The next generation of iron fertilization experiments in the ...
Upload
independentCategory
view
1download
0
DEVELOPMENTAL BIOLOGY kg, 7346 (1969)
Polyribosome Formation and RNA Synthesis in the
Early Post-Fertilization Stages of the
Sea Urchin Egg
ANNA MARIA RINALDI AND ALBERTO MONROY
Laboratory of Comparative Anatomy, The University, and Research Unit for Molecular Embryology of the C.N.R., Palermo, ltaly
Accepted October 23, 196S
The rise of the rate of protein synthesis that follows fertilization in the sea urchin egg has been attributed to the removal of a translational inefficiency of the ribosomes of the unfertilized egg (Hultin, 1961; Monroy, Maggio and Rinaldi, 1965; Maggio, Vittorelli, Caffarelli- Mormino and Monroy, 1968). On the other hand there is satisfactory evidence that (1) in the unfertilized egg the amino acid activating enzymes, the amino-acyl-tRNA synthetases and the tRNA itself are not limiting and indeed their activity does not appear to change significantly at least for several hours after fertilization (Maggio and Catalano, 1963; Ceccarini, Maggio and Barbata, 1967); (2) that in the unfertilized egg there is a store of mRNA which is enough to support development at least until the blastula stage although most of it does not appear to be available for translation in the unfertilized egg (Gross and Cousineau, 1963; Maggio, Vittorelli, Rinaldi and Monroy, 1964; Slater and Spiegelman, 1966). How soon after fertiliza- tion RNA synthesis begins is uncertain; this is however well under- way during early cleavage stages. On account of its base composition (Gross, Kraemer and Malkin, 1965), of its sedimentation properties and of its hybridization with DNA this RNA may be classified as mRNA (Nemer, 1963; Wilt, 1963; Nemer and Infante, 1965). The early synthesized RNA has been described as being mainly associated with a class of light polyribosomes and it has been suggested that it is stored there in an inactive state until after the blastula stage (Spirin and Nemer, 1965).
Using the method recently described by Cohen and Iverson ( 1967) which allows the preparation of undegraded polyribosomes from sea
73

74 RINALDI AND MONROY
urchin eggs, we have now been able to show ( 1) that a small number of active polyribosomes are functioning in the unfertilized egg; (2) that the polyribosomal population increases in a characteristic way beginning by 2 minutes after fertilization; (3) that rapidly labeled heterogeneous RNA becomes attached to both heavy and light poly- ribosomes before the first cleavage.
METHODS
We have used the eggs of t!-e mediterranean sea urchin, Paracen- trotus Zividus. The culture of the embryos as well as the incubations with radioactive precursors were always carried out in the presence of penicillin and streptomycin (Dicrysticin, Erba 140 mg/lt) and sulfadiazin ( 50 mg/lt ) . The eggs were homogenized using the high-K buffer essentially as described by Cohen and Iverson (1967) (Tris- HCl buffer 0.05 M pH 7.6; KC1 0.4 M; Mg-acetate 0.018 M). Since, however, breaking the cells in this nearly isotonic buffer is difficult and requires a vigorous homogenization, we have found it advan- tageous to add to the medium 0.5 per cent Deoxycholate and 0.5 per cent Tween 40. One mg of low-speed bentonite (Watts and Mathias, 1967) was added per ml of solution. In this way two to five gentle strokes of a hand operated homogenizer with loosely fitted plunger were sufficient to ensure complete breakage of all the eggs and of the embryos as well. Usually a 1: 5 (w/v) ratio of eggs to homoge- nizing buffer was used. Before homogenization the eggs were washed once with Ca- and Mg-free sea water. The homogenate was centri- fuged at 7,000 x g for 10 minutes and 1 ml of the supernatant was layered on top of a 1750 per cent sucrose linear gradient with a 60 per cent sucrose cushion. The gradients were centrifuged at 52,000 x g for 3.5 hours (Spinco Rotor SW 25.1); they were collected in 1 ml fractions and the A,,, continuously recorded with an ISCO fraction collector.
Incorporation studies. For the assay of the amino acid incor- poration, each fraction with the addition of 100 pg of serum albumin per tube was precipitated with 5% (final cont. ) cold TCA. After standing overnight in refrigeration the tubes were heated at 90°C for 15 minutes and the precipitate collected on Millipore filters and washed with 50 ml of 5% cold TCA. The filters were then dried and counted on a Nuclear Chicago Scintillation Spectrometer using a POP-POPOP in toluene scintillation fluid. For the analysis of the

POLYRIBOSOMES AND RNA SYNTHESIS IN SEA URCHIN EGGS 75
‘H-Uridine incorporation in the RNA, the fractions were precipitated as described above omitting the hot-TCA extraction. For the extrac- tion of RNA from the polyribosomes, the fractions from three to six gradients were pooled and extracted as described by Penman ( 1966) with the addition of purified Paracentrotus total RNA as carrier. The RNA was then displayed on a 15-30 per cent sucrose gradient in SDS buffer (Penman, 1966) at 50.000 g for 14 hours at 25°C. The radioactivity of the fractions was counted in Bray’s solution. Addi- tional technical details will be given in the legends of the figures. The radioisotopes were obtained from Radiochemical Center, Amersham, England.
RESULTS
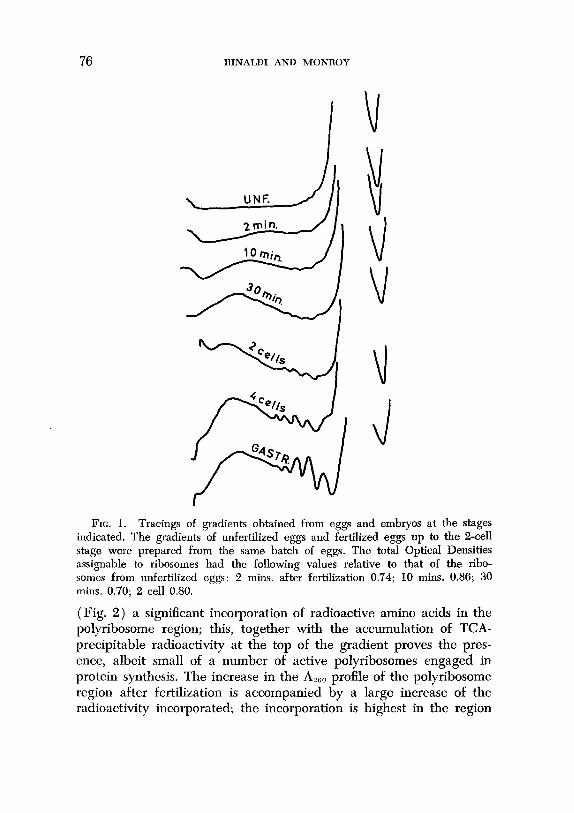
Fig. 1 presents the sedimentation profiles of polyribosomes of eggs collected at close time intervals after fertilization and from embryos at a later stage of development for comparison. The tracings provide evi- dence first of all that polyribosomes are already present 2 minutes after fertilization and that the area of the heavy polyribosomes in- creases rapidly during the first 30 minutes after fertilization. In spite of the difficulties of obtaining precise estimates, we have consistently observed an increase of the polyribosomes: monoribosomes ratio dur- ing these first 30 minutes after fertilization (from about 0.12 in the unfertilized eggs to about 0.18 at 10 minutes after fertilization and 0.30 at 30 minutes after fertilization; this ratio reaches 0.7 in the gastrula; see also Monroy and Tyler, 1963). It further appears from the tracings that the center of gravity of the heavy polyribosomes is progressively displaced toward the heavy regions of the gradient until approximately the end of the first cell cycle and then remains prac- tically unchanged. A group of smaller polyribosomes becomes notice- able 10 minutes after fertilization and becomes more prominent con- currently with the overall increase of the polyribosomal population. We have not observed a differential increase of either one of the two groups of polyribosomes at a particular stage (see Infante and Nemer, 1967). It must be noted that a mild degradation of the poly- ribosomes is first indicated by a decrease of the area of the heavy polyribosomes and a corresponding increase of the peaks of the lighter polyribosomes.
Gradients obtained from pulse-labeled unfertilized eggs show
76 RINALDI AND MONROY
FIG. 1. Tracings of gradients obtained from eggs and embryos at the stages indicated. The gradients of unfertilized eggs and fertilized eggs up to the 2-cell stage were prepared from the same batch of eggs. The total Optical Densities assignable to ribosomes had the following values relative to that of the ribo- somes from unfertilized eggs: 2 mins. after fertilization 0.74; 10 mins. 0.86; 30 mins. 0.70; 2 cell 0.80.
(Fig. 2) a significant incorporation of radioactive amino acids in the polyribosome region; this, together with the accumulation of TCA- precipitable radioactivity at the top of the gradient proves the pres- ence, albeit small of a number of active polyribosomes engaged in protein synthesis. The increase in the Az6,, profile of the polyribosome region after fertilization is accompanied by a large increase of the radioactivity incorporated; the incorporation is highest in the region

POLYRIROSOMES AND RNA SYNTHESIS IN SEA URCHIN EGGS 77
of the heavy polyribosomes and is much smaller in the region of the lighter ones. All the radioactivity over the polyribosomes is displaced to the monoribosome peak and lighter regions of the gradient by treatment with RNase; 0.01 M EDTA also releases more than 75 per cent of the radioactivity.
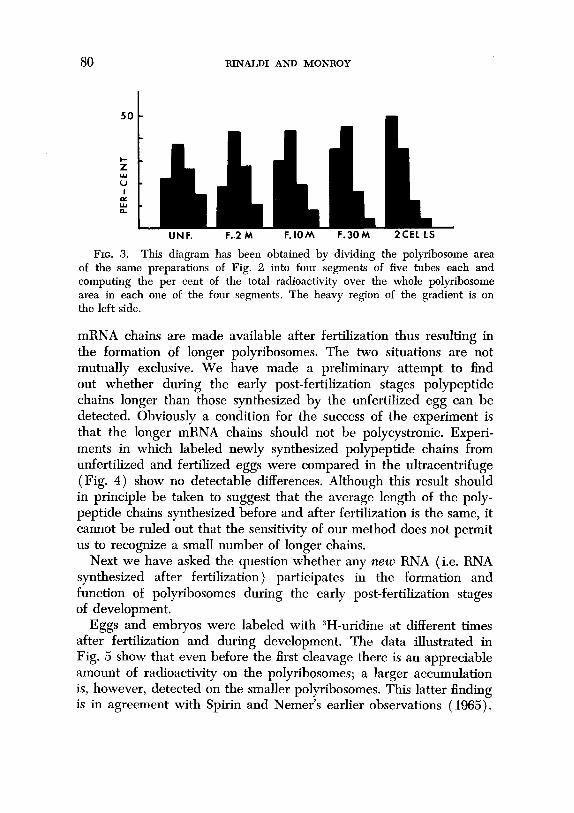
The diagram Fig. 3 shows the displacement of the relative in- corporating activity from the lighter to the heavier polyribosomes during the first cell cycle (one hour after fertilization). Thus both the A,,, tracings and the distribution of the radioactivity over the polyribosomes suggest a progressive increase of the average sedi- mentation value of the heavy polyribosomes. To account for this, two possibilities have been considered: (1) as a result of the removal of the translational inefficiency of the ribosomes after fertilization, the mRNA chains become more crowded with ribosomes; (2) longer
(2 - Li
90 ‘- 8.0 -- 7.0 -- UNFERTILIZED
60 .- 50 .-
4.0 .-
3.0 ‘: 20:-
5
5
10 1’5 10 1’5 20 25 20 25 FRACTIONS
FIG. 2. Distribution of radioactivity in gradients of eggs and embryos pulsed for 2 mins. with 10 PC of ‘“C reconstituted protein hydrolysate (spec. act. 1618 mC/mM ) per ml of egg suspension. Owing to the low level of uptake and of incorporation of the unfertilized eggs, a 2 minute pulse results in hardly detectable radioactivity over the polyribosomes. Hence the diagram has been combined from two experiments in one of which the eggs were pulsed for 2 minutes and in the other one for 10 minutes.

78 RINALDI AND MONROY
5 10 15 20 25 FRACTIONS
6.0
7.0
6.0
5.0 4.0 3.0
2.0
FERTILIZED 10 min
FRACTIONS
FIG. 2 (Continued)

1’OLYFiIBO‘OMES AND RNA SYNTHESIS IN SEA URCHIN FGGS 79
9.0
80
70
6.0
5.0
4.0
3.0
2.0
(2 -4)
FERTILIZED 30 min.
5 10 1’5 20 25
t=RACTIONS
I’
I 1 ,
9.0 ,- (2 - 5,
8.0 -- y, 70 -- :: 2 cell5
60.- : b4 50 -r : 9
I
I...!..,,!.,.,!‘.,.!....!...
5 10 15 20 25 FRACTIONS
FIG. 2 (Continmd)
30
0 20 I
:,
r4- ‘0
z
IO z
v
30
207 I
:,
c4- ‘0
x
10 r a 0
80 RINALDI AND MONROY
UN F. F.2 M F.lOM F.30M PCEL LS
FIG. 3. This diagram has been obtained by dividing the polyribosome area of the same preparations of Fig. 2 into four segments of five tubes each and computing the per cent of the total radioactivity over the whole polyribosome area in each one of the four segments. The heavy region of the gradient is on the left side.
mRNA chains are made available after fertilization thus resulting in the formation of longer polyribosomes. The two situations are not mutually exclusive. We have made a preliminary attempt to find out whether during the early post-fertilization stages polypeptide chains longer than those synthesized by the unfertilized egg can be detected. Obviously a condition for the success of the experiment is that the longer mRNA chains should not be polycystronic. Experi- ments in which labeled newly synthesized polypeptide chains from unfertilized and fertilized eggs were compared in the ultracentrifuge (Fig. 4) show no detectable differences. Although this result should in principle be taken to suggest that the average length of the poly- peptide chains synthesized before and after fertilization is the same, it cannot be ruled out that the sensitivity of our method does not permit us to recognize a small number of longer chains.
Next we have asked the question whether any new RNA (i.e. RNA synthesized after fertilization) participates in the formation and function of polyribosomes during the early post-fertilization stages of development.
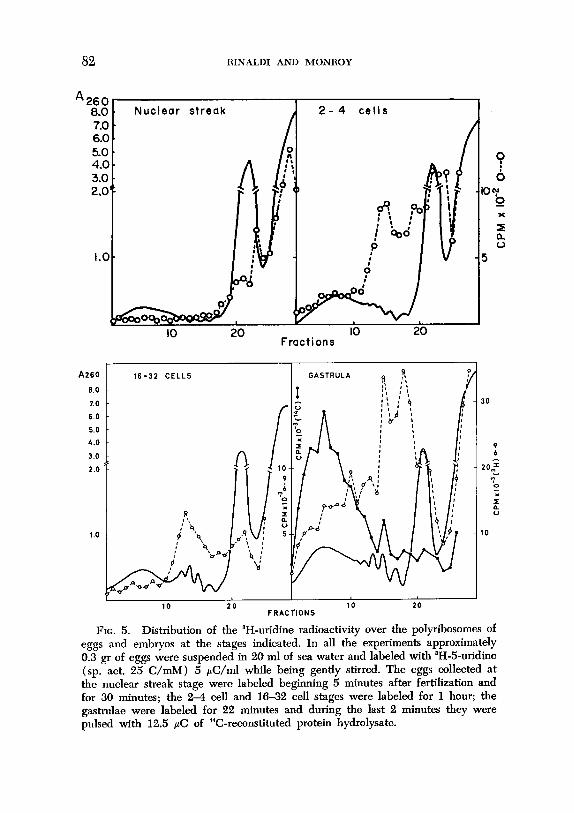
Eggs and embryos were labeled with 3H-uridine at different times after fertilization and during development. The data illustrated in Fig. 5 show that even before the first cleavage there is an appreciable amount of radioactivity on the polyribosomes; a larger accumulation is, however, detected on the smaller polyribosomes. This latter finding is in agreement with Spirin and Nemer’s earlier observations ( 1965).

POLYRLBOSOMES AND RNA SYNTHESIS IN SEA URCHIN EGGS 81
Fractions FIG. 4. Unfertilized eggs and eggs immediately after fertilization were pulsed
with radioactive leucine: the unfertilized eggs were pulsed with “C-1-leucine (sp. act. 165 mC/mM), 10 &/ml and the fertilized eggs with 3H-l-leucine (sp. act, 200 mC/mM), 10 &/ml. After 30 minutes the eggs were washed and homogenized in Tris-HCl 0.05 M pH 7.6, KC1 0.025 M and Mg-acetate 0.004 M. The homogenates were centrifuged at 7.000 X g for 10 minutes and the super- natant centrifuged again at 138.000 X g for 50 minutes. The upper 2/3 of the supernatants were now mixed and SDS and mercaptoethanol added to a final concentration, of 1% of each and heated at 37’C for 3 hours. The solution was then dialysed against the same buffer containing SDS and mercaptoethanol 0.1% at 18°C for 18 hours. The solution was concentrated to small volume and 0.2 ml layered on top of a 5-20 per cent sucrose gradient prepared in the same buffer containing 0.1% of SDS and mercaptoethanol and centrifuged in the SW 39 rotor of the Spinco ultracentrifuge at 38.000 rpm for 38 hours at 24“C. Three drop fractions were collected from the bottom of the tube and the radioactivity measured in Bray’s solution. The diagram indicated the per-cent distribution of the 14C (0) and ‘H (0) counts in each fraction.
Comparison of the diagrams shows (1) that before the first cleav- age the peak of radioactivity is within the area of the monoribosomes; actually, it is somewhat displaced toward the leading edge of the monoribosome peak; (2) and that the accumulation of the 3H-uridine
82 RINALDI AND MONROY
*2:;
;.( 61 5S 4.t 3.C 2.C
I.(
A260
8.0
7.0
6.0
5.0
4.0
3.0
2.0
Nuclear streak 2-4 cells
Fractions
16-32 CELLS GASTRULA
10 20 10 20 FRACTIONS
FIG. 5. Distribution of the aH-uridine radioactivity over the polyribosomes of
eggs and embryos at the stages indicated. In all the experiments approximately
0.3 gr of eggs were suspended in 20 ml of sea water and labeled with ‘H-5-uridine
(sp. act. 25 C/mM) 5 &/ml while being gently stirred. The eggs collected at
the nuclear streak stage were labeled beginning 5 minutes after fertilization and for 30 minutes; the 24 cell and 16-32 cell stages were labeled for 1 hour; the
gastrulae were labeled for 22 minutes and during the last 2 minutes they were pulsed with 12.5 PC of “C-reconstituted protein hydrolysate.
POLYRIROSOMES AND RNA SYNTHESIS IN SEA URCHIN EGGS 83
over the small polyribosomes becomes very prominent after the first cleavage.
The radioactivity is completely removed by RNase. The radio- activity over the polyribosomes region is also largely (about 70 per cent) removed by 0.01-0.02 M EDTA. However, the results have been rather variable and complicated by the concurrent breakdown of the ribosomes. It has to be noted that even at the gastrula stage, when the amount of the rapidly labeled newly synthesized RNA attached to the polyribosomes is very large, the modal RNA radio- activity is still over the small polyribosomes.
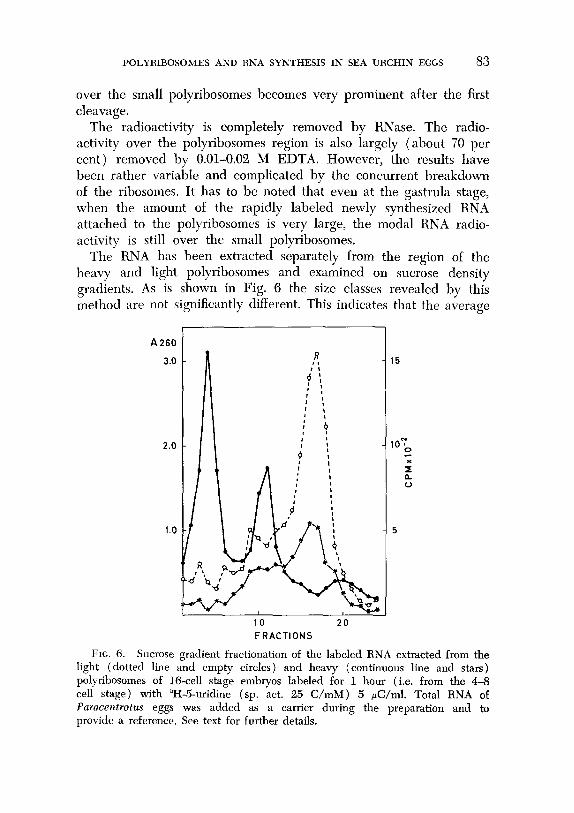
The RNA has been extracted separately from the region of the heavy and light polyribosomes and examined on sucrose density gradients. As is shown in Fig. 6 the size classes revealed by this method are not significantly different. This indicates that the average
A260
3.0 15
1or z
; 0 ”
5
10 20 FRACTIONS
FIG. 6. Sucrose gradient fractionation of the labeled RNA extracted from the light (dotted line and empty circles) and heavy (continuous line and stars) polyribosomes of 16-cell stage embryos labeled for 1 hour (i.e. from the 4-6 cell stage) with “H-5-uridine (sp. act. 25 C/mM ) 5 &/ml. Total RNA of Paracentrotus eggs was added as a carrier during the preparation and to provide a reference. See text for further details.

84 RINALDI AND MONROY
length of the newly synthesized RNA molecules is the same on the heavy and light polyribosomes. The diagram shows further that this is a heterogeneous, non-ribosomal RNA containing only a minimum amount of light (i.e. less than 8 S) components. Bacterial contamina- tion (Glisin and Glisin, 1965) should have been detected by radio- active peaks at 23 and 16 S.
Actinomycin D in the range from 10 to 40 pg/ml reduces the in- corporation by 50 to 75 per cent. This treatment does not appear to alter significantly the polyribosome profiles nor the ratio of the heavy to light polyribosomes.
CONCLUSIONS
The present results thus show first that in the unfertilized egg there is a small number of polyribosomes active in protein synthesis. Mobil- ization of ribosomes which become attached to the mRNA in rapidly increasing number follows upon fertilization. As to the mechanism of the increase of the polyribosome population after fertilization, the present experiments do not provide any evidence concerning the relative contributions of the mRNA already being translated in the unfertilized egg and the mRNA present in a non- translatable condition which becomes available only after fertiliza- tion. On the other hand, we find that very soon after fertilization, and in fact even before the first cleavage, a rapidly labeled, non- ribosomal polydisperse RNA begins to be transcribed which is at- tached to the polyribosomes, particularly to the lighter ones. Studies in which the kinetics of labeling of the nuclear and cytoplasmic RNA has been investigated (Giudice and Mutolo, to be published) suggest that the double detergent method used in these experiments prevents nuclear breakage and significant leakage of RNA. In all stages in- vestigated, including gastrula, i.e. when the rate of synthesis of mRNA is very high, 3H-uridine incorporation is the highest in the region of the light polyribosomes. (Considering the short period of labeling in this experiment, i.e. 22 minutes, it appears very unlikely that any radioactivity present may have been due to ribosomal RNA; see Giudice and Mutolo, 1967). This seems to speak against the possi- bility that the RNA that accumulates on the light polyribosomes may be an inactive mRNA stored to be utilized for the post-blastular development (Spirin and Nemer, 1965). In any case the significance and the function of this early synthesized RNA is at present obscure.

POLYRIBOSOMES AND RNA SYNTHESIS IN SEA URCHIN EGGS 85
Robbins and Borun (1967) report that in HeLa cells the small poly- ribosomes are the site of synthesis of histones. We have tested this possibility using essentially the double labeling technique suggested by these workers, (pulse labeling with I’C-tryptophane and “H- lysine). In our material however the ratio of the two isotopes is the same throughout the whole polyribosome region.
We wish to thank Drs. G. Giudice and S. Penman for valuable dis- cussions and suggestions and Drs. P. R. Gross, P. Marks and L. Nel- son for the critical reading of the manuscript. The technical assistance of Mr. 0. A. Oliva is gratefully acknowledged.
SUMMARY
In the unfertilized egg of the sea urchin Paracentrotus Zividus it is possible to detect a small number of polyribosomes active in protein synthesis. The polyribosomal population starts to increase rapidly very soon after fertilization and the polyribosomes become progressively heavier during the first cell cycle. It is suggested that as a result of the removal of the translational inefficiency of the ribosomes follow- ing fertilization the mRNA chains become more crowded with ribosomes.
Very shortly after fertilization, indeed even before the first cleav- age, a rapidly labeled non-ribosomal heterogeneous RNA is synthe- sized that becomes attached to the polyribosomes.
REFERENCES
CECCARINI, C., MAGGIO, R., and BARBATA, G. (1967). Amino-acyl-sRNA syn- thetases as possible regulators of protein synthesis in the embryo of the sea urchin, Pamcentrotus lividus. Proc. Natl. AC. SC. Washington, 58, 2235-2239.
COHEN, G. H., and IVERSON, R. M. (1967). High-resolution density gradient analysis of sea urchin polysomes. Bioch. Bioph. Res. Comm. 29, 349-355.
GIUDICE, G., and MUTOLO, V. ( 1967). Synthesis of ribosomal RNA during sea urchin development. Bioch. Bioph. Acta, 138, 276-285.
GLISIN, V. R., and GLISIY, M. V. (1964). Ribonucleic acid metabolism following fertilization in sea urchin eggs. PTOC. Natl. AC. SC. Washington, 52, 1548-1553.
GROSS, P. R., and COUSINEAU, G. H. ( 1963). Effects of Actinomycin D on macro- molecule synthesis and early development in sea urchin eggs. Bioch. Bioph. Res. Comm. 10, 321-326.
GROSS, P. R., KRAEMER, K., and MALKIX, L. J. (1965). Base composition of RNA synthesized during cleavage of the sea urchin embryo. Bioch. Bioph. Res. Comm. 18, 569-575.
HULTIX, T. ( 1961). Activation of ribosomes in sea urchin eggs in response to fertilization. Exptl. Cell Res. 25, 405417.

86 RINALDI AND MONROY
INFANTE, A. A., and NEMER, M. (1967). Accumulation of newly synthesized RNA templates in a unique class of polyribosomes during embryogenesis. Proc. Natl. AC. SC. Washington, 58, 681-688.
MAGGIO, R., and CATALANO, C. (1963). Activation of amino acids during sea urchin development. Arch. Bioch. and Bioph., 103, 164-167.
MAGGIO, R., VITTORELLI, M. L., RINALDI, A. M., and MONROY, A. (1964). In vitro incorporation of amino acids into proteins stimulated by RNA from un- fertilized sea urchin eggs. Bioch. Bioph. Res. Comm., 15, 436-441.
MAGGIO, R., VITTORELLI, M. L., CAFFARELLI-MORMINO, I., and MONROY, A. ( 1968). Dissociation of ribosomes of unfertilized eggs and embryos of sea urchin. J. Mol. Biol. 31, 621-626.
MONROY, A., h’lAGGI0, R., and RINALDI, A. M. (1965). Experimentally induced activation of the ribosomes of the unfertilized sea urchin egg. Proc. Natl. AC. SC. Washington, 54, 107-111.
MONROY, A., and TYLER, A. ( 1963). Formation of active ribosomal aggregates (Polysomes) upon fertilization and development of sea urchin eggs. Arch. Bioch. Bioph. 103, 431435.
NEMER, M. ( 1963). Old and new RNA in the embryogenesis of the purple sea urchin. Proc. Natl. AC. SC. Washington, 50, 230-235.
NEMER, M., and INFAPL’TE, A. A. (1965). Messenger RNA in early sea urchin embryo; size classes. Science 150, 217-221.
PENMAN, A. ( 1966). RNA metabolism in the HeLa cell nucleus. J. Mol. Biol. 17, 117-130.
ROBBINS, E., and BORUN, T. W. (1967). The cytoplasmic synthesis of histones in HeLa cells and its temporal relationship to DNA replication. Proc. Natl. AC. SC. Washington, 57, 409416.
SLATER, D. W., and SPIEGELMAN, S. ( 1966). An estimation of genetic messages in the unfertilized echinoid egg. Proc. Natl. AC. SC. Washington, 56, 164-171.
SPIRIK, A. S., and NEMER, M. (1965). Messenger RNA in early sea urchin embryos; cytoplasmic particles. Science, 150, 214-217.
WATTS, R. L., and MATHIAS, A. P. (1967). The use of bentonite in the isolation of plant polyribosomes. Bioch. Bioph. Acta, 145, 828-831.
WILT, F. H. ( 1963). The synthesis of ribonucleic acid in sea urchin embryos. Bioch. Bioph. Res. Comm., 11, 447451.
Copyright © 2022 FDOKUMEN