
Population Structure in Zamia debilis (Zamiaceae). I. Size Classes, Leaf Phenology, and Leaf...
14
Population Structure in Zamia debilis (Zamiaceae). I. Size Classes, Leaf Phenology, and Leaf Turnover Vivian Negrón-Ortiz; Gary J. Breckon American Journal of Botany, Vol. 76, No. 6. (Jun., 1989), pp. 891-900. Stable URL: http://links.jstor.org/sici?sici=0002-9122%28198906%2976%3A6%3C891%3APSIZD%28%3E2.0.CO%3B2-2 American Journal of Botany is currently published by Botanical Society of America. Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available at http://www.jstor.org/about/terms.html. JSTOR's Terms and Conditions of Use provides, in part, that unless you have obtained prior permission, you may not download an entire issue of a journal or multiple copies of articles, and you may use content in the JSTOR archive only for your personal, non-commercial use. Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained at http://www.jstor.org/journals/botsam.html. Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printed page of such transmission. The JSTOR Archive is a trusted digital repository providing for long-term preservation and access to leading academic journals and scholarly literature from around the world. The Archive is supported by libraries, scholarly societies, publishers, and foundations. It is an initiative of JSTOR, a not-for-profit organization with a mission to help the scholarly community take advantage of advances in technology. For more information regarding JSTOR, please contact [email protected]. http://www.jstor.org Mon Oct 29 14:38:17 2007
Transcript of Population Structure in Zamia debilis (Zamiaceae). I. Size Classes, Leaf Phenology, and Leaf...

Population Structure in Zamia debilis (Zamiaceae). I. Size Classes, Leaf Phenology,and Leaf Turnover
Vivian Negrón-Ortiz; Gary J. Breckon
American Journal of Botany, Vol. 76, No. 6. (Jun., 1989), pp. 891-900.
Stable URL:
http://links.jstor.org/sici?sici=0002-9122%28198906%2976%3A6%3C891%3APSIZD%28%3E2.0.CO%3B2-2
American Journal of Botany is currently published by Botanical Society of America.
Your use of the JSTOR archive indicates your acceptance of JSTOR's Terms and Conditions of Use, available athttp://www.jstor.org/about/terms.html. JSTOR's Terms and Conditions of Use provides, in part, that unless you have obtainedprior permission, you may not download an entire issue of a journal or multiple copies of articles, and you may use content inthe JSTOR archive only for your personal, non-commercial use.
Please contact the publisher regarding any further use of this work. Publisher contact information may be obtained athttp://www.jstor.org/journals/botsam.html.
Each copy of any part of a JSTOR transmission must contain the same copyright notice that appears on the screen or printedpage of such transmission.
The JSTOR Archive is a trusted digital repository providing for long-term preservation and access to leading academicjournals and scholarly literature from around the world. The Archive is supported by libraries, scholarly societies, publishers,and foundations. It is an initiative of JSTOR, a not-for-profit organization with a mission to help the scholarly community takeadvantage of advances in technology. For more information regarding JSTOR, please contact [email protected].
http://www.jstor.orgMon Oct 29 14:38:17 2007

Amer. J. Bot. 76(6): 891-900. 1989.
POPULATION STRUCTURE IN ZAMIA DEBILIS (ZAMIACEAE) I. SIZE CLASSES, LEAF PHENOLOGY, AND LEAF TURNOVER1
Departamento de Biologia, Universidad de Puerto Rico, Recinto Universitario de Mayagiiez, Mayagiiez, Puerto Rico 00709-5000
A B S T R A C T
Population structure, leaf phenology and leaf turnover were followed over a 29-month period in Zarnia debilis L.f. ex Aiton (Zamiaceae), an understory species in the Cambalache Forest in northern Puerto Rico. It was not possible to determine plant age or to measure the subterranean stems; size classes based on leaf number and leaf x leaflet number indices were used to determine population structure. Despite seasonal and year to year fluctuations in leaf number at the individual and population level, population profiles remained relatively constant. At any one time, over 50% of the population was composed of unbranched individuals with one or two leaves. Only 7% of the plants were branched. Plants with seven or more leaves comprised at a maximum 8% of the population, but accounted for 28% of the total foliage. Size classes based on leaf number and on a leaf x leaflet index gave approximately reverse J-shaped curves typical of trees with shade tolerant seedlings and saplings. New leaves emerged throughout the year, with a peak at the beginning of the rainy season in May or June and lowest production during the dry months of February through April. .4verage leaf life expectancy was approximately 2.3 years. Leaf death occurred over an extended period of time by the loss of individual leaflets. Patterns in leaf production and loss differed between few- and many-leaved plants. On the average, as the number of mature leaves on a plant increased, time between emergence of new leaves decreased. In many-leaved plants more than one event of new leaf emergence per year was common. Individuals with one to three mature leaves and individuals with four or more mature leaves differed in their response to water stress: few-leaved plants generally reduced the rate of new leaf production and retained old leaves longer. Plants with more than three leaves continued to produce new leaves, but the rate of leaf mortality increased so that most had a net leaf loss. There was no evidence that leaf emergence or retention were affected by cone production or seed maturation.
THEREIS LITTLE PUBLISHED On leaf phenology Chamberlain (I 935) and Stevenson (1 98 I) note for cycads growing in nature; most reports are that cone production may inhibit leaf produc- based on cultivated plants (Chamberlain, 1935; tion the following year, and Tang (1986) states Stevenson, 198 1). Chamberlain (1935) states that leaf loss may occur in plants maturing that leaf production in cycads remains more seeds. or less constant every year and that in Dioon Life span in evergreen species, i.e., species edule Lindl. and D. spinulosum Dyer a new that retain a cohort of leaves for more than crown of leaves is produced every other year. one year (Bell and Bliss, 1979) varies from 13 Tang (1986) gives a generalized diagram of to 14 months for most tropical trees (Longman cone and leaf phenology for Zamia pumila L. and Jenik, 1974; Richards, 1966) to 25 years in Florida, and Hallman (1976) reports that in Araucaria (Chabot and Hicks, 1982) and 45 the plants produce five to seven leaves an- years in Pinus longaeva (Ewers and Schmid, nually. With the exception of Stangeria, which 198 1). From Chamberlain's (1 93 5) data, one produces leaves one at a time throughout the can calculate that the leaf life span for D. edule year, cycads in cultivation generally produce is two years. There are no other references to an annual flush of leaves (Stevenson, 1981). leaf life span for cycads (see Read and Solt,
1 9 86, for general bibliography on cycads). Ecological research on cycads has been lim-
Received for publication 2 February 1988; revision accepted 4 January 1989. ited primarily to habitat and distribution de-
This study represents part of a thesis submitted in partial scriptions (for example: Dressler, 1975; De fulfillment of the requirements for the M.S. degree in Bi- Luca et al., 1980; De Luca, Sabato, and VSz- ology at the University of Puerto Rico, Mayagiiez. quez Torres, 1980, 1981a, b, 1982, 1984; Ste-
We wish to thank Drs. N. Dengler, D. Kolterman, A. Lewis, and R. Memll for critically reading the manuscript venson, 1982; Stevenson, Sabato, and VAzquez and making helpful suggestions. Cricket Yoshioka pre- Torres, 1986). Except for Chamberlain's (1 935) pared the computer-generated graphics. age estimate for an individual of Dioon, noth-
I

892 AMERICAN JOURNAL OF BOTANY [Vol. 76
ing is known about age or size structure in cycad populations.
The purpose of the present research was to determine leaf phenology, leaf turnover rates, and the population structure in a natural pop- ulation of Zamia in Puerto Rico. The correct specific epithet for the taxon studied remains confused. It has long gone under the name Za- rnia latifoliata Prenl. Eckenwalder (1980) in- cluded it, along with all the Caribbean popu- lations, in Z. pumila L. Using narrower species concepts, Liogier and Martorell (1 982) applied Z. debilis L.f. ex Aiton to the taxon. Stevenson (1 987) rejected both Z. latifoliata and Z. debilis for nomenclatural reasons and coined a new epithet, Z. amblyphyllidia for the broad-leaved Zamia from northern Puerto Rico. Despite the taxonomic interest in the Caribbean cycads, there are still problems of circumscription and typification to be resolved before the nomen- clature will be stabilized. For this reason, we prefer to use the more widely recognized name, Z. debilis, for now.
subtropical forests (Fig. 1). It occurs on lime- stone derived soils that are characterized by having a neutral to alkaline pH and being well- drained. The plants have a fleshy, turbinate underground stem (Fig. 2) which may be branched. It is not possible to use leaf scars for age estimates in Z. debilis, as has been done for palms (Bannister, 1970; Piiiero, SarukhSn, and GonzSlez, 1977) and for Dioon (Cham- berlain, 1935). Contractions by the root and stem (Stevenson, 1980) obliterate the bound- aries between leaf scars (Negr6n-Ortiz, 1987). The leaves are pinnately compound and may reach 1.5 m in length. Our preliminary and subsequent observations of over 25 excavated wild plants and numerous cultivated individ- uals support the assumption that plants with small stems have fewer leaves, smaller leaves, and fewer leaflets per leafthan plants with large stems. The only way we could estimate plant size without excavating the plants was by leaf and leaflet number, the latter being correlated with leaf length (Negrbn, 1987).
The plants are dioecious. Pollination occurs MATERIALSAND METHODS-D~SCY~P~~OY~of from mid-January through February, with the
site-The study was conducted in the Camba- lache Commonwealth Forest (ca. 18'26'30"N, 66'36'30") in the municipality of Arecibo on the north coast of Puerto Rico. The forest, which is on Karst Formation, encompasses 3 80 ha and varies in elevation from five to 50 m above sea level (DRN, 1976).
Annual rainfall is reported to be 1,400 to 1,500 mm and seasonal in occurrence (DRN, 1976) with the rainy season beginning in May or June, and reaching a maximum during Sep- tember and October. Rainfall in November, December, and January is erratic, and pro- longed dry and wet periods may occur. Feb- ruary, March, and April are the dry months (DRN, 1976), and effects of water stress, i.e., leaf loss and wilting of foliage, can be seen in the forest during this time (personal observa- tion). The climatological data from the Cam- balache station (NOAA, 1982-1 984) is too in- complete to use for purposes of quantitative correlation. Therefore, the climatic conditions described for the two-year study are based on our own observations.
Cambalache falls within Holdridge's Sub- tropical Moist Forest plant formation (Ewe11 and Whitmore, 1973). Although the majority of the species are evergreen (DRN, 1976), the canopy is more open during the dry season, when leaf litter accumulates on the forest floor (personal observation).
Description of species -In Puerto Rico Za- rnia debilis grows in the understory of lowland
seeds maturing between seven and 11 months later (Breckon and Negr6n-Ortiz, 1983; Ne- gr6n-Ortiz, 1987).
Nonrandom permanent plots-Four adja-cent hillsides, each facing a cardinal compass point, were selected in a relatively isolated area in the forest. Between April 1982 and October 1982, five 1 m x 5 m plots were established on each hillside, with the long axis of the plot perpendicular to the slope. Plots were selected so as to be accessible while including a rea- sonable number of ramets (average of 18.3 ra- mets per plot, range 9-39). Each ramet was labelled and attachments between ramets were determined by careful, shallow excavation around closely proximal ramets. Excavation was done so as to limit disturbance to the plants and, to the best of our knowledge, no roots were broken in the process.
Markers were put next to individuals con- sidered to be seedlings from the 1982 seed crop. Plants were considered seedlings if their old seed coats were still attached or if their leaves were 10-12 cm long with only two to four leaflets.
Plots were visited at irregular intervals (av- erage 32 days between visits, range 9-1 21 days) and ramets scored for the following: number of new, mature and dead leaves, number of leaflets on the largest leaf, and cone production and status. A leaf was consider dead after all of the leaflets were shed from the persistant rachis.

p g s g q s ou SEM aJayL -(p .%!A) s1msm1 mop -m ay) p m slo~d wopm~uou a q ur sarai~ad ml!w~s paMoys 'ppwlpul lad m p n u pa1 Lq paxapu? st! 'sa~n)Dw)s uogapdod 3!113~
esay3mq Jnoj jo auo pue saq3neJq aaJq jo XIS 'say3ua~q OM) jo pa)sIs -uo3 91 '£2 asaw jo -paq3ua~q alaM way1 jo %6'9 JO £2 Lpo 'slo~d wopua~uou aq) ur siuald S C ~ ayl jo -Sanaa1 Maj y i y siuvld payaumq -un Lq paleupop s e ~ a q 3 a p q w ~ la uog -apdod v!?!utv~ aqL-a.mim.us ~0?l~lndOd
'(~861 6 '£861 3% OZ '£861 S ~ V 02 '£861 unf 0€ '2861 3- ~2 "%'a) syaad pa1al)ms awos y l p ~ '~aad a q lnoy%no~y) paunmo qeap j e q JO p d v y%no~y) hraruqad woy a~ua%lawa pa)yg h r a ~ y ) y sr~)uow lawwns ayi %uunp payaad uorynpo~d jaaI '~eaL a q )noy%no~yl pa%~awa saAaal Mau a p w 3861 1aq0130 y%no~y) 2861 1udv WOJj p0uad ylUOUI-62 I? JaAO y)eapjaal pule aua%Jawajaaljo Bo1ouayd ay) s ~ o y s am% y - Doloua ydJva7- s ~ ~ n s q
.pap.103a~ osla s e ~ s%uqpaas jo Jaqurnu ayL 'laha1 IawaJ ay) )a auop SEM %uqd -wes 'pay3m)e aJaM spnppupq asop lay)ayM %uyuluua)ap uy L ~ p g ~ p ay) 01 ana . s n l s p m x x p n u auo3 p m 'pal isa8uol a¶) uo sial -3aa~jo laqwnu 'sa~aaljo Jaqwnu :pasual) ayl u! IanppIpur y3oa JOJ paJo3s aJaM % u y o ~ ~ o j
aqL -1msna~1 auo n e q aJow ur pa.un3m JaAa pnpppu! ou )nq 'do) a q )a aZba~uo3 o) papuai spasm) a q adays ado~s 01 ana -(w 26 o) pz a%m) walaw 8-sp SEM y)%ua~ pasum) a%t?la~a ayL 'lu~od %ugmts snopia~d a jo walaw OM) u g ) p ~ aq pp-103 lured %u!pms ou )ay) UO!SMOJ~
ayl y l p ~ ado~s lad pawalas aJaM s)u!od 8up.113~ wopua~ aAg .~saay~nos-y~nos %ur3ej Jaylo ayl '~sa~ypou-ypou %upaj auo 'sapis11ry %ursoddo OMIJO y3aa uo do) 01 wouoq w o ~ j apaw aJaM 'apy w I y3aa 'spasum) aAy '£86 1 hrmuef pz -L I jo yaaM ay) %uuna .slo~d ay) y1p~ asoy) 01 1ua3arpa sapIsI1ry OM) uo apew s e ~ aldwas uoyqndod u ~ o p u a ~ a 'a~n1m.q~ uo~)elndod ayl paldwes stold luauemad pal3alas ay) IIaM M O ~ auruualap 01 laplo UI - si~asuv.ti uopuvy
epeap pa~aprsuoa s a ~ jaaI ayl q 3 g ~ UI auo aylo) sno!aa~d snsua3 a q q ~uasald saAaa1 % q ~ y jo ~aqwnu ay) )su~&a palo3s SEM L I ! ~ -low j a q .paJalunoDua lug s a ~ a%agoj Mau s)! uayM snsua3 a $a Ianp!Arpur m uo )uasa~d saaaaI amlawjo Jaqwnu ay) )supBe palox s a ~ uoy3npo~d jeaI 'puo3as '1786 1 Ja90130-£86 I ~aqo130 '£86 I ~aq0130-~86 I ~aq0130 ''a'! 'pouad L11oaL y3ea %uunp saAeaI plo jo y)aap ayllo saAaa1 Maujo uo!)3npo~d ayl JOJ pa.xo3s aJaM slenpr~g)u! '£861 pue 2861 JOJ sasns - u a ~aqo130 ayl u~ luasa~d saneal jo Jaqwnu ayl y ~ y % u p q s 'ISJIJ :SLEM OM) UI paupual -ap aJaM Li!~truow joal put? uog3npold j o q
-031
894 AMERICAN JOURNAL OF BOTANY [Vol. 76
a A M J J A S O N D J F M A M J J A S O N D J F M A M J J A S O
MONTH
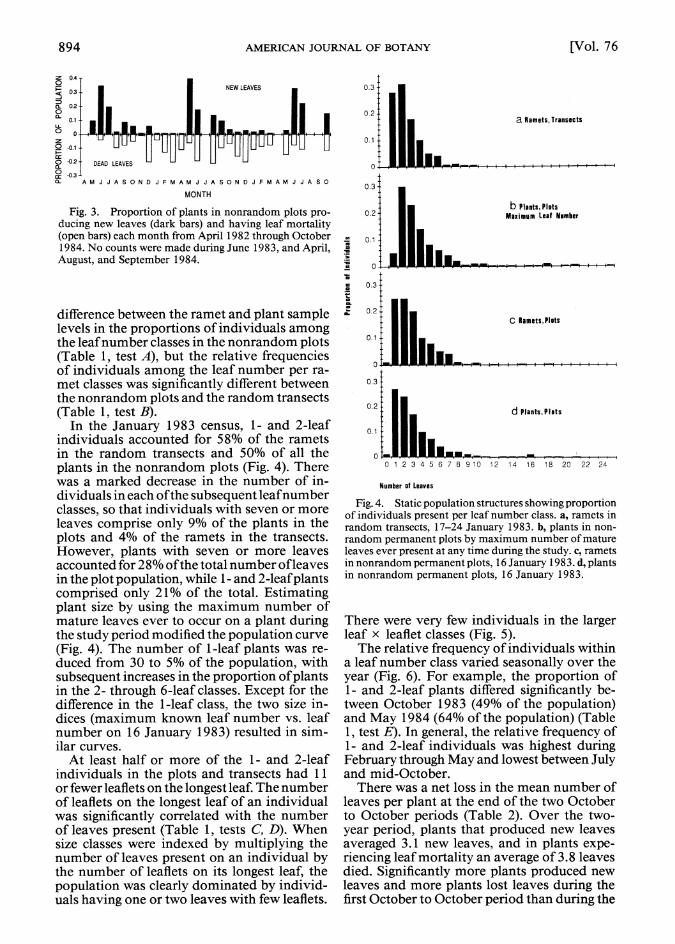
Fig. 3. Proportion of plants in nonrandom plots pro- ducing new leaves (dark bars) and having leaf mortality (open bars) each month from April 1982 through October 1984. No counts were made during June 1983, and April, August, and September 1984.
difference between the ramet and plant sample levels in the proportions of individuals among the leaf number classes in the nonrandom plots (Table 1, test A), but the relative frequencies of individuals among the leaf number per ra- met classes was significantly different between the nonrandom plots and the random transects (Table 1, test B).
In the January 1983 census, 1- and 2-leaf individuals accounted for 58% of the ramets in the random transects and 50% of all the plants in the nonrandom plots (Fig. 4). There was a marked decrease in the number of in- dividuals in each of the subsequent leafnumber classes, so that individuals with seven or more leaves comprise only 9% of the plants in the plots and 4% of the ramets in the transects. However, plants with seven or more leaves accounted for 28% of the total number ofleaves in the plot population, while 1- and 2-leaf plants comprised only 21% of the total. Estimating plant size by using the maximum number of mature leaves ever to occur on a plant during the study period modified the population curve (Fig. 4). The number of I-leaf plants was re- duced from 30 to 5% of the population, with subsequent increases in the proportion of plants in the 2- through 6-leaf classes. Except for the difference in the 1-leaf class, the two size in- dices (maximum known leaf number vs. leaf number on 16 January 1983) resulted in sim- ilar curves.
At least half or more of the 1- and 2-leaf individuals in the plots and transects had 11 or fewer leaflets on the longest leaf. The number of leaflets on the longest leaf of an individual was significantly correlated with the number of leaves present (Table 1, tests C, D). When size classes were indexed by multiplying the number of leaves present on an individual by the number of leaflets on its longest leaf, the population was clearly dominated by individ- uals having one or two leaves with few leaflets.
b plants, tots Maximum Leaf Number
11 0 1
v c 0
Number of Leaves
Fig. 4. Static population structures showing proportion of individuals present per leaf number class. a, ramets in random transects, 17-24 January 1983. b, plants in non- random permanent plots by maximum number of mature leaves ever present at any time during the study. c, ramets in nonrandom permanent plots, 16January 1983. d, plants in nonrandom permanent plots, 16 January 1983.
There were very few individuals in the larger leaf x leaflet classes (Fig. 5).
The relative frequency of individuals within a leaf number class varied seasonally over the year (Fig. 6). For example, the proportion of 1- and 2-leaf plants differed significantly be- tween October 1983 (49% of the population) and May 1984 (64% of the population) (Table 1, test E). In general, the relative frequency of 1- and 2-leaf individuals was highest during February through May and lowest between July and mid-October.
There was a net loss in the mean number of leaves per plant at the end of the two October to October periods (Table 2). Over the two- year period, plants that produced new leaves averaged 3.1 new leaves, and in plants expe- riencing leaf mortality an average of 3.8 leaves died. Significantly more plants produced new leaves and more plants lost leaves during the first October to October period than during the

June 1 9 8 91 NEGRON-ORTIZ AND BRECKON -POPULATION STRUCTURES IN ZAMIA 895
second period (Table 1, tests F and G; Table 2). The majority (59%) of the plants produced new leaves both years, and half of them (5 1 %) lost leaves both years (Table 3). Only 5% failed to produce a new leaf, and 5% failed to lose at least one leaf during the two-year period (Table 3). One plant was static, experiencing neither leaf production nor leaf mortality. The average number of leaves produced and/or lost by plants experiencing leaf production or loss was not significantly different between the two Oc- tober to October periods (Table 1, tests H, I). No relationship was evident between leaf turn- over and the production and maturation of cones (Table 1, test J).
The frequency of new leaf production was significantly different among the plants, de- pending on the number of mature leaves pres- ent at the time of emergence (Table 1, test K). Plants having no leaves and plants with seven or more mature leaves had a higher occurrence of new leaves emerging than did plants in the other leaf number classes (Table 1, tests L, M, Table 4, rows A, B). In general, the number of times that a plant produced new leaves in- creased with increasing maximum leaf number (Table 4, row N). Thus, in larger plants more than one episode of leaf emergence per plant per year was not uncommon.
On a yearly basis there was no statistical difference among leaf number classes as to whether new leaves were produced (Table 1, test N) although the low rate of production during year two by the 1 - and 2-leaf classes is noteworthy (Table 4, rows G, H). The number of new leaves produced per individual tended to be directly related to the number of mature leaves present at the time of production (Table 4, row C) and to the number of leaves present at the beginning of the year (Table 4, row I).
Calculated average leaf life expectancy among the leaf number classes varied from 1.8 to 3 years, averaging approximately 2.28 years over the two-year period (Table 4, row L). There was a statistically significant difference among the leaf number classes as to the frequency of leaf death on a per census basis (Table 1, test 0; Table 4, row E) or over each October to October period (Table 1, test P; Table 4, row J). The frequency of leaf mortality (Table 4, row E), as well as the average number of dead leaves at a census (Table 4, row F), or per year (Table 4, rows J, K), were correlated with the number of leaves present on the plant.
At the end of the two October to October periods, plants starting with two or more leaves had more individuals with a net leaf loss than a net gain (Table 5). Further, as shown in Table
~ L E1. Null hypothesis and test results for population structure and leafproduction and loss. Tests and tables used are from Rohlfand Sokal(1981) and Sokal and Rohlf(1981). Year one, October 1982-October 1983; year two, October 1983-October 1984. Asterisks in- dicate rejection of the null hypothesis at the following levels ofsign$cance: *= 0.05, **= 0.01, *** = 0.001; ns indicates that the null hypothesis could not be re- jected at the 0.05 level ofsign$cance
Ramet and plant frequencies were the same among the leaf number classes in the nonrandom plots, 16 January 1983; G = 10.4014 ns, ~~,. , ,(10) = 18.307. The frequency oframets among the leaf number classes was the same in the nonrandom plots and random transects in January 1983; G = 24.6264**, ~~ , , , , (10 ) = 23.209, x2,005(10) = 25.188. The number of leaflets on the longest leaf of a ramet was independent of the number of leaves on the ramet (nonrandom plots, 16 January 1983); F = 236.83**, F,,,, = 6.63, multiple r = 0.63 1. The number of leaflets on the longest leaf of a ramet was independent of the number of leaves on the ramet (random transects, January 1983); F= 3 1 1.37**, F,,,, = 6.63, multiple r = 0.578. The total number of 1- and 2-leaf plants during Oc- tober 1983 and May 1984 is not significantly different between the total number of plants having three or moreleaves. G = 10.563**,x2 ,,,, (1) = 7.879;x2,,,,, (1) = 10.828. There was no difference in the number of plants that produced leaves in year one vs. year two; G = 12.93***, x20001(l)= 10.8. There was no difference in the number of plants that experienced leaf mortality in year one vs. year two; G = 5.69*, ~ ~ ~ ~ ~ ( 1 ) 3.841.=
There was no difference in the mean number of leaves produced by plants producing leaves in year one vs. year two; t,,, = 1.2 ns, to,,= 1.645. There was no difference in the mean number of leaves dying in plants experiencing leaf mortality in year one vs. year two; t,,, = 0.606 ns, t o , = 1.645. Leaf production and mortality in fertile ramets were the same as in sterile ramets in a matched paired comparison between ramets from the same plots and having simlar numbers of leaves and leaflets; G,,,, ,, ,,,ti,, ., ,, = 0.9416 ns; x2,,,,(1) = 0.2504 ns; G,,,,, s,,,
= 3.841. The frequency of new leaf production was indepen- dent of the number of mature leaves present at the time of emergence; G = 89.66***, x2,.,,,(7) = 24.322. The frequency of new leaf production was the same for plants with or without leaves; G = 30.03***, x20.00,(l)= 10.828. The frequency of new leaf production in plants having one through six leaves was the same as that in plants having seven or more leaves; G = 15.78***, x2,,,,,(1) = 10.828. Whether a ramet produced new leaves over a one- year period was independent of the number of mature leaves present at the beginning of the year; year one, G=10.783ns;year2,G=5.856ns;x2,,,,(6)= 12.592. The frequency of leaf mortality was independent of the number of leaves present at the previous census; G = 346.99***, x2,,,,(7) = 24.322. Whether a plant lost leaves during a one-year period was independent of the number of mature leaves pres-
5, the net loss of leaves increased as the leaf ent at the beginning of the year; year one, G = 42.980;
number class increased. year two, G = ,,,,x266.109; (6) = 22.458.

896 AMERICAN JOURNAL OF BOTANY [Vol. 76
LVS. X L F L E T S .
Fig. 5. Population structure as indexed by number of leaves on an individual times the number of leaflets on its longest leaf. Data from January 1983 census of random transects.
Total leaf loss followed by a dormant period occurred in 44 individuals and lasted for an average of 105 days. The longest period a plant remained dormant was 297 days. At the end of the study 13 plants were leafless; five of these had not produced leaves after 297 days and were considered dead.
DISCUSSION-Phenologyand leaf turnover- Leaf production in 2.debilis was in part cor- related with seasonal rainfall. The May peak of leaf emergence in 1983 followed the onset of the rainy season, while in 1982 and 1984 the first rains and leaf emergence were in June. Under cultivation in Florida, 2. pumila ex-hibits leaf emergence in late spring and early summer (Tang, 1986). Tang's data differ from those for the Cambalache population, where new leaves emerged throughout the summer rainy season and, at a much lower rate,
throughout the dry season. The leaf emergence pattern observed in 2.debilis is apparently intermediate between that in Stangeria, where leaves are produced throughout the year and that reported for most other cycads, where pro- duction is by a single annual or biennial flush of new leaves (Chamberlain, 1935; Foster and Gifford, 1974; Hallman, 1976; Stevenson, 198 1; Tang, 1986). In general, individuals of 2.debilis with one or two leaves only produced leaves once a year, while those with seven or more leaves produced leaves two or more times a year. However, we could not tell if each ep- isode of leaf emergence was preceded by the production of new leaf primordia or if we wit- nessed prolonged, episodic emergence from one meristematic event of primordia production.
Seasonality, water availability, and temper- ature (Piiiero et al., 1977; Jow, Bullock, and Kummerow, 1980; Kikuzawa, 1982), light (Bannister, 1970; Barbour, Burk, and Pitts, 1980, p. 305; Nilsen, 1986), nutrient stress (Bazzaz and Harper, 1977), and carbon ratios (Chabot and Hicks, 1982) have all been used to explain leaf life span. In both 1982 and 1983 the island experienced drought conditions, with a prolonged dry period beginning each year in November. During 1982 there was a wet period in late January and early February; in 1983 no such rainy period occurred and the forest ap- peared to be more stressed by the dry season. The two drier-than-normal years probably ac- count for the net loss of leaves in the popu- lation, with the difference in leaf turnover be- tween the two years probably due to an additive effect of two dry years plus a more prolonged period of water stress in the second year. Leaf mortality may be more correlated with water stress than is shown in Fig. 3, but data are needed at the leaflet level in order to determine whether there is a seasonal pattern to foliage
A F 4 J J A S N D J F M A M J J A S O N D J F M A M J J A S O MONTH
Fig. 6. Proportion of plants per leaf number class in nonrandom plots during the 29 months of the study. No counts were made during June 1983 and April, August, and September 1984.

897 June 1 98 91 NEGRON-ORTIZ AND BRECKON- POPULATION STRUCTURES IN ZAMIA
TABLE 2. Summary of annual leaf production and mortality from 21 October 1982 through 21 October 1983, and from 21 October 1983 through 21 October 1984. Production and loss data are given as mean number of leaves per plant excluding plants that did not produce new leaves or did not experience leaf mortality. Population N = 327 plants
Leaves present 2 1 Oct Leaves produced Oct to Oct Leaves lost Oct to Oct
Year P x SD A' -2 + SD iV P i SD
mortality. Although the entire leaf was the unit of leaf production, mortality occurred at the leaflet level, with individual leaflets dying and being shed from a leaf over a long period of time. One or two persistent leaflets were often retained for months before finally being lost, at which point the leaf, e.g., the persistent ra- chis, was considered dead. The pattern of leaf death we observed in 2.debilisis different from that we have seen in cultivated individuals of Cycas revoluta, C. circinalis, and species of Dioon, where the entire leaf dies and is shed as a unit.
In seasonal subtropical and tropical rain- forests, compound leaves are more abundant in the upper canopy and uncommon in the understory (Beard, 1944, 1955; Givnish, 1978). The understory habitat and pattern of leaf loss of 2.debilis does not fit Givnish's (1 978) adap- tive model for compound leaves as throwaway branches. The adaptative value claimed by Givnish for compound leaves in trees is two- fold: first, they allow for rapid vertical growth; second, shedding of the rachis reduces water loss. Givnish's (1 978) argument that the com- pound leaf in herbaceous perennials results in a "consolidation of support tissue" and per- mits "greater mechanical efficiency in covering space near a plant's stemM-although undoc- umented-is probably a more reasonable ex- planation for the adaptive value of the com- pound leaf in 2.debilis.
As was true for leaf phenology, leaf produc- tion and mortality were not uniform within the population, but varied according to leaf num- ber class. The higher frequency of leaf pro- duction by 0-leaf plants and the low rate of plants entering the 0-leaf class is logical: a leaf- less individual in an otherwise evergreen pop- ulation would be at a disadvantage, being un- able to photosynthesize, yet still having to respire (Janzen and Wilson, 1974). In most cases, the individuals of 2. debilis observed entering the 0-leaf class probably did so due to water stress. Those with enough stored food were capable of undergoing dormancy and later producing a new leaf. The five plants experi-
encing mortality were small individuals that had not reached reproductive maturity. All were 1- or 2-leaf plants with few leaflets. It might be argued that some of these were senescent individuals whose leaf and leaflet number had declined to values typical of juvenile plants. Such a decline, however, did not occur in the few cases where we have observed death in mature cultivated cycads, including 2.debilis. Rather, the last leaves produced were typical of mature individuals.
Few-leaved individuals and many-leaved individuals differed in their responses to water stress. The lower frequency of leaf production during the second year was restricted to the 1-, 2-, and 3-leaf classes, which along with the 4-leaf class had a lower leaf loss rate in year two. Compared to the larger leaf number classes, members of the smaller leaf number classes had longer leaf turnover times, longer periods between new leaf production, and rel- atively fewer individuals showing a net leaf loss at the end of two years. Thus, few-leaved individuals responded to the drought condi- tions by leaf hoarding, while many-leaved in- dividuals maintained leaf production but re- duced their total leaf number through increased leaf mortality.
Based on the patterns observed, we hypoth- esize that leaf production in 2.debilis is con- trolled by some threshold system: a certain level of stored photosynthate must be available before leaf production will occur. Such a trig- gering system is reasonable because the plants occur in dense shade, where daily photosyn- thetic rates are low.
TABLE3. Proportion of individual plants producing and losing leaves annually in nonrandom permanentplots, N = 324plants. Year one, October 1982-October 1983; year two, October 1983-October 1984
Both Neither Year one Year two years year
New leaves 0.24 0.12 0.59 0.05 Dead leaves 0.25 0.19 0.51 0.05

898 AMERICAN JOURNAL OF BOTANY [Vol. 76
TABLE4. Leafproduction and loss as a function of leaf number. Data from plants in nonrandom permanent plots. A-C, leafproduction per census grouped by the number of mature leaves present on the plant during census when new leaves occurred (April 1982-1 984). D-F, leaf mortality per census grouped by the number of mature leaves present on the plant in the census previous to the one when mortality was scored. The dzference in counts between rows A and D are due to the scoring methods. G-L, annual turnover as afunction of leaf number at beginning of year. In each pair of rows, upper row is for year one, October 1982-October 1983, and lower row of each pair is for year two, October 1983-October 1984. M-P, leaf emergence pattern as a function of the maximum number of mature leaves occurring on the plant during study
No. of leaves 0 I 2 3 4 5 6 7 + Leaf production per census
A. No. of plants encoun- tered in class during all censuses
B. Frequency of plants with new leaves
C. Average number of leaves produced
LRaf mortality per census D. No. of plants encoun-
tered in class during all censuses
E. Frequency of plants with dead leaves
F. Average no. of leaves lost
Annual leaf turnover G. No. of plants in class at
start of year H. Proportion producing
new leaves during year I. Average no. leaves pro-
duced per individual in row H
J. Proportion losing leaves during year
K. Average no. leaves lost per individual in row J
L. Average leaf life expec- tancy in years
Leaf emergence pattern M. No. of plants in class N. Average no. of leaf
emergence events per individual in 29 months
Leaf survivorship may be affected by nu- or abort cones and seeds in order to retain trients being preferentially shunted to deve- foliage (Negrbn-Ortiz, 1 98 7). loping fruits (Street and Opik, 1984). Tang (1986) stated that in female Zamia pumila Population structure -The similarity in pop- plants, especially those on poor soils, leaf loss ulation structure at the ramet level and at the and often dormancy may be associated with plant level in the nonrandom plots was no doubt expanding cones. The absence of any differ- due to the small number of branched individ- ences in leaf loss or leaf production between uals encountered. The ramet population mea- fertile and similar-sized sterile plants (Table 1, sured in the random transects was probably test F)at Cambalache may be due to satisfac- more indicative of the population structure of tory soil conditions. Alternatively, it may be Zamia in the forest as a whole. The selected that 2.debiliswill forego cone production and/ plots, which were located in areas with many

899 June 1 9 8 91 NEGRON-ORTIZ AND BRECKON-POPULATION STRUCTURES IN ZAMIA
TABLE5. Net change in the number of leaves per plant from October 1982 through October 1984. Data is given as the proportion of individual plants in each leaf number category
No. ofleavest'plant 21 Oct 1982
Net change as 0 1 2 3 4 5 6 7+ of Oct 1984 N = 3 N = 75 N = 90 N = 55 N = 30 N = 23 N = 18 N = 32
Zamia plants, resulted in also selecting for larg- er plants (Breckon and Negr6n-Ortiz, unpub- lished data).
Chamberlain (1935) related the increase in leaf number in cycads to age and size. Neither leaf scars nor absolute leaf number have proved to be useful for age measurement in Z . debilis (Negr6n-Ortiz, 1987). Between year and sea- sonal fluctuations reduced the accuracy of leaf number or leaf x leaflet number as an absolute measure of size for individual plants; however, they are reasonable estimators of relative be- low ground plant size and vigor. Reproduction in plants is more a function of size than age (Hickman, 1979; Willson, 1983), and in Z. debilis cone production increases with leaf number and with leaf x leaflet number (Ne- gr6n-Ortiz, 1987; Breckon and Negr6n-Ortiz, in preparation). But these indices must be used with caution: leaf number alone may not dis- tinguish between immature and mature plants. Some mature plants with cones had three or fewer leaves, a number typical for seedlings. The index classes based on leaf x leaflet num- ber are arbitrary, and individuals with an index of 100 or less included nearly half the fertile plants (Negr6n-Ortiz, 19 87).
The relative frequencies of individuals in the various leaf number or leaf x leaflet classes did give a reasonable description of the overall ~ o ~ u l a t i o nstructure. i.e.. numerous small in- di;iduals with few large individuals. Further- more, the curve remained relatively stable over time despite the individual fluctuations in leaf number.
Both indices showed reversed-J shaped
Sarukhin (1 980) considered the reverse-J curve to be typical of tropical trees whose seedlings and saplings can persist for long periods in the understory. Most emergent tropical trees have short-lived seedlings that are renewed at rel- atively frequent intervals and are lacking in intermediate stages (Sarukhin, 1980). Zamia debilis, which must be shade tolerant through- out its life and has seedlings with large food reserves (Negr6n-Ortiz and Breckon, 1985; Negr6n-Ortiz, 1987), better fits the syndrome reported for shade-tolerant trees. Because it cannot be determined how long an individual of Z . debilis can remain at a particular size, there is no evidence for a suppression stage such as occurs in the shade-tolerant, phase-gap species Prestoea montana (R. Graham) Nich- ols. (Bannister, 1970) and Astrocaryum mex- icanum Liebm. (Sarukhin, 1980). The lack of literature on size or age structure or dynamics in populations of tropical understory species makes any generalizations as to a typical shade- tolerant syndrome for the group unwarranted.
LITERATURE CITED
BANNISTER,B. A. 1970. Ecological life cycle of Euterpe globosa Gaertn. In T. H. Odum [ed.], A tropical rain forest. A study of irradiation and ecology at El Verde, Puerto Rico, B-299-314. Div. Tech. Inform. U.S. Atomic Energy Commission, Oak Ridge, TN.
BARBOUR,M. G., J. H. BURK,AND W. D. PITTS. 1980. Terrestrial plant ecology. Benjamin/Cummings, Men- lo Park, CA.
Buur, F, A,, AND J, =, 977, Demographic analysis of the growth of Linum usitatissimum, New ~ h y t o l . 78: 193-208.
BEARD,J. S. 1944. Climax vegetation in tropical Amer- curves (Barbour et al., 1908; Sarukhin, 1980). ica. ~co logy 25: 127-158.

900 AMERICAN JOURNAL OF BOTANY [Vol. 76
. 1955. The classification of tropical America K. 1982. Leaf survival of woody plants in KIKUZAWA, vegetation types. Ecology 36: 89-100. deciduous broad leaved forest. 1. Tall trees. Canad.
BELL, K. L., AND L. C. BLISS. 1979. Autecology of Ko- J. Bot. 61: 2133-2139. bresia gellardii: why winter snow accumulation limits LIOGIER, H. A., AND L. F. MARTORELL. 1982. Flora of local distribution. Ecol. Monogr. 49: 377-402. Puerto Rico and adjacent islands: a systematic syn-
BRECKON,G. J., AND V. NEGRON. 1983. Pollination of opsis. Editorial de la Universidad de Puerto Rico, Rio Zamiapumila by fungusgnats. Amer. J. Bot. 70: 106- Piedras, Puerto Rico. 107. LONGMAN,K. A,, AND J. JENIK. 1974. Tropical forest
CHABOT,B. F., AND D. J. HICKS. 1982. The ecology of and its environment. Longman, London. leaf life spans. Annual Rev. Ecol. Syst. 13: 229-259. NATIONAL OCEANIC ADMINISTRATIONAND ATMOSPHERIC
CHAMBERLAIN,C. J. 1935. Gymnosperms, structure and (NOAA). 1982-1 984. Climatological data. Puerto evolution. University of Chicago Press, Chicago. Rico and Virgin Islands.
DE LUCA, P., A. MORETTI, S. SABATO, AND M. V ~ Q U E Z NEGRON-ORTIZ,V. 1987. Study of natural history and TORRES. 1980. Dioon rzedowskii (Zamiaceae), a new species from Mexico. Brittonia 32: 225-229.
-, S. SABATO, TORRES. 1980. DioonAND M. V ~ Q U E Z caputoi (Zamiaceae), a new species from Mexico. Brit- tonia 32: 43-46. --, AND -. 198la. Dioon merolae (Za-
miaceae), a new species from Mexico. Brittonia 32: 179-185. --, AND -. 198 1 b. Dioon holmgrenii
(Zamiaceae), a new species from Mexico. Brittonia 33: 552-555. --, AND -. 1982. Distribution and vari-
ation of Dcoon edule (Zamiaceae). Brittonia 34: 355- 362. --, AND -. 1984. Dioon tomaselii (Za-
miaceae), a new species with two varieties from west- ern Mexico. Brittonia 36: 223-227.
DEPARTAMENTODE RECURSOSNATURALES. 1 976. The Master plan for the Commonwealth forest of Puerto Rico. Rio Piedras, Puerto Rico.
DRESSLER,R. L. 1975. It grows up in the trees, really it does. Marie Selby Bot. Gard. Bull. 2: 22-23.
ECKENWALDER, 1980. Taxonomy ofthe West Indian J. E. cycads. J. Arnold Arbor. 61: 701-722.
EWELL,J. J., AND J. L. WHITMORE. 1973. The ecological life zones of Puerto Rico and the U.S. Virgin Islands. Institute of Tropical Forestry, Rio Piedras, Puerto Rico.
EWERS,F. W.,AND R. SCHMID. 198 1. Longevity ofneedle fascicles of Pinus longaeva (bristlecone pine) and oth- er North American pines. Oecologia 5 1 : 107-1 15.
FOSTER,A. S., AND E. M. GIFFORD, JR. 1974. Compar-ative morphology of vascular plants. W. H. Freeman, San Francisco.
GIVNISH, T. J. 1978. On the adaptive significance of compound leaves, with particular reference to tropical trees. In P. B. Tomlinson and M. H. Zimmermann [eds.], Tropical trees as livingsystems, 35 1-380. Cam- bridge University Press, Cambridge, MA.
HALLMAN,E. N. 1976. Notes on the natural history and lore of Zamia in Florida. Proceedings of the 5th Conf. on Cycad Toxicity:I-1-1-10,
HICKMAN,J. C. 1979. The basic biology ofplantnumbers. In 0 . T. Solbrig, J. Subodh, G. B. Johnson, and P. H. Raven [eds.], Topics in plant population biology, 232-263. Columbia University Press, New York.
JANZEN,D. H., AND D. E. WILSON. 1974. The cost of being dormant in the tropics. Biotropica 6: 260-262.
Jow, W. M., S. H. BULLOCK, AND J. KUMMEROW. 1980. Leaf turnover rates of Adenostoma fascicuIatu~?z (Ro-saceae). Amer. J. Bot. 67: 256-261.
ecology of Zamia in Puerto Rico. M.S. thesis. Recinto Universitario de Mayaguez. Mayaguez, Puerto Rico.
, AND G. J. BRECKON. 1985. Seed dispersal, via- bility and survivorship in Zamia debilis in Puerto Rico. Assoc. Trop. Biol. Abstracts, 17-18.
NILSEN, E. T. 1986. Quantitative phenology and leaf survivorship ofRhododendron maximum in contrast- ing irradiance environments of the southern Appa- lachian mountains. Amer. J. Bot. 73: 822-83 1.
PIRERO, D., J. SARUKHAN, AND E. GONZALEZ. 1977. Es- tudios demograficos en plantas. Astrocaryum mexi- canum Liebm. I. Estructura de las poblaciones. Bol. Soc. Bot. Mdxico 37: 69-1 18.
READ, R. W., AND M. L. SOLT. 1986. Bibliography of the living cycads (annotated). Lyonia 2: 33-199.
RICHARDS,P. W. 1966. The tropical rain forest-an eco-logical study. Cambridge University Press, Cam- bridge, MA.
ROHLF, F. J., AND R. R. SOKAL. 198 1. Statistical tables. W. H. Freeman, San Francisco.
S A R U K H ~ , 1980. Demographic problems in tropical J. systems. In 0 . T. Solbrig [ed.], Demography and evo- lution in plant populations, 161-188. University of California Press, Berkeley and Los Angeles.
SOKAL, R. R., AND F. J. ROHLF. 198 1. Biometry: the principles and practice of statistics in biological re- search. W. H. Freeman, San Francisco.
STEVENSON,D. 1980. Observations on root and stem contraction in Cycads with special reference to Zar~zia purnila. J. Linn. Soc., Bot. 81: 275-282.
-. 198 1. Observation on ptyxis, phenology and tri- comes in the Cycadales and their systematics impli- cations. Amer. J. Bot. 68: 1104-1 114.
. 1982. A new species of Cerarozamia (Zamiaceae) from Chiapas, Mexico. Brittonia 34: 181-184.
-. 1987. Again the West Indian zamias. Fairchild Trop. Gard. Bull. 42: 23-27.
-, S. SABATO, TORRES. 1986. AND M. V ~ Q U E Z A new species of Ceratozamia (Zamiaceae) from Vera Cruz, Mexico with comments on species relations, habitats and vegetative morphology in Ceratozamia. Brittonia 38: 17-26.
STREET,H. E., AND H. OPIK. 1984. The physiology of flowering plants, 3d ed. Edward Arnold, Baltimore, MD.
TANG, W. 1986. Sex change experiment with Zamia pumila. Cycad Soc. Newsl. 9: 9-10.
WILLSON,M. F. 1983. Plant reproductive ecology. John Wiley, New York.

You have printed the following article:
Population Structure in Zamia debilis (Zamiaceae). I. Size Classes, Leaf Phenology, andLeaf TurnoverVivian Negrón-Ortiz; Gary J. BreckonAmerican Journal of Botany, Vol. 76, No. 6. (Jun., 1989), pp. 891-900.Stable URL:
http://links.jstor.org/sici?sici=0002-9122%28198906%2976%3A6%3C891%3APSIZD%28%3E2.0.CO%3B2-2
This article references the following linked citations. If you are trying to access articles from anoff-campus location, you may be required to first logon via your library web site to access JSTOR. Pleasevisit your library's website or contact a librarian to learn about options for remote access to JSTOR.
Literature Cited
Demographic Analysis of the Growth of Linum usitatissimumF. A. Bazzaz; John L. HarperNew Phytologist, Vol. 78, No. 1. (Jan., 1977), pp. 193-208.Stable URL:
http://links.jstor.org/sici?sici=0028-646X%28197701%2978%3A1%3C193%3ADAOTGO%3E2.0.CO%3B2-C
Climax Vegetation in Tropical AmericaJ. S. BeardEcology, Vol. 25, No. 2. (Apr., 1944), pp. 127-158.Stable URL:
http://links.jstor.org/sici?sici=0012-9658%28194404%2925%3A2%3C127%3ACVITA%3E2.0.CO%3B2-%23
The Classification of Tropical American Vegetation-TypesJ. S. BeardEcology, Vol. 36, No. 1. (Jan., 1955), pp. 89-100.Stable URL:
http://links.jstor.org/sici?sici=0012-9658%28195501%2936%3A1%3C89%3ATCOTAV%3E2.0.CO%3B2-4
The Ecology of Leaf Life SpansBrian F. Chabot; David J. HicksAnnual Review of Ecology and Systematics, Vol. 13. (1982), pp. 229-259.Stable URL:
http://links.jstor.org/sici?sici=0066-4162%281982%2913%3C229%3ATEOLLS%3E2.0.CO%3B2-B
http://www.jstor.org
LINKED CITATIONS- Page 1 of 3 -

Dioon Rzedowskii (Zamiaceae), a New Species from MexicoPaolo de Luca; Aldo Moretti; Sergio Sabato; Mario Vazquez TorresBrittonia, Vol. 32, No. 2. (Apr. - Jun., 1980), pp. 225-229.Stable URL:
http://links.jstor.org/sici?sici=0007-196X%28198004%2F06%2932%3A2%3C225%3ADR%28ANS%3E2.0.CO%3B2-M
Dioon caputoi (Zamiaceae), a New Species from MexicoPaolo de Luca; Sergio Sabato; Mario Vazquez TorresBrittonia, Vol. 32, No. 1. (Jan. - Mar., 1980), pp. 43-46.Stable URL:
http://links.jstor.org/sici?sici=0007-196X%28198001%2F03%2932%3A1%3C43%3ADC%28ANS%3E2.0.CO%3B2-N
Dioon Merolae (Zamiaceae), a New Species from MexicoPaolo de Luca; Sergio Sabato; Mario Vazquez TorresBrittonia, Vol. 33, No. 2. (Apr. - Jun., 1981), pp. 179-185.Stable URL:
http://links.jstor.org/sici?sici=0007-196X%28198104%2F06%2933%3A2%3C179%3ADM%28ANS%3E2.0.CO%3B2-H
Dioon holmgrenii (Zamiaceae), a New Species from MexicoPaolo de Luca; Sergio Sabato; Mario Vazquez TorresBrittonia, Vol. 33, No. 4. (Oct. - Dec., 1981), pp. 552-555.Stable URL:
http://links.jstor.org/sici?sici=0007-196X%28198110%2F12%2933%3A4%3C552%3ADH%28ANS%3E2.0.CO%3B2-E
Distribution and Variation of Dion Edule (Zamiaceae)Paolo de Luca; Sergio Sabato; Mario Vazquez TorresBrittonia, Vol. 34, No. 3. (Jul. - Sep., 1982), pp. 355-362.Stable URL:
http://links.jstor.org/sici?sici=0007-196X%28198207%2F09%2934%3A3%3C355%3ADAVODE%3E2.0.CO%3B2-O
Dion Tomasellii (Zamiaceae), a New Species with Two Varieties from Western MexicoPaolo de Luca; Sergio Sabato; Mario Vazquez TorresBrittonia, Vol. 36, No. 3. (Jul. - Sep., 1984), pp. 223-227.Stable URL:
http://links.jstor.org/sici?sici=0007-196X%28198407%2F09%2936%3A3%3C223%3ADT%28ANS%3E2.0.CO%3B2-Y
http://www.jstor.org
LINKED CITATIONS- Page 2 of 3 -

The Cost of Being Dormant in the TropicsDaniel H. Janzen; Don E. WilsonBiotropica, Vol. 6, No. 4. (Dec., 1974), pp. 260-262.Stable URL:
http://links.jstor.org/sici?sici=0006-3606%28197412%296%3A4%3C260%3ATCOBDI%3E2.0.CO%3B2-L
A New Species of Ceratozamia (Zamiaceae) from Chiapas, MexicoDennis Wm. StevensonBrittonia, Vol. 34, No. 2. (Apr. - Jun., 1982), pp. 181-184.Stable URL:
http://links.jstor.org/sici?sici=0007-196X%28198204%2F06%2934%3A2%3C181%3AANSOC%28%3E2.0.CO%3B2-A
A New Species of Ceratozamia (Zamiaceae) from Veracruz, Mexico with Comments onSpecies Relationships, Habitats, and Vegetative Morphology in CeratozamiaDennis Wm. Stevenson; Sergio Sabato; Mario Vazquez TorresBrittonia, Vol. 38, No. 1. (Jan. - Mar., 1986), pp. 17-26.Stable URL:
http://links.jstor.org/sici?sici=0007-196X%28198601%2F03%2938%3A1%3C17%3AANSOC%28%3E2.0.CO%3B2-Q
http://www.jstor.org
LINKED CITATIONS- Page 3 of 3 -




















