
Positions of Chromosome 3pl'L2 Fragile Sites (FRA3B) within the FHIT Gene
6
1997;57:1166-1170. Cancer Res Drazen B. Zimonjic, Teresa Druck, Masataka Ohta, et al. Gene FHIT the ) within FRA3B Positions of Chromosome 3p14.2 Fragile Sites ( Updated version http://cancerres.aacrjournals.org/content/57/6/1166 Access the most recent version of this article at: E-mail alerts related to this article or journal. Sign up to receive free email-alerts Subscriptions Reprints and . [email protected] Department at To order reprints of this article or to subscribe to the journal, contact the AACR Publications Permissions . [email protected] Department at To request permission to re-use all or part of this article, contact the AACR Publications on March 17, 2014. © 1997 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from on March 17, 2014. © 1997 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
-
Upload
independent -
Category
Documents
-
view
5 -
download
0
Transcript of Positions of Chromosome 3pl'L2 Fragile Sites (FRA3B) within the FHIT Gene

1997;57:1166-1170. Cancer Res Drazen B. Zimonjic, Teresa Druck, Masataka Ohta, et al.
GeneFHITthe ) withinFRA3BPositions of Chromosome 3p14.2 Fragile Sites (
Updated version
http://cancerres.aacrjournals.org/content/57/6/1166
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
To request permission to re-use all or part of this article, contact the AACR Publications
on March 17, 2014. © 1997 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from on March 17, 2014. © 1997 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

(CANCERRESEARCH57. 1166-1 170, March15. 1997)
constitutive familial clear-cell renal carcinoma-associated translocation t(3;8)(pl4.2;q24) (1 1, 12) has been found to be indistinguishablefrom the most frequent of the common fragile sites, FRA3B (13).Nevertheless, evidence implicating fragile sites in the process ofneoplastic development has been circumstantial. It has indeed beenargued that it is very unlikely that breaks in a fragile site should beassociated with cancer (14).
The constitutive ape-inducible fragile site at chromosome band3pl4.2 (FRA3B) is of special interest, because it is the most commonly expressed in humans (15), and the region 3pi4.2 is involved indeletions in a variety of histologically different cancers (16—18),thet(3;8) translocation in familial RCC, and Kaposi's sarcomas (19). Inaddition, consistent with previous cytogenetic observations linkingviral integration sites and fragile sites (8, 20), the first molecularevidence for HPV integration in FRA3B was provided recently (21).The positions of some ape-induced gaps and breaks in FRA3B havebeen determined by isolation of YAC clones encompassing the breaks(22—25)and mapping ape-induced hybrid breaks (21, 26) or plasmidintegration sites (27, 28) to regions of the YAC map.
We have recently isolated the FHJT gene, which encompassesperhaps 1 Mb, straddling the t(3;8) break, the FRA3B, and cancercell-specific homozygous deletions (29), and have suggested that theFHIT gene may be the target of FRA3B-induced deletions in cancercells. The FlIT gene is ex@pressedin all tissues thus far examined(29), including normal lymphocytes (30) and ape-induced lymphocytes, in which the normal-sized FHJT transcript was observed (datanot shown).
To determine the relationship of the FHJT gene to cancer celldeletions and FRA3B breaks and gaps, we have determined the positions of FHlTexons relative to the translocation, viral integration site,cancer cell deletions, and FRA3B breaks described previously, andhave then used FHIT cosmid probes to determine the positions ofspecific FHJT exons relative to the ape-induced gaps and breaks in
human lymphocyte metaphase chromosomes.
MATERIALS AND METHODS
Localization of STSs by PCR. Primer pair sequences for amplification ofSTSs were obtained from the Genome Data Base or were designed frompublished sequences in GenBank; the numbers beginning with U are GenBankaccession numbers of the sequences. U39799 and U39804 primer pairs are
listed in Druck et a!. (30). Other newly designed primer pairs are: U39793F(5'-TGGATTGAmCACACC-3') and U39793R (5'-GCCAGAGGGAGA ri-i-i- iC-3') at bp 104 and 265 of the U39793 sequence, respectively;U061 l8F (5'-CGUGTAGTFCTGA'ITFGCA-3') and U06l l8R (5'-GGATA1T1'GTCACTGCC1TF-3') at nucleotides 20 and 330, respectively;U60203F (5'-GTGCTCAGAGATFACATGC-3' and U60203R (5'-GTATGTCAGCCAAACACC-3') at positions 786 and 1069, respectively; and cSB1AF(5'-CAAAGCAGAAATAAAACTGC-3') and c5B 1AR (5'-AlTFCCAACAAGCFGCCA-3'), which amplify a —2l8-bpproduct containing a TAdinucleotide repeat (locus name D3S4490; GenBank accession no. U76285).
Each primer pair was used in a PCR reaction with template DNA containingsome portion of the FHIT genomic locus. PCR amplifications were carried outin 12.5-pJ reaction volumes with 10 ng of cloned genomic DNA or 100 ng of
total genomic DNA, 20 ng of each primer, 10 mMTris-HCI (pH 8.3), 50 mr@iKC1,0.1 mg/rn)gelatin, 1.5 mMMgCl2,0.2 mMeach deoxynucleotidetriphos
1166
Positions of Chromosome 3pl'L2 Fragile Sites (FRA3B) within the FHIT Gene'
Drazen B. Zimonjic, Teresa Druck, Masataka Ohta,2 Kumar Kastury,3 Carlo M. Croce, Nicholas C. Popescu,
and Kay Huebner4
Laboratory of Experinwntal Carcinogenesis. Division of Basic Sciences, National Cancer Institute, Bethesda, Maryland 20892 (D. B. 1, N. C. P.!; Kimmel Cancer institute,JeffersonMedicalCollege.ThomasJeffersonUniversity.Philadelphia,Pennsylvania19107fT. D..M.0.. K.K., C.M.C.,K.H.J
ABSTRACT
The FHIT gene spans approximately 1 Mb of DNA at chromosomeband 3p14.2, which includes the famffial renal ceHcarcinoma chromosometranslocation breakpoint (between FHIT exons 3 and 4), the most Irequently expressed human constitutive chromosomal fragile site (FRA3B,telomeric to the t(3;8) translocation), and numerous homozygous deletionsin various human cancers, frequently involving FHIT exon 5. The FRA3Bhas previously been shown to represent more than one specific site, andsome specific representatives of FRA3B breaks have been shown to fall intwo regions, which we know to be in FHIT introns 4 and intron 5. Becausebreakage and integration of exogenous DNA in this chromosome region isfrequent in aphidicolin-treated somatic cell hybrids, cancer cells, and,
presumably, aphidicolin-treated normal lymphocytes that exhibit gaps orbreaks, we determined by one- and two color fluorescence in situ hybridization, using cosmids covering specific regions of the FHIT gene, thatmost of the aphidicolin-induced gaps at FRA3B fall within the FHIT gene,
with the higjtest frequency ofgaps falling in intron 5 ofthe FHIT gene, lessthan 30 kb telomeric to FHIT exon 5. Gaps also occur in intron 4, wherea human papillomavirus 16 integration site has been localized, and inintron 3, where the t(3;8) break point is located. These results suggest thatthe cancer-specific deletions, which frequently involve introns 4 and 5,originated through breaks in fragile sites.
INTRODUCTION
Chromosomal fragile sites are regions susceptible to breakageunder specific experimental conditions. Antifolates, fluorodeoxyuridine plus methotrexate, and ape,5 a DNA polymerase inhibitor (1),induce the expression of fragile sites during metaphase as nonstaininggaps or breaks, usually involving both chromatids (for review, seeRef. 2). The latest compendium comprises 103 fragile sites classifiedas common and rare, constitutive and inherited, that have been localized in all human chromosomes except chromosomes 21 and Y (3).Although fragile sites are dispersed throughout the genome, they tendto cluster at G-light chromosome bands (4) that are known to containGC-nch Alu repeats and are constitutionally more relaxed and unfolded during transcription (5). The location of fragile sites has beencorrelated cytogenetically with locations of recurrent translocationand deletion breakpoints in cancer cells, as well as the integration sitesof oncogenic viruses (6—9).Moreover, structural rearrangements induced by mutagens or carcinogens through fragile sites can be cytogenetically indistinguishable from those in cancer cells (10), and the
Received I 1126/96;accepted 1/13/97.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordance with18 U.S.C. Section 1734 solely to indicate this fact.
I This work was supported partially by United States Public Health Service Grants
CA51083, CA39860, and CA21124, and a gift from R. R. M. Carpenter Ill and Mary K.Carpenter.
2 Present address: Banyu Tsukuba Research Institute, Okubo 3, Tsukuba 300-33 Japan.
Phone: 81-298-77-2000; Fax: 81-298-77-2024.L1 Present address: Department of Endocrinology. Charles R. Drew University of
Medical Science, Room 3069, 162 1 E. I20th Street, MPO2, Los Angeles, CA 90059.Phone:(213)563-5959.
4 To whom requests for reprints should be addressed, at Kimmel Cancer Institute,
Room 1008, Jefferson Medical College, Thomas Jefferson University, 233 South 10thStreet, Philadelphia, PA 19107. Phone: (215) 503-4656; Fax: (215) 923-4498.
5 The abbreviations used are: ape. aphidicolin; HPV, human papillomavirus; YAC,
yeast artificial chromosome; STS, sequence-tagged site; BAC, bacterial artificial chromosome; RCC, renal cell carcinoma; FISH, fluorescence in situ hybridization.
on March 17, 2014. © 1997 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

POS@ONS OF FRA3B SITES WITHIN THE FH!T GENE
Telomere4-
Centromere-@
) 00 kb
Fig. 1. Map of the Fllfflocus. The chromosome 3pl4.2 region (—1Mb) is illustrated with approximate placement of the cosmid and BAC clones relative to each other. Positionsof relevant STSs were determined by PCR amplification with cosmid and BAC DNA templates. Localization of the telomeric side of the lung carcinoma deletion (U60203), the pSV2neoplasmid integrations, the c13break, the HPVI6 integration site, the t(3;8) break, as well as 5'-PTPRG are shown (arrows). D3S4260 and IA6G/H primers are derived from the cosmid1A6, to which 10 fragile site breaks in hybrid cells have been mapped (37).
5'PTPRG
‘I,LI)U
@8E8@ll14—@—14-c06#7c05#9c05#1 E3E2El.-4j..cX3—O-'cX2—II-cXl c19-20 —
@!Af@ O@AAb
phate, and 0.5 units of Taq polymerase. Amplifications were for 30 cycles of94°Cfor 30 s, 57°Cfor 30 s, and 72°Cfor 30 s. Products were analyzed byelectrophoresis in ethidium bromide-stained 1.5% agarose gels. Presence of anamplification product determined localization of a STS to a particular genomic
clone.Isolation of Genomic Clones. New cosmid clones were isolated as de
scribed previously (29), and BAC clones were identified by PCR amplificationof specific STS fragments from pooled BAC library template DNAs (ResearchGenetics, Huntsville, AL); individual BAC clones were then ordered fromResearch Genetics, single colonies for specific STSs were identified andgrown, and end sequences were determined, as for cosmids.
FISH Analysis. Normal peripheral lymphocytes and lymphoblastoid cellsderived from a member of the RCC family with the t(3;8) translocation werecultivated in RPM! 1640 supplemented with 15% fetal bovine serum, penicil
lin-streptomycin (50 mg/mI), and 0.2 m@i glutamine. The expression of FRA3B
was induced by ape (0.2 p.M;Sigma Chemical Co., St. Louis, MO) dissolvedin DMSO (31), or by combined ape and ethanol (0.2%) during the last 26 h of
culture (32). Metaphase chromosomes were obtained after a 1-h colcemid (50ILg/ml) treatment by standard KC1 hypotonic incubation and acetic acidmethanol fixation. Six previously described cosmid probes (25, 29) [19-20(representing the PTPRG 5' end, as shown in Fig. 1), cX3 (including FHJTexon 3), cX4 (exon 4), c76 (exon 5), c36, and c63 (intron 5)] labeled withbiotin or digoxigenin, as well as chromosome 3 and 8 painting probes (Oncor,Gaithersburg, MD) were used for in situ hybridization. The conditions for
FISH, detection ofthe signal, and digital imaging were carried out as describedpreviously (33, 34).
RESULTS
The FHIT/FRA3B Map. We have previously described a YACand partial cosmid contig for the FlIT locus, which extends frommore than 300 kb centromeric to the t(3;8) break to an estimated 700kb telomeric to the t(3;8) (25, 29). A map of the FHITlocus is shownin Fig. 1, on which we have placed a number of relevant markers andthe cosmid probes and the position of the FH!T exons; the relativedistances of the FHIT exons 1, 2, and 3 relative to each other and the5' end of the PTPRG gene is not known, nor is the position of c5BlArelative to exon 1 known. The positions of markers IA6G/H andD3S4260shownbetweenexons4 and5 (Fig.1,E4andES,respectively) were deduced by PCR amplification from our YAC, BAC, andcosmid templates, with pairs of primers derived from published se
quences. The positions of the U39793, U39804, and U39799 markers,which represent the HPV16 integration site (21), were deduced byamplification of appropriate fragments from the BAC clone (shown inFig. 1) and from knowledge of their positions relative to the nearbyMiul site and other markers that we mapped within the exon 4cosmid. The hybrid cl3 break, representing our 3pl4.2 hybrid de novo
break (35, 36), had previously been placed telomeric to FHJT exon 5based on Southern blot data (29), but we now know the break falls inintron 4, as shown here. The U60203 marker falls in cosmid cP4 inintron 5, based on amplification of a fragment from this template. Thismarker represents the telomeric side of a homozygous deletion observed in a lung carcinoma.6 Human sequences U40597 and U40401,representing portions of plasmid integration flanking sites (27, 28)were mapped within 4 kb telomenc of FHIT exon 5 and in cosmidsc36 and c76, respectively, by sequence identity; sequence U061l8,another plasmid flanking sequence, was mapped to cosmid c36; seeFig. 1 for positions of each of these sequences in intron 5. Additionally, numerous tumor-specific deletion end points fall within introns 4and 5 (30), and seven hybrid breakpoints identified by Paradee et a!.(37) are in intron 5. The relationship of these various chromosomelandmarks to each other and to the FH!T gene is summarized inFig.1.
Position of Exons Relative to FRA3B Gaps. Metaphase spreadsprepared from ape-treated peripheral lymphocytes derived from twonormal donors were hybridized with the six cosmid probes; 19-20,cX3, cX4, c76, c36, and c63 probes that span the t(3;8) and FRA3Bsites (25, 29); see Fig. 1 for a map of the region. To obtain a sufficientnumber of metaphases with informative signals at FRA3B, two orthree slides were hybridized with each cosmid probe. Only symmetrical signals were included in statistical analysis, although, due to thechromatid displacement at expressed fragile sites, single fluorescentspots were observed frequently. The location of the hybridizationsignal was assigned as medial (crossing the gap or break), proximal
6 T. 0. Ong, K. Fong, S. Bader, J. Minus, M. Le Beau, T. McKeithan, and F. Rassool.
Precise localization of the common fragile site at 3pl4.2 to the putative tumor suppressorgene FHJT and characterization of homozygous deletions in tumor cell lines withinFRA3B affecting FHIT transcription, unpublished, obtained from GenBank (accessionnumber U6O203).
1167
pSVZn.o HPV16
Q)0)Q)
@ + @+J!@@
F:::I 118k;@@ I II@I@@ C,)
I I I I II
63 E4
.I—c84 .—ll-cX4—c76@
—c36 BAC
on March 17, 2014. © 1997 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from
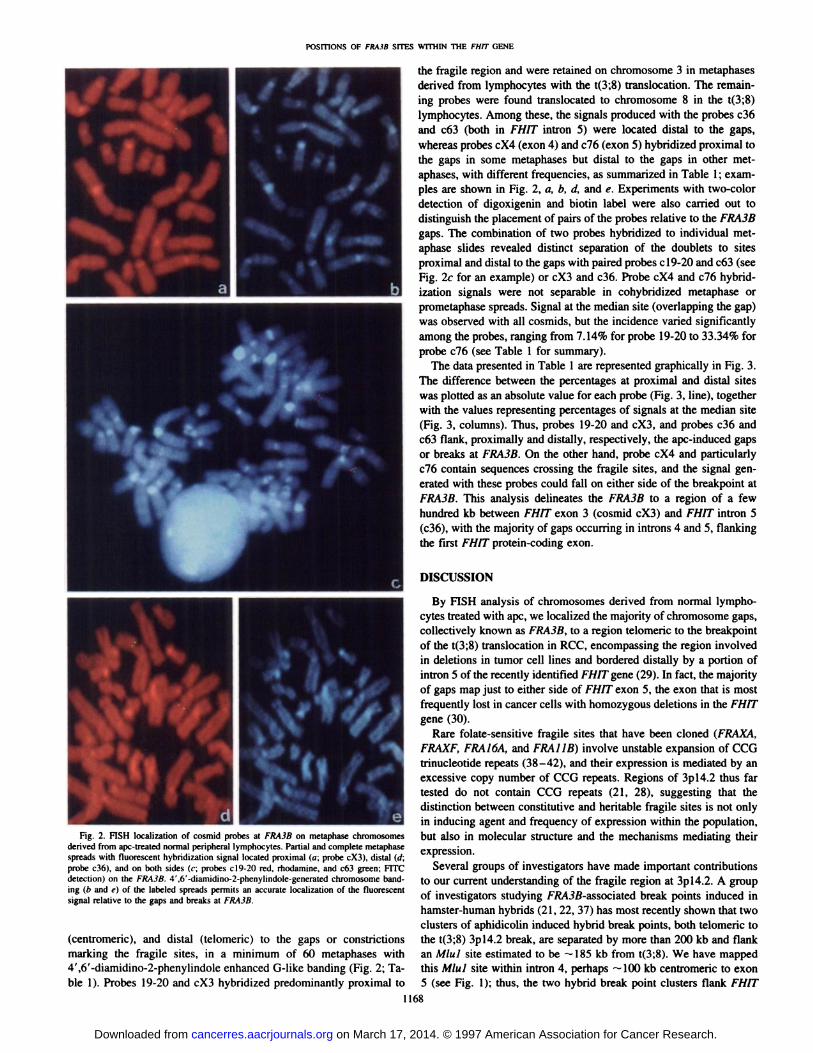
posmoNs OFFRA3BSITESWITHINThE FHITGENE
the fragile region and were retained on chromosome 3 in metaphasesderived from lymphocytes with the t(3;8) translocation. The remaining probes were found translocated to chromosome 8 in the t(3;8)lymphocytes. Among these, the signals produced with the probes c36and c63 (both in FHJT intron 5) were located distal to the gaps,whereas probes cX4 (exon 4) and c76 (exon 5) hybridized proximal tothe gaps in some metaphases but distal to the gaps in other metaphases, with different frequencies, as summarized in Table 1; exampies are shown in Fig. 2, a, b, d, and e. Experiments with two-colordetection of digoxigenin and biotin label were also carried out todistinguish the placement of pairs of the probes relative to the FRA3Bgaps. The combination of two probes hybridized to individual metaphase slides revealed distinct separation of the doublets to sitesproximal and distal to the gaps with paired probes cl9-20 and c63 (seeFig. 2c for an example) or cX3 and c36. Probe cX4 and c76 hybridization signals were not separable in cohybridized metaphase orprometaphase spreads. Signal at the median site (overlapping the gap)was observed with all cosmids, but the incidence varied significantlyamong the probes, ranging from 7.14% for probe 19-20 to 33.34% forprobe c76 (see Table 1 for summary).
The data presented in Table 1 are represented graphically in Fig. 3.The difference between the percentages at proximal and distal siteswas plotted as an absolute value for each probe (Fig. 3, line), togetherwith the values representing percentages of signals at the median site(Fig. 3, columns). Thus, probes 19-20 and cX3, and probes c36 andc63 flank, proximally and distally, respectively, the ape-induced gapsor breaks at FRA3B. On the other hand, probe cX4 and particularlyc76 contain sequences crossing the fragile sites, and the signal gencrated with these probes could fall on either side of the breakpoint atFRA3B. This analysis delineates the FRA3B to a region of a fewhundred kb between FHIT exon 3 (cosmid cX3) and FHJT intron 5(c36), with the majority of gaps occurring in introns 4 and 5, flankingthe first FHIT protein-coding exon.
DISCUSSION
By FISH analysis of chromosomes derived from normal lymphocytes treated with ape, we localized the majority of chromosome gaps,collectively known as FRA3B, to a region telomeric to the breakpointof the t(3;8) translocation in RCC, encompassing the region involvedin deletions in tumor cell lines and bordered distally by a portion ofintron 5 of the recently identified FH!T gene (29). In fact, the majorityof gaps map just to either side of FHJT exon 5, the exon that is mostfrequently lost in cancer cells with homozygous deletions in the FHJTgene (30).
Rare folate-sensitive fragile sites that have been cloned (FRAXA,FRAXF, FRAJ6A, and FR@411B)involve unstable expansion of CCGtrinucleotide repeats (38—42),and their expression is mediated by anexcessive copy number of CCG repeats. Regions of 3pl4.2 thus fartested do not contain CCG repeats (21, 28), suggesting that thedistinction between constitutive and heritable fragile sites is not onlyin inducing agent and frequency of expression within the population,but also in molecular structure and the mechanisms mediating theirexpression.
Several groups of investigators have made important contributionsto our current understanding of the fragile region at 3pl4.2. A groupof investigators studying FRA3B-associated break points induced inhamster-human hybrids (21, 22, 37) has most recently shown that twoclusters of aphidicolin induced hybrid break points, both telomeric tothe t(3;8) 3pl4.2 break, are separated by more than 200 kb and flankan Miul site estimated to be —185kb from t(3;8). We have mappedthis M!ul site within intron 4, perhaps —100kb centromeric to exon5 (see Fig. 1); thus, the two hybrid break point clusters flank FH!T
1168
a b
C
d eFig. 2. FISH localization of cosmid probes at FRA3B on metaphase chromosomes
derived from ape-treated normal peripheral lymphocytes. Partial and complete metaphasespreads with fluorescent hybridization signal located proximal (a; probe cX3), distal (d;probe c36), and on both sides (c; probes c19-20 red, rhodamine, and c63 green; FITCdetection) on the FRA3B. 4',6'-diamidino-2-phenylindole-generated chromosome banding (b and e) of the labeled spreads permits an accurate localization of the fluorescentsignal relative to the gaps and breaks at FRA3B.
(centromeric), and distal (telomenc) to the gaps or constrictionsmarking the fragile sites, in a minimum of 60 metaphases with4',6'-diamidino-2-phenylindole enhanced G-like banding (Fig. 2; Table 1). Probes 19-20 and cX3 hybridized predominantly proximal to
on March 17, 2014. © 1997 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

Table 1Distribution (percentage) offluorescent signals generated by cosmid probes spanning theFRA3BProbeNaN
(%) P N (%) D N (%) M(P -D)19-208478
(92.86) 0(0.00) 6(7.14)92.86cX36248(77.42) 2 (3.83) 12(19.35)73.59cX48860(68.18) 0 (0.00) 28(31.82)68.18c766614(21.21)
30(45.45)22(33.34)24.24c36760(0.00) 64 (84.21) 12(15.79)84.21c63
a Number of metaphase72 spreads examined0
(0.00) 60 (83.33) 12 (16.66)
; P. proximal; D, distal; M, medial; P —D, percentage proximal minus percentage distal in absolute numbers.83.33
100
90
80
70
60
50
40
30
20
10
0
POS@ONSOFFRA3BSITESWITHINThEFHJTGENE
exon 5. In fact, the cluster of fragile sites represented by 10 hybridbreaks in the proximal cluster map very close to FHJT exon 4, asshown in Fig. 1 by the position of markers D3S4260 and IA6GIH. Wewere also able to determine that the HPV16 integration site (21, 37)falls in FHJT intron 4 as shown in Fig. 1; integration of HPV16 in thissite resulted in a large deletion (21) in the cervical carcinoma FlITlocus, which included exon 5. Interestingly, our own cl3 humanhamster hybrid with a de novo break in 3pl4.2 (35, 36) also falls inintron 4, probably not far telomeric to the HPV16 integration site andbetween the two hybrid breakpoint clusters defined by Paradee et a!.(37).
A second group of investigators has been isolating DNA sequencesfrom FRA3B by cloning plasmid integration sites (27) and sequencingand mapping the flanking regions (28). This group had shown previously that, under conditions of fragile site induction, the pSV2neoplasmid preferentially integrates into chromosomal regions harboringfragile sites (27). These investigators reported isolation and sequencing of two plasmid integration sites that they mapped to the telomericend ofthe 850A6 YAC, which carries the t(3;8) break and the FRA3B.We have mapped one of the sequences flanking the centromeric sideof a plasmid integration, U40597, to 4 kb telomenc to FHIT exon 5.Additionally, a sequence cloned by this group (GenBank accessionno. U60203), encompassing one side of a tumor cell deletion, maps inintron 5, —100kb from exon 5, as shown in Fig. 1. Thus, the clonedfragile sites probably fall within a region ofless than 200 kb, flankingFHIT exon 5, the protein coding exon most frequently homozygouslydeleted in cancer cell lines. It also follows that the cosmids c36 andc63 in intron 5 are telomeric to many of the identified fragile sites, inagreement with our finding that these probes are almost always foundby FISH to be distal to the gaps induced in 3pl4.2. Wilke et a!. (21)
and Rassool et a!. (28) have also performed FISH analyses usingprobes from regions near their identified fragile sites, and their resultsare largely in agreement with ours. Interestingly, c76 and cX4 cosmids, containing exons 5 and 4, respectively, can be proximal or distalto the gaps or can overlap the gaps, as summarized in Fig. 3. Mostgaps occur within cosmid c76, the exon 5-containing cosmid, mirroring the findings in tumor cells (30), but a number of gaps fall withinthe exon 4 cosmid and even in the exon 3 cosmid, two cosmids thatflank the t(3;8) break, suggesting that the t(3;8) translocation, as wellas the loss and rearrangement of FHIT exons 3 and 4 observed incancer cells (30), could have derived from breaks in sequences that are“fragile,―although the actual fragile sequences, the bases for breaksand gaps in this region, have not yet been identified.
Fig. 3. Graphical representation of the data presented in Table 1 (see “Results―fordetails). Difference between the percentages of fluorescent signals at proximal and distalsites was plotted as an absolute value for each probe (line), together with the valuesrepresenting percentages of signals at the median site (columns).
1169
REFERENCES
FRA3B Probe Distribution
1. Glover, T. W., Berger, C., Coyle, J.. and Echo, B. DNA polymerase a inhibition byaphidicolin induced gaps and breaks at common fragile sites in human chromosomes.Hum. Genet., 67: 136—142,1984.
2. Sutherland, G. R. Chromosomal fragile sites. Genet. Anal. Tech. Appl., 8: 161—166,1991.
3. Sutherland, G. R., and Ledbetter, D. H. Report of the committee on cytogeneticmarkers. Cytogenet. Cell Genet., 51: 452—458,1989.
4. Hecht, F. Fragile sites, cancer chromosome breakpoints, and oncogenes all cluster inlight G bands. Cancer Genet. Cytogenet., 31: 17—24,1988.
5. Manuelidis, L. A view of interphase chromosomes. Science (Washington DC), 250:1533—1540,1990.
6. Yunis, J. J., and Soreng, A. L. Constitutive fragile sites and cancer. Science(Washington DC), 226: 1199—1204, 1984.
7. Le Beau, M. M., and Rowley, J. D. Heritable fragile sites and cancer. Nature (Lond.),12: 607—608,1984.
8. Popescu, N. C., Zimonjic, D., and DiPaolo, J. A. Viral integration, fragile sites andprotooncogenes in human neoplasia. Hum. Genet., 84: 383—386,1989.
9. Popescu, N. C. Chromosome fragility and instability in human cancer. Crit. Rev.Oncog., 5: 121—140,1994.
10. Yunis, J. J., Soreng, A. L., and Bowe, A. E. Fragile sites are targets of diversemutagens and carcinogenesis. Oncogene, 1: 59—69,1987.
11. Cohen, A. J., Li, F. P., Berg, S., Marchetto, D. J., Ted, S., Jacobs, S. C., and Brown,R. S. Hereditary renal-cell carcinoma associated with a chromosomal translocation.N. Engl.J.Med.,301:592—595,1979.
12. Wang, N., and Perkins, K. L. Involvement of band 3pl4 in hereditary renal carcinoma. Cancer Genet. Cytogenet., 11: 479—481, 1984.
@ 13. Glover, T. W., Coyle-Morris, J. F., Frederick, P. L., Brown, R. S., Berger, C. S.,
@‘N@ Gemmill,R.M.,andHecht,F.Translocationt(3;8)(pl4.2;q24.l)inrenalcell@@ carcinomaaffectsexpressionofthecommon fragilesiteat3pl4(FRA3B)inlym
‘N,. phocytes.CancerGenet.Cytogenet.,31:69—73,1988.14. Sutherland, G. R. Fragile sites and cancer breakpoints: the pessimistic view (Edito
rial). Cancer Genet. Cytogenet., 31: 5—7,1988.15. Smeets, D. F. C. M., Scheres, J. M. J. C., and Hustinx, T. W. J. The most common
fragile site in man is 3pl4. Hum. Genet., 72: 215—220,1986.16. Sandberg, A. A. The Chromosome in Human Cancer and Leukemia, Ed. 2. New
York: Elsevier Science Publishing Co., Inc., 1990.17. Solomon, E., Borrow, J., and Goddard, A. D. Chromosome aberrations and cancer.
Science (Washington DC), 254: 1153—1160, 1991.18. Rabbitts, T. H. Chromosomal translocations in human cancer. Nature (Land.), 372:____________ 143—149,1994.19. Popescu, N. C., Zimonjic, D. B., Levinton-Kriss, S., Bryant, J. L., Lunardi-Iskander,
Y., and Gallo, R. C. Deletion and translocation involving chromosome 3 (p14) in twoI@ , _i@ I I I I tumorigenicKaposi'ssarcomacelllines.J.Nail.CancerInst.,88:450—455,1996.
20. Popescu, N. C., DiPaolo, J. A., and Amsbaugh, S. C. Integration sites of human19 X3 X4 76 36 63 papillomavirus18 DNA sequencesof Hela cell chromosomes.Cytogenet.Cell
PROBES Genet., 44: 58—62,1987.21. Wilke, M. C., Hall, B. K., Hoge, A., Paradee, W., Smith, D. I., and Glover, T. W.
FRA3B extends over a broad region and contains a spontaneous HPV integration site:direct evidence for the coincidence of viral integration sites and fragile sites. Hum.Mol. Genet., 5: 187—195,1996.
22. Wilke, C. M., Guo, S-W., Hall, B. K., Boldog, F., Gemmill, R. M.,
on March 17, 2014. © 1997 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from

POSITIONS OF FRA3B SITES WITHIN THE FHIT GENE
Chandrasekharappa, S. C., Barcroft, C. L., Drabkin, H. A., and Glover, T. W.Multicolor FISH mapping of YAC clones in 3pl4 and identification of a YACspanning both FRA3Band the t(3;8) associated with hereditary renal cell carcinoma.Genomics, 22: 319—326, 1994.
23. Michaelis, S. C., Bardenheuer, W., Lux, A., Schramm, A., Gockel, A., Sieber, R.,Willers, C., Schmidtke, K., Todt, B., van der Hout, A. H., Buys, C. H. C. M.,Heppell-Parton,A.C.,Rabbius,P.H.,Ungar,S.,Smith,D.,LePaslier,D.,Cohen,D.,Opalka. B., and SchUtte, J. Characterization and chromosomal assignment of yeastartificial chromosomes containing human 3pl3—p2l-specific sequence tagged sites.Cancer Genet. Cytogenet., 81: 1—12,1995.
24. Boldog, F. L., Gemmill, R. M., Wilke, C. M., Glover, T. W., Nilsson, A-S.,Chandrasekharappa, S. C., Brown, R. S., Li, F. P., and Drabkin, H. A. Positionalcloning of the hereditary renal carcinoma 3;8 chromosome translocation breakpoint.Proc. Nail. Acad. Sci. USA, 90: 8509—8513, 1993.
25. Kastury, K., Ohta, M., Lasota, J., Moir, D., Dorman, T., LaForgia, S., Dnick, T., andHuebner, K. Structure of the human receptor tyrosine phosphatasc y gene (PTPRG)and relation to the familial RCC t(3;8) chromosome translocation. Genomics, 32:225—235,1996.
26. Paradee, W., Mullins, C., Zhanquan, H.. Glover, T., Wilke, C., Opalka, B., Schutte,J., and Smith,D. I. Preciselocalizationof aphidicolin-inducedbreakpointson theshon arm of human chromosome 3. Genomics, 27: 358—361,1995.
27. Rassool, F. V., McKeithan, T. W., Neilly, M. E., van Melle, E., Espinosa, R., and LeBeau, M. M. Preferential integration of marker DNA into the chromosomal fragilesite at 3pl4.2: a novel approach to cloning fragile sites. Proc. Nail. Aced. Sci. USA88: 6657—6661, 1991.
28. Rassool, F. V., Le Beau, M. M., Shen, M-L., Neilly, M. E., Espinosa R. Ill, Ong,S. T., Boldog, F., Drabkin, H., McCarroll, R., and McKeithan, T. W. Direct cloningof DNA sequences from the common fragile site region at chromosome band 3pl4.2.Genomics, 35: 109—I17, 1996.
29. Ohta. M., Inoue, H., Cotticelli, M. G., Kastury, K., Baffa, R., Palazzo, J., Siprashvili,z.,Mori,M.,McCue,P.,Dnick,T.,Croce,C.M.,andHuebner,K.TheFH1Tgene,spanning the chromosome 3pl42 fragile site and renal carcinoma-associated t(3;8)breakpoint, is abnormal in digestive tract cancers. Cell, 84: 587—597,1996.
30. Druck, T., Hadaczek, P., Fu, T-B., Ohta, M., Siprashvili, Z., Baffa, R., Negrini, M.,Kastury, K., Veronese, M. L., Rosen, D., Rothstein, J., McCue, P., Cotticelli, M. G.,Inoue, H., Croce, C. M., and Huebner, K. Structure and expression of the humanFHIT gene in normal and tumor cells. Cancer Res., 57: 504—512.
31. Wang, N-D., Tests. J. R., and Smith, D. I. Determination of the specificity ofaphidicolin-induced breakage of the human 3pl4.2 fragile site. Genomics, 17: 341—347, 1993.
32. Kuwano, A., and Kajii, T. Synergistic effect of aphidicolin and ethanol on theinduction of common fragile sites. Hum. Genet., 75: 75—78,1987.
33. Zimonjic, D. B., Popescu, N. C., Matsui, T., Ito, M., and Chihara, K. Localization of
the human cholecystokinin-B/gastrin receptor gene to chromosome I lplS.4-l5.5 byfluorescence in situ hybridization. Cytogenet. Cell Genet., 65: 184—185,1994.
34. Zimonjic, D. B., Rezanka, L., and Popeacu, N. C. Refined localization of the erbB-3protooncogene by direct visualization of FISH signals on LUT-inverted and contrastenhanced digital images of DAPI-banded chromosomes. Cancer Genet. Cytogenet.,80: 100—102,1985.
35. LaForgia, S., Morse, B., Levy, J., Bamea, G., Cannizarro, L A., Li, F., Nowell, P. C.,Boghosian-Sell, L., Glick, J., Weston, A.. Harris, C. C., Drabkin, H., Patterson, D.,Croce, C. M., Schlessinger, J., and Huebner. K. Receptor protein-tyrosine phosphatase y is a candidate tumor suppressor gene at human chromosome region 3p21. Proc.Nail. Aced. Sci. USA, 88: 5036—5040, 1991.
36. LaForgia,S., Lasota,J., Latif,F., Boghosian-Sell,L., Kastury,K.,Ohta,M.,Druck,T., Atchison, L., Cannizzaro, L. A., Barnea, G., Schiessinger, J., Modi, W., Kuzmin,I.. Tory, K., Thar, B., Croce, C. M., Lerman, M., and Huebner, K. Detailed geneticandphysicalmapof the 3p chromosomeregionsurroundingthe familialrenalcellcarcinomas chromosome translocation, t(3;8Xp14.2;q24.l). Cancer Res., 53: 3118—3124, 1993.
37. Paradee. W., Wilke, C. M., Wang, L., Shiidhar, R., Mullins, C. M., Hoge, A., Glover,T. W., and Smith, D. I. A 350-kb cosmid contig in 3pl4.2 that crosses the t(3;8)hereditary renal cell carcinoma translocation breakpoint and 17 aphidicolin-inducedFRA3B breakpoints. Genomics 35: 87—93,1996.
38. Verkerk, A. J. M. H., Pieretti, M., Sutciffe, J. S., Fu, Y-H., Kuhi, D. P. A., PIZZUti,A., Reiner, 0., Richards, S., Victoria, M. F., Zhang, F., Eussen, B. E., van Ommen,G-J. B., Blonden, L A. J., Riggens, G. J., Chastain, J. L, Kunst, C. B., Galjaard, H.,Caskey, C. T., Nelson, D. L., Oostra, B. A., and Warren, S. 1. Identification ofa gene(FMR-I) containing a CGG repeat coincident with a breakpoint cluster regionexhibiting length variation in fragile site syndrome. Cell, 65: 905—914,1991.
39. Knight, S. J. L, Flannery. A. V., Hirst, M. C., Campbell, L, Middleton-Piice, H. R.,Barnicoat, A., Pembrey. M. E., Holland, J., Oostra, B. A., Bobrow, M., and Davies,K. E. Trinucleotiderepeatamplificationand hypermethylationof a CpG islandinFRAXE mental retardation. Cell, 74: 127—134,1993.
40. Famish,J. E., Oostra,B. A., Verkerk,A. J. M. H., Richards,C. S., Reynolds,J.,Spikes, A. S., Stuffer, L G., and Nelson, D. L Isolation of a CGG repeat showingexpansion in FRAXF, a fragile site distal to FRAXA and FRAXE. Nat. Genet., 8:229—235,1994.
41. Nancarrow, J. K., Kremer, E., Holman, K., Eyre, H., Doggett, N. A., LePaslier, D.,Callen, D. F., Sutherland, G. R., and Richards, R. I. Implications ofFRAI6A structurefor the mechanism of chromosomal fragile site genesis. Science (Washington DC),264: 1938—1941,1994.
42. Jones, C., Slijepcevic. P., Marsh, S., Baker, E., Langdon, W. Y., Richards, R. I., andTunnacliffe, A. Physical linkage of the fragile site FRA1IB and the Jacobsensyndrome chromosome deletion breakpoint in I 1q23.3. Hum. Mol. Genet., 3: 2123—2130, 1994.
I170
on March 17, 2014. © 1997 American Association for Cancer Research.cancerres.aacrjournals.org Downloaded from




















