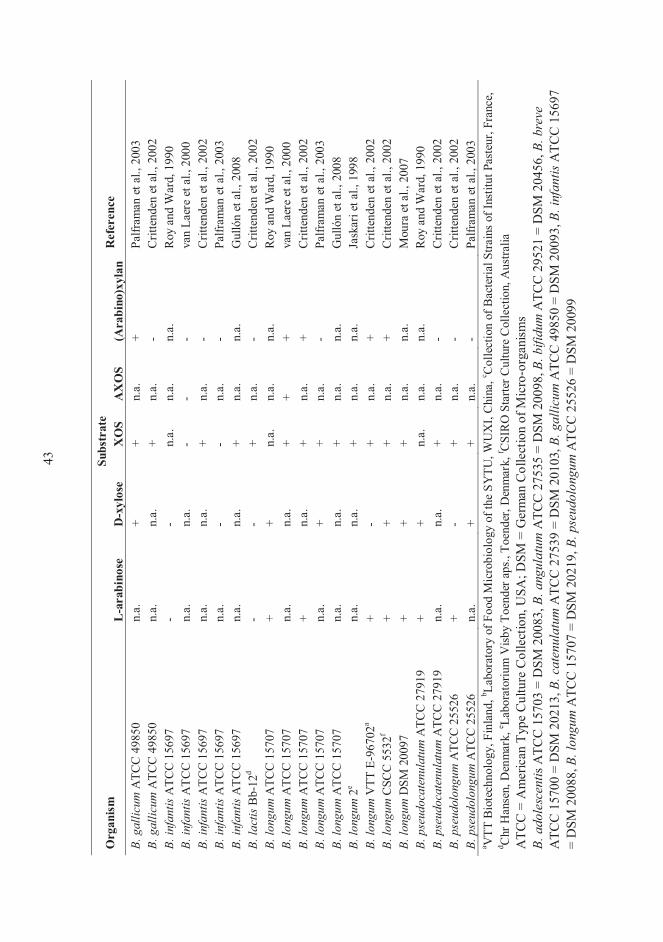
43
Org
anis
m
Subs
trat
e R
efer
ence
L
-ara
bino
se
D-x
ylos
e X
OS
AX
OS
(Ara
bino
)xyl
an
B. g
allic
um A
TCC
498
50
n.a.
+
+ n.
a.
+ Pa
lfram
an e
t al.,
200
3 B.
gal
licum
ATC
C 4
9850
n.
a.
n.a.
+
n.a.
-
Crit
tend
en e
t al.,
200
2 B.
infa
ntis
ATC
C 1
5697
-
- n.
a.
n.a.
n.
a.
Roy
and
War
d, 1
990
B. in
fant
is A
TCC
156
97
n.a.
n.
a.
- -
- va
n La
ere
et a
l., 2
000
B. in
fant
is A
TCC
156
97
n.a.
n.
a.
+ n.
a.
- C
ritte
nden
et a
l., 2
002
B. in
fant
is A
TCC
156
97
n.a.
-
- n.
a.
- Pa
lfram
an e
t al.,
200
3 B.
infa
ntis
ATC
C 1
5697
n.
a.
n.a.
+
n.a.
n.
a.
Gul
lón
et a
l., 2
008
B. la
ctis
Bb-
12d
- -
+ n.
a.
- C
ritte
nden
et a
l., 2
002
B. lo
ngum
ATC
C 1
5707
+
+ n.
a.
n.a.
n.
a.
Roy
and
War
d, 1
990
B. lo
ngum
ATC
C 1
5707
n.
a.
n.a.
+
+ +
van
Laer
e et
al.,
200
0 B.
long
um A
TCC
157
07
+ n.
a.
+ n.
a.
+ C
ritte
nden
et a
l., 2
002
B. lo
ngum
ATC
C 1
5707
n.
a.
+ +
n.a.
-
Palfr
aman
et a
l., 2
003
B. lo
ngum
ATC
C 1
5707
n.
a.
n.a.
+
n.a.
n.
a.
Gul
lón
et a
l., 2
008
B. lo
ngum
2e
n.a.
n.
a.
+ n.
a.
n.a.
Ja
skar
i et a
l., 1
998
B. lo
ngum
VTT
E-9
6702
a +
- +
n.a.
+
Crit
tend
en e
t al.,
200
2 B.
long
um C
SCC
553
2f +
+ +
n.a.
+
Crit
tend
en e
t al.,
200
2 B.
long
um D
SM 2
0097
+
+ +
n.a.
n.
a.
Mou
ra e
t al.,
200
7 B.
pse
udoc
aten
ulat
um A
TCC
279
19
+ +
n.a.
n.
a.
n.a.
R
oy a
nd W
ard,
199
0 B.
pse
udoc
aten
ulat
um A
TCC
279
19
n.a.
n.
a.
+ n.
a.
- C
ritte
nden
et a
l., 2
002
B. p
seud
olon
gum
ATC
C 2
5526
+
- +
n.a.
-
Crit
tend
en e
t al.,
200
2 B.
pse
udol
ongu
m A
TCC
255
26
n.a.
+
+ n.
a.
- Pa
lfram
an e
t al.,
200
3 a V
TT B
iote
chno
logy
, Fin
land
, b Labo
rato
ry o
f Foo
d M
icro
biol
ogy
of th
e SY
TU, W
UX
I, C
hina
, c Col
lect
ion
of B
acte
rial S
train
s of
Inst
itut P
aste
ur, F
ranc
e,
d Chr
Han
sen,
Den
mar
k, e La
bora
toriu
m V
isby
Toen
der a
ps.,
Toen
der,
Den
mar
k, f C
SIR
O S
tarte
r Cul
ture
Col
lect
ion,
Aus
tralia
A
TCC
= A
mer
ican
Typ
e C
ultu
re C
olle
ctio
n, U
SA; D
SM =
Ger
man
Col
lect
ion
of M
icro
-org
anism
s B.
ado
lesc
entis
ATC
C 1
5703
= D
SM 2
0083
, B. a
ngul
atum
ATC
C 2
7535
= D
SM 2
0098
, B. b
ifidu
m A
TCC
295
21 =
DSM
204
56, B
. bre
ve
ATC
C 1
5700
= D
SM 2
0213
, B. c
aten
ulat
um A
TCC
275
39 =
DSM
201
03, B
. gal
licum
ATC
C 4
9850
= D
SM 2
0093
, B. i
nfan
tis A
TCC
156
97
= D
SM 2
0088
, B. l
ongu
m A
TCC
157
07 =
DSM
202
19, B
. pse
udol
ongu
m A
TCC
255
26 =
DSM
200
99

44
3. AIMS OF THE STUDY
The overall aim of this thesis was to prepare and purify different short-chain arabinoxylo-
oligosaccharides (AXOS) from cereal arabinoxylans (AX) and to elucidate the structures
and to study prebiotic potential of the AXOS manufactured. Enzyme action on AX was
investigated through AXOS produced in the hydrolysis, and the studies were also carried
out to obtain information on the behaviour of structurally different AXOS in
chromatography.
The main objectives were:
- To enzymatically prepare short-chain AXOS and separate them from the other hydrolysis
products, using chromatographic methods (I-III)
- To fully analyse the structures of the purified AXOS (I-III)
- To evaluate the function of the Shearzyme used in the enzymatic hydrolysis (I-III)
- To specify the elution order of the AXOS and determine the responses of the different
AXOS on the HPAEC-PAD system used (II, III)
- To determine the structure of several different cereal AX (I-III) and the occurrence of �-
D-Xylp-(1�2)-�-L-Araf-(1�3) side chain in them (III)
- To study the prebiotic potential of the AXOS in in vitro studies, and in particular to
examine the influence of specific �-L-Araf substitutions on growth of bifidobacteria (IV)

45
4. MATERIALS AND METHODS
This section summarizes the materials and methods presented in more detail in the original
articles I-IV.
4.1 Materials
4.1.1 Carbohydrates
The arabinoxylans used in studies I-IV are listed in Table 5. Some of the AX used were
purchased and a few were obtained as gifts. The remaining AX were extracted from
different cereal samples. The AX were used as starting materials for preparation of AXOS
(I-IV) and in in vitro fermentation studies (IV) for substrate production.
Table 5. Arabinoxylans used in studies I-IV. Origin Abbr. Manufacturer Study Rye flour (high-viscosity) RAX Megazyme
(Bray, Ireland) I, IV
Wheat flour (high-viscosity) WAX-hv Megazyme I Wheat flour (medium-viscosity) WAX-mv Megazyme II, IV Wheat flour (low-viscosity) WAX-lv Megazyme I Oat spelts OSAX Sigma-Aldrich Chemie
(Steinheim, Germany) I, III
Corncob CCAX Dr. Ramírez/Dr. Saake (University of Guadalajara, Mexico /vTI, Hamburg, Germany)
III
Wheat straw WStAX VTT, Finland/Dr. Jürgen Puls (vTI) III Arabinoxylan origin Abbr. Manufacturer of cereal product Study Barley husks BHAX* Husks from Altia Corp.
(Koskenkorva, Finland) III
Oat brans OBAX* Brans from Raisio Oyj (Raisio, Finland)
III
Rice husks RiHAX* Husks from Prof. Parajó (University of Vigo, Spain)
III
Rye brans RBAX* Brans from Korpela Mill (Jalasjärvi, Finland)
III
Wheat brans WBAX* Brans from Raisio III *Arabinoxylans were extracted in our laboratory
Monosaccharides, L(+)-arabinose, D(+)-xylose, and D(+)-glucose from Merck
(Darmstadt, Germany) and linear XOS (1,4-�-D-xylobiose, 1,4-�-D-xylotriose and 1,4-�-
D-xylotetraose) from Megazyme were used as standards in thin layer chromatography

46
(TLC) and HPAEC-PAD analysis (I-IV). The preceding monosaccharides were also used
as standards in gas chromatography (GC) analysis (I-III). XOS from Wako Pure Chemical
Industries (Osaka, Japan), FOS from BioCare (Birmingham, UK) and monosaccharides
(arabinose and xylose) were used in the fermentation studies (V).
4.1.2 Enzymes
The commercial xylanase preparation Shearzyme®, which contains the Aspergillus
aculeatus GH family 10 endo-1,4-�-D-xylanase as the main enzyme (49 100 nkat/ml), was
used to prepare short-chain AXOS from the AX (I-IV). Shearzyme 500 L was obtained
from Novozymes A/S. The purified enzymes were used to produce various AXOS from
AX and to evaluate the structures of the AXOS formed with enzyme-aided methods (I-
IV). The purified enzymes used are listed in Table 6.
Table 6. Purified enzymes used in studies I-III. Purified enzyme Organism EC
Number GH Family
Activity (nkat/ml) Source Study
AXH-m (E-AFASE) Aspergillus niger 3.2.1.55 54* 6970 Megazyme I
AXH-d3 (E-AFAM2)
Bifidobacterium adolescentis 3.2.1.55 43 3340 Megazyme II
�XH-m Unknown 3.2.1.55 14590 Novozymes II, III �-xylosidase Trichoderma reesei 3.2.1.37 3 2340 VTT I endoxylanase Aspergillus oryzae 3.2.1.8 10* 1850 VTT I endoxylanase Aspergillus aculeatus 3.2.1.8 10 121530 Novozymes I exoxylanase Trichoderma reesei 3.2.1.156 5 9.7† VTT II * Belong to certain GH families, based on their action; † = mg protein/ml AXH-m and AHX-d3 are �-L-arabinofuranosidases (Table 6); AXH-m acts on both
(1�2)- and (1�3)-linked Araf units on monosubstituted �-D-Xylp residues, while AXH-
d3 is able to release only (1�3)-linked �- L-Araf units from disubstituted �-D-Xylp
residues. �-Xylosidase (exo-1,4-�-D-xylosidase) hydrolyses the xylose chain from the
nonreducing end, and xylanases (endo-1,4-�-D-xylanases) act in the middle of the xylose
chain. The GH family 5 xylanase is an exoenzyme that releases xylose units from the
reducing end of the xylose chain.

47
4.1.3 Bacteria used in the fermentation experiments
The pure bacterial strains used in study IV were B. adolescentis ATCC 15703 (DSM
20083), B. breve ATCC 15700 (DSM 20213) and B. longum ATCC 15707 (DSM 20219)
purchased from DSMZ (Braunsweig, Germany). For faecal microbiota fermentation
studies, the human faeces samples were obtained from a single healthy donor on four
occasions. The faeces samples were mixed in sterilized anaerobic 0.1 M PBS buffer
(phosphate-buffered saline). All the bacteria (pure strains of bifidobacteria and bacteria
from faecal microbiota) were cultivated anaerobically in nutrient-poor BA media
described by Angelidaki et al. (1990) with the modification that yeast extract (1 g/l;
Merck) was added to the medium. The vitamin solution was prepared according to Wolin
et al. (1963) and added prior to inoculation.
4.2 Experimental
4.2.1 Extraction of AX (III)
The arabinoxylans were either purchased or obtained as purified, or extracted from various
cereal materials in our laboratory (Table 5). The AX (III)were extracted from rye, wheat
and oat brans and rice husks with saturated Ba(OH)2 after enzymatic pretreatment by a
method described previously by Virkki et al. (2005). Barley husk AX was previously
extracted by Pitkänen et al. (2008), using the same method.
4.2.2 Monosaccharide composition analysis of AX (I, III)
To evaluate the monosaccharide composition and to calculate the Ara/Xyl ratio of the
isolated polymeric AX, two different methods were used to degrade AX into
monosaccharides: 1) sulphuric acid (H2SO4) hydrolysis (I) and 2) acid methanolysis (III).
The hydrolysis conditions for AX degradation by H2SO4 were chosen in the preliminary
tests (results not shown).

48
1) H2SO4 hydrolysis:
Rye flour AX was hydrolysed using 1 M H2SO4 for 30 min at 120 oC (I). The H2SO4
method was modified from that used by Lebet et al. (1997). The hydrolysis were carried
out with four samples in parallel. After the hydrolysis, the pH was adjusted to 7-9 with 10
M NaOH. The standards (25 mg/ml) D-xylose, L-arabinose and D-glucose, were treated
the same way as the AX samples. The dilutions for the standard curve samples were made
from these monosaccharide solutions. The HPAEC-PAD method for monosaccharide
analysis (section 4.2.6.2) was used to analyse the samples.
2) Acid methanolysis:
Wheat, rye and oat bran AX, rice and barley husk AX, oat spelt AX and wheat straw AX
were degraded into monosaccharides, using acid methanolysis (III) as described by
Sundberg et al. (1996). AXOS were also degraded into monosaccharides, using acid
methanolysis (III). The silylated monosaccharides were analysed with GC (section
4.2.6.3). The monosaccharide standards were also treated with acid methanolysis.
4.2.3 Enzymatic production and chromatographic purification of AXOS (I-III)
AXOS were produced enzymatically from the extracted AX on a preparative scale. The
commercial enzyme Shearzyme was used to hydrolyse the linear AX main chain (I-III).
The AX (5 g/l; 1.5 g of rye flour AX (I) or 1.5 g of wheat flour AX (II) or 5 g of oat spelt
AX (III)) in 20 mM sodium acetate (NaOAc) buffer, pH 5.0, were incubated with
Shearzyme (10 000 nkat/g AX (I, II) and 50 000 nkat/g AX (III)) at 40 oC for 48 h. The
enzymes were used stepwise for the production of AXOS (II). The substitution pattern of
AX was first modified using �-arabinofuranosidase AXH-m (5000 nkat/g AX at 40 oC for
24 h, pH 5) followed by Shearzyme treatment. Some of the L-Araf substituents were
further removed by AXH-d3 treatment (1000 nkat/g AX at 40 oC for 24 h, pH 6.5). All the
hydrolyses were terminated by keeping the samples in a boiling-water bath for 10 min.
After the enzymatic treatments, the hydrolysis products were separated (I-III) according
to Figure 4.
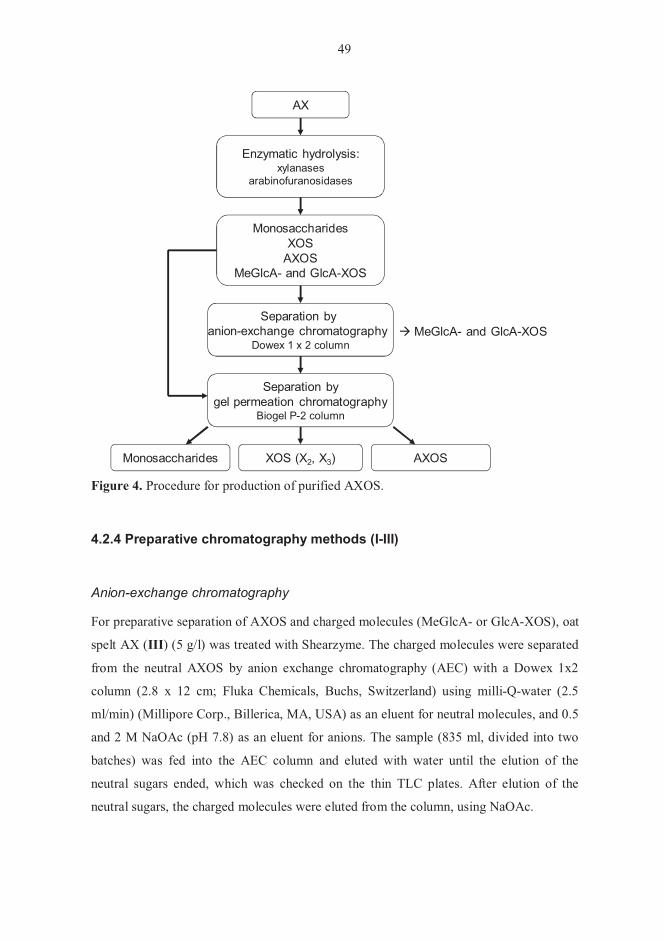
49
� MeGlcA- and GlcA-XOS
AX
Enzymatic hydrolysis:xylanases
arabinofuranosidases
MonosaccharidesXOS
AXOSMeGlcA- and GlcA-XOS
Separation bygel permeation chromatography
Biogel P-2 column
Separation byanion-exchange chromatography
Dowex 1 x 2 column
Monosaccharides XOS (X2, X3) AXOS
Figure 4. Procedure for production of purified AXOS.
4.2.4 Preparative chromatography methods (I-III)
Anion-exchange chromatography
For preparative separation of AXOS and charged molecules (MeGlcA- or GlcA-XOS), oat
spelt AX (III) (5 g/l) was treated with Shearzyme. The charged molecules were separated
from the neutral AXOS by anion exchange chromatography (AEC) with a Dowex 1x2
column (2.8 x 12 cm; Fluka Chemicals, Buchs, Switzerland) using milli-Q-water (2.5
ml/min) (Millipore Corp., Billerica, MA, USA) as an eluent for neutral molecules, and 0.5
and 2 M NaOAc (pH 7.8) as an eluent for anions. The sample (835 ml, divided into two
batches) was fed into the AEC column and eluted with water until the elution of the
neutral sugars ended, which was checked on the thin TLC plates. After elution of the
neutral sugars, the charged molecules were eluted from the column, using NaOAc.

50
Gel permeation chromatography
For preparative isolation of AXOS, rye flour AX (I), wheat flour AX (II), and oat spelt
AX (III) (5 g/l) were treated with Shearzyme. AEC was used for the oat spelt AX
hydrolysis sample (III) to separate MeGlA- and GlCA-XOS from the neutral
carbohydrates before the gel permeation chromatography (GPC). For GPC, each
hydrolysed sample was concentrated to a volume of 40-50 ml with rotavapour. The AXOS
were separated by a Biogel P2 column (8.9 x 135 cm, BioRad, Hercules, CA, USA) eluted
with milli-Q-water at 10 ml/min and 27-ml fractions were collected (I, II). Several 5-ml
fractions were collected after separation of AXOS with a smaller Biogel P2 column (5 x
95 cm; Bio-Rad), using milli-Q-water (2 ml/min) as eluent (III). Mono- and
oligosaccharides were detected from the fractions with TLC and HPAEC-PAD (I-III).
4.2.5 Quantification of AXOS (I-III)
The isolated AXOS were hydrolysed into monosaccharides with �-L-arabinofuranosidase
(AXH-m; 5000 nkat/g of substrate) and �-xylosidase (1000 nkat/g substrate) for 24 h at 40 oC in 20 mM NaOAc buffer, pH 5.0 (I). The enzyme mixture (II) was used to hydrolyse
AXOS totally into monosaccharides, as described by Virkki et al. (2008). The enzyme
mixture used was dosed according to the �-L-arabinofuranosidase activity (1000 nkat/g
AX), which was regarded as the limiting enzyme activity in the mixture. Acid
methanolysis was used (III) for degrading AXOS, as described above for AX degradation.
After degradation treatment of the purified AXOS, the samples were analysed first with
the HPAEC-PAD method for oligosaccharides to check the completeness of the
hydrolysis. Thereafter, the monosaccharides were analysed with the HPAEC-PAD method
for monosaccharides (I) or with GC (II, III) as described later.
The Ara/Xyl ratio was determined from the results of the HPAEC-PAD or GC analysis.
The concentrations of the AXOS were calculated, based on the monosaccharide yield and
the known AXOS structure. After quantification of the AXOS sample, dilutions for the
standard curve samples were made from the quantified sample. AXOS standard curves
were used late to quantify the corresponding AXOS from other samples.

51
4.2.6 Analytical chromatographic methods (I-IV)
TLC, HPAEC-PAD and GC were used to separate different mono-, di- and
oligosaccharides. Furthermore, after complete structural analysis on mass spectrometry
(MS) and nuclear magnetic resonance (NMR) spectroscopy, chromatographic methods
were used to identify various compounds based on their retention times in HPAEC-PAD
or retention factors (Rf) on TLC.
4.2.6.1 TLC
The TLC was carried out (I-III), using silica gel 60 plates (Merck). A mixture of 1-
butanol/ethanol/water (3:2:2 v/v) was used as an eluent. The carbohydrates were detected
by spraying with 2% (w/v) orsinol dissolved in a solution of ethanol/H2SO4/H2O
(80:10:10 vol%), after which the plates were heated at 105 °C for 10 min.
4.2.6.2 HPAEC-PAD
HPAEC-PAD was used to analyse the mono- and oligosaccharides after enzymatic or acid
hydrolysis of AX (I-IV), to identify and quantify the purified AXOS (I-III), and to
compare the carbohydrates before and after fermentation experiments (IV). For
monosaccharides HPAEC-PAD was used only in I, since in later studies the
monosaccharides were analysed using GC. For oligosaccharides, the HPAEC-PAD
method is invaluable and was used in all studies (I-IV).
The HPAEC-PAD system was equipped with an SSI pulse equalizer (Scientific Systems,
Inc., model LP 21; State College, PA, USA), two Waters 515 HPLC pumps, a PC Waters
pump control module and a cooling Waters 717 autosampler using Millenium32 software
(Waters Corporation, Milford, MA, USA) for instrument control and data handling. The
analytical CarboPac PA-1 (I) and CarboPac PA-100 (I-IV) columns (250 x 4 mm, i.d.),
and the guard column PA-1 (I) and PA-100 (I-IV), respectively, (25 x 3 mm, i.d.)
(Dionex, Sunnyvale, CA, USA) were maintained at 30 oC. All the samples were filtered,
using a 0.45-�m Acrodisc® syringe filter with nylon membrane (Pall Corporation, Ann
Arbor, MI, USA) and the injection volume was typically 10 �l in the measurements.
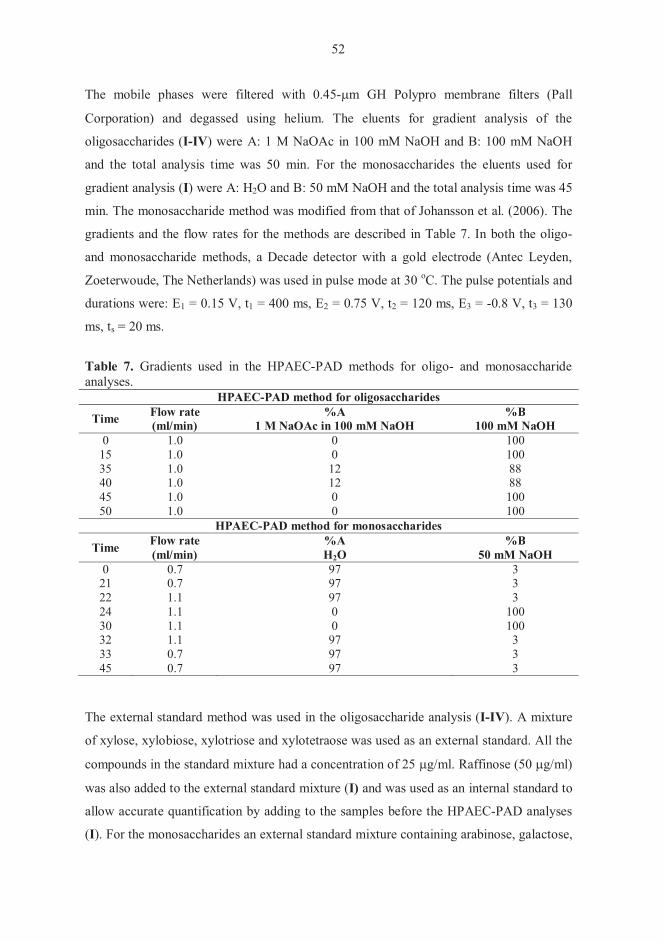
52
The mobile phases were filtered with 0.45-�m GH Polypro membrane filters (Pall
Corporation) and degassed using helium. The eluents for gradient analysis of the
oligosaccharides (I-IV) were A: 1 M NaOAc in 100 mM NaOH and B: 100 mM NaOH
and the total analysis time was 50 min. For the monosaccharides the eluents used for
gradient analysis (I) were A: H2O and B: 50 mM NaOH and the total analysis time was 45
min. The monosaccharide method was modified from that of Johansson et al. (2006). The
gradients and the flow rates for the methods are described in Table 7. In both the oligo-
and monosaccharide methods, a Decade detector with a gold electrode (Antec Leyden,
Zoeterwoude, The Netherlands) was used in pulse mode at 30 oC. The pulse potentials and
durations were: E1 = 0.15 V, t1 = 400 ms, E2 = 0.75 V, t2 = 120 ms, E3 = -0.8 V, t3 = 130
ms, ts = 20 ms.
Table 7. Gradients used in the HPAEC-PAD methods for oligo- and monosaccharide analyses.
HPAEC-PAD method for oligosaccharides
Time Flow rate (ml/min)
%A 1 M NaOAc in 100 mM NaOH
%B 100 mM NaOH
0 1.0 0 100 15 1.0 0 100 35 1.0 12 88 40 1.0 12 88 45 1.0 0 100 50 1.0 0 100
HPAEC-PAD method for monosaccharides
Time Flow rate (ml/min)
%A H2O
%B 50 mM NaOH
0 0.7 97 3 21 0.7 97 3 22 1.1 97 3 24 1.1 0 100 30 1.1 0 100 32 1.1 97 3 33 0.7 97 3 45 0.7 97 3
The external standard method was used in the oligosaccharide analysis (I-IV). A mixture
of xylose, xylobiose, xylotriose and xylotetraose was used as an external standard. All the
compounds in the standard mixture had a concentration of 25 �g/ml. Raffinose (50 �g/ml)
was also added to the external standard mixture (I) and was used as an internal standard to
allow accurate quantification by adding to the samples before the HPAEC-PAD analyses
(I). For the monosaccharides an external standard mixture containing arabinose, galactose,

53
glucose, xylose (50 �g/ml), and fructose and mannose (100 �g/ml) was used; an internal
standard was not used in the monosaccharide analysis (I). The external standards were
injected after every three samples in both methods to monitor the retention times and the
detection responses.
For identification and quantification of the respective saccharides in the hydrolysates (I-
IV) and samples after fermentations (IV), the commercial monosaccharides X2, X3 and X4
were used. Identification and quantification of AXOS (I-IV) was done, using pure A3X (I-
IV), A2X, A2XX, A2+3X, A2+3XX (II-IV), D2,3X (III) (chemical structures of AXOS are
presented in section 5.2, Figure 6). Standard curves were made for quantification from
each standard compound with duplicate or triplicate samples at 4-6 concentration levels,
as described in section 4.2.4. The peak heights (mV) were used in quantification of the
carbohydrates. XA3X and U4m2XX were used as standards for identification (III), and
XA3XX to determine the elution order of AXOS in HPAEC-PAD. Purified XA3X and
XA3XX were obtained from rye flour AX that was hydrolysed with GH11 T. reesei endo-
1,4-�-D-xylanase and U4m2XX from birch xylan hydrolysed with Shearzyme.
4.2.6.3 GC
GC analysis of monosaccharides
GC methods were used to analyse the monosaccharide composition of AX after acid
methanolysis, according to the method of Sundberg et al. (1996) (I, III), and after total
enzymatic hydrolysis of AX, using the method of Virkki et al. (2008) (IV) (Table 8). GC
methods were also used to analyse the monosaccharides after degradation of AXOS (II,
III). The monosaccharides obtained with acid methanolysis were silylated and sorbitol
was used as an internal standard. The monosaccharides obtained from the enzymatic
hydrolysis were reduced and acetylated, and myoinositol was used as an internal standard.
The monosaccharide derivatives were then analysed with GC, using one of the three
methods presented in Table 8. Quantification was performed, using six concentration
levels of each sugar (arabinose, xylose and glucose) and all the samples and standards
were analysed in duplicate or triplicate.

54
Table 8. Summary of the GC methods used in the monosaccharide composition analysis of AX and AXOS (I-III). I II III Sample-degrading method
Acid methanolysis (Sundberg et al., 1996)
Enzymatic hydrolysis (Virkki et al., 2008)
Acid methanolysis (Sundberg et al., 1996)
Monosaccharide pretreatment
Silylation Acetylation Silylation
GC instrument Hewlett Packard HP5859 series II system
Hewlett Packard HP5859 series II system
Agilent 6890N system
Detector FID FID FID Injector/autosampler HP7673 series
injector and autosampler (Hewlett Packard)
HP7673 series injector and autosampler (Hewlett Packard)
Agilent 7683 series injector and autosampler
Column HP-5 (30 m, 0.32-mm i.d., 0.25-�m film thickness; Agilent Technologies)
HP-5 (30 m, 0.32-mm i.d., 0.25-�m film thickness; Agilent Technologies)
DB-1 (30 m, 0.25-mm i.d., 0.25-�m film thickness; Agilent Technologies)
Temperature programme
Gradient system: 150 oC for 5 min, 2 oC/min to 260 oC
Isothermal programme: 200 oC for 20 min
Gradient system: 150 oC for 3 min 1) 2 oC/min from 150 oC to 186 oC 2) 1 oC/min from 186 oC to 200 oC 3) 20 oC/min from 200 oC to 325 oC
Run time 60 min 20 min 41.25 min Injector temperature 225 oC 225 oC 225 oC Detector temperature 280 oC 280 oC 280 oC Injection volume 1 �l 1 �l 1 �l Split ratio 1:30 1:30 1:20 Flow rate 1 ml/min 1 ml/min 0.8 ml/min Carrier gas He He He
GC analysis of VFA
Volatile fatty acids (VFA) were analysed with GC (IV). Prior to quantification of VFA the
samples were acidified to pH < 2 with 17% phosphoric acid (H3PO4) and then analysed
using GC (Hewlett Packard 5890 series II; Hewlett Packard, Palo Alto, CA, USA) with a
flame ionization detector (FID) and a capillary column (Hewlett Packard FFAP 30 m,

55
inner diameter 0.53 mm, film 1 �m). The temperatures of the injector and detector were
175 oC and 200 oC, respectively. The temperature gradient was as follows: the initial
temperature was 110 oC, ramped at 5 oC/min to 140 oC, maintained at 140 oC for 6 min,
ramped at 45 oC/min to 200 oC and maintained at 200 oC for 3 min. The injection volume
of the sample was 10 �l. The quantified VFA were analysed as acids: acetic, propionic,
isobutyric, butyric, isovaleric and valeric acids. The standards used in the VFA analyses
were the same acids as mentioned above with concentrations of 1-40 m�. Later these
compounds are referred to as VFA although they appeared as acid anions (acetate,
propionate, isobutyrate, butyrate, isovalerate and valerate) at physiological pH.
4.2.7 Structural elucidation of AXOS (I-III)
Chromatographic methods provide preliminary information on the AXOS sizes and
monosaccharide composition, and thus are part of structural elucidation. Enzyme-aided
methods were also used in structural elucidation of AXOS. Using partial enzymatic
hydrolysis, more information on the structures of the purified AXOS was obtained. The �-
Araf side groups were removed, using �-arabinofuranosidases, which revealed the length
of the xylose main chain in the following TLC or HPAEC-PAD analysis (III). The AXOS
were totally hydrolysed into monosaccharides, using a combination of �-xylosidase and �-
arabinofuranosidase, and the Ara/Xyl ratio was determined by GC or HPAEC-PAD
analysis (I, II). MS and NMR spectroscopy were used for complete structural analysis of
the AXOS. The analytical steps taken to fully characterize the structures of AXOS are
presented in Figure 5.

56
MS
First screening of AXOSwith TLC
GC analysis
Second screening of AXOSwith HPAEC-PAD
Degradation of AXOS into monosaccharides,
using enzymatic hydrolysisor acid methanolysis NMR
Choosing interestingsamples
Choosing the bestsamples for structural
analysis
Mw of AXOSAra/Xyl ratio Structure
Figure 5. Procedure for elucidation of AXOS structures.
4.2.7.1 Mass spectrometry
MS was used in structural analysis of the purified AXOS (I-III). Matrix-assisted laser
desorption/ionization time-of-flight mass spectrometry (MALDI-TOF-MS) analyses (I, II)
were performed, using an Ultraflex instrument (Bruker Daltonics, Wormer, The
Netherlands) equipped with a nitrogen laser at 337 nm. The mass spectrometer was
selected for positive ions, which were accelerated to a kinetic energy of 12 000 V after a
delayed extraction time of 200 ns. Hereafter, the ions were detected using the reflector
mode. The lowest laser power required to obtain good spectra was used, and at least 100
spectra were collected. The mass spectrometer was calibrated with a mixture of
maltodextrin standards (mass range 365-2309).
The matrix solution was prepared by dissolving 10 mg 2,5-dihydroxybenzoic acid in 1 ml
30% (v/v) acetonitrile-water solution. The samples (1 �l) and the matrix (1 �l) were
pipetted onto a MALDI-TOF plate (Bruker Daltonics) and the mixtures were allowed to
dry under a constant stream of warm air.

57
Electrospray ionization mass spectrometry (ESI-MS) analysis (III) was performed, using a
liquid chromatography/mass selective detector (LC/MSD-Trap-XCT Plus; Agilent
Technologies, Santa Clara, CA, USA) instrument in the positive ion mode. The purified
sample in 0.1% formic acid (HCOOH) and isopropyl alcohol (IPA), at a ratio of 1:1 (v/v),
was directly injected into the mass spectrophotometer, using the LC autosampler and the
instrument was operated without a column. The nebulizer gas was N2 and the capillary
voltage 3500 V at 350 oC. The spectra were recorded with a scanning range from 50 to
1000 m/z.
4.2.7.2 NMR spectroscopy
Prior to the NMR experiments, the samples were exchanged three or four times with
deuterium oxide (D2O) (99.8% D, Merck), filtered with 0.45-�m and/or 0.10-�m
Acrodisc® syringe filters with a GHP membrane (Pall Corporation) and finally dissolved
in 40 �l (I) or 0.5 ml (II, III) of D2O. The NMR spectra were acquired on a Varian Unity
500 spectrometer (Varian NMR Systems, Palo Alto, CA, USA) operating at 500 MHz for 1H. The measurements were performed using a gHX nano-NMR probe (I), or a 5-mm
triple-resonance pulse-field gradient (PFG) probe or 5-mm cryoprobe (I, II). The
measurements for the polysaccharide samples were performed at 50 ºC (III) and for the
oligosaccharides at 23 ºC or 25 oC (I-III). The chemical shifts were reported relative to an
internal acetone standard at 2.225 ppm for 1H (II, III) and 31.55 ppm for 13C (I-III). The 1H chemical shifts were referenced to an HDO signal at 4.78 ppm (I).
The one-dimensional (1D) 1H spectra were accumulated with spectral widths of 6000 or
8000 Hz and with acquisition times of 2-4 s. In recording the spectra, a modification of the
water-eliminated Fourier transform (WEFT) sequence was used. The two-dimensional
(2D) 1H-1H and 13C-1H correlation experiments were DQF-COSY (double-quantum
filtered correlation spectroscopy), HSQC (heteronuclear single-quantum correlation),
HMQC (heteronuclear multiple-quantum correlation) and HMBC (heteronuclear
multibond connectivity).

58
4.2.8 Microbiological and molecular biological methods (IV)
4.2.8.1 Fermentation experiments
The prebiotic properties of the AXOS prepared in earlier studies (I, II) were tested (IV).
Fermentation experiments were performed, using three different pure bifidobacterial
strains or mixed faecal microbiota in in vitro incubations with various carbohydrates as
substrates. The carbohydrates tested were: L-arabinose, D-xylose, XOS, FOS (reference
substrate), the RAX and WAX hydrolysates prepared by Shearzyme, and purified A2XX
and A2+3XX. RAX and WAX were also used as such as substrates for faecal
fermentations. The conditions of the fermentation experiments and the measured growth
indicators are summarized in Table 9.
Table 9. In vitro fermentation experiments of potential prebiotic AXOS by pure bifidobacteria and mixed faecal microbiota. Experiment Pure culture fermentations Faecal microbiota fermentations Bacterial strains Bifidobacterium adolescentis
B. breve B. longum
Mixed strains from intestinal microbiota
Growth medium BA medium + 1 g/l yeast extract + vitamin solution
BA medium + 1 g/l yeast extract + vitamin solution
Starting pH 7 7 Experiment volume 3 ml 2 ml Reductant - Na2S Incubation 140 h (~ 6 d) 48 h 37 oC in the dark, no shaking 37 oC in the dark, no shaking Measured growth indicators
pH VFA Carbohydrate consumption OD Contamination (microscopy)
pH VFA Carbohydrate consumption H2 Pressure
4.2.8.2 DNA extraction, purification, PCR and t-RFLP
At the beginning of the incubation period and after 48 h, 1-ml samples were taken from
the faecal samples (experiment volume 2 ml) for DNA extraction. At the same time points,
4-ml samples were taken from the reference fermentations (experiment volume 10 ml;
control samples without added carbon source). The samples were centrifuged for 2 min at
10 000 G at 4 oC, after which the supernatant was disposed and the pellet suspended in 1
ml 0.01-M PBS buffer, and this treatment was repeated three times. After the third

59
centrifugation, the pellet was suspended in 1.5 ml ASL buffer (stool lysis buffer; Qiagen
GmbH, Hilden, Germany) and bead-beaten (0.1-mm zirconia/silica beads) for 3 min,
followed by centrifugation at 16 000 G at 4 oC for 3 min. The subsequent DNA
purification was carried out, following the Qiagen kit instructions for "DNA purification
from stool samples" (Qiagen, Valencia, CA, USA).
The bifidobacterial 16S ribosomal RNA (rRNA) genes were amplified by a polymerase
chain reaction (PCR) using a bifidobacteria-specific primer set. The total volume of the
PCR mixture was 50 �l and contained milli-Q-water, reaction buffer, Mg2+ (25 mM),
deoxyribonucleotide triphosphate (dNTP) (2 mM), forward primer (25 pmol/�l; TET-
Bifido 144f NF, MWG-Biotech AG, Ebersberg, Germany, 5´-
CCGGAATAGCTCCTGGAAAC-3´, based on the Pbi F1 primer described by Roy and
Sirois (2000), reverse primer (25 pmol/�l; rP2, MWG-Biotech-AG, 5´-
ACGGCTACCTTGTTACGACTT-3´ (Weisburg et al., 1991), bovine serum albumin
(BSA; 2%), polymerase and template. The PCR (iCycler Thermal Cycler; Bio-Rad) was
started with an initial denaturation step at 94 oC for 5 min, followed by 30 cycles
comprising denaturation at 94 oC for 30 s, annealing at 59 oC for 30 s, elongation at 72 oC
for 90 s and final extention at 72 oC for 5 min. Agarose gel (1%) was loaded with the PCR
products and gel electrophoresis was run for 30 min at 80 V (Bio-Rad/PowerPac 300
power supply) and the bands on the gel were visualized, using the Bio-Rad Gel Doc™
2000 documentation system (darkroom cabinet, UV transilluminator, camera and
software). The band containing the PCR product of interest was purified, using gel
extraction according to the Gel-Out procedure (A&A Biotechnology, Gdynia, Poland).
After the PCR, the 16S rRNA gene products were terminally fluorescent-labelled. This
PCR product (10 �l) was digested with 1 �l of the enzyme MboI (10 U/�l; Fermentas
International Inc., Burlington, Ontario, Canada) for 6 h at 37 oC, after which the enzyme
was inactivated at 65 oC for 20 min. The labelled fragments were analysed by
electrophoresis in an ABI PRISM 373 DNA sequencer in Gene-Scan mode and the
fragment sizes were determined, using DNA fragment standards (GS-500 ROX and GS-
1000 ROX; PE Biosystems, Foster City, CA, USA). Terminal restriction fragment length
polymorphism (t-RFLP) analyses were used together with a specific Bifidobacterium

60
primer set to demonstrate the population changes in the faecal samples due to addition of
the xylan-derived carbohydrates.
61
5. RESULTS
5.1 Carbohydrate composition of starting materials (I-IV)
The composition of the neutral monosaccharides of the AX used was determined (I-IV).
The highest Ara/Xyl ratio was found in WBAX (0.60) and the lowest in OSAX (0.09)
(Table 10). In all the studied AX extracted from brans (WBAX, RBAX and OBAX) and in
most AX from flours (RAX, WAX-hv and WAX-mv), the Ara/Xyl ratios were relatively
high, indicating highly substituted AX structures. In contrast, in OSAX, RiHAX and
WStAX a low degree of arabinose substitution was detected (Ara/Xyl ratio 0.09-0.19).
Table 10. Composition of neutral monosaccharides of AX extracted from different cereal materials.
AX source Ara
(wt%) Xyl
(wt%) Glc
(wt%) Ara/Xyl Method Study
RAX 27 63 10 0.42 Acid methanolysis I 30 65 5 0.46 H2SO4 hydrolysis I, IV WAX-hv 30 66 5 0.45 Acid methanolysis * WAX-mv 32 68 - 0.47 Acid methanolysis II 36 64 - 0.56 Enzymatic hydrolysis§ IV WAX-lv 21 76 3 0.28 Acid methanolysis * WBAX 35 57 8 0.60 Acid methanolysis III RBAX 26 62 12 0.43 Acid methanolysis III OBAX 26 47 27 0.53 Acid methanolysis III RiHAX 16 74 10 0.19 Acid methanolysis III BHAX 21 73 6 0.28 Acid methanolysis† III OSAX 7 82 11 0.09 Acid methanolysis III 10 87 3 0.11 H2SO4 hydrolysis * CCAX 24 73 3 0.33 H2SO4 hydrolysis‡ III WStAX 10 85 5 0.12 Acid methanolysis III * This thesis (not published), § analysed by Virkki et al. (2008), † analysed by Pitkänen et al. (2008), ‡ analysed by Ramírez and Saake (unpublished results).
5.2. Structures of enzymatically prepared AXOS (I-III)
Six different AXOS were prepared and isolated (I-III). Similar methods were used with
some minor modifications in manufacturing various AXOS. The results of structural
elucidation of AXOS are shown in Table 11. The A3X, A2+3X, A2+3XX, A2XX and D2,3X

62
appeared quite pure, but A2X was not purified completely and it occurred together with
A2XX. The mixture of A2X and A2XX was further treated with T. reesei GH5 xylanase
which liberates one xylose residue from the reducing end of A2XX, forming A2X. A2X
was not further separated and thus analysis to determine the Ara/Xyl ratio of A2X was not
done.
Table 11. Structural elucidation of purified AXOS prepared from cereal xylans.
Study AXOS tR (min)�
Mw (Na adduct) Ara/Xyl AX material Enzymes
I A3X 31.0 437 0.49 RAX Shearzyme II
A2+3X A2+3XX
A2X A2XX
33.4 34.4
28.4 30.1
569 701
437† 569
0.93 0.73
n.a. 0.36
WAX-mv
WAX-mv
AXH-m + Shearzyme AXH-m + Shearzyme + AXH-d3
III D2,3X 30.2 569 0.24 OSAX Shearzyme � = tR from HPAEC-PAD analysis vary slightly between studies I-III but those reported here are adopted from Figure 9. † = analysed from a mixture of two AXOS n.a. = not analysed
The AXOS were analysed by HPAEC-PAD, from which they elute with different
retention times (tR). The molecular weights (Mw) were obtained by MS analysis and they
are presented as Na-adducts (437 = sodium adduct of pentotriose, 569 = sodium adduct of
pentotetraose, 701 = sodium adduct of pentopentaose). The Ara/Xyl ratio was determined,
using either total enzymatic hydrolysis or acid methanolysis to degrade AXOS into
monosaccharides that were quantified, using GC methods. The AXOS studied were
composed of one or two arabinose units and two or three xylose residues. Finally, the
detailed structures of AXOS were studied, using NMR spectroscopy. Detailed NMR
results (chemical shifts and spectra) can be found in studies I-III. The structures of AXOS
are shown in Figure 6. In addition to the six AXOS that were fully analysed, the structures
of XA3XX and XA3X were elucidated, using MS and enzyme-aided methods, but their
structures were not confirmed with NMR analysis.

63
OO O
O O
O O
OH
OHHOH2C
OO
OH
HO CH2OH
HOHO
HO
OH
OH
OHOO
O
O O
OH
OHHOH2C
OO
OH
HO CH2OH
HOHO
OH
OH
OO O
O O
O O
OH
OHHOH2C
HOHO
HO
OH
OH
OHHOOO
O
O O
OH
OHHOH2C
HOHO
OH
OHHO
OO
O
OO
O
HO CH2OH
HOHO
OH
OHOH
OHO
OHOH
OO
O
OO
OH
HO CH2OH
HOHO
OH
OHOH
A3X
A2+3X
A2X
�-L-Araf-(1�3)-�-D-Xylp-(1�4)-D-Xyl
�-L-Araf-(1�2)-[�-L-Araf-(1�3)]-�-D-Xylp-(1�4)-D-Xyl
�-L-Araf-(1�2)-�-D-Xylp-(1�4)-D-Xyl
D2,3X
�-D-Xylp-(1�2)-�-L-Araf-(1�3)-�-D-Xylp-(1�4)-D-Xyl
�-L-Araf-(1�2)-[�-L-Araf-(1�3)]-�-D-Xylp-(1�4)-�-D-Xylp-(1�4)-D-Xyl
�-L-Araf-(1�2)-�-D-Xylp-(1�4)-�-D-Xylp-(1�4)-D-Xyl
A2+3XX
A2XX
a
b
c
d
e
f
Figure 6. Chemical structures of the isolated AXOS.
Purification of the AXOS was performed on a preparative scale where 1.5 g (I, II) or 5 g
(III) of starting material were hydrolysed and fractionated. The purest and most
concentrated fractions, which were used for the structural analysis, were quantified. The
concentrations of the best AXOS samples were: A3X 132 �g/ml (0.32 �mol/ml), A2+3X
215 �g/ml (0.39 �mol/ml), A2+3XX 461 �g/ml (0.68 �mol/ml), A2XX 433 �g/ml (0.79
�mol/ml) and D2,3X 443 �g/ml (0.81 �mol/ml). The content of A2X was not determined.
The dilutions for standard curve samples were made from these fractions. The standards
were used for identification and quantification of other samples containing the
corresponding AXOS.
A total of 1.5 g of the RAX sample with 87 wt% purity (contained 1.3 g of pure AX), was
hydrolysed (I). The quantified products accounted for 28% of the AX, and the yield of
A3X was 12% of the quantified products (3% of AX). Similarly, 1.5 g of the WAX sample
with 84 wt% purity (contained 1.26 g of pure AX) was hydrolysed, and the hydrolysis

64
products A2+3X and A2+3XX comprised in all 17% of the AX (II). In an other hydrolysis
from the same starting material, 11.7% A2XX of AX was obtained (II). The AXOS yields
were calculated from the isolated fractions (I, II). The hydrolysis products (III)
corresponded to 23% of the OSAX and were quantified from the hydrolysate instead of
the isolated fractions, as in I and II. The yield of AXOS was 12% from the quantified
products and 4% were D2,3X.
Structural elucidation of monosubstituted AXOS from wheat flour AX is presented in
Figure 7 as an example. WAX-mv was hydrolysed stepwise with enzymes into mono-, di-
and oligosaccharides (Figure 7a). After the hydrolysis and chromatographic (GPC)
separation of AXOS, TLC was used for the first screening of AXOS (Figure 7b). The most
promising samples (fractions 141-150) were chosen for HPAEC-PAD analysis, in which
the most representative sample (fraction 143) for the structural elucidations was selected
(Figure 7c).
The Mw of the monosubstituted AXOS as Na adduct was 569 (Figure 7d), which indicates
the structure consisted of four pentose units. The AXOS was then degraded totally into
monosaccharides and the HPAEC-PAD chromatogram in Figure 7e shows that no
oligosaccharides were left undegraded. The Ara/Xyl ratio was determined with GC to be
0.36, which indicates that the AXOS is composed of one arabinose residue and three
xylose units. The amount of AXOS was then calculated, based on the monosaccharide
concentration. Using the enzyme AXH-m, which removes the �-L-Araf units from
monosubstituted �-D-Xylp units, we confirmed that the length of the xylose chain consists
of three residues (Figure 7f). With NMR spectroscopy (Figure 7g) the AXOS structure
was specified to have a single (1�2)-linked �-L-Araf residue (A2) at the nonreducing �-
D-Xylp unit (X3) of xylotriose (Figure 7h). Thus, the complete structure was �-L-Araf-
(1�2)-�-D-Xylp-(1�4)-�-D-Xylp-(1�4)-D-Xyl (A2XX).
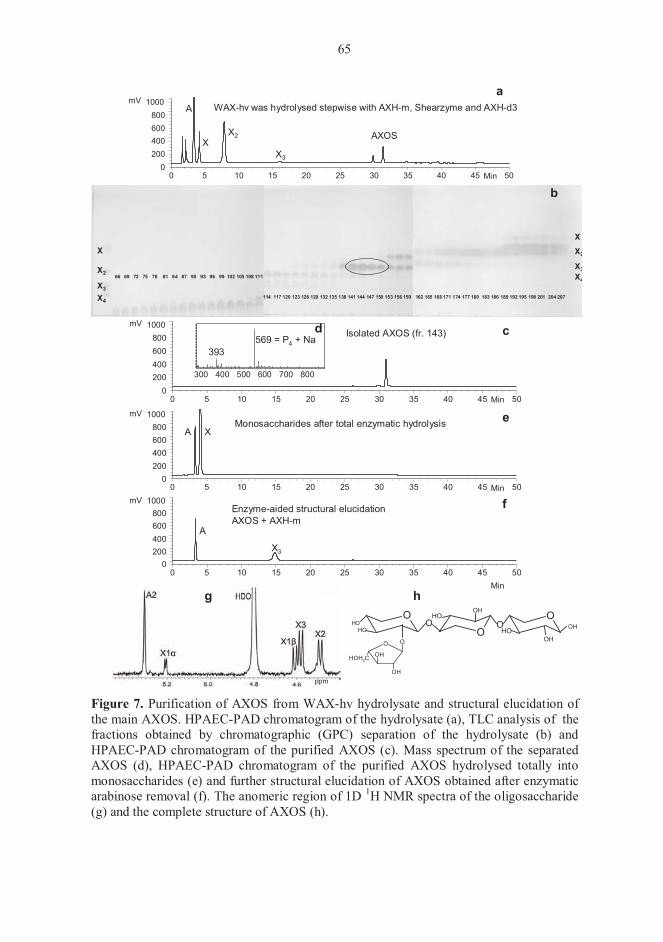
65
0200400600800
1000
0 5 10 15 20 25 30 35 40 45 50Min
WAX-hv was hydrolysed stepwise with AXH-m, Shearzyme and AXH-d3
X2X
A
X3
AXOS
a
1
X
X2
X3
X4
66 69 72 75 78 81 84 87 90 93 96 99 102 105 108 111
114 117 120 123 126 129 132 135 138 141 144 147 150 153 156 159 162 165 168 171 174 177 180 183 186 189 192 195 198 201 204 207
X
X2
X3X4
b
mV
0200400600800
1000
0 5 10 15 20 25 30 35 40 45 50
0200400600800
1000
0 5 10 15 20 25 30 35 40 45 50
Isolated AXOS (fr. 143)
300 400 500 600 700 800
393569 = P4 + Na
Min
0200400600800
1000
0 5 10 15 20 25 30 35 40 45 50Min
Monosaccharides after total enzymatic hydrolysisA X
c
e
f
d
Enzyme-aided structural elucidationAXOS + AXH-m
X3
A
mV
mV
mV
Min
OO O
O O
O O
OH
OHHOH2C
HOHO
HO
OH
OH
OHHO
g h
ppm Figure 7. Purification of AXOS from WAX-hv hydrolysate and structural elucidation of the main AXOS. HPAEC-PAD chromatogram of the hydrolysate (a), TLC analysis of the fractions obtained by chromatographic (GPC) separation of the hydrolysate (b) and HPAEC-PAD chromatogram of the purified AXOS (c). Mass spectrum of the separated AXOS (d), HPAEC-PAD chromatogram of the purified AXOS hydrolysed totally into monosaccharides (e) and further structural elucidation of AXOS obtained after enzymatic arabinose removal (f). The anomeric region of 1D 1H NMR spectra of the oligosaccharide (g) and the complete structure of AXOS (h).
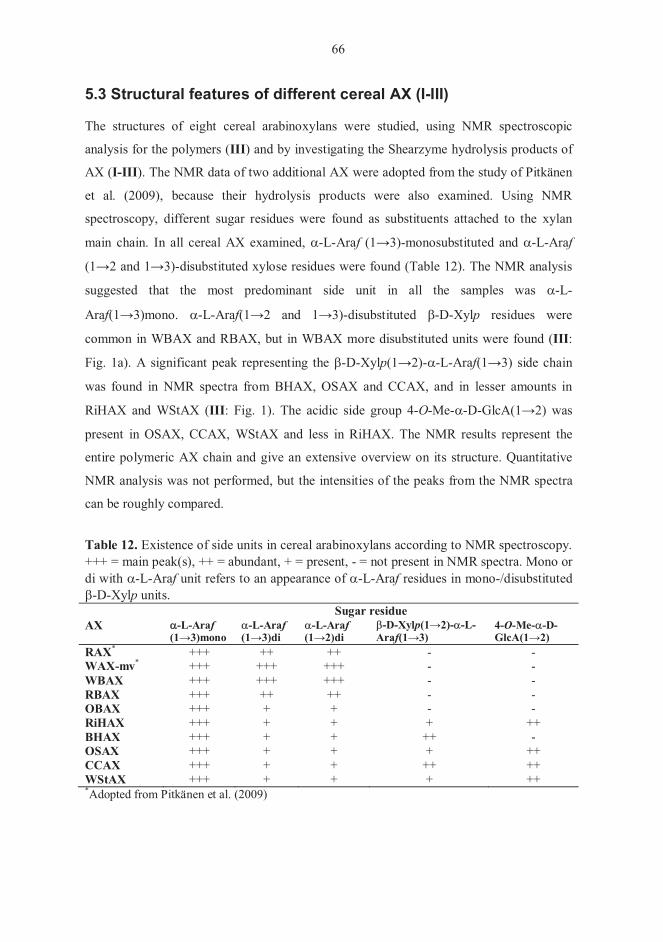
66
5.3 Structural features of different cereal AX (I-III)
The structures of eight cereal arabinoxylans were studied, using NMR spectroscopic
analysis for the polymers (III) and by investigating the Shearzyme hydrolysis products of
AX (I-III). The NMR data of two additional AX were adopted from the study of Pitkänen
et al. (2009), because their hydrolysis products were also examined. Using NMR
spectroscopy, different sugar residues were found as substituents attached to the xylan
main chain. In all cereal AX examined, �-L-Araf (1�3)-monosubstituted and �-L-Araf
(1�2 and 1�3)-disubstituted xylose residues were found (Table 12). The NMR analysis
suggested that the most predominant side unit in all the samples was �-L-
Araf(1�3)mono. �-L-Araf(1�2 and 1�3)-disubstituted �-D-Xylp residues were
common in WBAX and RBAX, but in WBAX more disubstituted units were found (III:
Fig. 1a). A significant peak representing the �-D-Xylp(1�2)-�-L-Araf(1�3) side chain
was found in NMR spectra from BHAX, OSAX and CCAX, and in lesser amounts in
RiHAX and WStAX (III: Fig. 1). The acidic side group 4-O-Me-�-D-GlcA(1�2) was
present in OSAX, CCAX, WStAX and less in RiHAX. The NMR results represent the
entire polymeric AX chain and give an extensive overview on its structure. Quantitative
NMR analysis was not performed, but the intensities of the peaks from the NMR spectra
can be roughly compared.
Table 12. Existence of side units in cereal arabinoxylans according to NMR spectroscopy. +++ = main peak(s), ++ = abundant, + = present, - = not present in NMR spectra. Mono or di with �-L-Araf unit refers to an appearance of �-L-Araf residues in mono-/disubstituted �-D-Xylp units.
AX Sugar residue
��-L-Araf (1�3)mono
�-L-Araf (1�3)di
�-L-Araf (1�2)di
�-D-Xylp(1�2)-�-L-Araf(1�3)
4-O-Me-�-D-GlcA(1�2)
RAX* +++ ++ ++ - - WAX-mv* +++ +++ +++ - - WBAX +++ +++ +++ - - RBAX +++ ++ ++ - - OBAX +++ + + - - RiHAX +++ + + + ++ BHAX +++ + + ++ - OSAX +++ + + + ++ CCAX +++ + + ++ ++ WStAX +++ + + + ++ *Adopted from Pitkänen et al. (2009)
67
In addition to the side units presented in Table 12, �-L-Araf(1�2)mono residue may be
found from BHAX, WBAX, and RBAX. The broad double signals originating presumably
from the side units �-L-Araf(1�2)mono and �-L-Araf(1�3)di were found from these
samples in NMR spectrometry (III, Fig. 1). However, based on NMR spectroscopy, the
presence of �-L-Araf(1�2)mono side units in the AX examined is still uncertain.
The same AX samples were also hydrolysed with Shearzyme (I-III) and the hydrolysis
products were identified and quantified. HPAEC-PAD chromatograms are presented in I:
Fig. 2, II: Fig. 2 and III: Fig. 4. In all cereal AX hydrolysates examined, monosubstituted
A3X was formed abundantly (Table 13). Most of all A3X is present in flour- and bran-
originated AX hydrolysates (RAX, WAX-mx, WBAX, RBAX and OBAX), and also as
the main AXOS in RiHAX, OSAX and WStAX hydrolysates. D2,3X is the most common
oligosaccharide in BHAX and CCAX and is also found in lesser amounts in OSAX,
RiHAX and WStAX. Furthermore, doubly substituted AXOS A2+3X and A2+3XX were
detected in all samples, except for RiHAX. In BHAX and CCAX, a small peak
representing A2+3XX was found in the chromatograms, but the amounts were too small for
quantification. AXOS with �-L-Araf(1�2) monosubstituted �-D-Xylp units (A2X or
A2XX) were not identified from the hydrolysates. The total quantified amount of the
hydrolysis products of the total weighed amount of AX was 10-40%. The highest amount
was quantified in WStAX and the lowest in WBAX.
Table 13. Distribution of monosaccharides, linear XOS and AXOS in hydrolysates obtained by incubating cereal AX with Shearzyme. Distribution (wt%) of carbohydrates was calculated from the quantified hydrolysis products in HPAEC-PAD analysis.
AX Hydrolysis products
Distribution of the quantified carbohydrates (wt%) A X X2 X3 X4 D2,3X A3X A2+3X A2+3XX
RAX 2 15 56 - - - 20 4 3 WAX-mv 2 12 47 - - - 14 9 16 WBAX 2 17 51 - - - 16 7 7 RBAX 3 18 49 - - - 22 5 3 OBAX 4 19 45 - - - 21 5 6 RiHAX 1 18 61 - - 5 15 - - BHAX - 12 66 1 - 14 4 3 n.q. OSAX - 10 68 9 1 4 6 1 1 CCAX 1 13 61 1 - 15 7 2 n.q. WStAX - 12 72 3 2 1 7 1 2 n.q. = not quantified

68
Similar features of AX structures were found when the NMR results (Table 12) and
information obtained from the hydrolysates (Table 13) were compared. No doubly
substituted AXOS were found in the RiHAX hydrolysate, although in NMR analysis they
were present. Otherwise the same structural elements occurred in the corresponding cereal
AX in both analyses.
5.4 Action of Shearzyme (I-III)
Shearzyme, with A. aculeatus GH10 endo-1,4-�-D-xylanase as the main enzyme,
produced short-chain AXOS with �-L-Araf(1�3)mono, �-L-Araf(1�2)mono, �-L-
Araf(1�2 and 1�3)di or �-D-Xylp(1�2)-�-L-Araf(1�3) substituents in the
nonreducing end of the xylose chain. The formation of A2X and A2XX was best shown
with WAX-lv hydrolysate (I; Fig. 1) but these AXOS were not identified until study II.
The majority of AXOS produced in the Shearzyme hydrolysis carry the nonreducing end
substituent and only small amounts of AXOS are substituted internally (I; Fig. 7). Despite
the side activities (�-xylosidase, �-arabinofuranosidase, �-glucanase, and �-glucosidase)
in Shearzyme, the hydrolysates by pure A. aculeatus endo-1,4-�-D-xylanase and
Shearzyme are similar (I; Fig. 7) and thus, for the production of AXOS, only the action of
endo-1,4-�-D-xylanase in Shearzyme is significant. For Shearzyme to act, at least one
unsubstituted �-D-Xylp unit between substituted �-D-Xylp units is needed, since the
shortest AXOS formed in the hydrolysis are A3X, A2X, and D2,3X. In addition to AXOS,
xylose and xylobiose were the major hydrolysis products in extensive hydrolysis.
5.5 Behaviour of AXOS in chromatography (I-III)
5.5.1 Separation of AXOS on TLC
TLC was used in the first screening of the fractions obtained from the separation of the
hydrolysed AX samples (I-III) with AEC and/or GPC. In GPC the elution order of the
carbohydrates is mainly based on their sizes (Figure 7b), but in TLC the elution rate is also
affected by the varying adsorbtion of the carbohydrates to the silica plate. Ascending of

69
eight AXOS was examined on TLC plate and the Rf for each AXOS was determined
(Table 14). In TLC analysis the Rf values of the AXOS increased in the order: XA3XX,
A2XX and A2+3XX, D2,3X and XA3X, A2X and A2+3X, A3X (Figure 8). Thus, of the
AXOS studied, A3X ascended the farthest (Rf 0.57) on the TLC plate and XA3XX the least
(Rf 0.41). However, some of the Rf values of the AXOS were so similar that the
distinction between AXOS based only on Rf values was difficult.
Table 14. Rf values obtained from TLC analysis. Carbohydrate Rf value X 0.60 X2 0.52 X3 0.45 X4 0.37 XA3XX 0.41 A2XX 0.45 A2+3XX 0.46 D2,3X 0.48 XA3X 0.48 A2X 0.51 A2+3X 0.52 A3X 0.57
X
X2
X3
X4
X
X2
X3
X4
XA3XX A2+3XX XA3X A2+3XA2XX D2,3X A2X A3X
Figure 8. TLC analysis of AXOS.
5.5.2 Elution order of AXOS and their responses in HPAEC-PAD
Purified AXOS (0.011 �mol of each) were mixed in a solution and the elution order of the
AXOS (I-III) and their responses in the HPAEC-PAD system used were studied. The
elution order in HPAEC-PAD was (Figure 9a): A2X, A2XX, D2,3X, A3X, A2+3X and
A2+3XX. Although A2XX and D2,3X could not be separated entirely from each other in the
mixture, a peak with split ends was seen at 30 min and the elution order was determined
from separate injections of these AXOS. Linear XOS (X-X4; 0.011 �mol of each) were
also mixed with six AXOS (Figure 9b). Furthermore, two more AXOS (XA3X and
XA3XX) were mixed with the solution of six AXOS and the elution order of these eight
AXOS was examined (Figure 9c). Elution order of the eight AXOS was: A2X, A2XX,
D2,3X, A3X and XA3X, XA3XX, A2+3X, A2+3XX. The tR of A3X and XA3X were exactly
the same, and their elution order in this HPAEC-PAD system could not be determined.
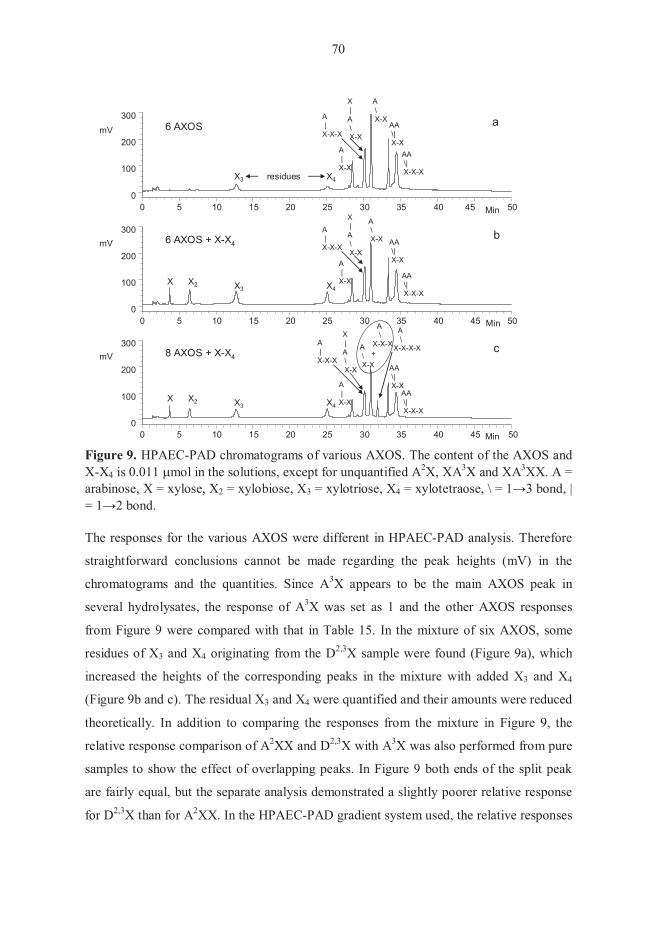
70
0
100
200
300
0 5 10 15 20 25 30 35 40 45 50
0
100
200
300
0 5 10 15 20 25 30 35 40 45 50
0
100
200
300
0 5 10 15 20 25 30 35 40 45 50
Min
Min
Min
X3
X3
X3X
X
X2
X2
X4
X4
X4
residues
A|
X-X
A|
X-X-X
X|
A\X-X
A\X-X
AA\|X-X
AA\|X-X-X
A|
X-X
A|
X-X
A|
X-X-X
A|
X-X-X
X|A\X-X
X|
A\X-X
A\X-X
A\X-X
AA\|X-X
AA\|X-X
AA\|X-X-X
AA\|X-X-X
A\
X-X-X-X
A\
X-X-X+
a
b
c
mV
mV
mV
6 AXOS
6 AXOS + X-X4
8 AXOS + X-X4
Figure 9. HPAEC-PAD chromatograms of various AXOS. The content of the AXOS and X-X4 is 0.011 �mol in the solutions, except for unquantified A2X, XA3X and XA3XX. A = arabinose, X = xylose, X2 = xylobiose, X3 = xylotriose, X4 = xylotetraose, \ = 1�3 bond, | = 1�2 bond.
The responses for the various AXOS were different in HPAEC-PAD analysis. Therefore
straightforward conclusions cannot be made regarding the peak heights (mV) in the
chromatograms and the quantities. Since A3X appears to be the main AXOS peak in
several hydrolysates, the response of A3X was set as 1 and the other AXOS responses
from Figure 9 were compared with that in Table 15. In the mixture of six AXOS, some
residues of X3 and X4 originating from the D2,3X sample were found (Figure 9a), which
increased the heights of the corresponding peaks in the mixture with added X3 and X4
(Figure 9b and c). The residual X3 and X4 were quantified and their amounts were reduced
theoretically. In addition to comparing the responses from the mixture in Figure 9, the
relative response comparison of A2XX and D2,3X with A3X was also performed from pure
samples to show the effect of overlapping peaks. In Figure 9 both ends of the split peak
are fairly equal, but the separate analysis demonstrated a slightly poorer relative response
for D2,3X than for A2XX. In the HPAEC-PAD gradient system used, the relative responses

71
for X-X4 appeared to be poorer than for AXOS, while variation in responses was also
observed amongst different AXOS.
Table 15. Responses of xylose, linear XOS and AXOS compared with the response of A3X. The content of the carbohydrates is similar in moles (0.011 �mol). Response = height (mV) of the peak in HPAEC-PAD analysis (Figure 9a and b).
Carbohydrate Relative response (carbohydrate/A3X)
X 0.2 X2 0.2 X3 0.1 X4 0.1 A2XX 0.5 (0.4*) D2,3X 0.5 (0.3*) A3X 1 A2+3X 0.7 A2+3XX 0.5 * Calculated from separate chromatograms in addition to the mixture where these AXOS overlap
5.6 Fermentation experiments (IV)
5.6.1 Carbohydrate composition of XOS and AX hydrolysates
The carbohydrate compositions of a commercial XOS mixture and RAX and WAX
hydrolysates, used as substrates in the fermentation experiments (IV), were analysed. The
commercial XOS mixture contained mainly xylobiose (26 wt%) and smaller amounts of
xylotriose and xylotetraose (Table 16, IV: Fig. 1a). Thus, in this mixture only 37% of the
amount weighed was detected as linear XOS. In addition to the XOS identified, the
commercial XOS product contained some longer unidentified oligosaccharides with tR
between 30 and 35 min. The RAX and WAX hydrolysates were made from commercial
AX extracted from rye and wheat flour (RAX and WAX-hv, respectively). The identified
and quantified carbohydrates in the RAX hydrolysate were 14 wt% and in the WAX
hydrolysate 20 wt% (Table 16, IV: Figure 1b and 1c). The two hydrolysates contained
similar amounts of xylose, xylobiose and A3X, but the WAX hydrolysate had more of the
disubstituted oligosaccharides A2+3X and A2+3XX.

72
Table 16. Carbohydrate composition of the mixture of commercial xylo-oligosaccharides (XOS), rye AX hydrolysate and wheat AX hydrolysate.
Carbohydrate X X2 X3 X4 A3X A2+3X A2+3XX Total wt% of AX
XOS - 26 8 3 - - - 37 RAX hydrolysate (aq.) 2 8 - - 4 n.q. n.q. 14 WAX hydrolysate (aq.) 2 9 - - 4 2 3 20 aq. = aqueous solution (not buffered), n.q. = not quantified
5.6.2. Growth of pure bifidobacterial cultures
The substrate preferences of three different bifidobacterial species (Bifidobacterium
adolescentis, B. breve and B. longum) were studied by incubating the bacteria with D-
xylose, L-arabinose, XOS, RAX and WAX hydrolysates and FOS (control substrate). The
fermentation of pure singly and doubly substituted AXOS was studied by cultivating a
mixture of the three preceding bifidobacteria with A2XX and A2+3XX. The utilization of
different substrates is presented in Table 17.
Table 17. Growth of three bifidobacterial species and their mixture. Growth of Substrate B. adolescentis B. breve B. longum ATCC 15703 ATCC 15700 ATCC 15707 60 h 140 h 60 h 140 h 60 h 140 h D-xylose + +++ - - - - L-arabinose - - - - +++ +++ XOS +++ +++ - - - - RAX hydrolysate + + - + + + WAX hydrolysate + ++ - - + + FOS +++ +++ ++ ++ +++ +++ Substrate Growth of the mixture of B. adolescentis, B. breve and B. longum 60 h 140 h A2XX +++ +++ A2+3XX - - FOS +++ +++ Scale: OD < 0.1 (- = no growth), 0.1-0.39 (+ = mild growth), 0.4-0.69 (++ = moderate growth), 0.7-1 (+++ = strong growth). Acetate content, pH and consumption of carbohydrates in the samples were also measured and considered in defining growth intensities. XOS = xylo-oligosaccharides, RAX hydrolysate = rye arabinoxylan hydrolysate, WAX hydrolysate = wheat arabinoxylan hydrolysate, FOS = fructo-oligosaccharides (control substrate).

73
Bifidobacterium adolescentis was able to grow on D-xylose, XOS and RAX and WAX
hydrolysates, whereas B. longum utilized L-arabinose and hydrolysates. Bifidobacterium
breve showed only mild growth on RAX hydrolysate after 140 h. With longer
fermentation time, some changes were also observed in the growth of B. adolescentis on
D-xylose and WAX hydrolysate, since the growth increased from mild at 60 h to strong or
moderate at 140 h. The growth of B. adolescentis and B. longum was strong on control
substrate FOS, while B. breve was able to grow moderately on FOS. A bifidobacteria
mixture utilized pure singly substituted AXOS almost completely, but pure AXOS with a
doubly substituted xylose residue was not fermented.
Even more detailed information on the substrate preferences of these three bifidobacteria
was obtained by analysing the substrates before and after the incubation, using HPAEC-
PAD chromatography. Quantification of substrates gave better insight into the fate of
mono- and oligosaccharides in the fermentations (Figure 10). Bifidobacterium
adolescentis fermented all AXOS (Figure 10b) but consumed mainly XOS, leaving L-
arabinose and some D-xylose unutilized. Bifidobacterium longum also fermented AXOS,
but utilized them with a strategy different from that of B. adolescentis, since it consumed
the L-arabinose released but not XOS (Figure 10c). Bifidobacterium breve utilized the
RAX hydrolysate poorly and no growth was observed with the WAX hydrolysate as a
substrate (Figure 10d).

74
0
50
100
150
0 5 10 15 20 25 30 35 40 45 50
X62�g/ml
X2316 �g/ml A3X
121 �g/ml
A2+3X72 �g/ml A2+3XX
91 �g/ml
WAX hydrolysatea
0
100
200
300
400
0 5 10 15 20 25 30 35 40 45 50
0
100
200
300
400
0 5 10 15 20 25 30 35 40 45 50
0
100
200
300
400
0 5 10 15 20 25 30 35 40 45 50
B. adolescentisX135 �g/ml
b
B. longum c
B. breve d
X21115 �g/ml
X2310 �g/ml
X3471 �g/ml
X84 �g/ml
Min
Min
Min
A3X84 �g/ml
A2+3X76 �g/ml A2+3XX
112 �g/ml
MinA
265 �g/ml
mV
mV
mV
mV
Figure 10. Carbohydrate analysis by HPAEC-PAD of WAX hydrolysate a) before fermentation and b) after 140-h fermentation by Bifidobacterium adolescentis ATCC 15703, c) B. longum ATCC 15700 and d) B. breve ATCC 15707.
5.6.3 Fermentations with faecal microbiota
In the fermentations with faecal microbiota, the measured factors providing information
on growth of the bifidobacteria were pH, pressure, and formation of VFA and H2. After
the incubation period, changes in pH were observed, indicating that the mixture of faecal
microbiota fermented the carbohydrates provided. In the samples with FOS, L-arabinose,
D-xylose, XOS, and RAX and WAX hydrolysates, the end pH was 5. With polymeric
RAX and WAX as carbon sources, the pH was somewhat higher (5.5) at the end of the
fermentation period. With the purified oligosaccharides A2XX and A2+3XX, the pH was 6
and 4, respectively. Pressure increased in all the samples (0.8-2.2 psi), and the least
change was detected in the samples grown on A2+3XX and the greatest with RAX
hydrolysate as a carbon source (Table 18). Hydrogen formation (0.2-3.8 v/v %) was
detected in the samples with all the other carbon sources except A2+3XX, in which no

75
hydrogen was found. Most of the hydrogen was formed in the samples grown on xylose
and FOS. With faecal samples the change in acetate concentration after the incubation
period, compared with the control sample after 48 h, was typically about 15 mM, but in
the samples with A2XX and A2+3XX, the change in acetate concentration was 22 and 31
mM, respectively. In most of the samples, some changes in propionate and butyrate
concentrations were also detected (0.3-6.9 and -1.3-5.8 mM, respectively).
Table 18. Formation of H2, pressure and VFA (acetate, propionate and butyrate) during the incubation period (48 h) in faecal microbiota cultivations.
Carbohydrate End pH H2 Pressure Acetate Propionate Butyrate (v/v %) (� psi) (� mM) (� mM) (� mM)
D-xylose 5 3.8 1.8 15.3 6.9 0.4 L-arabinose 5 2.0 1.9 15.4 2.6 2.3 XOS 5 2.1 2.0 16.7 4.3 4.0 RAX hydrolysate 5 1.4 2.2 13.9 2.2 4.9 WAX hydrolysate 5 2.0 2.0 13.4 1.9 2.9 RAX 5.5 0.2 1.1 14.4 2.3 3.5 WAX 5.5 1.0 1.6 16.0 2.9 5.8 A2XX 6 2.1 1.6 22.1 4.6 5.4 A2+3XX 4 0 0.8 31.2 0.3 -1.3 FOS 5 4.0 2.0 13.5 1.8 3.3 Pressure (� psi) and VFA (� mM): Pressure and VFA formation are compared with control sample at 48 h.
The utilization of various carbohydrates was analysed with the HPAEC-PAD method, and
the changes in bifidobacteria distribution were studied, using t-RFLP analysis. The L-
arabinose, D-xylose, XOS, FOS, WAX and RAX hydrolysates, and polymeric WAX and
RAX were utilized almost completely and only traces of these compounds (with polymers,
traces of X-X4) were seen in the HPAEC-PAD chromatograms. Purified monosubstituted
AXOS (A2XX) was also completely fermented and only traces of xylose were found in the
chromatograms (Table 19). In the case of doubly substituted purified AXOS (A2+3XX), the
oligosaccharide was not consumed completely (Table 19, IV: Figure 4) and small amounts
of arabinose, xylose, xylobiose, xylotriose and AXOS (A2+3XX and A2XX) were detected
after a 48-h fermentation. The results clearly show that in these samples the A2XX was
formed during the incubation from A2+3XX by the bacteria in the faecal microbiota.
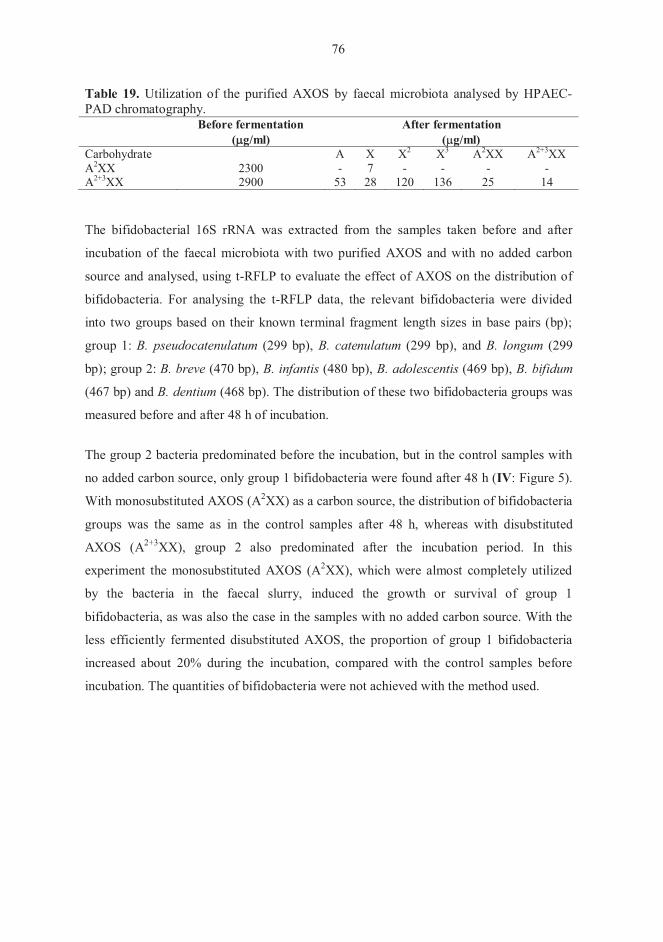
76
Table 19. Utilization of the purified AXOS by faecal microbiota analysed by HPAEC-PAD chromatography. Before fermentation
(��g/ml) After fermentation
(�g/ml) Carbohydrate A X X2 X3 A2XX A2+3XX A2XX 2300 - 7 - - - - A2+3XX 2900 53 28 120 136 25 14
The bifidobacterial 16S rRNA was extracted from the samples taken before and after
incubation of the faecal microbiota with two purified AXOS and with no added carbon
source and analysed, using t-RFLP to evaluate the effect of AXOS on the distribution of
bifidobacteria. For analysing the t-RFLP data, the relevant bifidobacteria were divided
into two groups based on their known terminal fragment length sizes in base pairs (bp);
group 1: B. pseudocatenulatum (299 bp), B. catenulatum (299 bp), and B. longum (299
bp); group 2: B. breve (470 bp), B. infantis (480 bp), B. adolescentis (469 bp), B. bifidum
(467 bp) and B. dentium (468 bp). The distribution of these two bifidobacteria groups was
measured before and after 48 h of incubation.
The group 2 bacteria predominated before the incubation, but in the control samples with
no added carbon source, only group 1 bifidobacteria were found after 48 h (IV: Figure 5).
With monosubstituted AXOS (A2XX) as a carbon source, the distribution of bifidobacteria
groups was the same as in the control samples after 48 h, whereas with disubstituted
AXOS (A2+3XX), group 2 also predominated after the incubation period. In this
experiment the monosubstituted AXOS (A2XX), which were almost completely utilized
by the bacteria in the faecal slurry, induced the growth or survival of group 1
bifidobacteria, as was also the case in the samples with no added carbon source. With the
less efficiently fermented disubstituted AXOS, the proportion of group 1 bifidobacteria
increased about 20% during the incubation, compared with the control samples before
incubation. The quantities of bifidobacteria were not achieved with the method used.

77
6. DISCUSSION
6.1 Carbohydrate composition of starting materials
The carbohydrate compositon of 10 arabinoxylans extracted from different cereal plant
parts was analysed, using various methods. In the beginning of the study, H2SO4
hydrolysis was used, because in the preliminary tests higher monosaccharide yield was
obtained with H2SO4 hydrolysis than with acid methanolysis in the degradation of RAX
(I). Later (II, III), acid methanolysis was chosen as the routine method for most of the
samples since the monosaccharide yields were high and the repeatability of the results was
better than with H2SO4 hydrolysis. WAX-mv was hydrolysed, using an enzyme mixture
designed especially for wheat flour AX (Virkki et al., 2008) (IV). Enzymatic hydrolysis is
a milder choice than acid hydrolysis or acid methanolysis for depolymerization of AX
since no unwanted destruction of the liberated monosaccharides occurs. Thus, enzymatic
hydrolysis is a good choice for selective depolymerization of AX, into monosaccharides,
but the challenge is to make the mixtures by including all enzymes necessary for complete
AX degradation. Acid methanolysis is currently more easily adapted for degradation of
different AX and other possible polysaccharides present in the sample.
The contents of arabinose, xylose and glucose were determined and the Ara/Xyl ratio was
calculated for all AX. The Ara/Xyl ratio represents the number of �-D-Xylp units
substituted with �-L-Araf residues; the higher the Ara/Xyl ratio found, the higher the
substitution degree of AX is. However, the DS does not specify the distribution of �-L-
Araf units (quantity of mono- and disubstituted �-D-Xylp units) in the xylose main chain.
The Ara/Xyl ratio affects the behaviour and characteristics of AX, such as water solubility
and viscosity of AX solutions. Highly substituted AX are challenging for endo-1,4-�-D-
xylanases to degrade and high DS reduces the yield of AXOS obtained in the hydrolysis.
The monosaccharide composition of commercial WAX-mv indicates a relatively highly
substituted AX structure, although slight variation was observed between the results
obtained with acid methanolysis and enzymatic hydrolysis (Ara/Xyl ratios 0.47 and 0.56,
respectively). The results are in accordance with the Ara/Xyl ratio reported by the
manufacturer (Megazyme; 0.61). In the previous literature the Ara/Xyl ratios reported for

78
wheat flour AX ranged between 0.52 and 0.80 (MacArthur and D´Appolonia, 1980;
Gruppen et al., 1991). The Ara/Xyl ratio for RAX (acid methanolysis; 0.42 and H2SO4
hydrolysis; 0.46) was lower than the manufacturer (Megazyme) states (0.64) or reported
by Cyran et al. (2003 and 2004) (0.67-0.75). The Ara/Xyl ratio of OSAX (0.09)
corresponds well with the Ara/Xyl ratio reported by the manufacturer (Sigma-Aldrich
Chemie; 0.08) and Gruppen et al. (1991) (0.12).
The highest Ara/Xyl ratio of all AX examined was found in WBAX (0.60), but other
studies have reported even higher Ara/Xyl ratios for alkali-extracted wheat bran AX (0.73
and 0.81) (Gruppen et al., 1991; Nandini and Salimath, 2001). From rye bran both highly
branched AX (Ara/Xyl ratio 0.87) and AX with low DS (Ara/Xyl ratio 0.18) were alkali-
extracted by Hromádková et al. (1987) and the result obtained in the present study (0.43)
were found in the middle of that range. Higher Ara/Xyl ratios were also reported for the
alkali-extracted OBAX (1.07) (MacArthur and D´Appolonia, 1980) than were detected in
this study (0.53). Moderate arabinose substitution was found in CCAX (Ara/Xyl ratio of
0.33), which was significantly higher than that reported earlier for alkali-extracted corn
cob AX (0.06-0.11) (Kabel et al., 2002a; Ramirez et al., 2008). The lowest Ara/Xyl ratios
were detected in OSAX and WStAX (0.09 and 0.12, respectively), which correspond well
with previous results observed for oat spelt AX and wheat straw AX (0.13 and 0.11,
respectively) (Puls, 1993; Sun et al., 2002).
The natural variations in Ara/Xyl ratios are wide (Izydorczyk and Biliaderis, 1995), which
partially explains the differences in Ara/Xyl ratios detected in various studies.
Furthermore, various processes are used in the separation of cereal grain tissues, resulting
in slightly different plant parts (remnants from other than the desired layers), which may
result in differences in the extracted AX structures. The differences can also be due to
different cereal varieties, as well as diverse extraction and analytical methods.
6.2 Structural features of different cereal AX
The structures of eight alkali-extracted AX from different cereal by-products and two
commercial AX were studied, using NMR analysis for polymeric AX and by hydrolysing
the AX with enzymes and studying the AXOS produced with HPAEC-PAD

79
chromatography. NMR analysis of the polymeric samples provides information on the
entire molecule and, thus, is an important part of the structural elucidation of AX.
However, there are some restrictions in NMR techniques; for quantitative results, total
solubility is needed, good resolution of the signals is required, the sample content must be
sufficient and the NMR-run conditions must be optimal. In the present study, the sample
concentrations were adequate, but not all the samples were totally soluble in D2O and the
resolution of the peaks was not complete, since some of the peaks overlapped.
Overlapping of the signals is typical for the complex structures of AX, and originates from
the �-L-Araf units attached to two consecutively substituted xylose residues (Hoffmann et
al., 1992). Despite this, valuable structural information on the water-soluble part of the
alkali-extracted AX was obtained, even though the analysis conditions were not
satisfactory for quantitative analysis.
The hydrolysis products of the AX were identified and quantified. However, the AXOS
obtained and analysed were only the short oligosaccharides from the hydrolysed part of
the AX molecule; thus the degradation of the AX may not always be complete. Some of
the AX are highly substituted, or may alternate between highly branched and less
branched regions (Ewald and Perlin, 1959; Gruppen et al., 1993), of which highly
substituted areas restrict the action of the enzymes. The quantified hydrolysis products
from Shearzyme hydrolysis accounted for 28 wt% of the AX (I), whereas in study II over
40 wt% was quantified. The higher yield in study II was achieved at least partially due to
the selective removal of some of the �-L-Araf substituents from the AX by the AXH-m
treatment, revealing more sites on the xylose backbone accessible for the endo-1,4-�-D-
xylanase. In study III, the highest total quantified amount of hydrolysis products was 40%
of the total amount of AX weighed. This was achieved on WStAX with low degree of
substitution. The yield of quantified hydrolysis products was also surprisingly low in the
hydrolysates of AX with low DS. Even though the shortcoming in total solubility of AX
did not greatly affect successful enzymatic hydrolysis, complete solubility may be more
important than previously considered. In addition the hydrolysis products identified, some
minor unidentified longer AXOS are also often found. By combining the results from AX
hydrolysis with NMR analysis, better insight into the structures of AX is obtained.

80
The intensities of the NMR signals were compared within the samples and all the AX
studied possessed �-L-Araf(1�3) monosubstituted side chains. Reviewing the hydrolysis
products (Table 13), the oligosaccharide A3X commonly occurs and clearly is the most
predominating AXOS in RAX, WBAX, RBAX, OBAX, RiHAX and WStAX
hydrolysates. In NMR analysis, similarities in substitution profiles among the bran-
originated AX (Table 12) were observed, but differences in the amounts of the side chains
were found. NMR spectroscopy showed that, WAX-mv (Pitkänen et al., 2009) and
WBAX contained one third of the �-L-Araf (1�3) singly substituted �-D-Xylp residues
and two thirds were �-L-Araf (1�2) and (1�3) doubly substituted �-D-Xylp units (III;
Figure 1), whereas RAX (Pitkänen et al., 2009) and RBAX contained two thirds of the �-
L-Araf (1�3) monosubstituted �-D-Xylp residues and one third of the doubly substituted
units. These results correspond with previous reports on commercial wheat and rye flour
AX (Sørensen et al., 2006; Höije et al., 2008). Although wheat and rye flour and bran AX
have similar Ara/Xyl ratios, major differences in the distribution of the substituents were
found. Doubly substituted �-D-Xylp units were more infrequent in OBAX than in rye and
wheat AX based, on NMR sprectra (III; Figure 1). All the AX studied contained �-L-Araf
disubstituted �-D-Xylp units according to NMR analysis, but only in the hydrolysis
products of RiHAX were disubstituted AXOS not found.
In WAX-mv hydrolysate, 14 wt% of the quantified products were monosubstituted A3X,
while 25 wt% accounted for the doubly substituted oligosaccharides A2+3X and A2+3XX,
but in WBAX the same amounts of mono- and disubstituted AXOS (16 wt% and 14 wt%,
respectively) were detected. Approximately three quarters of the AXOS in RAX and
RBAX were monosubstituted and one quarter disubstituted, whereas two thirds of the
quantified AXOS were mono- and one third disubstituted in OBAX hydrolysate.
Straightforward comparison between NMR spectrometry results and the AXOS quantified
from the hydrolysates could not be made, because not all the hydrolysis products were
identified and AX was not hydrolysed totally. Based on NMR spectroscopy, OBAX
(Ara/Xyl 0.53) contains fever doubly substituted units than RBAX (Ara/Xyl 0.43), but the
proportion of disubstituted AXOS in the hydrolysates is higher in OBAX. This may
indicate more even distribution of �-L-Araf substituents in the xylose chain of OBAX.
The yield of disubstituted AXOS remained surprisingly low in RBAX hydrolysate,

81
probably because large amount of doubly substituted xylose units are situated in those
areas with high DS and with no sites on the xylan backbone accessible to the enzyme.
The existence of a fairly uncommon �-D-Xylp(1�2)-�-L-Araf(1�3) side chain was
previously reported in corn cob AX and barley husk AX (Aspinall, 1959; Ebringerová et
al., 1992; Höije et al., 2006). In addition to CCAX and BHAX, �-D-Xylp(1�2)-�-L-
Araf(1�3) side chain was here also discovered in RiHAX, OSAX and traces in WStAX,
using NMR spectroscopy. These findings are supported by studies of AX hydrolysis
products, since the oligosaccharide D2,3X was identified from the very same cereal-based
materials by HPAEC-PAD as in NMR analysis. Surprisingly, in BHAX and CCAX
hydrolysates, D2,3X is the most predominating AXOS.
In addition to the side chains identified, indications for the presence of �-L-Araf (1�2)
monosubstituted �-D-Xylp residues in BHAX were found by NMR spectroscopy (III:
Figure 1, signal at 5.30 ppm in BHAX). The peak presumably presenting �-L-Araf
(1�2)mono overlapped with the signals originating from �-L-Araf(1�3)di and/or 4-O-
Me-�-D-GlcpA(1�2), and accurate analyses were not possible. However, in BHAX the
sizes of the signals, indicating the presense of �-L-Araf(1�3)di and �-L-Araf(1�2)di
side chains, were not equal as they should have been if originating only from �-L-Araf
residues involved in double substitution. Differences in peak size strongly indicate the
presence of �-L-Araf (1�2)mono side chain. However, in the hydrolysed samples
analysed with HPAEC-PAD, not enough AXOS with �-L-Araf(1�2) monosubstituted �-
D-Xylp units (A2X or A2XX) were formed in any of the samples for definite
identification. A2XX overlapped with D2,3X, which made the identification even more
difficult. No evidence for the oligosaccharides with 4-O-Me-�-D-GlcpA residues in
BHAX was found, but in RiHAX, OSAX, CCAX and WStAX the oligosaccharide
U4m2XX was identified. Thus, 4-O-Me-�-D-GlcpA side units were found in the same AX,
using HPAEC-PAD analysis and NMR spectroscopy.
The occurrence of side units, attached to the xylose residues of the AX main chain found
in this study, is in line with the appearance of AX substituents in the corresponding cereal
plant parts reported in the literature. However, the existence of the more complex �-D-

82
Xylp(1�2)-�-L-Araf(1�3) side chain in RiHAX, OSAX and WStAX, in addition to the
previously known sources of BHAX and CCAX, was reported here for the first time.
Straws, spelts, husks and corncobs are agricultural by-products that are not consumed as
such, and hence they may be valuable raw materials for AXOS. In the hydrolysis of AX,
extracted from these by-products, large amounts of complex, possibly slowly fermenting
AXOS with the side group �-D-Xylp(1�2)-�-L-Araf(1�3) (D2,3X) are produced. On the
other hand, high amounts of AXOS, prepared from flour and bran AX, are doubly
substituted, which is also interesting due to their slow fermentability. However, since
flours are utilized as food ingredients, and brans as well can be exploited in human
nutrition, they are not the most reasonable source for extracting AX for AXOS production.
Thus, the AX present in flours and brans, could be hydrolysed into AXOS in situ without
extracting AX first.
6.3 Production of AXOS on a preparative scale
Six different AXOS (A3X, A2+3X, A2+3XX, A2X, A2XX, D2,3X) were prepared
enzymatically from RAX, WAX-mv and OSAX on a preparative scale (starting from 1.5
to 5.0 g of AX). The AXOS were purified and their structures fully analysed (I-III).
Shearzyme, with Aspergillus aculeatus GH10 endo-1,4-�-D-xylanase as the main enzyme,
possesses high activity towards the xylosidic linkage before an �-L-Araf-substituted �-D-
Xylp residue, thus forming AXOS with the substituents attached to the nonreducing end of
the short linear xylose chain. The oligosaccharides A2X and D2,3X are novel and the
detailed structural H1 and/or C13 NMR spectroscopy of A2+3X and A3X was presented for
the first time in this study.
Doubly substituted AXOS were produced for the first time in good yields (A2+3XX 11.8
wt% of AX (0.22 mmol) and A2+3X 5.6 wt% of AX (0.13 mmol)), and the corresponding
singly �-L-Araf-(1�2) substituted AXOS could also be prepared in similar yields by
treating the doubly substituted AXOS further with AXH-d3. Of the AX hydrolysed, 11.7
wt% (0.22 mmol) was A2XX. The amount of A2X was not quantified, due to lack of pure
sample. However, if A2X is similarly transformed from A2+3X, its amount can be
estimated to be 0.13 mmol. The yield of A3X (3% of AX, 0.11 mmol) was relatively low,

83
because RAX (with Ara/Xyl 0.46) was hydrolysed with endo-1,4-�-D-xylanase without
pretreatments, and presumably highly substituted areas were left unhydrolysed, since the
densely branched substrate does not fit in the active site of the enzyme. The other AXOS
with evidently higher yields (A2X, A2XX, A2+3X, and A2+3XX) were prepared by treating
AX first with �-arabinofuranosidase (AXH-m) to decrease the DS before endo-1,4-�-D-
xylanase hydrolysis. Purified D2,3X was only quantified after fractionation from the best
sample, which contained 443 �g/ml (3.9 �mol in one 5-ml fraction). Most of the previous
studies reporting purified AXOS have been carried out on an analytical scale. Wheat flour
AX (80 mg) was hydrolysed by A. awamori endo-1,4-�-D-xylanase I in the study of
Kormelink et al. (1993b), and 50-72 wt% of the hydrolysis products (monosaccharides,
XOS, and AXOS) were quantified. From barley malt hydrolysate, a maximum of 26 nmol
of the main purified AXOS (A2XX, A2+3XX, and XA2+3XX) were quantified (Broberg et
al., 2000). The study of Ordaz-Ortiz et al. (2004) was performed on a preparative scale
and from wheat flour AX (2.5 g) from which 8 wt% of AXOS were produced with the two
most predominating oligosaccharides, XA3XX and XA2+3XX, representing 3 wt% (0.11
mmol) of AX and 2.3 wt% (0.07 mmol) of AX, respectively.
Shearzyme is a suitable commercial preparation for the production of AXOS, because it is
abundantly available at relatively low prices compared with the purified enzymes. Purified
AXOS, produced on a milligram scale, were used as standard samples for the
identification and quantification of corresponding AXOS from the hydrolysates in which
all the hydrolysis products were present as a mixture. Self-made standards were essential
because commercial AXOS standards are not available. Pure AXOS were also used as
substrates for fermentation studies in which the influence of different substitution patterns
was evaluated on the ability of bifidobacteria to utilize AXOS. To understand the role of
individual AXOS in the fermentation, purified short-chain AXOS are good models to use,
even though future prebiotic product AXOS with various DP and DS as mixtures may be
used.

84
6.4 Challenges in identification and quantification of AXOS
After the AXOS were purified and their structures fully characterized, identification with
TLC or HPAEC-PAD could be done from the hydrolysates with mixtures of several
AXOS, using purified AXOS as standard compounds. However, identification of AXOS
from the hydrolysates, using only one method, is challenging since on TLC the Rf values
for A2XX and A2+3XX (0.45 and 0.46), D2,3X and XA3X (0.48), and A2X and A2+3X (0.51
and 0.52) are similar (Table 14), while in HPAEC-PAD chromatography A2XX elutes
together with D2,3X (30.1 and 30.2 min), and A3X and XA3X (31.0 min) co-elute with the
same retention times (Figure 9). The use of these two methods is advantageous for correct
identification, because different AXOS, which co-elute in HPAEC-PAD, have similar Rf
values on TLC. The correct identification of AXOS is important for quantification,
because in HPAEC-PAD each oligosaccharide requires the corresponding standard
carbohydrate.
Formation of arabinoxylotriose (XA3X) is difficult to analyse by HPAEC-PAD, due to its
almost simultaneous elution with arabinoxylobiose (A3X) (Kabel et al., 2002a) (I). In
Shearzyme hydrolysates, some XA3X in addition to A3X can be formed. The formation of
XA3X in addition to A3X was shown on TLC plate (I: Figure 7). The major AXOS formed
in the Shearzyme hydrolysis carry �-L-Araf unit(s) at the nonreducing end of the xylose
main chain, and thus A3X is evidently produced most and only minor amounts of XA3X
are usually formed, as analysed using TLC.
Minor amounts of �-L-Araf(1�2)-monosubstituted AXOS (A2X and A2XX) may be
produced during hydrolysis of AX, because all cereal AX contain naturally small amounts
of �-L-Araf(1�2)mono substituents (Izydorczyk and Biliaderis, 1995). However, in
NMR spectroscopy only BHAX, WBAX and RBAX of the AX studied may contain �-L-
Araf(1�2)mono side units, but the identification was uncertain due to overlapping of the
peaks of �-L-Araf(1�2)mono and �-L-Araf(1�3)di (III; Fig. 1). Only BHAX of these
three AX also contains �-D-Xylp(1�2)-�-L-Araf(1�3) side chains, and the
oligosaccharide D2,3X is produced in the Shearzyme hydrolysis of BHAX. If A2XX is
present in the BHAX hydrolysate, it co-elutes with D2,3X in HPAEC-PAD. The peak
intensities in NMR spectroscopy show that �-D-Xylp(1�2)-�-L-Araf(1�3) side chains

85
are more common in BHAX than �-L-Araf(1�2)mono side units, but in HPAEC-PAD
A2XX can be left unidentified due to co-elution with D2,3X. However, A2X can be
identified in HPAEC-PAD and therefore existence of �-L-Araf(1�2)mono side units can
also be shown from the hydrolysis products.
Water-unextractable wheat flour AX contains highly branched and less branched regions.
The highly branched regions are enriched with both �-L-Araf(1�2 and 1�3)
disubstituted and �-L-Araf(1�2) monosubstituted xylose units, but the latter are absent in
less branched areas (Gruppen et al., 1993). Highly branched regions are more challenging
for the enzymes to act on and thus this may also result in underestimation of the amount of
�-L-Araf(1�2) monosubstituted xylose units in AX while examining the hydrolysis
products. However, �-L-Araf(1�2)mono substituents are usually not predominant side
groups since they contribute under 10% of the substituents in water-unextractable wheat
flour AX (Gruppen et al., 1993) and only traces in water-soluble rye flour AX (Vinkx et
al., 1995). The �-L-Araf(1�2)mono substituents are most abundant in alkali-soluble
barley grain and flour AX (22-28% of the substituents) (Viëtor et al., 1992; Trogh et al.,
2005; Pitkänen et al., 2008).
The arabinoxylo-oligosaccharides A3X, XA3X, XA3XX, A2+3X, A2+3XX, A2X, A2XX and
D2,3X were successfully identified, using a combination of TLC and HPAEC-PAD.
Quantification succeeded in A3X, A2+3X, A2+3XX, A2XX, and D2,3X in this study.
However, if both A2XX and D2,3X are identified from the same hydrolysed sample, their
quantification is not accurate, due to simultaneous elution. Analogously, the presence of
A3X and XA3X in the same sample impedes accurate quantification, due to co-elution.
6.5 Behaviour of AXOS in HPAEC-PAD system
6.5.1 Elution order of AXOS
The elution order of the AXOS in HPAEC-PAD system is difficult to predict, since
several interactions between column, eluent and eluting carbohydrates affect elution.
HPAEC-PAD can separate oligosaccharides based on slight variations in their net charge,

86
branching positions or linkage isomerism (Thayer et al., 1998). Carbohydrates are fully
dissociated in the high pH that is used in the HPAEC-PAD method. Thus, essential for
retention in the anion exchange chromatography column is the presence of –OH groups
available for dissociation and interactions with the column material. The tR and pKa values
of carbohydrates correlate inversely and the tR increases with growing molecular mass
(Antec Leyden). Linear XOS differ according to the homologous oligosaccharide series,
and the shorter the XOS are, the earlier they elute from the column.
The arabinose (�-L-Araf) substituents appeared to increase the retention time of the
molecules since the arabinoxylobiose eluted later than xylotriose and xylotetraose.
Furthermore, the increasing amount of �-L-Araf side chains increased retention in the
column material. Thus arabinoxylobioses (A2X and A3X) eluted before diarabinoxylobiose
(A2+3X), while arabinoxylotrioses (A2XX and XA3X) eluted before diarabinoxylotriose
(A2+3XX). The AXOS with the �-L-Araf(1�2)mono unit linked with the �-D-Xylp unit at
the nonreducing end of the xylose chain showed lower retention in the column material
than the corresponding AXOS with �-L-Araf(1�3)mono substituents. Actually, the
linkage type of (1�3) increased the tR substantially, which corresponds well with the
stronger acidity of the 2-OH group than the 3-OH group reported previously (Roberts et
al., 1971). Thus, the substituent at the O-2 of �-D-Xylp prevents the interaction of the 2-
OH group and column, and with the substituent at O-3 of �-D-Xylp, the more acidic 2-OH
group is free to react. With similarly substituted AXOS, the ones with shorter xylose main
chain eluted first. With the �-L-Araf unit attached to the nonreducing terminal of the
xylose chain, the oligosaccharide appeared to elute later than the corresponding AXOS
with internal substituent. This was concluded from the simultaneous elution of A3X and
XA3X.
6.5.2 Responses of AXOS in PAD
The responses for the various AXOS are different in pulsed amperometric detection. The
use of PAD is advantageous, since derivatization of the carbohydrates studied is not
needed and the method is very sensitive. Although carbohydrates can be detected at the
picomolar level, the response is not the same for those oligosaccharides carrying similar

87
substituents attached to different positions of the oligosaccharide main chain (Lee, 1990).
Therefore a quantification standard for each of the different oligosaccharides to be
determined is needed. In this study, the best relative PAD response on a molar basis was
obtained for A3X and the worst for xylotriose and xylotetraose under the HPAEC-PAD
analysis conditions used when the heights of the peaks were compared. In AXOS, with
double �-L-Araf substitution at the nonreducing end, the increasing length of the xylose
chain decreased the PAD response; thus the relative response for A2+3X was better than for
A2+3XX. Furthermore, the lengthening main chain in AXOS with various substitution
patterns caused decreases in PAD response. However, longer AXOS also eluted later,
which may have broadened the peaks and lowered the peak heights. The weakest relative
response of the AXOS studied was obtained for D2,3X. The position of the �-L-Araf
substituent between two xylose units in D2,3X, compared with terminal �-L-Araf unit(s) in
the other AXOS, may have affected the low response. Little information on the PAD
responses of AXOS is found in the literature.
6.6 Fermentation experiments
6.6.1 Utilization of substrates
In the fermentation experiments, the growth of B. adolescentis ATCC 15703, B. longum
ATCC 15707, B. breve ATCC 15700 and faecal microbiota on L-arabinose, D-xylose,
XOS, AXOS and on their mixtures (WAX and RAX hydrolysates) was tested, and the
prebiotic potentials of AXOS were evaluated. The indications of clear substrate
preferences among the various bifidobacterial strains were found. Bifidobacterium
adolescentis was able to grow on XOS, slowly on D-xylose but not on L-arabinose. In
contrast, B. longum preferred L-arabinose and did not grow on pure D-xylose or XOS.
Both strains were able to utilize AXOS but with different strategies, since after the
cleavage of L-arabinose, B. adolescentis consumed the XOS formed, whereas B. longum
fermented the L-arabinose released. Bifidobacterium breve grew poorly on all of the
substrates provided, although it may be able to slowly utilize some xylobiose and AXOS
from a mixture, since some reduction in their quantities was detected after incubation,
compared with the starting situation. Bifidobacterium adolescentis and B. longum were

88
able to grow about equally on AX hydrolysates, but the growth observed was clearly less
than that on FOS, indicating more restricted fermentability of a mixture rich in XOS and
AXOS. All the substrates provided were almost completely utilized by the bacteria from
the faecal microbiota.
The growth of Bifidobacterium adolescentis ATCC 15703, B. longum ATCC 15707, and
B. breve ATCC 15700 on D-xylose, L-arabinose, XOS, and AXOS was previously
reported by several groups and the results obtained in the present study are generally in
agreement with these. B. adolescentis ATCC 1570 earlier showed modest growth on
xylose (Palframan et al., 2003; Moura et al., 2007), although Crittenden et al. (2002) and
Gullón et al. (2008) speculated on the inability of B. adolescentis to utilize xylose. In the
present study, clear growth on pure xylose was detected after a prolonged incubation time
(140 h). Furthermore, HPAEC-PAD analysis showed that pure xylose was consumed by
B. adolescentis ATCC 15703; however, no reduction in xylose content was detected in the
AX hydrolysates. This difference may actually explain the diverging conclusions on the
ability of B. adolescentis ATCC 15703 to ferment xylose. In previous studies, the growth
of B. longum ATCC 15707 was detected with pure xylose (Palframan et al., 2003),
whereas in the present study xylose was utilized by B. longum ATCC 15707 only when
present in a mixture with xylobiose and AXOS. The result that B. longum ATCC 15707
can, but B. adolescentis ATCC 15703 cannot, utilize L-arabinose is well in agreement
with published data (Roy and Ward, 1990; van Laere et al., 1999; Moura et al., 2007).
Previous reports also indicated the inability of B. breve ATCC 15700 to grow on pure
xylose and L-arabinose (Mitsuoka, 1982; Roy and Ward, 1990; Palframan et al., 2003).
The strong growth of B. adolescentis ATCC 15703 on commercial XOS (X2, X3, and X4)
is in line with several previous reports (van Laere et al., 1999, 2000; Moura et al., 2007;
Gullón et al., 2008). The main diverging findings between this study and previous results
were found in the ability of XOS to promote growth of B. longum ATCC 15707. In the
study of Crittenden et al. (2002), B. longum ATCC 15707 (VTT E-96664) was able to
grow effectively on a mixture of XOS (Xylo-oligo 70, Suntory Co. Ltd., Osaka, Japan)
which, however, contained over 40% free monosaccharides, including xylose and some
glucose. Thus, the growth of B. longum could have occurred due to the utilization of the
monosaccharides instead of XOS. Gullón et al. (2008) reported modest growth of B.

89
longum ATCC 15707 on XOS prepared from rice husks. In addition to XOS, the product
contained some free arabinose and xylose, and due to the raw material used for XOS
production, it could also have contained some AXOS, all of which could have promoted
the growth of B. longum ATCC 15707 (Gullón et al., 2008). However, van Laere et al.
(2000) reported the ability of B. longum ATCC 15707 to grow on a xylose-free XOS
mixture (X2, X3 and X4). In the present study, XOS as the only carbon source was too
challenging for B. breve ATCC 15700, which was also observed in the study by Palframan
et al. (2003), while Gullón et al. (2008) reported on the ability of the same strain to grow
on XOS. In the commercial XOS mixture (Wako) which was used as a substrate in pure
bifidobacterial and faecal microbiota fermentations, some of the unidentified
oligosaccharides may have been AXOS, since Moura et al. (2007) reported the presence of
AXOS in the commercial XOS product (Xylo-oligo 95P; Suntory). However, the possible
presence of AXOS is not desired in XOS, since it may affect the ability of bifidobacteria
to utilize XOS when the utilization of linear XOS is evaluated.
Studies with AXOS are rarer than with XOS. van Laere et al. (2000) reported that AXOS
(mainly L-Araf doubly substituted XOS) were fermented partially by B. adolescentis
ATCC 15703 and completely by B. longum ATCC 15707. This supports our data that both
strains are able to consume AXOS, but we demonstrated partial utilization of AXOS by
both species; i.e. B. adolescentis ATCC 15703 used the backbone XOS but not the
released L-Araf substituents, and B. longum ATCC 15707 utilized the L-Araf released but
not the residual XOS.
In the present study, a small-scale method and extremely nutrient-poor medium were
applied. The use of this small-scale method was necessary for testing pure AXOS that are
difficult to obtain in large amounts. To obtain comparable results, all the experiments were
performed on a small scale. The downscaling of the cultivation volume of the pure
cultures had to be performed in test tubes where normally 10 ml of medium are added to
secure the preservation of anaerobic conditions and the reliable measurement of OD. The
small (3 ml) volume changed the proportions of the medium and the airspace, and possibly
made it more challenging to obtain completely anaerobic conditions. Furthermore, this
small volume hindered the usage of the reducing agent sodium sulphide (Na2S), which
may also have changed the growth environment. Long cultivation times, up to 140 h, were

90
used in the present study to ensure growth under the somewhat atypical conditions
applied. In other reported studies much shorter fermentation times were used, the purity
and composition of the XOS and AXOS varied and different neutral basal media were
applied. Variations between this and previous studies could have partially occurred for
these reasons.
6.6.2 Substrate utilization strategies of pure bifidobacteria
Bifidobacteria are presumed to utilize oligosaccharides either by transporting small
oligosaccharides into the cell and hydrolysing them further with intracellular enzymes, or
by hydrolysing the oligosaccharides with extracellular enzymes and transporting the
monosaccharides formed into the cell (Amaretti et al., 2006). Stronger growth of B.
adolescentis ATCC 15703 on XOS than on xylose indicated that short XOS (X2, X3) are
transported into the cell rather than hydrolysed extracellularly to xylose. Similar
preferences of the same bacterial strain for di- and oligosaccharides over monosaccharides
were also noted in previous studies (Crittenden et al., 2002; Palframan et al., 2003;
Amaretti et al., 2006). Furthermore, Palframan et al. (2003) showed that B. adolescentis
ATCC 15703 produces extracellular �-D-xylosidase activity when cultivated on XOS. The
presence of extracellular �-D-xylosidase was also indicated in the present study, since the
xylose formed was detected after cultivation of B. adolescentis ATCC 15703 on XOS and
AX hydrolysates. An increase in XOS chain length to DP 4-6 reduces the efficacy of B.
adolescentis ATCC 15703 in utilizing these XOS (Moura et al., 2007; Gullón et al., 2008),
most probably because longer XOS need to be first hydrolysed by an extracellular enzyme
to shorter XOS. In contrast to B. adolescentis ATCC 15703, Palframan et al. (2003)
reported that B. longum ATCC 15707 showed no extracellular �-D-xylosidase activity,
which agrees with the present study in which no consumption of xylobiose was detected.
From the data obtained it seems evident that AXOS are hydrolysed extracellularly by �-L-
arabinofuranosidases. Both B. adolescentis ATCC 15703 and B. longum ATCC 15707
were able to cleave the linkage between L-arabinose and xylose, since no short-chain
AXOS were detected after cultivations with AX hydrolysates. Bifidobacterium
adolescentis ATCC 15703 is known to express two distinct extracellular �-L-
arabinofuranosidases (van Laere et al., 1999; van Den Broek and Voragen, 2008).

91
Bifidobacterium longum ATCC 15707 undoubtedly also produces �-
arabinofuranosidase(s) analogously to other B. longum strains (Crittenden et al., 2002;
Gueimonde et al., 2007), although the presence of �-L-arabinofuranosidase-encoding
genes has not yet been shown.
Bifidobacterium breve K-110, isolated from human intestine, also produces �-D-
xylosidase and �-L-arabinofuranosidase (Shin et al., 2003a, 2003b). On the other hand,
preliminary results from the genome sequence of B. breve UCC2003 show the absence of
AX-degrading enzymes (van Den Broek and Voragen, 2008). In the present and previous
studies, B. breve ATCC 15700 was unable to grow on xylose or L-arabinose (Mitsuoka,
1982; Roy and Ward, 1990; Palframan et al., 2003). Degnan and Macfarlane (1993)
showed that to transport L-arabinose, B. breve NCFB 2257 requires glucose in the
cultivation medium, indicating cotransportation. The lack of cotransportation could be, in
addition to lack of enzymes for the hydrolysis of (A)XOS, another potential explanation
for the poor growth of B. breve ATCC 15700 when xylose, L-arabinose, and (A)XOS
were the only carbohydrates present in the cultivation medium.
After incubation of B. adolescentis with the WAX and RAX hydrolysates, twice as much
free L-arabinose was found in the samples than could be formed from the short, quantified
AXOS in the hydrolysates. This indicates that L-arabinose is also liberated from the
longer, unidentified AXOS present in the samples. Furthermore, B. longum was not able to
utilize xylobiose and xylotriose from the WAX and RAX hydrolysates, and the amounts
of these short XOS detected in the samples after fermentation were, surprisingly, 7-8 times
more than could be released from the quantified AXOS. Additional xylobiose and
xylotriose may also have originated in this case from longer AXOS. After releasing the L-
Araf substituents, B. longum may be able to hydrolyze the resulting XOS further, e.g.
using specific (exo)xylanase as is produced by Aeromonas caviae ME-1 (Kubata et al.,
1994). The formation of more xylobiose and xylotriose during growth of B. longum is
rather interesting, because it does not consume these carbohydrates.
Some of the results concerning the utilization of D-xylose, L-arabinose and XOS were
inconsistent with the literature, which may have been due to several reasons, from
different experimental conditions to various substrates and difficulties in interpretation of

92
the results. In the present study, bifidobacteria were able to utilize some substrates only
under extreme conditions, when more suitable substrates were not available, or on the
other hand, only when other substrates were also present to activate the utilization of more
challenging carbohydrates. The initial carbohydrate content of the substrates was
investigated, and in addition to observing only the growth of bacteria, the concentration of
individual carbohydrates was followed by HPAEC-PAD to furnish proof of which
carbohydrates induced the growth.
6.6.3 Specific features in utilization of purified AXOS
The fermentation experiments with pure AXOS revealed that monosubstituted A2XX is an
easier substrate than the doubly substituted A2+3XX, since the former was completely
fermented both by a mixture of pure bifidobacteria and by faecal microbiota, whereas the
latter was utilized only by the faecal bacteria mixture. However, even after the growth of
the faecal microbiota some mono- and oligosaccharides were detected, including A2XX
formed after cleavage of a single L-Araf unit from the doubly substituted D-Xylp residue
in A2+3XX. Removal of the first L-Araf unit appears to be challenging, but after this,
complete fermentation of the singly substituted AXOS formed can be achieved.
Consumption of the doubly substituted AXOS was, however, slow since carbohydrates
still detected after a 48-h incubation of A2+3XX were readily utilized in other
fermentations. Thus, it can be assumed that only a few bacterial species in the human
faecal microbiota were able to liberate �-1�3 -linked L-Araf units from the doubly
substituted D-Xylp residue. Furthermore, after fermentation of faecal microbiota on
A2+3XX, the pH was 4 and the growth of intestinal bifidobacterial species at pH lower than
4.5 is very limited (Biavati and Mattarelli, 2006), suggesting clearly that bacteria other
than bifidobacteria ferment disubstituted AXOS. This is supported by the altered
fermentation pattern seen in the VFA distribution in Table 18.
The t-RFLP analysis of faecal samples indicated that various AXOS may result in
different distributions of bifidobacteria, since not all AXOS support growth of the same
bifidobacterial species. With doubly substituted AXOS as substrates, it is unclear if the
slight change in distribution of bifidobacteria was due to more rapid death of group 2
bacteria or increased growth of group 1 bacteria. The t-RFLP analysis did not provide

93
quantities of bifidobacteria, but decrease in the bifidobacterial amount is presumable if
other bacteria were utilizing pure disubstituted AXOS, as suggested above. Since the
doubly substituted AXOS were consumed by pure B. adolescentis ATCC 15703 and B.
longum ATCC 15707 from the WAX hydrolysate, the pure A2+3XX may not have been
able to induce the production of essential �-L-arabinofuranosidases. Gueimonde et al.
(2007) reported that the GH 51 �-L-arabinofuranosidase of B. longum NIZO B667 was
induced by xylose and to a lesser extent by L-arabinose.
Interestingly, B. adolescentis ATCC 15703 did not ferment L-arabinose, although it has �-
L-arabinofuranosidases. The need for this organism to produce these enzymes may be to
degrade AXOS to obtain xylose, not L-arabinose. It is also tempting to hypothesize that
the liberation of L-arabinose as a result of the action of B. adolescentis �-L-
arabinofuranosidases may provide a carbon source for other bifidobacteria, e.g. B. longum,
indicating the complex synergisms of probiotic bacteria. This was indeed observed in the
present study when pure A2XX was completely consumed by the mixture of B.
adolescentis, B. breve and B. longum. The other indication of a possible versatile cross-
synergy between B. adolescentis and B. longum was the buildup of xylobiose and
xylotriose by B. longum when grown on AX hydrolysates, because the short XOS are the
preferred substrates of B. adolescentis.
Detailed chromatographic analysis of mono- and oligosaccharides after the fermentations
revealed new insight into carbohydrate, particularly AXOS, consumption by the
bifidobacteria studied. New knowledge of the inability of B. adolescentis ATCC 15703 to
consume the L-arabinose it cleaved from various AXOS was also obtained. Based on our
results and combined with previous studies by other groups, short (A)XOS support growth
of B. adolescentis ATCC 15703, whereas highly substituted AXOS, and even polymeric
AX, support the growth of B. longum ATCC 15707, utilizing primarily the L-Araf
substituents. AXOS are clearly potential candidates for selective prebiotics, but the
fermentation efficiency and the prebiotic effect appear to be dependent on the type of L-
Araf substitution. AXOS with doubly substituted D-Xylp were fermented less efficiently
than AXOS with singly substituted D-Xylp. Although isolated A2+3XX was not fermented
by the mixture of pure bifidobacterial strains, it was consumed by the versatile faecal
microbiota. Doubly substituted AXOS were also utilized by B. adolescentis ATCC 15703

94
and B. longum ATCC 15707 when present in the hydrolysate together with xylose, XOS,
and monosubstituted AXOS. Thus, a mixture of different singly and doubly substituted
AXOS could function as a suitable, slowly fermenting, prebiotic substance. The results
also reflected a complex cooperation between bifidobacteria in the consumption of AXOS.
In the present study, pure AX-derived saccharides were used as the only carbon source,
but in the colon there are also other carbohydrates available. This may contribute to
fermentation of the challenging AXOS through the activation of hydrolytic enzymes,
transport systems or activation of the bifidus pathway. Thus, further research is needed on
the interactions of different putative prebiotics in relation to the growth of probiotic
bacteria.

95
Conclusions
Cereal AX are efficiently hydrolysed by Shearzyme (A. aculeatus GH 10 endo-1,4-�-D-
xylanase) into xylose, XOS and AXOS. This enzyme is active on the xylosidic linkage at
the nonreducing end of the xylose chain, prior to both mono �-L-Araf(1�2)- and �-L-
Araf(1�3)-substituted �-D-Xylp units, as well as doubly �-L-Araf(1�2)- and (1�3)-
substituted �-D-Xylp residue and also 2-O-�-D-Xylp-�-L-Araf side chain attached to the
�-D-Xylp unit. Thus, Shearzyme produces short-chain AXOS with nonreducing end
substituents. The yield of the XOS and AXOS was amplified, using a combination of
different �-L-arabinofuranosidases (AXH-m and AXH-d3) to modify the substitution
pattern of AX before the endo-1,4-�-D-xylanase treatment. Furthermore, diversely
substituted AXOS were prepared with stepwise hydrolysis by a combination of preceding
enzymes.
Differences in the structures of several AX extracted from various cereal plant parts were
detected by studying the structure of polymeric AX with NMR spectroscopy and by
examining the hydrolysis products with HPAEC-PAD. These two methods are supporting,
since NMR spectroscopy provides information on the entire AX molecule from the water-
soluble part of the sample, but information on the water-insoluble part is lost. Using
hydrolysis products in the elucidation of the AX, complete solubility is not vital in
preparing the hydrolysates, but the most highly substituted areas of AX probably remain
intact by the enzyme. WAX-mv, WBAX, RAX, RBAX and OBAX are fairly similar with
relatively high DS levels. These flour and bran AX contain high amounts of �-L-
Araf(1�3)mono substituents and �-L-Araf(1�2 and 1�3)di residues. Both BHAX and
CCAX contain abundant less common �-D-Xylp(1�2)-�-L-Araf(1�3) side chains.
RiHAX, WStAX and OSAX have low DS levels of �-L-Araf mono- and disubstituted
xylose residues, but they carry some �-D-Xylp(1�2)-�-L-Araf(1�3) substituents. In
contrast to these, flour and bran AX, RiHAX, OSAX, CCAX and WStAX contain
abundant 4-O-Me-�-D-GlcA(1�2) units.
The purified AXOS prepared in good yields in this study were: A3X, A2X, A2XX, A2+3X,
A2+3XX and D2,3X. With doubly substituted AXOS, A2+3XX is the main oligosaccharide
formed in the hydrolysis and A2+3X is a minor product, in contrast to the corresponding

96
monosubstituted AXOS from which A2X is the major and A2XX the minor hydrolysis
product. The purified AXOS were invaluable tools for studing the elution order and
response in HPAEC-PAD, and they were further used as standard compounds in TLC and
HPAEC-PAD analysis.
Complete separation of the AXOS is essential for correct quantification. With the
HPAEC-PAD method used, good separation of A3X, A2X, A2+3X and A2+3XX was
obtained, but A2XX and D2,3X were not totally separated. Usually A2XX is rarely
obtained, since �-L-Araf(1�2)mono is a minor substituent in AX. A2XX and D2,3X were
separated on TLC, which can be used in combination with HPAEC-PAD to identify the
AXOS. For the quantification of AXOS and other carbohydrates produced in the
hydrolysis, corresponding standards are needed for each compound because the peak
heights cannot be compared as such, since the responses to the different monosaccharides,
XOS and AXOS vary in PAD.
For the first time, the prebiotic potential of the purified AXOS were studied in in vitro
fermentation experiments. AXOS, L-arabinose, D-xylose, XOS, RAX and WAX
hydrolysates, and FOS (positive control) were used as substrates for pure bifidobacterial
and faecal microbiota. Based on the results obtained, B. adolescentis ATCC 15703 is able
to utilize XOS, some D-xylose, but not L-arabinose. B. longum ATCC 15707 prefers L-
arabinose, but is not able to grow on pure D-xylose or XOS. B. breve ATCC 15700 grew
poorly on all of the substrates provided. Detailed new information on the fate of individual
AXOS during fermentation was obtained. Both B. adolescentis and B. longum were able
to grow on AXOS, but they fermented oligosaccharides with differing strategies, since
after cleavage of the L-arabinose B. adolescentis utilized the XOS formed, whereas B.
longum utilized the L-arabinose released. The results show that the ability to grow on
certain AXOS does not always mean complete utilization. Pure AXOS with doubly
substituted D-Xylp were fermented less efficiently than AXOS with singly substituted D-
Xylp by the mixture of three bifidobacteria. However, doubly substituted AXOS were also
utilized by B. adolescentis and B. longum when present in a hydrolysate together with
xylose, XOS and monosubstituted AXOS. Isolated doubly substituted AXOS were
fermented by versatile faecal microbiota. Thus, a mixture of different singly and doubly
substituted AXOS could function as a suitable, slowly fermenting prebiotic substance.

97
References
AACC. 2001. The definition of dietary fibre. Cereal Foods World 46, 112-126.
Alonso, J.L., Domínguez, H., Garrote, G., Parajó, J.C. and Vázquez, M.J. 2003. Xylooligosaccharides: properties and production technologies. Electron. J. Environ. Agric. Food Chem. 2.
Amaretti, A., Tamburini, E., Bernardi, T., Pompei, A., Zanoni, S., Vaccari, G., Matteuzzi, D. and Rossi, M. 2006. Substrate preference of Bifidobacterium adolescentis MB 239: compared growth on single and mixed carbohydrates. Appl. Microbiol. Biotechnol. 73, 654-662.
Andersson, R. and Åman, P. 2001. Cereal arabinoxylan: Occurrence, structure, and properties. Adv. Dietary Fibre Technol. 301-314.
Andrewartha, K.A., Phillips, D.R. and Stone, B.A. 1979. Solution properties of wheat flour arabinoxylans and enzymatically modified arabinoxylans. Carbohydr. Res. 77, 191-204.
Angelidaki, I., Petersen, S.P. and Ahring, B.K. 1990. Effects of lipids on thermophilic anaerobic digestion and reduction of lipid inhibition upon addition of bentonite. Appl. Microbiol. Biotechnol. 33, 469-472.
Antec Leyden. http://www.jasco.hu/konyvtar/220_002_01.pdf.
Aspinall, G.O. 1959. Structural chemistry of the hemicelluloses. Adv. Carbohydr. Chem. 14, 429-468.
Aspinall, G.O. 1970. Arabinans and Xylans. In: Polysaccharides. Pergamon Elmsford, New York, pp. 103-115.
Aspinall, G.O. 1980. Chemistry of cell wall polysaccharides. Biochem. Plants 3, 473-500.
Aspinall, G.O. and Ferrier, R.J. 1957. The constitution of barley-husk hemicellulose. J. Chem. Soc. 4188-4194.
Aspinall, G. and Ferrier, R.J. 1958. Synthesis of 2-[O-(�-D-xylopyranosyl)]-L-arabinose and its isolation from the partial hydrolysis of esparto hemicellulose. J. Chem. Soc. 1501-1505.
Bacic, A. and Stone, B. 1980. A (1�3)- and (1�4)-linked �-glucan in the endosperm cell-walls of wheat. Carbohydr. Res. 82, 372-377.
Bacic, A. and Stone, B.A. 1981. Chemistry and organization of aleurone cell wall components from wheat and barley. Aust. J. Plant Physiol. 8, 475-495.
Bataillon, M., Mathaly, P., Nunes Cardinali A.-P. and Duchiron, F. 1998. Extraction and purification of arabinoxylan from destarched wheat bran in a pilot scale. Ind. Crop. Prod. 8, 37-43.

98
Beaugrand, J., Chambat, G., Wong, V.W.K., Goubet, F., Rémond, C., Paës, G., Benamrouche, S., Debeire, P., O’Donohue, M. and Chabbert, B. 2004. Impact and efficiency of GH10 and GH11 thermostable endoxylanases on wheat bran and alkali-extractable arabinoxylans. Carbohydr. Res. 339, 2529-2540.
Beldman, G., Osuga, D. and Whitaker, J.R. 1996. Some characteristics of �-xylopyranosidases, �-arabinofuranosidases and an arabinoxylan �-arabinofuranohydrolase from wheat bran and germinated wheat. J. Cereal Sci. 23, 169-180.
Biavati, B. and Mattarelli, P. 2006. The family bifidobacteriaceae. In: The Prokaryotes. Dworkin, M., Falkow, S., Rosenberg, E. and Schleifer, K.-H. (Eds.), Springer, New York, pp. 3322-3382.
Bicking, M.K.L. 2006. Integration errors in chromatographic analysis, Part I: Peaks of approximately equal size. (Cover story). LC-GC North America 24, 402-414.
Bielecka, M., Biedrzycka, E. and Majkowska, A. 2002. Selection of probiotics and prebiotics for synbiotics and confirmation of their in vivo effectiveness. Food Res. Int. 35, 125-131.
Biely, P. 1985. Microbial xylanolytic systems. Trends Biotechnol. 3, 286-290.
Biely, P., Vrsanska, M., Tenkanen, M. and Kluepfel D. 1997. Endo-�-1,4-xylanase families: differences in catalytic properties. J. Biotechnol. 57, 151-166.
Brillouet, J.M., Joseleau J.P., Utille J.P. and Lelievre D. 1982. Isolation, purification and characterization of a complex heteroxylan from industrial wheat bran. J. Agric. Food Chem. 30, 488-495.
Broberg, A., Thomsen, K.K. and Duus, J.O. 2000. Application of nano-probe NMR for structure determination of low nanomole amounts of arabinoxylan oligosaccharides fractionated by analytical HPAEC-PAD. Carbohydr. Res. 328, 375-382.
Campbell, J.M., Fahey G.C., Jr. and Wolf B.W. 1997a. Selected indigestible oligosaccharides affect large bowel mass, cecal and fecal short-chain fatty acids, pH and microflora in rats. J. Nutr. 127, 130-136.
Campbell, J.M., Fahey, G.C., Jr, Demichele, S.J. and Garleb, K.A. 1997b. Metabolic characteristics of healthy adult males as affected by ingestion of a liquid nutritional formula containing fish oil, oligosaccharides, gum arabic and antioxidant vitamins. Food Chem. Toxicol. 35, 1165-1176.
Carpita, N.C. and Gibeaut, D.M. 1993. Structural models of primary cell walls in flowering plants: consistency of molecular structure with the physical properties of the walls during growth. Plant J. 3, 1-30.
CAZy - Carbohydrate Active enZymes. www.cazy.org
Cleemput, G., van Laere, K., Hessing, M., Van Leuven, F., Torrekens, S. and Delcour, J.A. 1997. Identification and characterization of a novel arabinoxylanase from wheat flour. Plant Physiol. 115, 1619-1627.

99
Cloetens, L., De Preter, V., Swennen, K., Broekaert, W.F., Courtin, C.M., Delcour, J.A., Rutgeerts, P. and Verbeke, K. 2008. Dose-response effect of arabinoxylooligosaccharides on gastrointestinal motility and on colonic bacterial metabolism in healthy volunteers. J. Am. Coll. Nutr. 27, 512.
Collins, M.D. and Gibson, G.R. 1999. Probiotics, prebiotics, and synbiotics: approaches for modulating the microbial ecology of the gut. Am. J. Clin. Nutr. 69, 1052S-1057.
Collins, T., Gerday, C. and Feller, G. 2005. Xylanases, xylanase families and extremophilic xylanases. FEMS Microbiol. Rev. 29, 3-23.
Courtin, C.M. and Delcour, J.A. 2001. Relative activity of endoxylanases towards water-extractable and water-unextractable arabinoxylan. J. Cereal Sci. 33, 301-312.
Crittenden, R.G. and Playne, M.J. 1996. Production, properties and applications of food-grade oligosaccharides. Trends Food Sci. Technol. 7, 353-361.
Crittenden, R., Karppinen, S., Ojanen, S., Tenkanen, M., Fagerström, R., Mättö, J., Saarela, M., Mattila-Sandholm, T. and Poutanen, K. 2002. In vitro fermentation of cereal dietary fibre carbohydrates by probiotic and intestinal bacteria. J. Sci. Food Agric. 82, 781-789.
Cyran, M., Courtin, C.M. and Delcour, J.A. 2003. Structural features of arabinoxylans extracted with water at different temperatures from two rye flours of diverse breadmaking quality. J. Agric. Food Chem. 51, 4404-4416.
Cyran, M., Courtin, C.M. and Delcour, J.A. 2004. Heterogeneity in the fine structure of alkali-extractable arabinoxylans isolated from two rye flours with high and low breadmaking quality and their coexistence with other cell wall components. J. Agric. Food Chem. 52, 2671-2680.
De Preter, V., Geboes, K., Verbrugghe, K., De Vuyst, L., Vanhoutte, T., Huys, G., Swings, J., Pot, B. and Verbeke, K. 2004. The in vivo use of the stable isotope-labelled biomarkers lactose-[N]ureide and [H4]tyrosine to assess the effects of pro- and prebiotics on the intestinal flora of healthy human volunteers. Br. J. Nutr. 92, 439.
Degnan, B.A. and Macfarlane, G.T. 1993. Transport and metabolism of glucose and arabinose in Bifidobacterium breve. Arch. Microbiol. 160, 144-151.
Dekker, R.F.H. and Richards, G.N. 1975. Purification, properties, and mode of action of hemicellulase II produced by Ceratocystis paradoxa. Carbohydr. Res. 42, 107-123.
Delcour, J.A., Rouseu, N. and Vanhaesendonck, I.P. 1999. Pilot-scale isolation of water-extractable arabinoxylans from rye. Cereal Chem. 76, 1-2.
Dervilly, G., Leclercq, C., Zimmermann, D., Roue, C., Thibault, J.F. and Saulnier, L. 2002. Isolation and characterization of high molar mass water-soluble arabinoxylans from barley and barley malt. Carbohydr. Polym. 47, 143-149.

100
Dervilly-Pinel, G., Rimsten, L., Saulnier, L., Andersson, R. and Åman, P. 2001. Water-extractable arabinoxylan from pearled flours of wheat, barley, rye and triticale. Evidence for the presence of ferulic acid dimers and their involvement in gel formation. J. Cereal Sci. 34, 207-214.
Duncan, S.H., Louis, P. and Flint, H. 2004. Lactate-utilizing bacteria, isolated from human feces, that produce butyrate as a major fermentation product. Appl. Environ. Microb. 70, 5810-5817.
Dyson, N.A. 1998. Peak area vs peak height. In: Chromatographic integration methods. Humana Press, Totowa, N.J. USA, pp. 83-88.
Ebringerová, A., Hromádková, Z., Petráková, E. and Hricovíni, M. 1990. Structural features of a water-soluble L-arabino-D-xylan from rye bran. Carbohydr. Res. 198, 57-66.
Ebringerová, A., Hromádková, Z., Alfoldi, J. and Berth, G. 1992. Structural and solution properties of corn cob heteroxylans. Carbohydr. Polym. 19, 99-105.
Ebringerová, A., Hromádková Z., Burchard W., Dolega R. and Vorwerg W. 1994. Solution properties of water-insoluble rye-bran arabinoxylan. Carbohydr. Polym. 24, 161-169.
Ebringerová, A. and Hromádková Z. 1997. The effect of ultrasound on the structure and properties of the water-soluble corn hull heteroxylan. Ultrason. Sonochem. 4, 305-309.
Ebringerová, A. and Heinze, T. 2000. Xylan and xylan derivatives - biopolymers with valuable properties. 1. Naturally occurring xylans structures, isolation procedures and properties. Macromol. Rapid Commun. 21, 542-556.
EFSA (European Food Safety Authority). 2009. Scientific opinion (draft); Dietary reference values for carbohydrates and dietary fibre. EFSA J. 1-76.
Ewald, C.M. and Perlin, A.S. 1959. Arrangement of branching in an arabino-xylan from wheat flour. Can. J. Chem. 37, 1254-1259.
Faulds, C.B., Sancho, A.I. and Bartolomé, B. 2002. Mono- and dimeric ferulic acid release from brewer's spent grain by fungal feruloyl esterases. Appl. Microbiol. Biotechnol. 60, 489-494.
Fauré, R., Courtin, C.M., Delcour, J.A., Dumon, C., Faulds, C.B., Fincher, G.B., Fort, S., Fry, S.C., Halila, S., Kabel, M.A., Pouvreau, L., Quemener, B., Rivet, A., Saulnier, L., Schols, H.A., Driquez, H. and O´Donohue, M.J. 2009. A brief and informationally rich naming system for oligosaccharide motifs of heteroxylans found in plant cell walls. Aust. J. Chem. 62, 1-5.
Ferré, H., Broberg, A., Duus, J.O. and Thomsen, K.K. 2000. A novel type of arabinoxylan arabinofuranohydrolase isolated from germinated barley. Analysis of substrate preference and specificity by nano-probe NMR. Eur. J. Biochem. 267, 6633-6641.
Fincher, G.B. 1975. Morphology and chemical composition of barley endosperm cell walls. J. Inst. Brewing 81, 116-122.

101
Fujimoto, Z., Kaneko, S., Kuno, A., Kobayashi, H., Kusakabe, I. and Mizuno, H. 2004. Crystal structures of decorated xylooligosaccharides bound to a family 10 xylanase from Streptomyces olivaceoviridis E-86. J. Biol. Chem. 279, 9606-9614.
Fulcher, R.G. and Duke, T.K.R. 2002. Whole-grain structure and organization: Implications for nutritionists and processors. In: Whole-grain foods in health and disease. Marquart, L., Slavin, J.L. and Fulcher, R.G. (Eds.), American Association of Cereal Chemists, St. Paul, Minnesota, USA, pp. 9-45.
Garrote, G. and Parajó, J.C. 2002. Non-isothermal autohydrolysis of Eucalyptus wood. Wood Sci. Technol. 36, 111-123.
Gibson, G.R. and Roberfroid, M.B. 1995. Dietary modulation of the human colonic microbiota: introducing the concept of prebiotics. J. Nutr. 125, 1401-1412.
Gibson, G.R., Probert, H.M., van Loo, J., Rastall, R.A. and Roberfroid, M.B. 2004. Dietary modulation of the human colonic microbiota: updating the concept of prebiotics. Nutr. Res. Rev. 17, 259-275.
Glitsø, L.V. and Bach Knudsen, K.E. 1999. Milling of whole grain rye to obtain fractions with different dietary fibre characteristics. J. Cereal Sci. 29, 89-97.
Grootaert, C., Delcour, J.A., Courtin, C.M., Broekaert, W.F., Verstraete, W. and Van de Wiele, T. 2007. Microbial metabolism and prebiotic potency of arabinoxylan oligosaccharides in the human intestine. Trends Food Sci. Tech. 18, 64-71.
Grootaert, C., Van den Abbeele, P., Marzorati, M., Broekaert, W.F., Courtin, C.M., Delcour, J.A., Verstraete, W. and Van de Wiele, T. 2009. Comparison of prebiotic effects of arabinoxylan oligosaccharides and inulin in a simulator of the human intestinal microbial ecosystem. FEMS Microbiol. Ecol. 69, 231-242.
Gruppen, H., Hamer, R.J. and Voragen, A.G.J. 1991. Barium hydroxide as a tool to extract pure arabinoxylans from water-insoluble cell wall material of wheat flour. J. Cereal Sci. 13, 275-290.
Gruppen, H., Hoffmann, R.A., Kormelink, F.J., Voragen, A.G., Kamerling, J.P. and Vliegenthart, J.F. 1992. Characterisation by 1H NMR spectroscopy of enzymically derived oligosaccharides from alkali-extractable wheat-flour arabinoxylan. Carbohydr. Res. 233, 45-64.
Gruppen, H., Kormelink, F.J.M. and Voragen, A.G.J. 1993. Water-unextractable cell wall material from wheat flour. 3. A structural model for arabinoxylans. J. Cereal Sci. 18, 111-128.
Gueimonde, M., Noriega, L., Margolles, A. and de los Reyes-Gavilán, C. 2007. Induction of �-L-arabinofuranosidase activity by monomeric carbohydrates in Bifidobacterium longum and ubiquity of encoding genes. Arch. Microbiol. 187, 145-153.
Gullón, P., Moura, P., Esteves, M.P., Girio, F.M., Domínguez, H. and Parajó, J.C. 2008. Assessment on the fermentability of xylooligosaccharides from rice husks by probiotic bacteria. J. Agric. Food Chem. 56, 7482-7487.

102
Hardy, M.R. and Townsend, R.R. 1988. Separation of positional isomers of oligosaccharides and glycopeptides by high-performance anion-exchange chromatography with pulsed amperometric detection. PNAS, Proc. Natl. Acad. Sci. USA 85, 3289-3293.
Härkönen, H., Pessa, E., Suortti, T. and Poutanen, K. 1997. Distribution and some properties of cell wall polysaccharides in rye milling fractions. J. Cereal Sci. 26, 95-104.
Henry, R.J. 1985. A comparison of the non-starch carbohydrates in cereal grains. J. Sci. Food Agric. 36, 1243-1253.
Hoffmann, R.A., Leeflang, B.R., de Barse, M.M., Kamerling, J.P. and Vliegenthart, J.F. 1991. Characterisation by 1H-n.m.r. spectroscopy of oligosaccharides, derived from arabinoxylans of white endosperm of wheat, that contain the elements �4)[�-L-Araf-(1�3)]-�-D-Xylp-(1� or �4)[�-L-Araf-(1�2)][�-L-Araf-(1�3)]-�-D-Xylp-(1�. Carbohydr. Res. 221, 63-81.
Hoffmann, R.A., Kamerling, J.P. and Vliegenthart, J.F.G. 1992. Structural features of a water-soluble arabinoxylan from the endosperm of wheat. Carbohydr. Res. 226, 303-311.
Höije, A., Gröndahl, M., Tømmeraas, K. and Gatenholm, P. 2005. Isolation and characterization of physicochemical and material properties of arabinoxylans from barley husks. Carbohydr. Polym. 61, 266-275.
Höije, A., Sandström C., Roubroeks J.P., Andersson R., Gohil S. and Gatenholm P. 2006. Evidence of the presence of 2-O-�-D-xylopyranosyl-�-L-arabinofuranose side chains in barley husk arabinoxylan. Carbohydr. Res. 341, 2959-2966.
Höije, A., Sternemalm, E., Heikkinen, S., Tenkanen, M. and Gatenholm, P. 2008. Material properties of films from enzymatically tailored arabinoxylans. Biomacromolecules 9, 2042-2047.
Hollmann, J. and Lindhauer, M.G. 2005. Pilot-scale isolation of glucuronoarabinoxylans from wheat bran. Carbohydr. Polym. 59, 225-230.
Holzapfel, W.H. and Schillinger, U. 2002. Introduction to pre- and probiotics. Food Res. Int. 35, 109-116.
Hopkins, M.J., Englyst, H.N., Macfarlane, S., Furrie, E., Macfarlane, G.T. and McBain, A.J. 2003. Degradation of cross-linked and non-cross-linked arabinoxylans by the intestinal microbiota in children. Appl. Environ. Microbiol. 69, 6354-6360.
Hromádková, Z. and Ebringerová, A. 1987. Isolation and characterization of hemicelluloses from rye bran. Nahrung 31, 149-156.
Hromádková, Z., Ebringerová, A., Petráková, E. and Schraml, J. 1987. Structural features of a rye-bran arabinoxylan with a low degree of branching. Carbohydr. Res. 163, 73-79.
Hromádková, Z. and Ebringerová, A. 1995. Isolation and characterization of hemicelluloses of corn hulls. Chem. Pap. 49, 97-101.

103
Hughes, S.A., Shewry, P.R., Li, L., Gibson, G.R., Sanz, M.L. and Rastall, R.A. 2007. In vitro fermentation by human fecal microflora of wheat arabinoxylans. J. Agric. Food Chem. 55, 4589-4595.
Imaizumi, K., Nakatsu, Y., Sato, M., Sedarnawati, Y. and Sugano, M. 1991. Effects of xylooligosaccharides on blood glucose, serum and liver lipids and cecum short-chain fatty acids in diabetic rats. Agric. Biol. Chem. 55, 199-205.
Ishii, T. 1997. Structure and functions of feruloylated polysaccharides. Plant Sci. 127, 111-127.
Izydorczyk, M.S. and Biliaderis, C.G. 1993. Structural heterogeneity of wheat endosperm arabinoxylans. Cereal Chem. 70, 641-646.
Izydorczyk, M.S. and Biliaderis, C.G. 1995. Cereal arabinoxylans: advances in structure and physiochemical properties. Carbohydr. Polym. 28, 33-48.
Jaskari, J., Kontula, P., Siitonen, A., Jousimies-Somer, H., Mattila-Sandholm, T. and Poutanen, K. 1998. Oat beta-glucan and xylan hydrolysates as selective substrates for Bifidobacterium and Lactobacillus strains. Appl. Microbiol. Biotechnol. 49, 175-181.
Jeffries, T.W. 1990. Biodegradation of lignin-carbohydrate complexes. Biodgradation, 1, 163-176.
Johansson, L., Virkki, L., Anttila, H., Esselström, H., Tuomainen, P. and Sontag-Strohm, T. 2006. Hydrolysis of �-glucan. Food Chem. 97, 71-79.
Johnson, D.C. and LaCourse, W.R. 1990. Liquid chromatography with pulsed electrochemical detection at gold and platinum electrodes. Anal. Chem. 62, 589A-597A.
Johnson, D.C., Dobberpuhl, D., Roberts, R. and Vandeberg, P. 1993. Pulsed amperometric detection of carbohydrates, amines and sulfur species in ion chromatography. The current state of research. J. Chromatogr. 640, 79-96.
Kabel, M.A., Carvalheiro, F., Garrote, G., Avgerinos, E., Koukios, E., Parajó, J.C., Girio, F.M., Schols, H.A. and Voragen, A.G.J. 2002a. Hydrothermally treated xylan rich by-products yield different classes of xylo-oligosaccharides. Carbohydr. Polym. 50, 47-56.
Kabel, M.A., Kortenoeven, L., Schols, H.A. and Voragen, A.G.J. 2002b. In vitro fermentability of differently substituted xylo-oligosaccharides. J. Agric. Food Chem. 50, 6205-6210.
Kabel, M.A., Schols, H.A. and Voragen, A.G.J. 2002c. Complex xylo-oligosaccharides identified from hydrothermally treated Eucalyptus wood and brewery's spent grain. Carbohydr. Polymer., 50, 191-200.
Karppinen, S., Kiiliäinen, K., Liukkonen, K., Forssell, P. and Poutanen, K. 2001. Extraction and in vitro fermentation of rye bran fractions. J. Cereal Sci. 34, 269-278.

104
Kellett L. E. 1990. Xylanase B and an arabinofuranosidase from Pseudomonas fluorescens subsp. cellulosa contain identical cellulose-binding domains and are encoded by adjacent genes. Biochem. J. 272, 369.
Khandke, K.M., Vithayathil, P.J. and Murthy, S.K. 1989. Degradation of larchwood xylan by enzymes of a thermophilic fungus, Thermoascus aurantiacus. Arch. Biochem. Biophys. 274, 501-510.
Koizumi, K., Kubota, Y., Tanimoto, T. and Okada, Y. 1991. High-performance anion-exchange chromatography of homogeneous D-gluco-oligosaccharides and -polysaccharides (polymerization degree � 50) with pulsed amperometric detection. J. Chromatogr. A 464, 365-373.
Kontula, P., Suihko, M.-L., Suortti, T., Tenkanen, M., Mattila-Sandholm, T. and von Wright, A. 2000. The isolation of lactic acid bacteria from human colonic biopsies after enrichment on lactose derivatives and rye arabinoxylo-oligosaccharides. Food Microbiol. 17, 13-22.
Kormelink, F.J.M., Hoffmann, R.A., Gruppen, H., Voragen, A.G.J., Kamerling, J.P. and Vliegenthart, J.F.G. 1993a. Characterisation by 1H NMR spectroscopy of oligosaccharides derived from alkali-extractable wheat-flour arabinoxylan by digestion with endo-(1�4)-beta-D-xylanase III from Aspergillus awamori. Carbohydr. Res. 249, 369-382.
Kormelink, F.J.M., Gruppen, H., Viëtor, R.J. and Voragen, A.G.J. 1993b. Mode of action of the xylan-degrading enzymes from Aspergillus awamori on alkali-extractable cereal arabinoxylans. Carbohydr. Res. 249, 355-367.
Kormelink, F.J.M., Gruppen, H. and Voragen, A.G.J. 1993c. Mode of action of (1�4)-�-D-arabinoxylan arabinofuranohydrolase (AXH) and �-L-arabinofuranosidases on alkali-extractable wheat-flour arabinoxylan. Carbohydr. Res. 249, 345-353.
Kormelink, F.J.M. and Voragen, A.G.J. 1993. Degradation of different [(glucurono)arabino]xylans by a combination of purified xylan-degrading enzymes. Appl. Microbiol. Biotechnol. 38, 688-695.
Kubata, B.K., Suzuki, T., Horitsu, H., Kawai, K. and Takamizawa, K. 1994. Purification and characterization of Aeromonas caviae ME-1 xylanase V, which produces exclusively xylobiose from xylan. Appl. Environ. Microbiol. 60, 531-535.
Kulkarni, N., Shendye, A. and Rao, M. 1999. Molecular and biotechnological aspects of xylanases. FEMS Microbiol. Rev. 23, 411-455.
Kusakabe, I., Ohgushi, S., Yasui, T. and Kobayashi, T. 1983. Studies on the xylanase system of Streptomyces. Part X. Structures of the arabinoxylo-oligosaccharides from the hydrolytic products of corncob arabinoxylan by a xylanase from Streptomyces. Agric. Biol. Chem. 47, 2713-2723.

105
Lebet, V., Arrigoni, E. and Amadò, R. 1997. Neutral-sugar determination by HPAEC-PAD to characterize polysaccharide degradation by colonic bacteria. Zeitschrift fuer Lebensmittel-Untersuchung und -Forschung A: Food Res. Technol. 205, 257-261.
Lee, Y.C. 1990. High-performance anion-exchange chromatography for carbohydrate analysis. Anal. Biochem. 189, 151-162.
Lee, Y.C. 1996. Carbohydrate analyses with high-performance anion-exchange chromatography. J. Chromatogr. A, 720, 137-149.
MacArthur, L.A. and D`Appolonia, B.L. 1980. Comparison of nonstarchy polysaccharides in oats and wheat. Cereal Chem. 57, 39-45.
Maes, C. and Delcour, J.A. 2002. Structural characterisation of water-extractable and water-unextractable arabinoxylans in wheat bran. J. Cereal Sci. 35, 315-326.
Maes, C., Vangeneugden, B. and Delcour, J.A. 2004. Relative activity of two endoxylanases towards water-unextractable arabinoxylans in wheat bran. J. Cereal Sci. 39, 181-186.
Margolles-Clark, E., Tenkanen, M., Luonteri, E. and Penttilä, M. 1996. Three �-galactosidase genes of Trichoderma reesei cloned by expression in yeast. Eur. J. Biochem. 240, 104-111.
Markwalder, H.U. and Neukom, H. 1976. Diferulic acid as possible crosslink in hemicelluloses from wheat germ. Phytochemistry, 15, 836-837.
Maslen, S.L., Goubet, F., Adam, A., Dupree, P. and Stephens, E. 2007. Structure elucidation of arabinoxylan isomers by normal phase HPLC–MALDI-TOF/TOF-MS/MS. Carbohydr. Res. 342, 724-735.
McNeil, M., Alberheim, P. Taiz, L. and Jones, R.L. 1975. Structure of plant cell walls. VII. Barley aleurone cells. Plant Physiol. 55, 64-68.
Meyer, V.R. 1995. Quantitation of chromatographic peaks in the 0.1 to 1.0 % range. Chromatographia, 40, 15-22.
Mitsuoka, T. 1982. Recent trends in research on intestinal flora. Bifidobacteria Microflora, 1, 3-24.
Modler, H.W. 1994. Bifidogenic Factors - Sources, Metabolism and Applications. Int. Dairy J. 4, 383-407.
Moura, P., Barata, R., Carvalheiro, F., Gírio, F., Loureiro-Dias, M.C. and Esteves, M.P. 2007. In vitro fermentation of xylo-oligosaccharides from corn cobs autohydrolysis by Bifidobacterium and Lactobacillus strains. LWT - Food Sci. Technol. 40, 963-972.
Moure, A., Gullon, P., Dominguez, H. and Parajo, J.C. 2006. Advances in the manufacture, purification and applications of xylo-oligosaccharides as food additives and nutraceuticals. Process Biochem. 41, 1913-1923.

106
Nandini, C.D. and Salimath, P.V. 2001. Carbohydrate composition of wheat, wheat bran, sorghum and bajra with good chapati/roti (Indian flat bread) making quality. Food Chem. 73, 197-203.
Nilsson, M., Andersson, R., Andersson, R.E., Autio, K. and Åman, P. 2000. Heterogeneity in a water-extractable rye arabinoxylan with a low degree of disubstitution. Carbohydr. Polym. 41, 397-405.
Okazaki, M., Fujukawa, S. and Matsumoto, N. 1990. Effects of xylooligosaccharide on growth of bifidobacteria. Nippon Eiyo, Shokuryo Gakkaishi 43, 395-401.
Ordaz-Ortiz, J.J., Guillon, F., Tranquet, O., Dervilly-Pinel, G., Tran, V. and Saulnier, L. 2004. Specificity of monoclonal antibodies generated against arabinoxylans of cereal grains. Carbohydr. Polym. 57, 425-433.
Ordaz-Ortiz, J.J., Devaux, M.F. and Saulnier, L. 2005. Classification of wheat varieties based on structural features of arabinoxylans as revealed by endoxylanase treatment of flour and grain. J. Agric. Food Chem. 53, 8349-8356.
Palframan, R., Gibson, G.R. and Rastall, R.A. 2003. Carbohydrate preferences of Bifidobacterium species isolated from the human gut. Curr. Iss. Intest. Microbiol. 4, 71-75.
Pascal, G. 2008. Prebiotics and food safety. In: Handbook of Prebiotics. Gibson, G.R. and Roberfroid, M.B. (Eds.), CRC Press, Taylor & Francis Group, Boca Raton, USA, pp. 449-470.
Pell, G., Taylor, E.J., Gloster ,T.M., Turkenburg, J.P., Fontes, C.M.G.A., Ferreira, L.M.A., Nagy, T., Clark, S.J., Davies, G.J. and Gilbert, H.J. 2004. The mechanisms by which family 10 glycoside hydrolases bind decorated substrates. J. Biol. Chem. 279, 9597-9605.
Pitkänen, L., Tuomainen, P., Virkki, L., Aseyev, V. and Tenkanen, M. 2008. Structural comparison of arabinoxylans from two barley side-stream fractions. J. Agric. Food Chem. 56, 5069-5077.
Pitkänen, L., Virkki, L., Tenkanen, M. and Tuomainen, P. 2009. Comprehensive multidetector HPSEC study on solution properties of cereal arabinoxylans in aqueous and DMSO solutions. Biomacromolecules 10, 1962-1969.
Poutanen, K., Sundberg, M., Korte, H. and Puls, J. 1990. Deacetylation of xylans by acetyl esterases of Trichoderma reesei. Appl. Microbiol. Biotechnol. 33, 506-510.
Puchart, V. and Biely P. 2008. Simultaneous production of endo-�-1,4-xylanase and branched xylooligosaccharides by Thermomyces lanuginosus. J. Biotechnol. 137, 34-43.
Puls, J., Schmidt, O. and Granzow, C. 1987. �-Glucuronidase in two microbial xylanolytic systems. Enzyme Microb. Technol. 9, 83-88.
Puls, J. 1993. Substrate analysis of forest and agricultural wastes. In: Bioconversion of forest and agricultural plant residues. Saddler, J.N. (Eds.), CAB International, Wallingford, UK, pp. 13-32.

107
Puls, J., Schroeder, N., Stein, A., Janzon, R. and Saake, B. 2006. Xylans from oat spelts and birch kraft pulp. Macromol. Symp. 232, 85-92.
Puls, J. and Schuseil, J. 1993. Chemistry of hemicelluloses: relationship between hemicellulose structure and enzymes required for hydrolysis. In: Hemicellulose and hemicellulases. Coughlan, M.P. and Hazlewood, G.P. (Eds.), Portland Press, London, U.K. pp. 1-28.
Ramirez, F., Puls, J., Zúñiga, V. and Saake, B. 2008. Sorption of corn cob and oat spelt arabinoxylan onto softwood kraft pulp. Holzforschung 62, 329-339.
Roberts, E.J., Wade, C.P. and Rowland, S.P. 1971. Effect of base concentration upon the reactivities of the hydroxyl groups in methyl d-glucopyranosides. Carbohydr. Res. 17, 393-399.
Roubroeks, J.P., Andersson, R. and Åman, P. 2000. Structural features of (1�3),(1�4)-�-D-glucan and arabinoxylan fractions isolated from rye bran. Carbohydr. Polym. 42, 3-11.
Roy, D. and Ward, P. 1990. Evaluation of rapid methods for differentiation of Bifidobacterium species. J. Appl. Bacteriol. 69, 739-749.
Roy, D. and Sirois, S. 2000. Molecular differentiation of Bifidobacterium species with amplified ribosomal DNA restriction analysis and alignment of short regions of the ldh gene. FEMS Microbiol. Lett. 191, 17-24.
Salminen, S., Bouley, C.H., Boutro-Ruault, M.C., Cummings, J.H., Franck, A., Gibson, G.R., Isolauri, E., Moreau, M.C., Roberfroid, M.B. and Rowland, I. 1998. Functional food science and gastrointestinal physiology and function. Br. J. Nutr. 80, S147-S171.
Saulnier, L., Marot, C., Chanliaud, E. and Thibault, J. 1995a. Cell wall polysaccharide interactions in maize bran. Carbohydr. Polym. 26, 279-287.
Saulnier, L., Vigoroux, J. and Thibault, J.-F. 1995b. Isolation and partial characterization of feruloylated oligosaccharides from maize bran. Carbohydr. Res. 272, 241-253.
Scharlau, D., Borowicki, A., Habermann, N., Hofmann, T., Klenow, S., Miene, C., Munjal, U., Stein, K. and Glei, M. 2009. Mechanisms of primary cancer prevention by butyrate and other products formed during gut flora-mediated fermentation of dietary fibre. Mutat. Res./Rev. Mutat. Res. 682, 39-53.
Schell, M.A., Karmirantzou, M., Snel, B., Vilanova, D., Berger, B., Pessi, G., Zwahlen, M., Desiere, F., Bork, P., Delley, M., Pridmore, R.D. and Arigoni, F. 2002. The genome sequence of Bifidobacterium longum reflects its adaptation to the human gastrointestinal tract. Proc. Natl. Acad. Sci. U. S. A. 99, 14422-14427.
Scheppach, W., Bartram H.P. and Richter F. 1995. Role of short-chain fatty acids in the prevention of colorectal cancer. Eur. J. Cancer 31, 1077-1080.
Scheppach, W., Luehrs, H. and Menzel, T. 2001. Beneficial health effects of low-digestible carbohydrate consumption. Br. J. Nutr. 85, S23-S30.

108
Schooneveld-Bergmans, M.E.F., Beldman, G. and Voragen, A.G.J. 1999. Structural features of (glucurono)arabinoxylans extracted from wheat bran by barium hydroxide. J. Cereal Sci. 29, 63-75.
Shibuya, N., Misaki, A. and Iwasaki, T. 1983. The structure of arabinoxylan and arabinoglucuronoxylan isolated from rice endosperm cell wall. Agric. Biol. Chem. 47, 2223-2230.
Shibuya, N. and Misaki, A. 1978. Structure of hemicellulose isolated from rice endosperm cell wall: Mode of linkages and sequences in xyloglucan, �-glucan and arabinoxylan. Agric. Biol. Chem. 42, 2267-2274.
Shibuya, N., Nakane, R., Yasui, A., Tanaka, K. and Iwasaki, T. 1985. Comparative studies on cell wall preparations from rice bran, germ, and endosperm. Cereal Chem. 62, 252-258.
Shiiba, K., Yamada, H., Hara, H., Okada, K. and Nagao, S. 1993. Purification and characterization of two arabinoxylans from wheat bran. Cereal Chem. 70, 209-214.
Shin, H., Lee, J., Lee, J., Han, Y., Han,, M.J. and Kim, D. 2003a. Purification and characterization of ginsenoside Ra-hydrolyzing �-D-xylosidase from Bifidobacterium breve K-110, a human intestinal anaerobic bacterium. Biol. Pharm. Bull. 26, 1170-1173.
Shin, H., Park, S., Sung, J.H. and Kim, D. 2003b. Purification and characterization of �-L-arabinopyranosidase and �-L-arabinofuranosidase from Bifidobacterium breve K-110, a human intestinal anaerobic bacterium metabolizing ginsenoside Rb2 and Rc. Appl. Environ. Microbiol. 69, 7116-7123.
Sjöström, E. 1981. Wood chemistry. Fundamentals and applications. Academic Press, Inc., New York,
Smith, M.M. and Hartley, R.D. 1983. Occurrence and nature of ferulic acid substitution of cell-wall polysaccharides in graminaceous plants. Carbohydr. Res. 118, 65-80.
Snyder, L.R. 1972. Rapid approach to selecting the best experimental conditions for high-speed liquid chromatography. 1. Estimating initial sample resolution and the final resolution required by a given problem. J. Chromatogr. Sci. 10, 200-212.
Sørensen, H.R., Jørgensen, C.T., Hansen, C.H., Jørgensen, C.I., Pedersen, S. and Meyer, A.S. 2006. A novel GH43 �-L- arabinofuranosidase from Humicola insolens: mode of action and synergy with GH51 �-L- arabinofuranosidases on wheat arabinoxylan, Appl. Microbiol. Biotechnol. 73, 850-861.
Sternemalm, E., Höije, A. and Gatenholm, P. 2008. Effect of arabinose substitution on the material properties of arabinoxylan films. Carbohydr. Res. 343, 753-757.
Sun, R., Lawther, J.M. and Banks, W.B. 1996. Fractional and structural characterization of wheat straw hemicelluloses. Carbohydr. Polym. 29, 325-331.

109
Sun, H., Yoshida, S., Park, N. and Kusakabe, I. 2002. Preparation of (1�4)-�-xylooligosaccharides from an acid hydrolysate of cotton-seed xylan: suitability of cotton-seed xylan as a starting material for the preparation of (1�4)-�-xylooligosaccharides. Carbohydr. Res. 337, 657-661.
Sundberg, A., Sundberg. K., Lillandt. C. and Holmbom. B. 1996. Determination of hemicelluloses and pectins in wood and pulp fibers by acid methanolysis and gas chromatography. Nord. Pulp Pap. Res. J. 11, 216-219, 226.
Suwa, Y., Koga, K., Fujikawa, S., Okazaki, M., Irie, T. and Nakada, T. 1999 Bifidobacterium bifidum proliferation promotion containing xylooligosaccharide, U.S. Patent 5939309.
Swennen, K., Courtin, C.M., Lindemans, G.C. and Delcour, J.A. 2006. Large-scale production and characterisation of wheat bran arabinoxylooligosaccharides. J. Sci. Food Agric. 86, 1722-1731.
Tenkanen, M. 2004. Enzymatic tailoring of hemicelluloses. ACS Symp. Ser. 864, 292-311.
Tenkanen, M., Luonteri, E. and Teleman, A. 1996. Effect of side groups on the action of �-xylosidase from Trichoderma reesei against substituted xylo-oligosaccharides. FEBS Lett. 399, 303-306.
Tenkanen, M. and Siika-aho, M. 2000. An �-glucuronidase of Schizophyllum commune acting on polymeric xylan. J. Biotechnol. 78, 149-161.
Tenkanen, M., Bürgermeister, M., Vršanská, M., Biely, P., Saloheimo, M. and Siika-aho, M. 2003. A novel xylanase XYN IV from Trichoderma reesei and its action on different xylans. In: Recent advances in enzymes in grain processing. Courtin, C., Veraverbeke, W. and Delcour, J. (Eds.). ACCO, Leuven, Belgium, pp. 41-46.
Thayer, J.R., Rohrer, J.S., Avdalovic, N. and Gearing, R.P. 1998. Improvements to in-line desalting of oligosaccharides separated by high-pH anion exchange chromatography with pulsed amperometric detection. Anal. Biochem. 256, 207-216.
Trogh, I., Croes, E., Courtin, C.M. and Delcour, J.A. 2005. Enzymic degradability of hull-less barley flour alkali-solubilized arabinoxylan fractions by endoxylanases. J. Agric. Food Chem. 53, 7243-7250.
Van Craeyveld, V., Swennen, K., Dornez, E., Van de Wiele, T., Marzorati, M., Vestraete, W., Delaedt, Y., Onagbesan, O., Decuypere, E., Buyse, J., De Ketelaere, B., Broekaert, W.F., Delcour, J.A. and Courtin, C.M. 2008. Structurally different wheat-derived arabinoxylooligosaccharides have different prebiotic and fermentation properties in rats. J. Nutr. 138, 2348-2355.
Van Craeyveld, V., Holopainen, U., Selinheimo, E., Poutanen, K., Delcour, J.A. and Courtin, C.M. 2009. Extensive dry ball milling of wheat and rye bran leads to in situ production of arabinoxylan oligosaccharides through nanoscale fragmentation. J. Agric. Food Chem. 57, 8467-8473.

110
Van den Broek, L.A.M., Lloyd, R.M., Beldman, G., Verdoes, J.C., McCleary, B.V. and Voragen, A.G.J. 2005. Cloning and characterization of arabinoxylan arabinofuranohydrolase-D3 (AXHd3) from Bifidobacterium adolescentis DSM20083. Appl. Microbiol. Biotechnol. 67, 641-647.
Van den Broek, L.A.M., Hinz, S.W.A., Beldman, G., Vincken, J.-P. and Voragen, A.G.J. 2008. Bifidobacterium carbohydrases-their role in breakdown and synthesis of (potential) prebiotics. Mol. Nutr. Food Res. 52, 146-163.
Van den Broek, L.A.M. and Voragen, A.G.J. 2008. Bifidobacterium glycoside hydrolases and (potential) prebiotics. Innov. Food Sci. Emerg. Technol. 9, 401-407.
Van Laere, K.M.J., Beldman, G. and Voragen, A.G.J. 1997. A new arabinofuranohydrolase from Bifidobacterium adolescentis able to remove arabinosyl residues from double-substituted xylose units in arabinoxylan. Appl. Microbiol. Biotechnol. 47, 231-235.
Van Laere, K.M.J., Voragen, C.H.L., Kroef, T., Van den Broek, L.A.M., Beldman, G. and Voragen, A.G.J. 1999. Purification and mode of action of two different arabinoxylan arabinofuranohydrolases from Bifidobacterium adolescentis DSM 20083. Appl. Microbiol. Biotechnol. 51, 606-613.
Van Laere, K.M.J., Hartemink, R., Bosveld, M., Schols, H.A. and Voragen, A.G.J. 2000. Fermentation of plant cell wall derived polysaccharides and their corresponding oligosaccharides by intestinal bacteria. J. Agric. Food Chem. 48, 1644-1652.
Van Loo, J., Cummings, J., Delzenne, N., Englyst, H., Franck, A., Hopkins, M., Kok, N., Macfarlane, G., Newton, D., Quigley, M., Roberfroid, M., van Vliet, T. and van den Heuvel, E. 1999. Functional food properties of non-digestible oligosaccharides: a consensus report from the ENDO project (DGXII AIRII-CT94-1095). Br. J. Nutr. 81, 121-132.
Vardakou, M., Katapodis, P., Samiotaki, M., Kekos, D., Panayotou, G. and Christakopoulos, P. 2003. Mode of action of family 10 and 11 endoxylanases on water-unextractable arabinoxylan. Int. J. Biol. Macromol. 33, 129-134.
Vardakou, M., Palop C.N., Gasson, M., Narbad, A. and Christakopoulos, P. 2007. In vitro three-stage continuous fermentation of wheat arabinoxylan fractions and induction of hydrolase activity by the gut microflora. Int. J. Biol. Macromol. 41, 584-589.
Vardakou, M., Palop, C.N., Christakopoulos, P., Faulds, C.B., Gasson, M.A. and Narbad, A. 2008. Evaluation of the prebiotic properties of wheat arabinoxylan fractions and induction of hydrolase activity in gut microflora. Int. J. Food Microbiol. 123, 166-170.
Vázquez, M.J., Alonso, J.L., Domínguez, H. and Parajó, J.C. 2000. Xylooligosaccharides. Manufacture and applications. Trends Food Sci. Technol. 11, 387-393.
Verwimp, T., Van Craeyveld, V., Courtin, C.M. and Delcour, J.A. 2007. Variability in the structure of rye flour alkali-extractable arabinoxylans. J. Agric. Food Chem. 55, 1985-1992.

111
Viëtor, R.J., Angelino, S.A.G.F. and Voragen, A.G.J. 1992. Structural features of arabinoxylans from barley and malt cell wall material. J. Cereal Sci. 15, 213-222.
Viëtor, R.J., Hoffmann, R.A., Angelino, S., A.G.F., Voragen, A.G.J., Kamerling, J.P. and Vliegenthart, J.F.G. 1994. Structures of small oligomers liberated from barley arabinoxylans by endoxylanase from Aspergillus awamori. Carbohydr. Res. 254, 245-255.
Vinkx, C.J.A., Delcour, J.A., Verbruggen, M.A. and Gruppen, H. 1995. Rye water-soluble arabinoxylans also vary in their 2-monosubstituted xylose content. Cereal Chem. 72, 227-228.
Virkki, L., Johansson, L., Ylinen, M., Maunu, S. and Ekholm, P. 2005. Structural characterization of water-insoluble nonstarchy polysaccharides of oats and barley. Carbohydr. Polym. 59, 357-366.
Virkki, L., Maina, H.N., Johansson, L. and Tenkanen, M. 2008. New enzyme-based method for analysis of water-soluble wheat arabinoxylans. Carbohydr. Res. 343, 521-529.
Visek, W. 1978. Diet and cell growth modulation by ammonia. Am. J. Clin. Nutr. 31, 216S-220.
Voragen, A.G.J. 1998. Technological aspects of functional food-related carbohydrates. Trends Food Sci. Technol. 9, 328-335.
Weisburg, W.G., Barns, S.B., Pelletier, D.A. and Lane, D.J. 1991. 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 173, 697-703.
Wende, G. and Fry, S.C. 1997. 2-O-�-D-xylopyranosyl-(5-O-feruloyl)-L-arabinose, a widespread component of grass cell walls. Phytochem. 44, 1019-1030.
Westerlund, E., Andersson, R. and Åman, P. 1993. Isolation and chemical characterization of water-soluble mixed-linked �-glucans and arabinoxylans in oat milling fractions. Carbohydr. Polym. 20, 115-123.
Whistler, R.L. and McGilvray, D.I. 1955. 2-O-�-D-Xylopyranosyl-L-arabinose from hemicellulose-B of corn cob1,2. J. Am. Chem. Soc. 77, 1884-1885.
Wilkie, K.C.B. 1979. The hemicelluloses of grasses and cereals. Adv. Carbohydr. Chem. Biochem. 36, 215-264.
Wolin, E.A., Wolin, M.J. and Wolfe, R.S. 1963. Formation of methane by bacterial extracts. J. Biol. Chem. 238, 2882-2886.
Wollowski, I., Rechkemmer G. and Pool-Zobel, B.L. 2001. Protective role of probiotics and prebiotics in colon cancer. Am. J. Clin. Nutr. 73, 451S-455.
Wong, J.M.W.R.D., de Souza, R.R.D., Kendall, C.W.C., Emam, A. and Jenkins, D.J.A. 2006. Colonic health: Fermentation and short chain fatty acids. J. Clin. Gastroenterol. 40, 235-243.

112
Yuan, X., Wang, J. and Yao, H. 2005. Feruloyl oligosaccharides stimulate the growth of Bifidobacterium bifidum. Anaerobe 11, 225-229.
Copyright © 2022 FDOKUMEN




















