
The diversity of harmful algal blooms: a challenge for science and management
Upload
independentCategory
view
1download
0
Date: Tue, 30 May 2000 14:32:15 -0400From: OAO [email protected]>To: [email protected]: WWW ILL Request [email protected] (OAO) sent the following ILL request:____------------------------------------------------------------Author: Bode, A., M. Varela, et alBook Title: Journal of the Marine Biological Association of the United KingdomJournal Article: "Preliminary studies on the export of organic matter duringphytoplankton blooms off La Coruna (north-western Spain)"Year: 1998Volume: 78(l)Pages: l-15.114Location:Rush: onNeeded By:User Name: OAOE-Mail address: [email protected] #: 30-05-2000____---------------------------------------------------- -___----
Server Protocol: HTTP/1.1Remote Host: (200.10.149.11)

1. Mar. Rio!. Ass. U.K. (1998), 78, l-15 1t Printed in Great Britain
PRELIMINARY STUDIES ON THE EXPORT OF ORGANIC MATTERDURING PHYTOPLANKTON BLOOMS OFF LA CORUfiA (NORTH-
WESTERN SPAIN)
ANTONIO BODE, MANUEL VARELA, SUSANA BARQUERO, Ma TERESAALVAREZ-OSSORIO AND NICOLAS GONZALEZ
Institute Esptiol de Oceanograia, Centro Costero de La Coruiia, Apartado 130, E-15080 La Corufia, Spain
Sinking of particles was measured at a coastal site near La Coruna (north-western Spain)using sedimentation traps placed at the base of the euphotic zone during four shortperiods between March and June 1995. In addition, measurements of water columnproperties and particles were made at five stations. The cases studied were representativeof the main oceanographic situations that occur in this area and were related to the wind-induced upwelling. Phytoplankton blooms ( > 100 mg chlorophyll-a m-‘) were observedon two occasions in April and June, both during favourable upwelling conditions. The twoother cases were studied during low phytoplankton biomass concentrations in the watercolumn.
The mean values of sinking carbon varied between 1203 and 323 mg C me2 d-‘. Theminimum values were measured during the upwelling case in June, and may beexplained in part by the offshore displacement of surface water caused by the upwelling.The contribution of phytoplankton to the exported carbon was low (1045% of thetotal particulate carbon), and only a small fraction corresponded to Livingphytoplankton (24%). Most of the particles collected in the traps appeared to be ofpelagic origin, mainly because low C:N ratios, dominance of phytoplanktonic overphytobenthic species and the abundance of zooplankton faecal pellets. The trap-collectedmaterial displayed a sample of the phytoplankton species present, but some speciesappeared with higher frequencies than expected from their abundances in the watercolumn. In addition, the sedimented material showed a high abundance of faecal pellets.The abundance of the smaller faecal pellets’( < 200 pm) decreased during the study, whilethe larger pellets ( > 200 pm) were more abundant in May, matching the variation in theabundance of herbivorous and omnivorous copepods in the water column.
INTRODUCTION
The sinking of organic materials is the link between pelagic productivity and thefood supply to the benthos, but it also represents a sink for the excess of carbonaccumulated at the surface of the oceans. The recognition of this key process producedan increase of biogeochemical studies in recent years (Berger et al., 1989). In manycases, the sinking material is collected using sediment traps. These traps have beendeployed in a large variety of depths, areas and oceanic zones. In general, the export ofmaterials from the euphotic zone from oceanic areas with marked seasonality orfrequent inputs of new nutrients are characterized by pulses, following the signal ofthe primary production in the water coluinn In boreal and temperate areas, the main

2 ANTONIO BODE ET AL.
I
productivity and export pulse occurs at the time of the spring blooms (Smetacek, 1980;Wassmann, 1991; Asper et al., 1992; Bender et al., 1992; Olesen & Lundgaard, 1995).Similar coupling between pelagic productivity pulses and vertical export of biogenicmaterials have been reported in coastal upwelling areas (Wefer & Fisher, 1993).
The upwelling system of north-western Spain is characterized by the episodicenrichment of nutrients into the coastal zone and by its accompanying elevated rates ofprimary productivity (Femandez & Bode, 1991; Bode et al., 1994a; Tenore et al., 1995).The high primary productivity of this region allows for important pelagic fisheries onthe shelf (Tenore et al., 1995) and shellfish farming in the estuarine rias (Blanton et al.,1987). At the same time, it is known that large accumulations of organic matteroccurred in shallow areas of the continental shelf, particularly along the western coastnear the more productive rias (Lopez-Jamar et al., 1992). These accumulations ofsediment can be related to the organic matter exported from the rias, where primaryproduction is enhanced by the upwelling (Fraga, 1976; Tenore et al., 1982).
The export of particles from the euphotic zone was measured in the coast of Asturias(southern Bay of Biscay), in the eastern limit of this upwelling region, during no-upwelling conditions in summer 1993 (Fernandez et al., 1995). The results indicate thata large proportion of material collected in traps placed at the base of the euphotic zonewas of phytoplanktonic origin, and therefore a close coupling between phytoplanktonproduction pulses at the surface and vertical export was inferred. However, there are nocomparable results for the Galician coast, in the north-western coast of Spain, where theupwelling lasts for a long time and reaches higher intensity (Fraga, 1981; Botas et al.,1990). Ln this paper we report the flux of particulate material collected with sedimenttraps deployed during short time-scales at a coastal station off La Coruiia (north-western Spain), between March and June 1995. Complementary and descriptive aspectsof the physical, chemical and biological oceanography of this area can be found inValdes et al. (1991); Bode et al. (1994a) and Casas et al. (1997). These results are the firstpublished for this highly productive continental shelf region and allow for a firstanalysis of the relationships between the biomass and composition of plankton andvertical export of particles during caSe events in the season of high productivity.
MATERIALS AND METHODS
Water column observations were made at five stations off La Coruria (Figure 1) on15 March, 11 April, 16 May and 13 June 1995. Temperature, salinity, photosynthe-tically available radiation (PAR), and in situ fluorescence were measured with aconductivity, temperature and depth probe (CID) Seabird SBE-25, equipped with alight PAR sensor and an in viva fluorometer. Water dynamics were inferred from dailyaveraged values of the upwelling index (Bakun, 1973), computed from local windsmeasured every six hours at the Centro Meteorologico Zonal of La Coruria (InstituteEspaiiol de Meteorologia). The method of computation of this index is described inLavin et al. (1991). This index estimates the Ekman offshore transport of surface waterin a transect parallel to the shoreline. Surface water can be transported offshoreduring upwelling events (positive values of the index) or accumulated towards thecoast when downwelling conditions prevail (negative values of the index).

ORGANIC EXPORT OFF LA CORmA 3
/1OOm
f
8” 27’ 8” 24’ W
43” 28’
43’ 24’
43O 20’
Figure 1. Map of the studied stations. The position of the trap was marked as station T.
Bottle casts were made at station 2 to obtain water samples for chlorophyll-a andphaeopigments, phytoplankton counts, particulate carbon and nitrogen, and primaryproductivity measurements at the depths of 100,50,25,10 and 1% of PAR received atthe surface. An extra bottle sampled water at 70 m depth. Chlorophyll-u (Chl-a) andphaeopigment (Phaeo) concentrations were determined by fluorometry using acetonicextracts and Whatman GF/F filters (UNESCO, 1994). Aliquots for the determinationof total carbon and nitrogen content were filtered through Whatman GF/F filters andanalysed using a Perkin Elmer CHN analyser. Primary production was estimated bythe C-14 method, using incubations of 2 h in simulated in situ conditions as describedin Bode et al. (1994a). Incubations were terminated by filtration of samples throughWhatman GF/F filters. Daily primary production rates were calculated by multi-plying hourly rates by the total number of sun hours during the day ( * 8 h in Marchand April, and 10 h in May and June). Samples for microscopic examination werepreserved in Lugol’s solution and examined with an inverted microscope forrecognizable phytoplankton cells.
Zooplankton was sampled at station 2 from the surface to + 70 m depth usingdouble oblique hauls with a WP~ plankton net (25~pm mesh size). Zooplanktonabundance was measured in samples preserved with 4% buffered formalin asdescribed in Valdes et al. (1991). Zooplankton biomass was estimated firstly as dryweight (DWz) of non-preserved aliquots filtered through pre-weighed Whatman GF/C

4 ANTONIO BODE ET AL.
filters, dried at 60°C for 48 hours. Dry samples were cornbusted for 1 h at 5OO’C andreweighed for determination of the ash-free dry weight. Dry weight was converted tocarbon (Czl) using the formula of Wiebe (1988):
log (DWz)=0.499 + 0.991 log (Czl) (1)
Other aliquots of dry zooplankton were analysed for total carbon (Cz2) and nitrogen(Nz) in the CHN analyser. In addition, a multitrap collector system was deployed at50 m and anchored to the sea-floor at station T (total depth 63 m) for periods of two orthree days including the dates when the entire transect of stations was sampled. Forlogistic reasons, the trap had to be located at 1 nautical mile off station 2. The trapsystem was described in Knauer et al. (1979) and consisted of four transparentPlexiglass collection cylinders of 6 cm diameter each. Prior to the deployment, thecylinders were filled with a saline solution (sea-water filtered through Whatman GF/F filters to which 35 g of NaCl were added per litre of solution) to prevent the escapeof particles from the traps. No preservatives were used in the traps.
Immediately after recovery, aliquots of the material collected by the traps werefiltered through Whatman GF/F filters for the determination of total carbon andnitrogen, chlorophvll-a and phaeopigments as described above. Phytoplankton-derived carbon was calculated as in Femandez et al. (1995). First, chlorophyll-equivalent concentrations (Cl-&a,+,) were computed as:
Chl-a~~i,=Chl-a+I.51 Phaeo (2)
where l-51 is the ratio of molar weights of Chl-a and Phaeo. Total phytoplankton carbon(C-Chl) was estimated from the product of Chla,gti, by the ratio 50 g C (g CM-u)-‘.Similarly, carbon from living or non-degraded phytoplankton was estimated usingChl-u and the above ratio.
Samples of trap-collected material for microscopic examination were preserved ineither Lugol’s solution or 5?0 glutaraldehyde. Lugol’s preserved samples wereexamined with an inverted microscope for recognizable phytoplankton cells andzooplanktonic faecal pellets. Pellets were classified in three size groups: pellets< 50 pm; pellets between 50 and 200 pm; and pellets >200 pm. The samplespreserved with glutaraldehyde were stored for 24 h, stained with DAPI (2-diamidinophenyl-indol), and filtered through black-stained polycarbonate filters of 0.2 pm pore-size. Filters were stored frozen until observation of bacteria with an epifluorescencemicroscope (Bode et al., 1994b). The frequency of dividing bacteria was recorded as anindicator of bacterial productivity (Newell & Christian, 1981).
RESULTS
Water column conditions
The direction of prevailing winds changes seasonally in the study area (Figure 2).Winds blowing from the east and north-east are favourable to the upwelling andbecome increasingly dominant during spring and early summer. Downwellingconditions prevail during winter. The studied events are illustrative of the different

4000
2000
EuE.P‘S 0
4
-2000
4ooo
ORGANIC EXPORT OFF LA CORmA
13-June
5
0 3 0 60 9 0 1 2 0 150 180
Julian day (1995)
Figure 2. Variation of the daily upwelling index (m 3 s -I km-‘) during the studv. The sampling dateswere marked as circles.
situations that occurred during the spring and early summer. The situation of 15March 1995 was a moderate upwelling pulse, sampled a few days after the maximumintensity peak. T’he case of 11 April corresponded to a relatively calm situation afterseveral weeks of weak upwelling. A very different situation was found on 16 Maywhere downwelling conditions followed several short upwelling-favourable periods.The samples of 13 June were collected during upwelling conditions, but in this casethere was a period of approximately two weeks of relatively strong upwelling-favourable winds before sampling.
Stratification of the surface upper layers was the main characteristic of the studiedoffshore transect on 15 March (Figure 3). This stratification was caused mainly bywater of continental origin near the coast, with low salinity values (not shown), whilewater below 20 m had density values typical of shelf waters in this area (Rios et al.,1992). The phytoplankton biomass, indicated by in vivo fluorescence, was low ingeneral, except for a small increase at the surface of station 2. However, thechlorophyll-a concentration at this station reached the minimum value recordedduring this study (Table 1). The surface stratification on 11 April was almost reducedto the coastal station and shelf waters predominated in the water column along thetransect (Figure 3). By comparison with the situation on 15 March, water with thesame st value was l&ated closer to the surface on 11 April (Figure 3), indicating arecent upwelling. In addition, phytoplankton biomass was high in the area,Particularly near station 2 and the location of the trap.
Low densitv waters extended well below the surface and the phytoplanktonbiomass was higher near the coast on 16 May (Figure 4). The pattern of the lines ofequal density and in vivo fluorescence suggest sinking of surface waters near the

i
6 ANTONIO BODE ET AL.
Table 1. Integrated values of chlorophyll-a (Chl-a), phaeopigments (Phaeo), chlorophyll-aequivalents (Chl-a,q,iz, 1, particulate carbon CC) and nitrogen (N), C:N molar ratio, andprimary production (PP) in the upper 50 m of the water column at station 2. Water columnintegrated values of dry weight (DWz) and ash-free dry weight of mesozooplankton (AFDWz),estimates of mesozooplankton carbon (Czl) as in Wiebe (1988), and mean f&SD) directmeasures of mesozooplankton carbon (Cz2) and nitrogen (Nz) at station 2.
Chl-lJ (mg me21 1640 10383 3263P h a e o (mg m-‘) 10.10 76 .53 2588Chk?qUi” (mg m-*1 3165 219 .39 71.71C 6-q me*) 1293.00 4353.00 3254.75N (mg me2) 175.75 640.50 495.75C : N by atoms 7.36 6.80 657PP (mg me2 d“) 99940 864268 919.50DWz (mg me*) 300 .30 1142 .96 4134 .16AFDWz (mg me21 246 .40 868 .40 3551 .36Czl (mg me*) 95-45 368-71 1347.16Cz2 ( + SD) (mg mw2) 12460 +OdO 450,32 + 2.94 1796.76 + 18.41Nz (&SD) (mg m-‘) 20-30 1oGo 69.94 r 1.10 292.02 2 6.51
15 March 11 April 1 6 M a y 13 June
A2 30 3c 3t1 4
i
7 1Mdafcb1995 (
3
840-
‘ i n vivo’ fluofmnce1 SMarch- 1995
196.1630.78
2426461894094640
6547369.85
875 .03640 .24281 .55
-
i / 1 l-April-1895
Figure 3. (A&C) Distribution of water density (St); (B&D) in vivo fluorescence along the studiedtransect on 15 March and 11 April 1995.
location of the trap, between stations 3a and 2. Sinking of surface waters was expectedin the measured downwelling conditions (Figure 2). In contrast, upwelling of shelfwaters on 13 June was indicated by the ascent of the isopycnal line of 27.0 unitstowards the coast (Figure 4). There was a stratification of the upper layer caused
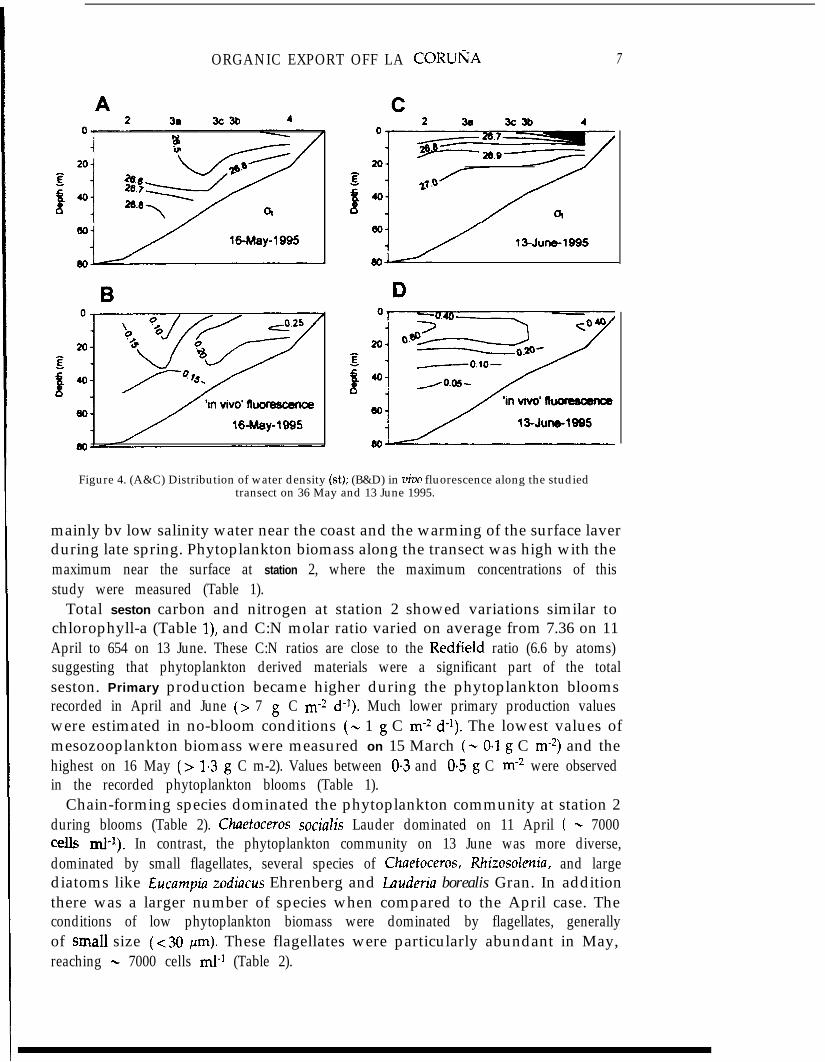
ORGANIC EXPORT OFF LA CORUI;JA 7
“1 / 1%June1995
Figure 4. (A&C) Distribution of water density (St); (B&D) in viva fluorescence along the studiedtransect on 36 May and 13 June 1995.
mainly bv low salinity water near the coast and the warming of the surface laverduring late spring. Phytoplankton biomass along the transect was high with themaximum near the surface at station 2, where the maximum concentrations of thisstudy were measured (Table 1).
Total seston carbon and nitrogen at station 2 showed variations similar tochlorophyll-a (Table l), and C:N molar ratio varied on average from 7.36 on 11April to 654 on 13 June. These C:N ratios are close to the Redfield ratio (6.6 by atoms)suggesting that phytoplankton derived materials were a significant part of the totalseston. Primary production became higher during the phytoplankton bloomsrecorded in April and June (> 7 g C m-2 d-r). Much lower primary production valueswere estimated in no-bloom conditions (- 1 g C m-2 d-i). The lowest values ofmesozooplankton biomass were measured on 15 March (h 0.1 g C m-‘) and thehighest on 16 May (> 1.3 g C m-2). Values between 0.3 and O-5 g C m-2 were observedin the recorded phytoplankton blooms (Table 1).
Chain-forming species dominated the phytoplankton community at station 2during blooms (Table 2). Chaetoceros socialis Lauder dominated on 11 April ( z 7000cells ml-l). In contrast, the phytoplankton community on 13 June was more diverse,dominated by small flagellates, several species of Chaetocerus, Rhizosolerzia, and largediatoms like Eucumpia zodiacus Ehrenberg and Lauderia borealis Gran. In additionthere was a larger number of species when compared to the April case. Theconditions of low phytoplankton biomass were dominated by flagellates, generallyof small size (~30 pm). These flagellates were particularly abundant in May,reaching h 7000 cells ml-i (Table 2).

8 ANTONIO BODE ET AL.
Table 2. Mean abundance C + SE cells ml-‘) of the main phytoplankton taxa at station 2. Onlygroups with >40 cells ml-’ are listed.
DinophyceaeCachonina hallii
Freudentahl & LeeSmall dinoflagellates ( ~30 pm)
DiatomophyceaeSmall centric diatoms ( < 30 pm)Chaetoceros afinis
Lauder+uar willei GranChaetoceros compressus LauderChaetoceros gracilis SchiittChaetoceros pelagicus CleveChaetoceros socialis LauderEucampia zodiacus Ehrenbergkzuderia borealis GranRhizosolenia fragilissima BergonR. set&era Brightwell
PrymnesiophyceaePhaeocystis poucheti
(Hariot) LagerheimCryptophyceaeFlagellates
15 March 11 April
177.9 *1344 -
373.1 f 307.7 468 57.7
- -
- 77.7 k32.1- --- 6877.8 + 2669.2- _I --
--
-
606.9 & 355.3
10234 2 178.5 2079.2 5 422.2Small flagellates (3-5 pm)Medium-sized flagellates (5-8 pm) 3440 573.2 240.1 f Y3.5Large flagellates ( > 8 pm) 228.9 + 1884 -
16 May 13 June
1856 i98-l 41-6 * 19.8
768.6 5 301.2 63.0 ill.7
- 79.2 +9.2- 69.2 & 17.3
- -- 117.0 * 754- 56.0 to.0- 209.5 k 173.1
56,O +O.O- 60.0 * 16.6- 1826-l _+8834- 12543 + 39.1
404 5 145
901.6 2 4615 504 k7.3
4669.8 = 2148 23956 & 373.710664 i_ 114.1 2147 2 15.211844 e 306.5 684 * 15.7
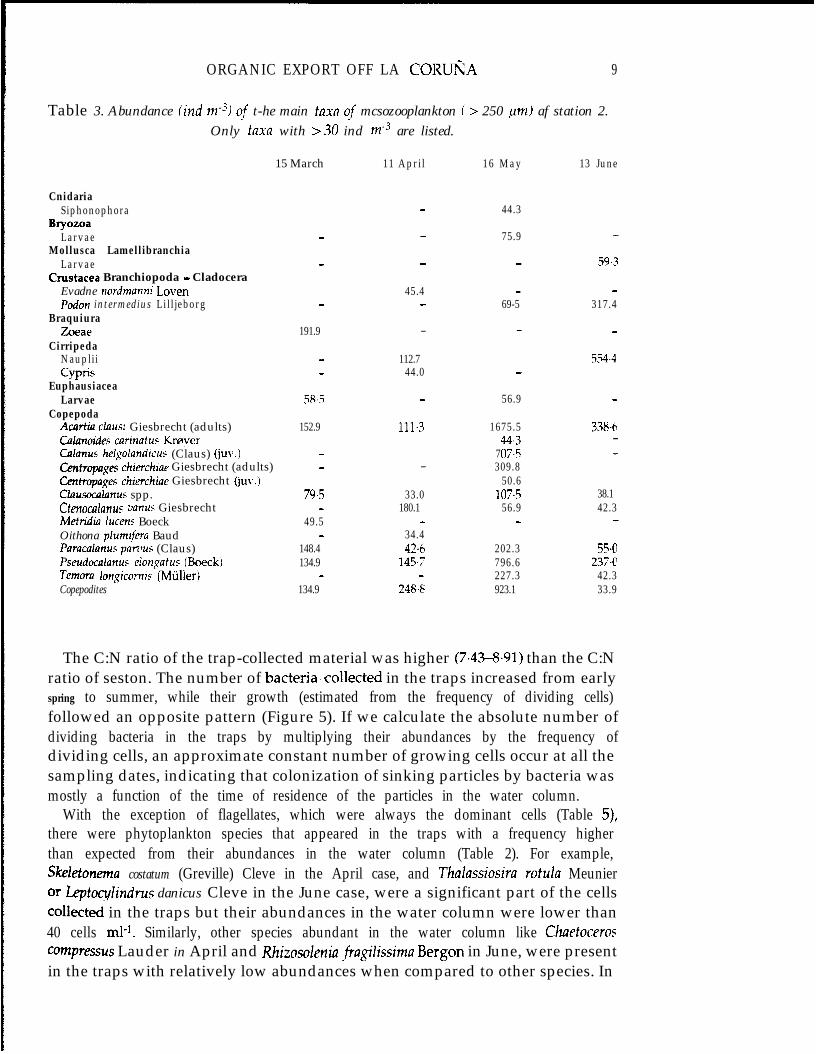
The mesozooplankton community was characterized by the appearance of peaks inthe abundance of meroplanktonic larvae of several taxa (Table 3), but also by thedominance of calanoid copepods in all the studied cases. The highest abundance anddiversity of copepods was found on 16 May, when there was also a large number ofcopepodites and young individuals of several species, indicating populations in agrowing stage. Most of these copepods were herbivores or omnivores (Valdes, 1993).These variations are typical of the spring and early summer season, as describedpreviously in this area by Valdks et al. (1991) and Valdes (1993).
Sediment- traps
Phytoplankton pigment concentrations in the trap-collected material were alwayslow (Table 4) when compared to those measured in the water column at station 2(Table 1). The total contribution of phytoplankton materials to the sinking matter, asestimated by the chlorophyll-equivalents, decreased from the March to the June case,and the same occurred with the fluxes of carbon and nitrogen. The phytoplankton-derived carbon constituted a small fraction of the sinking carbon, even at times of highchlorophyll-a concentration in the water column. Only 45% of the carbon fluxmeasured in the April case could be related to phytoplankton remains, and thisfraction was reduced to 10% in the June case. The contribution of livingphytoplankton to the total flux of sinking carbon never exceeded 8%, but maximumvalues were estimated when phytoplankton abundance was high.
ORGANIC EXPORT OFF LA CORURA 9
Table 3. Abundance (ind rne3) qf t-he main taxa qf mcsozooplankton i > 250 ,um) af station 2.Only taxa with >30 ind me3 are listed.
15 March 11 April 1 6 M a y 13 June
CnidariaSiphonophora
BryozoaLarvae
Mollusca LamellibranchiaLarvae
Crustacea Branchiopoda - CladoceraEvadne nordmanni LollenPodon intermedius Lill jeborg
BraquiuraZoeae
CirripedaNaupliiCYPG
EuphausiaceaLarvae
CopepodaAcarfia clausi Giesbrecht (adults)Calanoides carinatus KreyerCalanus helgolandicus (Claus) (jw.)Cenfrqqes chierchiae Giesbrecht (adults)Cenfropages chierchiae Giesbrecht (juv.)Clausocalanus spp.Cfenocalanus vanm GiesbrechtMefridia lucens BoeckOithona plumtfera BaudParacalanus panw (Claus)Pseudocalanus elongatus (Boeck)remora longicornw (Miiller)Copepodites
-
-
-
191.9
--
58.5
152.9
--
79.5-
49.5-
148.4134.9
-134.9
- 44.3
-
45.4-
112.744.0
-
117.3
33.0180.1
-34.442.6
1457-
248.E
75.9
-
-69-5
-
56.9
1675.544.3
7 07.5309.8
50.6107.5
56.9-
202.3796.6227.3923.1
-317.4
-
5-54.4
-
3386
38.142.3
55.0237.0
42.333.9
The C:N ratio of the trap-collected material was higher (7.43-8.91) than the C:Nratio of seston. The number of bacteria,collected in the traps increased from earlyspring to summer, while their growth (estimated from the frequency of dividing cells)followed an opposite pattern (Figure 5). If we calculate the absolute number ofdividing bacteria in the traps by multiplying their abundances by the frequency ofdividing cells, an approximate constant number of growing cells occur at all thesampling dates, indicating that colonization of sinking particles by bacteria wasmostly a function of the time of residence of the particles in the water column.
With the exception of flagellates, which were always the dominant cells (Table 5),there were phytoplankton species that appeared in the traps with a frequency higherthan expected from their abundances in the water column (Table 2). For example,SkeZetonema costatum (Greville) Cleve in the April case, and Thlassiosira rotula Meunieror Leptocylindrus danicus Cleve in the June case, were a significant part of the cellscollected in the traps but their abundances in the water column were lower than40 cells ml-l. Similarly, other species abundant in the water column like Chetoceroscompressus Lauder in April and Rhizosolenia fiagilissima Bergon in June, were presentin the traps with relatively low abundances when compared to other species. In

1 I
10 ANTONIO BODE ET AL.
-; 500 25
g 400 20s
** 300 8 15s-8
200 g 10
3 100 5s2 0 0 . . . . ,
Mar APT May Jun Mar APT May Jun
Dates Dates
Figure 5. (A) Mean (&SD) abundance of bacteria iri ‘the material collected in the traps. (A) Valueswere normalized to one day and expressed in x 103 cells ml-’ of water inside the trap. (B) Frequencyof dividing cells (FDC) inside the traps. Values normalized to one day of collection and expressed aspercentages of total bacteria abundance.
Table 4. Vertical fluxes ( k SD, N=4) of phytoplankton pigments and particulate materialcollected by the traps. Phytoplankton-derived carbon calculated as in Ferndndez et al. (1995).
Chl-a &SDPhaeo ?SDChl-aq,,, & SDC *SDN k!SDC:N rc_SDC-Chl-XC + SDC-Chl-u,,,:C + SD
(mg mW2 d-‘) 046 kO.19 la9 k1.10 0.43 *ool(mg mm2 d-t) 4.30 k1.12 3.35 *1-20 2.28 kO.23(mg me2 d-‘) 6-96 f 1.58 6.15 +240 3.88 kO.33(mg me2 d-‘) 120269 k 6034 780.42 + 336.01 627.79 z 10046(mg me2 d-‘) 158.07 + 14.61 110.89 + 53.31 88-29 & 13.41
By atoms 8.91 10.53 x.33 TO.35 8.29 t0.18“/o 1.95 io85 7.68 -7.49 3.54 20.77O/o 29.02 * 7.15 45.44 + 21-85 31.37 c -4.89
16 March 12 April 17 May 14 June
0.34 kO.340.19 io.070.63 *0,33
322.78 k 16.935035 + 4427.43 rO.38xl9 24.949.62 k4.74
C-Chl-a:C is the percentage of the carbon flux that can be attributed to living phytoplankton. C-C~~-U.+~:Cis the percentage of total phytoplankton in the carbon flux.
contrast, Chaetoceros socialis was well represented in both the water column and thecollected material in the April case. Some species of benthic or benthopelagic diatomslike Trachyneis sp., Grammatophora sp., Gyros&a balticurn (Ehrenberg) Cleve andNitzschia sigmoidea (Nitzch) W. Smith, were observed in the traps but not in the watercolumn. However, their abundances were low (from O-1 to 84x106 cells rns2 d-l) whencompared with planktonic species and were not included in Table 5.
Faecal pellets were collected in the traps in all the studied cases. The smaller pellets( < 200 pm) were more abundant in the March case, approximately twice the numberfound in all other cases (Figure 6). The larger pellets were first detected in the Aprilcase and reached their maximum in the May case, when the abundance of copepodsin the water column was also maximum (Table 3).

ORGANIC EXPORT OFF LA CORmA 11
Mar Apr MayDates
Jun Mar Apr MayDates
Jun
Figure 6. Mean ( *SD) abundance of faecal pellets collected in the traps: (A) pellets < 200 pm; (B)pellets >200 pm.
Table 5. Mean abundance (mean & SE; x 106 cells m-2 d-l) of the main phytoplankton taxacollected by the traps during the study. Only groups with > x301 cells rn-? d-l are listed.
16 March 12 April 17 May 14 June
DinophyceaeSmall dinoflagellates ( ~30 pm) 12.8 t-3.0
DiatomophyceaeSmall centric diatoms ( ~30 jlrn) -Small pennate diatoms ( ~30 pm) 7oe5 24.4Chaetoceros @finis Lauder+var willei Gran -Chaetoceros gracilzs SchiittChaetoceros socialis Lauder -Chmtoceros spp (spores)Lauderin borealis GranLeptocylindrus danicus CleveKhizosolenia setigera Brightwell -Skeletonema cost&urn (Greville) Cleve -Thalassiosira rotula Meunier ,. -Cyptophyceae -
FlagellatesSmall flagellates (3-5 pm) 171.0 k80.3Medium-sized flagellates (!+8 pm) 160.6 & 37.9Large flagellates ( > 8 pm) 14.3 k2.5
17.1 k4.3
1759 + 56.217.1 +0-o
-
-135.1 k86.6
-
249.8 k 123.689.2 &- 15.517.1 +3.5
15.2 51.7
27.2 +4.9
731.5 -t 270.889.2 k 17.932.9 5 6.4
421.5 k 34.8121.9 f 43.6
21.7 k2.4
28.9 +4.8
19.3 kO.0-
24.8 2 10.8-
13.4 & 3.4
17.4 +3.933.9 +9.2
44.2 k32.5
22.8 +8.8-
DISCUSSION
The measured rates of exported materials from the euphotic zone off La Coruna,ranging from 323 to 1203 mg C m-2 d-i, are high when compared to other studies.Femandez et al. (1995) reported for a shelf-break station in the nearby Cantabrian Sea(southern Bay of Biscay) a range of 173 to 236 mg C mw2 d-i. Our values were higheven when compared to eutrophic areas, like the VERTEX stations V5c (333 mg C m-2d-‘) and Vl (212 mg C me2 d-l) as reported in Knauer et al. (1984), the oceanic spring-bloom in the North Atlantic (305 mg C m-2 d-i, Bender et al., 1992), or the spring-bloom in the Kattegat (up to 700 mg C m-2 d-i, Olesen & Lundgaard, 1995). However,

12 ANTONIO BODE ET AL.
our measured export rates during phytoplankton blooms are comparable to thoseobserved in a similar system on the coast of the North Pacific (432 mg C me2 d-l,Knauer et al., 1979), and do not exceed the large values reported for the mussel raft-culture areas in the southern Galician rias (north-west Spain), where sinking rates ofup to 24-6 g mm2 d-l (Cabanas et al., 1980) and 200-2400 me2 d-l (Tenore et al., 1982)were estimated from trap measurements.
Although the possibility of sediment resuspension by storms and water advectionoccurring in the study area can not be completely ruled out, there are severalindications that most of the matter collected by our traps during the study have apelagic origin. First, concentrations of more than 500 mg C m-3 have been measuredthrough the euphotic zone during spring blooms at the same station. Casas et al. (1997)reported carbon concentrations higher than 900 mg C me3 at station 2 in March 1992.The vertical distribution of particulate carbon was related to that of chlorophyll-a andboth concentrations decreased towards the bottom. Second, the C:N molar ratioobserved in the trap-collected material (74-8.9 by atoms) was only slightly higher thatthe C:N ratio of seston (65-7.4), and resulted low when compared to values measuredin sediments of the area ( % 9.3 by atoms; A. Bode & M. Came, unpublished data).Higher values were observed in the material collected by traps in the Cantabrian Sea( > 9 by atoms; Femandez et al., 1995). As Olesen & Lundgaard (1995) noted in theirstudy of the Kattegat, this indicates that most of the material collected in the traps wasof pelagic origin, since allochtonous materials usually have a C:N ratio greater thannine (Lancelot & Billen, 1985). Ln addition, the abundance of recognizable benthicmicroalgae in the traps was low during our study.
Resuspension of sedimented materials could be expected at times of activeupwelling, when deep waters flow over the bottom from the outer shelf and ascendtowards the coast. However, the lowest vertical export fluxes of organic matter in ourstudy were recorded in the June case, suggesting that horizontal advection was moreimportant than resuspension for particle export. The offshore movement of surfacewater can transport a significant amount of the organic matter and organismsproduced during the bloom. These materials sink to the bottom in outer-shelf zones ofthis region, as described by Varela et al. (1991) and Castro et al. (1994). A similarmechanism originates the accumulations of organic-rich sediments outside the highlyproductive rias of this region (Lopez-Jamar et al., 1992). While this mechanism canexplain the low sedimentation rates during the early phases of upwelling, whenphysical advection is high, biological remineralization of the organic matter can beresponsible of lower sedimentation rates at the end of the bloom induced by theupwelling. Bode & Varela (1994) have estimated that most of the phytoplanktonbiomass produced during a summer upwelling event off La Con&a may beremineralized in a few days by the microheterotrophic community in the watercolumn.
Most of our results indicate an uncoupling between organic production in the watercolumn and sedimentation of particles. This may be in part a consequence of the shorttime-scales employed in the study. More coupling is expected on monthly or annualtime-scales (Berger et al., 1989; Asper et al., 1992). The traps deployed for a period ofup to three days may be recording the consequences of past productive events, while

ORGANIC EXPORT OFF LA CORUNA 13
the activity of the plankton during the time of deployment may be completelydifferent. This may be particularly true for the initial stages of a bloom, when there isless chance of cell and particle sinking, or the final stages when production is low andthe sinking rates are high (Bender et al., 1992; Bode et al., 1994b). The occurrence ofcells of some phytoplankton species in the traps, that were not particularly abundantin the water column at the time of trap deployment, like Skeletonema costatum in Aprilor 7hulassiosiru r&la in June, may be an indication of past blooms dominated by thesespecies.
Phytoplankton-derived carbon in the trap-collected material never exceeded 50% ofexported carbon in our study. Femandez et al. (1995) reported phytoplanktoncontributions from 50 to 70% to exported carbon in the Cantabrian Sea duringsummer, although the fraction of living phytoplankton was < 10%. Low phytoplank-ton contribution to total carbon export was expected in coastal zones with largeconcentrations of detrital particles in the seston. Olesen & Lundgaard (1995) reportedthat phytoplanktonic materials accounted for 30% of total exported carbon during thestratification period in the Kattegat, however there was a rapid sinking of cells afterblooms and the fraction of living phytoplankton was 18%.
There are indications that some particles present in the euphotic zone of our studyarea are quickly transferred to the sediment. Large phytoplankton cells, chains andaggregates were well represented in the traps. Even small phytoplankton andnanoflagellates of <8 pm were abundant through the study. This suggests the rapidformation of cell aggregates that would increase the sinking speed of individual cells.A phytoplankton cell of 20 pm in diameter will sink 1.7 m d-i, while a particle of200 pm in diameter will sink ‘c 178 m d-i, if we do not account for particledecomposition during settling (Csanady, 1986). This aggregation process may explainthe presence of abundant faecal pellets in the traps. Zooplankton faecal pellets, most ofthem produced by copepods, are one of the main vehicles of particle aggregation andtransfer from the surface to the sea bottom (Bathman et al., 1987). Most faecal pelletsproduced by zooplankton ~200 pm are thought to be recycled within the euphoticzone (Bathman et al., 1987; Small et al., 1987). However, we found a large number ofpellets ~200 pm in the traps, particularly in the early-spring samples. The variationsin the number of these small pellets closely followed those in total carbon and nitrogenin the traps, which suggests that they are a significant part of the exported material.The pellets > 200 pm were more abundant in May, when most herbivorous copepodsreached their maximum abundances. These large pellets may also be a significantcontribution to the carbon export because of their high potential sinking speeds. Usingthe equations of Csanady (1986) we can estimate that a faecal pellet of 200 pm indiameter, even being partlv decomposed by attached bacteria while sinking, can reachan average sinking speed* of -90 m d-i. This means that most of the large pelletscollected in our traps were produced at the time-scale of the trap measurement.
In conclusion, the studied site was characterized by a large export of carbon,nitrogen and biogenic materials. Most of the particles collected in the traps appearedto be of pelagic origin, mainly because of low C:N ratios, dominance ofphytoplanktonic over phytobenthic species and the abundance of zooplankton faecalPellets. The contribution of nhvtonlankton to the exported carbon was generally low,

14 ANTONIO BODE ET AL.
but the values measured in this study were similar to those reported for other coastalareas. The minimum value of phytoplankton-derived carbon in the trap-collectedparticles was recorded in the case studied in June. This may be due in part to theoffshore displacement of surface water caused by the upwelling.
We are indebted to the crew of the RV ‘Lura’ for their collaboration during trap deploymentand recovery. We also thank the assistance of Femanda Martin and Isabel Gonz6lez for fieldsampling and laboratory processing of seston and plankton samples, and the help of Ma LuisaFermoso with the CHN analysis. Wind data were kindly provided by the Centro Metorol6gicoZonal at La Conuia (Institute Espafiol de Meteorologia) and the upwelling indices werecomputed with the help of Jorge Lorenzo. This work was funded in part by the Spanish researchprojects IEO-1007 and CICYT AMB93-0014. S.B. acknowledges the Spanish Ministry of Scienceand Education for a FPI fellowship.
REFERENCES
Asper, V.L., Deuser, W.G., Knauer G.A. & Lohrenz, S.E., 1992. Rapid coupling of sinkingparticle fluxes between surface and deep ocean waters. Nature, London, 357, 670-672.
Bakun, A., 1973. Coastal upwelling indices, west coast of North America, 1946-1971.Atmospheric Administration Technical Report NMFS, no. SSRF-671.
Bathmann, U.V., Noji, T.T., Voss, M. & Peinert, R., 1987. Copepod faecal pellets: abundance,sedimentation and content at a permanent station in the Norwegian Sea in May/June 1986.Marine Ecology Progress Series, 38, 45-51.
Bender, M., Ducklow, H., Kiddon, J., Marra, J. & Martin, J., 1992. The carbon balance during the1989 spring bloom in the North Atlantic Ocean, 47’N, 2O;‘W. Deep-Sea Research, 39,1707-1725.
Berger, W.H., Smetacek, V.S. & Wefer, G., 1989. Ocean productivity and paleoproductivity - anoverview. In Productivity of the ocean: present and past. Report qf the Dahlem Workshop onProductivity of the ocean: present and past, Berlin, 24-29 April 1988 (ed. W.H. Berger et al.),pp. l-34. New York: John Wiley & Sons. [Life Sciences Research Paper no. 44.1
Blanton, J.O., Tenore, K.R., Castillejo, F., Atkinson, L.P., Schwing, F.B. & Lavin, A., 1987. Therelationship of upwelling to mussel production in the rias of the western coast of Spain.lournal qf Marine Research, 45, 497-511.
Bode, A., Casas, B. & Varela, M., 1994a. Size-fractionated primary productivity and biomass inthe Galician shelf (NW Spain): netplankton versus nanoplankton dominance. ScientiaMarina, 58, 131-141.
Bode, A. et al., 1994b. Biological characteristics of the plankton associated to a shelf-break frontoff the Galician coast. Gaiu, 8, 9-18.
Bode, A. & Varela, M., 1994. Planktonic carbon and nitrogen budgets for the N-NW Spanishshelf: the role of pelagic nutrient regeneration during upwelling events. Scientia Marina, 58,221-231.
Botas, J.A., Femindez, E., Bode, A. & Anad&, R., 1990. A persistent upwelling off the centralCantabrian coast (Bay of Biscay). Estuarine, Coastal and Shelf Science, 30, 185-199.
Cabanas, J.M., Gonzalez, J.J., MariIio, J., Perez, A. & Roman, G., 1980. Estudio de1 mejillbn y desu epifauna en 10s cultivos flotantes de la Ria de Arosa. III. Observaciones previas sobre laretenci6n de particulas y la biodeposici6n de una batea. Boletin de1 Institute Espariol d eOceanograjia, 5, 45-50.
Casas, B., Varela, M., Canle, M., Gor&lez, N. & Bode, A., 1997. Seasonal variations of nutrients,seston and phytoplankton, and upwelling intensity off La Coruiia (NW Spain). Estuarine,Coastal and Shelf Science, 44, 767-778.
Castro, C.G., Perez, F.F., Alvarez-Salgado, X.A., Ros6n, G. & Rios, A.F., 1994. Hydrographicconditions associated with the relaxation of an upwelling event off the Galician coast (NWSpain). \oumal of Geophysical Research, 99C, 5135-5147.
Csanady, G.T., 1986. Mass transfer to and from small particles in the sea. Limnology andOceanography, 31,237-248.

ORGANIC EXPORT OFF LA CORURA 15
Femandez, E. & Bode, A., 1991. Seasonal patterns of primary production in the centralCantabrian Sea (Bay of Biscay). Scimtia Marina, 55, 629-636.
Femandez, E., Mar-anon, E., Cabal, J., Alvarez, F. & Anadon, R., 1995. Vertical particle flux in outershelf waters of the southern Bay of Biscay in summer 1993. Oceunolo@ca Actu, 18, 379-384.
Fraga, F., 1976. Fotosintesis en la Ria de Vigo. lnz~estigacion Pesquera. Barcelona, 45, 151-167.Fraga, F., 1981. Upwelling off the Galician coast. In Coastal upwelling. JDOE Jnternational
Symposium on coastal upwelling, Los Angeles, California, 4 February 1980 (ed. F.A. Richards),pp. 176-182. Washingt on, DC: American Geophysical Union.
Knauer, G.A., Martin, J.H. & Bruland, K.W., 1979. Fluxes of particulate carbon, nitrogen andphosphorus in the upper water column of the northeast Pacific. Deep-Sea Research, 26,97-108.
Knauer, G.A., Martin, J.H., & Karl, D.M., 1984. The flux of organic material out of the euphoticzone. In Global ocean fJux studies. Proceedings of a Workshop, pp. 136-150. Woods Hole:National Academy Press.
Lancelot, C. & Billen, G., 1985. Carbon-nitrogen relationships in nutrient metabolism of coastalmarine ecosystems. Advances in Aquntic Microbiology, 3, 263-321.
Lavin, A., Diaz de1 Rio, G., Cabanas, J.M. & Casas, G., 1991. Afloramiento en el noroeste de laPeninsula Iberica. Indices de afloramiento para el punto 43-N 11’W. Period0 1966-1989.lnforme Tecnico. lnstituto Espariol de Oceanogr@a, 91, l-39.
Lopez-Jamar, E., Cal, R.M., Gonzalez, G., Hanson, R.B., Rey, J., Santiago, G. & Tenore, K.R..1992. Upwelling and outwelling effects on the benthic regime of the continental shelf offGalicia, NW Spain. Journal @Marine Research, 50, 465488.
Newell, S.Y. & Christian, R., 1987. Frequency of dividing cells as an estimator of bacterialproductivity. Applied and Enz~ironmental Mxrobiology, 42, 23-31.
Olesen, M. & Lundsgaard, C., 1995. Seasonal sedimentation of autochtonous material from theeuphotic zone of a coastal system. Estuarine, Coastal and Shelf Science, 41, 475490.
Rios, A.F., Perez, F.F. & Fraga, F., 1992. Water masses in the upper and middle North AtlanticOcean east of the Azores. Deep-Sea Research, 39, 645-658.
Small, L.F., Knauer, G.A. & Tuel, M.D., 1987. The role of sinking fecal pellets in stratifiedeuphotic zones. Deep-Sea Research, 34, 1705-1712.
Smetacek, VS., 1980. Annual cvcle of sedimentation in relation to plankton ecology in westernKiel Bight. Opheliu, 1, supplement, 65-76.
Tenore, K.R. et al., 1982. Coastal upwelling in the Rias Bajas, NW Spain: contrasting the benthicregimes of the Rias de Arosa and de Muros. journal qf Marine Research, 40, 701-772.
Tenore, K.R. et al., 1995. Fisheries and oceanography off Galicia, NW Spain: mesoscale spatialand temporal changes in physical processes and resultant patterns of biologicalproductivity. ]ournaJ of Geophysical Research, lOOC, 10943-10966.
Unesco, 1994. Protocols for the Joint Global,Ocean Flux Study (JGOFS) core measurements.Manuals and Guides, 29, l-170.
Valdes, L., 1993. Composition, abundancia y distribucio’n de1 mesozooplancton en la plataformacontinentaJfiente u La Coruria. PhD thesis, Universidad de Oviedo, Spain.
Valdes, L., Alvarez-Ossorio, M.T., Lavin, A., Varela, M. & Carballo, R., 1991. Ciclo anual depar&netros hidrograficos, nutrientes y plancton en la plataforma continental de La Corur?a(NO, Espaiia). Boletin de1 Jnstituto Espariol de Oceanografi, 7, 91-138.
Varela, M., Diaz-del-Rio, G., Alvarez-Ossorio, M.T. & Costas, E., 1991. Factors controllingphytoplankton size class distribution in the upwelling area of the Galician continental shelf(NW Spain). Scientia Marina, 55, 505-518.
Wa~smann, P., 1991. Dynamics of primarv production and sedimentation in shallow fjords andpolls in western Norway. Oceanography and Marine Biology. Annual Review, 29, 87-154.
Wefer, G. & Fisher, G., 1993. Seasonal patterns of vertical particle flux in equatorial and coastalupwelling areas of the eastern Atlantic. Deep-Sea Research, 40, 1613-1645.
wiebe, P.H., 1988. Functional regression equations for zooplankton displacement volume, wetweight, and carbon: a correction. Fishery Bulletin. National Oceanic and AtmosphericAdministration. Washington, DC, 86, 833-835.
Submitted 9 February 1996. Accepted 27 October 1996.
Copyright © 2022 FDOKUMEN




















