
Plato's theory of ideas revisited
168
Table of Contents for Neural Networks Volume 10, Issue 7, 1 October 1997 Consciousness for Neural Networks [Abstract] [Full text] (PDF 225.6 Kb) 1173-1173 Walter J. Freeman Three Centuries of Category Errors in Studies of the Neural Basis of Consciousness and Intentionality [Abstract] [Full text] (PDF 118.1 Kb) 1175-1183 J. G. Taylor and H.-W. Mueller-Gaertner Non-Invasive Analysis of Awareness [Abstract] [Full text] (PDF 142.8 Kb) 1185-1194 James Newman, Bernard J. Baars and Sung-Bae Cho A Neural Global Workspace Model for Conscious Attention [Abstract] [Full text] (PDF 416.7 Kb) 1195-1206 J.G. Taylor Neural networks for consciousness [Abstract] [Full text] (PDF 1.6 Mb) 1207-1225 Edmund T. Rolls Consciousness in Neural Networks? [Abstract] [Full text] (PDF 401.2 Kb) 1227-1240 Erich Harth From Brains to Neural Nets to Brains [Abstract] [Full text] (PDF 744.8 Kb) 1241-1255 Jeffrey A. Gray, Catalin V. Buhusi and Nestor Schmajuk The Transition from Automatic to Controlled Processing [Abstract] [Full text] (PDF 381.8 Kb) 1257-1268 Javier Ropero Pelaez Plato's theory of ideas revisited [Abstract] [Full text] (PDF 1.2 Mb) 1269-1288 M. Nakao, I. Honda, M. Musila and M. Yamamoto Metastable associative network models of dream sleep [Abstract] [Full text] (PDF 1.4 Mb) 1289-1302 Table of Contents for Neural Networks http://www.elsevier.nl/gej-ng/10/33/54/15/18/show/toc.htt (1 of 2) [29.6.2001 18:38:41]
Transcript of Plato's theory of ideas revisited

Table of Contents forNeural NetworksVolume 10, Issue 7, 1 October 1997
Consciousness for Neural Networks
[Abstract] [Full text] (PDF 225.6 Kb) 1173-1173
Walter J. Freeman
Three Centuries of Category Errors in Studies of the Neural Basis ofConsciousness and Intentionality
[Abstract] [Full text] (PDF 118.1 Kb)
1175-1183
J. G. Taylor and H.-W. Mueller-Gaertner
Non-Invasive Analysis of Awareness
[Abstract] [Full text] (PDF 142.8 Kb)1185-1194
James Newman, Bernard J. Baars and Sung-Bae Cho
A Neural Global Workspace Model for Conscious Attention
[Abstract] [Full text] (PDF 416.7 Kb)1195-1206
J.G. Taylor
Neural networks for consciousness
[Abstract] [Full text] (PDF 1.6 Mb)1207-1225
Edmund T. Rolls
Consciousness in Neural Networks?
[Abstract] [Full text] (PDF 401.2 Kb)1227-1240
Erich Harth
From Brains to Neural Nets to Brains
[Abstract] [Full text] (PDF 744.8 Kb)1241-1255
Jeffrey A. Gray, Catalin V. Buhusi and Nestor Schmajuk
The Transition from Automatic to Controlled Processing
[Abstract] [Full text] (PDF 381.8 Kb)1257-1268
Javier Ropero Pelaez
Plato's theory of ideas revisited
[Abstract] [Full text] (PDF 1.2 Mb)1269-1288
M. Nakao, I. Honda, M. Musila and M. Yamamoto
Metastable associative network models of dream sleep
[Abstract] [Full text] (PDF 1.4 Mb)1289-1302
Table of Contents for Neural Networks
http://www.elsevier.nl/gej-ng/10/33/54/15/18/show/toc.htt (1 of 2) [29.6.2001 18:38:41]

Chris Browne, Richard Evans, Nick Sales and Igor Aleksander
Consciousness and neural cognizers: a review of some recentapproaches
[Abstract] [Full text] (PDF 441.4 Kb)
1303-1316
Ron Sun
Learning, action and consciousness: a hybrid approach towardmodelling consciousness
[Abstract] [Full text] (PDF 309.2 Kb)
1317-1331
Robert Alan Brown
Consciousness in a Self-Learning, Memory-Controlled, CompoundMachine
[Abstract] [Full text] (PDF 903.1 Kb)
1333-1343
© Copyright 2001, Elsevier Science, All rights reserved.
Table of Contents for Neural Networks
http://www.elsevier.nl/gej-ng/10/33/54/15/18/show/toc.htt (2 of 2) [29.6.2001 18:38:41]

Introduction: 1997 Special Issue
Neural Networks for Consciousness
Consciousness research is now coming to the
forefront in cognitive neuroscience. The instruments
of non-invasive imagingÐPET, fMRI, EG andMEG
are now being deployed to attempt to tease out how,
when and where consciousness emerges in the brain
and how it is then used to provide the e�ciency of
processing able to achieve acts of great creativity and
reasoning. Yet, the wealth of data pouring in will not
be able to be correlated together and used to build a
clear picture of how the brain works, and in particular
how consciousness itself is supported, without there
being a corresponding underpinning at a theoretical
level. This must be ultimately at the level of neural
networks. That is why it is our pleasure to welcome
you to this special issue on ``Neural Networks for
Consciousness''. The issue contains contributions
from across the range of workers in the ®eldÐfrom
experimental neuroscientists to cognitive scientists to
engineers to mathematicians. This breadth alone
shows the vitality of the ®eld and the great interest
in it from across these disciplines. It also shows the
strength future research can draw on since all of these
disciplines can, in principle, contribute to solving the
problems that consciousness raises. The problems
themselves are very deep indeed, and we do not claim
that we have assembled the last word on the subject.
However, the acumen and experience in thinking
about consciousness that the contributors undoubt-
edly possess will make this issue of both general
interest and also of relevance to those concerned
either in their own thinking processes or creating
machines which can emulate them. We wish you
enjoyment in reading the contributions.
John G. Taylor
London
Walter Freeman
Berkeley, CA
Neural Networks, Vol. 10, No. 7, pp. 1173, 1997
Copyright# 1997 Published by Elsevier Science Ltd. All rights reserved
Printed in Great Britain
0893±6080/97 $17.00+.00
PII: S0893-6080(97)00095-6
Pergamon
1173

1997 SPECIAL ISSUE
Non-Invasive Analysis of Awareness
J. G. Taylor and H.-W. Mueller-Gaertner
Institute for Medicine, Research Centre-Juelich, D-52425
(Received30 September1996;accepted2 May 1997)
Abstract—A program is outlined which indicates in what manner awareness may be probed by the recently developednon-invasive instruments (PET, fMRI, MEG and EEG). The nature of the neural systems needed for the creation ofawareness is reviewed, leading to a set of characteristics required for the crucial experiments able to uncover theprocesses involved. The assessment of experiments possible to probe the emergence of perception is then surveyed interms of these criteria, and those which appear to be most effective are delineated. Brief conclusions are then drawn as tothe viability of the program.q 1997 Elsevier Science Ltd.
1. INTRODUCTION
There is considerable interest in the use of non-invasiveinstruments to detect the emergence of phenomenalawareness, say of a percept. The new machines givehope of being able to localise, both spatially and tempo-rally, the neural processes which underpin such an event.However, there are numerous questions which must beanswered before the search can be made well-enoughdefined to design paradigm experiments which can beexpected to be of high import in this task. It is the pur-pose of this paper to explore these questions and come toconclusions as to the viability of this program.
Amongst these questions are:
(a) what is the nature of phenomenal awareness itself?(b) how can the ‘‘explanatory gap’’ (which separatesthe internal experience from the external neuralpatterns of the behavioural response) be bridged?(c) what is the expected spatial localisation of theprocess?(d) what are the expected dynamical neural processes?
Some of these questions are very difficult, especially(a) and (b), but without at least a preliminary attempt toanswer them it may be difficult to answer questions (c)and (d), and so give guidance to the proposed experi-mental program.
There is a considerable lack of agreement as to theanswers to (a) and (b). However, it is becoming
recognised that awareness involves use of previouslyencoded material to give content and meaning to newinputs (Baars, 1988; Baddeley, 1993; Edelman, 1989;Taylor, 1973, 1991). Initial processing before the aware-ness of percepts occurs is at the lowest level of coding,that of feature encoding and filtering in the early andassociative cortices. At the next stage, higher levelobject representations are expected to be activatedwhich begin to ‘‘fill out’’ and bind together the lowerlevel activations into meaningful percepts. At variousstages there would also be feedback helping to guideactivation of the best representation able to fit the totalinput (Bullier, 1995). Awareness of a percept, say of ageometrical shape, of a colour or a moving region, willbe expected to occur after the lowest level of coding hasbeen achieved, so at the object (or equivalent) level,although exactly where that occurs is difficult to pindown, and is the object of the experimental programbeing discussed here. However, persistence of suitableencoded activity is expected to be essential. The contin-ued activity arises in modules with the special property ofbeing able to hold activity for a suitably long temporalduration. Such modules are different from those used inthe first (non-conscious) stage of the processing.
This leads to a working definition of awareness at thelowest level as some form of encoded input which isavailable, over a suitably extended period of time, forfurther processing, and in particular for ‘‘filling out’’,by using closely related stored representations ofexperiences from the past, so as to be useable at higherlevels (for action responses, reasoning, planning, forexample; Taylor, 1998). This definition needs to bedeveloped more fully in terms of detailed results from
Neural Networks, Vol. 10, No. 7, pp. 1185–1194, 1997q 1997 Elsevier Science Ltd. All rights reserved
Printed in Great Britain0893–6080/97 $17.00+.00
PII: S0893-6080(97)00055-5
Pergamon
1185
Acknowledgements: J. G. Taylor would like to thank his co-author(H.-W. Mueller-Gaertner) for the excellent support given him whilsthe was a guest scientist at the Institute of Medicine, ResearchCentre-Juelich where this work was performed.

the experimental program, but should help in initiallyclarifying what is to be looked for.
What results from the above brief sketch of the natureof awareness is a two-stage model for consciousness witha simplified flow chart as in Figure 1. In the figure, the‘‘Non A’’ module performs the low-level coding ofwhich a person is unaware. The output from this moduleis then passed on to the awareness module ‘‘A’’ or canbypass it so as to go directly to the motor responsesystem. This two-stage model is to be distinguishedfrom the continuous or one-stage model, in which con-sciousness arises in the same modules as were used in thenon-conscious processing. The reason for such extraawareness of the activity would, for some reason orother in this one-stage model, be due to increased activityin the relevant modules.
There is much support for the two-stage model. ThusWilliam James (1950) wrote ‘‘the distribution of con-sciousness shows it to be exactly such as we mightexpect in an organadded for the sake of steering anervous system grown too complex to regulate itself’’.Also, from evidence associated with certain forms ofprosopagnosia, Young (1994) wrote ‘‘activation mustcross some form of threshold before it can result inawareness’’. Further evidence from neglect led Bisiachand Berti (1995) to note ‘‘patients may be apparentlyunaware of what lies on the side opposite to the brainlesion despite its having been processed up to a highlevel, perhaps, at which meaning is captured’’. Finally,there is evidence from study of the blindsightedsubject GY that ‘‘the visual input can reach V5 withoutpassing first through V1 and that such input is suffi-cient for both discrimination and conscious awareness
of the visual input’’ (Weiskrantz et al., 1995). Thesequotations, especially the last two, indicate that thereare cortical modules needed for the production of aware-ness beyond those involved in low-level processing.
The answer to (b) above, as to the bridging of theexplanatory gap, is already indicated by the aboveanswer to (a). It will be in terms of the manner inwhich the preprocessed neural activity representing aninput is able to be made available for further transforma-tions which lead to a higher cognitive level. Such avail-ability will require suitable temporal duration of therepresentations so that they can be involved in activitiesin regions crucially concerned with such higher levelprocesses, such as the frontal lobes.
It is natural to consider possible ‘‘buffer’’ posteriormemory sites, holding activity encoded at object level,as the first candidates for sites of this availability. Thesesites sustain temporally extended activity, as is knownfrom psychological tests of short-term memory function-ing in the presence of distractors to prevent long-termmemories from forming (Baddeley, 1986). Such short-term memory buffers have been observed directly bynon-invasive techniques by a number of groups (Paulesuet al., 1993; Salmon et al., 1996; Smith and Jonides,1995) after the initial proposal for their existence basedon psychological data; the present evidence is sum-marised in Table 1. These sites give a tentative firstanswer to question (c). They also allow the model ofFigure 1 to be extended to that of Figure 2.
In Figure 2, the highest level of coding at a non-awarelevel is denoted SM (for semantic memory in wordprocessing) which feeds automatically to the workingmemory WM. It is in the latter that awareness arises,through lateral inhibitory and excitatory connections onthe WM module. In Figure 3 are shown two of thecoupled SM/WM pairs for different codes (denoted Aand B). The manner in which awareness arises fromjoint activity of these two sets of modules has been pro-posed to involve a more global control system involvingthe thalamus and the nucleus reticularis thalami (a sheetof inhibitory neurons with good lateral connections;Taylor, 1992). It is through this sheet that a competitioncan be run between the two (or more) coupled SM/WMsystems shown in Figure 2.
1186 J. G. Taylor and H.-W. Mueller-Gaertner
FIGURE 1. The hierarchical two-stage model of consciousness inwhich the early processing by the ‘‘Non A’’ modules is only atpreconscious level whilst that by the ‘‘A’’ modules supports theemergence of consciousness.
TABLE 1PET results on posterior sites of working memory
Task Brain area Task/Code
Spatial discrimination Left 40 (posterior parietal)a Object shapeRight 40/19 (occipito/parietal)a
Right 19 (occipital)b Orientation discrimination
Object discrimination Left 40 (posterior parietal)a ObjectLeft 37 (inferotemporal)a
Phonemes/Words Left 40 (posterior parietal)c Word disambiguation

In a similar manner, the coupled SM/WM modulepairs of Figure 2 provide a first guess at the temporaldynamics of the initial stages of awareness, question(d), as has been recently discussed by Taylor (1996b).This gave an analysis, for a lexical decision task (Marcel,1980), of the manner in which subliminally processeddata can retard or speed up processing of later data. Itposited that the initial emergence of awareness of wordsarises from their persistence on the phonological store inBrodmann’s area 40 (Paulesu et al., 1993), where theydecay away over a fixed time of several seconds.
In this model, the initially unperceived neural activityat the highest level of preprocessing (in the lexical caseup to the level of the semantic coding in Wernicke’sarea) has numerous representations active, correspond-ing to possibly contradictory interpretations. It is onlyat the next level up, that of the phonological store,that a competition is run between these various possi-bilities, and one is singled out by the use of previousactivities there which had earlier reached awareness orhad buffered subliminal activity. Thus, perception isregarded as the filter to achieve disambiguation betweenalternative interpretations based on earlier decisionmade there.
It is unclear at this stage exactly how to extend thismodel to other modalities, especially to vision, wherenumerous separate codes exist and the problem of thecombination of the resulting activity may be mostextreme. It is the purpose of the following discussionto consider several possibilities of doing that whichwill delineate features needing to be probed by suggestedexperiments, and hence give valuable criteria to applyto their choice.
2. NEURAL SUPPORTS AND CRITERIA
There appear to be two extremes of neural support whichmay be suggested as being at the basis of the emergenceof perception:
1. in a single module (as suggested by the model of wordperception in Taylor, 1996b)
2. as a distributed activity over a set of coupled modu-les.The second model is more likely to be valid invision, given the number of codes (colour, motion,shape, texture, stereoscopy) and the level of intercon-nection between the various visual areas (Fellemanand Van Essen, 1991). However, it is still to beexpected that there will be certain of these visualareas which are playing a more decisive part in theemergence of a visual percept in some particular code,with characteristics as indicated at the end of the pre-vious section.
Whilst the questions (c) and (d) above about the spatialand temporal characteristics of neural activity cruciallyinvolved in perception were posed separately, it is clearthat the answers to them must be combined in somemanner. This is achieved on positing (Taylor, 1996b)that the ‘‘buffering’’ memory sites be regarded as candi-dates for emergence of percepts in that particular code.Thus we are lead to the first criterion:
1. To search for those posterior cortical sites which aremost effective in temporally sustaining activity as apercept becomes experienced.
There may well arise a set of sites satisfying the abovecriterion, in which there may be a spread of temporalduration over the various modules, but they all musthave several seconds of duration for neural activity.How do we regard the emergence of the percept: fromall of the modules conjointly or only, say, from thatmodule which holds activity for the longest time? Thisis partly an experimental question, but there is greatrelevance to perform the related psychophysical experi-ments to ascertain the time of first experience by thesubject of the percept.
An example of such a combination of psychophysicaland non-invasive instrument experiments is that of themotion after-effect (Tootall et al., 1995), in whichthe experiential decay of adaptation was found to
1187Non-Invasive Analysis of Awareness
FIGURE 2. Competition for emergence into consciousness in thetwo-stage model where there is lateral inhibition on the second‘‘A’’ or working memory (WM) stage modules producing con-sciousness from the earlier preprocessing on the ‘‘Non A’’ orsemantic (SM) modules.

match that of blood flow activation as observed by fMRIin suitable visual cortical regions. However, it is neces-sary to note if there are any regions in which activitypersists even longer than that of the subjective experi-ence. The model of lexical decision of (Taylor, 1996b)would indicate that activity on the phonological storepersists longer than the subjective experience itself.This arose, according to the model, when the neuralactivity in the buffer site became stronger than somecriterial value. The activity then decays away over afew seconds, and so should leave a signal of its activationtrace lasting longer than the subjective experience. Thuswe are led to the second criterion:
2. Attempt to order the modules observed to haveextended temporal activity according to the duration ofthe observed activity (beyond that experienced by thesubject); the best candidate for the emergence ofperception will be that module with the longest temporalduration.
It is still not clear that the module holding neuralactivity for the longest time during the development ofa percept will be the only one involved in the emergenceof that percept in the subject’s experience. There mayindeed be others which hold activity for a very similar,but slightly shorter, duration. In that case it would benecessary to measure the manner in which there is acausal flow of activity between the modules to determineif one is dominating the others. Thus we must impose thethird criterion:
3. The most suitable candidate(s) will be those moduleswhich are causally the dominant one(s) in the temporaldynamics causing the emergence of the percept.
To apply this criterion, it will be necessary to measureaccurately both the temporal dynamics of the neuralactivity as well as that of the subjective experience.This criterion puts a requirement on the appropriatemachines to use as those with fast temporal sensitivity,i.e. EEG and MEG. It may also be possible to use suitablesequencing of the slicing in echo-planar imaging in fMRIto follow the dynamics of activity through the brain(Shah, 1997).
3. SURVEY OF RELEVANT EXPERIMENTS
The purpose of this section is to consider how effectivevarious experiments will be in achieving the aim to cap-ture the cortical sites of the emergence of perceptionunder various visual (and other modality) stimulationconditions. The basic question that has to be addressedin this assessment is:
Does the experiment under consideration reveal areaswhich give ‘‘added value’’ to cortical processing
beyond the level of non-conscious processing alreadyknown to occur, say at the retina or at the level of theLGN? More specifically do the areas satisfy the abovethree criteria?
The non-conscious processing areas have therefore tobe delineated and separated from those involved in theemergence of a percept. That an experiment cannot dothis will be taken as indication it will not be a crucial onefor the program. However, the results could still be ofgreat importance in determining the details of prepro-cessing necessary to build a more complete picture,both experimental and model-based, which can be usedto probe the emergence of a percept in more detail.
3.1. Visual Modality Experiments
3.1.1. Motion After-Effect (MAE).This paradigm usesthe relation between the perceptual and blood flow timecourses of the decay of the adaptation to continuousmotion, as in the case of looking at a waterfall for 30seconds and then viewing the adjacent stationary rockface; the latter appears to move upwards. We have con-jectured in the previous section that the MAE perceptarises in that particular region which has the slowestsuch decay. It may be that further regions, with bloodflow activity decaying as slow as or slower than theperceptual one, are crucially involved in percept forma-tion; it is the observation of these further regions thatmust be done in more detail. That may also be observedby MEG analysis of the faster processing between theregions and the more detailed nature of that interaction.
In any case the experimental paradigm is satisfactory,as far as criteria 1 to 3 above are concerned (and hasalready been used to good effect in Tootall et al.,1995). It involves direct comparative analysis of timingeffects across different areas.
3.1.2.Orientation-Specific Adaptation.Continued view-ing of oriented stripes (for over 5 or so seconds) bringsabout adaptation to them, as observed by an increase ofthe contrast threshold for detection of stripes oriented inthat direction as compared with those in a perpendiculardirection (He et al., 1996). This effect has been shown,by means of ‘‘crowding’’—the input of further sets oforiented stripes in adjacent regions of visual space so asto cause a loss of awareness of the direction of orienta-tion but not loss of non-conscious knowledge of thatorientation—to be present in spite of the loss of theconscious experience by human observers of the particu-lar orientation to which adaptation had occurred. Thepreservation of orientation adaptation knowledge, inspite of the loss of awareness due to crowding, indicatesthat ‘‘visual awareness, and the crowding that blocks it,occur after orientation analysis in the visual informationprocessing stream’’ (He et al., 1996). Moreover, theasymmetry of crowding (being stronger in the upperhalf visual field) can only arise from areas after V1
1188 J. G. Taylor and H.-W. Mueller-Gaertner

(and even V2) since these areas are both symmetricalbetween the upper and lower half fields.
These effects—the differential placement of orienta-tion adaptation (in V1 and V2) and crowding in the visualprocessing stream—lead to search for the placementof the computation of crowding itself. Let us supposethat it can be determined to be in a particular visualarea, Vn say. Does that imply that consciousness ofcrowding must also occur in Vn? It is possible thatthe crowding effect is itself computed in an earliermodule than would correspond to the working memorysite of consciousness of the two-stage model of Figure 1.In other words, consciousness could arise at a laterstage than the initial computation of the crowding effect,so that the first module performing the computation ofthe effect is not a ‘‘tag’’ for the emergence of awareness.It would be necessary to determine the time course ofactivity in order to pin down which module (or modules)were more precisely involved in the emergence of aware-ness of AM, following the criteria set out in Section 2.
In conclusion, the adaptation paradigm itself is notsatisfactory to obtain the source of awareness. Thecrowding effect, probed as to its temporal course acrossvarious modules involved in its computation, does seemto be so. In that form it comes under subliminal pro-cessing, and as such will be considered in more detailin Section 3.3.
3.1.3. Apparent Motion (AM).This is a well-knownparadigm in which a spot of light, say being shone at aparticular position on a screen, is deleted and almostimmediately (within about 300 msec) shone at anearby point. This produces in the subject the experi-ence of motion of the spot from the first to the secondposition.
The difficulty with the experiment as one for detectingthe emergence of the awareness of motion is that it usesthe motion pathway all the way up from V1 (and below,in LGN) due to the rapid onset and offset of the twostimuli, so that even when the experience of motionarises in the subject (when the spots have been movedclose enough together for the motion percept to occur)there may be use of areas already activated by the suddenonset and offset which are involved in both non-con-scious and conscious processing. The times of onsetand offset might be used to separate out the motioneffects they cause from the activation due to the furthermotion experience. However, this may need high spatialresolution.
There is also the difficulty that the initial lowestlevel computation of the track of motion may beachieved at a lower level than the computation of aware-ness; this is the same problem as that met above inorientation adaptation. It would be necessary to use thefurther criterion of temporal dynamics to ascertain themodules turned on by the AM signal which also held itfor a suitably long temporal duration.
In conclusion, the AM paradigm is only satisfactoryif its temporal dynamics is also probed with suitableprecision.
3.1.4.Blindsight.The case of GY (or similar subjects),for whom there is a separation of ‘‘contentless aware-ness’’ as compared with knowledge without awareness,as stimulus speeds are altered (Weiskrantz et al., 1995),is an excellent one to observe possible regions which areactivated by one paradigm compared with the other.This corresponds precisely to the case of subliminalversus conscious perception considered below. Thereis, however, the concern that there may be a considerabledifference in the modules involved in such subjects ascompared with those in the emergence of perception innormal humans. A blindsighted person may well havedeveloped new connections and usage of modules thatare different than those used in normally sighted peoplein the creation of awareness.
In conclusion, the paradigm is satisfactory (modulothe problem raised above).
3.1.5.Normal Blindsight.The phenomenon of blindsight,in which there is knowledge without awareness, is onewhich has been activated in normal subjects by means ofthe experimental paradigm of Kolb and Braun (1995). Inthis paradigm, a field of short oriented bars (or oppositelymoving dots), all with the same orientation (or directionof motion), is embedded in a set of similar but oppositelyoriented (or moving) objects. There is knowledge ofthe position of the small embedded subset of stimuluspatterns, but no awareness occurs. It would be possibleto modify the stimulus parameters, by increasing thesize of the region of the embedded stimuli, so as tocause the emergence of perceptual experience in a con-trolled manner.
This experimental paradigm is seen to suffer from thedifficulty that there is already considerable activationalong the whole of the dorsal pathway caused by thesurrounding pattern, of which the subject is clearlyaware. The modification of this activity when there is‘‘normal blindsight’’, as compared with when thereis not, will therefore be submerged in a large level ofactivity involved with normal perception.
In conclusion, the paradigm is not satisfactory (due tothe low signal for the signature of the emergence ofawareness). It is also relevant to note that the paradigmis part of that of subliminal processing, so will be con-sidered in more detail in Section 3.3.
3.1.6. Rivalry/Binocular Rivalry.Rivalry of concepts,such as arises in the Necker cube, has a long history ofpsychological investigation. The case of binocular riv-alry has also been used in humans (Kolb and Braun,1995) and in monkeys (Logothetis and Schall, 1989;see also Sheinberg and Logothetis, 1997) to attempt toascertain where the computation occurs in cortex for the
1189Non-Invasive Analysis of Awareness

production of the rivalrous percepts as they alternate.This has not yet proved possible. Yet there is a similarproblem to that raised above for orientation adaptationand AM: it is necessary to probe the temporal dynamicson the modules involved very carefully.
In the case of switching between the two images in theNecker cube, the computation could occur at a levelbelow that of awareness. Thus if the mechanism wereof simple adaptation of one percept to give way to theother, for example through some form of competitiveprocess, such adaptation may take place on a module atthe NonAware level of Figure 1, with output being usedto feed onto the further Awareness module A in thatfigure. This would not correspond precisely to the elabo-rated two-stage model of Figure 2, where disambigua-tion takes place only at the same level as awareness onthe working memory site WM in that figure. However,it may be that there is some decoupling betweendisambiguation and temporal duration in the visualmodality, especially due to the large number of codesinvolved as compared with the auditory one. The crucialtag of consciousness was claimed in Section 2 to be thatof the extended temporality of neural activity; the dis-ambiguation process is not necessarily associated withthe onset of awareness (nor will it be so easy to identify).
In conclusion, it would be necessary to determinedetailed temporal characteristics of neural activity duringrivalrous phenomena; those modules in which there istemporal duration highly correlated with the oscillationbetween the concepts (but having the highest such dura-tion, even extending beyond the switch-over times)would be the best candidates for the site of awareness.The observation of such sites may be difficult, however,due to highly similar coding which may occur for therivalrous concepts. Thus, the experimental probing ofsuch phenomena is of great interest but not at the topof the list of crucial experiments to probe awareness.
3.1.7. Stabilised Images.When images are stabilisedon the retina they are know to fade (Pritchard, 1963).The manner in which they do so has very interestingcharacteristics, such as would seem to imply that bothGestalt and Hebbian ensemble actions are at work. Thishas been explained by Taylor (1996a) in terms ofseparate loss of activation of the ‘‘what’’ and ‘‘where’’pathways. It would be possible to attempt to determinewhich regions lose activation as the percept fades fromawareness.
It is unclear, when a stabilised image drops out ofperception, that it does so only to the next lower levelof non-conscious activation. It may be that there is noactivity even from LGN, or from any other low-levelmodule, where the most crucial levels of adaptationmay occur. The discovery of the set of modifications(which area is on, which off, when the stabilised imageblinks off) is of great interest, but not able to provide thedesired answer.
In conclusion, it can only be said, for the above reason,that the paradigm is not satisfactory.
3.2. Auditory Modality
3.2.1. The Double Click Experiment.The double clickexperiment involves a subject attempting to detectthose double clicks (with interstimulus delivery timesof 0, 6, 12, 18 and 24 msec) presented to the earswhich are detectable as double clicks and not singleones. At an interstimulus interval larger than about12 msec there appears to be both double click detectionand a phase shift in the observed 40 Hz signal observedby MEG (Joliot et al., 1994). Let us suppose it to bepossible to detect the region involved in the computationof the double click interstimulus interval to produce sucha phase shift. Is that necessarily the region where aware-ness of the double click arises?
The answer is no, since it is possible to consider thatthe computation of the interstimulus intervals was per-formed at a lower non-aware level and sent on to theAwareness module of Figure 2. Such a possibilitycould be guarded against if there was a control experi-ment performed which required the subject to be dis-tracted by a different stimulus load (say by countingdown by threes from one hundred). The resulting activitywould show whether or not the activity was still the same(the phase shift still having occurred at about 12 msecinterstimulus interval). If the activity were still in thesame cortical area then the computation of the ISIwould clearly have been at a non-aware level. But ifthere were a difference in the sites of activity betweenthe attended and distracted conditions it would stillnot be the case that the residual activity, after subtractingthe aware from the distracted condition, would leavemodules activated solely at an aware level. The distrac-tion could have prevented processing in some parts ofthe non-aware circuitry but not throughout it all; the restcould still be producing the stimulus timing signal tosend up to the awareness level.
In conclusion, again temporal duration must be usedas a critical tag to separate non-aware from awareprocessing.
3.2.2.Binaural Fusion.This phenomenon involves theobservation and comparison by means of MEG of thoseregions activated either by a tone or by a set of harmonicsof the tone which lead to the same percept of the soundin a subject (Pantev et al., 1989). This experimentalparadigm has only been performed under attentive pro-cessing conditions, so it is unclear that the crucial regionswere at an aware level. If the experiment were also to beperformed, as in the double click experiment, under adistracting load then one would still have the difficultyraised in the previous paradigm, that the computation,after subtracting the activity that was occurring underthe distracting load, would not necessarily be at the
1190 J. G. Taylor and H.-W. Mueller-Gaertner

level of the awareness circuitry of Figure 1, but couldstill be down in the non-aware modules.
In both of these paradigms it would be necessary todetermine timing effects with care. Thus in conclusion,the paradigm is interesting but does not single themout as leading automatically to the non-aware/awareboundary.
3.3. Subliminal Perception
There are different regions devoted to supporting sub-liminal as compared with aware activity, as Figure 1denotes. Thus confounding the two forms of activitywill make the flow of information in the brain very diffi-cult to understand. This is particularly important as moredata is gathered from the non-invasive instruments. Itwill become more and more difficult to comprehendwithout a proper distinction between low and top-levelprocessing being introduced. Otherwise the data will justpile up to increase confusion over its interpretation.
Such seems to be the case in lexical processing(though not involving non-invasive studies yet), whererecent results (Levelt et al., 1991) seem to contradict thesimple connectionist approaches to lexical priming (inparticular, the expected broad spread of activation atphonological level to semantic associates of an inputword or target to be recognised). Yet these analysesand theoretical frameworks are not based on standardunderstanding of the phonological loop and its relevanceto conscious experience. That needs to be developed onsure experimental grounds. The same situation will occurin vision and other modalities, although the theoreticalstructures are not as well developed to be able to appreci-ate the problem as effectively.
3.3.1. Fully Subliminal Versus Partly SubliminalParadigms.One experimental paradigm to determinethe extra machinery which divides perception withoutawareness from that with it would thus be to subtractresults obtained from experiments on stimulus detectionin which there is only a subliminally perceived stimulusfrom those obtained when awareness has occurred. Theneuronal modules activated in giving the extra feature ofaware beyond that of non-aware processing would thenbe detectable as having higher activation levels after sub-traction, or more general comparison through a suitablecorrelational technique.
An alternative or additional paradigm is to consider anexperiment in which there are different effects arisingfrom stimuli perceived at the two levels (as in the Stroopeffect). The results of such an experiment will have to beinterpreted with care so as to be able to distinguishbetween the aware level of processing and that at anon-conscious level; certain paradigms should allowthe separation of modules involved at the two differentlevels by means of use of different paths in the brain (asin foveal versus parafoveal experiments) or different
parts of a module (as in the ‘‘filling in’’ phenomenonin audition).
The first of these paradigms will be termed ‘‘fullysubliminal’’ experience, the second ‘‘partly subliminal’’experience. These lead clearly to different experimentalparadigms and to different criteria on the stimulusdelivery systems. That will be explored below. Thepartly-subliminal is a paradigm which will be discussedfurther in Section 3.3.3 after the consideration of the fullsubliminal stimulus paradigm in the next section.
3.3.2. Fully Subliminal Stimuli.The basic idea behindsuch a visual stimulus delivery paradigm is that they bepresented to subjects on a screen in front of them so thatsome of the stimuli are experienced at a subliminal level,whilst, in comparison and at different times, other stimuliare to be fully experienced. To ensure that a visualstimulus is subliminally experienced it is necessary topresent the stimulus for a brief time, say 50 msec, andthen flash another stimulus, called the ‘‘masking stimu-lus’’, on the screen at a controlled time afterwards so asto persist for, say 500 msec. The time between the offsetof the first and onset of the masking stimulus, the so-called stimulus onset asynchrony (SOA), needs to bevaried in steps of 5 msec to be effective in changingthe level at which a given subject is able to perform atjust above chance level in a lexical decision task (decid-ing whether the letter string composing the first stimulusis a word or not; Marcel, 1980).
Fully subliminal perception occurs in at least themodalities of touch, vision and audition, and there is aconsiderable psychological literature on the nature ofthese phenomena (Merikle, 1992).
The important problem of stimulus attention (so as tokeep attention aroused even in the apparent absence ofany stimulus and allow stimulus timing to be effective)may be approached by the following choice of paradigm:require the subject to respond, after the offset of themask, to one of the questions:
1. was there a stimulus preceding the mask?2. to which of two probe words (presented after offset of
the mask) was that which preceded the mask mostsimilar graphically?
3. towhich word (as in theparadigm for (2)above) was theword preceding the mask most similar semantically?
In this manner, attention can be kept focused onto agiven task (the final response) yet neither the subliminalstimulus nor the mask should be perturbed by theresponse of the subject.
This is valid for visual stimuli, but can be extended tosomatosensory or auditory stimuli (in the former casewith stimuli possibly delivered to one or other hand, orto two sites on the hand, and ask which hand or which sitewas stimulated, whilst for audition the stimulus couldbe delivered to either ear, with response to the questionas to which ear).
1191Non-Invasive Analysis of Awareness

3.3.3.Partly-Subliminal or Unattended Stimuli.A formof this paradigm particularly relevant to fMRI experi-mentation is that of two words presented to a subject,one foveally and the other parafoveally, for 120 msec,with attention being paid to the foveally presented word.At an inter-stimulus interval (ISI) of 200 or 2000 msec atarget word is presented with the request to classify thestimulus as an animal or not by pressing one or other oftwo buttons, in the paradigm of DiPace et al. (1991).Reaction time measurements to the response at the earlierISI should show priming from words semanticallyrelated to either the foveally or the parafoveally pre-sented stimuli; at the later ISI only the foveally presentedprime is effective. This was found to be so experimen-tally. The activation of the parafoveally presented wordhas died away before it has had time to reach awareness.This was also determined by asking the subjects if theyhad experienced the parafoveal word; in only 15% ofcases did this happen.
There are many other stimuli which produce partlysubliminal stimuli; in vision, the Stroop test has beenmentioned. In audition, the use of a prime, which isattended to, and a probe which can then be used as asubliminal modifier of response to the attended prime,is common in lexical analysis, and again there is a veryextensive literature. The accuracy of timing in theselatter cases is only at the 20 msec level or above.
3.3.4.Conclusions on Subliminal Stimuli.There has beenconsiderable analysis of the phenomenon of subliminalperception, in which there is knowledge acquired for
inputs below a so-called ‘‘objective threshold’’ butonly awareness when the input is above a higher ‘‘sub-jective threshold’’ (Cheesman and Merikle, 1986;Merikle, 1992). Thus the phenomenon would itselflead to the analysis of the emergence of perceptionby attempting to detect, under one of the numerous
1192 J. G. Taylor and H.-W. Mueller-Gaertner
FIGURE 3. Coupling between two sets of two-stage modules of Figure 1 and Figure 2 to achieve competition between the activities on thesecond stages through the interaction of those stages with the NRT-thalamic system. There is also support from episodic memoriesencoded in the store denoted E in the figure.
FIGURE 4. The difference between subliminal processing (a) andaware processing (b); in the former activity fails, for some reason(low level, degradation, backward masking), to reach the second,working memory stage of the construction of consciousnessdenoted in Figures 1–3.

paradigms in which it has been observed, the differencebetween those areas involved in the subliminal level ofprocessing and those involved in the further creationof awareness in the subject.
The nature of these paradigms, in terms of the two-stage processing model of Figure 2, is as shown inFigure 4. The first figure shows purely subliminalprocessing, in which the awareness modules ‘‘A’’ ofFigure 2 are not activated; the second figure indicateswhat happens when activity also arises at the awarelevel during conscious processing.
Theseseemtobethebestofall possibleparadigms,sincethey directly attack the separation of tasks into thoseinvolved with subliminal, just pre-conscious, activationsfrom those in which there is awareness of the inputs.
In conclusion, these subliminal paradigms appear to behighly satisfactory. The optimal ones involve the fullysubliminal stimuli of Section 3.3.2, since there is thenless activity expected to arise at the aware level in theprocessing of the purely subliminal stimuli. In the partlysubliminal stimulus case, there would be expected to bepossible interference of the attended stimulus processingat the subliminal level. It would be relevant to developparadigms in which such interference is expected to beminimised.
4. CONCLUSIONS
Of a range of experimental paradigms analysed to probethe emergence of perception, very few were found tosatisfy the criteria presented above in Sections 2 and 3.A summary of the results of the above discussions isgiven in Table 2. In particular, the remaining successfulexperiments were:
1. MAE2. subliminal processing3. dissociated blindsight4. comparison of activations caused by various inputs
between attentive and non-attentive conditions.
Of these, (1) and (2) seem to be the most precise andclean, since they involve direct measurement of regionsof long holding of the neural activity directly causing theemergence of the relevant percept. In (3), there is also thedanger that blindsighted subjects could have reroutedtheir activity to other modules, so the neural sites dis-cerned would not necessarily be those used by sightedsubjects. In (4), there is the danger of only partiallyattended or unattended processing.
Given that there are two paradigms which wouldappear to be effective, we can conclude that thereare some paradigms which satisfy all of the criteria ofSections 2 and 3. The next steps are (a) to perform andanalyse the experiments in detail and (b) to model thedetailed dynamics observed, so as to obtain a betterunderstanding of the complex phenomena involved, andso as to make detailed predictions, from the models, inorder to extend the experimental understanding.
REFERENCES
Baars, B. (1988).A cognitive theory of consciousness. Cambridge:Cambridge University Press.
Baddeley, A. D. (1986).Working memory. Oxford: Oxford UniversityPress.
Baddeley, A. (1993). Working memory and conscious awareness. InS. E. Gathercole, A. F. Collins, M. A. Conway, & P. E. Morris(Eds.),Theories of memory(pp. 11–28). Hillsdale, NJ: Erlbaum.
Bisiach, E., & Berti, A. (1995). Consciousness in dyschiria. InThecognitive neurosciences(chap. 88, pp. 1331–1340). Cambridge,MA: MIT Press.
Bullier, J. (1995). Parallel versus serial processing: New vistas on thedistributed organization of the visual system.Current Biology, 5,497–503.
Cheesman, J., & Merikle, P. M. (1986). Distinguishing conscious fromunconscious perceptual processes.Can. J. of Psychology, 40,343–367.
DiPace, E., Longoni, A. M., & Zoccolotti, P. (1991). Semantic pro-cessing of unattended parafoveal words.Acta Psychologica, 77,21–34.
Edelman, G. (1989).The remembered present. New York: Basic Books.Felleman, D. J., & Van Essen, D. C. (1991). Distributed hierarchical
processing in the primate cerebral cortex.Cerebral Cortex, 1, 1–47.
1193Non-Invasive Analysis of Awareness
TABLE 2
Paradigm G/NG Reason/Comments
1) Blindsight G But may have altered circuits2) MAE G Area with longest duration of adaptation (& longer than
psychophysical effects)?3) Binocular rivalry NG Adaptation in module
Concept rivalry NG Below creation of ‘‘A’’?4) Orientation-specific adaptation NG Adaptation not create ‘‘A’’5) Double click NG Where is ‘‘A’’?
G If use non-attentive conditions in addition6) Filling in NG As for double click
G If use non-attentive conditions in addition7) Non-attentive G Must ensure full non-attention (but attentive ‘‘A’’)8) Subliminal processing (many paradigms) G Must ensure only subliminal experience is occurring
G denotes ‘‘good’’, indicating that the paradigm is effective in discerning the location of the emergence of phenomenal awareness; NG denotes ‘‘no good’’).

He, S., Cavenagh, P., & Intrilligator, J. (1996). Attentional resolutionand the locus of visual awareness.Nature, 383,334–337.
James,W. (1950).Theprinciplesofpsychology.NewYork:DoverBooks.Joliot, M., Llinas, R., & Ribary, U. (1994). Human oscillatory activity
near 40 Hz coexists with cognitive temporal binding.Proc. Nat.Acad. Sci. (USA), 91, 11748–11751.
Kolb, F. C., & Braun, J. (1995). Blindsight in normal observers.Nature,377,336–338.
Levelt, W. J. M., Schriefers, H., Vorberg, D., Meyer, A. S., Pechmann,T., & Havinga, J. (1991). The time course of lexical access in speechproduction: A study of picture naming.Psychological Review, 98,122–142.
Logothetis, N.K., & Schall, J.D. (1989). Neuronal correlates of subjec-tive visual perception.Science, 245,761–763.
Marcel, A. (1980). Conscious and preconscious recognition on poly-semous words: Locating selective effects of prior verbal contexts. InR. S. Nickerson (Ed.),Attention and performance VIII. Hillsdale,NJ: Erlbaum.
Merikle, P. M. (1992). Perception without awareness.AmericanPsychologist, 47, 792–795.
Pantev, C., Hoke, M., Lutkenhoner, & Lehnertz, K. (1989). Tonotopicorganization of the auditory cortex: Pitch versus frequencyrepresentation.Science, 246,486–488.
Paulesu, E., Frith, C., & Frakowiak, R. S. J. (1993). The neural corre-lates of the verbal components of working memory.Nature, 362,342–345.
Pritchard, R.M. (1963). Stabilised images on the retina.ScientificAmerican, 204,72–78.
Salmon, E., Van der Linden, M., Collette, F., Delfiore, G., Maquet, P.,Degueldre, C., Luxen, A., & Franck, G. (1996). Regional brainactivity during working memory tasks.Brain, 119,1617–1625.
Shah, J. (1997). Private communication.Sheinberg, D. L., & Logothetis, N. K. (1997). The role of temporal
cortical areas in perceptual organisation.Proc. Nat. Acad. Science(USA), 94, 3408–3413.
Smith, E. E., & Jonides, J. (1995). Working memory in humans:Neurophysiological evidence. In M. Gazzaniga (Ed.),The cognitiveneurosciences(chap. 66, pp. 1090–1020). Cambridge, MA: MITPress.
Taylor, J. G. (1973). Seminar, Tubingen University (unpublished).Taylor, J. G. (1991). Can neural networks ever be made to think?.
Neural Network World, 1, 4–11.Taylor, J. G. (1992). Towards a neural network model of mind.Neural
Network World, 2, 797–812.Taylor, J. G. (1996a). Invited talk, WCNN96, San Diego, CA.Taylor, J. G. (1996b). Breakthrough to awareness: A preliminary neural
network model of unconscious perception in word processing.Biol.Cybernetics, 75, 59-72.
Taylor, J. G. (1998).The race for consciousness. Boston, MA: MITPress.
Tootall, R. B. H., Reppas, J. B., Dale, A. M., Look, R. B., Sereno, M. I.,Malach, R., Brady, T. J., & Rosen, B. (1995). Visual motion after-effect in human cortical area MT revealed by functional magneticresonance imaging.Nature, 375,139–141.
Weiskrantz, L., Barbur, J. L., & Sahrie, A. (1995). Parameters affectingconscious versus unconscious visual discrimination with damageto the visual cortex (V1).Proc. Nat. Acad. Sci. (USA), 92,6122–6126.
Young, A. W. (1994). Neuropsychology of awareness. In A. Revonsuo& M. Kamppinen (Eds.), Consciousness in philosophy andcognitive neuroscience(chap. 8, pp. 173–204). Hillsdale, NJ:Erlbaum.
1194 J. G. Taylor and H.-W. Mueller-Gaertner

1997 SPECIAL ISSUE
Three Centuries of Category Errors in Studies of the NeuralBasis of Consciousness and Intentionality
Walter J. Freeman
University of California at Berkeley
(Received2 September1996;accepted20 January1997)
Abstract—Recent interest in consciousness and the mind–brain problem has been fueled by technological advances inbrain imaging and computer modeling in artificial intelligence: can machines be conscious? The machine metaphororiginated in Cartesian ‘‘reflections’’ and culminated in 19th century reflexology modeled on Newtonian optics. Itreplaced the Aquinian view of mind, which was focused on the emergence of intentionality within the body, with controlof output by input through brain dynamics. The state variables for neural activity were identified successively withanimal spirits, elan vital, electricity, energy, information, and, most recently, Heisenbergian potentia. The source ofdynamic structure in brains was conceived to lie outside brains in genetic and environmental determinism. An alternativeview has grown in the 20th century from roots in American Pragmatists, particularly John Dewey, and Europeanphilosophers, particularly Heidegger and Piaget, by which brains are intrinsically unstable and continually createthemselves. This view has new support from neurobiological studies in properties of self-organizing nonlinear dynamicsystems. Intentional behavior can only be understood in relation to the chaotic patterns of neural activity that produce it.The machine metaphor remains, but the machine is seen as self-determining.q 1997 Elsevier Science Ltd.
Keywords—Brain dynamics, Chaos, Consciousness, Existentialism, Information, Intentionality, Nerve energy.
1. INTRODUCTION
Studies of the neural basis of consciousness haverecurred in the biomedical literature for 2500 years,beginning with Hippocrates:
One ought to know that on the one hand pleasure, joy, laugh-ter, and games, and on the other, grief, sorrow, discontent,and dissatisfaction arise only from [the brain]. It is espe-cially by it that we think, comprehend, see, and hear, that wedistinguish the ugly from the beautiful, the bad from thegood, the agreeable from the disagreeable... Furthermore,it is by [the brain] that we are mad, that we rave, thatfears and terrors assail us—be it by night or by day—dreams, untimely errors, groundless anxiety, blunders,awkwardness, want of experience. We are affected by allthese things when the brain is not healthy, that is, when it istoo hot or too cold, too moist or too dry, or when it has
experienced some other unnatural injury to which it is notaccustomed (Clarke and O’Malley, 1968, pp. 4–5).
The last strong outpouring from biologists came 40years ago following the discovery of the midbrain andthalamic reticular activating systems and their roles inarousal and attention (Adrian et al., 1954). In the pasttwo decades, contributions have come from researchersin brain imaging, psychology, psychiatry, neurology,philosophy, mathematics, physics, computer science,and artificial intelligence. As a result, a shift of immensemagnitude is taking place in our understanding of our-selves, but none of us has the perspective yet to grasp itsnature and significance. The limitation stems from thecircumstance that the bulk of new data has been obtainedwithin the confines of the machine metaphor for mind/brain function. This essay aims to explore the origin ofthat metaphor from what preceded it, and to indicate anew approach to mind/brain studies.
2. THE ORIGIN OF BRAIN DYNAMICS IN THEMACHINE METAPHOR
Behaviorists have a long history of using natural science
Neural Networks, Vol. 10, No. 7, pp. 1175–1183, 1997q 1997 Elsevier Science Ltd. All rights reserved
Printed in Great Britain0893–6080/97 $17.00+.00
PII: S0893-6080(97)00054-3
Pergamon
1175
Acknowledgements: This work was supported by a grant MH06686from the National Institute of Mental Health. The paper was read atthe First Tucson Conference on ‘‘Toward a Scientific Basis forUnderstanding Consciousness’’ on 13 April 1994. It providedmaterials for Chapter 2 in ‘‘Societies of Brains’’ (1995), and is givenhere with the permission of the publisher, Lawrence ErlbaumAssociates, Hillsdale, NJ.
E-mail:[email protected].

to explain what they can observe among animals andtheir fellow humans. The Greeks invented a chemistryof the soul, in which the four elements (air, earth, fireand water) combined to make the four humors of thebody (phlegm, blood, black bile and yellow bile), whencethe Hippocratic temperaments: phlegmatic, sanguine,melancholic and choleric. Aristotle applied his physicsto conceive the brain as a radiator to cool the blood.The foundation for a dynamics of behavior was laidby Descartes, who proposed that the brain operated asa pump for fluids termed ‘‘animal spirits’’, which flowedthrough the ventricles from the brain into the spinalcord and out into the muscles. The valves such as thepineal gland were controlled by the soul in humansbut were automatic in animals, they being soullessmachines.
The seminal importance of this formulation can beseen from the following standpoint. In a dynamicalsystem some material thing moves with time. SinceNewton and Leibniz the preferred description hasbeen a differential equation, which relates time as itelapses independently (in either direction) tosomething in the system that is changing. That ‘‘some-thing’’ is the state of the system, and its descriptor is a‘‘state variable’’. It is essential to measure accuratelyboth the time lapse and the state variable, because theequation gives the relation between the numbers thatrepresent time and the state variables. In Cartesianstudies of the brain, the state variable would have repre-sented the flow of animal spirits, had the equationsbeen written. No one did this. The difficulty was thatthere was no way to measure the flow of animal spiritsthrough the nerves. Descartes postulated that the muscleswere shortened by being pumped full of the fluid like aballoon. Physiologists tested this prediction by invent-ing the plethysmograph to measure the volume of musclebefore and during contraction. There was no increasebut, in fact, a slight decrease owing to expulsion ofblood from the veins by muscle contraction, whichshowed that animal spirits had no volume to measure.Hence animal spirits could not be represented by a stringof numbers in a model. This is an example of whatGilbert Ryle (1949) called a category error, in thiscase the assignment of the physical property of volumeto an immaterial entity. There were more such errorsto follow.
Late in the 18th century a Bohemian ophthalmologistnamed Giri Prochaska made a remarkable discovery.Until that time most scientists had assumed that thebrain was a source of animal spirits. Prochaska observedthe behavior of newborn anencephalics and found theirbehavior to be entirely normal. As we now know thehuman cerebrum is essentially nonfunctional at birth.His findings led him to propose that sensory receptorsare the source of animal spirits, which are released by theaction of stimuli from the environment. He then drewexplicitly on Newtonian optics to formulate a theory of
reflex action:
The reflection of sensory impressions into motor...is notperformed solely according to physical laws...but followsspecial laws inscribed, as it were, by nature on the medullarypulp... The general law...is that of our preservation: so thatcertain motor impressions may follow external impressionsabout to harm our body and produce movements aimed atwarding off and removing the harm...or sensory impressionsabout to be favorable to us, and produce movement tendingto preserve that pleasant condition longer (Prochaska, 1784,p. 116).
The brain was seen as complex but passive mirror.‘‘Reflections’’ became reflexes. Three decades laterProchaska made another category error in identifyinghis ‘‘sensory impressions’’ with electricity newlydiscovered by Galvani and Volta. This hypothesis wasdisputed by Carlo Matteucci, who maintained thatnerves carried spiritual force, that came to be identifiedwith ‘‘e lan vital’’.
3. NERVE ENERGY REPLACES VIS NERVORUM
The hegemony of physics was re-established by theYoung Turks. Du Bois-Reymond discovered the injurycurrent and the ‘‘negative variation’’ (the nerve actionpotential). Helmholtz measured its conduction velocity.Sechenov developed an animal model for anencephalyby pithing a frog to study reflexes. The centerpiece ofthis antivitalist movement was the experimental demon-stration of the First Law of Thermodynamics, the con-servation of energy. In the grandest category error ofthem all, the animal spirits and e´lan vital were replacedwith nerve forces and nerve energies, which flowedfrom the environment through the sensory receptorsinto the brain and back out again through the muscles,after being stored in nerve cells and then ‘‘liberated’’.The new doctrine was announced by Herbert Spencer(1863, p. 109):
[It is]...an unquestionable truth that, at any moment, theexisting quantity of liberated nerve-force, which in aninscrutable way produces in us the state we call feeling,must expend itself in some direction—must generate anequivalent manifestation of force somewhere... [A]n over-flow of nerve-force, undirected by any motive, will mani-festly take the most habitual routes; and, if these do notsuffice, will next overflow into the less habitual ones.
Charles Darwin (1872, p. 70), continued:
This involuntary transmission of nerve force may or may notbe accompanied by consciousness. Why the irritation ofnerve-cells should generate or liberate nerve force is notknown; but that this is the case seems to be the conclusionarrived at by all the greatest physiologists such as Mueller,Virchow and Bernard, and so on.
The application of Newtonian dynamics was alsoexplicit in the writings of J. Hughlings Jackson (1884,
1176 W. J. Freeman

pp. 42–44):
...we speak of the dynamics of the nervous system... Anormal discharge starting in some elements of the highestcentres overcomes the resistance of some of the middle, nextthe resistance of some of the lowest centers, and the musclesare moved... A fit of epilepsy is an excessive caricatureof normal physiological processes during what is called avoluntary action... We have, in the case of ‘‘discharginglesions,’’ to consider not only the quantity of energy liber-ated, but the rate of its liberation...Resistanceswill beconsidered later.
A note in Jackson’s handwriting was later found in themargin of this text: ‘‘No more of this was published.’’ Itmay have already become clear that while nerve tissuedid have electrical resistance, the barrier to the flow of‘‘nerve energy’’ was not so simple. The principle of theconservation of momentum was also used by SigmundFreud as a foundation for his project of a scientificpsychology, in which he confused his neuronic inertiawith dendritic current:
This line of approach is derived directly from pathologicalclinical observations, especially those concerned withexcessively intense ideas... These occur in hysteria andobsessional neurosis, where, as we shall see, the quantitativecharacteristic emerges more plainly than in the normal...What I have in mind is the principle of neuronic inertia,which asserts that neurones tend to divest themselvesof quantity (Q)... We arrive at the idea of a ‘‘cathected’’neurone (N) filled with a certain quantity... The principleof inertia finds expression in the hypothesis of a current,passing from the cell-processes or dendrites to the axone...The secondary function [memory] is made possible bysupposing that there are resistances which oppose dis-charge...in the contacts [between the neurones] which thusfunction as barriers. The hypothesis of ‘‘contact-barriers’’ isfruitful in many directions (Freud, 1895, pp. 356–359).
Two years later these barriers were named by MichaelFoster and Sir Charles Sherrington:
Such a special connection of one nerve-cell with anothermight be called a synapsis (Foster and Sherrington, 1897,p. 929).
Some four decades later the hypothesis of synapticresistance was undermined by Otto Loewi’s discoveryof chemical neurotransmission, though it persists inmore realistic treatments of electrical synapses, inwhich it refers to electrical current and not to nerveenergy.
Another physical principle, the field of potential thatwas developed by Michael Faraday to explain electricaland magnetic forces, was coopted by Gestalt psycholo-gists to explain their data from studies in perception:
...let us think of the physiological processes not as molecu-lar, but as molar phenomena... Their molar properties will bethe same as those of the conscious processes which they aresupposed to underlie (Koffka, 1935, p. 57).
The task of psychology...is the study of behavior in itscausal connection to the psychophysical field (Koffka,1935, p. 67).
The environment was conceived as a source of nerveenergy, which flowed through the sensory receptors intothe brain with striking motivational consequences:
...things in our environment tell us what to do with them...Their doing so indicates a field of force between theseobjects and our Egos...which...leads to action. ...A handlewants to be turned, ...chocolate wants to be eaten, ...(Koffka, 1935, p. 353).
One of the principle architects of Gestalt psychology,Wolfgang Kohler, presented meticulous studies of inter-active phenomena in perception:
Our present knowledge of human perception leaves nodoubt as to the general form of any theory which is to dojustice to such knowledge: a theory of perception must be afield theory. By this we mean that the neural functions andprocesses with which the perceptual facts are associated ineach case are located in a continuous medium (Ko¨hler,1940, p. 55).
He pressed further into physics by identifying theperceptual fields with the electrical fields of the newlydiscovered electroencephalogram. This hypothesis wasidentical in form to the category error of Prochaska. Itwas quickly disproved by Roger Sperry (1958), whoplaced strips of mica and silver needles in the visualcortex of trained cats and monkeys and showed that theresulting distortions in electrical fields had negligibleeffects on behaviors involving visual perception. Unfor-tunately, for this and other reasons, the body of Gestalttheory was discredited among neurobiologists.
With continuing advances in the analysis of anatomi-cal pathways in the cerebrum it became increasinglyobvious that the concept of mass flow of energy madeno sense. According to Lashley (1942, pp. 302–306):
Generalization [stimulus equivalence] is one of the primi-tive basic functions of organized nervous tissue. ...Here isthe dilemma. Nerve impulses are transmitted...from cellto cell through definite intercellular connections. Yet allbehavior seems to be determined by masses of excitation...What sort of nervous organization might be capable ofresponding to a pattern of excitation without limitedspecialized paths of conduction? The problem is almostuniversal in the activities of the nervous system.
He had already noted the difficulty of finding usefulconcepts:
...expressions like mass action, stress patterns, dynamiceffects, melodies of movement, vigilance, or nervous energy[are] all highly metaphorical and unproductive of experi-mental problems (Lashley, 1929, p. 254).
Yet he continued to borrow from the physical sciences
1177Three Centuries of Category Errors in Studies

fuzzy concepts such as ‘‘reverberatory circuits’’,‘‘equivalent nervous connections’’, ‘‘systems of spacecoordinates’’, ‘‘wave interference patterns’’, ‘‘tunedresonating circuits’’, etc. (Lashley, 1950).
4. INFORMATION REPLACES NERVE ENERGY
The repeated failure of the energy metaphor opened theway for a new approach that came from the communi-cation sciences. Basing their work on Golgi analyses ofthe entorhinal cortex by Rafael Lorente de No´ (1934),Warren McCulloch and Walter Pitts introduced the con-cept of nerve cells operating as binary switches in neuralnetworks to compute Boolean algebra. John vonNeumann used this concept to develop programmabledigital computers. Shannon and Weaver developed thetheory of information bydivorcing it from meaning.This led to the replacement of ‘‘energy’’ by ‘‘informa-tion’’ as a descriptor of neural activity. Informationand energy are both conceived as flows from environ-mental ‘‘sources’’. They are transduced through sensorysystems, transmitted by axonal tracts as channels, carriedby action potentials (bits), transformed (processed) inbrains by synapses working as binary switches, storedas fixed patterns (representations), recalled by read-outunder constraints of finite channel capacities and entropiclosses, like the content addressable memories in compu-ters, and matched or cross-correlated with new inputpatterns.
Information metaphors are found at four levels. At thelevel of nerve cells the single neuron is seen as generat-ing a pulse train to represent a meaning correspondingto the Logical Positivist element of a word or an object,such as a grandmother. The frequency of the train repre-sents the probability that the object is present (Barlow,1972). Members of a distributed collection of neuronsthat symbolize the same object are called ‘‘cardinalcells’’, deriving from a College of Cardinals runningthe brain, as distinct from the pontifical cell decried bySherrington (1940, pp. 177–178):
In the great head end which has been mostly darkness springup myriads of twinkling stationary lights and myriads oftrains of moving lights of many different directions... Thebrain is waking and with it the mind is returning. It is as ifthe Milky Way entered upon some cosmic dance. Swiftlythe head-mass becomes an enchanted loom where millionsof flashing shuttles weave a dissolving pattern though neveran abiding one; a shifting harmony of subpatterns.
An alternative formulation holds a neuron to be a‘‘feature detector’’ by virtue of its afferent synapticconnections, which are modified and adaptively shapedby prior learning. A collection of feature detectorsdefines an object when their pulse trains become syn-chronized, a proposed solution to ‘‘the binding problem’’(Milner, 1974; von der Malsburg, 1983) of gettingfeature detectors to work together. Large arrays of such
neurons form Hebbian nerve cell assemblies and neuralnetworks, which provide the basis for neurocomputationor computational neural science. A well-known exampleis the tensor system for coordination of hand movementsunder visual guidance by cerebellar circuitry developedby Pellionisz and Llina´s and reviewed by Churchland(1986).
At the behavioral level and among successors of theGestalt school, most notable is J.J. Gibson, in whosework the ‘‘affordances’’ denote information that flowsinto the brain from outside the body through exterocep-tors and from inside the body through the proprioceptors:
...the affordance, being invariant, is always there to be per-ceived. An affordance is not bestowed upon an object by aneed of an observer and his act of perceiving it. The objectoffers what it does because it is what it is. ...But this does notin the least imply separate realms of consciousness andmatter, a psychophysical dualism. It says only that the infor-mation to specify the utilities of the environment is accom-panied by information to specify the observer himself...[E]xteroception is accompanied by proprioception...to per-ceive is to coperceive oneself (Gibson, 1979, p. 139).
Information is delivered into resonant circuits in thebrain, and it flows out again as ‘‘effectivities’’ frommuscles and glands in object-oriented actions. Accordingto Shaw et al. (1990, pp. 586–587):
Gibson, like Tolman, would disagree with [the view of]Skinner...that the organism is merely a ‘‘through-putsystem’’. For Tolman, cognition can embellish the stimulus,while for Gibson, stimulus must be informative about theenvironment in ways that a stimulus, as a physiological‘‘goad;’’ or a reflexive ‘‘force’’, could never be. Theyboth endow [the organism] with a complex interior—which Tolman cites as the residence of cognitive functionsand Gibson as the seat of a tunable (not necessarily linear)information detection operator which resonates to qualita-tive environmental properties (i.e. affordances). For Gibson,the environment that surrounds an organism is real andobjective for each given organism.
The metaphorical ‘‘resonance’’ is reminiscent ofKarl Lashley’s ‘‘tuned resonating circuits’’. It is meta-phorical, because no physiological embodiment has yetbeen demonstrated. The difficulties of pursuing thisline of theory have been well formulated by Shaw andhis colleagues. For example, the infinite complexity of‘‘objects’’ in unstructured environments is treated byconverting objects to numbers:
...by following Cantor’s fractalization rule, we have a wayto rescale continuous geometric objects so that theirdimensionality is reduced. Moreover, by following it withfinite recursion, we find that there are objects withoutinteger dimensions... These are Mandelbrot’s fractals...(Shaw and Kinsella-Shaw, 1988, pp. 197–198).
They resolve the paradox that is inherent in combiningpast experiences and future goals by postulating a dual
1178 W. J. Freeman

Minkowski space with the two cones of past and futuremelded at the apex of the present, and they propose:
...an environment of information which dynamically links asocially invariant exterior with both a biologically invariantinterior frame, on the one hand, and with a still more exteriorphysically invariant frame on the other. That psychologicalinverse dynamics must couple energy with informationacross a frame exterior (observable) to one and interior(controllable) to the other, and vice-versa, defines what ismeant by an ecological map (Shaw et al., 1990, p. 587).
Their proposed exploration of the role of intentionalityin purposive behavior by ecological psychology willdepend heavily on the development of graphical compu-ter displays for the three levels of maps and the deriva-tion of equations to describe the operations required fortransfer of information between them. At present, theirefforts are remote from direct observations made on thedynamics of brains.
At the subneuronal level, the discovery of DNA as thebasis for transmission of genetic information has alsostimulated search for stores of experiential informationin the form of ‘‘memories’’ in RNA molecules as wellas synapses. The search for the ‘‘molecular alphabet’’ oflearning is now among the hottest areas of neurobiologi-cal research (Alkon, 1992), although studies of synapticmodification have not yet progressed beyond habituation,facilitation, and simple go/no go reflex arcs. HolgerHyden demonstrated a change in RNA in the brains ofrats trained to climb a wire and suggested that it indicatedthe storage of a procedural memory in the neurons ofthe vestibular nuclei. This line of thinking culminatedin studies by worm runners to transfer the memory ofworking a T-maze from trained Planarian flatworms totheir naive, cannibalistic siblings. After initial successthis hypothesis failed in the hands of trained scientists,but it is still being ‘‘replicated’’ in high school sciencefairs annually across the country.
At the submolecular level is a variant of the informa-tion hypothesis, in which ‘‘quanta of sentience’’ emergeas Heisenbergian potentia from a ‘‘sea of universal con-sciousness’’ (Herbert, 1993, p. 26):
Though materialists agree that mind (defined as ‘‘innerexperience’’) is no more than a particular motion of matter,they differ concerning how complex matter’s movementmust be actually to produce a noticeable sensation, togenerate what might be called a ‘‘quantum of sen-tience’’...analogous to physicist Max Planck’s famousquantum of action.
The main hope that drives these investigations is fordiscovery of new laws of physics (Penrose, 1989), whichwill explain such paranormal phenomena as teleporta-tion, precognition, distance viewing, and related formsof extrasensory perception (Herbert, 1993, p. 248):
Most quantum models of consciousness are similar toCrookes’s coherer proposal in that they consider the synapse
to be a sensitive receiver of mental messages that originateoutside the brain. The main difference between the coherermodel of mind and quantum consciousness models is that...-mind is somehow resident in Heisenberg’s quantum potentiarather than in electromagnetic ether.
Criticisms that brains, neurons, organelles and recep-tor molecules are neither small enough nor cold enoughto afford quantum coherence have been met with therejoinder that superconductivity is a macroscopic stateof coherence that has already been achieved at tempera-tures approaching Siberian winter nights, and that it maysoon be found also at normal brain temperatures. Lesseasily handled have been the criticisms that, for all itspower in chemistry, quantum mechanics is a linear, first-order, discrete approximation for protons, that it is eveninadequate to describe the collapse of the wave function,and that it is poorly suited for describing the nonlinearcontinuous time dynamics displayed by the nervoussystem at all levels. More to the point, this line of thoughtis new wine in an old bottle. The same properties areinvoked as for energy and information: environmentalsources of input, sinks for output, tuned receptors toresonate with selected inputs, and connectionist mechan-isms for storage and retrieval. The elemental buildingblocks (reflexes, action potentials, bits, words, symbols,primitives, and quanta) change with the centuries, but theunderlying concepts have been passed whole from onegeneration to the next.
5. THE UNIQUENESS OF BRAIN FUNCTION
Three insights are lacking from these input–outputapproaches to brain dynamics. The first insight is thatthe tissue formed by neurons in animal brains is unique.There is no other substance like neuropil in the knownuniverse. Phylogenetically it has emerged by evolutionrepeatedly and independently in the brains of molluscs,crustaceans, and vertebrates, always as the basis foradaptive, goal-directed behavior. Being unlike anythingelse, it offers us the opportunity to discover its ‘‘laws’’,which might constitute the ‘‘new laws of physics’’sought by Penrose (1989). Second, it follows that themachine metaphor cannot be serve to identify brainstate variables with the state variables of any othermachine. Third, brains organize their own goals, whichmachines cannot now do.
The second insight holds that, while neural activity isbased in flows of transmitter molecules, inorganic ions,and electric currents fueled by metabolic energy andcontrolled by conformational structural changes in cellmembranes, and while it carries both meaning and infor-mation, it cannot be defined by any of these physicalor conceptual quantities. As noted by Karl Lashley theterms ‘‘nerve force’’, ‘‘nerve energy’’, ‘‘information’’,and ‘‘representation’’ aremetaphorsand not measur-able descriptors of brain events. The spatiotemporaldynamic patterns of neuroactivity are observed by its
1179Three Centuries of Category Errors in Studies

electrochemical manifestations (‘‘signs’’ according toAdrian, 1947), but neuroactivity cannot be observeddirectly, and models of it must include transforms thatdescribe postulated relations of the signs to the activity(Freeman, 1975). The contents of the signs are inferredfrom behavioral correlates and introspection (Freeman,1991, 1995), which are used to characterize and modelthe operations by which brains constructmeaningsfromsensory stimulation.This is not ‘‘information process-ing’’. The problem is, how to define neuroactivity? Justas ‘‘force’’ in physics is defined as a relation betweenmass, time and distance, ‘‘neural activity’’ must bedefined by relations between its electrochemical signsand overt, measured behaviors. Neuroactivity does notflow across the receptors, the muscles, or the blood–brain barrier as energy, matter and information do.Brains are closed systems with respect to meaning,though not to energy or information. This enclosureexplains the inaccessibility of qualia between brains,because the feelings and the associations in experiencesthat come in each brain with expectations, actions, andstimuli are rooted in its undivided body of past learningand its present chemical state.
The third insight is that the patterns of neural activityare endogenous. Their structure emerges from within andis not imposed solely by flows of energy, information orquanta from the bodies. The theory of chaos in nonlineardynamical systems offers a set of techniques for describ-ing the conditions required for emergence of self-organized patterns (Freeman, 1992, Skarda and Freeman,1987). The process of self-determination is inherent inthe Aquinian concept of intentionality (Martin, 1988,Pegis, 1948) by which each soul (mind/brain) intends(‘‘stretches forth’’) outside itself and into the world. Itacquires knowledge by shaping itself (learning) inaccordance with the effects (sensory feedback) of itsendogenous action. The soul creates itself and its virtueby its own actions. Descartes discarded this medievaldoctrine and mechanized the body by relegating per-ceptual events to the status of ‘‘representations’’ of theworld, so the soul understood through logic, not pre-logical learning. Kant deliberately revolutionized themechanization by postulating that the rules of knowingwere embedded as absolute ideas in human nature. Hehad no framework in Newtonian science to cope withthe emergent processes of intentionality. Brentano andHusserl reintroduced the word as denoting what theKantian representations were ‘‘about’’, whether or notthe objects or events so represented actually existed inthe world (Freeman, 1995). This meaning now consti-tutes the mainstream interpretation among analyticphilosophers (Putnam, 1990).
6. NONLINEAR BRAIN DYNAMICS ANDNEUROEXISTENTIALISM
The neo-Aristotelian philosophy of the later Middle Ages
brooked no doubts about the unique causal efficacy ofeach person. The machine metaphor undermined thatcertainty and induced conflicts between ideas of freewill and universal determinism that persist into themodern era. Several philosophers in the present centuryconstructed powerful theories that avoided the machinemetaphor and did not lead to the cruel dichotomy. Theearliest were John Dewey with American pragmatism,Henri Bergson with ‘‘Creative Evolution’’, MartinHeidegger with ‘‘Being and Time’’, and Jean Piagetwith developmental psychology. Sir Frederic Bartlett(1932) described the problem from the viewpoint of hisstudies on remembering:
...some widely held views [of memory] have to be com-pletely discarded, and none more completely than thatwhich treats recall as the re-excitement in some way offixed and changeless ‘‘traces’’ (p. vi).
The picture is one of human beings confronted by a worldin which they can live and be masters only as they learnto match its infinite diversity by increasing delicacy ofresponse, and as they discover ways of escape from thecomplete sway of immediate circumstances (p. 301).
There is one way in which an organism could learn how todo this. It may be the only way... An organism has somehowto acquire the capacity to turn round upon its own ‘‘sche-mata’’ and to construct them afresh. This is a crucial step inorganic development. It is where and why consciousnesscomes in; it is what gives consciousness its most prominentfunction... I wish I knew exactly how it was done (p. 206).
Jason Brown (1977) described it from a philosophicalviewpoint:
The structural organization of cognition is no less dynamicthan the psychological systems it supports... The incessantflow of cognition, the continual appearance and disappear-ance of new form at each moment of our waking and sleep-ing life, are manifestations of the activity of the structureas a whole as it achieves one or another level of realization(pp. 2–11).
Affect is not an energy that invades and charges an idea...There is no need for the concept of psychic energy (instinct,motivation) as a motivating force in cognition. The orderlysequence and unfolding of cognitive levels repeats andextends the phylogenetic and ontogenetic pattern. The pro-gression from depth to surface, the incessant repetition ofdevelopmental form, and the striving toward higher levelsare all part of an evolutionary trend that leads in a forwarddirection simply because it is in the nature of the organiza-tion to unfold in this manner (pp. 127–133).
Ilya Prigogine (1980) has applied his theory of ‘‘dis-sipative structures’’, which feed on energy and evolvecomplex patterns in states that are far from equilibrium,to understand the nonlinear dynamics of brains. HermannHaken (1983) has applied his theory of synergetics to
1180 W. J. Freeman

comprehend the physical principles, by which masses ofneurons can interact to generate spatiotemporal patternsof activity.
Only one of several theories was effectively linkedto neurobiology by its author. Maurice Merleau-Ponty (1942, 1945) drew heavily on work in clinicalneurology from the First World War, particularlydescribing the phenomena now known as phantomlimb and sensory neglect in brain-damaged indi-viduals. With careful reasoning over voluminous bio-logical details, he discarded the ‘‘materialist’’ viewthat minds were the results of linear causal chains ofreflexes and chemical reactions. He likewise dismissedthe ‘‘idealist’’ Kantian and Husserlian views of mindsconsisting of collections of representations, that wereprocessed according to logical algorithms. He pro-posed instead the ‘‘existentialist’’ view, taken fromhis teacher, Heidegger, and classmate, Sartre: mind is‘‘the structure of behavior’’, that creates itself bycircular causality in its own ‘‘action–perception cycle’’(Freeman, 1995).
7. THE NEUROBIOLOGY OF INTENTIONALITY
These several theories provide the warp and woof withwhich to weave the pattern of a strong neurobiologicaltheory of self-organization of mind and brain. Biologistsoffer observations of the space–time patterns from brainimaging of human and animal brains during the per-formance of intentional behavior. Psychologists offerthe measurements and analyses of behavior, in order toprovide the essential behavioral structures that are to becorrelated with the brain data. Physicists offer the dyna-mical systems theory by which to model the data andverify the capacity for brains to create and evolve theirown unique space–time patterns of neural activity.Philosophers offer the conceptual framework requiredto bring the large picture into focus. The questionremains: how do brains work? After three centuries ofdynamics, answers are still elusive.
Nonlinear dynamics gives the technical toolsneeded to learn how it is done. The concept of theself-organizing brain, with its ever-shifting basins andattractors, its evolving trajectories (Tsuda, 1991), and itsglobal cooperativity, enables us to model brain func-tions that transcend the present limitations of computa-tional and representational schemata, and enter intothose domains of nonrational and nonlogical construc-tion from which consciousness emerges.
The complementary foundation in the mental scienceshas been built in the past century by outstandingphilosophers and psychologists, who can be groupedunder the term ‘‘existentialists’’, and whose work hasremained outside the main stream of modern neuro-biology, owing to the unsolved problem of self-organization. The essential message of existentialism isthat humans—and animals—create themselves by their
actions. This insight has been arrived at independentlyin the 20th century by (among others):
Dewey (1914): Pragmatism—‘‘Actions are not reac-tions to stimuli; they are actions into the stimuli.’’Heidegger (1927) and his students, Sartre andMerleau-Ponty (1942): Existentialism—‘‘Mind isthe structure of behavior.’’Koffka (1935): Gestalt psychology—‘‘a field of forcebetween objects and our Egos...leads to action.’’Piaget (1930): The cycle of ‘‘action, assimilation, andadaptation’’ in the sensorimotor stage of childhooddevelopment.Gibson (1979): Ecopsychology—‘‘An affordance...ofan object offers what it does because it is what it is.’’
In each of these systems sensation takes place as partof an ‘‘action–perception cycle’’ that Merleau-Ponty(1942) described as ‘‘circular causality’’ to contrast itwith the ‘‘linear causality’’ of conditioned reflex chainsand machine metaphors of brain function, such as clocks,telegraph nets, thermodynamic engines, chemical reac-tion systems, computers and holographs. Animals andhumans receive and perceive stimuli as the end resultof goal-oriented search for knowledge in the environ-ment, and they learn about the world and shape them-selves accordingly entirely in terms of the consequencesof their own actions. The word ‘‘intentionality’’ hasthree widely accepted meanings. In analytic philosophyit means that a thought, belief, word, phrase or mentalact is ‘‘about’’ something, whether an object or a personor a state of affairs, whether in the world or in the mind.In the psychological sciences it means that a thought,action or speech has a purpose, goal or intent, which isboth outwardly directed toward manipulating objectsin the world and inwardly directed toward satisfyingbiological drives, needs or instincts. In medicine itrefers to the process of healing from injury, the re-estab-lishment of wholeness of the body (Freeman, 1995). Allthe meanings stem from Medieval philosophy, whichwas synthesized in the 13th century by Aquinas. Themind is conceived as having unity that serves to distin-guish itself from nonself; wholeness that expresses itsdirection of growth to maturity and the full realizationof its potential; and intent (‘‘stretching forth’’), by whichmind thrusts itself into the nonself by the actions ofits body, and learns about the world by shaping itselfin accordance with the outcomes of its actions, namelyby learning from the sensory stimuli that were soughtby its own actions (Freeman, 1995).
The neural mechanisms for intentionality in inverte-brate animals and humans clearly reside in the limbicsystem. The evidence for this conclusion comes fromdiverse areas of study of animal and human behavior.Comparative neuroanatomists have shown that theforebrain of the most primitive surviving vertebratesrepresentative of the ancestral line is composed ofthe essential sensory, motor and associational parts
1181Three Centuries of Category Errors in Studies

of the limbic system, including the primordial hippo-campus, septum and amygdala (Herrick, 1948). Selectivelesions brains have shown that removal of the entireneocortex but sparing the ancient limbic structuresimpairs sensory guidance and elaboration of behavior,but the impoverished actions are clearly identified asintentional (Broca, 1973, Goltz, 1874). Intentionalactions must take place within a space–time matrix forspatial orientation (the ‘‘cognitive map’’) and temporalintegration (‘‘short-term memory’’). Electrophysiologi-cal investigations of the hippocampus (O’Keefe andNadel, 1978) combined with studies of selective lesionsof the hippocampal formation (Milner, 1966) have shownthe importance of the limbic system for this matrix.An essential role in intentional action is played by‘‘corollary discharge’’ (Sperry, 1950) and reafference(Kay, 1994) first identified by von Helmholtz (1879),which clearly is focused in the entorhinal cortex throughits dense reciprocal connections both with the hippo-campus and with all primary sensory cortices (Lorentede No, 1934) and the frontal lobes (Freeman, 1995).
8. CONCLUSIONS
Electrophysiological studies of visual, auditory, somaticand olfactory EEGs (Barrie et al., 1996) have shown thatspatial patterns of neural activity emerge by constructionwith each act of perception, and that they depend on thecontext of the present and equally on the past experienceof each subject, not merely on the stimuli. This is areflection of the unity of function of the forebrain.
The implication is that each perceptual act has beenorganized in the context of the present state of the limbicsystem, expressing a desired state that is elaborated intoa plan of action and an expectancy of the sensory conse-quences of that action (Freeman, 1995). All past experi-ence is available and operating in the global dynamicalstate of the forebrain with each act in a continual succes-sion of acts. The objective aspect that is observable bybehaviorists is the flexible and adaptive sequence of actsfrom which intent is inferred. The subjective aspect isconsciousness within the individual of the context thatis brought to bear in each moment of choice with eachact.
By this interpretation there is little further to be saidabout the biology of consciousness, because animals can-not describe their states of awareness using language, andthe requisite electrophysiological studies in humans arenot admissible. Problems abound in the biology of inten-tionality, including the dynamics by which structureemerges from the chaos of neural activity, by whichlimbic activity patterns are re-shaped into commandsinto the motor systems and corollary discharges intothe sensory systems, and how the chain of nuclei in thebrain stem that provide the neurohormones bathingthe entire forebrain are integrated into the intentionalstructure of brain activity. These may not constitute
‘‘the hard problem’’ of Chalmers (1996), but as severalauthors have commented (Hameroff et al., 1996), theyare hard enough for this generation of researchers.
REFERENCES
Adrian, E. D. (1947).The physical background of perception. Oxford,UK: Clarendon Press.
Adrian, E. D., Bremer, F., & Jasper, H. H. (1954).Brain mechanismsand consciousness. Oxford, UK: Blackwell.
Alkon, D. (1992).Memory’s voice: Deciphering the mind–brain code.New York: HarperCollins.
Barlow, H. B. (1972). Single units and sensation: A neuron doctrine forperceptual psychology?Perception, 1, 371–394.
Barrie, J. M., Freeman, W. J., & Lenhart, M. (1996). Spatiotemporalanalysis of prepyriform, visual, auditory and somesthetic surfaceEEGs in trained rabbits.Journal of Neurophysiology, 76, 1–20.
Bartlett, F. C. (1932).Remembering. Cambridge, UK: CambridgeUniversity Press.
Broca, P. (1973).Memoires d’Anthropologie. Paris: Reinwald.Brown, J. (1977).Mind, brain and consciousness. New York: Academic
Press.Chalmers, D. J. (1996). Facing up to the problem of consciousness. In S.
R. Hameroff, A. W. Kaszniak, A. C. Scott (Eds.),Toward a scienceof consciousness(chap. 5, pp. 5–28). Cambridge, MA: MIT Press.
Churchland, P. S. (1986).Neurophilosophy: Toward a unified science ofthe mind–brain. Cambridge, MA: MIT Press.
Clarke, E., & O’Malley, C. D. (1968).The human brain and spinalcord: A historical study illustrated by writings from antiquity tothe twentieth century. Los Angeles: University of California Press.
Darwin, C. (1872).The expression of emotion in man and animals.London: Murray.
Dewey, J. (1914). Psychological doctrine in philosophical teaching.Journal of Philosophy, 11, 505–512.
Foster, M., & Sherrington, C. S. (1897).A textbook of physiology. III:The central nervous system(7th ed., p. 929). London: MacMillan.
Freeman, W. J. (1975).Mass action in the nervous system. New York:Academic Press.
Freeman, W. J. (1991). The physiology of perception.ScientificAmerican, 264, 78–85.
Freeman, W. J. (1992). Tutorial in neurobiology: From single neuronsto brain chaos.International Journal of Bifurcation and Chaos, 2,451–482.
Freeman, W. J. (1995).Societies of brains: A study in the neuroscienceof love and hate. Hillsdale, NJ: Lawrence Erlbaum.
Freud, S. (1895/1954). The project of a scientific psychology. InM. Bonaparte, A. Freud, E. Kris (Eds.), E. Mosbacher, J. Strachey(Trans.),The origins of psycho-analysis. New York: Basic Books.
Gibson, J. J. (1979).The ecological approach to visual perception.Boston, MA: Houghton Mifflin.
Goltz, F. (1874). U¨ ber die funktionen des Lendenmarks des Hundes.Pflugers Archiv gesamte Physiologie, 8, 460–498.
Haken, H. (1983).Synergetics: An introduction. Berlin: Springer.Hameroff, S. R., Kaszniak, A. W., & Scott, A. C. (Eds.), (1996).
Toward a science of consciousness. Cambridge, MA: MIT Press.Heidegger, M. (1927/1962).Being and time (J. Macquarrie,
E. Robinson, Trans.). New York: Harper.von Helmholtz, H. (1879/1925).Treatise on physiological optics: Vol. 3.
The perceptions of vision(J.P.C. Southall, Trans.). Rochester, NY:Optical Society of America.
Herbert, N. (1993).Elemental minds: Human consciousness and thenew physics. New York: Dutton, Penguin.
Herrick, C.J. (1948).The brain of the tiger salamander. Chicago, IL:University of Chicago Press.
Jackson, J. H. (1884/1958). Evolution and dissolution of the nervoussystem. Lecture III. In J. Taylor (Ed.),Selected writings. New York:Basic Books.
1182 W. J. Freeman

Kay, L. (1994). Distribution of gamma and beta oscillations in olfactoryand limbic structures during olfactory perception in rats: Evidencefor reafference.Proceedings of the World Conference on NeuralNetworks, WCNN’94(vol. 2, pp. 675–680).
Koffka, K. (1935).Principles of gestalt psychology(pp. 7, 353). NewYork: Harcourt Brace.
Kohler, W. (1940).Dynamics in psychology. New York: Grove Press.Lashley, K. S. (1929).Brain mechanisms of intelligence. Chicago:
University of Chicago Press.Lashley, K. S. (1942). The problem of cerebral organization in vision.
In J. Cattell (Ed.), Biological symposia VII (pp. 301–322).Lancaster, PA: Cattell Press.
Lashley, K. S. (1950). In search of the engram.Symposia. Society ofExperimental Biology, 4, 454–482.
Lorente de No´, R. (1934). Studies in the structure of the cerebral cortex:I. The area entorhinalis.Journal von Psychologie und Neurologie,45, 381–438.
Martin, C. (Ed.) (1988).The philosophy of Thomas Aquinas: Introduc-tory readings. New York: Routledge.
Merleau-Ponty, M. (1942/1963).The structure of behavior(A.L. Fischer, Trans.). Boston, MA: Beacon Press.
Merleau-Ponty, M. (1945/1962).Phenomenology of perception(C. Smith, Trans.). New York: Humanities Press.
Milner, B. (1966). Amnesia following operation on the temporal lobes.In C. W. M. Whitty, O. M. Zangwill (Eds.),Amnesia(pp. 109–133).London: Butterworths.
Milner, P. M. (1974). A model for visual shape recognition.Psycho-logical Review, 81, 521–535.
O’Keefe, J., & Nadel, L. (1978).The hippocampus as a cognitive map.Oxford, UK: Clarendon.
Pegis, A. C. (Ed.) (1948).Introduction to Saint Thomas Aquinas. NewYork: Modern Library.
Penrose, R. (1989).The emperor’s new mind. Oxford, UK: OxfordUniversity Press.
Piaget, J. (1930).The child’s conception of physical causality. NewYork: Harcourt Brace.
Prigogine, I. (1980).From being to becoming: Time and complexity inthe physical sciences. San Francisco: W.H. Freeman.
Prochaska, G. (1784).Adnotationum academicarum. Prague: W. Gerle.See also Unzer, J. A. (1851).The principles of physiology by GeorgeProchaska(T. Laycock, Trans.). London: Sydenham Society.
Putnam, H. (1990).Realism with a human face. Cambridge, MA:Harvard University Press.
Ryle, G. (1949).The concept of mind. New York: Barnes and Noble.Shaw, R. E., & Kinsella-Shaw, J. (1988). Ecological mechanics: A
physical geometry for intentional constraints.Human MovementScience, 7, 155–200.
Shaw, R. E., Kugler, P. N., & Kinsella-Shaw, J. M. (1990). Recipro-cities of intentional systems. In R. Warren, A. Wertheim (Eds.),Control of self-motion(chap. 22, pp. 579–619). Hillsdale, NJ:Lawrence Erlbaum.
Sherrington, C.S. (1940/1951).Man on his nature(2nd ed.). Cam-bridge, UK: Cambridge University Press.
Skarda, C.A., & Freeman, W.J. (1987). How brains make chaos in orderto make sense of the world.Behavioral and Brain Sciences, 10,161–195.
Spencer, H. (1863).Essays: Moral, political, and aesthetic. New York:Appleton.
Sperry, R. W. (1950). Neural basis of the spontaneous optokineticresponse.Journal of Comparative Physiology, 43, 482–489.
Sperry, R. W. (1958). Physiological plasticity and brain circuit theory.In H. F. Harlow, C. N. Woolsey (Eds.),Biological and biochemicalbases of behavior. Madison, WI: University of Wisconsin Press.
Tsuda, I. (1991). Chaotic itinerancy as a dynamical basis of hermeneu-tics in brain and mind.World Futures, 32, 167–184.
von der Malsburg, C. (1983). How are nervous structures organized? InE. Basar, H. Flohr, H. Haken, A. J. Mandell (Eds.),Synergetics ofthe brain(pp. 238–249). Berlin: Springer-Verlag.
1183Three Centuries of Category Errors in Studies

1997 SPECIAL ISSUE
A Neural Global Workspace Model for Conscious Attention
James Newman,1 Bernard J. Baars,2 and Sung-Bae Cho3
1Colorado Neurological Institute, Denver,2The Wright Institute, Berkeley and3Department of Computer Science, Yonsei University
(Received9 July 1996;accepted24 April 1997)
Abstract—Considerable progress is being made in interdisciplinary efforts to develop a general theory of the neuralcorrelates of consciousness. Developments of Baars’ Global Workspace theory over the past decade are examples of thisprogress. Integrating experimental data and models from cognitive psychology, AI and neuroscience, we present aneurocognitive model in which consciousness is defined as a global integration and dissemination system — nested in alarge-scale, distributed array of specialized bioprocessors — which controls the allocation of the processing resourcesof the central nervous system. It is posited that this global control is effected via cortical ‘gating’ of a strategic thalamicnucleus. The basic circuitry of this neural system is reasonably well understood, and can be modeled, to a firstapproximation, employing neural network principles.q 1997 Elsevier Science Ltd.
Keywords—Attention, Binding, Consciousness, Global Workspace theory, Nucleus reticularis thalami, Programmableblackboard, Thalamocortical circuit, Thalamus.
1. INTRODUCTION
Consciousness has been widely portrayed as an intract-able or irrelevant problem for cognitive science (e.g.Harnad, 1994; Penrose, 1994; O’Nuallain et al., 1997).Certainly its serious consideration is a fairly recentdevelopment (Jackendorf, 1987; Baars, 1988; Johnson-Laird, 1988; Edelman, 1989; Crick and Koch, 1990a),although a brief enthusiasm for the subject surfaced,and submerged, three decades ago (Eccles, 1966;Penfield, 1975). While it is not widely realized, theexperimental neuroscience which served as the basisfor that earlier enthusiasm is proving increasinglyrelevant to the present recrudescence of interest inconscious processes (see e.g. Stryker, 1989; Newman,1995a, 1997). Beginning with historical developmentsin both AI and neuroscience, this paper reviews agrowing body of evidence that some of the basicmechanisms underlying consciousness can be modeled,to a first approximation, employing variations uponcurrent neural network architectures (see also Taylor,1992; Taylor, 1996; Baars et al., in press; Newmanet al., 1997).
Baars (1983, 1988, 1992, 1994) has developed a setof ‘Global Workspace Models’, based upon a unifyingpattern, and addressing a substantial domain of evidence
explicitly related to conscious experience. These modelsexplicate an architecture in which many parallel,non-conscious experts interact via a serial, consciousand internally consistent Global Workspace (GW), orits functional equivalent. GW, or blackboard, architec-tures were first developed by cognitive scientists in the1970s and this framework is closely related to the UnifiedTheories of Cognition of Simon, Newell and Anderson(see Newell, 1992).
The HEARSAY model of speech understanding(Reddy et al., 1973) was one of the earliest attemptsto simulate a massively parallel/interactive computingarchitecture. The notion of a global workspace wasinitially inspired by this architecture, consisting of alarge number of knowledge modules, or ‘local experts’,all connected to a single ‘blackboard’, or problem-solving space. Activated experts could compete to post‘messages’ (or hypotheses) on the blackboard for allthe other experts to read. Incompatible messages wouldtend to inhibit each other, while the output of cooperat-ing experts would gain increasing access to the black-board until a global solution emerged. Blackboardarchitectures are relatively slow, cumbersome anderror-prone, but are capable of producing solutions toproblems too novel or complex to be solved by anyextant modular knowledge source. Once such ‘globalsolutions’ are attained, however, the original problemscan be allocated to modular processors for ‘non-conscious’ solution.
Neural Networks, Vol. 10, No. 7, pp. 1195–1206, 1997q 1997 Elsevier Science Ltd. All rights reserved
Printed in Great Britain0893–6080/97 $17.00+.00
PII: S0893-6080(97)00060-9
Pergamon
1195
Corresponding Author, Present address: 740 Clarkson Street, Denver,CO 80218, USA; Tel.: (303) 832-9062; e-mail: [email protected].

McClelland (1986) attested to the significance of thisset of models to subsequent developments in cognitivescience when he described HEARSAY, not only as ‘‘aprecursor of the interactive activation model’’, but ‘‘ofthe approach that underlies the whole field of paralleldistributed processing’’ (p. 122). We considerMcClelland’s own ‘Programmable Blackboard Modelof Reading’ as a connectionist example of a global work-space architecture, and discuss its applicability tomodeling conscious processes in a concluding section.
Another class of models that may turn out to be com-patible with GW theory comes from ‘distributed artificialintelligence’ (DAI), which Durfee (1993) characterizesas the study of ‘‘how intelligent agents coordinate theiractivities to collectively solve problems that are beyondtheir individual capabilities’’ (p. 84). He cites examplesof DAI applications, such as generic conflict resolution,unified negotiation protocols, and search-based modelsof coordination/cooperation. DAI applications appearto more closely approximate human interpersonalbehaviour than purely logic-driven AI. They requirethat agents learn to be ‘‘knowledgeable and skilled ininteracting with others’’ (p. 86). DAI models wouldappear to reflect an intelligent balance between compe-titive self-interest and cooperative problem-solving thatis essential to optimizing overall outcomes in complex‘social’ organizations. This, like GW theory, is consis-tent with other well-known metaphors in cognitivescience, such as Minsky’s ‘Society Theory’ (Minsky,1979) and Gazzaniga’s ‘Social Brain’ (Gazzanigga,1985).
A similar, globally-integrative balancing of prioritiesappears to characterize the optimal processing of con-scious information. Conscious percepts are characterizedby unified gestalts of shape, texture, color, location andmovement, despite the fact that these contributions toperception are initially processed in parallel areas ofthe cortex, in both hemispheres. Moreover, consciousintentions are generally single-minded and goal-directed.Of course, conflicts can and do arise, but a centralpurpose of consciousness seems to be resolving suchconflicts (employing both integrative and inhibitoryalgorithms).
While such global states can be highly adaptive —indeed, are essential to explicit learning — GW theorymaintains that the vast majority of cognitive tasks per-formed by the human brain are automatic, and largelynon-conscious (Baars, 1988; Newman and Baars, 1993;Baars, 1997; Newman, 1997). Consciousness generallycomes in play when stimuli are assessed to be novel,threatening, or momentarily relevant to active schemasor intentions.
The defining properties of stimuli which engage consciousattention (i.e. the global allocation of processing resources)are that they: 1) vary in some significant degree fromcurrent expectations; or 2) are congruent with the current,predominant intent/goal of the organism. In contrast, the
processing of stimuli which are predictable, routine orover-learned is automatically allocated to non-conscious,highly modularized cognitive systems (Newman, 1995b,p. 691).
Generally, we are conscious of what has the highestrelevance to us at that moment. This may be a momen-tary threat, a sudden insight, a pleasant sensation, etc. (inrelaxed moments, there may be no particular focus orintent, simply a stream of associations). Yet, while therange of our awareness is immense (limited only by ourmost developed cognitive capacities), we contend thatthe basic mechanism for the allocation of these capacitiesremains constant under virtually all contingencies; andthe basic neural circuitry of that resource-allocationmechanism is reasonably well understood. Indeed, insubsequent sections, we suggest how it might be modeledbased upon already existing neural network simulations(McClelland, 1985; Hampshire and Waibel, 1992;Taylor and Alavi, 1993; Llinas et al., 1994).
The relevance of Global Workspace theory extendsbeyond NN modeling, however. Indeed, it bears uponcentral philosophical problems in consciousness studies,such as thehomunculusand Cartesian theater. The twoare, of course, related. The image of a ‘little man in ourhead’ observing and manipulating the play of consciousimages is beguiling, but absurd. For who is this strangebeing lodged in our brains? And who is watchinghim?
In Global Workspace theory the single homunculus isreplaced by a large ’audience of experts’. The ‘theater ofconsciousness’ then becomes a workspace, with stage(Baars, 1997). Almost everyone in an audience haspotential access to center stage (although most prefer tosimply observe, or exert indirect influences). The focusof conscious activity, at any moment, corresponds to the‘work’ produced by the most active coalition of experts,or modular processors: whoever has managed to win thecompetition for ‘the spotlight’. There is no fixed, super-ordinate observer. Individual modules can pay as muchor as little attention as suits them, based upon their par-ticular expertise. At any one moment, some may bedozing in their seats, others busy on stage. In thissense, the global workspace resembles more a delibera-tive body than a theater audience. Each expert has acertain degree of ‘influence’, and by forming coalitionswith other experts can contribute to deciding whichissues receive immediate attention and which are ‘sentback to committee’. Most of the work of this deliberativebody is done ‘off stage’ (i.e. non-consciously). Only mat-ters of greatest relevance in-the-moment gain access toconsciousness.
While the GW is a teaming multiplicity, what isexplicitly represented in consciousness is largely coher-ent and adaptive. The overall workspace serves as a‘global integration and dissemination system’, in whichall experts can participate, but only select coalitionsdominate, momentarily, producing an orderly succession
1196 J. Newman et al.

of global representations. The stream of consciousnessarises out of the operations of the GW system — and,over time, our sense of being a coherent ‘I’ (the memoryand intention systems vital to this aspect are beyond thescope of this paper; see Baars et al., in press; Newman,1997). It is this unitary awareness, not any agent orhomunculus, that is globally superordinate. Of course,such a system is prone to inefficiencies and pathologicalperturbations, but this is consistent with the scientificliterature concerning human consciousness (see Baars,1988).
If we are to proceed beyond pleasing metaphors, how-ever, it is necessary to operationalize the GW model inexplicit neurocognitive terms. This process begins in thenext section. To introduce it, we offer the followingworking definition:
consciousness reflects the operations of a global inte-gration and dissemination system, nested in a large-scale, distributed array of specialized bioprocessors;among the various functions of this system are theallocation of processing resources based, first, uponbiological contingencies of novelty, need or potentialthreat and, secondly, cognitive schemas, purposes andplans.
2. MODELING GLOBAL, COMPETITIVEATTENTION
We have introduced the theoretical background for themodel. Newman and Baars (1993) and Newman (1997)present detailed accounts of its neural architecture. Wewould stress, however, that consciousness is a dynamicprocess, not a static structure. Also, it is not localized tosome ‘brain center’, but arises out of the coordinatedactivities of widely distributed networks of neurons.Resource allocation is integral to these activities. Theneural bases of resource allocation, or attention, havebeen extensively explored (see, e.g. Heilman et al.,1985; Mesulam, 1985; Posner and Rothbart, 1991;Posner, 1994; LaBerge, 1990, 1995). But, of course,not all forms of attention are conscious. As an examplefrom AI, McClelland (1986) notes that in simulationsof reading, activated modules must be ‘sticky’, that is‘‘interactive activation processes continue in olderparts of the programmable blackboard while they arebeing set up in newer parts as the eye moves along...’’(pp. 150–151). This ‘stickiness’ would seem to entail atype of attention. It normally proceeds quite auto-matically, however, both in a reading machine and in aliterate human being. Only when the process is disruptedby, say, a mis-spelled or unknown word, does that wordbecomes the focus of our conscious awareness.Normally, we are only conscious of the overall senseof the passage of text, and the images and thoughts itevokes, not particular semantic or syntactical operations.These linguistic processes became second nature to us
long ago. Such ‘particular operations’ are hardly trivialaspects of language acquisition, but as Kihlstrom (1987)noted, in humans they tend to be ‘‘automatized throughexperience and thus rendered unconscious’’ (p. 285).
Conscious awareness clearly involves a higher orderof resource allocation, which Newman and Baars (1993)call ‘global attention’. The term ‘‘refers to a level ofcognitive processing at which a single, coherent streamof information emerges out of the diverse activities of theCNS’’ (p. 258). The focus of that stream could (underatypical circumstances) be an individual word; but theconscious mind seldom confines itself to the processingof such rudimentary representations. Rather it seems to bedecisively biased towards multifaceted, yet unifiedimages. Thus, we are able to perceive a Necker Cubeas projecting out of a two-dimensional page, alternatelyto the left, then to the right; but we are curiously incapableof perceiving these two perspectives simultaneously.
The processing load of global attention (like workingmemory), is both highly chunked and highly restricted(Baars, 1988). The non-conscious allocation of proces-sing resources operates under no such constraints. Forexample, neuroscience has shown that specialized areasin the visual cortex process, in parallel, the contour,movement, color, spatial location, etc. of a stimulus(LaBerge, 1995). Yet our awareness is of a single, coher-ent object (and often includes tactile, auditory and asso-ciative aspects). Thus, neuroscience is faced with the’binding problem’ of how these multifarious representa-tions, generated by widely separated areas, are integratedinto real-time ‘objects’ of perception (see Crick andKoch, 1990a; Newman and Baars, 1993).
One would expect the neural mechanism for globalattention to be complex, and widely distributed, whichit is. But the basic circuitry can be described, to a firstapproximation, in terms of repeating, parallel loops ofthalamo–cortico–thalamic axons, passing through athin sheet of neurons known as thenucleus reticularisthalami (nRt). The loops are formed by long-axoned,excitatory neurons. The neurons of nRt are largelyGABA-ergic, inhibitory neurons. Most, if not all, of thelooping axons give off collaterals as they pass throughnRt, while nRt neurons themselves project mainly tocells of the particular thalamic nucleus lying directlybeneath them. There is an orderly topography to thisarray of axon collaterals and underlying thalamic nuclei(Scheibel and Scheibel, 1966; Mitrofanis and Guillery,1993). It essentially mirrors, in miniature, the modulararchitecture of the cortex (see Newman and Baars, 1993;LaBerge, 1995; Newman, 1997, for reviews).
Evidence for the central role of this ‘thalamocorticalcircuit’ (LaBerge, 1995) in attention and conscious-ness has been accumulating for decades (Jasper, 1960;Scheibel, 1980; Jones, 1985; Steriade and Llinas, 1988;Llinas and Pare, 1991). Skinner and Yingling (1977) firstproposed a neural model for its role in selective attention.Our ‘wagon wheel’ model (next section) represents a
1197Neural Global Workspace Model

synthesis of both the accumulated evidence, and relatedmodels (Skinner and Yingling, 1977; Scheibel, 1980;Crick, 1984; Taylor and Alavi, 1993; Llinas et al.,1994; LaBerge, 1995). These related models vary intheir details, as the precise connectivities andphysiology of the thalamocortical circuit are not fullyworked out.
Most attentional models are based upon conventionalsimulations of mechanisms such as center-surround inhi-bition, or winner-take-all (WTA) competitions, amonglocal circuits. Various researchers have described thenetwork of nRt neurons as a mosaic, or array, of neural‘gatelets’ acting to selectively filter the flow of sensoryinputs to the cortex (Skinner and Yingling, 1977;Scheibel, 1980; Crick, 1984). The WTA dynamic mayseem analogous to the ‘competition’ posited by GWtheory. The problem with such conventional networksis that they are poorly suited to global forms of competi-tion, because prohibitively long-range and geometricallyincreasing numbers of connections would be required.Moreover, most long-range, reciprocal connections inthe CNS are excitatory. Inhibitory effects tend to belocal.
Taylor and Alavi (1993), however, have modeled a
competitive network for global attention based upona highly simplified version of the ‘thalamus–NRT–cortex complex’. Their model is unique, in that it takesinto account the effects of dendro–dendritic interactionsthroughout nRt. The dendrites of nRt cells project outtangentially within the reticular sheet, bidirectionally.The physiology of information processing in dendritictrees is highly complex, and not well understood (Mel,1994); but Koch and Poggio (1992) review evidence forthe dendritic trees playing a role in several types ofsecond-order, multiplicative computations. We willhave more to say about this subsequently.
Figure 1 [taken from Taylor and Alavi (1993)] illus-trates three thalamocortical circuits, as well as the non-linear, dendro–dendritic connections between N1, N2,N3,... within NRT. We would refer the reader to theoriginal paper for a detailed description of the simula-tions carried out, employing a network of 100 thalamo-cortical loops. To briefly summarize the results, theaddition of dendro–dendritic connections to the loopingcircuits provided ‘‘the basis for a simple version of theglobal gating model... that instantiates a form of compe-tition in the spatial wavelength parameters of incominginputs...’’ (p. 352). In this version of the model, the entirenRt network oscillates with a wavelength,
with the net strength given by the component of the inputwith the same wavelength.
The way in which global control arises now becomesclear. Only those inputs which have special spatial wave-length oscillations are allowed through to the cortex, orare allowed to persist in those regions of the cortexstrongly connected to the NRT: the thalamus–NRT sys-tem acts as a spatial Fourier filter (p. 353).
Simulation runs demonstrated the global, wave-likeproperties of the competitive model. The overall patternof activation in cortical units was shown to be exclu-sively dependent upon the wave pattern spanning acrossall of the NRT units (Figure 2). As LaBerge (1995) notes,the actual physiology of nRt gating in alert states remainsunclear, but it is firmly established that nRt is the sourceof global oscillatory activity (at 8–13 Hz) initiating thedescent into sleep.
The RN cells are known to inhibit each other, and wheninhibition hyperpolarizes an RN cell sufficiently, it producesa rebound burst. In this way a network of connected RNinhibitory cells can spread activity to every cell within thenetwork, apparently without decrement in the intensity ofthe activity (p. 184).
Here then, is a plausible circuitry for a global, winner-take-all competition among the large array of specializedcortical processing areas.
Llinas et al. (1994) offer an interesting variation uponthis circuitry, in which thalamocortical loops of the ‘non-specific’ intralaminar nuclei operate in parallel with thespecific (input) loops described above. The synchronous
1198 J. Newman et al.
FIGURE 1. The wiring diagram of the main model of the thala-mus–NRT–cortex complex. Input I j is sent both to the thalamicrelay cell T j and the inhibitory interneuron INj, which latter cellalso feeds to Tj. Output from Tj goes up to the correspondingcortical cell C j, which returns its output to T j. Both the axons T jCj
and CjT j send axon collaterals to the corresponding NRT cell Nj.There is axonal output from Nj to INj, as well as collaterals toneighbouring NRT cells. There are also dendro–dendriticsynapses between the NRT cells (from Taylor and Alavi, 1993).

activation of specific and non-specific loops is postulatedto provide a basis for ‘‘perceptual unity... by whichdifferent sensory components are gathered into oneglobal image’’ (p. 251). Their modeling is concernedwith high-frequency EEG oscillations (and omitsdendro–dendritic connections), yet appears to parallelmuch of what we discuss above.
When the interconnectivity of these nuclei is combined withthe intrinsic properties of the individual neurons, a networkfor resonant neuronal oscillations emerges in which specificcorticothalamic circuits would tend to resonate at 40 Hz.According to this hypothesis, neurons at the differentlevels, and particularly those in the reticular nucleus,would be responsible for the synchronization of 40-Hzoscillations in distant thalamic and cortical sites...theseoscillations may be organized globally over the CNS, espe-cially as it has been shown that neighboring reticular cellsare linked by dendrito–dendritic and intranuclear axoncollaterals (Deschenes et al., 1985; Yen et al., 1985,pp. 253–254).
3. A NEURAL MODEL FOR GLOBAL RESOURCEALLOCATION
We have introduced a set of convergent models for thebasic circuitry of a Global Workspace system involved inthe integration and dissemination of the processingresources of the nervous system. This ‘bare bones’version accounts for how a global, winner-take-all com-petition might be mediated between various externalinputs and cortical modules, to produce ‘‘a single, coher-ent stream of information out of the diverse activities of
the CNS’’ (Newman and Baars, 1993). There remains tobe explained how the thalamocortical circuit fits in withthe second half of our working definition for the con-scious system: the allocation of processing resources
1199Neural Global Workspace Model
FIGURE 2. One of 15 simulation runs for the thalamus–NRT–cortex model showing full global control with semi-constant spatial input.Note that cortex activity is influenced by the NRT alone (from Taylor and Alavi, 1993).
FIGURE 3. ‘Wagon wheel’ model of CNS systems contributing toglobal attention and conscious perception. A1, primary auditoryarea; BG, basal ganglia; g c, ‘closed’ nRt gate; g o, ‘open’ nRt gate;MRF, midbrain reticular formation; nRt, nucleus reticularis tha-lami; PFC, prefrontal cortex; S1, primary somatosensory area;Th, ventral thalamus; V1, primary visual cortex (from Newmanet al., 1997).

based, first, upon biological contingencies of novelty,need or potential threat and, secondly, cognitive sche-mas, purposes and plans. In keeping with our definition,we will first add a sub-cortical component for orienting to‘‘novelty, need, or potential threat’’, and then discuss themuch more complex aspects of cortically-mediatedeffects upon the system.
This extended version of the model is schematicallyillustrated in Figure 3 as a ‘wagon wheel’, with thethalamus (Th) as its ‘hub’. The reticular nucleus (nRt)corresponds to the metal sleeve fitted around the hub.The upper rim of the wheel represents the cerebral cortex(PFC/S1...V1), and closely associated basal ganglia(BG). The lower half shows the major sensory systemsand subcortical nuclei whose projections converge uponthe thalamus. The outer ‘spokes’ represent the sensorypathways for vision, audition and the bodily senses.These project, in an orderly topography, to modality-specific nuclei in the thalamic ‘hub’. As they ascendtowards the thalamus, these pathways give off collateralsto the midbrain reticular formation (MRF) (see alsoNewman et al., 1997). Scheibel (1980) reviewed threedecades of experimental evidence indicating that thesemidbrain collaterals serve as the basis for an initial‘spatial envelope’, or global map, of the environmentsurrounding the animal.
Most reticular [MRF] neurons...appear multimodal,responding to particular visual, somatic and auditorystimuli, with combinations of the last two stimuli mostnumerous. The common receptive fields of typical bimodalcells in this array show a significant degree of congruence.For instance a unit responding to stimulation of the hindlimb will usually prove maximally sensitive to auditorystimuli originating to the rear of the organism. These twinsomatic and auditory maps retain approximate register andoverlap the visuotopic map laid down in the...superiorcolliculus...These data might be interpreted to mean thateach locus maps a point in the three-dimensional spatialenvelope surrounding the organism. Further studies suggestthe presence of a deep motor map closely matching and inapparent register with the sensory map. (p. 63)
More recent research has supported Scheibel’s por-trayal of the superior colliculus as the visual componentof what Crick and Koch (1990b) termed a ‘saliency map’for eye movements, involved in orienting the animal tobiologically relevant stimuli. Subsequent findings haveboth confirmed Scheibel’s analysis, and revealed anumber of ‘top-down’ projections that modulate activ-ities in MRF. LaBerge (1995) writes:
the superficial area [of the superior colliculus] receivesstrong cortical inputs from V1, V2 and V3 [primary andsecondary visual cortex], the deep layers in the monkeySC receive their main cortical inputs from the posteriorparietal area (Lynchet al., 1985), from the prefrontalareas (Goldman and Nauta, 1976), and the frontal eye fields(Leichnetzet al., 1981). The deep layers contain a map ofvisual space that is stacked adjacent to maps for auditory
and somatosensory spaces in a manner that cells correspond-ing to points in space lie along the same vertical axis(Merideth and Stein, 1990). Stimulation of these cells bymicroelectrodes produces movements of eyes and head...(LaBerge, 1995, p. 145)
LaBerge goes on to describe basal ganglia inputs that‘‘are of particular importance because they tonicallyinhibit activity in the SC cells’’. It has long beenknown that the frontal eye fields, and posterior parietalarea ‘‘exert strong influences on eye movements andmust be considered together with the superior colliculusin accounting for...orienting of attention’’ (p. 142). Thesefacts emphasize two key aspects of the ‘conscioussystem’ we are modeling: 1) it is polymodal, integratingnot just visual, auditory and somatosensory inputs, butmotor and ‘higher-order’ cortical effects; and 2) it isextended, with input/output relations reaching from thebrain stem core to association cortices. Indeed, the gen-eral term we have used to describe it elsewhere is the’extended reticular–thalamic activation system’, or’ERTAS’ (Baars, 1988; Newman and Baars, 1993;Newman, 1995a, 1995b, 1997).
The third key aspect of the system (as exemplified bythe ’wagon wheel’ model) is that it converges on thethalamus. We have already discussed this in terms ofthe thalamocortical circuit, which connects to ‘‘virtuallyevery area of the cerebral cortex’’ (LaBerge, 1995,p. 221). Scheibel (1980) described the MRF portion ofthe system as:
sweep[ing] forward on a broad front, investing the [intra-laminar complex of the] thalamus and nucleus reticularisthalami. The investiture is precise in the sense that thesites representing specific zones of the spatial envelope(receptive field) project to portions of the nucleus reticularisconcerned with similar peripheral fields via projections fromboth sensory thalamus and sensory association cortices(p. 62).
The fact that Scheibel’s (1980) ‘spatial envelope’projects with some topographic precision upon nRt,would appear to enable it to disinhibit particular arraysof nRt gatelets, selectively enhancing the flow of sensoryinformation to the cortex. The ’intralaminar complex’(Newman and Baars, 1993) is also integral to theERTAS system, as the non-specific portion of the thala-mocortical circuit. It is intralaminar projections whichrelay MRF activation to the cortex (illustrated by thevertical MRF-Th projection, above which it branchesout to all areas of CORTEX). As noted above, Llinaset al. (1994) hypothesize the perceptual unity of con-sciousness (binding) to be brought about by the globalsynchronization of specific and non-specific circuits vianRt. Scheibel (1980) earlier concluded as much concern-ing the role of this extended activation system in ‘selec-tive awareness’:
From these data, the concept emerges of a reticularis
1200 J. Newman et al.

complex [nRt] selectively gating interaction between speci-fic thalamic nuclei and the cerebral cortex under the opposedbut complementary control of the brain stem reticular core[MRF] and the frontal granular cortex [PFC]. In addition,the gate is highly selective; thus, depending on the nature ofthe alerting stimulus or central excitation, only that portionof the nucleus reticularis will open which controls the appro-priate subjacent thalamic sensory field. The reticularis gate[thus] becomes a mosaic of gatelets, each tied to somespecific receptive zone or species of input. Each is underthe delicate yet opposed control of: (a) the specificallysignatured sensory input and its integrated feedback fromcortex [S1...V1]; (b) the reticular core [MRF] with its con-cern more for novelty (danger?) than for specific detailsof experience; and (c) the frontal granular cortex-medialthalamic system [PFC/BG] more attuned to upper levelstrategies of the organism, whether based on drive mechan-isms (food, sex) or on still more complex derivative phe-nomenon (curiosity, altruism). Perhaps here resides thestructuro- functional substrate for selective awareness, andin the delicacy and complexity of its connections, our sourceof knowing, and of knowing that we know (p. 63).
Here, as well, is a summary description of a neuralsubstrate for the global allocation of the processingresources of the CNS. All that it lacks is the mechanismsfor a global competition (Taylor and Alavi, 1993) andbinding (Llinas et al., 1994) introduced in the previoussection. But we must tie the operations of this thalamus-centered system more closely to those of the cortex andbasal ganglia, or most of the functions routinely studiedby cognitive science have no place in the model. Thisintroduces an exponentially higher level of complexity(one of the hazards of dealing with global systems).
One of the values of GW theory, however, is that itprovides a framework for understanding this complexity.First, it holds that the vast majority of cognitive functionsare carried out, non-consciously, via changing arrays ofspecialized, modular processors. This is reflected, anato-mically, in the immense number of cortico–cortical con-nections in the human brain, outnumbering those withsubcortical nuclei by nearly ten to one. Thalamocorticalprojections are comparatively sparse, but serve at leasttwo essential functions: 1) transmitting sensory inputs tothe primary cortical areas (S1, A1, V1, Figure 3); and 2)providing a means to selectively amplify/synchronizecortex-wide activation (Section 2).
GW theory also reminds us that conscious functionsoperate upon an information load about the size of work-ing memory. Thus, we are talking of a highly coarse-grained level of processing. In this context, globalattention is (at least) a second-order operation, actingupon a highly selective stream of information. All thisis to say that a relatively low density of widely distrib-uted, yet highly convergent, circuits could be all that arerequired to create a conscious system; and these are thevery characteristics of the neural model we havedescribed.
However, most neural network modelers take a
cortically-centered view of cognition, from which thebrain stem functions so far described probably seemrather primitive or trivial (i.e. orienting, controlling eyemovements) when compared to cortically-mediated pro-cesses such as language acquisition, pattern recognition,motor planning, etc. What evidence is there that cortical(and other forebrain systems) depend upon projectionsto the thalamus for effecting high-level cognitiveprocesses?
Early support for such effects, mediated by prefrontalprojections, was provided by animal experiments under-taken by Skinner and Yingling (1977). They found thatselective activation of one portion of a fronto-thalamictract could shut down sensory processing in visual,but not auditory, cortex. Activation of another ‘spoke’of the prefrontal-thalamic tract shut down auditory pro-cessing, but allowed visual inputs to reach posteriorcortex. Skinner and Yingling wrote ‘‘This result impliesthat selective attention emerges via selective inhibitionin certain sensory channels that the animal mustknow inadvanceare irrelevant to its situation’’ (p. 54). To inhibitorienting based upon advanced knowledge is clearly asophisticated use of cognition. Several lines of researchhave converged in recent years to support this concept.Summarizing the current state of knowledge of prefrontalregulation of subcortical systems, Newman (1997)wrote:
It is now generally accepted that the prefrontal lobes (withthe cingulate cortex) constitute an ‘executive’ over thelimbic system mediating such functions as working mem-ory, inhibition of conditioned responses, and goal-directedattention (see Fuster, 1980; Goldman-Rakic, 1988b;Damasio, 1994; Posner, 1994). More recent research onthe basal ganglia (see reviews by Groenewegen andBerendse, 1994; Parent and Hazrati, 1995) have suggestedthat they constitute a ‘motor programming extension’ ofthe frontal lobes as well — routed through the thalamus(p. 112–113).
Newman (1997) goes on to cite evidence (Parent andHazrati, 1995) that the BG ‘extension’ (like the thalamo-cortical loops) sends rich, collateral projections to nRtthat effect not only its ‘gating’ of motor programs, buthippocampal-mediated episodic memory functions (seealso Newman, 1995b).
Finally, we would note that cortico-thalamic projec-tions to nRt and associated specific nuclei are both moretopographically precise (Mitrofanis and Guillery, 1993)and more pervasive than had once been thought (Jones,1985). Llinas and Pare (1991) estimate that, for everyaxon the thalamus sends to the cortex, the cortical areait projects to reciprocates with ten. Given the modulararchitecture of the neocortex, one might reasonablypredict that these cortico-thalamic projections exerthighly differentiated influences upon the flow of infor-mation through the thalamus. Efforts by experimentalneuroscience throughout the 1980s to elucidate theprecise effects of cortico-thalamic projections were
1201Neural Global Workspace Model

frustratingly inconclusive. But a recent review by Buserand Rougeul-Buser (1995) notes:
The situation has however recently swung back, due to somenew and perhaps consistent findings, indicating that thevisual cortex appears to have a major action down ontothe lateral geniculate nucleus, which may generatethalamic oscillations (Funke and Eyse, 1992; McCormickand Krosigk, 1992; Krosigk et al., 1993; Sillito et al., 1994)(p. 252).
While additional research is clearly needed, theserecent findings suggest that Scheibel’s (1980) earlymodel of the converging influences of projections upona thalamic hub — with the addition of basal gangliainputs to nRt and the intralaminar complex — remainsa viable model for ‘‘global attention’’, including theinfluences of cortically generated ‘‘schemas, purposesand plans’’. Newman (1997) discusses the contributionsof the ‘cortico-basal ganglia–thalamo–cortical loop’ tomemory and volitional processes in greater detail. Thecomplexities of this system are beyond the scope of themodels presented here, although Monchi and Taylor(1995) and Taylor and Michalis (1995), among others,have developed neural models simulating functions ofthe BG and hippocampal systems.
What we propose to do instead is present a muchsimpler, but highly relevant, connectionist model thatsimulates the sorts of second-order operations onewould predict in a GW system employing a gating net-work to selectively filter and integrate inputs as a func-tion of central knowledge stores. The basic heuristic forthis type of model is described in Newman and Baars(1993). It posits that
prefrontal cortex acts as an executive attentional system byactively influencing information processing in the posteriorcortex through its effects upon the nucleus reticularis. In thismanner, the highly parallel [processing] functions of theposterior cortex are brought into accord with increasinglycomplex and intentional cognitive schemes generatedwithin the prefrontal regions of the brain (p. 281).
A defining property of an executive system is that itacts upon other sub-systems, modifying their inputs forits particular purposes. Posterior cortical areas act morelike arrays of quasi-autonomous processing modules (orlocal experts) — the bread and butter of NN simulations.Note that an executive system is not anessentialrequire-ment for consciousness. That this is the case is illustratedby the literature on extensive damage to the frontallobes of the brain. PFC damage results in significantdeficits in such purposeful activities as: the inhibitionof inappropriate responding; switching of response set,planning and monitoring of actions, etc.; but produceslittle or no alteration in basic mental status. Indeed,many patients with frontal lobe pathology perform atpre-morbid levels on intelligence tests (Walshe, 1978;Damasio, 1994). In terms of the GW model we have
presented, it is not executive attentional processes, butthe selective binding of coalitions of active corticalmodules via a thalamocortical competition which is thesine qua nonfor the generation of a coherent stream ofconscious representations. Examples of these aspectsof the GW model have already been offered.
4. SECOND-ORDER MODELS FORGLOBAL GATING
Let us return to the ‘wagon wheel’ model illustrated inFigure 3, and transform its components into a connec-tionist GW, with an executive system. To simplifythings, the network will have only two sensory modules,one for processing auditory (A1) inputs, and one forvisual (V1). In order to provide second-order controlover processing in both modules, we will add a gatingmodule (nRt) with the same number of units as connec-tions in each sensory module. Each gating unit sends itsoutput to a corresponding connection in A1 and V1. Theconnections between the gating units and sensory unitsare multiplicative. As Rummelhart et al. (1986) writeabout such connections:
if one unit of a multiplicative pair is zero, the other memberof the pair can have no effect, no matter how strong itsoutput. On the other hand, if one unit of a pair has value1, the output of the other passe[s] unchanged to the receivingunit....In addition to their use as gates [such] units can beused to convert the output level of a unit into a signal thatacts like aweight connecting two units (p. 73).
In this manner, a fully connected gating module canactually program the connection strengths of one or moreinput modules to process a particular type of input,for example phonemes, or letters, into words. For maxi-mum flexibility, it is preferable that the gating modulenot have fixed connections either, but simply relaygating (connection strength) information from a centralmodule to which its units are connected. The centralmodule contains (in this case) word-level knowledgeneeded to program the sensory modules to processwords. Another central module might be specializedfor knowledge for processing visual scenes or tactileshapes. To complete the system, each programmableinput unit sends a corresponding connection to a centralmodule unit.
The highly simplified network just described is reallya variation on a ‘Programmable Blackboard Modelfor Reading’ developed by McClelland (1985, 1986).Its four modules correspond to those labeled inFigure 4: a Central Module (PFC); Connection Activa-tion System (PFC-nRt); and two Programmable Modules(A1, V1). The connections described above are shownin Figure 5 (note: McClelland’s modules are identical,and used only for reading (not hearing) words, but theo-retically they could be programmed to processany typeof input).
1202 J. Newman et al.

In the brain, of course, the primary areas (A1, V1, S1)send no direct projections to PFC; but they do sendconvergent projections (as in Figure 5) to secondaryassociation areas, which send projections directly toPFC (as well as posterior association areas). Althoughthese feed-forward projections to PFC are less topo-graphically precise (e.g. the receptive fields of visualneurons in the secondary areas are much larger), theymaintain a fair degree of parallel distribution, indicatingthat much of the prefrontal cortex is as modular in itsorganization as the posterior ‘association’ cortex. More-over, PFC ‘modules’ reciprocate these parallel, feed-forward projections, although in a more divergent pattern(Goldman-Rakic, 1988a; LaBerge, 1995). Interestingly,this convergence/divergence pattern is paralleled bythe connections in Figure 5 for the central module.
In the actual prefrontal cortex there are hundreds (ifnot thousands) of ‘central modules’. Feed-forward inputsallow them to use and store highly processed infor-mation from the posterior (sensory) cortex. Of course,feedback (or re-entrant) connections enable PFC to influ-ence processing in the posterior areas as well. But suchdivergent and indirect feedback pathways are poorlysuited to exercising momentary, direct effects upon pro-cessing at the input level. Nor could such centrally-storedknowledge be employed to guide, or anticipate, how
inputs are processed (re the ‘knowing in advance’Skinner and Yingling (1977) attributed to PFC-Thcircuits). This is where direct projections to the primaryprocessing areas (actually the thalamocortical circuit)could prove quite valuable. Instead of the sensory inputunits (A1-Th; V1-Th) responding based upon fixedconnection strengths, a central module could programinput modules to process (i.e. pay attention to) particularcategories of inputs. McClelland (1986) calls this formof activation ‘connection information distribution’ (CID)and compares its benefits to those of:
the invention of the stored program....The use of centrallystored connection information to program local processingstructures is analogous. It allows the very same processingstructures to be programmed to perform a very wide range ofdifferent tasks.... [CID] also carries out a form of what isknown in production systems as ‘resolution’, binding theright tokens in the blackboard together into higher-orderstructural patterns (p. 165).
Finally, he notes analogous aspects in the CID’s opera-tions to ‘working memory’, a process which has been tiedby neuroscientists to a prefrontal/thalamic/hippocampalsystem (e.g. Fuster, 1980; Goldman-Rakic, 1988b).These comparisons between the Wagon Wheel and Pro-grammable Blackboard models, of course, have purelyheuristic value (although McClelland’s (1986) PABLOsimulation of his model contained a sufficient program-mable blackboard to read lines of text up to 20 characterslong). But the use of gating networks to generate useful‘higher-order structural patterns’ is fairly widespread.
For engineering problems such as object recognitionand robot motion control, the concept of combiningmodular networks using gating connections has beenactively exploited to develop highly reliable systems(Jacobs et al., 1991; Hampshire and Waibel, 1992;
1203Neural Global Workspace Model
FIGURE 4. A simplified example of a Connection InformationDistributor (CID), sufficient for simultaneous bottom-up process-ing of two two-letter words. The programmable modules consistof the programmable letter (PL) and programmable word (PW)nodes, and programmable connections between them (opentriangles). The central module consists of a set of central letter(CL) nodes and a set of central word (CW) nodes, and hard-wiredconnections between them (filled triangles). The connectionactivation system includes the central word nodes, a set of con-nection activator (CA) nodes, and hard-wired connectionsbetween them. Connections between the central knowledgesystem (central module plus connection activation system)and the programmable blackboard are shown in Figure 6 (fromMcClelland, 1985).
FIGURE 5. Each CA node projects to the corresponding connec-tion in both programmable modules, and each central letter nodereceives projections from the corresponding programmableletter node in both programmable modules. The inputs to twocentral letter nodes, and the outputs from two CA nodes areshown (from McClelland, 1985).

Jacobs and Jordan, 1993; Cho and Kim, 1995). The keyissue in this approach is how to combine the resultsof the individual networks to give the best estimate ofthe optimal overall result. Architectures used in thisapproach consist of two types of networks: an expertand a gating network. Basically, the expert networkscompete to learn the training instances, and the gatingnetwork facilitates cooperation by the overall mediationof this competition. The expert networks may be trainedseparately using their own preassigned sub-tasks anddiffering modalities (e.g. vision and touch), or the samemodality at different times (e.g. the consecutive 2-Dviews of a rotating 3-D object). The gating networkneed only have as many output units as there are expertnetworks.
To train such a gating network, Hampshire and Waibel(1992) developed a new form of multiplicative connec-tion, which they call the ‘Meta-Pi’ connection. Its func-tion is closely aligned with predecessors described inMcClelland (1986). The final output of the overallsystem is a linear combination of the outputs of theexpert networks, with the gating network determiningthe proportion of each local output in the linearcombination. Figure 6 illustrates this architecture withthree expert networks.
The final output of the overall system is a linearcombination of the outputs of the expert networks, withthe gating network determining the proportion of eachlocal output in the linear combination. The Meta-Pigating network allocates appropriate combinations ofthe expert networks when stimuli are assessed to benovel, while an automatic (‘non-conscious’) decisionprocess operates in instances where a single expert canexecute the task. This coupling of modular, expertnetworks and gating controls produces new levels ofcooperative behavior. The expert networks are local inthe sense that the weights in one network are decoupledfrom the weights in other expert networks. However,
there is still some indirect coupling because if someother network changes its weights, it may cause thegating network to alter the responsibilities that getassigned to the expert network.
These examples from engineering applications ofmultiplicative, gating networks are not based upon theWagon Wheel model or, for that matter, any specificneural circuitry. Yet Koch (1997) notes that
Multiplication is one of the most common operations carriedout in the nervous system (for example, for estimatingmotion or the time-to-contact with an approaching stimulus)(p. 208).
We are not aware of any studies of either the axoncollateral or dendro–dendritic projections in nRt demon-strating multiplicative properties, but Mel (1994) hasmodeled such connections in the NMDA-rich dendritictrees of cortical pyramidal cells. He postulates that theyperform nonlinear pattern discrimination and correlativeoperations. Given the role of the bidirectional dendritictrees of nRt cells in globally synchronizing the thalamo-cortical circuit (Taylor and Alavi, 1993; LaBerge, 1995),it seems likely that they will eventually be found to haveimportant computational functions as well.
Even if it transpires that synchronous oscillations, notmultiplicative connections, are the basis for the ‘gating’functions of nRt upon the thalamocortical circuit, NNmodels based upon Meta-Pi connections may still beuseful for simulating global workspace systems. Theuse of Meta-Pi connections has already been extendedto synchronous oscillators in modular cortical neuralnetworks. Indeed, computational simulations of phase-locked oscillations characteristic of neurons involvedin the ‘binding’ of visual (Grossberg and Somers,1991; Sompolinsky et al., 1991) and auditory (Vibertet al., 1994) features of an attended object have alreadybeen extended to synchronous oscillators using Meta-Piconnections. Such oscillatory circuits have also been
1204 J. Newman et al.
FIGURE 6. Schematic diagram of modular neural networks with three expert networks and a gating network. The output of the entirearchitecture, denoted by Y, is Y ¼ g 1y 1 þ g 2y 2 þ g 3y 3, where y 1 denotes the output of the ith expert network.

employed in modeling storage and retrieval in patternrecognition tasks (Yao and Freeman, 1990).
In this paper, we have introduced a collection ofneuroscience and NN models for attention and binding,resource allocation, and second-order gating, whichshare important features and parallels with a NeuralGlobal Workspace System for conscious attention(Newman and Baars, 1993). While the NN models wehave presented only implement partial aspects of theGW system, and even our Wagon Wheel model largelyneglects the influences of memory and affective systemsupon the stream of consciousness, the outlines of ageneral framework for understanding conscious pro-cesses should be discernable (see Newman, 1997 for afuller account). This is certainly great progress, giventhe virtual terra incognita consciousness has been formost of the history of science.
REFERENCES
Baars, B. J. (1983). How does a serial, integrated and very limitedstream of consciousness emerge out of a nervous system that ismostly unconscious, distributed, and of enormous capacity? InG. R. Brock & J. Marsh (Eds),CIBA Symposium on Experimentaland Theoretical Studies of Consciousness(pp. 282–290). London:John Wiley and Sons.
Baars, B. J. (1988).A cognitive theory of consciousness. Cambridge:Cambridge University Press.
Baars, B. J. (1992).Experimental slips and human error: Exploring thearchitecture of volition. New York: Plemun Press.
Baars, B. J. (1994). A global workspace theory of conscious experience.Baars, B. J. and Newman, J. (1994). A neuro-biological inter-pretation of a Global Workspace theory of consciousness. InA. Revonsuo and M. Kamppinen (Eds),Consciousness in philo-sophy and cognitive neuroscience. Hillsdale, NJ: Erlbaum.
Baars, B. J. (1997).In the theatre of consciousness: The workspace ofthe mind. Oxford: Oxford University Press.
Baars, B. J., Newman, J. & Taylor, J. G. (in press). Neuronal mechan-isms of consciousness: A Relational Global Workspace framework.In S. Hameroff, A. Kaszniak, J. Laukes, & A. Scott (Eds),Towardsa science of consciousness: The second Tucson discussion anddebates. Cambridge, MA: MIT Press.
Buser, P., & Rougeul-Buser, A. (1995). Do cortical and thalamic bio-electric oscillations have a functional role? A brief survey.Journalof Physiology (Paris), 89, 249–254.
Cho, S.-B., & Kim, J. H. (1995). Multiple network fusion using fuzzylogic. IEEE Trans. Neural Networks, 6, 497–501.
Crick, F. (1984). Function of the thalamic reticular complex: the search-light hypothesis.Proceedings of the National Academy of Sciences,USA, 81, 4586–4590.
Crick, F., & Koch, C. (1990a). Towards a neurobiological theory ofconsciousness.Seminars in the Neurosciences, 2, 263–275.
Crick, F., & Koch, C. (1990b). Some reflections on visual awareness.Cold Spring Harbor Symposium on Quantitative Biology, 15,953–962.
Damasio, A. R. (1994).Descartes’ Error. New York: G.P. Putnam Sons.Deschenes, M., Madariage-Domich, A., & Steriade, M. (1985).
Dendrodendritic synapses in the cat reticularis thalami nucleus: Astructural basis for thalamic spindle synchronization.BrainResearch, 334,165–168.
Durfee, E.H. (1993). Cooperative distributed problem solving between(and within) intelligent agents. In P. Rudominet al. (Eds),Neuroscience: From neural networks to artificial intelligence(pp. 84–98). Heidelberg: Springer-Verlag.
Eccles, J. C. (1966).Brain and conscious experience. Heidelberg:Springer-Verlag.
Edelman, G. M. (1989).The remembered present, a biological theory ofconsciousness. New York: Basic Books.
Funke, K., & Eyse, U. T. (1992). EEG-dependent modulation ofresponse dynamics of cat dLGN relay cells and the contributionof corticogeniculate feedback.Brain Research, 573,217–227.
Fuster, J. M. (1980).The prefrontal cortex. New York: Raven Press.Gazzanigga, M. S. (1985).The social brain, discovering the networks of
the mind. New York: Basic Books.Goldman-Rakic, P. S. (1988a). Changing concepts of cortical connec-
tivity: parallel distributed cortical networks. In P. Rakic andW. Singer (Eds),Neurobiology of the cortex(pp. 177–202).Berlin: John Wiley and Sons Ltd.
Goldman-Rakic, P. S. (1988b). The prefrontal contribution to workingmemory and conscious experience. In O. Creutzfeld and J. Eccles(Eds), The brain and conscious experience. Rome: PontificalAcademy.
Groenewegen, H. J., & Berendse, H. W. (1994). The specificity of the’nonspecific’ midline and intralaminar thalamic nuclei.Trends inNeuroscience, 4(2), 52–58.
Grossberg, S., & Somers, D. (1991). Synchronized oscillations duringcooperative feature linking in a cortical model of visual perception.Neural Networks, 4, 452–466.
Hampshire II, J.B., & Waibel, A. (1992). The Meta-Pi network: Build-ing distributed knowledge representations for robust multisourcepattern recognition.IEEE Trans. Pattern Analysis and MachineIntelligence, 14, 751–769.
Harnad, S. (1994). Guest editorial — Why and how we are not zombies.Journal of Consciousness Studies, 1(2), 164–168.
Heilman, K. M., Watson, R. T. & Valenstein, E. V. (1985). Neglect andrelated disorders. In K.M. Heilman & E.V. Valenstein (Eds),Clinical neuropsychology. New York: Oxford University Press.
Jackendorf, R. (1987).Consciousness and the computational mind.Cambridge, MA: MIT Press.
Jacobs, R. A., Jordan, M. I., Nowlan, S. J., & Hinton, G. E. (1991).Adaptive mixtures of local experts.Neural Computation, 3, 79–87.
Jacobs, R. A., & Jordan, M. I. (1993). Learning piecewise controlstrategies in a modular neural network architecture.IEEE Trans.Systems, Man, and Cybernetics, 23, 337–345.
Jasper, H. H. (1960). Unspecific thalamocortical relations. In J. Field,H. W. Magoun & V. E. Hall (Eds),Handbook of neurophysiology,Vol. 1 (pp. 1307–1322). Washington, DC: American PhysiologicalSociety.
Johnson-Laird, P. N. (1988).The computer and the mind. Cambridge,MA: Harvard University Press.
Jones, E. G. (1985).The thalamus. New York: Plenum Press.Kihlstrom, J. F. (1987). The cognitive unconscious.Science, 237,
285–292.Koch, C. (1997). Computation and the single neuron.Nature, 385,207–
210.Koch, C., & Poggio, T. (1992). Multiplying with synapses and neurons.
In T. McKenna, J. Davis & S. F. Zornetzer (Eds),Single neuroncomputation(pp. 315–345). Boston, MA: Academic Press.
Krosigk, von M., Bal, T., & McCormack, D. (1993). Cellular mechan-isms of a synchronized oscillation in the thalamus.Science, 261,361–364.
LaBerge, D. L. (1990). William James symposium: Attention.Psycho-logical Science, 1(3), 156–162.
LaBerge, D. L. (1995).Attentional processing: The brain’s art ofmindfulness. Cambridge, MA: Harvard University Press.
Llinas, R. R., & Pare, D. (1991). Commentary: of dreaming andwakefulness.Neuroscience, 44(3), 521–535.
Llinas, R., Ribary, U. Joliot, M. & Wang, X-J. (1994). Content andcontext in temporal thalamocortical binding. In G. Busakiet al.(Eds), Temporal coding in the brain(pp. 251–272). Heidelberg:Springer-Verlag.
McClelland, J. L. (1985). Putting knowledge in its place: A scheme for
1205Neural Global Workspace Model

programming parallel processing structures on the fly.CognitiveScience, 9, 113–146.
McClelland, J. L. (1986). The programmable blackboard model of read-ing. In J. L. McClelland & D. E. Rummelhart (Eds),Paralleldistributed processing, Vol. 2 (pp. 122–169). Cambridge, MA:MIT Press.
McCormick, D. A., & Krosigk, M. (1992). Corticothalamic activationmodulates thalamic firing through glutamate metabotropicreceptors.Proceedings of the National Academy of Science USA,89, 2774–2778.
Mel, B. W. (1994). Information processing in dendritic trees.NeuralComputation, 6, 1031–1085.
Mesulam, M. (1985).Principles of behavioral neurology. Philadelphia:F.A. Davis.
Minsky, M. (1979). The society theory. In P. H. Winston & R. H. Brown(Eds),Artificial intelligence, an MIT perspective, Vol. 1(pp. 423–450). Cambridge, MA: MIT Press.
Mitrofanis, J., & Guillery, R. W. (1993). New views of the thalamicreticular nucleus in the adult and developing brain.Trends inNeuroscience, 16, 240–245.
Monchi, O. & Taylor, J. G. (1995). A model of the prefrontal loop thatincludes the basal ganglia in solving a recency task,Proceedings ofthe International Neural Network Society Annual Meeting, July1995. Washington, DC: International Neural Network SocietyPress.
Newell, A. (1992). SOAR as a unified theory of cognition: Issues andexplanations.Behavioral and Brain Sciences, 15(3), 464–492.
Newman, J. (1995a). Review: Thalamic contributions to attention andconsciousness.Consciousness and Cognition, 4(2), 172–193.
Newman, J. (1995b). Reticular-thalamic activation of the cortexgenerates conscious contents.Behavioral and Brain Sciences,18(4), 691–692.
Newman, J. (1997). Putting the puzzle together: Towards a generaltheory of the neural correlates of consciousness.Journal of Con-sciousness Studies, 4 (1 and 2), 47–66 and 99–120.
Newman, J., & Baars, B. J. (1993). A neural attentional model foraccess to consciousness: A Global Workspace perspective.Con-cepts in Neuroscience, 4(2), 255–290.
Newman, J., Baars, B. J. & Cho, S-B. (1997). A neurocognitive modelfor attention and consciousness. In S. O’Nuallain, P. McKevitt &E. MacAogdin (Eds),Two sciences of mind: Readings in cognitivescience and consciousness. Philadelphia, PA: John Benjamins ofNorth America.
O’Nuallain, S., McKevit, P. & MacAogdin, E. (1997).Two sciencesof mind: Readings in cognitive science and consciousness.Philadelphia, PA: John Benjamins of North America.
Parent, A., & Hazrati, L.-N. (1995). Functional anatomy of the basalganglia. I. The cortico-basal ganglia–thalamo–cortical loop.BrainResearch Reviews, 20, 91–127.
Penfield, W. (1975).The mystery of the mind: A critical study ofconsciousness and the human brain. Princeton, NJ: PrincetonUniversity Press.
Penrose, R. (1994).Shadows of the mind—In search of the missingscience of consciousness. Oxford: Oxford University Press.
Posner, M. I. & Rothbart, M. K. (1991). Attentional mechanisms andconscious experience. In A. D. Milner & M. D. Rugg (Eds),Theneuropsychology of consciousness(pp. 11–34). London: AcademicPress.
Posner, M. I. (1994). Attention: The mechanisms of consciousness.Proceedings of the National Academy of Science USA, 91,7398–7403.
Reddy, D. R., Erman, L. D., Fennell, R. D., & Neely, R. B. (1973). TheHearsay speech understanding system: An example of the recogni-tion process.Proceedings of the International Conference onArtificial Intelligence, 185–194.
Rummelhart, D. E., Hinton, G. E. & McClelland, J. L. (1986). A generalframework for parallel distributed processing. In J. L. McClelland &D. E. Rummelhart (Eds),Parallel distributed processing, Vol. 1(pp. 43–76). Cambridge, MA: MIT Press.
Scheibel, M. E., & Scheibel, A. B. (1966). The organization of thenucleus reticularis: A Golgi study.Brain Research, 1, 43–62.
Scheibel, A. B. (1980). Anatomical and physiological substrates ofarousal: A view from the bridge. In J. A. Hobson & M. A. B. Brazier(Eds), The reticular formation revisited(pp. 55–66). New York:Raven Press.
Sillito, A., Jones, H., Gerstein, G., & West, D. (1994). Feature-linkedsynchronization of thalamic relay cell firing induced by feedbackfrom the visual cortex.Nature, 369,479–482.
Skinner, J. E. & Yingling, C. D. (1977). Central gating mechanisms thatregulate event-related potentials and behavior. In J. E. Desmedt(Ed.), Progress in clinical neurophysiology: Attention, voluntarycontraction and event-related cerebral potentials, Vol. 1(pp. 30–69). Basel: Karger.
Sompolinsky, H., Golomb, D., & Kleinfeld, D. (1991). Cooperativedynamics in visual processing.Physical Review A, 43(12),6990–7011.
Steriade, M., & Llinas, R. R. (1988). The functional states of thethalamus and the associated neuronal interplay.PhysiologicalReviews, 68(3), 649–742.
Stryker, M. P. (1989). Is grandmother an oscillation?Nature, 338,297–337.
Taylor, J. G. (1992). Towards a neural network model of the mind.Neural Network World, 2, 797–812.
Taylor, J. G. (1996). A competition for consciousness?Neuro-computing, 11, 271–296.
Taylor, J. G. & Alavi, F. N. (1993). Mathematical analysis of a compe-titive network for attention. In J. G. Taylor (Ed.),Mathematicalapproaches to neural networks(pp. 341–382). Amsterdam:Elsevier Science Publishers B.V.
Taylor, J. G. & Michalis, L. (1995). The functional role of thehippocampus in the organization of memory.Proceedings ofthe International Neural Network Society Annual Meeting, July1995. Washington, DC: International Neural Network SocietyPress.
Vibert, J., Pakdaman, K., & Azmy, N. (1994). Interneural delaymodification synchronizes biologically plausible neural networks.Neural Networks, 7, 589–607.
Walshe, K. W. (1978).Neuropsychology, A clinical approach.Edinburgh: Churchill Livingstone.
Yao, Y., & Freeman, W.J. (1990). Model of biological pattern recog-nition with spatially chaotic dynamics.Neural Networks, 3,153–170.
Yen, C. T., Conely, M., Hendry, S. H. C., & Jones, E. G. (1985). Themorphology of physiologically indentified GABAergic neuronsin the somatic sensory part of the thalamic reticular nucleus in thecat.Journal of Neuroscience, 5, 2254–2268
1206 J. Newman et al.

1997 SPECIAL ISSUE
Neural Networks for Consciousness
J. G. Taylor
Department of Mathematics, King’s College and Institute for Medicine, Research Centre Juelich
(Received9 March 1997;accepted28 June1997)
Abstract—The paper outlines a three-stage neural network model for (i) the emergence of consciousness at its lowestlevel of phenomenal experience, (ii) the development of actions on the emerged conscious activity so as to generatehigher-order consciousness. In the model, the lower first stage involves modules transforming inputs into various codes.It is only at the second level that further modules are activated in a manner which brings about the emergence ofphenomenal awareness. The evidence in support of the initial two-stage model is briefly summarised. The two-stagemodel is then developed by means of a detailed neural framework which allows analysis of the extended duration ofactivity in posterior working memory sites.The third stage is then described in terms of known neural structures in thefrontal lobes, and the manner they might support the creation of higher consciousness is briefly presented.q 1997Elsevier Science Ltd.
1. INTRODUCTION
The race for consciousness has started (Taylor, 1998).There is a large amount of interest in the neuroscienceand neural network community on the topic. Variousgroups are seriously trying to track it down by the useof non-invasive instruments (PET, fMRI, EEG andMEG) by observing the networks of connected moduleswhich function so as to enable various high level cogni-tive tasks to be solved by the brain.
The current body of knowledge on consciousness israpidly increasing. There is already a large reservoir ofmaterial on it discovered by psychologists over the lastcentury, and this is now being combined with the resultscoming from non-invasive instruments, and at single celllevel in monkeys, to attempt to indicate where and howconsciousness arises, and is supported in the brain.
Various models of consciousness have been suggestedin the past. These give answers to the two questionsindicated in Figure 1 associated with the questionmarks inserted there. The figure shows schematicallythe preprocessing of input by the modules labeled 1,and its further analysis by the modules 2. It is the latter
which are supposed to support the initial appearanceof consciousness of the incoming stimuli. The twoquestion marks attached to the flow of activity inFigure 1 are related to the following basic questionsabout consciousness:
1. Q1: how is content supplied to the conscious experi-ence once it has been created?
The second question mark is attached to the ‘‘hard’’question of consciousness (Levine, 1983):
2. Q2: how does the transformation of activity from themodules 1 to those in 2 cause phenomenal awarenessto arise in the first place?
These two questions are closely related to each other,but are logically distinct in that content may have manypossibilities, but the underlying phenomenal experienceitself appears to have a relatively constant set of attri-butes (which will be considered in due course). Question1 attempts to explain how these variations of awarenessoccur, whilst question 2 explores the principles behindthat emergence in the first place. It might be suggestedthat one should answer question 2 before question 1,but indeed the former is very hard to give a convincingexplanation. That is why a number of reasonablyacceptable proposals have been made to answer question1, but with no acceptable solution to question 2 in sight.
The most promising of the answers to question 1 is interms of some form of feedback from earlier memories,both of semantic and episodic form. Thus, there is the‘Remembered Present’ (Edelman, 1989), the ‘Relational
Neural Networks, Vol. 10, No. 7, pp. 1207–1225, 1997q 1997 Elsevier Science Ltd. All rights reserved
Printed in Great Britain0893–6080/97 $17.00+.00
PII: S0893-6080(97)00064-6
Pergamon
1207
Acknowledgements: The author would like to thank Prof. H.-W.Mueller-Gaertner, Director of the Institute for Medicine, forhospitality at the Institute where this paper was written, and to hiscolleagues A. Ionnides, L. Jaencke, S. Posse and J. Shah forstimulating conversations. I would also like to thank S. Petersenfor the simulation of Figure 10.
Requests for reprints should be sent to Dept of Maths, King’sCollege, Strand, UK; E-mail: [email protected].

Mind’ (Taylor, 1973, 1991) and the ‘Predicted Present’(Gray, 1995) as candidate versions using, in one way oranother, relational structures involving past memoriesin one form or another. However, these approaches donot seem to help solve question 2. It is the purpose ofthe first part of this paper to propose the bare bonesof such a solution.
There is also the unresolved question as to how furtherhigher level cognitive experience arises and is related tothat of phenomenal experience. This leads to our thirdquestion:
Q3: Is there a dissociation between higher level cog-nitive processes (thinking, planning, self-awareness) andthe lower level phenomenal experience, and if so whatare the additional neural structures and mechanismswhich support the higher level processes?
There has already been acceptance of some level ofdissociation between passive awareness and controlledcognition in presenting the two-stage model of Figure 1,since otherwise it would not be possible to leave outthe higher levels. The three stage model of Figure 2does assume some dissociation but may simply beregarded as a summary of known anatomy, with themodules at stage 2 being in posterior cortex and stage-three modules in frontal lobe. However, the process ofmodeling the actions of the various parts of the morecomplete structures of Figure 2 would need to be per-formed with some knowledge of the level of any suchdissociation. That will be considered at a later stageof the paper; for the moment we will assume that themethodology we are adopting, of modeling the lowertwo stages before attacking the third stage, will havesome chance of being successful.
The detailed contents of the paper are as follows. Itcommences in Section 2 with an outline of the two-stageneural model for the emergence of consciousness. In thismodel the lower first stage involves modules transform-ing inputs into codes in the visual or other modalities. Itis only at the second higher level that further modules areactivated in a manner which brings about the emergenceof phenomenal awareness. The evidence in support ofsuch a two-stage model will be briefly summarised inSection 3.
The two-stage model is then developed in Section 4by means of a detailed neural framework which allowsanalysis of the extended duration of activity in posteriorworking memory sites (which are posited as at the second
stage in the model) using the notion of ‘bubbles’ of activ-ity which are formed in neural models with enoughneurons and suitably strong recurrent excitatory andinhibitory connections (Amari, 1977; Taylor, 1997a).Such models have recently been suggested as explainingthe development of observed orientation sensitivity inprimary visual cortex (Douglas et al., 1995; Somerset al., 1995; Ben-Yishai et al., 1995). The purpose ofSection 4 is to discuss the implications of the resultingdynamical activity for the two-stage model of theemergence of phenomenal consciousness.
Detailed neural models are then briefly summarisedfor
1. the initial emergence of consciousness in terms ofsemi-autonomous ‘bubbles’ of activity in a neuralsystem of the form of a continuum neural fieldtheory (CNFT), as developed in a two-dimensionalcontext (Taylor, 1997a);
2. application of CNFT to give brief initial explanationsof various visual phenomena: apparent motion (due tomoving bubbles) and stabilised image effects(destruction of bubbles);
3. how working memory modules, with extended tem-poral activity, can arise as from CNFT for suitablyhigh density neuronal content (together with suitableionically-driven adaptation);
The initial two-stage model is then extended to a thirdstage, as shown in Figure 2, in Section 5. There is muchinvestigation presently as to the extent of possible disso-ciations between posterior sites of working memory, the
1208 J. G. Taylor
FIGURE 1. The basic two-stage model of awareness. Module 1performs preprocessing in various modalities and codes;activity in module 2 is that on which phenomenal awareness isbased. The two question marks denote the questions Q1 and Q2stated in the text.
FIGURE 2. The three-stage model for the creation and control ofawareness. The modules at the lowest or first stage are denotedA 1 , A 2 , ... and are involved only in low level feature analysis inthe appropriate modality. A number of modules are involvedin such processing. At the next stage are sites of ‘buffer’ memoryin posterior cortex, denoted B 1, B 2, .... They are fed by prepro-cessed activity from first stage modules, and are coupled to eachother so as to run a competition by lateral inhibition so as toselect the most appropriate percept, given earlier context stillpresent in activity traces on these modules, to enter phenomenalawareness. The final third stage is composed of modules whichare frontal and possess attentional feedback control both tothe second stage modules, so as to refresh activity there,and to the first stage modules to reduce or increase stimulusselectivity. Feedback is shown explicitly from the third to lowerstages although it will also be present from the second to firststage (not all feedback is shown explicitly).

so-called ‘buffer sites’ of short term memory, and theanterior ‘active memory’ sites (Fuster, 1993). Thenature of this evidence will be summarised and a third-stage model suggested in Section 6 which appears to beconsistent with presently know data.
The paper finishes with a discussion, particularly onhow the anterior activations can create the experiencedfeatures of higher consciousness.
2. THE TWO-STAGE MODEL OF AWARENESS
The two-stage model of the emergence of awareness inthe brain is shown in Figure 1. There is supposedly noawareness of activity in stage 1 modules, whilst it is inmodules at stage 2 that activity can enter awareness.What sort of neural networks would be suitable forsuch a separation? Some features of them have beensummarised in Table 1, derived from analysis of intro-spection by psychologists and philosophers (and sum-marised in Metzinger, 1995). As seen from the table,there are spatial and temporal features of the networkswhich are of great importance. Thus, there are spatialfeatures which require localised representations in setsof well-connected modules which are also well coupledto those modules involved in higher cognitive processes(the modules C of the third stage of processing inFigure 2).
The temporal features are also very specific, requiringsuitably long temporal duration of activity (of the orderof 300–500 ms) for phenomenal experience (PE) to arisebut with little time between one experience and thenext. Finally, there are emergent features which arealso non-trivial, involving one-way and rapid emergenceof PE at the highest level of a processing hierarchywith no ability to gain awareness of the activity oflower modules of stage 1.
Coding at the highest level of activity emerginginto PE is most simply obtained by assuming that itarises from those first stage modules which are alsocoded at the second-stage level. The other most
important feature in Table 1, that of the temporal aspectof duration, is a crucial part of the general features ofthe buffer stores in the psychologically-based workingmemory model (Baddeley and Hitch, 1974; Baddeley,1986). These buffers have input from semantic-levelcoding, where all possible interpretations (in the caseof words) have already been accessed in the semanticmemory, so at the end of the stage 1 processing, beforethe buffer store is attained (Marcel, 1980; Levelt et al.,1991).
The two-stage model of Figure 1 has been developedin a neural network implementation elsewhere (Taylor,1996a). It uses semantic-level coding of activity at stage1 and then buffered activity at stage 2, with lateralinhibition at this stage to achieve singling out of thecontextually appropriate representation at the higherstage. It is this activity which then enters awarenessby being broadcasting around the ‘global workspace’ of(Baars, 1988) or, in terms of the three stage modelof Figure 3 gaining access to the anterior sites of workingmemory. Once at that third and highest level the materialcan be rehearsed or manipulated as desired. A neuralnetwork model of this higher level processing will alsobe described later in the paper.
Besides the construction of a model of the two-stageprocess, and due to the plethora of buffer stores nowobserved in posterior cortex in various modalities andcodes (Salmon et al., 1996; Paulesu et al., 1993; Smithand Jonides, 1995) there must be some way of combiningtheir various activities so as to achieve a unified experi-ence of consciousness. Control structures in the brainare thereby needed which produce global correlationsbetween the various component working memories. Inparticular, these control structures should support com-petition between various inputs, when suitably encoded,and only allow certain, most relevant, memories to beactivated and related to the corresponding winning input.Such structures may also be involved with assessingthe level of discrepancy of new incoming input withthat predicted from later parts of activated stored patternsequences. These aspects were considered (Taylor, 1992,1993, 1994; Taylor and Villa, 1997), in terms of possiblenetworks which could perform pattern matching andsustain competition. One of these involved the nucleusreticularis thalami (NRT), a sheet of mutually inhibitoryneurons interposed between thalamus and cortex (seeFigure 3).
It was suggested that the thalamus-nucleus reticularisthalami(NRT)-cortex complex may support such activ-ities. This is due to the fact that the NRT is composedalmost entirely of inhibitory neurons which are fed byactivity coming from thalamus up to cortex and alsoby reciprocal cortico-thalamic connections. Since NRTalso sends inhibitory inputs down onto thalamus, it isclear that such inhibition (which could also function asa release from inhibition if the NRT targets inhibitoryinterneurons in thalamus more effectively than the
1209Neural Networks for Consciousness
TABLE 1Criterial features on neural networks to support PE
Spatial features:(a) localised representations in localised modules(b) modules well-coupled together(c) modules well-coupled to higher-level modules
Temporal features:(a) temporal continuation of activity(b) time required to achieve PE activation(c) no gap between different PE activations
Emergent features:(a) one-way creation of PE activation(b) PE created at highest level of a hierarchy(c) rapid emergence once begun(d) no ability to probe lower levels supporting PE but below PE
creation level

excitatory relay cells to cortex) could exert a powerfulcontrol influence on cortical activity. This has beenshown experimentally to be the case, with global effectsof the NRT sheet especially being observed in themanner in which NRT controls the nature of corticalpatterns of activity in sleep. There is also some evidencefor a similar global form of control by NRT on allowedcortical activity in the non-sleep states (Skinner andYingling, 1977; Villa, 1988).
It is possible, in a general manner, to understandthis global control achieved by NRT of cortical activityin the following manner. Since any localised activity on
the NRT will try to damp down activity on it elsewhereby lateral inhibition, the NRT can sustain ‘bunched’spatially inhomogeneous activity, in which competitionbetween neighbouring thalamic or cortical inputs onto itis occurring. This spatially structured activity may occurglobally over the whole NRT sheet if it is well enoughconnected laterally, as is seen to be the case in spindlegeneration in sleep (Steriade et al., 1990). In this mannerthe NRT may function as a global controller of corticalactivity. As such it appears of great relevance to includein models of the control circuitry for consciousness(Taylor, 1992, 1993, 1996b; Alavi and Taylor, 1992,1993, 1995; Baars and Newman, 1993, 1994; Harth,1995; Kilmer, 1996). Known local circuitry (Steriadeet al., 1990) was used to construct a simulation ofthe coupled thalamo-NRT-cortical system followingthe circuit of Figure 4; the resulting activity arisingfrom a simulation of 100 cortical neurons is shown inFigure 5. The resulting extended two-stage model isshown in Figure 6.
In summary the proposed two-stage model has thefollowing features:
1. coding occurs up to semantic level (denoted SM) inthe first (preprocessing) stage in Figure 6,
2. there is feedforward transfer of activity from the firstto the second stage at approximately the same level ofcoding, with the feedforward map being
3. SM → WM4. in a given code (as shown in Figure 6),5. duration of activity is longest of all posterior modules
in the WM of the second stage,6. there is a process of global competition between
1210 J. G. Taylor
Cortex
Thalamus
Main Nuclei of Thalamus
Nucleus Reticularis Thalami Cerebellum
FIGURE 3. The Nucleus Reticularis Thalami (NRT) is a sheetof inhibitory neurons surrounding the upper and lateral partsof the thalamus. It only gives output to the thalamus as well asby lateral connections to other parts of itself, whilst receivingcollaterals from both thalamo-cortical and cortico-thalamicaxons.
C1
C2
C3
Cortex NRT Thalamus
f1
f2
f3
T1
T2
IN3
N1
N2
N3 T3
IN3
IN3
FIGURE 4. Circuit used to simulate the global competitive controlaction of the NRT over thalamic inputs to cortex, and hence tocortical activity itself. Input I enters both the main relay cells,denoted T, and the associated inhibitory interneurons IN in tha-lamus, and then feeds topographically to cortical cells C. Thereare collaterals from the cortico-thalamic axons and the thalamo-cortical ones feeding the NRT cells N, whcih are also laterallyconnected. These latter also feed back to both the inhibitoryand relay cells in the thalamus, having greatest inhibitory effecton the former (from Alavi and Taylor, 1993).
Cor
tex
(vol
ts)
NR
T
(vol
ts)
OU
T
(Tha
lam
.)O
UT
(I
nhib
.)In
put
(vol
ts)
FIGURE 5. Results of a simulation of the circuit of Figure 4 for aline of 100 cells, for an input given on the bottom line. There is aglobal wave of activity set up over the NRT, as seen in the activa-tion curve iin the second line of the figure. This causes a reducedlevel of input to cortical cells, as their activity indicate, shown onthe top line.

activities on different WMs, supported by theTH/NRT/C system,
7. there is a strong reciprocal connectivity between theposterior WM sites and frontal areas at the highestlevel of coding (to allow rapid and effective accessto the frontal ‘active’ working memory sites). Suchconnections are observed neuroanatomically.
The above model of the two-stages of phenomenalawareness leaves unanswered many detailed points. Inparticular:
1. what is the dynamical origin of the temporallyextended activity traces on the working memorysites?
2. how are the codes in the WMs built up?3. how do features become bound across codes and
modalities?4. what is the temporal dynamics of the emergence of
awareness?
Answers to some of these questions (especially the 1st,2nd and 4th) will be developed in Section 4. Before thatwe turn in the next section to describe some of the experi-mental basis for the separation of the emergence ofawareness in two parts, as the 2-stage model claims.
3. EXPERIMENTAL SUPPORT FOR THETWO-STAGE MODEL.
The two-stage model supposes that there are two sorts ofcortical processing: firstly at the level of the stage-1modules in Figure 1 and supporting neural activity ofwhich there is no direct awareness, and secondly that atthe level of stage-2 modules of Figure 1, of which there isdirect awareness. This is not a new idea although it hasbeen severely challenged in the past.
The reason for such a challenge is not too difficultto discern. If the two-stage model is true then theremust be some properties possessed by the stage-2modules which are not by the stage-1 modules. Suchproperties should be open to direct analysis by experi-ment and modeling, and perhaps allow the manner inwhich awareness is created to be determined. Onthe other hand, if there were no special modules in thebrain supporting awareness then consciousness couldpreserve its monolithic mystery; it would still be someunknown feature possessed by a complex enough set ofnerve cells but not one we can yet (if ever) see how toprobe. Of course the two-stage model shades into theone-stage, monolithic one as the preprocessing stagesof Figure 1 reduce in size and importance. Howeverthe existence of any modules at the non-aware levelwould still destroy the monolithic approach and makethe task of probing consciousness more hopeful.
There are two main sources of support for the two-stage model: altered experience brought about by defi-cits, and that caused by suitably subtle paradigms fornormal people. The first class contains modification ofmany forms of experience due to brain damage broughtabout by accident or disease. All of these are germane tothe general program of explaining the mind in terms ofthe brain but some are more crucial to the task at hand,that of supporting the two-stage model, so will bedescribed briefly in this section.
The two most important deficits to be discussed hereare those of neglect and of blindsight. Neglect involvesthe inability to notice inputs to one side of the body.Thus, a patient will be unable to notice one side oftheir field of view; if such a patient is shown a picturecontaining two houses, placed one above the other, withone of the houses having flames emerging from its leftside, then the patient may well prefer the house withoutthe flames if asked to choose between them. This andother tests (Bisiach, 1988) indicate that the patient hasknowledge up to a high level about the neglected side oftheir field of view but it is not part of their phenomenalexperience.
Blindsight is a similar phenomenon in which there isknowledge without awareness. It is a phenomenon whichhas been well documented (Weiskrantz, 1986), andoccurs to those who have lost a part of the primaryvisual cortex and so cannot see things in that part ofthe visual field. Yet they have knowledge of movingspots of light in their blindfield, since they can guessabove chance as to where a spot of light has movedacross a screen in front of them. It has even beenfound, in some cases, that the patient possesses someform of awareness of the movement-termed ‘contentlessawareness’ (Weiskrantz et al., 1995), if it is fast enough.Thus, it would appear as if there is still the ability ofmodules after V1 (the primary visual area) to createawareness provided there is a large enough input bysome alternate route.
1211Neural Networks for Consciousness
FIGURE 6. The resulting more detailed first two stages ofthe networks of the 3-stage model. The various features arediscussed in the text.

Besides the cases brought about by damage anddisease there is also the phenomenon of ‘normal blind-sight’ (Kolb and Braun, 1995). A subject is required topick out from an array of short bars oriented in a parti-cular direction a small subset oriented in the perpendi-cular one. It is found that, provided the field of oppositelyoriented bars is small enough, the subject is not aware ofthem but can be correct as to where they are more oftenthan by chance alone. Here again there is knowledgewithout awareness, so that the orientation analysis, per-formed very likely in V1, is producing knowledge whichis below the level of awareness.
Further support for V1 being below the level of aware-ness has also been given by an experiment which usesthe adaptation of subjects to oriented bars, which takesplace over about 5 s of viewing the bars (He et al., 1996).Once such adaptation has taken place, and before recov-ery, the contrast threshold for detecting oriented bars ofthe same orientation is found to have increased consider-ably. If the bars were ‘crowded’ by having bars of oppo-site orientation surrounding them in the field of view,loss of awareness of the particular bars now occurs butthere is still an increase of the threshold for detectingbars presented at a variable contrast thresholds. The phe-nomenon of crowding is also found to be asymmetricbetween the upper and lower halves of the field ofview, an asymmetry not possessed by V1. This meansthat V1 is involved in the adaptation to the orientationanalysis but not in the final production of awareness. Asthe authors concluded ‘activation of neurons in V1 isinsufficient for conscious perception’.
This is further supported by the so-called waterfalleffect or ‘motion after-effect’ (MAE). This occurs asan adaptation after about 30 seconds of viewing a movingscene; on cessation of the motion there is an apparentmotion of the now static scene in the opposite direction.The time course of this has been investigated under fMRI(Tootell et al., 1996a) where they detected in subjectscontinued activity in the motion-sensitive area V5 forabout the same length of time that the MAE persists inhumans (about 9 s). even more interestingly theyobserved (Tootell, 1996b) a longer time course of theeffect (by about 3 s) in higher visual areas beyond V5.This supports the notion that awareness arises in siteswith about 3 seconds of duration of neural activations.
Single cell data from monkeys are also relevant to thesearch for the modules involved in the creation of con-sciousness. Binocular rivalry (in which different imagesare presented to the two eyes and the percept will switchevery few seconds between the two possibilities) hasbeen used in monkeys to determine in which area thereis a predominance of neurons signaling the percept whenit is experienced (which they can be trained to do faith-fully). The use of this technique, over several years, hasled Logethitis and colleagues (Logothetis and Schall,1989; Leopold and Logothetis, 1996) to be able toshow that, in vision, certain earlier areas (V1, V2, V4)
have about equal numbers of neurons firing both forthe percept they support (in terms of direction ofmotion, for the paradigm in which the rivalry is betweentwo sets of moving bars going vertically upwards versusmoving downwards; a neuron supports a movementdirection if it fires preponderantly when that directionof movement is presented) and the opposite one. Asthey wrote, ‘The majority of cells in all areas continueto respond even when it is perceptually suppressed’.However their most recent analysis in the inferotemporalarea (Sheinberg and Logothetis, 1997) has shown that inareas TE and TEO there is now a majority of activeneurons in favor of the percept being experienced.Where the change-over area is sited is unclear and veryimportant to discover.
There is also support from psychophysical experi-ments on word processing, in which the processing of aword presented visually is probed by a further auditorily-presented probe word (Levelt et al., 1991). The conclu-sion of the study was that a two-stage process occurs inwhich at the first stage there is automatic processing ofall meanings of the word, which at a later stage arereduced to a suitably unique phonological representation.This is similar to the earlier two-stage ‘logogon’ modelof (Morton, 1969, 1979) both supporting the proposalof the two-stage model of Section 2 that awarenessarises at the second stage.
There is also increasing evidence in support of the
1212 J. G. Taylor
FIGURE 7. The levels of activation of the various modules of the3-stage model for a given input under the three conditions of a)ignore b) passive awareness c) directed attention. Under condi-tion (b) both the first two stage modules are activated, whilstunder (c) all are on. There will be differences expected betweenthe total levels of activation across the conditions due to feed-back modulation, so the levels in the different conditions arenormalized.

2-stage model from non-invasive measurements. Thus, ifa stimulus is ignored (by means of a suitably distractingtask in another modality) the activity in cortical areas canbe compared with that arising from passive awareness ofthe stimulus itself without any distractor. The 2-stagemodel would lead to the expectation of differences inactivation along the lines of that shown in Figure 7.These differences in activation level have been seen inan auditory listening task with an added attentional load,as shown in Figure 8 (Taylor et al., 1997). This is takenfrom an fMRI study of five subjects in which there iscomparison between subjects ignoring a sequence of syl-lables and listening passively to them. There is clearactivation of regions in BA44/45 (Broca’s area) andauditory areas when one passes from the ignore condition
to the passive listening condition; these are candidatesfor the emergence of phenomenal awareness of syllables.The activation in Broca’s area may be part of attetionalprocessing and not due to phenomenal awareness.
Similar support arises from a recent MEG experiment(Vanni et al., 1996) in which subjects were presentedwith line drawings of objects or of non-objects, with avisual mask then delivered 30, 45 or 106 ms later. Thelevel of the peak amplitude in only one visual area, theright lateral occipital (LO) area, was found to be signifi-cantly correlated with the level of awareness of theobject drawings. This result indicates that the rightcortical visual area LO is importantly involved inthe creation of awareness of the objects described bythe line drawings.
1213Neural Networks for Consciousness
FIGURE 8. Differences in activation at various levels of the brain from an fMRI experiment (Taylor, Jaenke et al, 1997) in which the passivelevels had subtracted from them levels due to ignore conditions for listening to a sequence of syllables. Note the non-zero levels ofactivity in various sites (threshold set at 5 leads to significance at p , 0.01).

There is also a set of recent fMRI studies determiningareas activated by various working memory tasks, suchas then-back task (Cohen et al., 1996). In this task asubject observes a sequence of letters or numbers pre-sented one per second and has to respond when the samesymbol occurs after n presentations (wheren has beentold to the subject by the experimenter before the parti-cular run, with values ofn ¼ 1 to 4). Various areas werefound to be significantly active compared to a back-ground task, in particular posterior sites in BA 40 andBA 18/19, as well as prefrontal sites whenn became 2 orgreater. This was especially so for the dorsolateral pre-frontal cortex (DLPFC), a fact important for our analysisof the third stage modules of Figure 2. However this areawas not active forn ¼ 1, so corresponding to posteriorlysited buffer memory (holding an object in memory over10 s), and supporting the dissociation between frontal(stage 3) and posterior (stage 2) cortical areas underlow-load conditions.
The conclusion of this section is that there is strongexperimental support for
1. the existence of two separate stages at the lowest levelin Figure 1,
2. the localisation of activity associated with the secondstage of Figure 1 to specialised sites in posteriorcortex, usually termed sites of ‘buffer memory’,
3. the determination of sites also involved in the stage-3modules of Figure 3, involved in the creation ofhigher -order consciousness.
4. dissociation of the third from the lower stages underlow attentional load.
4. A NEURAL NETWORK FOR THE TWO-STAGE MODEL
4.1. The Basics of Bubbles
We now turn to the development of a neural model tohelp support the two-stage model of Figure 1 and use it togive tentative answers to the questions raised at the endof Section 2. In particular we will consider the questions:
1. what is the dynamical origin of the temporallyextended memory traces residing in the short-termworking memory sites considered in this section?
2. how are the codes in these sites constructed?3. what are the neural principles behind the creation of
phenomenal awareness in these sites?
As stated in the Introduction we will employ conti-nuum neural field theory (CNFT) whcih has been usedover many years as a first approximation to cortical struc-ture (Amari, 1977; Beurle, 1956; Ermentrout and Cowan,1978). One of the important results of that period is thediscovery of cortical ‘bubbles’ of activity which areinitially created by input but which persist after stimulusoffset (Amari, 1977). A brief introduction to bubbles
and some of their properties in CNFT is given in thissubsection, and relevant applications developed in thefollowing one.
CNFT is based on a model of cortex as a continuoustwo-dimensional sheet of neurons with a lateral connec-tion with the shape of a ‘Mexican hat’. This allows thecreation of localised activity, the bubbles, which canpersist, and remain localised in spite of the absence ofinput due to the recurrent activity produced initiallycausing repeated firing of the neurons initially activated.The basic CNFT equation is constructed in terms ofthe membrane potential of a neuron and denoted byu(x,t), at the pointx and timet. It will be assumed thatthere is lateral connectivity other neural sheet definedby the lateral connection weight functionw(x ¹ x9)between the two neurons at the relevant pointsx. Theconnection weight will be usually taken to be of Mexicanhat form as a function of the Euclidean distance |x ¹ x9|.There is also an afferent connection weight functions(x, y) from the thalamic positiony to the corticalpoint x. The response function of a neuron will betaken to be determined by its mean firing rate,which is given as some functionf of the membranepotential u of the relevant cell.
The membrane potentialu(x, t) will satisfy the CNFTequation (Amari, 1977)
t]u(x, t)=]t ¼ ¹ u(x, t) þ
∫dx9w(x¹ x9)f [ u(x9, t) ]
þ
∫dy s(x,y) I (y, t) þ h ð1Þ
where I(y, t) is the input to the thalamic positiony attime t, h is the neuron threshold and the integrationover the lateral connection weight is over the manifoldM of neurons.
There are well-known autonomous solutions to (1) inthe case whenM is one-dimensional (Amari, 1977). Inthat case equation (1), for a static solution and with noinput, becomes:
u(x) ¼
∫w(x¹ x9) 1[u(x9)] dx9 þ h (2)
where the sharp threshold response functionf ¼ 1 (thestep function) has been assumed in (2). A ‘bubble’ isdefined to have a positive membrane potential over aninterval, independent of input. This is formalised as
4.1.1.Definition.A ‘bubble’ of neural activity is a loca-lised persistent solution to the CNFT equations. Its size isin general dependent on input, but its continued existenceis not (to within effects of adaptation).
Let us consider the bubble extending fromx ¼ 0 tox ¼ a:
u(x) . 0 , 0 , x , a; u(0) ¼ u(a) ¼ 0 (3)
and otherwiseu , 0. Then from (2) and (3), u is obtained
1214 J. G. Taylor

explicitly as
u(x) ¼
∫a
0w(x¹ x9) dx9 þ h¼ W(x) ¹ W(x¹ a) (4)
where the functionW is defined by
w(x) ¼
∫x
0w(x9) dx9 (5)
Necessary conditions for the bubble to exist are that themembrane potential vanishes at the ends of the interval[0, a], so
u(0) ¼ u(a) ¼ 0¼ W(a)9 þ h (6)
It is then possible to show thatu(x) . 0 for0 , x , a if h , 0;u(x) , 0 otherwise.
Stability of the resulting solution then requires
dW(a)=da , 0, ∨ w(a) , 0 (7)
Thus, the one-dimensional bubble exists under the con-ditions (6) and (7).
There are a number of further important resultsderived in Amari (1977) concerning the nature ofbubble solutions and their extension to input dependencewhich will be briefly summarised here:
1. the parameter ranges forh and for the parameters inWcan be determined so as to allow for autonomoussolutions of various types (B or the trivial one,`or the constant non-zero one, an a-solution as thebubble of finite length a described above, and a spa-tially periodic solution),
2. complete determination of those patterns which arestable and those which are unstable, from amongstthe stationary solutions described above,
3. response to input stimulus patterns: a bubble of finitelength moves to a position of maximum of the input,
4. two bubbles interact, if close, with attraction (fromthe Mexican hat connection weight function), ifmore distant with repulsion, and if very distant withno effect on each other,
5. there can occur spatially homogeneous temporaloscillations (between a layer of excitatory and oneof inhibitory cells)
6. traveling waves can persist.
Returning to the full two-dimensional bubble in theregion D (which will be called anR[2]-solution here),we define
W` ¼ limR→` W(R) (8)
where
W(R) ¼
∫D
w(x¹ x9)dx9 (9)
where x is only allowed to be on the boundary ofD,which has radiuslxl ¼ R.
It is now possible to extend the methods of Amari,
1977 to deduce the same results as in the one dimensionalcase for the questions (a) to (f) raised above. Using thesame enumeration:
1. Theorem 1. In the absence of input:
1. There exists aB solution iff h , 0.2. There exists an -solution iff W` . - h.3. There exists anR[2]-solution iff h , 0 andR . 0
satisfies
W(R) þ h ¼ 0 (10)
1. It is possible to extend the classification of the solu-tions for varying levels of the stimulus h. LetWm ¼
maxx W(x).
2. Theorem 2. The nature of the various solutions fordifferent parameter ranges is as in Figure 9.
1. To determine which of these solutions is stable it isnecessary to extend the one-dimensional discussion ofAmari (1977) to two (or higher) dimensions. From thetwo-dimensional extension of equation (6) the bound-ary of D, defined by the radiusR(t) at timet, satisfiesthe constraint
u(R(t), t) ¼ 0 (11)
1. On differentiation of equation (11) with respect totand use of equation (1) there results
dR=dt ¼ ¹ [G(R(t)) þ h]=tv (12)
2. wherev is gradient ofu normal to]D and is negative.The equilibrium case results on setting the right
1215Neural Networks for Consciousness
FIGURE 9. Parameter ranges for the existence of stable bubblesin two-dimensional CNFT (from Taylor, 1997a). The meaning ofthe paramteres is given in the text.

hand side of equation (12) to zero. The stability of thissolution is determined by the sign of dG/dR: dG/dR ,0 ⇔ stability. This leads to the stability classificationof the solutions as given in theorem 2.
3. The response to stationary inputs of anR[2]-solutioncan also be treated as in Amari (1977). Consider asmall stationary input« I(x), which is not assumedto be circularly symmetric so that the asymptoticbubble will not be circularly symmetric either.The equation of constraint is, following equation(12), that
dx=dt: =uþ ]u=]t ¼ 0 on ]D(t) (13)
1. Replacing the time derivative ofu on the left handside of equation (13) by equation (1) it is nowpossible to derive the condition forx(t) on the bound-ary of D(t),
dx=dt: =u ¼ (1t)[hþ «I þ G(lx(t)l)] (14)
2. On expanding in a perturbation series in the smallquantity«, with
llx(t)ll¼ R0 þ «(s, t) (15)
1. (wheres denotes a parameter specifying which pointon the boundary of the unperturbedR[2] is beingconsidered in the perturbation (15)) there results theconstraint
d«=dt: =u ¼ (1=t)[« þ «: =G(lx(t)l) ] (16)
2. where« ¼ «(s, t) is a vector describing the directionas well as the magnitude«(s, t) of the perturbation ofthe boundary pointx, and the derivatives in (16) areevaluated at« ¼ 0, so atx(t) ¼ R0 .
3. The result from the constraint (16) is that the netradial movement of the boundary ofD(t) is towardsthe region of largest input. There will be a movementof regions of]D(t) towards lower values of the input,if these are positive, but there will be a larger velocityof movement towards those regions of the boundarynearer the maxima ofI.
4. The one-dimensional result (d) above can also beextended in the same way, where the effect of oneregion (sayD1) on another (sayD2) is given, interms of the lateral interaction term in equation (2),as the effective input to a neuron inD2 at the pointxof amount
s(x) ¼
∫D1
w(lx¹ x9l)dx9 (17)
This will have the same effect as in the one-
dimensional case, with attraction between the bubblesat D1 and D2 if they are close enough (as deter-mined by s(x)), repulsion if the two regions arefurther separated, and ultimately no interactionbetween the bubbles at all if they are beyondthe range of the lateral interaction termw (if that isfinite).
5. The case of spatially homogeneous oscillationsextends immediately to the two-dimensional case,since only`-solutions are being considered.
6. This case involves temporal structure and is consid-ered more fully in Taylor (1997a).
4.2. Applications of Bubbles
We will apply the bubble solutions only to the cases ofapparent motion, the fading of stabilised images and thelifetime of bubbles in cortex. Bubbles have also beenapplied to a variety of other cortical processes: the devel-opment of topographic maps, both in the one dimensionalcase (Takeuchi and Amari, 1979) and for two dimensions(Taylor, 1997a), control of saccades by the superiorcolliculus, on which the bubbles are supposed to form(Kopecz and Schoner, 1995), the modifications of thesomatosensory constant topographic map by rapidunmasking due to removal of portion of an input(Petersen and Taylor, 1996a) or by relearning (Petersenand Taylor, 1996b), the guidance of head-directed cells(Zhang, 1996) and in explaining pre-attentive auditorymemory (May et al., 1995). The two cases to be consid-ered are closely related to features associated with thepossible manner in which bubbles could enter directlyinto perception, and so are most relevant for discussionhere.
4.2.1. Apparent Motion.This is the very well-knownphenomenon in which a spot of light appearing on ascreen in one place within 200 or so ms after the dis-appearance of one within 5 degrees of the second leadsto the experience to a viewer of the motion of the firstspot to the second. There has been considerable researchon this phenomenon (see, for example, Cavanagh andMather, 1990 and Ullman, 1979) and interesting attemptsto correlate and explain it (Dawson, 1991). Here CNFTwill be used to give an explanation at the level ofprinciple, without attempting to explain all of the detailsof the phenomenon by the model (which is discussedmore fully elsewhere).
The basic idea is the visual input from the initial spotcreates a bubble in a cortical area (very likely MT) whichis then dragged across the area by the appearance of thenew spot in the manner arising from the result (d) above.The disappearance of the first spot and the appearance ofthe second causes a change in the background in whichthe bubble created by the first spot resides. It is thusattracted to the central region of activity brought about
1216 J. G. Taylor

by the second spot on its appearance, following theabove argument. This process is difficult, although notimpossible, to analyse in detail using the time-dependentequation (1), but a one-dimensional simulation is shownin Figure 10. In this, a bubble is created by a spot attime t ¼ 0 at the position 30, using equation (1) withsuitable parameters for the input to cause the creationof a bubble. This initial input is then removed at time250 and a new input used at the position 70 at thattime. As seen from the figure the bubble created by thefirst spot reduces in size when its input is removed butpersists to move over to the position of the secondinput. It leaves a trail as it goes, causing the impressionof a moving input.
4.2.2.Stabilised Images.These are experienced when theretinal image is kept fixed on the eye and the imageultimately fades from view (Pritchard, 1963). The initialloss of image may be due to adaptation in the retinalpathway, so reducing the image able to reach laterstages. The process leads to very interesting patterns ofimage loss in which there are both Gestalt-type andensemble effects observed. One particularly interestingeffect was observed by (Crane and Piantanida, 1983) bystabilising the line between a rectangle of green and oneof red. Normally it is not possible to observe both greenand red together (they are opponent colours) but onstabilising the dividing line between them three different
percepts were observed by subjects:
1. an overall ‘red/green’ colored region they had neverexperienced before,
2. dots of red and green interspersed throughout thevisual field,
3. islands of red in a sea of green or vice versa.
It is possible to give a simple model possessing thesethree activations by means of two coupled CNFTs, onecoding for red, the other for green. The red and greenneurons at a given position are coupled inhibitorily toeach other. The resulting model is shown in Figure 11.The coupled equations for the membrane potentials u andv for the two sets of colours are
du=dt ¼ ¹ u þ wuu p v(u) ¹ wuv p v(v) þ hu þ Iu
(18a)
1217Neural Networks for Consciousness
FIGURE 10. The dragging of a bubble, due to a localised input at the point 30 at time 0-300, formed in a line of neurons when the new inputat the position 70 is switched on at the time 250. This is a proposed explanation of AM (see text).
FIGURE 11. The architecture used for modelling red/greenstabilised image percepts. See text for details.

dv=dt ¼ ¹ v þ wvv p v(v) ¹ wvu p v(u) þ hv þ Iv
(18b)
(where * denotes convolution over the cortical area,wuu and wvv are the usual Mexican hat functions andwuv, wvu are short range (excitatory) carriers ofinhibition to their opposite colour). The inputsI u and I v
are assumed to have been reduced on stabilisationof the image, so as to lead to a range of possiblesolutions to (18a) and (18b). It may be shown (Taylor,1997b) that there are at least three forms of solutionto (18):
1. `x` (in the notation of earlier in the section, where‘`’ denotes an infinitely extended bubble),
2. interdigitating spatial oscillations,3. solutions of the type {(¹ `, a1 ] ∪ [ a2, `} 3
[ a1, a2 ] (giving the support of the bubble solutionin the separate coordinates).
It is seen that these solutions correspond respectively
to the experiences of subjects listed above. The rangeof the parameter plane (I u þ hu, I v þ hv) for whichthese solutions (i)–(iii) exist is shown in Figure 12 andFigure 13.
4.2.3. Bubble Lifetimes.Having seen that bubbles canhelp explain some of the phenomena of perception (andother features of cortical processing as well, as notedin the earlier references) we now turn to the importantquestion as to how bubbles might disappear It is veryunlikely that they persist for ever, and if they did sothen they would present an ever increasing backgroundof ‘noise’ interfering with current ongoing processing.There is even some evidence for their possessing afinite lifetime from work of Lu et al. (1992). Theyexposed subjects to a sequence of sounds with the inter-stimulus interval (ISI) being gradually increased. Theydiscovered that the amplitude of the N100 response(100 ms after stimulus onset) reached saturation atsuitably long ISI in both primary and secondary auditorycortex, with the rise to saturation corresponding to adecaying trace with lifetime of about 3 s in primaryand 5 in secondary auditory cortex.
What mechanism could cause the decay of such atrace or more generally of the bubbles which theymight represent? The most likely answer is that of adap-tation of the responses of the neurons in the CNFT.Spike adaptation is a well-studied phenomenon (Connorset al., 1982; McCormick, 1986) arising from slowafter-hyperpolarizing currentsI AHP and I M, which canlast for several seconds (Schwindt et al., 1988). Theseafter-currents essentially raise the threshold for neuronalresponse. There may also be an effect from previoustraces of neural activity which shunts out later activation(May et al., 1997).
The results of Lu et al. (1992) and of Uusitalo and
1218 J. G. Taylor
FIGURE 12. The regions of the ( I u þ h u, I v þ h v )- planeassociated with the possible solutions to the model of figure11, as discussed in the text.
1.50
Po
ten
tia
l
5
input
store
probe
output
0.000
Potential
Time
FIGURE 13. The model of recency in working memory (Hastings and Taylor, 1994). The nodes are dedicated to a given input item, and theresulting activity decays as corresponding to a leaky-integrator neuron. Probe re-activation has to lead to neuron activity above a criteriallevel before reponse. The resulting ‘universal forgetting’ formula for the reaction time to a new input as having been in the previouslypresented list fits known data well.

Williamson (1996) will be interpreted here in terms ofthe decay of a lasting trace of activity in the parti-cular region from which the magnetic field was beingmeasured (auditory or primary visual cortex respec-tively). This will be shown by using a very simplemodel of the response of a neuron, that of the leakyintegrator (Hodgkin–Huxley) neuron, but with an after-hyperpolarizing current as observed in Schwindt et al.(1988). The equation for the membrane potentialu ofthis neuron will be
t du=dt ¼ ¹ u þ I (t) ¹l
∫0
exp[ ¹ (t ¹ t9)=t9] f [u(t9)]dt9
(19)
whereI(t) is the input current at timet, l is the strengthof an after-hyperpolarising current dependent on theresponsef[u(t)] integrated over the time the neuron isactive with an exponential decay of lifetimet9, and t
is the intrinsic lifetime of activity on the surface of theneuron (assumed to have no internal structure).
Under the experimental paradigm of Lu et al. (1992)the input is on for the timeT and off for the time whichwe denote by the value ISI.
During the period that the input is on,u builds up itsvalue driven by the inputI. Let us suppose that the hyper-polarising lifetimet9 is much longer than the timeT ofduration of the input. This is valid for the long-lastingpotassium-dependent hyperpolaristions mentioned bySchwindt et al. (1988), which we assume to be the oneof equation (19). At the end of the input period the mem-brane potential begins to decay exponentially, so behavesas exp[¹ t/t] (multiplied by a suitable constant, and towithin an additive constant).
We now assume that the term on the left-hand side of(19) can be neglected (in other words the time constantt
is relatively short) with the result that the time depend-ence ofu at (or close to) the beginning of the next input,say at the N100 response, has the value (to within aconstant initial value fromt ¼ T)
¹ l
∫0
exp[ ¹ (t ¹ t9)=t9] f [u(t9)]dt9 (20)
where t ¼ T þ ISI. For f chosen to be the semi-linearfunctionf(x) ¼ (x)þ (the positive part ofx) and assumingthat the membrane potential is positive through the ISI(so thatf is linear) then the membrane potential att ¼ T¼
ISI, is from (20) equal to
constant¹ lexp[ ¹ (T þ ISI)=t] (21)
For an ISI¼ 0 there will be no N100 so that (21) mustthen be equal to zero. Thus (21) becomes (for fixedT)
u(T þ ISI) ¼ A(1¹ exp[ ¹ ISI=t]) (22)
Formula (22) fits the results of Williamson and collea-gues (1992, 1996). This justifies interpreting the value ofthe time constant (as the duration of the neural traceof activity in the cortical site from which they weremeasuring. A more complete analysis can be given,
under other assumptions on the relation betweent andt9, with a similar result to that presented above.
There is the further question as to how neurons canpossess such large and variable time constants, bothacross areas and across subjects, as have been measuredby Williamson and colleagues (1992, 1996). We willnow show that these variations are to be seen as arisingfrom variable levels of recurrence, giving an increase tothe intrinsically identical time constants of each neuronby an amount dependent on the amount of recurrence;this level of recurrence can vary from one cortical region(and from one subject) to another, and is crucially depend-ant on the cell density in the short-term memory stores.
In order to consider bubble decay we will considerin more detail the effect of the long-lasting after-hyperpolarisation current used in equation (19) on bubblelifetime. The two-dimensional expression replacingequation (19) is
t]u(x, t)=]t ¼ ¹ u(x, t9) þ
∫dx9 w(x¹ x9) f [ u(x9, t) ]
þ
∫dy s(x,y) I (y, t) þ h ¹ l
∫a
0
3 exp[ ¹ (t ¹ t9)=t9] f [u(x, t9)] dt9 ð23Þ
wheret9 is a measure of the lifetime of the adaptationcurrent, andl denotes its strength. It is now necessary tocalculate the lifetime of a bubble created using equation(23). Let us first consider the one-dimensional case; thatfor two dimensions will follow straightforwardly.
A particular case of interest is when a bubble hasinitially been created by an input which is then removed.That could be due, for example, to the neural moduleacting as the source of the input having a shorter lifetimefor the persistence of bubbles than the one under con-sideration. It would also occur if the bubble is created in aprimary sensory module and the input itself has beenmodified.
To discuss this case it is appropriate to first reduceeven further to a single recurrent neuron. For that casethe membrane potential equation, from (23), is:
t]u(t)= ]t ¼ ¹ u(t) þ wv[ ut) ] þ h
¹ l
∫a
0exp[ ¹ (t ¹ t9)=t9] v[u(t9)] dt9
(24)
where a step function response has been taken for theneuron. From equation
u(t) ¼ u(0) þ [1¹ exp( ¹ t=t)][hþ w] (25)
¹ l
∫t
0exp[ ¹ (t ¹ t9)=t]dt9
∫t9
0exp[ ¹ (t9 ¹ t9)=t9]dt0
From (25), with u(0) . 0, u(t) will remain positiveinitially in time. Moreover (25) reduces to the expression
u(t) ¼ u(0) þ [1¹ exp( ¹ t=t)][hþ w¹ ltt9]
þ lt(t9)2[exp( ¹ t=t9) ¹ exp( ¹ t=t)]=(t9 ¹ t) ð26Þ
1219Neural Networks for Consciousness

where the last term on the right hand side of (26) isreplaced, fort ¼ t9, by the expressionltt9exp( ¹ t=t).The last term in (26) may be neglected ift9 , t, so that if
ltt9 . h þ w þ u(0) (27)
then for suitably larget, u(t) will become negative andthe firing of the neuron will then cease. If no newinput arrives then no further activity will ensue fromthe neuron.
The initial lifetime of the bubble is given by equatingthe right hand side of (26) to zero. Using the assumptionthat t9 q t in (26) gives the approximate value for thelife-time T as
T ¼ ¹ t91n{ 1 ¹ u(0)=t[ltt9 ¹ h ¹ w] } (28)
where the factor [ltt9 ¹ h ¹ w] is positive by (27).Equation (28) is the formula we wish to extend to thecase of a one- and then a two-dimensional CNFT.
Firstly the case of a bubble solution infinitely extendedin either dimension reduces to the above analysis withthe constantw in the single neuron case being replacedby the quantitiesw ¼
∫w(x)dx, w ¼
∫w(x)dx in the
one and two dimensional cases respectively.The relevant equation in one dimensions for a finite-
sized bubble solution (Taylor, 1997a) has the extra adap-tation term
¹ ltt9 (29)
(dropping the term of O(t9) in (26)) and the added initialvalueu(l(0), 0), wherel(t) is the size of the bubble at timet; the input term involvingShas also to be dropped. Thebubble will have a finite lifetime if the adaptation term isso negative that there exists a solution to the resultingequation for the asymptotic size of the bubble Amari,1977:
h ¹ ltt9 þ W(21(`)) ¼ 0 (30)
whereW(x) is the first integral of the connection weightw over the bubble domain. Such a solution could arise if
ltt9 ¹ h . Wm (31)
whereWm is the maximum value ofW. Thus if (31) istrue then the bubble will have a finite lifetime given,under the same approximation as for the single neuron,by
T ¼ t91n[ ¹ h(ltt9 ¹ h Wm)] (32)
This approximation should hold for both the one and twodimensional cases. In both cases we note that asWm isincreased, say by increase of cell density, the correspond-ing lifetime increases.
For the other extremet9 . . t thent andt9 must beinterchanged in the lifetime formulae (28) and (32).
In conclusion, for the caset . . t9 the bubble life-time is effectively proportional tot, so dependent onwhatever mechanism produces the bubble itself. In theopposite case the bubble lifetime is proportional tot9.
The latter quantity is expected to be an intrinsic charac-teristic of the single (pyramidal) neuron, so very likelyconstant throughout cortex. This characteristic differ-ence between the bubble lifetimes in the two cases (inthe former depending on the cortical area under con-sideration, in the latter constant throughout cortex) hasinteresting experimental consequences for corticalprocessing, which are explored in Taylor (1997a).
Finally the lifetime is seen to increase (logarithmi-cally) as the lateral connection strength (soWm in (32))increases. Such an increase is slow, according to theformula (32), but this may be only a result of the assump-tion of a hard limiting threshold output function for theneurons. A smooth sigmoid response (which can stillsupport bubble creation) can lead to a linear increase inlifetime. This may therefore explain the observedincrease of lifetime as observed in Lu et al. (1992), aswell as be the source of the buffer capability of workingmemory modules.
4.2.4. The Buffer Stores.A simple explanation of therecency effect, that more recent items in a short list areremembered better and faster, has been given by manygroups; one of these, with a useful ‘universal forgettingformula’ was in Hastings and Taylor (1994). This modelsthe short-term buffer store as a set of dedicated nodeswhich have decaying activity on them. A short list ofitems coded by these nodes has decaying activity onthe store in which the strongest activity is that for thelatest input. Recognition of the items presented thenoccurs from probe inputs which cause a temporalincrease in activity until it reaches some criterial thresh-old for response. The form of this reactivation is shownin Figure 12. It is clear from the figure that the mostrecent input reaches the criterial threshold soonest, theearliest one taking the longest time. The resulting setof reaction timesRT(n, N) for the n’th item in a list oflengthN may be shown to be given by the ‘forgetting’formula
RT(n, N) ¼ a:1n{ b þ c:exp[d(n¹ N)]} (33)
wherea, b, c, d are constants and in particulard is deter-mined by the decay constant of the nodes. A very good fitto the experimental observations leads to a value of thelifetime of the activations on the nodes of about 1.5 s,which is in the same ‘ball-park’ as the lifetimes observedby Williamson et al. noted above.
We conclude that there is support for the existence ofbubbles in cortex at the basis of phenomenal experienceand that these can help to explain the somewhat activity-independent lifetimes observed in short-term memorytests and modeled by very simple dedicated nodes withlifetimes of about 100 times the decay constant of thesingle neurons themselves. This latter feature is explic-able in terms of adaptation-driven bubble decay. Theresulting lifetime of a bubble depends on the densityof the cells in the area supporting it, with the longest
1220 J. G. Taylor

lifetime occurring for highest density. Finally we notethat these bubble may be used to help explain the natureof the ‘qualia’-like aspects of phenomenal experience, inparticular its apparently intrinsic and non-relationalcharacteristics Taylor (1997b).
5. THE THREE-STAGE MODEL
We now turn briefly to the third stage of the three-stagemodel of Figure 3. This stage receives conscious materialfrom the second stage discussed above and uses it toachieve goals by planning and reasoning. We wouldexpect that the regions of frontal cortex so involvedwould be the working memory sites of the prefrontalcortex, particularly areas 46 and 9, those observed tocause deficits in long-term planning and delayed taskswhen lost due to accident or disease. There is also aclear change in personality and social responsivenesswhen there is loss of the mesial oribtofrontal regions.Such regions would also be implicated in the construc-tion of higher order consciousness.
We have already given support, at the end of Section 3,to the suggested dissociability between the first twostages and that at the third, highest level assumed inthe three-stage model of Figure 3. There is considerablefurther material for such dissociation between the stagesfrom deficits of frontal patients, who appear to possesawareness of their deficits but not the concern thatwould be expected in their situations. Thus there is ‘adissociation between what the frontal patient knows orsays, and how he or she behaves’ (Stuss, 1991), a featureindicating dissociation between posterior knowledge andanterior action systems.
The earlier approach, in terms of the use of memorystructures, termed the ‘Relational Mind’ model in Taylor(1973, 1991) can also be used to explain how contentarises in higher consciousness. Thus there are repre-sentations of earlier experienced objects encoded inappropriate sites in frontal lobe and their re-activationsby later input gives content to that input. These inputsarise from the posterior buffer sites discussed earlier, sothat the coding of the frontal sites is guided by posterioractivity and at the same time feedback to those siteswould help their own representations. However the lateonset of prefrontal myelination would prevent much usebeing made of such frontal sites in the first few years oflife, a period when there is extensive coding of words andobjects in posterior sites. Thus the prefrontal representa-tions may only help in the development of moreadvanced and sophisticated concept representationsthan carried posteriorly.
There is now considerable experimental evidencefrom non-invasive instruments for the involvement ofthe above mentioned areas in so-called ‘working mem-ory’ tasks, those requiring the holding of activity overlong and variable periods of time for transformation andlater response. Such tasks, for example in the encoding
and later retrieval of words or pictures, have shown up anetwork of modules involving area 46 on both left andright as well as Broca’s speech area 44/45. These newmodules appear to have considerably longer lifetimesthan those of the posterior cortex (Goldman-Rakic,1992) so posing a question as to the mechanism behindsuch long activation. We will turn in the next section toconsider a possible model for such long and adaptivelifetimes.
6. A NEURAL NETWORK FOR THE THREE-STAGE MODEL.
One of the clearest differences between the frontal andposterior cortices is the existence of strong cortico-thalamic recurrence between frontal sites and themedio-dorsal thalamus. There is also known crucialinvolvement of the basal ganglia in motor and cognitiveprocessing, as defects brought about by loss of the basalganglia show in patients with Parkinson’s disease. Theproblem we are facing is that of modeling what Fuster(Fuster, 1993) calls ‘active memory’, that is persistentneural activities which can be acted upon and trans-formed into other ones so as to achieve some goal orother. It is useful to turn to consider what form represen-tations of motor actions take in motor and relatedcortices. Strong support for population vector coding inmotor and pre/supplementary motor cortices for outputresponses (coding of the direction of motor actions)has been presented in Georgopolous (1994). This alsogives a mechanism for allowing the active memoryregions of frontal cortex to be used as ‘blackboards’.Material can be written on these by input causing therecurrent lateral cortical and recurrent thalamo-corticalloops to tend rapidly to the relevant attractors of thepopulation coding; modulation by basal ganglia will actthereby to modify the thresholds and direct or changethe attractors.
A neural model of such processing has been presentedin the ACTION network of Taylor (1995) and Alavi andTaylor (1996), which has some similarity to models ofHouk and Wise (1993). It has been applied to modelingdelayed tasks and their deficits in frontal patients(Monchi and Taylor, 1995, 1997). There are also afurther range of neural models of frontal componentsof working memory (Carpenter and Grossberg, 1993;Dominey and Arbib, 1992; Dominey et al., 1995:Guigon et al., 1994; Kirillov et al., 1993; Zipser et al.,1993). We will concentrate here on the ACTION net-work as being most closely related to the neuroanatomyof the frontal cortex.
6.1. The Action Network
The ACTION net is composed of the basal ganglia actingin a disinhibitory manner on the thalamo-cortical recur-rent loops (with the presence of lateral cortico-cortical
1221Neural Networks for Consciousness

connections present as well), so as to cause an attractor tobe set up by an external input in the (possibly high)dimensional space given by the input features, asshown schematically in Figure 14. This generalises thetwo-dimensional population-vector action coding ofthe motor cortex to the other loops of the frontal system.The process of learning the correct connection weightsfor such an input set, so as to be able to write any materialfrom posterior WM and other memory sites onto thefrontal cortex, is presently under analysis (Taylor andTaylor, 1997), and has been considered in some detailin Dominey et al. (1995). These connections would allowthe active memories to have written on them materialfrom posterior buffer sites, from autobiographical/episodic memory databases related to this material andfrom goal or other memory buffered in hippocampus.
The process of ‘intermingling’ of these activitieswith ongoing activity so as to achieve higher level con-sciousness is expected to require competitive processingfor a similar reason to that discussed in connection withlow-level consciousness and for which the NRT wasemployed earlier in the paper. That such competitioncan be supported by the ACTION network was suggestedin Alavi and Taylor (1996), where the inhibitory nucleiin basal ganglia (striatum and globus pallidus) may beable to function in a similar manner to that of NRT. Thecompetition would then involve threshold-changingprocesses, as performed by the disinhibitory action ofstriatum on thalamus.
There are five possible ACTION networks, corres-ponding to the 5 great frontal loops of Alexander andcolleagues (Alexander et al., 1986) involving motor,premotor and supplementary motor cortex (actionsequences), limbic (affect), orbitofrontal (social), frontaleye fields (eye movements) and dorsolateral prefrontalcortex (cognitive/ attentional). Each of the corticalregions is expected to have suitable connections forwriting on them from posterior and limbic sites; thisseems to be the case from neuro-anatomical knowledge,as mentioned earlier, and so supporting the ‘well-connected character’ required for the modules supportingconsciousness noted in Table 1.
From the discussions so far it is clear that there-excitation of episodic memories from hippocampal
and mesio-orbital areas needs to be of a whole host ofearlier memories, which can give the consciousness‘color’ to experience. That might not best be achievedby a pattern completion or attractor network (Amit,1990), since only a single pattern would result at onetime. The most suitable memory structure is an associa-tive matrix memory for a feedforward net, in which agiven input pattern will excite as many outputs as pre-viously stored patterns with strong overlap with theinput. However the use of an attractor net may in facthelp explain the length of time taken in memory search,as various of the attractors are activated and then rejectedby comparison to some actively held template as part ofthe processing of the frontal lobes’ ACTION network.This has been discussed from an interesting neural basisin Amit (1995).
7. DISCUSSION
Having constructed some of the possible neural machin-ery to support higher consciousness, it has now to be putto work. In particular it is important to show, though inthe space available only very cursorily, how the frontalprocesses of attention, thinking, planning, prediction andschema learning and selection might be achieved. Alsothe place of emotions in such a galaxy of functionsneeds to be clarified. More general aspects of higherconsciousness, such as intentionality and introspection,are considered elsewhere (Taylor, 1997a).
A number of these processes have already been hintedat in Taylor (1995). Thus attention can be decomposedinto the component sub-processes of comparison, activa-tion/inhibition and monitoring. All of these can be per-formed by the ACTION network (Alavi and Taylor,1996), as can sequence learning and generation, at thebasis of schema learning and production. Prediction isalso involved in sequence generation, in which hippo-campal activity, as in Gray’s model (Gray, 1995;Kubota and Gabriel, 1995), should be included. Thinkinghas been discussed in Taylor (1995), using action trans-formations on representations written on prefrontalcortex so as to make such representations be closer,after comparison by the ACTION network, to a goalheld in the Papez circuit. Planning can use similar tech-niques to discover transformations taking a given state toa goal state. Note the difference between thinking andplanning; on this view the former uses whatever trans-formations that can be constructed so as to reach agoal—it is the sequence of states that are important—whilst the latter emphasises the set of transformationsand not the states.
Emotions are considered as global activations from thelimbic system when concerns (Frijda and Moffat, 1993),that is differences between desired and actual states,become large. Such comparisons (as concerns) arerelated to those of the model of Gray, but are used togive a global signal of affect to the cortical sites of
1222 J. G. Taylor
FIGURE 14. A schematic version of the ACTION network. See textfor details.

consciousness so as to cause modification of on-goingbehavior. This may be crucial in order to changeschemata (Frijda and Moffat, 1993) as well as havingan important effect in motivation and intelligence.Such an approach has been used in the neural modelsof delayed tasks and the Wisconsin card sorting task inMonchi and Taylor (1995, 1997) where there are threecoupled ACTION networks with switching betweeenthem according to valuation (concerns) by a net model-ing the amygdala.
The above models of neural networks for conscious-ness have not only been sketched very briefly but alsothere has been omission of models for the limbic system,in particular the hippocampus and related areas. Thereis presently considerable interest in such areas and theyclearly play an important role in determining the contentsof consciousness. However this is something that theinterested reader will have to look elsewhere (see, forexample, Taylor, 1996c, 1998).
REFERENCES
Alavi, F. and Taylor, J. G. (1992). A simulation of the gated thalamo-cortical model (pp 929–932) In I. Aleksander and J. G. Taylor(Eds),Artificial Neural Network 2, North Holland: Amsterdam.
Alavi, F. and Taylor, J. G. (1993). Mathematical analysis of a globalnetwork for Attention, In J.G. Taylor (Ed.),MathematicalApproaches to Neural Networks, Elsevier.
Alavi, F., & Taylor, J. G. (1995). A global competitive neural network.Biol Cybernetics, 72, 233–248.
Alavi, F. and Taylor, J. G. (1996). A basis for long range inhibitionacross cortex, In J. Sirosh, R. Miikulainen and Y. Choe (Eds.),Lateral Interactions in the Cortex: Structure and Function, Hper-text Book ISBN 0-9647060-8 (sited at http://www.cs.utexas.edu/users/nn/lateral-interaction-book/cover.html.
Alexander, G. E., DeLong, M. R., & Strick, P. L. (1986). Parallelorganisation of functionally segregated circuits linking basalganglia and cprtex.Annu Rev Neuroscience, 9, 357–381.
Amari, S.-I. (1977). Dynamical study of formation of cortical maps.Biological Cybernetics, 27, 77–87.
Amit, D. (1990).A Model of the Brain, Cambridge: Cambridge Uni-versity Press.
Amit, D. (1995). Psychology, neurobiology and modeling: the scienceof hebbian reverberations, In F. Ventriglia (Ed.), ch11NeuralModeling and Neural Networks, Oxford: Pergamon Press.
Baars, B. J., (1988).A Cognitive Theory of Consciousness, CambridgeUniversity Press.
Baars, B. J., & Newman, J. B. (1993). A neural attentional model foraccess to consciousness: a global workspace perspective.Conceptsin Neuroscience, 4, 255–290.
Baars, and B. J. and Newman, J. B. (1994). A neurobiological inter-pretation of global workspace theory, In A. Revonsuo andM. Kamppinenpp (Eds),Consciousness in Philosophy and Cogni-tive Neuroscience, (211–226) Hillsdale NJ: Lawrence Erlbaum.
Baddeley, A. D. and Hitch, G. (1974). Working memory, In G. A. Bower(Ed.), The Psychology of learning and motivation, 8, 47–89,Academic Press, New York.
Baddeley, A. D. (1986).Working Memory,Oxford: Oxford UniversityPress.
Ben-Yishai, R., Bar-Or, R.L., & Sompolinsky, H. (1995). Theory oforientation tuning in visual cortex.Proc. Nat Acad Sci (USA), 92,3844–3848.
Beurle, R. L. (1956). Properties of a mass of cells capable of regenerat-ing pulses.Trans. Roy. Soc. London B, 240,55–94.
Bisiach E. (1988) The (haunted) brain and consciousness, In A. J. Marceland E. Bisiach (eds.)Consciousness in Contemporary Science, NewYork: Oxford University Press, pp 101–120.
Cavanagh, and Mather, (1990). Motion: The long and short of it,SpatialVision, 4, 103–129.
Carpenter, G., & Grossberg, S. (1993). Normal and amnesic learning,recognition and memory by a neural model of cortico-hippocampalinteractions.Trends in neuroscience, 16, 131–140.
Cohen, J. D., Perlstein, W. M., Braver, T. S., Nystrom, L. E., Noll, D. C.,Jonides, J., & Smith, E.E. (1996). Temporal dynamics of brainactivation during a working memory task.Nature, 386,604–608.
Connors, B. W., Gutnick, M. J., & Prince, D. A. (1982). Electro-physiological properties of neocortical neurons in vitro.Journalof Neurophysiology, 48, 1302–1320.
Crane, H. D., & Piantanida, T. P. (1983). On seeing Reddish Green andYellowish Blue.Science, 221,1078–1080.
Dawson, M.R.W. (1991). The how and why of what went where inapparent motion: modeling solutions to the motion correspondenceproblem.Psychological Review, 98, 569–603.
Dominey, P., & Arbib, M.A. (1992). A cortico-subcortical model forgeneration of spatially accurate sequential saccades.CerebralCortex, 2, 153–173.
Dominey, P., Arbib, M., & Joseph, J.-P. (1995). A Model of cortico-striatal plasticity for learning oculomotor associations andsequences.J Cognitive Neuroscience, 7, 311–336.
Douglas, R. J., Koch, C., Mahowald, M., Martin, K. A. C., & Suarez,H. H. (1995). Recurrent excitation in neocortical circuits.Science,269,981–985.
Edelman, G. (1989).The Remembered Present,New York: BasicBooks.
Ermentrout, G. B. and Cowan, J. D. (1978).Studies in mathematics, TheMath Assoc of America, 15, 67–117.
Frijda, N.H. and Moffat, D. (1993). A model of emotions and emotioncommunication, InProc ROMAN ‘93: 2nd IEEE InternationalWorkshop on Robot and Human Communication, pp. 29–34.
Fuster, J.M. (1993). Frontal lobes.Current Biology, 3, 160–165.Georgopolous, A.P. (1994). New concepts in generation of movement.
Neuron, 13, 257–268.Goldman-Rakic, P., (1992).Working Memory and the Mind, Scientific
American, pp. 73–79.Gray, J. A. (1995). The contents of consciousness: a neurophysiolgical
conjecture.Behavioural and Brain Sciences, 18, 659–676.Guigon, E., Dorizzi, B., Burnod, Y. and Schultz, W. (1994). Neural
correlates of learning in the prefrontal cortex of the monkey,Cereb Cortex(in press).
Harth, E. (1995). The sketchpad model.Consciousness and Cognition,4, 346–368.
Hastings, S. and Taylor, J.G. (1994). Modeling the articulatory loop, P.Morasso and M. Marinaro (Eds),Proc Int Conf Art Neural Networks(ICANN’94), pp. 1452–1455.
He, S., Cavenagh, P., & Intrilligator, J. (1996). Attentional resolutionand the locus of visual awareness.Nature, 383,334–337.
Houk, J. C. and Wise, S. P. (1993). Outline for a theory of motorbehaviour: Involving cooperative actions of the cerebellum, basalganglia and cerebral cortex, In Rudomin, P., Arbib, M. A. andCervantes, F. (Eds.),From Neural Networks to Artificial Intelli-gence, Springer, pp. 452–470.
Kilmer, (1996). Global inhibition for selecting modes of attention.Neural Networks, 9(4), 567–573.
Kirillov, A. B., Myre, C. D., & Woodward, D. J. (1993). Bistabilityswitches and working memory in a two-neuron inhibitory-feedbackmodel.Biol Cybern., 68, 441–449.
Kolb, F.C., & Braun, J. (1995). Blindsight in normal observers.Nature,377,336–338.
Kopecz, K., & Schoner, G. (1995). Saccadic motor planning by inte-grating visual information and pre-information on neural dynamicfields.Biol. Cybern., 73, 49–60.
Kubota, Y., & Gabriel, M. (1995). Studies of the limbic comparator:
1223Neural Networks for Consciousness

limbic circuit training-Induced unit activity and avoidance beha-viour in rabbits with anterior dorsal thalamic lesions.BehavioralNeuroscience, 109,258–277.
Leopold, and Logothetis, (1996).Nature, 379, 549–553.Levelt, W. J. M., Schriefers, H., Vorberg, D., Meyer, A. S., Pechmann,
T., & Havinga, J. (1991). The time course of lexical access in speechproduction: a study of picture naming.Psychological Review, 98,122–142.
Levine, J. (1983). Materialism and qualia: the explanatory gap.PacificPhil. Quarterly, 64, 354–361.
Logothetis, and Schall, (1989).Nature, 245, 761–763.Lu, Z.-L., Williamson, S. L., & Kaufman, L. (1992). Human auditory
primary and association cortex have different lifetimes for activa-tion traces.Brain Research, 572,236–241.
Marcel, A. (1980). Conscious and preconscious recognition on poly-semous words: Locating selective effects of prior verbal contexts, InNickerson, R. S. ed.Attention and performance VIII, Hillsdale NJ:Erlbaum Assoc.
May, P., Himmelbach, M. and Taylor, J. G. (1997). (in preparation).McCormick, D. (1986). Membrane properties and neurotransmitter
actions, In: Sheperd (ed.)The Synaptic Organization of the Brain,3rd edition, Oxford: Oxford University Press.
May, P., Tiitinen, H., Winkler, I., Naatanen, R. and Taylor, J. G. (1995).A neural network model of mismatch negativity and the underlyingmemory trace, Soc for Neuroscience Abstracts, san Diego.
Metzinger, T. (1995). The Problem of Consciousness, In (ed.)T. Metzinger, Conscious Experience, (pp. 3–40) Paderborn:Schoningh.
Monchi, O. and Taylor, J. G. (1995). A model of the prefrontal loop thatincludes the basal ganglia in solving the recency task,ProcWCNN95, Washington, INNS Press.
Monchi, O. and Taylor, J. G. (1997). A model of coupled frontalworking memories for delayed response tasks and the wisconsincard sorting task, In (Ed.) P. P. Wang, (pp. 34–37) inIntJoint Conf in Information Sciences, 2nd ICCIN’97, Amsterdam:Elsevier.
Morton, J. (1969). Interaction of information in word recognition.Psychological Review, 76, 165–178.
Morton, J. (1979). Word recognition, In J. Morton and J. Marshall (eds.)Psycholinguistics: Series 2. Structures and processes, London:Elek, pp. 107–156.
Paulesu, E., Frith, C., & Frakowiak, R. S. J. (1993). The neural corre-lates of the verbal components of working memory.Nature, 362,342–345.
Petersen, R. S. and Taylor, J. G. (1996a). Reorganisation of somato-sensory cortex after tactile training, pp. 82–88 inProc Conf onNeural Information Processing NIPS ‘96, New York: Kaufmann.
Petersen, R. and Taylor, J. G. (1996b). Unmasking in somatosensorycortex,Neural Information Processing(in press).
Pritchard, R. M. (1963). Stabilized images on the retina.ScientificAmerican, 204,72–78.
Salmon, E., Van der Linden, M., Collette, F., Delfiore, G., Maquet, P.,Degueldre, C., Luxen, A., & Franck, G. (1996). Regional brainactivity during working memory tasks.Brain, 119,1617–1625.
Schwindt, P. C., Spain, W. J., Foehring, R. C., Stafstrom, C. E., Chubb,M. C., & Crill, W. E. (1988). Multiple potassium conductancesand their functions in neurons from cat sensorimotor cortex invitro. J Neurophysiol, 59, 424–449.
Skinner, J. E. and Yingling, C. D. (1977). Central gating mechanismsthat regulate even-related potentials and behaviour (pp. 30–69) InDesmedt, J.E. (ed.),Progress in clinical neurophysiology: Atten-tion, voluntary contraction and event -related potentials, Klinger, I.
Sheinberg, D. L., & Logothetis, N. K. (1997). The role of temporalcortical areas in perceptual organization.Proc Nat Acad Sci, 94,3408–3413.
Smith, E. E. and Jonides, J. (1995). Working memory in Humans:Neurophysiological Evidence, ch 66, pp. 1090–1020 inThe Cogni-tive Neurosciences.
Somers, D. C., Nelson, S. B., & Sur, M. (1995). An emergent model oforientation selectivity in cat visual cortical simple cells.J Neuroscience, 15, 5448–5465.
Steriade, M., Gloor, P., Llinas, R. R., Lopes da Silva, F. H., & Mesulam,M. M. (1990). Basic mechanisms of cerebral rythmic activities.Electroenc. and Clin. Neurophysiol, 76, 481–508.
Stuss, D. T. (1991). Disturbance of self-awareness after frontal systemdamage, in Awareness of Deficit After Brain Injury, (eds.)G. P. Prigatano and D. L. Schachter, New York: Oxford UniversityPress, pp. 63–83..
Takeuchi, A., & Amari, S.-I. (1979). Formation of topographic mapsand columnar microstructures in nerve fields.Biol Cybernetics, 35,63–72.
Taylor, J. G. (1973). A model of thinking neural networks, Seminar,Institute for Cybernetics, Univ of Tu¨binjen.
Taylor, J. G. (1991). Can neural networks ever be made to think?.Neural Network World, 1, 4–12.
Taylor, J.G. (1992). Towards a neural network model of the mind.Neural Network World, 2, 797–812.
Taylor, J. G. (1993). A global gating model of attention and conscious-ness, InNeurodynamics and Psychology, ed. Oaksford, M. andBrown, G., Academic Press, New York.
Taylor, J. G. (1994). Goals, drives and consciousness.Neural Networks,7, 1181–1190.
Taylor, J. G. (1995). Modules for the mind of PSYCHE,Proc WorldCongress on Neural Networks, Washington 1995, Hillsdale NJ: L.Erlbaum/INNS Press.
Taylor, J. G. (1996). Breakthrough to awareness.Biol Cybernetics, 75,59–72.
Taylor, J. G. (1996). A global competition for consciousness?Neurcomputing, 11, 271–296.
Taylor, J. G. (1996c). Modelling what it is like to be, InProc ArizonaConf on Scientific Basis of Consciousness, (ed.), Hammeroff, S.,Cambridge MA: MIT Press.
Taylor, J. G. (1997a). Neural ‘bubble’ dynamics in two dimensionsI: Foundations; ibid II: Applications to vision, KCL/IMEPreprints.
Taylor, J. G. (1997b). Cortical Activity and the Explanatory Gap,King’s College preprint.
Taylor, J. G. (1998).The Race for Consciousness, Camrbidge MA: MITPress.
Taylor, J. G., Jaencke, L., Shah, N. J., Noesselt, T., Schmitz, N.,Himmelbach, T. and Mueller-Gaertner, H.-W. (1997). A Three-Stage Model of Awareness: Formulation and Intial ExperimentalSupport, KFA Preprint, 1997 and Soc for Neurosci. Abstrs.
Taylor, N. and Taylor, J.G. (1997). (in preparation).Taylor, J. G. and Villa, A. E. P. (1997). The Conscious I- A Neuro-
Heuristic Approach to the Mind, to appear in The Brain of HomoSapiens, Rome: Istituto della Enciclopedia Italiana.
Tootell, R. B. H., Reppas, J. B., Dale, A. M., Look, R. B., Sereno, M. I.,Malach, R., Brady, T. J., & Rosen, B. R. (1996). Visual motionaftereffect in human cortical area MT revealed by functiona mag-netic resonance imaging.Nature, 375,139–141.
Tootell, R. B. H. (1996b). Functional MRI maps of human visualcortical activity and consciousness, abstract 104, ConsciousnessResearch Abstracts, J Consciousness Studies, 1996.
Ullman, S. (1979).The Interpretation of Visual Motion,CambridgeMass: MIT Press.
Uusitalo, M. and Williamson, S. J. (1996). Memory Lifetimes ofHuman Visual Cortex, Human Brain Mapping Conference AbstractS565.
Vanni, S., Revonsuo, A., Saarinen, J., & Hari, R. (1996). Visual aware-ness of objects correlates with activity of right occipital cortex.NeuroReport, 8, 183–186.
Villa, A. (1988). Influence de L’Ecorce Cerebrale sur L’ActiviteSpontanee et Evoque du Thalamus auditif du Chat, These, Univde Lausanne.
Weiskrantz, (1986).Blindsight, Oxford: Clarendon Press.
1224 J. G. Taylor

Weiskrantz, L., Barbur, J. L., & Sahrie, A. (1995). Parameters affectingconscious versus unconscious visual discrimination withdamage to the visual cortex (V1).Proc Nat Acad Sci (USA), 92,6122–6126.
Zhang, K. (1996). Representation of spatial orientation by the intrinsic
dynamics of the head—Direction cell ensemble: a theory.J.Neuroscience, 16, 2112–2126.
Zipser, D., Kehoe, B., Littlewort, G., & Fuster, J. (1993). A spikingnetwork model of short-term active memory.J. Neurosci., 13,3406–3420
1225Neural Networks for Consciousness

1997 SPECIAL ISSUE
Consciousness in Neural Networks?
Edmund T. Rolls
Department of Experimental Psychology, University of Oxford
(Received31 August1996;accepted15 January1997)
Abstract—A combined neurophysiological and computational approach is reviewed that leads to a proposal for howneural networks in the temporal cortical visual areas of primates could function to produce invariant object representa-tion and identification. A similar approach is then reviewed which leads to a theory of how the hippocampus couldrapidly store memories, especially episodic memories including spatial context, and how later recall of the informationto the neocortex could occur. Third, it is argued that the visual and memory mechanisms described could operate withoutconsciousness, and that a different type of processing is related to consciousness. It is suggested that the type ofprocessing related to consciousness involves higher-order thoughts ("thoughts about thoughts"), and evolved to allowplans, formulated in a language, with many steps, to be corrected. It is suggested that it would feel like something to be asystem that can think linguistically (using syntax) about its own thoughts, and that the subjective or phenomenal aspectsof consciousness arise in this way. It is further suggested that "raw sensory feels" arise in evolution because once sometypes of processing feel like something by virtue of a system capable of higher-order thoughts, it is then parsimonious topostulate that sensory and related processing, which has to be taken into account in that processing system, should feellike something. It is suggested that it is this type of processing, which must be implemented in neural networks, which isrelated to consciousness.q 1997 Elsevier Science Ltd.
Keywords—Consciousness, Hippocampus, Memory, Invariance, Visual recognition, Higher-order thoughts, Visualcortex.
1. INTRODUCTION
Advances are being made in understandinghowthe braincould perform some of the processing involved in per-ception and memory. These advances come in part fromneurophysiological experiments in which the processinginvolved in vision and memory is analysed by recordingthe activity of single neurons in primates during thesetypes of processing, and incorporating this informationinto computational models at the neuronal network levelwhich provide an account of the ways in which manyneurons in the networks found in different brain regions
could perform the required computations. Examples ofthis approach are described first in this paper.
Having considered brain mechanisms involved invisual object recognition and memory, I then considerwhether, once this processing is fully understood, wewill have produced an account of the brain mechanismsunderlying consciousness. I argue that we will not, andthat it is a different type of information processing that isinvolved in consciousness. I outline a theory of what theprocessing is that is involved in consciousness, of itsadaptive value in an evolutionary perspective, and ofhow processing in our visual and other sensory systemscan result in subjective or phenomenal states, the "rawfeels" of conscious awareness. These processes involvedin consciousness must themselves be implemented inneural networks, but before considering how these pro-cesses are implemented, it is useful to be clear aboutwhat processing must be implemented.
2. NEURONAL NETWORKS INVOLVED ININVARIANT VISUAL OBJECT RECOGNITION
2.1. Neurophysiology
The visual pathways project in primates by a number of
Neural Networks, Vol. 10, No. 7, pp. 1227–1240, 1997q 1997 Elsevier Science Ltd. All rights reserved
Printed in Great Britain0893–6080/97 $17.00+.00
PII: S0893-6080(97)00049-X
Pergamon
1227
Acknowledgements: The author has worked on some of theexperiments described here with G. C. Baylis, M. Booth, M. J.Burton, P. Georges-Franc¸ois, M. E. Hasselmo, C. M. Leonard, F.Mora, D. I. Perrett, R. G. Robertson, M. K. Sanghera, T. R. Scott, S.J. Thorpe, and F. A. W. Wilson, and their collaboration, and helpfuldiscussions with or communications from M. Davies and C. C. W.Taylor (Corpus Christi College, Oxford), and M. Stamp Dawkins, aresincerely acknowledged. Some of the research described was supportedby the Medical Research Council (PG8513579), and by The HumanFrontier Science Program.
Requests for reprints should be sent to Professor E. T. Rolls, Uni-versity of Oxford, Department of Experimental Psychology, SouthParks Road, Oxford OX1 3UD, UK; Tel.: +44-1865-271348; Fax:+44-1865-310447; e-mail: [email protected]

cortico-cortical stages from the primary visual cortexuntil they reach the temporal lobe visual cortical areas(see Figure 1, and for details of the neurophysiologysummarized next, see Rolls, 1991, 1992, 1994b, 1995b,1997). Along these pathways the receptive fields of neu-rons gradually become larger, as shown in Figure 1. (Thereceptive field of a neuron is the part of visual spacewithin which appropriate visual stimuli can activate theneuron.) Part of the basis for this is the convergence ontoneurons at any one stage of processing from a limitedarea of the preceding cortical area (see Figure 1). For thisto result in neurons at the final stages of visual processingresponding to the same object or stimulus independentlyof position on the retina, the appropriate connectionsmust be set up in the hierarchy. Ways in which the appro-priate synaptic weights to achieve this translation invar-iance could be learned are considered below.
The encoding that is provided of objects and faces atthe end of this processing in the inferior temporal visualcortex is distributed, in the sense that the representationis not local or "grandmother cell" like, but instead manyneurons are active to encode any one object (Rolls &Tovee, 1995; Rolls et al., 1996). Using an information-theoretic approach, it has been shown that the informa-tion available from the firing rates of a population ofneurons about which visual stimulus (which of 20 equi-probable faces) has been shown on a single 500 ms pre-sentation increases linearly with the number of neuronsin the sample (Abbott et al., 1996; Rolls et al., 1997b).Because information is a logarithmic measure, this indi-cates that the number of stimuli encoded rises approxi-mately exponentially, as the number of neurons in thesample increases. The consequence of this is that largenumbers of stimuli, and fine discriminations betweenthem, can be represented without (a receiving neuron)
having to measure the activity of an enormous numberof neurons. For example, the results of the experiments ofRolls et al. (1997b) indicate that the activity of 15 neu-rons would be able to encode 192 face stimuli (at 50%accuracy), of 20 neurons 768 stimuli, and of 25 neurons3072 stimuli (Abbott et al., 1996; the values are for anoptimal decoding case). This is strong evidence for dis-tributed encoding. This type of encoding makes brainconnectivity possible, in the sense that a receivingneuron can gain a great deal of information even whenit does not receive vast numbers of inputs. Another inter-esting aspect of this encoding is that the information justdescribed is available from the firing rates of the neurons,without taking into account the relative time at which theneurons fire. Thus temporal encoding is not an essentialpart of the code at this stage at least of visual informationprocessing (see further Rolls et al., 1997b; Tovee &Rolls, 1995; Tovee et al., 1993). Another interestingaspect of the encoding is that much of the informationfrom a population of neurons is available when thedecoding is a simple neuronally plausible decodinginvolving a dot product of the neuronal activity in thecurrent 500 ms (or 100 ms or 50 ms) presentation withthat which occurred previously in the population of neu-rons to a particular stimulus (Rolls et al., 1997b). Suchdecoding could be performed by neurons which calculatetheir activation by a weighted sum of their input activity,which is common in neural network modelling. The factthat the information is available in a form in which it canbe read out by this simple neuronally plausible dot pro-duct decoding with sampling from a limited set of neu-rons, and at the same time having the properties of aconstant sparseness of the representation, and providingfor generalization and graceful degradation, is probablywhat accounts for the fact that neurophysiologicallyinterpretable information is available in the responsesof single neurons about which stimulus has been seen(Rolls et al., 1997a; Tovee & Rolls, 1995; Tovee et al.,1993)1. This is one of the factors that allows singleneuron recording to be so useful in understanding brainfunction—a correlation can frequently be found betweenthe activity of even a single neuron and a subset of thestimuli being shown, of the motor responses being made,etc.
Some neurons in the temporal cortical visual areashave responses which are invariant not only for positionon the retina, but also for the size, contrast, spatial fre-quency, position on the retina, and even angle of view
1228 E. T. Rolls
FIGURE 1. Schematic diagram showing convergence achievedby the forward projections in the visual system, and the types ofrepresentation that may be built by competitive networks oper-ating at each stage of the system from the primary visual cortex(V1) to the inferior temporal visual cortex (area TE) (see text).LGN, Lateral geniculate nucleus. Area TEO forms the posteriorinferior temporal cortex. The receptive fields in the inferior tem-poral visual cortex (e.g. in the TE areas) cross the vertical midline(not shown). (Reprinted from Wallis & Rolls, 1997.)
1 The fact that the information increases approximately linearly withthe number of neurons in the sample implies that the neurons conveyalmost independent information (if the stimulus set size is sufficientlylarge). If local encoding were used, the information would increase inproportion to the logarithm of the number of cells. If, for example,binary encoding were used (as, for example, numbers are encoded ina computer word), then the sparseness of the representation wouldfluctuate wildly, any receiving neuron would need to receive from allthe input neurons, and generalization and graceful degradation wouldnot occur.

(see Rolls, 1992, 1994b, 1995b, 1997; Rolls et al., 1996).It is clearly important that invariance in the visual systemis made explicit in the neuronal responses, for this sim-plifies greatly the output of the visual system to memorysystems such as the hippocampus and amygdala, whichcan then remember or form associations aboutobjects.The function of these memory systems would be almostimpossible if there were no consistent output from thevisual system about objects (including faces), for then thememory systems would need to learn about all possiblesizes, positions, etc., of each object, and there would beno easy generalization from one size or position of anobject to that object when seen with another retinal sizeor position.
Other aspects of the neurophysiological findingswhich provide constraints on and guide the developmentof neural network theories about how the visual corticalareas involved in visual object recognition operate is thatlearning of new faces or objects can occur rapidly, withina few seconds; that the processing within any one corticalarea is fast, with sufficient processing being completedwithin 30 ms in each cortical area in the hierarchy tosubserve recognition; and that neurons in intermediatestages of processing (e.g. V2 and V4) respond to combi-nations of features present at earlier stages of processing(see Figure 1 and Rolls, 1992, 1994b, 1995b, 1997).
2.2. Computational Processes Involved in InvariantVisual Object Recognition
Cortical visual processing for object recognition is con-sidered to be organized as a set of hierarchically con-nected cortical regions consisting at least of V1, V2,V4, posterior inferior temporal cortex (TEO), inferiortemporal cortex (e.g. TE3, TEa and TEm), and anteriortemporal cortical areas (e.g. TE2 and TE1). (This streamof processing has many connections with a set of corticalareas in the anterior part of the superior temporal sulcus,including area TPO.) There is convergence from eachsmall part of a region to the succeeding region (orlayer in the hierarchy) in such a way that the receptivefield sizes of neurons (e.g. 18 near the fovea in V1)become larger by a factor of approximately 2.5 witheach succeeding stage (and the typical parafoveal recep-tive field sizes found would not be inconsistent with thecalculated approximations of, e.g. 88 in V4, 208 in TEO,and 508 in inferior temporal cortex; Boussaoud et al.,1991) (see Figure 1). Such zones of convergence wouldoverlap continuously with each other (see Figure 1). Thisconnectivity would be part of the architecture by whichtranslation invariant representations are computed (seeRolls, 1992, 1994b, 1995b, 1996a; Wallis & Rolls,1997). Each layer is considered to act partly as a set oflocal self-organizing competitive neuronal networks withoverlapping inputs. (The region within which competi-tion would be implemented would depend on the spatialproperties of inhibitory interneurons, and might operate
over distances of 1–2 mm in the cortex.) These compe-titive nets operate by a single set of forward inputs lead-ing to (typically non-linear, e.g. sigmoid) activation ofoutput neurons; of competition between the outputneurons mediated by a set of feedback inhibitory inter-neurons which receive from many of the principal (in thecortex, pyramidal) cells in the net and project back tomany of the principal cells, which serves to decrease thefiring rates of the less active neurons relative to the ratesof the more active neurons (i.e. soft competition); andthen of synaptic modification by a modified Hebb rule,such that synapses to strongly activated output neuronsfrom active input axons strengthen, and from inactiveinput axons weaken (see Rolls, 1989c; Rolls & Treves,1997). (A biologically plausible form of this learning rulethat operates well in such networks is
dwij ¼ k·mi(rj9 ¹ wij )
where k is a constant,dwij is the change of synapticweight, rj9 is the firing rate of thejth axon, andmi is anon-linear function of the output activation of neuroniwhich mimics the operation of the NMDA receptors inlearning; see Rolls, 1989a, b, c; Rolls & Treves, 1997).Related approaches to self-organization in the visualsystem are described by Linsker (1986, 1988) andMacKay & Miller (1990).
Translation invariance would be computed in such asystem by utilizing competitive learning to detect regu-larities in inputs when real objects are translated in thephysical world. The hypothesis is that because objectshave continuous properties in space and time in theworld, an object at one place on the retina might activatefeature analysers at the next stage of cortical processing,and when the object was translated to a nearby position,because this would occur in a short period (e.g. 0.5 s), themembrane of the postsynaptic neuron would still be in its"Hebb-modifiable" state, and the presynaptic afferentsactivated with the object in its new position would thusbecome strengthened on the still-activated postsynapticneuron. It is suggested (Rolls, 1992) that the short tem-poral window (e.g. 0.5 s) of Hebb-modifiability helpsneurons to learn the statistics of objects moving in thephysical world, and at the same time to form differentrepresentations of different feature combinations orobjects, as these are physically discontinuous and presentless regular correlations to the visual system. Foldiak(1991) has proposed computing an average activationof the postsynaptic neuron to assist with the same pro-blem. Another suggestion is that a memory trace for whathas been seen in the last 300 ms appears to be implemen-ted by a mechanism as simple as continued firing ofinferior temporal neurons after the stimulus has disap-peared, as we have shown in masking experiments (seeRolls & Tovee, 1994; Rolls et al., 1994b). This continuedfiring could be implemented by local attractor networksin columns or modules in the cerebral cortex imple-mented by the local recurrent collaterals of the cortical
1229Consciousness in Neural Networks?

pyramidal cells (Rolls & Treves, 1997). Other invar-iances, for example, size, spatial frequency, and rotationinvariance, could be learned by a comparable process. Itis suggested that this process takes place at each stage ofthe multiple-layer cortical processing hierarchy, so thatinvariances are learned first over small regions of space,and then over successively larger regions. This limits thesize of the connection space within which correlationsmust be sought. It is suggested that view-independentrepresentations could be formed by the same type ofcomputation, operating to combine a limited set ofviews of objects. Increasing complexity of represen-tations could also be built in such a multiple layerhierarchy by similar mechanisms. At each stage orlayer the self-organizing competitive nets would resultin combinations of inputs becoming the effective stimulifor neurons.
To test and clarify these hypotheses (see further Rolls,1992, 1994b, 1995b, 1997) about how the visual systemmay operate to learn invariant object recognition, wehave performed a simulation which implements manyof the ideas just described, and is consistent with andbased on much of the neurophysiology summarizedabove. The network simulated can perform object,including face, recognition in a biologically plausibleway, and after training shows, for example, translationand view invariance (Wallis & Rolls, 1997; Wallis et al.,1993).
In the four-layer network, the successive layers corre-spond approximately to V2, V4, the posterior temporalcortex, and the anterior temporal cortex. The forwardconnections to a cell in one layer are derived from atopologically corresponding region of the precedinglayer, using a Gaussian distribution of connection prob-abilities to determine the exact neurons in the precedinglayer to which connections are made. This schema isconstrained to preclude the repeated connection of anycells. Each cell receives 100 connections from the 32332 cells of the preceding layer, with a 67% probability
that a connection comes from within four cells of thedistribution centre. Figure 2 shows the general conver-gent network architecture used, and may be comparedwith Figure 1. Within each layer, lateral inhibitionbetween neurons has a radius of effect just greater thanthe radius of feedforward convergence just defined. Thelateral inhibition is simulated via a linear local contrastenhancing filter active on each neuron. (Note that thisdiffers from the global ‘winner-take-all’ paradigm imple-mented by Foldiak, 1991). The cell activation is thenpassed through a non-linear activation function (e.g. sig-moid), which also produces contrast enhancement of thefiring rates.
So that the results of the simulation might be madeparticularly relevant to understanding processing inhigher cortical visual areas, the inputs to layer 1 comefrom a separate input layer which provides an approxi-mation to the encoding found in cortical visual area 1(V1) of the primate visual system. These responsecharacteristics of neurons in the input layer are providedby a series of spatially tuned filters with image contrastsensitivities chosen to accord with the general tuningprofiles observed in the simple cells of V1.
The synaptic learning rule used in these simulations(VisNet) can be summarized as follows:
dwij ¼ kmirj9
and
mti ¼ (1¹ h)r (t)
i þ hm(t ¹ 1)i
whererj9 is thejth input to the neuron,ri is the output ofthe ith neuron,wij is the jth weight on theith neuron,hgoverns the relative influence of the trace and the newinput (typically 0.4–0.6), andm(t)
i represents the value ofthe ith cell’s memory trace at timet. In the simulationsthe neuronal learning was bounded by normalization ofeach cell’s dendritic weight vector.
To train the network to produce a translation invariantrepresentation, one stimulus was placed successively in asequence of nine positions across the input, then the nextstimulus was placed successively in the same sequenceof nine positions across the input, and so on through theset of stimuli. The idea was to enable the network to learnwhatever was common at each stage of the network abouta stimulus shown in different positions. To train on viewinvariance, different views of the same object wereshown in succession, then different views of the nextobject were shown in succession, and so on. It has beenshown that this network, inspired by Fukushima’s(Fukushima, 1980) neocognitron as well as by the neu-rophysiological data, can form cells in its final layer withtranslation, size and view invariant responses to stimulipresented on the ‘retina’ (Wallis & Rolls, 1997; Wallis etal., 1993).
These results show that the proposed learning mechan-ism and neural architecture can produce cells withresponses selective for stimulus type with considerable
1230 E. T. Rolls
FIGURE 2. Hierarchical network structure used in the model ofinvariant visual object recognition. (Reprinted from Wallis &Rolls, 1997.)

position, size or view invariance. The ability of the net-work to be trained with natural scenes is currently help-ing to advance our understanding of how representationsof objects are built and encoded in the primate visualsystem.
This combined neurophysiological and computationalapproach is thus leading to biologically plausible theoriesabout how the brain operates when it performs face orobject recognition. In addition, there is now considerableevidence about what happens in our higher cortical visualareas when we recognize faces, and about how informa-tion about at least some classes of object in the world isrepresented in the visual system. Yet does this under-standing of visual object recognition help us directlywith the problem of consciousness, of why it is that itfeels the way it does when we recognize a face? Would acomputer which operated in the way described above beconscious during object recognition? I suggest that itwould not be, and that for the object recognition pro-cesses to be conscious, including to feel like anything,
the information from the type of visual processingsystem I describe would have to be projected to a differ-ent brain system, the nature of which will be describedbelow. Before turning to that, some recent advances inunderstanding the brain processing that occurs when westore and then recall later everyday events are described,and I ask whether these memory processes are closer toconsciousness.
3. THE HIPPOCAMPUS AND MEMORY
The hippocampus is implicated in a particular type ofmemory, the memory for recent events and episodes, inwhich there is frequently a spatial aspect or context (seefor details Rolls, 1996b, d, 1997). In monkeys, a proto-typical memory task impaired by damage to the hippo-campal system is object-place memory, in which thelocations of objects in space must be remembered (seeGaffan, 1994). This impairment is analogous to thatshown by anterograde amnesic patients with damage to
1231Consciousness in Neural Networks?
FIGURE 3. Forward connections (continuous lines) from areas of cerebral association neocortex via the parahippocampal gyrus andperirhinal cortex, and entorhinal cortex, to the hippocampus; and backprojections (dashed lines) via the hippocampal CA1 pyramidalcells, subiculum, and parahippocampal gyrus to the neocortex. There is great convergence in the forward connections down to thesingle network implemented in the CA3 pyramidal cells, and great divergence again in the backprojections. Left: block diagram. Right:more detailed representation of some of the principal excitatory neurons in the pathways. D, Deep pyramidal cells; DG, dentate granulecells; F, forward inputs to areas of the association cortex from preceding cortical areas in the hierarchy; mf, mossy fibres; PHG,parahippocampal gyrus and perirhinal cortex; pp, perforant path; rc, recurrent collateral of the CA3 hippocampal pyramidal cells; S,superficial pyramidal cells; 2, pyramidal cells in layer 2 of the entorhinal cortex; 3, pyramidal cells in layer 3 of the entorhinal cortex. Thethick lines above the cell bodies represent the dendrites.

the hippocampus and nearby areas who cannot rememberthe locations of objects on a tray.
On the basis of these findings in humans and otheranimals, the hypothesis is suggested that the importanceof the hippocampus in spatial and other memories is thatit can rapidly form event or "episodic" representations ofinformation originating from many areas of the cerebralcortex. In rats, hippocampal pyramidal cells (e.g. CA3and CA1 neurons) respond when the rat is in a particularplace in a spatial environment. In monkeys, it has beenshown that there is a rich representation of space outsidethe monkey implemented by "spatial view" cells (seeRolls, 1996b, 1996d). These would provide an excellentrepresentation of the spatial information needed to form amemory of where an object had been seen in space. It issuggested that an autoassociation network implementedby the CA3 cells of the hippocampus brings together theobject information represented in temporal corticalvisual areas, and spatial information represented in par-ietal areas, so that associations can be formed betweenobjects and places (see Figure 3 and Rolls, 1989a, b, c,1990a, 1996a,b).
A theory of how the networks shown in Figure 3 couldoperate, not only to store memories of events, but also torecall them to the neocortex via the backprojection path-ways, has been developed (see Rolls, 1989a, b, 1996a;Rolls & Treves, 1997; Treves & Rolls, 1992, 1994). Away in which such recall could be useful in the cortex forbuilding long-term semantic memories has beendescribed by McClelland et al. (1995). A comparisonof these approaches with others (for example by Burgesset al., 1994; and Hasselmo & Bower, 1993) is providedby Rolls (1996a), Rolls & Treves (1997) and Treves &Rolls (1994).
4. CONSCIOUSNESS
It would be possible to build a computer which wouldperform all the above functions of visual object recogni-tion, memory storage and recall to the neocortex, andeven emotion (Rolls, 1990b, 1995c), using the samecomputational principles described above, and yet wemight not want to ascribe subjective or phenomenalstates, which I shall call qualia, to this computer. Wemight not want to say that it feels like something to thecomputer when the computer is performing these func-tions. This raises the issue of in which networks in thebrain would consciousness be represented. Because thetopic of subjective or phenomenal feels or feelings (thatit feels like something to be in that state) is of con-siderable current interest, and is for the present purposesthe defining aspect of consciousness, one view on con-sciousness, influenced by contemporary cognitiveneuroscience, is outlined next. However, this view isonly preliminary, and theories of consciousness arelikely to develop considerably. A reason for describingthis view of consciousness is that we need to be clear
aboutwhatmust be implemented before consideringhowit could be implemented in neural networks.
A starting point is that many actions can be performedrelatively automatically, without apparent consciousintervention. An example sometimes given is driving acar. Such actions could involve control of behaviour bybrain systems which are old in evolutionary terms such asthe basal ganglia. It is of interest that the basal ganglia(and cerebellum) do not have backprojection systems tomost of the parts of the cerebral cortex from which theyreceive inputs (see, e.g. Rolls, 1994a; Rolls & Johnstone,1992). In contrast, parts of the brain such as the hippo-campus and amygdala, involved in functions such asepisodic memory and emotion respectively, aboutwhich we can make (verbal) declarations (hence declara-tive memory, Squire, 1992) do have major backprojec-tion systems to the high parts of the cerebral cortex fromwhich they receive forward projections (Rolls, 1992;Rolls & Treves, 1997; Treves & Rolls, 1994; seeFigure 3). It may be that evolutionarily newer parts ofthe brain, such as the language areas and parts of theprefrontal cortex, are involved in an alternative type ofcontrol of behaviour, in which actions can be plannedwith the use of a (language) system which allowsrelatively arbitrary (syntactic) manipulation of semanticentities (symbols).
The general view that there are many routes tobehavioural output is supported by the evidence thatthere are many input systems to the basal ganglia (fromalmost all areas of the cerebral cortex), and that neuronalactivity in each part of the striatum reflects the activity inthe overlying cortical area (Rolls, 1994a; Rolls & John-stone, 1992). The evidence is consistent with thepossibility that different cortical areas, each specializedfor a different type of computation, have their outputsdirected to the basal ganglia, which then select the stron-gest input, and map this into action (via outputs directed,for example, to the premotor cortex) (Rolls & Johnstone,1992; Rolls & Treves, 1997). Within this scheme, thelanguage areas would offer one of many routes toaction, but a route particularly suited to planning actions,because of the syntactic manipulation of semantic enti-ties which may make long-term planning possible. Aschematic diagram of this suggestion is provided inFigure 4. Consistent with the hypothesis of multipleroutes to action, only some of which utilize language,is the evidence that split-brain patients may not beaware of actions being performed by the "non-domi-nant" hemisphere (Gazzaniga, 1988, 1995; Gazzaniga& LeDoux, 1978). Also consistent with multiple includ-ing non-verbal routes to action, patients with focal braindamage, for example to the prefrontal cortex, may emitactions, yet comment verbally that they should not beperforming those actions (Rolls et al., 1994a). In boththese types of patient, confabulation may occur, inthat a verbal account of why the action was performedmay be given, and this may not be related at all to the
1232 E. T. Rolls

environmental event which actually triggered the action(Gazzaniga, 1988, 1995; Gazzaniga & LeDoux, 1978). Itis possible that sometimes in normal humans whenactions are initiated as a result of processing in a specia-lized brain region such as those involved in some types ofrewarded behaviour, the language system may subse-quently elaborate a coherent account of why that actionwas performed (i.e. confabulate). This would be consis-tent with a general view of brain evolution in which asareas of the cortex evolve, they are laid on top of existingcircuitry connecting inputs to outputs, and in which eachlevel in this hierarchy of separate input–output pathwaysmay control behaviour according to the specialized func-tion it can perform (see schematic diagram in Figure 4).(It is of interest that mathematicians may have a hunchthat something is correct, yet not be able to verbalizewhy. They may then resort to formal, more serial andlanguage-like theorems to prove the case, and these seemto require conscious processing. This is a further indica-tion of a close association between linguistic processingand consciousness. The linguistic processing need not, asin reading, involve an inner articulatory loop.)
We may next examine some of the advantages andbehavioural functions that language, present as themost recently added layer to the above system, wouldconfer. One major advantage would be the ability toplan actions through many potential stages and to eval-uate the consequences of those actions without having toperform the actions. For this, the ability to form proposi-tional statements, and to perform syntactic operations onthe semantic representations of states in the world, wouldbe important. Also important in this system would be theability to have second-order thoughts about the type of
thought that I have just described (e.g. I think that hethinks that…), as this would allow much better modellingand prediction of others’ behaviour, and therefore ofplanning, particularly planning when it involves others.This capability for higher-order thoughts would alsoallow reflection on past events, which would also beuseful in planning. In contrast, non-linguistic behaviourwould be driven by learned reinforcement associations,learned rules, etc., but not by flexible planning for manysteps ahead involving a model of the world includingothers’ behaviour. (For an earlier view which is closeto this part of the argument, see Humphrey, 1980.)(The examples of behaviour from non-humans thatmay reflect planning may reflect much more limitedand inflexible planning. For example, the dance of thehoney-bee to signal to other bees the location of foodmay be said to reflect planning, but the symbol manip-ulation is not arbitrary. There are likely to be interestingexamples of non-human primate behaviour, perhaps inthe great apes, that reflect the evolution of an arbitrarysymbol-manipulation system that could be useful forflexible planning; see Cheney & Seyfarth, 1990.) It isimportant to state that the language ability referred tohere is not necessarily human verbal language (thoughthis would be an example). What it is suggested is impor-tant to planning is the syntactic manipulation of symbols,and it is this syntactic manipulation of symbols which isthe sense in which language is defined and used here.
It is next suggested that this arbitrary symbol-manip-ulation using important aspects of language processingand used for planning but not in initiating all types ofbehaviour is close to what consciousness is about. Inparticular, consciousness maybe the state which arisesin a system that can think about (or reflect on) its own (orother peoples’) thoughts, that is, in a system capable ofsecond- or higher-order thoughts (Rosenthal, 1986, 1990,1993; compare Dennett, 1991). On this account, a mentalstate is non-introspectively (i.e. non-reflectively) con-scious if one has a roughly simultaneous thought thatone is in that mental state. Following from this, intro-spective consciousness (or reflexive consciousness, orself consciousness) is the attentive, deliberately focusedconsciousness of one’s mental states. It is noted that notall of the higher-order thoughts need themselves be con-scious (many mental states are not). However, accordingto the analysis, having a higher-order thought about alower-order thought is necessary for the lower-orderthought to be conscious. (A slightly weaker positionthan Rosenthal’s on this is that a conscious state corre-sponds to a first-order thought that has thecapacity tocause a second-order thought or judgement about it—Carruthers, 1996). This analysis is consistent with thepoints made above that the brain systems that arerequired for consciousness and language are similar. Inparticular, a system that can have second- or higher-orderthoughts about its own operation, including its planningand linguistic operation, must itself be a language
1233Consciousness in Neural Networks?
FIGURE 4. Schematic illustration indicating many possibleroutes from input systems to action (output) systems. Corticalinformation processing systems are organized hierarchically,and there are routes to output systems from most levels of thehierarchy.

processor, in that it must be able to bind correctly to thesymbols and syntax in the first-order system. Accordingto this explanation, the feeling of anything is the statewhich is present when linguistic processing that involvessecond- or higher-order thoughts is being performed.
It might be objected that this captures some of theprocess aspects of consciousness, what it is good for inan information processing system, but does not capturethe phenomenal aspect of consciousness. I agree thatthere is an element of "mystery" that is invoked at thisstep of the argument, when I say that it feels like some-thing for a machine with higher-order thoughts to bethinking about its own first- or lower-order thoughts.But the return point is the following:if a human withsecond-order thoughts is thinking about his or her first-order thoughts, surely it is very difficult for us to conceivethat this would not feel like something? This isespecially the case when the first-order thoughts are lin-guistic, and are about (grounded in) the real world.
It is suggested that part of the evolutionary adaptivesignificance of this type of higher-order thought is that itallows correction of errors made in first-order linguisticor in non-linguistic processing. Indeed, the ability toreflect on previous events is extremely important forlearning from them, including setting up new long-termsemantic structures. It was shown above that the hippo-campus may be a system for such "declarative" recall ofrecent memories. Its close relation to "conscious" proces-sing in humans (Squire has classified it as a declarativememory system) may be simply that it allows the recallof recent memories, which can then be reflected upon inconscious, higher-order, processing. Another part of theadaptive value of a higher-order thought system may bethat by thinking about its own thoughts in a given situa-tion, it may be able to better understand the thoughts ofanother individual in a similar situation, and thereforepredict that individual’s behaviour better (Humphrey,1980).
As a point of clarification, I note that according to thistheory, a language processing system is notsufficientforconsciousness. What defines a conscious system accord-ing to this analysis is the ability to have higher-orderthoughts, and a first-order language processor (thatmight be perfectly competent at language) would notbe conscious, in that it could not think about its own orothers’ thoughts. One can perfectly well conceive of asystem which obeyed the rules of language (which is theaim of much connectionist modelling), and implementeda first-order linguistic system, that would not be con-scious. (Possible examples of language processing thatmight be performed non-consciously include computerprograms implementing aspects of language, or ritua-lized human conversations, e.g. about the weather.These might require syntax and correctly groundedsemantics, and yet be performed non-consciously. Amore complex example, illustrating that syntax couldbe used, might be: "If A does X, then B will probably
do Y, and then C would be able to do Z." A first-orderlanguage system could process this statement. Moreover,the first-order language system could apply the rule use-fully in the world, provided that the symbols in the lan-guage system (A, B, X, Y, etc.) are grounded (havemeaning) in the world.) In line with the argument onthe adaptive value of higher-order thoughts and thus con-sciousness given above, that they are useful for correct-ing lower-order thoughts, I now suggest that correctionusing higher-order thoughts of lower-order thoughtswould have adaptive value primarily if the lower-orderthoughts are sufficiently complex to benefit from correc-tion in this way. The nature of the complexity is specific:that it should involve syntactic manipulation of symbols,probably with several steps in the chain, and that thechain of steps should be a one-off set of steps, as in aparticular plan or sentence, rather than a set of well-learned rules. The first- or lower-order thoughts mightinvolve a linked chain of "if…then" statements thatwould be involved in planning, an example of whichhas been given above. It is partly because complexlower-order thoughts such as these, which involve syntaxand language, would benefit from correction by higher-order thoughts, that I suggest that there is a close linkbetween this reflective consciousness and language. Thehypothesis is that by thinking about lower-orderthoughts, the higher-order thoughts can discover whatmay be weak links in the chain of reasoning at thelower-order level, and having detected the weak link,might alter the plan, to see if this gives better success.In our example above, if it transpired that C could not doZ, how might the plan have failed? Instead of having togo through endless random changes to the plan to see ifby trial and error some combination does happen to pro-duce results, what I am suggesting is that by thinkingabout the previous plan, one might, for example, usingknowledge of the situation and the probabilities thatoperate in it, guess that the step where the plan failedwas that B did not in fact do Y. So by thinking about theplan (the first- or lower-order thought), one might correctthe original plan, in such a way that the weak link in thatchain, that "B will probably do Y", is circumvented. Todraw a parallel with neural networks: there is a "creditassignment" problem in such multistep syntactic plans, inthat if the whole plan fails, how does the system assigncredit or blame to particular steps of the plan? The sug-gestion is that this is the function of higher-orderthoughts and is why systems with higher-order thoughtsevolved. The suggestion I then make is that if a systemwere doing this type of processing (thinking about itsown thoughts), it would then be very plausible that itshould feel like something to be doing this. I even sug-gest to the reader that it is not plausible to suggest that itwould not feel like anything to a system if it were doingthis.
Two other points in the argument should be empha-sized for clarity. One is that the system that is having
1234 E. T. Rolls

syntactic thoughts about its own syntactic thoughtswould have to have its symbols grounded in the realworld for it to feel like something to be having higher-order thoughts. The intention of this clarification is toexclude systems such as a computer running a programwhen there is in addition some sort of control or evenoverseeing program checking the operation of the firstprogram. We would want to say that in such a situation itwould feel like something to be running the higher-levelcontrol program only if the first-order program was sym-bolically performing operations on the world and receiv-ing input about the results of those operations, and if thehigher-order system understood what the first-order sys-tem was trying to do in the world. The second clarifica-tion is that the plan would have to be a unique string ofsteps, in much the same way as a sentence can be aunique and one-off string of words. The point here isthat it is helpful to be able to think about particularone-off plans, and to correct them; and that this type ofoperation is very different from the slow learning of fixedrules by trial and error.
This analysis does not yet give an account for sensoryqualia ("raw sensory feels"; for example, why "red" feelsred), for emotional qualia (e.g. why a rewarding touchproduces an emotional feeling of pleasure), or for moti-vational qualia (e.g. why food deprivation makes usfeelhungry). The view I suggest on such qualia is as follows.Information processing in and from our sensory systems(e.g. the sight of the colour red) may be relevant to plan-ning actions using language and the conscious processingthereby implied. Given that these inputs must be repre-sented in the system that plans, we may ask whether it ismore likely that we would be conscious of them or thatwe would not. I suggest that it would be a very special-purpose system that would allow such sensory inputs,and emotional and motivational states, to be part of (lin-guistically based) planning, and yet remain unconscious.It seems to be much more parsimonious to hold that wewould be conscious of such sensory, emotional and moti-vational qualia because they would be being used (or areavailable to be used) in this type of (linguistically based)higher-order thought processing, and this is what Ipropose.
The explanation of emotional and motivational sub-jective feelings or qualia that this discussion has ledtowards is thus that they should be felt as consciousbecause they enter into a specialized linguistic symbol-manipulation system that is part of a higher-orderthought system that is capable of reflecting on and cor-recting its lower-order thoughts involved, for example, inthe flexible planning of actions. It would require a veryspecial machine to allow this higher-order linguisticallybased thought processing, which is conscious by its nat-ure, to occur without the sensory, emotional and motiva-tional states (which must be taken into account by thehigher-order thought system) becoming felt qualia. Thequalia are thus accounted for by the evolution of the
linguistic system that can reflect on and correct its ownlower-order processes, and thus has adaptive value.
This account implies that it may be especially animalswith a higher-order belief and thought system and withlinguistic symbol manipulation that have qualia. It maybe that much non-human animal behaviour, provided thatit does not require flexible linguistic planning and correc-tion by reflection, could take place according to reinfor-cement-guidance (using, e.g. stimulus-reinforcementassociation learning in the amygdala and orbitofrontalcortex, Rolls, 1990b, 1996c), and rule-following (imple-mented, e.g. using habit or stimulus-response learning inthe basal ganglia, Rolls, 1994a; Rolls & Johnstone,1992). Such behaviours might appear very similar tohuman behaviour performed in similar circumstances,but would not imply qualia. It would be primarily byvirtue of a system for reflecting on flexible, linguistic,planning behaviour that humans (and animals close tohumans, with demonstrable syntactic manipulation ofsymbols, and the ability to think about these linguisticprocesses) would be different from other animals, andwould have evolved qualia.
For processing in a part of our brain to be able to reachconsciousness, appropriate pathways must be present.Certain constraints arise here. For example, in the sen-sory pathways, the nature of the representation maychange as it passes through a hierarchy of processinglevels, and in order to be conscious of the informationin the form in which it is represented in early processingstages, the early processing stages must have access tothe part of the brain necessary for consciousness. Anexample is provided by processing in the taste system.In the primate primary taste cortex, neurons respond totaste independently of hunger, yet in the secondary tastecortex, food-related taste neurons (e.g. responding tosweet taste) only respond to food if hunger is present,and gradually stop responding to that taste during feedingto satiety (see Rolls, 1989d, 1993, 1995a). Now the qual-ity of the tastant (sweet, salt, etc.) and its intensity are notaffected by hunger, but the pleasantness of its taste isdecreased to zero (neutral) (or even becomes unpleasant)after we have eaten it to satiety. The implication of this isthat for quality and intensity information about taste, wemust be conscious of what is represented in the primarytaste cortex (or perhaps in another area connected to itwhich bypasses the secondary taste cortex), and not ofwhat is represented in the secondary taste cortex. In con-trast, for the pleasantness of a taste, consciousness of thiscould not reflect what is represented in the primary tastecortex, but instead what is represented in the secondarytaste cortex (or in an area beyond it). The same argumentarises for reward in general, and therefore for emotion,which in primates is not represented early on in proces-sing in the sensory pathways (nor in or before the inferiortemporal cortex for vision), but in the areas to whichthese object analysis systems project, such as the orbito-frontal cortex, where the reward value of visual stimuli is
1235Consciousness in Neural Networks?

reflected in the responses of neurons to visual stimuli (seeRolls, 1990b, 1995a, c). It is also of interest that rewardsignals (e.g. the taste of food when we are hungry) areassociated with subjective feelings of pleasure (see Rolls,1990b, 1993, 1995a, c). I suggest that this correspon-dence arises because pleasure is the subjective statethat represents in the conscious system a signal that ispositively reinforcing (rewarding), and that inconsistentbehaviour would result if the representations did not cor-respond to a signal for positive reinforcement in both theconscious and the non-conscious processing systems.
Do these arguments mean that the conscious sensationof, for example, taste quality (i.e. identity and intensity)is represented or occurs in the primary taste cortex, andof the pleasantness of taste in the secondary taste cortex,and that activity in these areas is sufficient for conscioussensations (qualia) to occur? I do not suggest this at all.Instead, the arguments I have put forward above suggestthat we are only conscious of representations when wehave high-order thoughts about them. The implicationthen is that pathways must connect from each of thebrain areas in which information is represented aboutwhich we can be conscious, to the system which hasthe higher-order thoughts, which, as I have arguedabove, requires language. Thus, in the example given,there must be connections to the language areas fromthe primary taste cortex, which need not be direct, butwhich must bypass the secondary taste cortex, in whichthe information is represented differently (see Rolls,1989d, 1995a). There must also be pathways from thesecondary taste cortex, not necessarily direct, to thelanguage areas so that we can have higher-order thoughtsabout the pleasantness of the representation in the sec-ondary taste cortex. There would also need to be path-ways from the hippocampus, implicated in the recall ofdeclarative memories, back to the language areas of thecerebral cortex (at least via the cortical areas whichreceive backprojections from the hippocampus, seeFigure 3, which would in turn need connections to thelanguage areas). A schematic diagram incorporating thisanatomical prediction about human cortical neuralconnectivity in relation to consciousness is shown inFigure 5.
One question that has been discussed is whether thereis a causal role for consciousness (e.g. Armstrong &Malcolm, 1984). The position to which the above argu-ments lead is that indeed conscious processing does havea causal role in the elicitation of behaviour, but onlyunder the set of circumstances when higher-orderthoughts play a role in correcting or influencing lower-order thoughts. The sense in which the consciousness iscausal is then, it is suggested, that the higher-orderthought is causally involved in correcting the lower-order thought; and that it is a property of the higher-order thought system that it feels like something whenit is operating. As we have seen, some behaviouralresponses can be elicited when there is not this type of
reflective control of lower-order processing, nor indeedany contribution of language. There are many brain pro-cessing routes to output regions, and only one of theseinvolves conscious, verbally represented processingwhich can later be recalled (see Figure 4).
Some of the brain systems involved in this type ofconscious processing that it is suggested has evolved tohelp the correction of plans are as follows. One module isa system that can implement syntax, because the manysymbols (e.g. names of people) that are part of the planmust be correctly linked or bound. Such linking might beof the form: "if A does this, then B is likely to do this, andthis will cause C to do this…". The requirement of syntaxfor this type of planning implies that an output to lan-guage systems in the brain is required for this type ofplanning (see Figure 4). Another building block for suchplanning operations in the brain may be the type of short-term memory in which the prefrontal cortex is involved.This short-term memory may be, for example, in non-human primates of where in space a response has justbeen made. A development of this type of short-termresponse memory system in humans to allow multipleshort-term memories to be held in place correctly, pre-ferably with the temporal order of the different items inthe short-term memory coded correctly, may be anotherbuilding block for the multiple step "if…then" type ofcomputation so as to form a multiple step plan. Suchshort-term memories are implemented in the (dorso-lateral and inferior convexity) prefrontal cortex of
1236 E. T. Rolls
FIGURE 5. Schematic illustration indicating that early corticalstages in information processing may need access to languageareas which bypass subsequent levels in the hierarchy, so thatconsciousness of what is represented in early cortical stages,and which may not be represented in later cortical stages, canoccur. Higher-order linguistic thoughts (HOLTs) could be imple-mented in the language cortex itself, and would not need a sepa-rate cortical area. Backprojections, a notable feature of corticalconnectivity, with many probable functions including recall(Rolls, 1989a, b, 1996a), probably reciprocate all the connectionsshown.

non-human primates and humans (see Goldman-Rakic,1996; Petrides, 1996), and may be part of the reason whyprefrontal cortex damage impairs planning (see Shallice& Burgess, 1996).
It is of interest to comment on how the evolution of asystem for flexible planning might affect emotions. Con-sider grief which may occur when a reward is terminatedand no immediate action is possible (see Rolls, 1990b,1995c). It may be adaptive by leading to a cessation ofthe formerly rewarded behaviour and thus facilitating thepossible identification of other positive reinforcers in theenvironment. In humans, grief may be particularly potentbecause it becomes represented in a system which canplan ahead, and understand the enduring implications ofthe loss. (Thinking about or verbally discussing emo-tional states may also in these circumstances help,because this can lead towards the identification of newor alternative reinforcers, and of the realization that, forexample, the negative consequences may not be as bad asfeared.)
This account of consciousness also leads to a sugges-tion about the processing that underlies the feeling of freewill. Free will would in this scheme involve the use oflanguage to check many moves ahead on a number ofpossible series of actions and their outcomes, and thenwith this information to make a choice from the likelyoutcomes of different possible series of actions. (If, incontrast, choices were made only on the basis of thereinforcement value of immediately available stimuli,without the arbitrary syntactic symbol manipulationmade possible by language, then the choice strategywould be much more limited, and we might not wantto use the term free will, as all the consequences ofthose actions would not have been computed.) It is sug-gested that when this type of reflective, conscious, infor-mation processing is occurring and leading to action, thesystem performing this processing and producing theaction would have to believe that it could cause theaction, for otherwise inconsistencies would arise, andthe system might no longer try to initiate action. Thisbelief held by the system may partly underlie the feelingof free will. At other times, when other brain modules areinitiating actions, the conscious processor may confabu-late and believe that it caused the action, or at least givean account (possibly wrong) of why the action wasinitiated. The fact that the conscious processor mayhave the belief even in these circumstances that itinitiated the action may arise as a property of it beinginconsistent for a system which can take overall controlusing conscious verbal processing to believe that it wasoverridden by another system.
In the operation of such a free will system, the uncer-tainties introduced by the limited information possibleabout the likely outcomes of series of actions, and theinability to use optimal algorithms when combining con-ditional probabilities, would be much more importantfactors than whether the brain operates deterministically
or not. (The operation of brain machinery must be rela-tively deterministic, for it has evolved to provide reliableoutputs for given inputs.)
These are my initial thoughts on why we have con-sciousness, and are conscious of sensory, emotional andmotivational qualia, as well as qualia associated withfirst-order linguistic thoughts. It is likely that theoriesof consciousness will continue to undergo rapid devel-opment, and current theories should not be taken to havepractical implications.
5. DISCUSSION
Some ways in which the current theory may be differentfrom other related theories follow. The current theoryholds that it is higher-order linguistic thoughts(HOLTs) that are closely associated with consciousness,and this may differ from Rosenthal’s higher-orderthoughts (HOTs) theory (Rosenthal, 1986, 1990, 1993),in the emphasis in the current theory on language. Simi-larly, the theory differs from suggestions for a function ofconsciousness in "monitoring" (e.g. Marcel, 1988), inthat a specification is given in the present theory of thetype of correction being performed of first-order linguis-tic thought processes, and of the computational advan-tages of this. Language in the current theory is defined bysyntactic manipulation of symbols, and does not neces-sarily imply verbal language. The reason that strongemphasis is placed on language is that it is as a resultof having a multistep flexible "on the fly" reasoning pro-cedure that errors which cannot be easily corrected byreward or punishment received at the end of the reason-ing, need ‘thoughts about thoughts’, that is, some type ofsupervisory and monitoring process, to detect whereerrors in the reasoning have occurred. This suggestionon the adaptive value in evolution of such a higher-order linguistic thought process for multistep planningahead, and correcting such plans, may also be differentfrom earlier work. Put another way, this point is thatcredit assignment when reward or punishment arereceived is straightforward in a one layer network (inwhich the reinforcement can be used directly to correctnodes in error, or responses), but is very difficult in amultistep linguistic process executed once "on the fly".Very complex mappings in a multilayer network can belearned if hundreds of learning trials are provided. Butonce these complex mappings are learned, their successor failure in a new situation on a given trial cannot beevaluated and corrected by the network. Indeed, the com-plex mappings achieved by such networks (e.g. back-propagation nets) mean that after training they operateaccording to fixed rules, and are often impenetrable andinflexible. In contrast, to correct a multistep, singleoccasion, linguistically based plan or procedure, recallof the steps just made in the reasoning or planning, andperhaps related episodic material, needs to occur, so thatthe link in the chain which is most likely to be in error
1237Consciousness in Neural Networks?

can be identified. This may be part of the reason whythere is a close relation between declarative memorysystems, which can explicitly recall memories, and con-sciousness.
Some computer programs may have supervisory pro-cesses. Should these count as higher-order linguisticthought processes? My current response to this is thatthey should not, to the extent that they operate withfixed rules to correct the operation of a system whichdoes not itself involve linguistic thoughts about symbolsgrounded semantically in the external world. If, on theother hand, it were possible to implement on a computersuch a high-order linguistic thought supervisory correc-tion process to correct first-order linguistic thoughts withsymbols grounded in the real world, then this processwould prima facie be conscious. If it were possible in athought experiment to reproduce the neural connectivityand operation of a human brain on a computer, thenprima facie it would also have the attributes of conscious-ness. It might continue to have those attributes for as longas power was applied to the system.
Another possible difference from earlier theories isthat raw sensory feels are suggested to arise as a conse-quence of having a system that can think about its ownthoughts. Raw sensory feels, and subjective states asso-ciated with emotional and motivational states, may notnecessarily arise first in evolution.
A property often attributed to consciousness is that it isunitary. The current theory would account for this by thelimited syntactic capability of neuronal networks in thebrain, which renders it difficult to implement more than afew syntactic bindings of symbols simultaneously (seeMcLeod et al., 1998; Rolls & Treves, 1997). This limita-tion makes it difficult to run several "streams of con-sciousness" simultaneously. In addition, given that alinguistic system can control behavioural output, severalparallel streams might produce maladaptive behaviour(apparent as, e.g. indecision), and might be selectedagainst. The close relation between, and the limited capa-city of, both the stream of consciousness, and auditory–verbal short-term memory, may be that both implementthe capacity for syntax in neural networks. Whethersyntax in real neuronal networks is implemented by tem-poral binding (see von der Malsburg, 1990) is still anunresolved issue (see Rolls & Treves, 1997). (For exam-ple, the code can be read off from the end of the visualsystem without taking the temporal aspects of the neuro-nal firing into account, as described above; much of theinformation about which stimulus is shown is availablein short times of 30–50 ms, and cortical neurons need firefor only this long during the identification of objects(Rolls & Tovee, 1994; Rolls et al., 1994b; Tovee &Rolls, 1995; Tovee et al., 1993) (these are rather shorttime windows for the expression of multiple separatepopulations of synchronized neurons); and oscillations,at least, are not an obvious property of neuronal firing inthe primate temporal cortical visual areas involved in the
representation of faces and objects (Tovee & Rolls,1992).)
The current theory holds that consciousness arises byvirtue of a system that can think linguistically about itsown linguistic thoughts. The advantages for a system ofbeing able to do this have been described, and this hasbeen suggested as the reason why consciousness evolved.The evidence that consciousness arises by virtue of hav-ing a system that can perform higher-order linguisticprocessing is, however, and I think may remain, circum-stantial. (Why must it feel like something when we areperforming a certain type of information processing? Theevidence described here suggests that it does feel likesomething when we are performing a certain type ofinformation processing, but does not produce a strongreason for why it has to feel like something. It justdoes, when we are using this linguistic processing systemcapable of higher-order thoughts.) The evidence, sum-marized above, includes the points that we think of our-selves as conscious when, for example, we recall earlierevents, compare them with current events, and plan manysteps ahead. Evidence also comes from neurologicalcases, from, for example, split brain patients (who mayconfabulate conscious stories about what is happening intheir other, non-language, hemisphere), and from casessuch as frontal lobe patients who can tell one consciouslywhat they should be doing, but nevertheless may bedoing the opposite. (The force of this type of case isthat much of our behaviour may normally be producedby routes about which we cannot verbalize, and are notconscious about.) This raises the issue of the causal roleof consciousness. Does consciousness cause ourbehaviour?2 The view that I currently hold is that theinformation processing which is related to consciousness(activity in a linguistic system capable of higher-orderthoughts, and used for planning and correcting the opera-tion of lower-order linguistic systems) can play a causalrole in producing our behaviour (see Figure 4). It is, Ipostulate, a property of processing in this system(capable of higher-order thoughts) that it feels like some-thing to be performing that type of processing. It is in thissense that I suggest that consciousness can act causally toinfluence our behaviour: consciousness is the propertythat occurs when a linguistic system is thinking aboutits lower-order thoughts. The hypothesis that it doesfeel like something when this processing is taking
1238 E. T. Rolls
2 This raises the issue of the causal relation between mental eventsand neurophysiological events, part of the mind–body problem. Myview is that the relation between mental events and neurophysiologicalevents is similar (apart from the problem of consciousness) to the rela-tion between the program running in a computer and the hardware in thecomputer. In a sense, the program causes the logic gates to move to thenext state. This move causes the program to move to its next state.Effectively, we are looking at different levels of what is overall theoperation of asystem, and causality can usefully be understood asoperating both within levels (causing one step of the program tomove to the next), as well as between levels (e.g. software to hardwareand vice versa).

place is at least to some extent testable: humans perform-ing this type of higher-order linguistic processing, forexample, recalling episodic memories and comparingthem with current circumstances, who denied being con-scious, would prima facie constitute evidence against thetheory. Most humans would find it very implausiblethough to posit that they could be thinking about theirown thoughts, and reflecting on their own thoughts, with-out being conscious. This type of processing does appearto be for most humans to be necessarily conscious.
Finally, I provide a short specification of what mighthave to be implemented in a neural network to imple-ment conscious processing. First, a linguistic system, notnecessarily verbal, but implementing syntax betweensymbols implemented in the environment would beneeded. Then a higher-order thought system also imple-menting syntax and able to think about the represen-tations in the first-order language system, and able tocorrect the reasoning in the first-order linguistic systemin a flexible manner, would be needed. So my answer tothe title of this paper is that consciousness can be imple-mented in neural networks (and that this is a topic worthdiscussing), but that the neural networks would have toimplement the type of higher-order linguistic processingdescribed in this paper.
REFERENCES
Abbott, L. A., Rolls, E. T., & Tovee, M. J. (1996). Representationalcapacity of face coding in monkeys.Cerebral Cortex, 6, 498–505.
Armstrong, D. M., & Malcolm, N. (1984).Consciousness and Causal-ity. Oxford: Blackwell.
Boussaoud, D., Desimone, R., & Ungerleider, L. G. (1991). Visualtopography of area TEO in the macaque.Journal of ComputationalNeurology, 306,554–575.
Burgess, N., Recce, M., & O’Keefe, J. (1994). A model of hippocampalfunction.Neural Networks, 7, 1065–1081.
Carruthers, P. (1996).Language, Thought and Consciousness. Cam-bridge: Cambridge University Press.
Cheney, D. L., & Seyfarth, R. M. (1990).How Monkeys See the World.Chicago: University of Chicago Press.
Dennett, D. C. (1991).Consciousness Explained. London: Penguin.Foldiak, P. (1991). Learning invariance from transformation sequences.
Neural Computation, 3, 193–199.Fukushima, K. (1980). Neocognitron: a self-organizing neural network
model for a mechanism of pattern recognition unaffected by shift inposition.Biological Cybernetics, 36, 193–202.
Gaffan, D. (1994). Scene-specific memory for objects: a model of epi-sodic memory impairment in monkeys with fornix transection.Journal of Cognitive Neuroscience, 6, 305–320.
Gazzaniga, M. S. (1988). Brain modularity: towards a philosophy ofconscious experience. In A.J. Marcel & E. Bisiach (Eds.),Con-sciousness in Contemporary Science(Ch. 10, pp. 218–238).Oxford: Oxford University Press.
Gazzaniga, M. S. (1995). Consciousness and the cerebral hemispheres.In M. S. Gazzaniga (Ed.),The Cognitive Neurosciences(pp. 1392–1400). Cambridge, MA: MIT Press.
Gazzaniga, M. S., & LeDoux, J. (1978).The Integrated Mind. NewYork: Plenum.
Goldman-Rakic, P. S. (1996). The prefrontal landscape: implications offunctional architecture for understanding human mentation and thecentral executive.Philosophical Transactions of the Royal Society,Series B, 351,1445–1453.
Hasselmo, M. E., & Bower, J. M. (1993). Acetylcholine and memory.Trends in Neuroscience, 16, 218–222.
Humphrey, N. K. (1980). Nature’s psychologists. In B. D. Josephson &V. S. Ramachandran (Eds.),Consciousness and the Physical World(pp. 57–80). Oxford: Pergamon.
Linsker, E. (1986). From basic network principles to neuralarchitecture.Proceedings of the National Academy of Sciences ofthe USA83, 7508–7512, 8390–8394, 8779–8783.
Linsker, E. (1988). Self-organization in a perceptual network.Computer, March 1988,105–117.
MacKay, D. J. C., & Miller, K. D. (1990). Analysis of Linsker’s simu-lation of Hebbian Rules.Neural Computation, 2, 173–187.
McClelland, J. L., McNaughton, B. L., & O’Reilly, R. C. (1995). Whythere are complementary learning systems in the hippocampus andneocortex: insights from the successes and failures of connectionistmodels of learning and memory.Psychological Review, 102,419–457.
McLeod, P., Plunkett, K. & Rolls, E. T. (1998).Introduction to Con-nectionist Modelling of Cognitive Processes. Oxford: Oxford Uni-versity Press.
Marcel, A. J. (1988). Phenomenal experience and functionalism. In A.J. Marcel and E. Bisiach (Eds.),Consciousness in ContemporaryScience(pp. 121–158). Oxford: Oxford University Press.
Petrides, M. (1996). Specialized systems for the processing of mnemo-nic information within the primate frontal cortex.PhilosophicalTransactions of the Royal Society, Series B, 351,1455–1462.
Rolls, E. T. (1989a). Functions of neuronal networks in the hippocam-pus and neocortex in memory. In J. H. Byrne & W. O. Berry (Eds.),Neural Models of Plasticity: Experimental and TheoreticalApproaches(pp. 240–265). San Diego, CA: Academic Press.
Rolls, E. T. (1989b). The representation and storage of information inneuronal networks in the primate cerebral cortex and hippocampus.In R. Durbin, C. Miall, & G. Mitchison (Eds.),The ComputingNeuron(pp. 125–159). Wokingham, UK: Addison Wesley.
Rolls, E. T. (1989c). Functions of neuronal networks in the hippocam-pus and cerebral cortex in memory. In R. M. J. Cotterill (Ed.),Models of Brain Function(pp. 15–33). Cambridge: CambridgeUniversity Press.
Rolls, E.T. (1989d). Information processing in the taste system ofprimates.Journal of Experimental Biology, 146,141–164.
Rolls, E. T. (1990a). Functions of the primate hippocampus in spatialprocessing and memory. In D. S. Olton & R. P. Kesner (Eds.),Neurobiology of Comparative Cognition(pp. 339–362). Hillsdale,NJ: Lawrence Erlbaum.
Rolls, E.T. (1990b). A theory of emotion, and its application to under-standing the neural basis of emotion.Cognition and Emotion, 4,161–190.
Rolls, E.T. (1991). Neural organisation of higher visual functions.Cur-rent Opinion in Neurobiology, 1, 274–278.
Rolls, E.T. (1992). Neurophysiological mechanisms underlying faceprocessing within and beyond the temporal cortical visual areas.Philosophical Transactions of the Royal Society, 335,11–21.
Rolls, E. T. (1993). The neural control of feeding in primates. In D. A.Booth (Ed.),Neurophysiology of Ingestion(pp. 137–169). Oxford:Pergamon.
Rolls, E.T. (1994a). Neurophysiology and cognitive functions of thestriatum.Revue Neurologique (Paris), 150,648–660.
Rolls, E.T. (1994b). Brain mechanisms for invariant visual recognitionand learning.Behavioural Processes, 33, 113–138.
Rolls, E. T. (1995a). Central taste anatomy and neurophysiology. InR. L. Doty (Ed.),Handbook of Olfaction and Gustation(pp. 549–573). New York: Dekker.
Rolls, E.T. (1995b). Learning mechanisms in the temporal lobe visualcortex.Behavioural Brain Research, 66, 177–185.
Rolls, E. T. (1995c). A theory of emotion and consciousness, and itsapplication to understanding the neural basis of emotion. In M. S.Gazzaniga (Ed.),The Cognitive Neurosciences(pp. 1091–1106).Cambridge, MA: MIT Press.
1239Consciousness in Neural Networks?

Rolls, E.T. (1996a). A theory of hippocampal function in memory.Hippocampus, 6, 601–620.
Rolls, E. T. (1996b). The representation of space in the primate hippo-campus, and episodic memory. In T. Ono, B. L. McNaughton, S.Molotchnikoff, E. T. Rolls, & H. Nishijo (Eds.),Perception,Memory and Emotion: Frontier in Neuroscience(pp. 567–579).Amsterdam: Elsevier.
Rolls, E.T. (1996c). The orbitofrontal cortex.Philosophical Transac-tions of the Royal Society, Series B, 351,1433–1444.
Rolls, E. T. (1996d). The representation of space in the primate hippo-campus, and its relation to memory. In K. Ishikawa, J. L. McGaugh& H. Sakata (Eds.),Brain Processes and Memory(pp. 203–227).Amsterdam: Elsevier.
Rolls, E. T. (1997). A neurophysiological and computational approachto the functions of the temporal lobe cortical visual areas in invar-iant object recognition. In L. Harris & M. Jenkin (Eds.),Computa-tional and Psychophysical Mechanisms of Visual Coding.Cambridge: Cambridge University Press.
Rolls, E. T., & Johnstone, S. (1992). Neurophysiological analysis ofstriatal function. In G. Vallar, S. F. Cappa, & C. W. Wallesch (Eds.),Neuropsychological Disorders Associated with Subcortical Lesions(pp. 61–97). Oxford: Oxford University Press.
Rolls, E.T., & Tovee, M.J. (1994). Processing speed in the cerebralcortex, and the neurophysiology of visual masking.Proceedingsof the Royal Society, Series B, 257,9–15.
Rolls, E.T., & Tovee, M.J. (1995). Sparseness of the neuronal repre-sentation of stimuli in the primate temporal visual cortex.Journal ofNeurophysiology, 73, 713–726.
Rolls, E. T., & Treves, A. (1997).Neural Networks and Brain Function.Oxford: Oxford University Press.
Rolls, E. T., Booth, M. C. A., & Treves, A. (1996). View-invariantrepresentations of objects in the inferior temporal visual cortex.Society for Neuroscience Abstracts, 22, 760.5.
Rolls, E.T., Hornak, J., Wade, D., & McGrath, J. (1994a). Emotion-related learning in patients with social and emotional changes asso-ciated with frontal lobe damage.Journal of Neurology, Neurosur-gery and Psychiatry, 57, 1518–1524.
Rolls, E.T., Tovee, M.J., Purcell, D.G., Stewart, A.L., & Azzopardi, P.(1994b). The responses of neurons in the temporal cortex of pri-mates, and face identification and detection.Experimental BrainResearch, 101,474–484.
Rolls, E. T., Tovee, M., Treves, A., & Panzeri, S. (1997a). Informationin the neuronal representation of individual stimuli in the primate
temporal visual cortex.Journal of Computational Neuroscience,inpress.
Rolls, E. T., Treves, A., & Tovee, M. J. (1997b). The representationalcapacity of the distributed encoding of information provided bypopulations of neurons in the primate temporal visual cortex.Experimental Brain Research, 114,149–162.
Rosenthal, D. M. (1986). Two concepts of consciousness.PhilosophicalStudies, 49, 329–359.
Rosenthal, D. (1990).A Theory of Consciousness(ZIF Rep. 40). Bie-lefeld, Germany: Zentrum fur Interdisziplinaire Forschung.
Rosenthal, D. M. (1993). Thinking that one thinks. In M. Davies &G. W. Humphreys (Eds.),Consciousness(pp. 197–223). Oxford:Blackwell.
Shallice, T., & Burgess, P. (1996). The domain of supervisory processesand temporal organization of behaviour.Philosophical Trans-actions of the Royal Society, Series B, 351,1405–1411.
Squire, L. R. (1992). Memory and the hippocampus: a synthesis fromfindings with rats, monkeys and humans.Psychological Review, 99,195–231.
Tovee, M. J., & Rolls, E. T. (1992). Oscillatory activity is not evident inthe primate temporal visual cortex with static stimuli.Neuroreport,3, 369–372.
Tovee, M. J., & Rolls, E. T. (1995). Information encoding in short firingrate epochs by single neurons in the primate temporal visual cortex.Visual Cognition, 2, 35–58.
Tovee, M. J., Rolls, E. T., Treves, A., & Bellis, R. P. (1993). Informa-tion encoding and the responses of single neurons in the primatetemporal visual cortex.Journal of Neurophysiology, 70, 640–654.
Treves, A., & Rolls, E. T. (1992). Computational constraints suggest theneed for two distinct input systems to the hippocampal CA3network.Hippocampus, 2, 189–199.
Treves, A., & Rolls, E. T. (1994). A computational analysis of the roleof the hippocampus in memory.Hippocampus, 4, 374–391.
von der Malsburg, C. (1990). A neural architecture for the representa-tion of scenes. In J. L. McGaugh, N. M. Weinberger and G. Lynch(Eds.),Brain Organization and Memory: Cells, Systems and Cir-cuits (pp. 356–372). New York: Oxford University Press.
Wallis, G., & Rolls, E. T. (1997). Invariant face and object recognitionin the visual system.Progress in Neurobiology, 51, 167–194.
Wallis, G., Rolls, E. T., & Foldiak, P. (1993). Learning invariantresponses to the natural transformations of objects.InternationalJoint Conference on Neural Networks, 2, 1087–1090
1240 E. T. Rolls

1997 SPECIAL ISSUE
From Brains to Neural Nets to Brains
Erich Harth
Physics Department, Syracuse University
(Received10 October1996;accepted6 January1997)
Abstract—The paper traces theoretical work concerning the understanding and simulation of brain functions fromearly studies of artificial neural nets to present considerations of human consciousness. The emphasis is on work carriedout since about 1963 at my laboratory in collaboration with my students. The discussion centers on sensory, especiallyvisual, information processing, some of the cerebral mechanisms involved, and current approaches to an understandingof conscious perception. The sketchpad model, in which the ubiquitous feedback pathways in the brain play a dominantrole, is described, together with a discussion of the meaning and applicability of scientific reductionism to the problem ofconsciousness.q 1997 Elsevier Science Ltd.
Keywords—Neurodynamics, Receptive fields, Thalamocortical pathways, Reticular formation, Hillclimbing processes,Consciousness, Reductionism.
1. INTRODUCTION
I was asked to review my work and ideas for this SpecialIssue on Consciousness. My aim here will be to retracemy own circuitous route that took me from studyingprimitive structures of formal neurons to that hardest ofhard questions, the nature of consciousness.
In the beginning, the problem of how mind arises fromthe physical brain seemed not too awesome a challengeto some of us. With enviable insouciance one theoreti-cian had declared that the constellation of neurons activein a neural net—both its biological prototype and theelectronic analogue—is quite simply… a thought! Wehave become more sophisticated both in our knowledgeof the brain and in constructing powerful neural net-works, but the question What, if anything, lies betweenthe machine and human thought? has continued to be anunresolved issue.
The impetus for the many studies of neural nets in thelate 1950s and early 1960s came mostly from a seminalpaper by McCulloch & Pitts (1943) in which neuronswere treated as simple threshold elements with all-or-none response. The authors showed that networks ofsuch formal neuronswere capable of carrying out anycomputation whose logical steps could be defined, andwere thus functionally equivalent to universal Turing
machines. The McCulloch–Pitts paper raised the hopethat all manner of brain functions could be under-stood—and eventually synthesized—by systems com-posed of such primitive logical elements, relying onlyon the sheer mass of the net and, perhaps, the delicacyof its interconnections.
The ability to undergo permanent or semi-permanentchanges is a further essential ingredient in a system thatis to mimic the brain. The psychologist Donald Hebb(1949) suggested that this plasticity resided in themodifiable strengths of neuronal interconnections,the synaptic couplings between neurons. He furtherassumed that these synaptic changes, calledsynapticfacilitation, result from strictly local effects, that is,from the coincidence of a presynaptic action potentialand a postsynaptic excitation of the target cell.
It remains to endow the net of real or simulatedneurons with structure. Here, the controversy raged fora long time, whether the biological neural net was hard-wired in the sense that the entire connectivity was bothdetermined and essential to its functioning, or whethergenetic determination left some of the details of thecircuitry to chance. The extreme assumption of a single,amorphous, randomly connected net was clearly inviolation of biological facts, but was nevertheless oftentaken as the starting point of theoretical discussions andthe study of artificial neural networks. At the otherextreme, the assertion of a rigidly deterministic circuitryin which all detail is specified, has been contradictedby numerous brain studies.
Neural Networks, Vol. 10, No. 7, pp. 1241–1255, 1997q 1997 Elsevier Science Ltd. All rights reserved
Printed in Great Britain0893–6080/97 $17.00+.00
PII: S0893-6080(97)00048-8
Pergamon
1241
Requests for reprints should be sent to: Erich Harth, Department ofPhysics, Syracuse University, Syracuse, NY 13244-1130, USA; E-mail:[email protected].

There is, finally, the question what we wish the net toaccomplish, or what output to expect, and in what code.Here, again, Hebb has shown the way. Meaning is to beattached not to the firing of single neurons (sometimescalled grandmother cells), but to subpopulations of theneural net that Hebb termedcell assemblies. A cellassembly may arise through synaptic facilitations andis capable of sustaining neural activity within itselfthrough reverberating circuits.
Hebb’s ideas of synaptic facilitation and cell assem-blies have been the cornerstone of practically all thinkingabout the brain’s ability to record, associate, and recallsensory events, as well as the chief ingredient in learningmachines we call neural nets. They appear over andover in most theories of brain functions from the mostprimitive to the elusive phenomena of human thoughtand consciousness.
Together with the concept of McCulloch–Pittsneurons, the Hebb rules make it appear that brain func-tions are mechanisms that are conceptually simple. Thishas given rise to bold attempts to simulate differentaspects of the brain by use of artificial nets of formalneurons, and to a general euphoria about their futurepotential. A word of caution is in order. It has becomeincreasingly clear just how ‘‘impoverished’’ artificialneurons are when compared to their biological arche-types. The proliferation of recognized neurotransmitters,neuromodulators, and neurohormones since about themid 1970s, all of which affect the behavior of neuronsbut take their information from widely different sources,has made us aware of the enormous functional complex-ity that exists already at the level of individual cells.
2. SIMPLE NEURAL NETS ANDBRAIN FUNCTIONS
The late 1050s saw a number of papers (Allanson, 1956;Rochester et al., 1956; Beurle, 1959) investigating thedynamics of populations of primitive neuron-like ele-ments, and incorporating the ideas proposed byMcCulloch & Pitts (1943) and Hebb (1949).
In 1958 Frank Rosenblatt (1958) initiated a newapproach in neural net technology. Rather than studyingthe dynamic properties of multiply connected networksof many formal neurons, he conceived a circuit consist-ing of an array of input units, connected through a set ofintermediate neurons to an array of output units. Theinputs and outputs are both binary words. He calledthis device a perceptron. Initial coupling strengths arechosen arbitrarily, and, for a particular task, a truthtable prescribes the ‘‘correct’’ output for every possibleinput. In a training procedure many different inputs arepresented to the perceptron. A simple training rule speci-fies the changes to be made to the coupling strengthsaccording to whether the net gave a correct or incorrectresponse. After training, the perceptron will in generalproduce correct answers a large fraction of the time.
Limitations of the original elementary perceptronswere pointed out by Minsky & Papert (1969); thesewere largely overcome by adding one or more layers ofhidden units to the device. The simple reinforcementrules for elementary perceptrons could no longer beapplied since the role of the hidden units in forming aparticular output is not readily ascertained. A procedurecalled back propagation (Rumelhart et al., 1986) solvesthis problem, but is time-consuming and biologicallyimplausible. Neither the elementary perceptron, nor itsmore sophisticated hidden-layer successor, containedneural feedback loops; hence no reverberatory activity.
2.1. Netlets
My own contribution to the field began with a paperpresented in 1965 at a conference on Automata Theoryheld in Ravello (Harth, 1966). I carried out computersimulations of a randomly connected net of 100 formalneurons whose structure was given by a ‘sparse’1003 100 asymmetric coupling matrix. Before learning,the net was amorphous and isotropic except for statisticalfluctuations. In addition, 25 input units were randomlyand diffusely connected into the net.
Stimuli were defined as 25-five digit binary words.The resulting state of the net—a 100-bit word—repre-sented the response. The application of Hebbian learningrules brought out two related results. When the net wastrained on the simultaneous presentation of two differentstimuli, then the subsequent presentation of just one ofthem would produce a response that contained elementsof the responses to both stimuli (association). Similarly,the presentation of parts of a stimulus would tend toproduce a response closer to that of the previouslylearned entire stimulus (pattern completion).
2.2. Associations and Reflexes
Harth & Edgar (1967) extended this study to associationnets of 1000 neurons. These were defined by such para-meters as thresholds, average number of efferent junc-tions per neuron, average coupling coefficients, and ratioof inhibitory to excitatory neurons. Although containingmany loops, reverberatory activities were kept down bysparse connectivity matrices and high firing thresholds.The response to a given input vector is the totality ofneurons that reach firing level following the stimulus.We demonstrated that, upon applying rules of synapticfacilitation, systems of this kind are able to perform aremarkable variety of ‘‘cognitive’’ tasks such as simpleconditioned reflexes, dual-conditioned reflexes, bidirec-tional, and reverse conditioned reflexes, as defined byAsratian (1965).
We assumed that subsets of the network acted asHebbian cell assemblies with fixed and arbitrarilyassigned meanings. Associations between different cellassemblies were accomplished by strengthening their
1242 E. Harth

interconnections. The previously observed effect ofpattern completion means that the association of anassemblyA with an assemblyB will trigger a portionof B wheneverA alone is activated, and vice versa.
In this approach the simple conditioned reflex is char-acterized schematically in Figure 1, which shows a partof the matrix of coupling coefficients. The domainsU, C,andR are cell assemblies representing the unconditionedand conditioned stimuli and the response, respectively.Here, each small square in Figure 1 is ann 3 n matrix ofcoupling strengths, wheren is the number of neuronsin the cell assembly. Before training, the only inhomo-geneities in the net are the shaded area indicatingenhanced coupling coefficients withinU, C, andR, andfrom U to R, causing activity inR wheneverU istriggered, but no activity inR when C is active. Thecross-hatched areas represent synaptic facilitation dueto learning, in this case the frequent coincidences ofstimuli U andC. Thus, triggering the conditioned stimu-lus C will cause activity to extend intoU and from thereinto R.
Figure 2 is a schematic diagram of the connectivitymatrix in a classification task. Eleven cell assemblies areshown here, representing different objects and propertiesas labeled. The initially random, homogeneous andisotropic connectivity matrix is shown after associations,i.e., enhancement of couplings between netlets. Theresulting matrix is seen to be macroscopically sym-metrical, but microscopically asymmetric. Cross-hatched
areas are domains of enhanced synaptic strengths result-ing from binary associations: FIRE ENGINE with RED,FIRE ENGINE with METAL, etc. The net is thusinstructed that fire engines are red and of metal, thatpencils are yellow and of wood, traffic signs yellowand of metal, that red, yellow, and blue are colors andplastic, wood and metal materials. With properly chosenparameters of thresholds and average normal andenhanced synaptic strengths, it will take more than onecell assembly to cause significant activity in an asso-ciated assembly. Taking advantage of the non-linearityof responses, the diagram in Figure 2 shows that the netwill respond with RED when the input is FIRE ENGINEplus COLOR, but shows weak activities in the otherassemblies. Similarly, the question ‘‘What is the materialof pencils?’’ is answered by WOOD, ‘‘What is yellowand made of metal?’’ by TRAFFIC SIGN, and so on.
In a computer simulation, each of the cell assemblieswas made up of 75 formal neurons. The question ‘‘Whatis the color of traffis signs?’’ produced the activitiesshown in Table 1. The answer is given by the strongactivity of 16 out of 75 in the YELLOW cell assembly.
2.3. Neurodynamics
The next two papers (Harth et al., 1970b; Anninos et al.,1970) examine the dynamics of structured neural netsmade up of randomly connected subsystems that arenon-randomly connected to one another. We introducedhere the concept of randomness-in-the-small and struc-ture-in-the-large (Anninos et al., 1970). For the randomsubsystems we chose the term netlets. In a sense, the cellassemblies arising in the originally homogeneous
1243Brains and Neural Nets
FIGURE 1. Matrix of coupling coefficients in simulation of a con-ditioned reflex. Each small square represents a netlet of about100 neurons. U—unconditioned stimulus; C—conditionedstimulus: R—unconditioned response. Shaded areas denotedomains of enhanced coupling coefficients within netlets U, C,and R, and the initially strong connections from U to R. Cross-hatched areas are enhanced due to learning.
FIGURE 2. Connectivity matrix (defined as in Figure 1) in acomputer simulation experiment. The 11 netlets are assignedmeanings as indicated, and cross-hatched areas are enhanced.

connectivity matrix (Figs 1 and 2) become netlets in thetrained net.
We classified netlets according to their excitability,that is their ability to sustain activity in the absence ofinput. The parameters defining a netlet were, amongothers, fraction of inhibitory neurons, thresholds, andaverage coupling strengths. In the formalism used theresults were independent of the total number of‘‘neurons’’ in the netlet, except for the size of the statis-tical fluctuations. Dynamics of these structures wereobtained from statistical arguments as well as by com-puter simulations, some of which are shown in Figure 3.Here, the abscissaan refers to the fraction of neuronsactive in thenth iteration, and the ordinate is the activityatn þ 1. (The drop-off of the curves asan approaches 1.0is due to an assumed refractory period.) Thus, a very highinitial activity will find most neurons refractory in thenext iteration, and the activity in the net will soon ter-minate. The curve labeledh ¼ 2 (a measure of thenetlet’s excitability) shows two stationary pointsA andB (in addition to the one at the origin). We deduced thatthis gives rise to a hysteresis effect: a momentary inputthat triggers more than about 15% of the neurons in thenetlet causes sustained activity at the level of pointB,
that is with around 30% of the neurons active in eachiteration.
The dynamics were extended to the case of netletshaving a steady excitatory or inhibitory input from theoutside. PointsO, A, andB are now functions of the inputas shown in the schematicphase diagramof Figure 4.For changing input levels the net now exhibitshysteresis,with the arrows in Figure 4 indicating the spontaneousshift in activity level. For a combination of excitatoryand inhibitory inputs, stationary states form a surfacein three dimensions (Figure 5).
The calculations and simulations were extended(Wong & Harth, 1973) to netlets of different temporalcharacteristics describable by finite difference equationsof order two and greater. This formalism can take intoaccount any combination of refractory periods, sum-mation times, and effective delays.
The dynamic characteristics here described weresimulated for netlets of 1000 neurons, but turned out tobe robust down to very small populations. Shaw et al.(1982) have shown that small netlets can act as func-tional units in the central nervous system, and that thecooperative phenomena described above were reliablyperformed with net sizes down to about 30 neurons.
With the approach of randomness-in-the-small andstructure-in-the-large, neural systems can be constructedto perform specific tasks. Harth et al. (1975) and
1244 E. Harth
TABLE 1Cell assembly responses to ‘‘What is the color of traffic signs?’’
Traffic sign Pencil Fire eng. Color Yellow Red Blue Material Plastic Wood Metal75 3 0 75 16 2 3 1 0 6 0
FIGURE 3. Dynamics of neural netlet, showing activity at timen þ 1 as function of activity at n. Points are results of repeatedcomputer simulations and solid lines are computed from a finitedifference equation. Different curves are for different excitabil-ities of the netlets. Points O, A, and B are stationary points (fordetails, see Anninos et al., 1970).
FIGURE 4. Hysteresis of netlets with steady input j. The unstablestationary state A is indicated by the dotted line. It is seen that,when excitatory input exceeds jþ
crit, activity will rapidly increaseto Bcrit. The input will now have to be reduced to the negative(inhibitory) value j¹
crit, before the activity is extinguished. Twonetlets of lower excitability are shown for comparison.

Csermely et al. (1973) attempted to account for theoscillatory behavior in the escape motion of the molluskTritonia gilberti. In the anatomical studies reported byWillows (1967), the system is composed of four smallneural populations (Figure 6), where TFN refers totrigger group neurons, GEN to general excitor neurons,and DFN and VFN to dorsal and ventral flexion neurons,respectively. In the mechanism proposed by Willows, thetrigger neurons excite the GEN pool causing alternatingactivities in DFN and VFN (Figure 7).
In our simulation ofT. gilberti’s motion, we havetaken the connectivity proposed by Willows, replacingeach of his neuron pools with a random netlet. No furtherdetails of connectivity are thus assumed. This contrastswith the approach of Selverston (1980), in which heproposes to account for a similar problem (the motionin the lobster stomatogastric ganglion) by invoking alldetails of neural connectivity. I discussed the funda-mental differences in the two approaches in a commen-tary to Selverston’s paper (Harth, 1980).
2.4. Visual Perception
Vision is the most extensively studied of the senses,and, in humans, the most valuable. A much simplified
schematic diagram of the visual pathway is shown inFigure 8. Here, visual information gathered by the retinais first conveyed to neurons in the lateral geniculatenucleus (LGN), a structure in the thalamus. From thereabout a million neural fibers go to a region in the cortexcalled V1, the first cortical area devoted to vision. Aprominent feature—but one rarely discussed by physio-logists—is the fact that perhaps 10 times as many fibersdescend from V1 and send information back to the LGN.From V1 the visual information travels along severalparallel pathways to centers called V2, V3, V4, andmany others. Some 30 different visual centers havebeen identified in monkey cerebral cortex (Felleman &Van Essen, 1991). Again, practically all connectionsbetween different cortical centers are reciprocal.
The ubiquity of such loops of connectivity in thecentral nervous system blurs the distinction betweenhigher and lower cortical centers. There is no stricthierarchy of sensory processing. The ‘‘higher’’ centersreceive information from the ‘‘lower’’ centers, but thesesources are modified, augmented, and perhaps censoredby the ‘‘higher’’ centers. The prominence of these returnpathways, especially the massive fiber bundle from V1 toLGN, has been one of the great puzzles of cerebral archi-tecture. It appears that the cortex is more bent on intro-spection and confabulation than on forming an unbiasedview of the outside world.
2.5. Receptive Fields
To understand the functioning of the visual system, wewould like to know what patterns of neural activity in thebrain follow different visual stimuli. This has been untilvery recently extremely difficult to determine, and so,experimenters have often chosen to investigate a relatedproblem: What is the visual stimulus pattern to which agiven single neuron exhibits its strongest response? Thestimulus, usually obtained by trial and error, is calledthe receptive field of that cell.
1245Brains and Neural Nets
FIGURE 5. Surface of stationary netlet activities in the presenceof excitatory ( jþ ) and inhibitory ( j¹ ) input.
FIGURE 7. Activities in DFN and VFN populations of swimmingT. gilberti (after Willows, 1967).
FIGURE 6. Connectivity between neuron populations in Tritoniagilberti, according to Willows (1967).
FIGURE 8. Schematic of visual pathways from retina to visualcortex.

In a slight departure from the netlet concept, Finetteet al. (1978) applied the dynamic principles we haddeveloped for homogeneous and isotropic netlets, tostructures with random but anisotropic connectivity.The attempt here was to account for the transition fromsmall, essentially circular, visual receptive fields inmammalian retina and lateral geniculate nuclei (LGN)to the elongated fields observed by Hubel & Wiesel(1962) in area V1 of visual cortex. It is customary toassume that these elongated fields are the result of multi-ple inputs to V1 cells from LGN cells whose fields liealong an oriented line.
By contrast, Finette et al. (1978) found that locallyanisotropic connectivity within V1 can give rise toresonant activity having the shapes of Hubel andWiesel fields. Although attempts to detect such aniso-tropies in rat visual cortex (Paldino & Harth, 1977)remain inconclusive, the ease with which these elongatedresponses arise, and the comparative simplicity of theneural circuitry required, make this model still aplausible candidate for explaining cortical receptivefields.
The study of receptive fields offers an interestingexercise in solving theproblem of the inverse. One isattempting to determine a visual pattern for which aparticular neuron (that is being monitored with a micro-electrode) shows the highest response. Clearly, it isimpractical to try all possible stimuli, hence the answerinvolves some reasonable guesses on the part of theexperimenter. There is never a certainty, however, thatsome modification of the input may not have producedan even higher response.
2.6. Alopex
In the early 1970s work began in my laboratory on aprocedure for determining visual receptive fields thatwould eliminate the guesswork and biases of the experi-menter. Receptive fields were to be mapped by ahill-climbing procedure in which the cell under observationgenerates its own receptive field. We accomplished thisby using an algorithm that maximizes the cell response(Harth & Tzanakou, 1974). In this procedure, calledAlopex, an animal viewed a computer-generated visualpattern displayed on a CRT while a neuron in the visualpathway was being monitored (Figure 9). The Alopexprocedure changes all pixel intensities on the CRTbefore each successive presentation depending on thecorrelation (or anticorrelation) between the precedingchanges in pixel intensity and cell response. The algo-rithm can be represented by the simple equation
x(n) ¼ x(n¹ 1) þ g(n) þ c[x(n¹ 1) ¹ x(n¹ 2)]·[R(n¹ 1) ¹ R(n¹ 2)](1)
In eqn (1) x(n) is the intensity of theith pixel in thestimulus array in thenth iteration,g(n) a random variable(noise),c an adjustable parameter, andR(n) the strength
of the cell’s response in thenth iteration.R is thus ascalar quantity that is to be maximized in this process.This is often called acost function.
The method was applied successfully to frog retinalganglion cells (Tzanakou et al., 1979) and cat corticalcells (Micheli-Tzanakou, 1984). In Figure 10 we showthe ‘‘optimal’’ pattern obtained by the Alopex proceduretogether with the receptive field arrived at by conven-tional methods for a neuron in V1 of cat.
The ease with which the Alopex algorithm reversedthe sensory process in our studies of visual receptivefields, spawned another idea (Harth, 1976). Could itbe that similar processes take place within the brain,whereby feature-detecting neural circuits in thecortex—such as the ‘‘face detectors’’ first reported byGross et al. (1969)—enhance or generate at more peri-pheral cerebral locations, those patterns to which theyare evidently tuned? If such a mechanism existed, itwould clearly add another dimension to visual percep-tion: the visual stimuli would no longer be transportedin one direction only, along the brain’s conveyor belts,being progressively fragmented and having variousfeatures extracted in what I have called theslaughter-house paradigm(Harth, 1993). Instead, visual perception
1246 E. Harth
FIGURE 9. Determination of visual receptive field by Alopexmethod. D—CRT display; M—microelectrode; C—computer.
FIGURE 10. Cortical visual receptive field of cat cortical neurondetermined by standard procedures (a), and by the Alopexmethod (b) (from Micheli-Tzanakou, 1984).

would now appear as a self-referent process in whichthe incoming stimulus is continuously recreated andreanalyzed by higher cortical centers.
2.7. Feedback Pathways
Neural connections in the brain are generally reciprocal.If a regionA sends fibers toB, then, very likely, there willbe return paths fromB to A. Thus, we speak not only ofafferent fibers, but also of descending, orcorticofugalfibers. The massive return paths from cortical visualarea V1 to the visual thalamic relay nucleus LGN hasbeen known to neuroanatomists for nearly a century.Livingston (1958) quotes Brower (1933) who com-mented: ‘‘We accept that there is also a centrifugalside in the process of sensation, of vision, of hearing,and so on. I believe that a further analysis of the descend-ing tracts to pure sensory centers will also help physio-logists and psychologists to understand some of theirexperiences’’.
Through all the years since then this goal has beenelusive. There have been many speculations concerningthe function of these return paths, but little consensus.Rothblatt & Pribram (1972) speculated that there shouldbe pattern-specific top-down modification of sensoryinputs, and were among the first to raise the questionby what neural mechanism this could be accomplished.
Apart from the cortico-thalamic tracts, it has beenrecognized for a long time that brainstem structuresalso communicate with all afferent sensory pathways.In an early symposium on the brainstem reticular forma-tion, Livingston (1958) talks about ‘‘the reticular forma-tion … rippling its fingers in some kind of play of…perceptive… composition’’.
In more recent work, Yingling & Skinner (1977)report that neurons of the nucleus reticularis thalami(NRT), which is interposed between thalamic sensoryrelay nuclei and sensory cortex, are inhibitory onthalamic relay cells. This inhibitory feedback is topo-graphically organized between NRT and thalamic cells,but the NRT cells, in turn, receive ‘‘non-selective,generalized’’ inhibitory input (Steriade et al., 1986)from the mesencephalic reticular formation (MRF).
Thus, if cortico-thalamic pathways and thalamic inputfrom the brainstem affect the pattern of incoming sensorystimuli—and it is difficult to avoid this conclusion—then these feedbacks are very likely to be feature-specific.
This reciprocity of connections in sensory pathways isalmost universal and has given rise to many speculations.Von Holst & Mittelstaedt (1950) speak of aprinciple ofreafference, by which the brain may generate its owninput without sensory stimulation. Edelman (1992)points to there-entry of information passed back andforth between sensorymaps. The resulting reverberationsare both conservative and creative, in the sense that theymay preserve a stimulus long enough to be contemplated
by ‘‘higher’’ cortical centers, and perhaps committed tomemory, and creative in that neural activity patternsevolveand link sensory information to past experiencesin novel combinations. Edelman believes that the modu-lation of images through extensive cross-talk betweensensory centers bestows a kind of unity on sensoryexperience.
The problem of preserving spatial mappings was dis-cussed by Harth et al. (1970a) in connection with lateralinhibition. This phenomenon is as ubiquitous in sensorysystems as the reciprocity between maps. What effectdoes lateral inhibition have on the information that ispassed back and forth between two maps, or passedsequentially from one map to the next?
It turned out that under repetitive mapping with aGaussian spread function, a sharply defined spot willrapidly blur and melt into background noise.
In Figure 11(a) we observe the repeated mappings ofan initial delta function by a Gaussian spread functionthat results in the expected progressive deterioration ofthe signal. In Figure 11(b) the spread function is alteredby adding a negative Gaussian of smaller amplitude butgreater width, which mimics the well known character-istic spread function for lateral inhibition. This results ina remarkable phenomenon. After some initial changes,the signal stabilizes and becomes essentially insensitiveto further blurring.
A graphic confirmation of this principle is shown inFigure 12. Here, a black spot is imaged repeatedly by aphotocopier whose spread function also showed aninhibitory surround. The succession of mappings at firstintroduced increasing amounts of noise. But, instead ofeventually overwhelming the signal, the pattern stabilizesand soon becomes immune to further corruption. Theinformation is now carried by a stable pattern of noise.
1247Brains and Neural Nets
FIGURE 11. Successive mappings of a delta function through (a)a Gaussian point spread function, (b) through a function withpositive Gaussian center and negative Gaussian surround.

In a subsequent paper (Harth & Pertile, 1972), weshowed that there exists a class ofideal mapping func-tions that would preserve information for an infinitenumber of successive mappings, and that one of thesehas the form(sinkx)=x, which is similar to the spreadfunction in lateral inhibition except for having furthernegative and positive peaks of diminishing amplitudes.
I believe that this form of signal stabilization due tolateral inhibition plays a role in most any sensory system,together with mechanisms for altering the signal insignificant ways, as proposed for example by Edelman(1992). The first must exist for the second to function.
2.8. Mental Images
Humans are able to construct visual images in theabsence of visual input. We speak ofmental images,and the controversy has been whether these imageshave the spatio-temporal characteristics of true visualstimuli, or whether they areneural codesappearing athigh level cortical areas. In the first alternative, we wouldexpect them to appear at more peripheral regions in thevisual pathway where neural representations are stillretinotopically organized.
This appears to be the case. In a series of PET scanstudies Kosslyn et al. (1993) have shown that mentalimagery is accompanied by marked activity in V1, andthat the spatial extent of the activity depends on the sizeof the object imagined. They propose that mentalimagery possesses characteristics resembling images ofreal objects, and that these images form the inputs tohigher cortical levels as in visual perception. In ananalogous description, Llina´s et al. (1994) call thebrain a ‘‘closed system capable of self-generated
activity’’ and a ‘‘reality-emulating system’’, thoughthey take the emulation to occur at higher corticallevels as a result of temporal scanning mechanisms.This point is admittedly still somewhat in dispute,since some data suggest that V1 activity is not essentialfor conscious cognition.
3. THE SKETCHPAD MODEL
The ideas presented here are taken chiefly from myrecent work (Harth, 1995, 1996a), but incorporatemany elements of the earlier papers.
The notion of internal sketchpads has its origin in theobservation that thought processes, especially creativeones, often involve a cyclic interplay between cerebraland external events: the artist sketches before puttingpaint to canvas, the composer needs a piano to composehis music, and all of us occasionally talk to ourselves.
Why do we bother to externalize for our own perusalsomething we already know? The answer is, I believe,not hard to find. Our nervous system finds it difficult tostore complex images of, say, a painting or a symphony,in a way that would allow us to contemplate and judgethe work in its entirety. Hence, the creation of such worksrequires temporary external storage of the nascent idea,sensory access to the image, and the ability to modify,elaborate and to perfect. This can be described as a hill-climbing procedure, in which the changes in the externalimage, or artifact, are guided by internal aesthetic orother criteria. Such processes are self-referent andinvolve positive feedback.
I have proposed thatmental images, though morerestrictive in their capacity to hold information, servefunctions that are very similar to the artist’s sketchpador the composer’s piano.
I came across this same idea in a most unlikely source.Tran Duc Thao (1984) quotes from Karl Marx’sDasKapital:
… what distinguishes the worst architect from the best ofbees is this, that the architect raises his structure in theimagination before he erects it in reality.
If, indeed, top-down mechanisms in the brain are togenerate ‘‘peripheral’’ neural activity patterns that havethe spatio-temporal characteristics of visual stimuli, thenwe may suppose that they can function as trial patterns,to be viewed and further developed, very much likepatterns we put on a sketchpad.
In developing this idea further, I proposed (Harth,1976) that a feedback to the LGN act as acost functionthat guides a pattern-generating optimization process. Ihave taken the example of the Alopex algorithm and ourreceptive field studies (Tzanakou et al., 1979) as generalguide for this model. Accordingly, the feedback wouldhave to be aglobal function, i.e., one that carries a simplefigure of merit expressing the degree to which the
1248 E. Harth
FIGURE 12. Successive mappings by a photocopier. (a) original;(b) 4th copy; (c) 13th copy; (d) 15th copy.

peripheral pattern satisfies some internal requirements.In the limit, a simple scalar quantity would suffice. Ibelieve this assumption of aglobal re-entry into thesensory pathway is somewhat related to Edelman’s(1992) concept ofglobal mapping.
In this first attempt (Harth, 1976), I viewed the LGNas the site of the internal sketchpad, and the cortico-thalamic feedback pathways as the carriers of the costfunction.
There are several difficulties with this scheme. Ratherthan carryingglobal information, the cortico-thalamicpathways are specific, connecting cells in V1 with theirretinotopic equivalents in the LGN (Yingling & Skinner,1977). These are more likely to cause reverberations, i.e.,a temporary stabilization of the stimulus pattern, whilethe ‘‘nonselective, generalized’’ input from MRF toNRT has more of theglobal qualitiesof a cost function.
3.1. Brainstem Control of Sensory Messages
The effect of brainstem neurons on thalamocortical trans-mission was considered in a revised sketchpad model(Harth & Unnikrishnan, 1985; Harth et al., 1986, 1987,1990).
The diagram in Figure 13 illustrates the anatomicalfeatures on which the model is based. Amapof thalamicrelay cells (G) is connected reciprocally to visual cortexV1. These connections are excitatory, and send exci-tatory collateral inputs to PGN cells (P). These areinhibitory on thalamic relay cells and are, in turn, inhib-ited by fibers from the MRF. Theglobal character ofthe latter is here represented by a single ascending line.
On the basis of this diagram, one would expect that aretinal input pattern would be quickly extinguished bythe PGN cells, except when strong brainstem inputinhibits the PGN cells. With the addition of randomnoise and the iterative character made possible by theretinotopic feedback from V1, this circuit is capable ofcarrying out the optimization algorithm defined byeqn (1).
3.2. Sketchpad Simulations
I present here one of the many results of computer simu-lations reported in previous publications (e.g., Harth,1995). In Figure 14, boxA represents an optimizer thatcarries out an algorithm such as the one defined by eqn(1), using the scalar variableF ascost function, and dis-playing the resulting pattern in an iterative fashion.These patterns areviewedby four feature analyzersB,whose responsesR1 to R4 reflect the similarity betweenthe observed pattern and the fixed template assigned to
1249Brains and Neural Nets
FIGURE 13. Schematic of connectivity in visual pathway,showing thalamocortical loop with collateral inputs to PGN,and PGN inhibition of LGN relay cells. G—thalamic relayneurons; P—neurons in PGN; C—cortical cells.
FIGURE 14. Schematic of computer simulation of sketchpadmechanism. A—hillclimbing algorithm modifying a peripheralpattern; B—array of feature analyzers producing responsesR1 ¹ R4; C—a non-linear integrator producing a cost function F .
FIGURE 15. Templates used in sketchpad simulation.

that analyzer. The analyzer responses are summed non-linearly in C to make up the cost functionF.
Figure 15 shows four such templates, and Figure 16(a)an initial state chosen for the system. This pattern isalmost random; it was obtained by superimposingrandom noise on the Einstein template. With the applica-tion of the hillclimbing algorithm, the pattern changedgradually and eventually became almost identical withthe Einstein template [Figure 16(b)].
Figure 17 shows the evolution of the analyzerresponses and the combined responseF. Note that theresponseR2 of the Einstein analyzer is initially higherthan that of the others, and begins to increase sharplyafter an initial period of uncertainty. The processspeeds up as the cost functionF becomes dominatedalmost exclusively byR2.
An optimization algorithm of the type shown here isthe most general way in which peripheral pattern genera-tion can be achieved in the sensory system by top-downcontrol. It is also the one requiring the least complexityof the necessary neural circuitry. The known connectiv-ity between LGN, PGN, V1, and BRF (Figure 13)appears to be eminently suited to carry out the necessaryalgorithm, but similar processes may take place at highervisual areas, as well as in other sensory modalities.
3.3. Summary of Model
Mental processes are appropriately calledreflective, andconcomitant neural activity is cyclic, self-referent. Whatdistinguishes the thinking brain from computers, andfrom most artificial neural networks, is the fact that inthought processes there are no final, oroutput states.
In summarizing the model, let me begin, arbitrarily, byconsidering the extensive, scattered activities in corticalareas, including sensory analyzers, association areas, and‘‘higher’’ areas such as theworking memory(Baddely,1986), or theglobal workspace(Newman & Baars,1993). These activities are in part elicited by currentsensory stimuli, in part due to entirely endogenous pro-cesses, including neural noise. Chaotic processes maywell be involved, allowing minute fluctuations to causemacroscopic activity changes (Harth, 1983; Freeman,1987; Freeman & Barrie, 1994).
From all this scattered activity, aglobal function isconstructed and, by way of brainstem nuclei, relayedto visual and/or other sensory projection areas. A hill-climbing process will then strengthen whatever featureshappen to dominate the global cost function.
There will be competition between such corticalfancyand ongoing sensory stimuli, with the latter generallydominating the process, except in the cases of hallucina-tions and dreams, and such creative activities we referredto above.
The transformation from the extended activities in theassociation areas and working memory to specific mentalimages may be likened to the collapse of a wave functionin quantum mechanics, although I hesitate to refer to thatmuch overused term. No quantum mechanical effect ishere implied.
I want to stress two significant characteristics of themodel.
1. Unification of sensory elements and stored memoriesand associations is achieved by reciprocal linking, inwhich those sensory elements are enhanced which
1250 E. Harth
FIGURE 16. (a) Initial stimulus pattern; (b) pattern after hillclimb-ing convergence.
FIGURE 17. Responses R1 ¹ R4 of feature analyzers and costfunction F in sketchpad simulation.
FIGURE 18. Periodic (ca. 30 Hz) post-stimulus histogram in frogretinal ganglion cell (from Stiles et al., 1985).

cause resonant activity at higher levels. (I leave openfor the time being the question of how a cost functionis constructed from what goes on in the workingmemory, and how this is conveyed to the brainstemreticular formation.)
2. The self-referent processes in which higher corticallevels observetheir own reflections, make unneces-sary the introduction of the pesky homunculus.
3.4. Limitations of the Model
In a critique of the sketchpad model, Francis Crick(private communication) has pointed out that hillclimb-ing processes such as Alopex are inherently slow, andcan therefore not account for mental phenomena likepattern recognition. Although some receptive fieldexperiments (Tzanakou et al., 1979) produced significantconvergence in less than 100 iterations, the simulationstudies (Harth et al., 1990) extended over thousands ofiterations. This would translate into many seconds oreven minutes of neural processing time. Another objec-tion to hillclimbing processes in general is their frequentfailure to arrive at theglobal maximum by becomingtrapped on secondary peaks.
I commented on these objections as follows (Harth,1996a):
1. In the computer simulations (Harth, 1980) the initialpatterns were either very noisy [Figure 16(a)] or com-pletely random. Perception would, in general, have todeal with much less ambiguous inputs.
2. Unlike the recognition of unambiguous stimuli, manycognitive processes are slow, This is especially truefor creative processes, for which the sketchpad modelis most applicable.
3. Convergence times may be shortened considerably ifwe allow a hierarchy of feature analyzers togetherwith a similar hierarchy of hillclimbing algorithms.
4. Recognition of ambiguous stimuli may require only aslight increase in global cost function, rather thancomplete convergence.
5. Other processes, such as matching activity in thethalamus–PGN–cortex–brainstem complex that wassuggested by Taylor (1992, 1996), Newman et al.(1996), and Baars (1996), may contribute to solvingthe inverse problem.
6. The random variable used in the hillclimbing proce-dure (the quantityg in eqn (1)) will tend toshaketheprocess off secondary maxima. On the other hand, weknow that thought processes are not unerringly goal-seeking, but that ‘‘getting sidetracked’’ is a frequent,and probably useful feature of mentation.
4. CONSCIOUSNESS
How does subjective awareness arise in the brain, and howcan we understand it in terms of a network of neurons?
Nobody likes to define consciousness, but everybodyexpects others to define it. I succumbed to this pressure,fully aware of the pitfalls of any such attempt, when Iwrote: ‘‘Consciousness is… a feelingof subjective andperceptual unity. It is as though a unique, but probablyfictitious center, theI, were the recipient of all thesensory inputs, past and present, and as though all mes-sages pertaining to an object of sensory reality werereunited somewhere in the mind to constitute a con-sciously perceived facsimile of the same reality’’(Harth, 1995).
4.1. The Binding Problem
One of the attributes I wanted to stress is thatconscious-ness is a feeling. The brain is a network of neurons, and itfeels. Whether a network of artificial neurons can alsohave feelings is one of the hotly debated issues. The otherpoint refers to what is now generally called thebindingproblem:what is it that confers unity, meaning, and theproperty of beingowned to the activities of countlessneurons that are scattered over large areas of the nervoussystem? Why is this thoughtmy thought, as WilliamJames mused? [I want to avoid here the much over-worked dichotomy of the problem into aneasyand ahard question(Chalmers, 1996)].
Binding poses an almost impossible task for the brain.It appears at first that the contents of consciousnesswould have to be brought together both spatially andtemporally to form a single, localized, and instantaneousglobalexpression. It is not clear, how anything other thana single bit of information can be so compressed.
If, on the other hand, the information is spreadspatially and/or temporally, we are left with the feelingthat—unless there is another observer, ahomunculus, tointegrate these data—we are back to the simplistic asser-tion that ‘‘a thought issimply a constellation of activeneurons’’ containing either a spatial or temporal code.(Whenever ‘‘simply’’ or ‘‘ nothing but’’ are used in adefinition, I suspect that a great deal of complexity isbeing swept under the rug.)
True spatialbinding appears to be out. Physiologistshave looked in vain for aconvergence zoneat the highestcortical levels such as the prefrontal cortex, althoughconvergence of a sort exists in the brainstem reticularformation through which all afferent and efferent infor-mation must pass.
This leaves us with temporal binding. Eckhorn et al.(1988) proposed that coherent oscillations of around40 Hz are the ‘‘feature linking mechanism’’ inthe brain. Such periodic oscillations were found in thevisual system of cats and monkeys (Doty & Kimura,1963), and of frogs (Stiles et al., 1985). Figure 18shows a post-stimulus spike histogram obtained in mylaboratory for frog retinal ganglion cells.
The idea of binding by temporally linking scatteredneural activities was taken up by Crick & Koch (1990)
1251Brains and Neural Nets

and Llinas et al. (1994). I have pointed out (Harth, 1995)that the difficulty in all these schemes comes from theassumption of a final, oroutput statearising spon-taneously or triggered by sensory stimuli. To makesuch final states account for consciousness, somehowconflicts with the notion of locality in physics. Eachneuron knows only what transmitters are coming acrossits synapses (if we can speak of a single cell asknowinganything). How, then does global knowledge arise fromlocal ignorance? Does neuronA know that neuronsB, C,andD are alsofiring at this moment? If not,whoor whatdoes? This problem has occasioned some investigators toresort to some form of non-local quantum mechanics(Penrose, 1994; Hameroff & Penrose, 1996).
In connection with the models of temporal bindingby phase-locked 40 hz activities, Freeman & Barrie(1994) pointed out that cortical neurons, unlike peri-pheral sensory neurons, generally do not exhibitperiodic activity, Instead, ‘‘local neighborhoods… canbe viewed as an equivalence class, in which the burdenof firing is rotated at random among the members’’. Thisis reminiscent of the dynamics of neuralnetlets(Harthet al., 1970b).
4.2. The Computer Analogy
The whole idea of binding derives, I believe, from theerroneous analogy between brains and computers, whichassigns in both cases initial and final states with inter-vening computation. In the case of computers these statesare written in a code, designed and read by an externalintelligence. In the case of the brain, the sensory stimuliare not really input states, since—as we have seen—theyare subject to modification by the very centers for whichthey are intended. It is even less appropriate to speak ofoutput states because activity in the cortex is contin-uous, widespread, and ever-changing. Nor is meaningassigned to these activities by some externaluser. Wecan simulate many brain functions on a computer, butthe brain does notcomputeany more than a telescopecomputes the trajectory of light passing through it (Harth,1986).
In my sketchpad model, by contrast, no final corticalstates can be defined, since the cortico-thalamic–reticular system is self-referent. It appears also, that thecortical activities following a given sensory stimulusare not invariant relative to that stimulus, but changewith every subsequent presentation as a function of theever-changing store of memories and associations(Freeman & Barrie, 1994). The picture I have drawn isperhaps complementary to Taylor’srelational model(Taylor, 1996), in which sensory inputs derive theirmeaning and significance from cortically storedmemories and associations. Here, these higher corticalactivities play a role in conscious perception throughtheir interaction with and modification of sensoryprimitives at peripheral levels.
5. REDUCTIONISM RE-EXAMINED
The problem of consciousness further raises these funda-mental questions: Is the problem amenable to a scientificapproach? and Can we apply the methodology of reduc-tionism? Are the two questions perhaps the same? Iaddressed that problem at a recent conference (Harth,1996b).
Much has been written about the scientific method, butthe concept of reductionism appears to be at its core. Tophysicists this term has a lucid meaning that has to dowith the flow of causality in the inanimate world, whichis from the small to the large, orbottom-up. Hence, tounderstand a phenomenon, we must go the other way,reducing it to its more elementary constituents. Weunderstand the formation of an ice crystal, for example,by examining the properties of water molecules and theirmutual interactions. The water molecule itself, or itsatomic constituents, are the same whether part of anicicle or a cloud of steam. There are, to be sure, situationsin which large-scale phenomena affect events in themicroworld—the cold weather causing the water tofreeze—but the explanation of that is againreducedto molecular events.
The enormous success of what I will callclassicaloratomistic reductionism—explaining the large by lookingat the small—suggests to many that the method shouldbe universally applicable, even to its ultimate challenge:consciousness. Thus, Francis Crick (1994) states that,
You, your joys and sorrows, your memories and your ambi-tions, your sense of personal identity and free will, are infact no more than the behavior of a vast assembly of nervecells and their associate molecules.
In a similar vein, the philosopher John Searle (1995)writes,
… our entire mental life is caused by the behavior ofneurons, and all they do is increase or decrease their rateof firing.
And this from the physicist Steven Weinberg (1995):
Phenomena like mind and life do emerge… the rules theyobey are not independent truths, but follow from scientificprinciples at a deeper level… The reductionist world viewrules out other sorts of understanding.
In 1895 Sigmund Freud argued for aProject for aScientific Psychology.With true reductionist fervor heproposed that human actions and emotions were to beunderstood by studying the underlying neural processes.He soon abandoned that approach.
Now, a hundred years later,emergentismis still verypopular among neuroscientists and philosophers.Mental phenomena are viewed asemergingout of thestaggering complexity of the human brain, but theirroots are sought in the electro-chemical pulsations ofthe nerve cell.
1252 E. Harth

But the where and how of this emergence hadremained obscure. Although we have learned muchabout brain function and neural dynamics, no satisfactorycausal connection has been established between brainprocesses and the manifestations of the mind. The bestwe have been able to do is point to some neural activitiesthat are observed when we experience certain emotionsor perform certain mental tasks. Physiologists speak oftheneural correlatesof mental phenomena. The findingof such correlates has been made possible by modernnon-invasive techniques such as PET scans, in whichshortlived radioactive substances trace enhanced bloodflow and, presumably, heightened neural activity. Thesubject is fully conscious and alert, and can be given avariety of mental tasks. But correlation alone does notproduce understanding. Apart from the fact that suchcorrelations are evanescent, and at best approximate,no such finding produces the much desiredAha! reaction.
Our frustration has to do, I believe, with the fact thatwe are looking for understanding in the wrong place. Acursory look at life processes will convince us that we arefacing here a situation that differs fundamentally fromwhat goes on in the world of physics. Where top-downcontrol was virtually absent in inanimate nature—or atleast transparently physical in its elementary details—itplays a dominant role where life is involved.
The downward sweep of causality is nowhere asapparent as in the interplay between an individual lifeform, its DNA blueprint, and its role in the ecologicalstruggle for existence. We may explain most physicalproperties of the individual through methods of atomisticreductionism, going all the way back to its submicro-scopic blueprint, the DNA molecule and studying thecopying, reading, and expressing of the genetic mes-sages. But if we wish to understandwhya given genomehas a certain nucleotide sequence, we must look farabove the molecular level. Unlike theemergentphenomenon of the ice crystal, the genome structure isa devolved principle. It was shaped in the battle forsurvival by complex ecological factors extending overeons of time.
The complexity of the situation increases drasticallywith the introduction of mental processes into the causalchain. Human speech requires precise timing in the acti-vation of many muscle groups, but if we want to under-stand what is being said and why it is being said, we mustanalyze theglobal sources (semantic coding, culturalbackground, motivation, etc.) rather than the neuronalfiring patterns. Similarly, the detailed neural processesin dreaming are under top-down control by memorystructures that involve the whole gamut of personalexperience and values. To explain a given human activityother than a simple reflex, we must invoke phenomenathat are themselves emergent principles of a higher order.If, instead, we tried delving into details of the underlyingcortical neurodynamics, reducing behavior to the actionof neurons that can only ‘‘increase or decrease their rate
of firing’’, we would soon be lost in a causal jungle thatis both impenetrable and unenlightening.
The absurdity of pushing atomistic reductionismbeyond its useful range becomes more and more apparentas we further increase the complexity of the system. In asociety of a few hundred million individuals the fourelementary interactions recognized in physics are oflittle help in tracing and solving societal problems.What guides the dynamics of a nation—what reallymovesit—are non-physical entities: political philoso-phies, the economy, the appeal, persuasion (and dupli-city) of its leaders. Clearly, we are no longer able toconstruct seamless causal chains made up entirely ofelementary physical interactions, though they undoubt-edly exist.
I believe that the concept of reductionism has to bebroadened. In fact, the Latin root of the word merelymeans tolead back. The reductio ad absurdumleadsus back to the original statement (and proves it wrong);it does not dissect an argument into mutually contradic-tory elements.
Mental activities, then, are what I have calleddevolvedphenomena. They are—in the sense of Taylor’s (1996)relational mind—tied inextricably to extensive memorystructures and associations that, in turn, are derived froma lifetime of interaction with our environment. Searchingthrough the underbrush of neural connectivity mayuncover regions of correlated activity, or even mechan-isms, but will not satisfy our curiosity regarding thenature of thought. If mental phenomena are under top-down control, then, to understand them, we must look athigher levels.
This new type ofnon-atomisticreductionism other-wise conforms to all requirements of a scientificapproach. It is just immeasurably more complex thanwhat the ‘‘nothing-but’’ school has been trying to tellus. Our one-hundred-billion-neuron brains with theirtrillions of modifiable synapses, their largely unexploredpharmacopoeia of neurotransmitters, neuromodulatorsand neurohormones, are the necessary instruments onwhich the mental symphonies are played, but therewould be little music without the greater complexity ofthe society of humans with its language, traditions, its art,and its history. This shows us also how very much furtherwe have to go before we can simulate by computersthe true nature of consciousness.
REFERENCES
Allanson, J. T. (1956). Some properties of a randomly connected neuralnetwork. In C. Cherry (Ed.),Information theory(pp. 303–313).New York: Academic Press.
Anninos, P. A., Beek, B., Csermely, T. J., Harth, E., & Pertile, G.(1970). Dynamics of neural structures.Journal of TheoreticalBiology, 26, 121–148.
Asratian, E.A. (1965).Compensatory adaptations, reflex activity,and the brain. English transl. by S.A. Corson, Oxford: PergamonPress.
1253Brains and Neural Nets

Baars, B. J. (1996). Sensory projection areas of the cortex may provide aconscious global workspace for a massively distributed unconsciousbrain. Paper presented at Tucson II conference: Toward a science ofconsciousness.
Baddely, A. D. (1986).Working memory. London: Oxford UniversityPress.
Beurle, R. L. (1959). Storage and manipulation of information in thebrain. I.E.E. Journal, 5, 75–82.
Brower, B. (1933). Centrifugal influence on centripetal systems in thebrain.Journal of Nervous & Mental Diseases, 77, 621.
Chalmers, D. J. (1996). Facing up to the problem of consciousness.In Hameroff et al. (Eds.),Toward a science of consciousness(pp. 5–28). Cambridge, MA: MIT Press.
Csermely, T. J., Harth, E. & Lewis, N. S. (1973). The netlet theory andcooperative phenomena in neural nets.Journal of Dynamic Systems,Measurement and Control, Transactions of the ASME, 315–320.
Crick, F. H. C. (1994).The astonishing hypothesis. New York:Scribners.
Crick, F. H. C., & Koch, C. (1990). Towards a neurobiological theory ofconsciousness.Seminars in Neuroscience, 2, 263–275.
Doty, R. W., & Kimura, D. S. (1963). Oscillatory potentials in the visualsystem of cats and monkeys.Journal of Physiology (London), 168,205–218.
Eckhorn, R., Bauer, R., Jordan, W., Brosch, M., Kruse, W., Munk, M.,& Reitboeck, H. J. (1988). Coherent oscillations: a mechanism offeature linking in the visual cortex?Biological Cybernetics, 60,121–130.
Edelman, G. M. (1992).Bright air, brilliant fire. New York: BasicBooks.
Felleman, D. J., & Van Essen, D. C. (1991). Distributed hierarchicalprocessing in the primate cerebral cortex.Cerebral Cortex, 1,1–47.
Finette, S., Harth, E., & Csermely, T. J. (1978). Anisotropic connectiv-ity and cooperative phenomena in visual cortex.BiologicalCybernetics, 30, 231–240.
Freeman, W. J. (1987). Simulation of chaotic EEG patterns with adynamic model of the olfactory system.Biological Cybernetics,56, 139–150.
Freeman, W. J., & Barrie, J. M. (1994). Chaotic oscillations and thegenesis of meaning in cerebral cortex. In G. Buzsa´ki et al. (Eds.),Temporal coding in the brain. Berlin–Heidelberg: Springer Verlag.
Gross, C. G., Bender, D. B., & Rocha-Miranda, E. C. (1969). Visualreceptive fields in inferotemporal cortex of the monkey.Science,N.Y., 166,1303–1305.
Hameroff, S. R., & Penrose, R. (1996). In S. R. Hameroff,A. W. Kasniak, & A. C. Scott (Eds.),Toward a science of con-sciousness(pp. 507–540). Cambridge, MA: MIT Press.
Harth, E. (1966). Brain models and thought processes. InE. R. Caianiello (Ed.),Automata theory(pp. 201–217). NewYork: Academic Press.
Harth, E. (1976). Visual perception: a dynamic theory.BiologicalCybernetics, 22, 169–180.
Harth, E. (1980). Must neural mechanisms be Newtonian?TheBehavioral and Brain Sciences, 3, 550–551.
Harth, E. (1983). Order and chaos in neural systems: an approach to thedynamics of higher brain functions.IEEE Transactions on Systems,Man, and Cybernetics, SMC-13,782–789.
Harth, E. (1986). Does the brain compute?Behavioral and BrainSciences, 9, 98–99.
Harth, E. (1993).The Creative loop. Reading, MA: Addison Wesley.Harth, E. (1995). The sketchpad model.Consciousness & Cognition, 4,
346–368.Harth, E. (1996a). Selfreferent mechanisms as the neural basis of
consciousness. In S. R. Hameroff, A. W. Kaszniak, & A. C. Scott(Eds.), Toward a science of consciousness(pp. 611–631).Cambridge, MA: MIT Press.
Harth, E. (1996b). The pros and cons of mind-brain reductionism.Paper presented at Tucson II conference: Toward a science ofconsciousness.
Harth, E., Beek, B., Pertile, G., & Young, F. (1970). Signal stabilizationand noise suppression in neural systems.Kybernetik, 7, 113–122.
Harth, E., Csermely, T. J., Beek, B., & Lindsay, R. D. (1970). Brainfunctions and neural dynamics.Journal of Theoretical Biology, 26,93–120.
Harth, E., & Edgar, S. L. (1967). Association by synaptic facilitation inhighly damped neural nets.Biophysical Journal, 7, 689–717.
Harth, E., Lewis, N. S., & Csermely, T. J. (1975). The escape ofTritonia. Dynamics of a neuromuscular control mechanism.Journalof Theoretical Biology, 55, 201–228.
Harth, E., Pandya, A.S., & Unnikrishnan, K.P. (1986). Perception as anoptimization process.Proceedings of IEEE Computer Society Con-ference on Computer Vision and Pattern Recognition(pp. 662–665). IEEE Computer Society, Washington, DC.
Harth, E., Pandya, A. S., & Unnikrishnan, K. P. (1990). Optimizationof cortical responses by feedback modification and synthesis ofsensory afferents. A model of perception and REM sleep.Conceptsin Neuroscience, 1, 53–68.
Harth, E., & Pertile, G. (1972). The role of inhibition and adaptation insensory information processing.Kybernetik, 10, 32–37.
Harth, E., & Tzanakou, E. (1974). Alopex: a stochastic method fordetermining visual receptive fields.Vision Research, 14, 1475–1482.
Harth, E., & Unnikrishnan, K.P. (1985). Brainstem control of sensoryinformation: a mechanism for perception.International Journal ofPsychophysiology, 3, 101–119.
Harth, E., Unnikrishnan, K.P., & Pandya, A.S. (1987). The inversion ofsensory processing by feedback pathways: a model of visual cog-nitive functions.Science, 237,184–187.
Hebb, D.O. (1949).The organization of behavior. New York: Wiley.Von Holst, E., & Mittelstaedt, H. (1950). Das Reafferenzprinzip.
Naturwissenschaften, 37, 464–476.Hubel, D. H., & Wiesel, T. N. (1962). Receptive fields, binocular inter-
action and functional architecture in the cat’s visual cortex.Journalof Physiology, London, 160,106–154.
Kosslyn, S.M., Alpert, N.M., Thompson, W.L., Maljkovic, V., Weise,S.B., Chabris, C.F., Hamilton, S.E., Rauch, S.L., & Buonanno, F.S.(1993). Visual mental imagery activates topographically organizedvisual cortex: PET investigations.Journal of CognitiveNeuroscience, 5, 263–287.
Livingston, R. B. (1958). Central control of afferent activity. InH.H. Jasper, L. D. Proctor, R. S. Knighton, W. C. Moshay, &R. T. Costello (Eds.),The reticular formation of the brain(pp. 177–185). New York: Little Brown.
Llinas, R., Ribary, U., Joliot, M., & Wang, X.-L. (1994). Content andcontext in temporal thalamocortical binding. In Buzsa´ki et al.(Eds.),Temporal coding in the brain.Berlin–Heidelberg: SpringerVerlag.
McCulloch, M. C., & Pitts, W. (1943). A logical calculus of the ideasimmanent in nervous activity.Bulletin of Mathematical Biophysics,5, 115–133.
Micheli-Tzanakou, E. (1984). An automated system for visual studies.Journal of Medical Systems, 8, 507–519.
Minsky, M., & Papert, S. (1969).Perceptrons: an introduction to com-putational geometry. Cambridge, MA: MIT Press.
Newman, J., & Baars, B.J. (1993). A neural attentional model for accessto consciousness: a global workspace perspective.Concepts inNeuroscience, 4, 255–290.
Newman, J., Baars, B.J., & Taylor, J.G. (1996). A neurocognitive modelfor consciousness and attention. Paper presented at Tucson II con-ference: Toward a science of consciousness.
Paldino, A. M., & Harth, E. (1977). Some quantitative results on Golgiimpregnated axons.Journal of Comparative Neurology, 176,247–261.
Penrose, R. (1994).Shadows of the mind. London: Oxford UniversityPress.
Rochester, N., Holland, J.H., Haibt, L.H., & Duda, W.L. (1956). Testsof a cell assembly theory of the action of the brain using a large
1254 E. Harth

digital computer.IRE Transactions on Information Theory, PGIT-2,80–93.
Rosenblatt, F. (1958). The perceptron, a probabilistic model for infor-mation storage and organization in the brain.Psychological Review,62, 386–408.
Rothblatt, L., & Pribram, K. H. (1972). Selective attention: input filteror response selection.Brain Research, 39, 427–436.
Rumelhart, D. E., Hinton, G. E., & Williams, R. S. (1986). Learn-ing representations by back-propagating errors.Nature, 323,533–536.
Searle, J. R. (1995, November 2). The mystery of consciousness.TheNew York Review.
Selverston, A. (1980). Are central pattern generators understandable?Behavioral and Brain Sciences, 3, 535–571.
Shaw, G. L., Harth, E., & Scheibel, A. B. (1982). Cooperativity andbrain function: assemblies of 30 neurons.Experimental Neurology,77, 324–358.
Steriade, M., Domich, L., & Oakson, G. (1986). Reticularis thalamineurons revisited: activity changes during shifts in states ofvigilance.Journal of Neuroscience, 6, 68–81.
Stiles, M., Tzanakou, E., Michalak, R., Unnikrishnan, K.P., Goyal, P.,& Harth, E. (1985). Periodic and nonperiodic burst responses in frog
(Rana pipiens) retinal ganglion cells.Experimental Neurology, 88,176–197.
Taylor, J. G. (1992). Towards a neural network model of the mind.Neural Network World, 2, 797–812.
Taylor, J. G. (1996). The relational mind. Paper presented at Tucson IIconference: Toward a science of consciousness.
Tran Duc Thao, (1984).Investigations into the origin of language andconsciousness. Dordrecht: D. Reidel.
Tzanakou, E., Michalak, R., & Harth, E. (1979). The Alopex process:visual receptive fields by response feedback.BiologicalCybernetics, 35, 161–174.
Weinberg, S. (1995, October 5). Reductionism redux.New York Reviewof Books, pp. 3–9.
Willows, A. O. D. (1967). Behavior elicited by stimulation of single,identifiable brain cells.Science, 157,570–574.
Wong, R., & Harth, E. (1973). Stationary states and transients in neuralpopulations.Journal of Theoretical Biology, 40, 77–106.
Yingling, C. D., & Skinner, J. E. (1977). Gating of thalamic input tocerebral cortex by nucleus reticularis thalami. In J. E. Desmedt(Ed.), Attention, voluntary contraction and event-related cerebralpotentials (pp. 70–96)Prog. clin. Neurophysiol.vol. 1. Basel:Karger.
1255Brains and Neural Nets

1997 SPECIAL ISSUE
The Transition from Automatic to Controlled Processing
Jeffrey A. Gray,1 Catalin V. Buhusi2 and Nestor Schmajuk2
1Department of Psychology, Institute of Psychiatry, De Crespigny Park and2Department of Psychology, Duke University
(Received12 September1996;accepted12 March 1997)
Abstract—The transition from automatic (unconscious) to controlled (conscious) processing is described in terms of aneural network model of classical conditioning (Schmajuk et al., 1996). In the framework of the network, an environ-mental stimulus is processed in controlled or conscious mode when Novelty and attention to the stimulus are large, andin automatic or unconscious mode otherwise. In the model, indirect dopamine (DA) agonists, such as amphetamine ornicotine, enhance the DA representation of Novelty, thereby increasing attention and engaging conscious processing ofenvironmental stimuli. By contrast, DA receptor antagonists, such as haloperidol, reduce the DA representation ofNovelty, thereby decreasing attention, and engaging unconscious processing of the stimuli.q 1997 Elsevier Science Ltd.
Keywords—Consciousness, Dopamine, Latent inhibition, Nucleus accumbens, Schizophrenia, Controlled processing,Automatic processing, Neural network.
1. INTRODUCTION
The starting point for the speculations contained in thispaper lies in the cognitive abnormalities characteristic ofthe acute phase of schizophrenic psychosis. Patients inthis stage of the illness typically display a variety ofpositive symptoms (Crow, 1980), such as hallucinations,delusions, fragmentation, intensification or loss of selec-tivity of attention, etc. These symptoms have been inter-preted as reflecting a breakdown in the normal control ofcurrent information processing by stored regularitiesof previous input (Hemsley, 1987, 1993, 1994; Grayet al., 1991a). One consequence of this breakdown isthat processing which, in normal individuals, takesplace in automatic, non-conscious mode has to be dealtwith in controlled or conscious mode (Schneider andShiffrin, 1977; see discussion in Gray et al., 1991b).We have for some years been studying the disruptionof latent inhibition (LI; see below) as a relativelysimple experimental model of Hemsley’s (1987) postu-lated breakdown in the control of current informationprocessing by stored regularities of previous input(Gray et al., 1991a, b, 1995), and have recently devel-oped a neural network model of LI (Schmajuk et al.,1996; Schmajuk, 1997). We speculate here how LI, theneural machinery that underlies LI, and our neural net-work model of LI may relate to the transition fromautomatic to controlled processing.
In so speculating, we make no claims to deal withthe Hard Question about consciousness (Chalmers,1996), i.e. the issue of how it is that brain processes,
the information processing that the brain performs,and/or the transactions between the organism and itsenvironment determined by such information processing– how any or all of these give rise to conscious experi-ences. One of us (Gray, 1995a, b) believes that thisis indeed a Hard Question that awaits a radically newtheory for its resolution; the others (Schmajuk andAxelrad, 1995; Buhusi and Schmajuk, 1996), that theHard Question will be resolved simply by the accumula-tion of new data and models within existing theoreticalmodels. We all believe, however, that the best (probablythe only) thing that can be done right now is to continuewith the accumulation of such data and the developmentof such models. It is in that spirit that the present paperis written.
2. LATENT INHIBITION AND SCHIZOPHRENIA
If a stimulus is repeatedly presented without otherconsequence (preexposure) and is subsequently used asthe conditioned stimulus (CS) in a standard Pavlovianconditioning paradigm, the preexposed (PE) CS developsa weaker association with the unconditioned stimulus(US), as measured by the strength of the ensuing condi-tioned response (CR), than does a non-preexposed (NPE)CS. This difference between the CRs evoked by PEand NPE CSs, respectively, is the phenomenon of latentinhibition. Most often, different groups of subjects areconditioned with PE and NPE CSs, respectively. In thiscase, LI is measured as the difference in the efficacy
Neural Networks, Vol. 10, No. 7, pp. 1257–1268, 1997q 1997 Elsevier Science Ltd. All rights reserved
Printed in Great Britain0893–6080/97 $17.00+.00
PII: S0893-6080(97)00058-0
Pergamon
1257

of conditioning between the preexposed and non-preexposed groups. This between-subjects approachmay, however, be complemented by within-subjectdesigns, in which all subjects receive both PE and NPECSs and LI is measured as the difference in the efficacyof conditioning between the two CSs (e.g. N. S. Grayet al., 1995; Killcross and Robbins, 1993).
First described by Lubow and Moore (1959), LI hassubsequently been the subject of considerable investi-gation (Lubow, 1989) in many species, including man.The experiments in Gray’s laboratory have used bothrats and human subjects. In the former case, we typicallyassess LI using an off-the-baseline conditioned emo-tional response (CER) procedure in animals licking forwater (e.g. Feldon and Weiner, 1991; Weiner et al.,1984). In this procedure, after initial baseline trainingto lick, CS preexposure and CS–footshock pairings areboth conducted without access to water, and the CS isthen presented to the rat while it is again licking; CRmagnitude is measured by the degree of lick suppressionduring the CS. LI consists of lower suppression of lickingin response to the CS in the PE as compared to the NPEanimals.
Another procedure used to study LI is nictitating mem-brane or eyeblink conditioning, in rabbits (Solomon andMoore, 1975) and rats (Schmajuk et al., 1994). In theseexperiments, animals in the PE condition are exposed tothe CS for several days. Animals in the control, NPEcondition remain in the conditioning chamber for anequivalent amount of time without CS presentations forthe same number of days. On the first conditioning day,animals in the PE condition are exposed to additionaltrials of CS-alone presentations, while animals in theNPE condition again remain in the conditioning chamberfor an equivalent time. Subsequently on the same day,when subjects in both conditions are trained in a delayconditioning paradigm, PE animals exhibit slower acqui-sition of the CR than NPE animals.
With human subjects, our usual procedure is basedupon that of Ginton et al. (1975). Subjects first listento a tape recording of nonsense syllables, with instruc-tions to count the number of times they recur. In the PEcondition, bursts of low-intensity white noise (the CS)are randomly superimposed on the recording. Subse-quently, still listening to the tape recording, subjectsare asked to predict when a counter display will be incre-mented; increments (the US) are preceded for allsubjects by the white noise CS, and the number oftrials taken to detect this contingency is the measureof conditioning.
The significance of disrupted LI for the deficit inthe ability to ignore irrelevant stimuli extensively docu-mented in schizophrenia (for review, see Hemsley, 1987)was initially pointed out by groups in Massachusetts(Solomon et al., 1981; Solomon and Staton, 1982) andTel Aviv (Weiner et al., 1981, 1984). These groupsboth reported that LI is abolished in the rat by systemic
administration of the indirect dopamine (DA) agonist,amphetamine. Neurochemically, the relevance to schizo-phrenia of this observation derives from the fact thatindirect DA agonists, including amphetamine (Meltzerand Stahl, 1976), have psychotomimetic effects, whileDA receptor antagonists, such as haloperidol, are usedtherapeutically as anti-psychotics (Carlsson, 1988;Haracz, 1982; Meltzer and Stahl, 1976; Swerdlow andKoob, 1987). Psychologically, its relevance lies in thefact that at least some positive psychotic symptoms(Crow, 1980) can be regarded as reflecting a diminishedtendency to screen out redundant stimuli from pro-cessing (Anscombe, 1987; Frith, 1979), a descriptionthat seems also to apply well to an amphetamine-treatedrat learning about a PE CS that is disregarded by aplacebo-treated control.
In the 15 years since publication of these seminalobservations, LI and its relation to schizophrenia havebecome the focus of increasingly intense interest (Dunnet al., 1993; Ellenbroek and Cools, 1990; Killcross et al.,1994; Weiner, 1990). This interest is largely based uponthe assumption that the blockade of LI by amphetaminein the rat presents a viable experimental model of posi-tive psychotic symptoms in man. We have recentlyembedded this assumption in a model of the neural sub-strate of LI, as well as in a general neuropsychologicaltheory of positively-symptomatic schizophrenia (Grayet al., 1991a, b; Weiner, 1990). This theory has in partbeen formulated mathematically as a neural network(Schmajuk et al., 1996; Buhusi and Schmajuk, 1996),considered in more detail below; and also related toclinically observed symptoms (Hemsley, 1993 Hemsley,1994) and, in outline, to the aberrations of consciousexperience that these symptoms reflect (Gray, 1993,1995a, b).
At the psychological level, our central hypothesis(Gray et al., 1991a; Hemsley, 1987; Weiner, 1990)states that the cognitive deficits of acute, positively-symptomatic schizophrenia stem from disruption ofthe normal ability to use past regularities as a guide tocurrent information processing. Within this framework,we treat LI as a case in which the initial regularity,to-be-CS leading to no consequence, is followed by therequirement to learn a conflicting regularity, which inour standard conditioned suppression paradigm withrats is CS-shockor, in our human paradigm (Baruchet al., 1988a, b),CS-counter increment. In the absenceof pathology or drug treatment, this sequence of conflict-ing regularities normally leads to retarded learning ofthe CS–US association (i.e. to LI). In animals whichfail to show LI (e.g. after amphetamine treatment), theinfluence of the past regularity is lost, and they are con-trolled by the most recent (i.e. the CS–US) regularity. Interms of symptomatology, Hemsley has, in a series ofpapers (Hemsley, 1987, 1993, 1994), provided supportfor the general theory by showing how, given this ana-lysis, blockade of LI can give rise to such positive
1258 J. A. Gray et al.

psychotic symptoms, well documented in the early stagesof schizophrenia, as hyper-vigilance; and similarlyhow blockade of the closely-related Kamin blockingeffect (Jones et al., 1992; Kamin, 1969) can give rise todelusional beliefs (elaborated in detail by Garety andHemsley, 1994).
In support of this theory, work in Gray’s laboratory hasdemonstrated loss of LI in normal human subjects afteroral administration of a low dose of amphetamine (N. S.Gray et al., 1992a; Thornton et al., 1996); and, critically,in medicated schizophrenics, provided that they aretested in the first two weeks of the current psychoticepisode (Baruch et al., 1988a; N. S. Gray et al.,1992b), before treatment with neuroleptics is able tobring symptoms under control. LI is also absent in unme-dicated schizophrenics, the loss of LI in this case lastingup to a year (N. S. Gray et al., 1995). In addition, a seriesof studies have reported that LI is reduced in normalsubjects who obtain high scores on questionnaire mea-surements of the tendency towards psychosis (Baruch etal., 1988b; De la Casa et al., 1993; Lipp and Vaitl, 1992;Lubow et al., 1992). At the same time, extensive studiesof the rat in a number of laboratories including our own(for review, see Gray et al., 1995) have done much toelucidate the neural basis of LI and its disruption.
3. THE NEURAL BASIS OF LATENT INHIBITION
As noted above, LI is abolished by increased DA release,as caused for example by systemic administration of theindirect DA agonist, amphetamine. Behaviourally, lossof LI occurs because conditioning is increased in PEsubjects, but not in NPE subjects, by indirect DAagonists. This effect is blocked by concomitant adminis-tration of DA receptor antagonists, such as haloperidol.In addition, these compounds, given on their own, induceLI under conditions (too few preexposure trials) in whichundrugged animals fail to display LI; this ‘‘potentiatedLI’’ is due to decreased conditioning induced by DAantagonists in PE but not NPE subjects (for review, seeGray et al., 1995). It has been proposed (Solomon et al.,1981; Solomon and Staton, 1982; Gray et al., 1991a) thatthese effects are due to alterations in dopaminergic trans-mission specifically at the terminals of the mesolimbicdopaminergic projection from nucleus (n.) A 10 in theventral tegmental area to n. accumbens in the ventralstriatal region of the basal ganglia. Thus, on this hypo-thesis, increased intra-accumbens DA release facilitates,while blockade of DA transmission reduces, condition-ing only in subjects that have had preexposure to theCS. Experiments using systemic administration of DAagonists and antagonists have further demonstrated thatboth blockade and potentiation of LI can be obtained ifthe relevant drug is administered only at the time ofconditioning, preexposure having taken place in thenormal state (Joseph et al., 1993; Peters and Joseph,1993; Moran et al., 1996). Thus, the hypothesis proposed
by Gray et al. (1991a, 1995) holds more specificallythat an increase in accumbens dopamine transmissionat the time of conditioning reverses, and a decrease inthis transmission boosts, the effects of CS preexposure;this hypothesis is consistent with Hemsley’s (1987)view of schizophrenic positive psychotic symptoms asreflecting a failure in the normal integration of currentinformation processing with previous regularities ofexperience.
The experiments with the rat have provided evidencein support of this hypothesis. These experiments haveshowninter alia the following:
1. In our standard CER paradigm (see above), a non-preexposed CS associated with the footshock UScomes to elicit conditioned DA release in n. accum-bens, but not in the dorsal striatum (target of thenigrostriatal dopaminergic projection). However, apreexposed CS (to which conditioning is substantiallyreduced) fails to elicit such conditioned DA release inn. accumbens (Young et al., 1993). Thus, accumbensDA release maps well on to the changes in condition-ing that constitute LI.
2. Destruction of dopaminergic terminals within n.accumbens potentiates LI (causing this phenomenonto appear with a number of PE trials too few to causeLI in normal animals); blockade of accumbens post-synaptic receptors by local injection of the DA recep-tor antagonist, haloperidol, at the time of conditioningonly similarly potentiates LI (S. Peters, G. Grigoryan,A. Young, M. H. Joseph and J. A. Gray, in prepara-tion). In both cases, these effects are due to changesin the preexposed animals only. Thus, in accord withthe hypothesis under test, decreased intra-accumbensDA transmission at the time of conditioning booststhe effects of CS preexposure, even though this hastaken place in the normal state.
3. The blockade of LI by systemic injection of the indir-ect DA agonists, amphetamine and nicotine, is in bothcases reversed by intra-accumbens injection of halo-peridol at the time of conditioning only (S. Peterset al., in preparation). Thus, the effect of theseagonists must be mediated by DA transmission in n.accumbens, showing that (in agreement with thehypothesis) an increase in such transmission reversesthe effect of CS preexposure even though this hastaken place in the normal state.
Other experiments, also offering support for theneuropsychological model of LI advanced by Grayet al. (1991a, b), have provided evidence (reviewedin Gray et al., 1995) that:
4. In general, but not always, LI depends upon the integ-rity of the hippocampal formation, and of the retro-hippocampal region (subicular area plus entorhinalcortex) reciprocally connected to the hippocampalformation.
5. The roles of the dopaminergic projections to n.
1259The Transition from Automatic to Controlled Processing

accumbens and the hippocampal system in LI areinterconnected; thus, for example, the loss of LIafter damage to the hippocampal formation or to theretrohippocampal region is in both cases restored bysystemic administration of haloperidol (Christiansenand Schmajuk, 1993; Yee et al., 1995).
Although the n. accumbens is usually thought of asbelonging to the motor programming circuitry of thebasal ganglia (in which, indeed, it clearly plays an impor-tant role; Swerdlow and Koob, 1987; Gray et al., 1991a),it appears to play a key role in the regulation of LI.Recently, Lavin and Grace (1994) have studied whathappens to the outputs from the n. accumbens. Usingelectrophysiological and tract-tracing techniques, theseworkers have demonstrated that the inhibitory GABA-ergic output from n. accumbens synapses, in the ventralpallidum, upon further GABA-ergic inhibitory neuronsthat project to the nucleus reticularis thalami (NRT). TheNRT is unusual among thalamic nuclei in that it tooconsists mainly of inhibitory GABA-ergic neurons;these project to a number of the surrounding thalamicnuclei whose job is to relay impulses originating inperipheral sense organs to the appropriate sensoryregions of the cerebral cortex (Jones, 1975). The possiblerole of the NRT in the selection of stimuli for attentionand conscious processing was first pointed out by Crick(1984), and has been incorporated into a neural networkmodel by Taylor and Alavi (1992). Note that, since thepallidal output to these neurons is itself inhibitory, itsactivation has the effect of disinhibiting these sensoryrelay pathways, i.e. increasing the entry to the cerebralcortex of those stimuli that are currently engaging the
thalamocortical loops. Figure 1 presents this circuitryin diagrammatic form.
Let us consider how the circuitry of Figure 1 would belikely to work under the conditions of an experiment inwhich an indirect DA agonist, such as amphetamine ornicotine, is used to block LI by causing DA release in n.accumbens. As we have seen, the basic phenomenon ofLI consists in the fact that the preexposed CS is slow toenter into an association with a Pavlovian US. Let usinterpret this, speculatively, as reflecting a lack of accessto conscious processing by the preexposed CS. If, how-ever, presentation of this CS is accompanied by enhancedDA release in n. accumbens (as induced pharmacologi-cally, by activation of the retrohippocampal input to n.accumbens, or during acute psychosis), LI is overcome,indicating ex hypothesithat the preexposed CS hasregained the capacity to engage conscious processing.The circuitry of Figure 1 constitutes a mechanism bywhich this effect can be produced. DA release within n.accumbens inhibits (by acting on DA D2 receptors;Robertson and Jian, 1995) the GABA-ergic pathway tothe ventral pallidum, thus disinhibiting the pallidalGABA-ergic pathway to NRT, which in turn inhibitsthe GABA-ergic projections from NRT to the ascendingthalamocortical sensory relay projections, so disinhibit-ing the latter. In this way, accumbal DA release shouldlead to an intensification of processing in whateverthalamocortical sensory relay projections were alreadyoperative in the prior instant of time. In the LI experi-ment, this intensification of sensory processing will allowthe preexposed CS (which otherwise would not havebeen fully processed) to enter more readily into asso-ciation with the US. This line of argument is consistent
1260 J. A. Gray et al.
FIGURE 1. Diagram of different brain regions involved in latent inhibition. Connections from the subiculum (sub) and entorhinal cortex(ERC) (‘‘retrohippocampal region’’) to the n. accumbens (NAC) and from the NAC to the nucleus reticularis thalami (NRT) and thalamo-cortical sensory pathways. PFC, prefrontal cortex; DM, dorsomedial thalamic area; VP, ventral pallidum; A 10, dopaminergic nucleusA 10 in the ventral tegmental area (VTA). Neurotransmitters: GLU, glutamate; DA, dopamine; GABA, gamma-aminobutyric acid. þ , ¹ :excitation and inhibition. I, II and III: feedback loops.

with the suggestion that, in acute schizophrenia, con-scious (or controlled) processing (Schneider and Shiffrin,1977) is dominant.
This analysis of how the circuitry of Figure 1 would beexpected to function when a supra-normal pulse of DA isinjected into it in n. accumbens needs to be understood inthe context of its likely continuing activity under morenormal conditions. A striking feature of this circuitry isthe imbalance between, on the one hand, the accumbensoutput to the thalamocortical sensory loops (3 steps) and,on the other, the dorsomedial thalamic-prefrontal motorloop (2 steps). This imbalance has the consequence that,
assuming that these pathways are activated either by thesame initial output from n. accumbens to the ventralpallidum or by two such outputs but simultaneously,the one set of thalamocortical loops will always beexcited when the other is inhibited. This arrangementseems ideally suited to produce a rapid oscillationbetween the taking in of information from the environ-ment and the organisation of a step in a motor program(see Gray et al., 1991a). Given other considerations asto the time-scale of events in these loops (Gray, 1982,1995a), we may suppose that these oscillations betweensensory-dominant and motor-dominant processing occur
1261The Transition from Automatic to Controlled Processing
FIGURE 2. Diagram of the SLG network. CS i, conditioned stimulus; t i, trace CS i; X i, internal representation of CS i, z i , attentionalmemory; Vi,US, CSi–US association; V i ;j , CSi–CSj association; B k, aggregate prediction of event k; BUS, aggregate prediction of theUS; CSk , average value of CS k; B k , average prediction of CS k; US, average value of the US; BUS, average prediction of the US; CR,conditioned response; OR, orienting response; A, Absolute values. Arrows represent fixed connections. Open triangles representvariable synapses.

every 50 ms or so. Figure 1 suggests, furthermore, that(in agreement with the hypothesis proposed by Weiner,1990) the switch between the one and the other form ofprocessing is controlled by the feedback loop from theventral pallidum via A 10 back to n. accumbens. Whenthe inhibitory step from n. accumbens to ventral pallidumis operated (leading to sensory-dominated processing),this simultaneously disinhibits the ventral pallidal inputto A 10 and so inhibits the A 10 to n. accumbens path-way. In this way accumbens inhibition of the ventralpallidum is reinstated, switching dominance back to thethalamo-prefrontal loop. Interpreted without furthergloss, this understanding of the circuitry of Figure 1would seem to predict that conscious experience(assumed here to be linked to processing in the sensorybut not the motor component of these linked loops,together with widespread activity in the overall compara-tor system, as described by Gray, 1995a) would be inter-rupted by recurrent periods of 50-ms blanks in every100 ms; which is clearly not the case. We need thereforeto suppose that, once the sensory processing that fillsconscious experience is initiated in the thalamocorticalloops, it continues until it is replaced, approximatelyevery 100 ms, by a new set of selected elements. Manyof these will presumably be the same as elements in theprevious set, so resulting in an apparently seamlessstream of consciousness.
4. A NEURAL NETWORK THEORY OF LATENTINHIBITION
In order to offer a formal description of the transitionfrom automatic to controlled processing, or from con-scious to unconscious modes, the present section intro-duces the neural network theory of LI offered bySchmajuk et al. (1996); (see also Schmajuk, 1997).Figure 2 shows a detailed diagram of the network,henceforth called the SLG model. The SLG network isareal-time modelwhich describes the unbroken temporaldynamics of behavioural variables and, therefore, itsoutput can be compared to behaviour as it unfolds inreal time. Furthermore, the dynamics of its interveningvariables can be contrasted with neural activity, provid-ing a basis for the study of the physiological founda-tions of behaviour. This dynamic description isformalized by a set of differential equations that depictchanges in the values of neural activities and connectiv-ities as a function of time. A formal description of themodel is presented in Appendix A.
The SLG network incorporates different types ofmemory: (1) a trace short-term memory (STM) of CSi,(2) an attentional long-term memory (LTM) for CSi, (3)associative CSi–CSk and CSi–US LTMs, and (4)intermediate-term memories of CSi, US, and theirpredictions.
Trace STM. Trace STM,t i(t), increases over time to amaximum during CSi presentation and then decays back
to zero (see Hull, 1943; Grossberg, 1975). In addition, weassume thatt i(t) increases also when CSi is predictedby other CSs (see eqn (1) in Appendix A).
Internal representation. The internal representation ofCSi, Xi, is proportional tot i and tot i multiplied by itsattentional LTM,zi (see eqn (2)). Internal representa-tions, Xi, determine both the read-in and read-out ofassociative LTM (see below). The magnitude of theinternal representation of CSi, Xi, modulates both (a)the rate of change of CSi–CSj and CSi–US associations(see eqns (4) and (49)), and (b) the magnitude of the CR(see eqns (3) and (9)). That is,Xi controls both storageand retrieval of CSi–CSj and CSi–US associations.Simultaneous control of memory read-in and memoryread-out byXi is a most important feature of the model.
Attentional LTM. Attentional LTM, zi, represents thepast experience that CSi, or its predictionBi, is accom-panied by a certain amount of NoveltySkllk ¹ Bkl(defined below) (see eqn (5)). This attentional LTM isestablished when Novelty temporally overlaps with theSTM of CSi, t i, or its predictionBi. LI is the consequenceof the depressedXi that results from the decreased atten-tion (small positivezi) or the inattention (negativezi) tothe preexposed CS. The magnitude of the LI effectdepends on the time needed to increaseXi by reversinginattention (negativezi) into attention (positivezi) duringconditioning.
Associative LTM. Associative LTM,Vi,k, representsthe past experience that CSi is followed by eventk(CSk or the US) (see eqns (4) and (49)). This associativeLTM is established when eventk temporally overlapswith the internal representation of CSi, Xi. By controllingthe magnitude ofXi (see eqn (2)), attentional LTMzi
controls the rate of change ofVi, US associations (seeeqn (4)). In addition, by controlling the magnitude ofXi ; zi also controls the size ofBUS (see eqn (39)) and thestrength of the CR (see eqn (9)).
Cognitive mapping. The network storing CS–CS andCS–US associations in Figure 2 is a recurrent autoasso-ciative network (Kohonen, 1977) capable of combiningmultiple associative values, a process similar to whatTolman (1932) called inference. Tolman hypothesizedthat a large number of associative values can be com-bined into acognitive map. The internal representationof CSi, Xi, becomes associated with the US in proportionto the difference (lUS ¹ BUS), whereBUS is the aggregateprediction of the US by all CSs with representationsactive at a given time, andlUS represents the intensityof the US. More generally, the aggregate prediction ofCSk by all CSs with representations active at a giventime, Bk, is given by Bk ¼ S i Xi Vi,k. Vi,k increaseswhen Xi(t) is active and eventk is underpredicted,i.e. (lk ¹ Bk) is greater than zero.Vi,k decreaseswhen Xi(t) is active and eventk is overpredicted, i.e.(lk ¹ Bk) is smaller than zero.Vi,k remains unchangedwheneverXi(t) is zero or eventk is perfectly predicted,i.e. (lk ¹ Bk) equals zero.
1262 J. A. Gray et al.

Novelty. Novelty of eventk is computed asllk ¹ Bkl,i.e. the absolute value of the difference betweenlk, therunning average of the observed value of eventk, andBk,the running average of the aggregate prediction of eventk. lk and Bk are computed by neural elements whoseoutputs are proportional to present and past values oflk andBk (see eqns (6) and (7)).
Novelty is given by the sum of the novelty of allstimuli present or predicted at a given time,Skllk ¹ Bkl(see eqn (8)). We assume that CSk can be predicted byother CSs, the context (CX), or itself. Therefore, eitherrepeated presentations of CSk in a given context orsimply repeated presentations of CSk lead to a decreasein CSk novelty. Whereas CSk–CSk associations decreaseCSk novelty in a context-independent manner, CSj–CSk
or CX–CSk associations decrease CSk novelty in a con-text-dependent way. Because, according to the SLGmodel, decrements in novelty are responsible for LI,CSk–CSk associations are responsible for context-nonspecific LI, whereas CSj–CSk or CX–CSk associa-tions are responsible for context-specific LI (see Goodand Honey, 1993).
Figure 2 shows that Novelty becomes associated withtracet i(t). This association is represented by the atten-tional memory,zi, which modulates the amplitude ofinternal representationXi, thereby modulating the rateof learning. Therefore, the SLG model suggests thatattentional memoryzi is CSi-specific but regulated bythe non-specific Novelty (see eqn (5) in Appendix A).
Orienting response (OR). Sokolov (1960) proposedthat the strength of the OR might be an index of theamount of processing afforded to a given stimulus, andthat this amount of processing is proportional to thenovelty of the stimulus. In line with Sokolov’s idea, weassume that Novelty,Skllk ¹ Bkl, determines the magni-tude of the OR (see eqn (9)).
Behavioural inhibition. Gray (1971) suggested that abehavioural inhibition system, activated by signals ofpunishment or non-reward, innate fear stimuli, or novelstimuli, gives rise to behavioural inhibition, increasedarousal, and increased attention to environmental stimuli.In the same vein, we suggest that CR amplitude (a)increases proportionally to the magnitude of the predic-tion of the US, and (b) decreases in proportion to themagnitude of the OR, CR¼ S iXiVi,k ¹ OR (see eqn (10)).
LI and the transition from controlled (conscious) toautomatic (unconscious) processing. According to themodel, at the beginning of acquisition of classical con-ditioning, Novelty andzi are large because both CSi andthe US are unexpected, but both Novelty andzi decreaseas conditioning progresses and both CSi and the USbecome expected. Pearce and Hall (1980) suggestedthat this transition from high to low levels of attentionwas akin to the concept of passage from controlled orconscious to automatic or non-conscious modes pro-posed by Schneider and Shiffrin (1977). At the beginningof conditioning, CSi is processed in controlled or
conscious mode, and later in training CSi is processedin automatic or unconscious mode.
In terms of the SLG model, LI is the consequence ofthe depressed representation of CSi, Xi, that results fromthe decreased Novelty that follows preexposure andfrom the decreased attention (smallzi) to the preexposedCS. This decreased attention can be interpreted, specula-tively, as reflecting a lack of access to conscious pro-cessing by the preexposed CS.
In summary, in the framework of the SLG model, CSi
is processed in controlled or conscious mode whenNovelty and zi are large, and in automatic or non-conscious mode when Novelty andzi are small.
5. A NEURAL SYSTEM INTERPRETATION OFTHE NETWORK MODEL
In this section we consider how to translate the variablesin the neural network model of latent inhibition into anequivalent set of functions on real-brain structures. Inattempting to do this, we limit ourselves to the variableNovelty. Although we have begun the task of makingappropriate translations for other variables within theneural network model, it is Novelty and its real-brainequivalent that are central to our present theme, the tran-sition from automatic to controlled processing. Let usnow consider how the critical variable Novelty in theSLG model can be mapped onto the circuitry ofFigure 1. Under the assumption that Novelty is repre-sented by the DA input to the accumbens, increases inNovelty result in increased DA release within n. accum-bens which inhibits the GABA-ergic pathway to theventral pallidum, thus disinhibiting the pallidal GABA-ergic pathway to NRT, which in turn inhibits the GABA-ergic projections from NRT to the ascending thalamo-cortical sensory projections, so disinhibiting the latter. Inthis way, increased Novelty increases accumbal DArelease, thus leading to an intensification of processingof the CSs that activate the thalamocortical sensory relayprojections, and to more conscious processing. After CSpreexposure, decreases in Novelty result in a reductionof the attentional memory and the processing of thepreexposed CS, and in less conscious processing.
Weiner et al. (1988) demonstrated that two adminis-trations of amphetamine, given prior to preexposure andprior to conditioning on successive days, are effective inimpairing LI. In contrast to the effect of amphetamineadministration, Weiner et al. (1987) reported that halo-peridol administered during both the preexposure andconditioning phases facilitates LI, but that this effect isabsent when drug administration is limited to the pre-exposure phase.
The top left panel in Figure 3 shows mean percentageof avoidance responses for preexposed and non-preexposed groups under saline or haloperidol adminis-tration (during preexposure and conditioning) as reportedby Weiner et al. (1987). Whereas animals receiving
1263The Transition from Automatic to Controlled Processing

saline (CON/NPE and CON/PE) show little difference inbehaviour, PE animals treated with haloperidol show aclear retardation in the acquisition of avoidance(HAL/NPE vs. HAL/PE). In addition, animals receivingsaline (CON/NPE and CON/PE) learn faster thananimals receiving haloperidol (HAL/NPE vs. HAL/PE).The bottom left panel in Figure 3 presents computersimulations obtained with the SLG network showingpeak CR amplitude after 5 CS preexposure trialsfollowed by 10 conditioning trials. Although simulatedHAL/PE animals show more learning than the experi-mental animals, in agreement with the experimentaldata, haloperidol administration facilitates LI.
Thetoprightpanel inFigure3showsmeanpercentageofavoidance responses for preexposed and non-preexposed
groups under saline or amphetamine administration (dur-ing preexposure and conditioning) as reported by Weineret al. (1988). Whereas animals treated with amphetamine(AMP/NPE and AMP/PE) show little difference inbehaviour, PE animals receiving saline show a clearretardation in the acquisition of avoidance (CON/NPEvs. CON/PE). In addition, animals receiving ampheta-mine (AMP/NPE and AMP/PE) learn faster than animalsreceiving saline (CON/NPE and CON/PE). The bottomright panel in Figure 3 presents computer simulationsobtained with the SLG network showing CR amplitudeafter 295 context preexposure trials representing appara-tus familiarization and 5 CS preexposure trials followedby 6 conditioning trials. In agreement with the experi-mental data, amphetamine administration impairs LI.
1264 J. A. Gray et al.
FIGURE 3. Effect of the administration of amphetamine and haloperidol on LI. Top left panel: (data from Weiner et al., 1987): Meanpercentage avoidance responses for saline-control (CON) and haloperidol-treated (HAL), preexposed (PE) and non-preexposed (NPE)rats. Lower left panel: (simulation) Percentage peak CR after 5 CS preexposure trials followed by 10 conditioning trials for saline-control(CON) and haloperidol-treated (HAL), preexposed (PE) and non-preexposed (NPE) rats. Top right panel: (data from Weiner et al., 1988)Mean percentage avoidance responses for saline-control (CON) and amphetamine-treated (AMPH), preexposed (PE) and non-preex-posed (NPE) rats. Lower right panel: (simulation) Percentage peak CR after 295 context-only preexposure trials, 5 CS preexposure trialsfollowed by 6 conditioning trials for saline-control (CON) and amphetamine-treated (AMPH), preexposed (PE) and non-preexposed (NPE)rats. Because experimental data show that CS–US classical associations (interpreted as fear of the US) modulate the generation ofavoidance responses (Rescorla, 1967; Rescorla and LoLordo, 1965; Weisman and Litner, 1969), we compare percent CR amplitude withthe mean percent of avoidance responses.

In addition to describing the effect of haloperidol andamphetamine administration on LI, the model alsodescribes LI facilitation produced by context preexpo-sure that precedes CS preexposure (compare LI in nor-mal preexposed animals without context preexposure(left panels) to LI in normal preexposed animals withcontext preexposure (right panels)).
In the model, indirect DA agonists, such as ampheta-mine or nicotine, enhance the DA representation ofNovelty reduced by CS preexposure, thereby increasingattention, overcoming LI, and engaging conscious pro-cessing of the preexposed CS. As mentioned, CSi isprocessed in controlled or conscious mode when Noveltyandzi are large, and in automatic or non-conscious modewhen Novelty andzi are small. Therefore, administrationof DA agonists force the system to process CSi in thecontrolled, conscious mode during conditioning. Bycontrast, DA receptor antagonists, such as haloperidol,further reduce the DA representation of Novelty alreadyreduced by CS preexposure, thereby additionallydecreasing attention, intensifying LI, and engagingunconscious processing of the preexposed CS.
6. CONCLUSION
In the preceding sections we have sketched out a neuralnetwork model of the basic behavioural phenomenon ofLI. As indicated in detail elsewhere (Schmajuk et al.,1996; Schmajuk, 1997), this model makes a very goodjob of accounting for the substantial corpus of empiricaldata concerning LI, as summarised in the book by Lubow(1989), as well as accommodating (and therefore, ineffect, predicting) a number of experimental observa-tions published since the model was completed. Wehave also, but more tentatively, indicated some possible‘‘translations’’ between the building blocks of the neuralnetwork (Figure 2), on the one hand, and systems in thereal brain that have been implicated in LI experi-mentally (as reviewed briefly above; and see Weiner,1990; Gray et al., 1991a, 1995). In doing so, we haveconcentrated here on only a few of these ‘‘translations’’,particularly those most closely related to our presenttheme regarding the transition from automatic to con-scious processing. We shall expand the list of suchpossible translations in a future paper (Schmajuk,Buhusi and Gray, in preparation).
These developments are part of normal science in therelevant fields: that is, they use a standard process ofreverse engineering in the attempt to account for obser-vations on the brain and behaviour that have beengathered in standard laboratory procedures. However,in terms of current scientific understanding, the relation-ship between observations of this kind, on the one hand,and conscious experience, on the other, is undefined andmust remain so until we are in possession of a theory thatprovides more than a series of brute correlations to linkthe two sides of this divide (see Nagel, 1993).
While awaiting the advent of that theory, it is non-theless worth speculating on just what might turn out tobe linked to what in the brain-consciousness conundrum.This paper has attempted to do this for the transitionbetween automatic and controlled processing, a transi-tion which appears to go badly awry in schizophrenia.
REFERENCES
Anscombe, F. (1987). The disorder of consciousness in schizophrenia.Schizophrenia Bulletin, 13, 241–260.
Baruch, I., Hemsley, D. R., & Gray, J. A. (1988a). Differential perform-ance of acute and chronic schizophrenics in a latent inhibition task.Journal of Nervous and Mental Disease, 176,598–606.
Baruch, I., Hemsley, D. R., & Gray, J. A. (1988b). Latent inhibition andpsychotic proneness in normal subjects.Personality and IndividualDifferences, 9, 777–783.
Buhusi, C., & Schmajuk, N. A. (1996). Attention, configuration andhippocampal function.Hippocampus, 6, 621–642.
Carlsson, A. (1988). The current status of the dopamine hypothesis ofschizophrenia.Neuropsychopharmacology, 1, 179–203.
Chalmers, D. J. (1996).The conscious mind: In search of a funda-mental theory. New York: Oxford University Press.
Christiansen, B. A., & Schmajuk, N. A. (1993). Latent inhibition:effects of haloperidol and hippocampal lesions.Society of Neuro-science Abstracts, 19, 798.
Crick, F. (1984). The function of the thalamic reticular complex: thesearchlight hypothesis.Proceedings of the National Academy ofSciences USA, 81, 4586–4590.
Crow, T. J. (1980). Positive and negative schizophrenic symptoms andthe role of dopamine.British Journal of Psychiatry, 137,383–386.
De la Casa, L. G., Ruiz, G., & Lubow, R. E. (1993). Latent inhibitionand recall/recognition of irrelevant stimuli as a function of pre-exposure duration in high and low psychotic-prone normals.BritishJournal of Psychology, 84, 119–132.
Dunn, L. A., Atwater, G. E., & Kilts, C. D. (1993). Effects of anti-psychotic drugs on latent inhibition: sensitivity and specificity of ananimal behavioural model of clinical drug action.Psychopharmacology, 112,315–323.
Ellenbroek, B. A., & Cools, A. (1990). Animal models with constructvalidity for schizophrenia.Behavioural Pharmacology, 1,469–490.
Feldon, J., & Weiner, I. (1991). The latent inhibition model of schizo-phrenic attention disorder: haloperidol and sulpiride enhance rats’ability to ignore irrelevant stimuli.Biological Psychiatry, 29,635–646.
Frith, C. D. (1979). Consciousness, information processing andschizophrenia.British Journal of Psychiatry, 134,225–235.
Garety, P.A. & Hemsley, D.R. (1994).Delusions: investigations intothe psychology of delusional reasoning. Maudsley Monographs, No.36. Oxford: Oxford University Press.
Ginton, A., Urca, G., & Lubow, R. E. (1975). The effects of preexpo-sure to a non-attended stimulus on subsequent learning: latentinhibition in adults.Bulletin Psychonomic Society, 5, 5–8.
Good, M., & Honey, R. (1993). Selective hippocampus lesions abolishcontextual specificity of latent inhibition and conditioning.Behavioural Neuroscience, 107,23–33.
Gray, J. A. (1971).The psychology of fear and stress. London:Weidenfeld and Nicholson.
Gray, J. A. (1982).The neuropsychology of anxiety. Oxford: OxfordUniversity Press.
Gray, J. A. (1993). Consciousness, schizophrenia and scientific theory.In G.R. Bock & J. Marsh (Eds),Experimental and theoreticalstudies of consciousness(pp. 263–272). Ciba Foundation Sympo-sium 174. Chichester: John Wiley and Sons.
Gray, J. A. (1995a). The contents of consciousness: a neuropsycho-logical conjecture.Behavioral and Brain Sciences, 18, 659–676.
1265The Transition from Automatic to Controlled Processing

Gray, J. A. (1995b). Consciousness and its (dis)contents.Behavioraland Brain Sciences, 18, 703–722.
Gray, J. A., Feldon, J., Rawlins, J. N. P., Hemsley, D. R., & Smith, A. D.(1991a). The neuropsychology of schizophrenia.Behavioral andBrain Sciences, 14, 1–20.
Gray, J. A., Hemsley, D. R., Feldon, J., Gray, N. S., & Rawlins, J. N. P.(1991b). Schiz bits: misses, mysteries and hits.Behavioral andBrain Sciences, 14, 56–84.
Gray, J. A., Joseph, M. H., Hemsley, D. R., Young, A. M. J., Warburton,E. C., Boulenguez, P., Grigoryan, G. A., Peters, S. L., Rawlins,J. N. P., Taib, C.-T., Yee, B. K., Cassaday, H., Weiner, I., Gal,G., Gusak, O., Joel, D., Shadach, E., Shalev, U., Tarrasch, R., &Feldon, J. (1995). The role of mesolimbic dopaminergic and retro-hippocampal afferents to the nucleus accumbens in latent inhibition:implications for schizophrenia.Behavioural Brain Research, 71,19–31.
Gray, N. S., Pickering, A. D., Hemsley, D. R., Dawling, S., & Gray, J. A.(1992a). Abolition of latent inhibition by a single 5 mg dose ofd-amphetamine in man.Psychopharmacology, 107,425–430.
Gray, N. S., Hemsley, D. R., & Gray, J. A. (1992b). Abolition of latentinhibition in acute, but not chronic, schizophrenics.Neurology,Psychiatry and Brain Research, 1, 83–89.
Gray, N.S., Pilowsky, L.S., Gray, J.A., & Kerwin, R.W. (1995). Latentinhibition in drug naive schizophrenics: relationship to duration ofillness and dopamine D2 binding using SPET.SchizophreniaResearch, 17, 95–107.
Grossberg, S. (1975). A neural model of attention, reinforcement anddiscrimination learning.International Review of Neurobiology, 18,263–327.
Haracz, J. L. (1982). The dopamine hypothesis: an overview of studieswith schizophrenic patients.Schizophrenia Bulletin, 8, 438–469.
Hemsley, D. R. (1987). An experimental psychological model forschizophrenia. In H. Hafner, W.F. Gattaz & W. Janzavik (Eds),Search for the causes of schizophrenia. Stuttgart: Springer-Verlag.
Hemsley, D. R. (1993). A simple (or simplistic?) cognitive model forschizophrenia.Behaviour Research and Therapy, 31, 633–645.
Hemsley, D. R. (1994). Cognitive disturbance as the link betweenschizophrenic symptoms and their biological bases.Neurology,Psychiatry and Brain Research, 2, 163–170.
Hull, C. L. (1943). Principles of behavior. New York: AppletonCentury Crofts.
Jones, E. G. (1975). Some aspects of the organisation of the thalamicreticular complex.Journal of Comparative Neurobiology, 162,285–308.
Jones, S. H., Gray, J. A., & Hemsley, D. R. (1992). Loss of the Kaminblocking effect in acute but not chronic schizophrenics.BiologicalPsychiatry, 32, 739–755.
Joseph, M. H., Peters, S. L., & Gray, J. A. (1993). Nicotine blocks latentinhibition in rats: evidence for a critical role of increased functionalactivity of dopamine in the mesolimbic system at conditioningrather than preexposure.Psychopharmacology, 110,187–192.
Kamin, L. J. (1969). Predictability, surprise, attention and conditioning.In B.A. Campbell & R.M. Church (Eds),Punishment and aversivebehavior(pp. 279–296). New York: Appleton Century Crofts.
Killcross, A. S., Dickinson, A., & Robbins, T. W. (1994). Ampheta-mine-induced disruptions of latent inhibition are reinforcermediated: implications for animal models of schizophrenic atten-tional dysfunction.Psychopharmacology, 115,185–195.
Killcross, A. S., & Robbins, T. W. (1993). Differential effects of intra-accumbens and systemic amphetamine on latent inhibition using anon-baseline, within-subject conditioned suppression paradigm.Psychopharmacology, 110,479–489.
Kohonen, T. (1977).Associative memory. Berlin: Springer-Verlag.Lavin, A., & Grace, A. A. (1994). Modulation of dorsal thalamic cell
activity by the ventral pallidum: its role in the regulation ofthalamocortical activity by the basal ganglia.Synapse, 18,104–127.
Lipp, O. V., & Vaitl, D. (1992). Latent inhibition in human Pavloviandifferential conditioning: effect of additional stimulation after
pre-exposure and relation to schizoptypal traits.Personality andIndividual Differences, 13, 1003–1012.
Lubow, R. E. (1989).Latent inhibition and conditioned attentiontheory. Cambridge: Cambridge University Press.
Lubow, R. E., Ingberg-Sachs, Y., Zalstein-Orda, N., & Gewirtz, J. C.(1992). Latent inhibition in high and low psychotic prone subjects.Personality and Individual Differences, 13, 563–572.
Lubow, R. E., & Moore, A. U. (1959). Latent inhibition: the effect ofnon-reinforced pre-exposure to the conditioned stimulus.Journal ofComparative and Physiological Psychology, 52, 415–419.
Meltzer, H. Y., & Stahl, S. M. (1976). The dopamine hypothesis ofschizophrenia: a review.Schizophrenia Bulletin, 2, 19–76.
Moran, P. M., Gray, J. A. & Joseph, M. H. (1996). Disruption of latentinhibition in the rat by a single administration of amphetamine.Journal of Psychopharmacology, 10, A53.
Nagel, T. (1993). What is the mind–body problem? In G. R. Bock &J. Marsh (Eds),Experimental and theoretical studies in conscious-ness(pp. 1–7). Chichester: John Wiley and Sons.
Pearce, J. M., & Hall, G. (1980). A model for Pavlovian learning:variations in the effectiveness of conditioned but not unconditionedstimuli. Psychological Review, 87, 532–552.
Peters, S. L., & Joseph, M. H. (1993). Haloperidol potentiation of latentinhibition in rats: evidence for a critical role at conditioning ratherthan preexposure.Behavioural Pharmacology, 4, 183–186.
Rescorla, R. A. (1967). Inhibition of delay in Pavlovian fearconditioning. Journal of Comparative and PhysiologicalPsychology, 64, 114–120.
Rescorla, R. A., & LoLordo, V. M. (1965). Inhibition of avoidancebehavior.Journal of Comparative and Physiological Psychology,59, 406–412.
Robertson, G. S., & Jian, M. (1995). D1 and D2 dopamine receptorsdifferentially increase fos-like immunoreactivity in accumbal pro-jections to the ventral pallidum and midbrain.Neuroscience, 64,1019–1034.
Schmajuk, N. A. (1997).Animal learning and cognition. Cambridge:Cambridge University Press.
Schmajuk, N. A., & Axelrad, E. (1995). Communication and conscious-ness: a neural network conjecture.Behavioral and Brain Sciences,18, 695–696.
Schmajuk, N. A., Lam, P., & Christiansen, B. A. (1994). Hippo-campectomy disrupts latent inhibition of the rat eyeblinkconditioning.Physiology and Behavior, 55, 597–601.
Schmajuk, N., Lam, Y.-W. & Gray, J. A. (1996) Latent inhibition: aneural network approach.Journal of Experimental Psychology(Animal Behaviour Processes), 22, 1029–1041.
Schneider, W., & Shiffrin, R. M. (1977). Controlled and automatichuman information processing: I. Detection, search and attention.Psychological Review, 84, 1–66.
Sokolov, E. N. (1960). Neuronal models and the orienting reflex. InM. A. B. Brazier (Ed.),The central nervous system and behaviour.New York: Josiah Macey Junior Foundation.
Solomon, P. R., & Moore, J. W. (1975). Latent inhibition and stimulusgeneralisation of the classically conditioned nictating membraneresponse in rabbits following dorsal hippocampal ablation.Journalof Comparative and Physiological Psychology, 89, 1192–1203.
Solomon, P. R., & Staton, D. M. (1982). Differential effects of micro-injections of d-amphetamine into the nucleus accumbens or thecaudate-putamen on the rat’s ability to ignore an irrelevantstimulus.Biological Psychiatry, 16, 519–537.
Solomon, P. R., Crider, A., Winkelman, J. W., Turi, A., Kamer, R. M.,& Kaplan, L. J. (1981). Disrupted latent inhibition in the rat withchronic amphetamine or haloperidol-induced supersensitivity: rela-tionship to schizophrenic attention disorder.Biological Psychiatry,16, 519–537.
Swerdlow, N., & Koob, G. F. (1987). Dopamine, schizophrenia, maniaand depression: toward a unified hypothesis of cortico-striato-pallidothalamic function.Behavioral and Brain Sciences, 10,197–245.
1266 J. A. Gray et al.

Taylor, J. G., & Alavi, F. N. (1992). Mathematical analysis of a com-petitive network for attention. In Taylor, J.G. (Ed.),Mathematicalapproaches to neural networks(pp. 341–382). Amsterdam:Elsevier Science Publishers.
Thornton, J. C., Dawe, S., Lee, C., Capstick, C., Corr, P. J., Cotter, P.,Frangou, S., Gray, N. S., Russell, M. A. H., & Gray, J. A. (1996).Effects of nicotine and amphetamine on latent inhibition in humansubjects.Psychopharmacology, 127, 164–173.
Tolman, E. C. (1932).Purposive behavior in animals and men. NewYork: Irvington Publishers.
Weiner, I. (1990). Neural substrates of latent inhibition: the switchingmodel.Psychological Bulletin, 108,442–461.
Weiner, I., Feldon, J., & Katz, Y. (1987). Facilitation of the expressionbut not the acquisition of latent inhibition by haloperidol in rats.Pharmacology, Biochemistry and Behavior, 26, 241–246.
Weiner, I., Lubow, R. E., & Feldon, J. (1981). Chronic amphetamineand latent inhibition.Behavioural Brain Research, 2, 285–286.
Weiner, I., Lubow, R. E., & Feldon, J. (1984). Abolition of the expres-sion but not the acquisition of latent inhibition by chronic amphe-tamine in rats.Psychopharmacology, 83, 194–199.
Weiner, I., Lubow, R. E., & Feldon, J. (1988). Disruption of latentinhibiton by acute administration of low doses of amphetamine.Pharmacology, Biochemistry and Behaviour, 30, 871–878.
Weisman, R. G., & Litner, J. S. (1969). Positive conditioned reinforce-ment of Sidman avoidance in rats.Journal of Comparative andPhysiological Psychology, 68, 597–603.
Yee, B., Feldon, J., & Rawlins, J. N. P. (1995). Latent inhibition in ratsis abolished by NMDA-induced dopamine loss in the retrohippo-campal region but this lesion effect can be prevented by systemichaloperidol treatment.Behavioral Neuroscience, 109,227–240.
Young, A. M. J., Joseph, M. H., & Gray, J. A. (1993). Latent inhibitionof conditioned dopamine release in the nucleus accumbens.Neuroscience, 54, 5–9
APPENDIX A:
A.1. A Formal Description of the SLG Model
This section formally describes the SLG model as depicted in Figure 2.
A.1.1. Short-Term Memory Trace of the CSCSi generate an STM trace,t i, according to
d(ti )=dt ¼ K1(CSi ¹ ti ) (1)
whereK1 represents the rate of increase and decay oft i, and the initialvalue oft i is zero.
A.1.2. Internal Representation of the CSThe combination oft i with the prediction of CSi, Bi, can be modified bythe attentional valuezi, to yield the internal representation of CSi, Xi,according to
Xi ¼ K2(ti þ K3Bi )(K4 þ zi ) (2)
whereK3 represents a reinjection coefficient forBi, K4 represents anunmodifiable connection between input,t i þ K3 Bi, andXi, andzi is theattentional memory defined by eqn (5). By eqn (2),Xi is active eitherwhen (a) CSi is present andt i is greater than zero, or (b) when CSi ispredicted by other CSs andBi is greater than zero. Increasing values ofzi increase the magnitude ofXi. Because increasing CSi–CSi associa-tions increaseBi, they also increase the magnitude ofXi.
A.1.3. Aggregate PredictionsTheaggregate predictionof eventk (CSk) by all CSs with representa-tions active at a given time,Bk, is given by
Bk ¼SiVi, kXi (3)
whereVi,k represents the association ofXi with CSk.
The aggregate prediction of the US by all CSs with representationsactive at a give time,BUS, is given by
BUS ¼SiVi, USXi (3’)
whereVi,US represents the association ofXi with the US.
A.1.4. Long-Term Memory CS–US AssociationsChanges in the association betweenXi and the US,Vi,US, are given by
d(Vi, US)=dt ¼ K5Xi (lUS ¹ BUS)(1¹ jVi, USj) (4)
whereXi represents the internal representation of CSi, lUS the intensityof the US,BUS the aggregate prediction of the US by allX’s active at agiven time. The term (1¹ lVi,USl) boundsVi,US between 1 and¹ 1. Byeqn (4), Vi,US increases wheneverXi is active andlUS . BUS anddecreases whenlUS , BUS. In order to prevent the extinction ofconditioned inhibition or the generation of an excitatory CS by present-ing a neutral CS with an inhibitory CS, we assume thatBUS ¼ 0 whenBUS , 0.
A.1.5. Long-Term Memory CS–CS AssociationsChanges in the association betweenXi and CSj, Vi,j, are given by
d(Vi, j )=dt ¼ K5Xi (lj ¹ Bj )(1¹ jVi, j j) (4’)
whereXi represents the internal representation of CSi, l j represents theintensity of CSj, Bj represents the aggregate prediction of eventj by allX’s active at a given time. The term (1¹ lVi,jl) boundsVi,j between 1and ¹ 1. By eqn (49), Vi,j increases wheneverXi is active andl j . Bj
and decreases whenl j , Bj. WhenBj , 0 thenBj ¼ 0. Vi,i representsthe association of CSi with itself.
A.1.6. Attentional MemoryChanges in the association between (t i þ K3 Bi) and Novelty,zi, aregiven by
d(zi )=dt ¼ (ti þ K3Bi )(K6Novelty(1¹ zi ) ¹ K7(1þ zi ) (5)
whereK6 represents the rate of increase,K7 the rate of decay ofzi, andNovelty is given by eqn (8). When Novelty is zero,zi decreases untilzi
¼ ¹ 1.We assume that whenzi # 0, thenXi ¼ K2(t i þ K3 Bi)K4 (see eqn
(2)). This means that whenzi becomes negative, input (t i þ K3 Bi)activatesXi only through the unmodifiable connnectionK4. By eqn (6),whereas a CSi with negativezi has a relatively smallXi, and therefore,its associations change slowly, a CSi with positivezi has a relativelylargeXi, and therefore, its associations change rapidly. LI is the con-sequence of the negative value acquired byzi during CS preexposure (asNovelty decreases) and, consequently, of the time needed to restorezi toits positive value (as Novelty increases) during conditioning. Interest-ingly, in line with Lubowet al.’s (1981) suggestion, positive values ofzi
can be interpreted as a measure of the attention directed to CSi, whereasnegative values ofzi can be interpreted as a measure of theinattentionto CSi.
A.1.7. Total NoveltyThe average observed value of eventk is given by
lk ¼ (1¹ lk)lk þ K8lk (6)
whereK8 represents the rate of decay oflk.The average prediction of eventk is given by
Bk ¼ (1¹ Bk)Bk þ K8Bk (7)
whereK8 represents the rate of decay oflk. Novelty is given by
Novelty¼Skjlk ¹ Bkj (8)
wherek includes all CSs and the US.
A.1.8. Orienting ResponseThe magnitude of the OR is given by
OR¼ Novelty2=(K2
9 þ Novelty2) (9)
1267The Transition from Automatic to Controlled Processing

A.1.9. Performance RulesThe amplitude of the CR is given by
CR¼ B9US(1¹ K10OR) (10)
whereB’ US is given byB’ US ¼ BUS2 /(K11
2 þ BUS2 ). According to eqn (9),
the magnitude of the CR increases with increasing predictions of theUS, and decreases with increasing ORs elicited by novelty.
A.1.10. Effects of Amphetamine and HaloperidolAdministrationIt is assumed that the administration of amphetamine or nicotineincreases, and administration of haloperidol decreases, the action ofthe VTA dopaminergic projection to the n. accumbens coding Novelty.Novelty is given by
Novelty¼ K12Skjlk ¹ Bkj (8’)
where K12 . 1 for DA agonist andK12 , 1 for DA antagonistadministration.
A.2. Simulation Parameters
In our computer simulations, CS-preexposure trials, context-preexposure trials, and conditioning trials last 600 time units. A CSof amplitude 1.0 is presented between time units 55 and 80, a US ofintensity 1.8 is applied between time units 75 and 80. Context amplitudeis 0.1. Parameter values used in all simulations areK1 ¼ 0.2,K2 ¼ 2,K3
¼ 0.4,K4 ¼ 0.1,K5 ¼ 0.005,K6 ¼ 0.02,K7 ¼ 0.005,K8 ¼ 0.005,K9 ¼0.75,K10 ¼ 0.7,K11 ¼ 0.15, andK12 ¼ 1. Administration of ampheta-mine was simulated by assumingK12 ¼ 2.0, and administration ofhaloperidol by assumingK12 ¼ 0.65.
A DOS version of the program used in the present paper is availableon request.
1268 J. A. Gray et al.

1997 SPECIAL ISSUE
Plato’s Theory of Ideas Revisited
Javier Ropero Pelaez
Department of Mathematical Engineering, Faculty of Engineering, The University of Tokyo, 7-3-1 Bunkyo-ku, 113,
(Received15 July 1996;accepted15 April 1997)
Abstract—According to Plato’s theory, every pattern perceived by the mind is a composition of raw concepts (ideals).In this paper it is argued that the thalamus can decompose input patterns into these ideals and, during development,generate them from incoming patterns. As the neuronal counterpart of a certain ideal is proposed to be the activation of areticular neuron inside a competitive layer in the thalamus, these ideals are not coincident over time. In order to recruitthe ideals of a certain pattern, ideals are delayed in the long dendrites of pyramidal neurons. When several parts of apattern are to be related, these are bound through the association of their equal constituent ideals. Coincidences of thesame ideals are stored through synaptic reinforcement in nearby spines of pyramidal neurons. After binding, if a partialpattern is input, it will be reconstructed by the recall of its ideals over the reinforced synapses.q 1997 Elsevier ScienceLtd.
Keywords—Thalamus, Cortex 4th layer, Pyramidal neurons, Spine, Temporal coding, Burst firing, Orthogonalization,Binding.
1. INTRODUCTION
The ‘Myth of the Cave’, found in Plato’s dialogue,TheRepublic, illustrates how, according to the philosopher,objects perceived by the senses are nothing but a com-bination of pure entities called ‘ideals’. It can beimagined that some people are trapped in an undergroundcave in such a way that they can only see the cave’s innerwall and cannot see the outside world. They think that theshadows projected onto the wall constitute the real world,and do not realize that these shadows are only poorreflections of radiant and perfect figures that exist outsidethe cave. In this way, every object of their world is amixture of the projections of these figures onto thecave wall. These figures are a metaphor for the pure‘ideals’ that, when combined, give rise to an entireworld represented by the shadows on the cave wall.For example, according to Plato, the idea of a real treewould be a combination of the ideas of an ideal tree, theidea of greenness, the idea of height, etc. However, these
ideals (the basic and distinctive characteristics that com-bine to give rise to the various phenomena we comeacross in nature) can be more abstract. For example, inthe brain there appears to exist a kind of process forthe decomposition of incoming visual patterns in sucha way that, at a cortical level, different groups of neuronsbecome specialized in detecting individual lines thatcompose the pattern.
The dwellers of the cave have no consciousness ofthis ideal world because they are unable to see beyondthe images on the cave wall. Indeed, we are incapableof perceiving the abstract seeds of knowledge that, as awhole, produce our consciousness. In the above examplewe do not appreciate a visual pattern as a group of dis-tinctive lines even though we appreciate it as a whole.Plato considered that the philosopher’s (or perhapsnowadays, the neuroscientist’s) task was to presenttheir findings to those people who seemed to be trappedat the entrance of the cave.
The author writer believes that the Platonic world ofideas exists within us. It is neither eternal nor immutable,as Plato thought, but rather is created through learningand experience. According to the model of the thalamusand cortex introduced in this paper, one of the functionsof the thalamus is to break down arriving informationinto simple and relevant components similar to Platonicideals. Through a binding process that will be explainedlater, these ideals are recomposed to generate similar orother related patterns. When a new pattern arrives, there
Neural Networks, Vol. 10, No. 7, pp. 1269–1288, 1997q 1997 Elsevier Science Ltd. All rights reserved
Printed in Great Britain0893–6080/97 $17.00+.00
PII: S0893-6080(97)00052-X
Pergamon
1269
Acknowledgements: The author expresses his gratitude to ProfessorShun-ichi Amari, director of the Brain Information Processing Groupin the RIKEN Institute, and Professor Kazu Aihara of the University ofTokyo for their valuable criticisms and suggestions. The author is alsograteful to Shirley F. Taniguchi for reviewing this contribution.
Requests for reprints should be sent to Javier Ropero Pela´ez,Department of Mathematical Engineering, Faculty of Engineering,The University of Tokyo, 7-3-1 Bunkyo-ku, Tokyo 113. Tel./fax03-5689-5752; e-mail: [email protected]

would be some plausible ‘candidate ideals’ which areused in combination by the brain to match the incomingpatterns. During this process a combination of candidateideals would win the competition for matching theincoming pattern. If there is not a perfect match, newideals will be created and stored for later use in dealingwith upcoming patterns.
Although input patterns are perceived as simultaneous,the thalamus will split them into a temporal sequence ofideals. These ideals will be synchronized again throughthe delays in the long dendrites of cortex pyramidalneurons. If two or more patterns are presented simulta-neously, their equal ideals will be associated throughsynaptic reinforcement in pyramidal neurons. Forexample, continuing with the Platonic metaphor, if abush and a tree are intended to be memorized as simul-taneous, the binding of both concepts must be done atthe level of the characteristics or ideals they share, suchas the idea of greenness, height, etc. We are not aware ofthis binding, that is performed in different temporallayers (Thomson & Deuchars, 1994). Therefore, thisinformation must be synchronized in order for us to beconscious of it. This synchronization is hypothesized tobe performed at a cortical level where asynchronous,non-overlapping information coming from the thalamusis delayed. The result of this process would be returnedas an input to the thalamus at the same level of otherinput patterns (Contreras et al., 1995).
Up to this point two processes have been discussed:the generation of ideals and the binding process. As willbe revealed, these two processes are closely related to thetwo modes of thalamic neuron firing: the tonic and burstmodes. An attempt will be made to explain the overallbehaviour of the thalamus and cortex in order to developa new paradigm of neural network (Ropero, Pelaez,1996a, b). This will include an examination of the rulesof interconnectivity of the thalamus and cortex, thesynchronizing role of the nonspecific thalamic nucleus,the modelization of synapses, the interplay between thetonic and bursting mode in thalamic neurons, the specificcomputation in every kind of neuron with a deep analysisof pyramidal processing, the mathematical algorithmsthat describe the ‘ideals’ extraction and the bindingproblem and, finally, the explanation of the neuralcounterpart that supports these algorithms.
2. NEUROLOGICAL FOUNDATIONS
2.1. Rules of Interconnectivity in the Thalamus andCortex
The majority of the information that arrives at the neo-cortex is relayed by the thalamus, an ovoid-shaped struc-ture in the middle of the inverted bowl formedby the cerebral hemispheres. Although most of theafferences from the senses to the cortex are relayedthrough the thalamus, the thalamus receives ten times
more information back from the cortex than from thesenses. Figure 1 schematically depicts the synaptic organi-zation of the thalamus (see Steriade & Llinas, 1988).
Neurons that arrive at the thalamus make contact withtwo different kinds of neuron: thalamocortical and localcircuit neurons. Local circuit neurons exert inhibitionover each other and over thalamocortical neurons. Thetwo types of thalamocortical neuron projections whichgo to cortical and to reticular neurons are excitatory.Each of the thalamic projections to the cortex is accom-panied by a reverse projection from cortex pyramidalneurons to the thalamus, with matching conductionvelocities. The spatial organization of the reverse projec-tions is in a fairly precise reciprocal relationship withthe forward spatial organization (see article by Crick &Asanuma in McClelland et al., 1986).
Reticular neurons in the reticular nucleus of thethalamus (RNT) possess dendrodendritic synapses andexert inhibition over each other, and over thalamocorticalneurons. The inhibition over thalamocortical neuronsis mediated in higher mammals’ LGN parvocellularpathway by inhibitory local circuit neurons whosehypothetical role in learning will be explained later.
The fourth layer of the cortex receives most of theafferences from the thalamus. 40 Hz sparsely inhibitoryspinous interneurons inhibit nearby pyramidal neuronsand seem to be restricted to this area. Chandelierneurons inhibit pyramidal neurons in their axon hillockwhich is the strategic point in which axon potentialsare generated. Basket neurons seem to regulate in acontrol loop manner the threshold of surrounding pyra-midal neurons.
2.2. Synchronization Role of Nonspecific ThalamicNucleus
Intralaminar and paralaminar thalamic nuclei are callednonspecific because their projections are not restricted toa unique sensorial modality. Intralaminar thalamocorti-cal neurons exhibit a firing frequency of 40 Hz (Steriadeet al., 1993). Paralaminar neurones receive inhibitionfrom reticular neurons and project to the thalamus andto cortical layers I and IV (Llinas et al., 1994). First layerprojections are performed mainly in the tuft of pyramidalneurons. Since branches in this tuft converge over thepyramidal dendritic shaft, excitations over the tuft areintegrated and generate a large response that will travelthrough the shaft and will reinforce other stimuli over a40 Hz basis.
2.3. Modelling Synaptic Weights
In the next example,A and B are the spikes in thepresynaptic and postsynaptic space respectively. Theconditional probability ofB given A, P(B/A), couldwell describe the weight which is the value of thereinforcement or weakening in the synapse. The above
1270 J. Ropero Pelaez
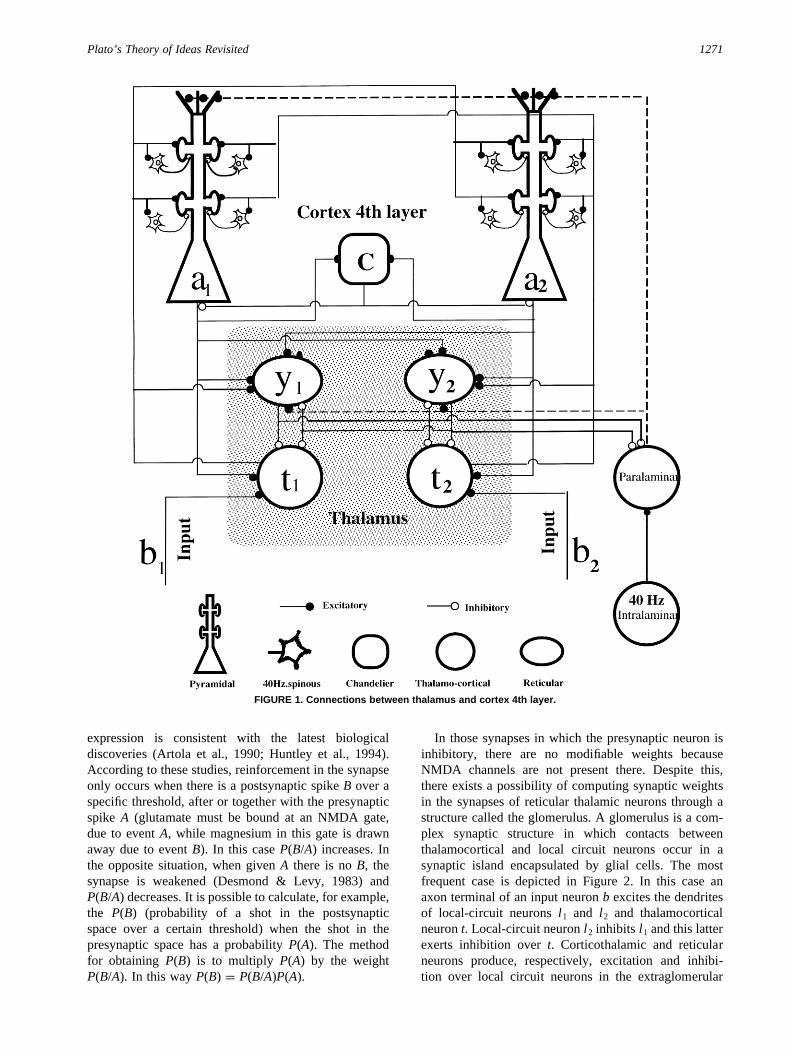
expression is consistent with the latest biologicaldiscoveries (Artola et al., 1990; Huntley et al., 1994).According to these studies, reinforcement in the synapseonly occurs when there is a postsynaptic spikeB over aspecific threshold, after or together with the presynapticspike A (glutamate must be bound at an NMDA gate,due to eventA, while magnesium in this gate is drawnaway due to eventB). In this caseP(B/A) increases. Inthe opposite situation, when givenA there is noB, thesynapse is weakened (Desmond & Levy, 1983) andP(B/A) decreases. It is possible to calculate, for example,the P(B) (probability of a shot in the postsynapticspace over a certain threshold) when the shot in thepresynaptic space has a probabilityP(A). The methodfor obtaining P(B) is to multiply P(A) by the weightP(B/A). In this wayP(B) ¼ P(B/A)P(A).
In those synapses in which the presynaptic neuron isinhibitory, there are no modifiable weights becauseNMDA channels are not present there. Despite this,there exists a possibility of computing synaptic weightsin the synapses of reticular thalamic neurons through astructure called the glomerulus. A glomerulus is a com-plex synaptic structure in which contacts betweenthalamocortical and local circuit neurons occur in asynaptic island encapsulated by glial cells. The mostfrequent case is depicted in Figure 2. In this case anaxon terminal of an input neuronb excites the dendritesof local-circuit neuronsl 1 and l 2 and thalamocorticalneuront. Local-circuit neuronl 2 inhibits l 1 and this latterexerts inhibition overt. Corticothalamic and reticularneurons produce, respectively, excitation and inhibi-tion over local circuit neurons in the extraglomerular
1271Plato’s Theory of Ideas Revisited
FIGURE 1. Connections between thalamus and cortex 4th layer.

neuropil (in Figure 2, inhibition from reticular neurony isrepresented). The following explanation shows thatthe structure in Figure 2a is equivalent to that inFigure 2b in which the weight in the inhibitory synapsefrom the reticular neuron is modelled asP(b/y). The netinput to the thalamic neuron can be measured as theprobability of the excitation fromb minus the probabilityof the inhibition fromy: P(b) ¹ P(b=y)P(y).
In Figure 2c, dotted areas represent depolarizedstructures. In this instance inputb depolarizesl 1 and l 2,but l 2 inhibition overl 1 neutralizesl 1 depolarization. Noinhibition from l 1 over t is produced.
Figure 2d shows what happens when there is areticular inhibition byy. In this instance,l 2 depolariza-tion is neutralized so that it cannot inhibitl 1. Because ofthis, an input fromb excitesl 1, which inhibits the thala-mocorticalt. This can lead us to a rule of thumb in which,when there is reticular inhibition fromy, which is madeeffective by inputb, thenl 1 is active as well. In this way,activity of l 1 is highly correlated to activity iny. Becauseof this, it is possible to model the weightw1 betweenl 1
and b as P(y=b). Thus the excitation fromb over l 1 isP(y=b)P(b), which can be rewritten asP(b=y)P(y). The
net input probability overt is P(b) ¹ P(b=y)P(y), as inFigure 2b.
2.4. Types of Neuronal Firing
2.4.1. Thalamocortical Neurons. Thalamocortical(thal-cor) neurons have two ways of firing (Llinas &Jahnsen, 1982). They may fire tonically when they areexcited above a certain threshold, as happens in the restof the neurons (Figure 3a). The frequency of firing ofthalamocortical neurons during their tonic mode dependson the summation of their inputs. In Figure 3a, thethalamocortical neuron would fire in this mode withprobability P(b) ¹ P(y), which would be the sum of theexcitatory and inhibitory input probabilities.
The other mode of firing is a high frequency( . 250 Hz) burst firing that occurs after the accomplish-ment of a precise sequence of facts. First, a smalldepolarization from rest is required. Afterwards, thisdepolarization must be reduced by an active hyper-polarization or from a reduction of a depolarizinginput. When the neuron is maintained in this state forat least 50–100 ms, the bursting mode is produced by a
1272 J. Ropero Pelaez
FIGURE 2. Equivalence between the hypothetical process carried out in thalamic glomeruli in which inhibitory synapses are notmodifiable (a) and in an artificial model (b) in which these synapses have a weight defined by P(b =y ): t ¼ thalamocortical dendrite; l 1
and l 2 ¼ local-circuit neurons’ dendrites; b ¼ input terminal; y ¼ reticular neuron. (c) and (d) On ‘ b’ being activated, l 1 is active only whenthere is inhibition from ‘ y’. This allows us to calculate weight w 1 as P(b =y ), the influence from ‘ b’ being P(b )P (b =y ) which can be rewrittenas P(y )P (y =b ). This expression is equivalent to an inhibition from ‘ y’ through a synaptic weight P (y =b ), as depicted in (b).

small depolarization, The probability of a high frequencyburst has been modelized in Figure 3b asP(b=y)P(y), Theexpression of the conditional probability ofb given y isused here because the condition forb to allow thalamo-cortical neuron burst firing is the previous inhibitionfrom the reticular neuron ‘y’, which has a probabilityP(y) (Figure 2b).
Reticular neurons exhibit not only the same kind offiring as thalamocortical neurons (Llinas, 1988) but alsoa unique pacemaker-like rhythmicity (Pinault &Deschenes, 1992). Their firing is also characterized bya second rebound in the burst mode whose importancewill be remarked upon during the course of the paper.Threshold accomodation is another specific character-istic of reticular neurons that is necessary to producealternation of reticular neuron activations in the NRT,as will be suggested.
2.4.2.40 Hz Inhibitory Stellate Spinous Neurons.40 Hzinhibitory stellate spinous neurons are present mainly inthe fourth layer of the cortex, which is the region thatreceives most of the afferences from the thalamus. Theirinputs come from the thalamus and their efferences goto the dendritic spines of pyramidal neurons. In vitrointracellular recordings demonstrate a 40 Hz oscillatoryresponse after depolarization. Another characteristicthat is relevant, due to its computational possibilities, isthat the response outlasts the first stimulus but comes toan abrupt cessation in the middle of a second one (Llinaset al., 1991) in a ‘flip-flop’ manner.
2.4.3. Pyramidal Neurons and Sequential Processes.Pyramidal cells represent the most abundant and charac-teristic neuronal type of the cerebral cortex (DeFelipe &Farinas, 1992). They have a long apical dendrite withbulbous-like synapses called dendritic spines connectedto the dendrite by a ‘neck’. One or two excitatory inputscould arrive at a dendritic spine mainly in the spine headand, in many cases, there is also an afference from aninhibitory input neuron near or on the spine neck. Inhibi-tion in this specific place could cut off most of a previous
excitation. In the cortex 4th layer, efferences from thethalamus are used to make contact over 40 Hz inhibitoryneurons and over pyramidal neurons. Looking carefullyat the disposition of the synapses over a spine in thecortex 4th layer (Figure 1) it is possible to hypothesizethat these inhibitory neurons are able to cut off the exci-tation from the thalamus. Spines have the property ofaccumulating calcium. Although this fact has an impor-tant biological meaning (calcium leads to cell death, sospines can be considered as isolated compartmentswhere high concentrations of Ca2þ are stored withoutdamage to the cell function), calcium would be storedas a reservoir of ions for boosting travelling potentialsthrough the dendrite. It also could provide an effectivepath for signal transmission and synaptic reinforcementbetween two nearby spines (Yuste & Denk, 1995; Yuste& Tank, 1996).
1273Plato’s Theory of Ideas Revisited
FIGURE 3. Two types of firing in thalamic neurons. (a) Theprobability of firing in tonic mode is the sum of the probabilitiesof the inputs. (b) In bursting mode, a necessary condition forbursting is the previous inhibition by neuron ‘ y’. Therefore theprobability of bursts in the thalamocortical neuron is modelledby the mathematical expression of the conditioned probability of‘b’ due to ‘ y’ multiplied by the probability of bursts in y.
FIGURE 4. Different theoretical situations in spine signalprocessing. (a) No excitation over the spines; calcium ionsremain inside. (b) Tonic excitation arrives at both spines.Although a little calcium leaves the spines, the frequency ofthe inputs is not high enough to allow the communicationof the two spines through a calcium ion’s path, nor to allowproperly the synchronicity of the spikes in both spines. Thereis not synaptic reinforcement R. (c) Every spine receives a burstof spikes. The high frequency of bursting allows the release ofcalcium to the dendrite and the reinforcement of both synapses.(d) A burst excitation is applied over the spines and over inhibi-tory neurons. The consequence is a depolarization that tries torelease calcium. However, when this ion tries to go to the den-drite the hyperpolarization due to the inhibitory neuron bringsback the calcium to the spine. Neither calcium path nor reinfor-cement is produced. (e) The same case as in (d), but there is abearer depolarization that comes through the dendrite. As it doesnot arrive at the inter-spine gap at the time that the spines receivebursting excitation, the result is as in (d). (f) In this case thebearer wave is synchronized with the spikes’ burst so that itarrives at the inter-spine gap at the same time as the burst firing.LTP is produced and the bearer wave is boosted due to the addedcalcium ion potential.

Taking into account these properties of dendriticspines, several theoretical situations of spine computa-tion have been depicted in Figure 4. These situations willallow us to understand temporal computation of inputsover dendrites. The conclusion is that high frequencybursts of spikes could promote reinforcement in nearbyspines (Figure 4c) while this situation is exceptionalwhen low frequency spikes (Figure 4b) arrive at thespine. When an input volley arrives simultaneously atthe spine and at a 4th layer inhibitory spinous neuron,the inhibitory neuron could hypothetically block both thesignal and calcium diffusion into the dendrite, makingdifficult the reinforcement of synapses of nearby spines,Figure 4d. However, when, synchronously with excita-tion of the spine, a big depolarization passes throughthe dendritic shaft invading the neck of these spines,excitation in a spine would be transmitted faithfully to
the other spines and reinforcement would be accom-plished, Figure 4f. Furthermore, a travelling depolariza-tion would be boosted. If the depolarization is enhancedalong its course by different active spines in the dendriticshaft, this depolarization can be seen as a bearer wavethat carries the information of the sum of different inputs.If every input is required to be summed once, and all theinputs are to be considered in the sum, a top-downsummation must be accomplished, from the most distalinput in the upper layer to the lowest one near the soma.In order to obtain this top-down summation, the inputsmust be ordered according to time; in other words, thefirst input must be in the upper part of the dendrite andthe last input in the lower part. If it is supposed thatevery event inside consciousness not only generates aspecific spatial pattern but also a sequentially orderedpattern, it can be seen that every input to the dendritemight be ordered according to time in a top-downmanner. According to the hypothesis stated below, this
1274 J. Ropero Pelaez
FIGURE 5. Retrograde action potentials can theoretically giverise to a hebbian learning that reinforces ( R) not only theappropriate synapses but also a temporal order of the synapticstimulus. Paralaminar thalamic burst depolarizes the branchesof the pyramidal tuft, and this depolarization is integrated to forma big ‘bearer wave’ that can be boosted if the spine’s synapticdepolarization occurs when this wave passes near the spine. Theconsequence of this boosting might be the reinforcement ofthe active synapses through a retrograde action potential. (a)Various responses due to a paralaminar thalamic neuron in thetuft of an apical dendrite are summed and generate a large wavethat travels through the dendritic shaft. We have considered that,because of the shaft cable properties, the amplitude will diminishand will not be large enough to generate an action potential afterreaching the soma. (b) Let us suppose that, as a consequenceof the paralaminar burst over cortical neurons, some of thesubsequent bursts will arrive at the neuron under study witha fixed delay after the intralaminar burst. Let us say that thesynchronizing burst arrives at time t ¼ 0 while the others willarrive with delays t 1 and t 2. In the figure b 1 and b 2 are orderedaccording to time, from the upper part of the dendritic shaft tothe lower part. In this way the first burst will take place nearer thetuft and the last one nearer the soma. The synaptic locations ofb 1 and b 2 are purposely placed at a proper distance so that the‘bearer wave’, the one originating at the tuft, could reach the firstsynapse at time t 1 and the second at time t 2. In this case, thebearer wave is boosted twice and will become big enough topromote a retrograde potential that will reinforce the previouslyexcited synapses (because glutamate appears to be bound at theNMDA gate, at least until the retrograde spike depolarizes thepostsynaptic membrane above a certain threshold). Although astimulation in spine S may not coincide with the passing of thebearer wave, it is also reinforced by means of the retrogradeaction potential. (c) This case could be helpful for understandingthose in Figure 7. In this case we have burst b 1 and b 2 goingthrough the same axon. They are produced at the same time asbursts b 19 and b 29 that are travelling through the axon below.Since this case is the same as that of Figure 4c, reinforcementis produced. In this particular case every synapse is reinforcedtwice, first by the pair b 1 and b 19 and secondly by b 2 and b 29. Ashas been explained in Figure 4c, the ‘bearer wave’ is notnecessary here for promoting reinforcement.

organization is achieved through a mechanism thatinvolves the reinforcement of the synapses that accom-plish this time-based top-down organization, asexplained in Figure 5.
In Figure 5a is represented the course of a big ‘bearerwave’ generated by the convergence of the 40 Hzparalaminar spikes over all the dendrites of the tuft of acertain pyramidal neuron. This response is weakenedgradually and if, when reaching the soma, it is not highenough it will not generate an action potential. Thisbearer wave can be amplified during its course if it iscoincident with other incoming inputs. In Figure 5b, theparalaminar burstb0 is supposed to produce excitation,not only in the tuft of the neuron but also in the other twoneurons that, due to burstb0, will subsequently fire withburstsb1 andb2 over the neurons depicted in the figurehaving at1 andt2 delay time respectively from burstb0.If the synapses are placed in a position that allowsthe coincidence of the bearer wave produced byb0 andthe excitations fromb1 and b2, the bearer wave willbe boosted twice and probably will be big enough toproduce an action potential in the soma. Together withthis action potential, a retrograde action potential (Stuart& Sakmann, 1994) will return to the apical dendrite,reinforcing the synapses that were previously active.This reinforcement is produced because the neurotrans-mitter glutamate that was previously released in thesynaptic gap is still bound to the NMDA gate whenthe retrograde action potential reaches the synapse(Lester et al., 1990). In this moment the strong potentialrepels the magnesium that was blocking the channel, andcalcium is able to enter the spine, promoting the synapticreinforcement. If, in the case of a retrograde potential,there were synapses that previously were not coincidentwith the bearer wave, they would be potentiated too. Inshort, potentiation will be produced in all the synapsesthat receive inputs if there is a retrograde action potentialdue to a suprathreshold activation of the soma producedby a minimal group of active inputs that boosted thebearer wave.
Another way of producing synaptic reinforcement isby means of the coincidence of impulses in nearbyneurons, Figure 5c. The situation in this figure anticipatessome concepts that will be seen later: two trains ofhighly correlated bursts will reinforce all the synapsesin the figure as many times as the number of bursts ineach train, without any consideration of the order ofbursts nor of the exact point where spines are placed.
Several other questions must be addressed. There isone way to avoid the reinforcement of the synapses inthis last case of coincidence of pulses, or even in the casewhere the proper order of inputs is accomplished. Sincethe action potential generated in the axon hillock is theone that, retrogradely, allows the reinforcement ofthe synapses, if an inhibition is applied in the hillock,no action potential is generated and no reinforcementoccurs. This could give some insight into the role of
certain inhibitory neurons, such as chandelier cells thatstrongly inhibit the cell at that precise point.
The top-down organization of synapses inside a pyra-midal neuron could give rise to an organization ofsequential firing in different pyramidal neurons. As ageneral idea, the order of the input neurons in the apicaldendrite will define the order in the sequence. In Figure 6,a simple case of sequentiality with three pyramidalneurons is shown with examples of clockwise, counter-clockwise, cluster and reverberant states. It tries to drawthe reader’s attention towards the importance of inhibi-tion in sequential processing in the cortex, and towardsthe possible role of inhibitory chandelier and basketneurons. According to this hypothesis, chandelier cellsavoid coincident activation over pyramidal neurons,allowing sequentiality in their inputs. It is remarkablethat chandelier cells inhibit the coincidence of the acti-vation of pyramidal neurons over other pyramidalneurons, but are not capable of avoiding thalamocorticalcoincident activation because afferences from the thala-mus over chandelier cells are unusual. In this way,pyramidal neurons seem to promote sequentialitythrough temporal integration, mainly at a cortical level,as well as detection of thalamocortical coincidences.This hibrid mode hypothesis integrates former ones(Konig et al., 1996).
Inhibition from basket neurons regulates the firingthreshold of pyramidal neurons. Consequently, if thereis little inhibition, neurons fire when receiving thesequential activation of very few neurons. The range ofinput neurons that, depending on the threshold, producethe pyramidal activation define which neurons take partin a certain sequence. If the inhibition is high, a lot ofinput neurons are necessary for reaching the firing thresh-old. In this case the sequence comprises more neurons.If the inhibition is low, very few inputs produce thepyramidal firing, and the sequences comprise onlythese few input neurons. In this case, different clustersof neurons can produce different sequential loops at thesame time.
In the cortex 4th layer, the present work suggests thatnot spines but rather clusters of spines (see Yuste &Tank, 1996) are also ordered through time in the wayexplained above. In this case, the reinforcement is notdue to a retrograde action potential but, instead, is dueto the simultaneous coincidence of the bearer wave andthe activations of the inputs over the same spine as inFigure 4f. While in the previous case there was rein-forcement in active spines whose inputs were not coin-cident with the bearer wave, in the present case of the 4thlayer, reinforcement does not occur if the activation ofthe inputs is not coincident with the synchronizingwave (Figure 7a). If the same thalamocortical axonsmake contact over different spines on the section of adendritic shaft in the 4th layer, the spatial organizationof the spines will mimic the temporal organization ofthe bursts in thalamocortical axons, Figure 7b and 7c.
1275Plato’s Theory of Ideas Revisited

The hypothesis that thalamocortical axons conveyprecise sequences of bursts will be explained in thiswork.
Sequentiality, as will be shown, is promoted not onlyat a cortical level; the thalamus could also promote
sequentiality due to the possibility that it could decom-pose input patterns into a sequence of raw ‘ideals’ thatare produced in order through an iterative sequentialmethod that will be explained.
These two reasons, sequentiality promoted by
1276 J. Ropero Pelaez
FIGURE 6. A temporal organization of synapses in the apical dendrite of pyramidal neurons could cause a group of these neurons to firesequentially. (a) In this case a neuron will fire if the 40 Hz neuron fires first and the other two inputs receive excitation in the appropriateorder (threshold ¼ 2; 2 inputs). In this group of neurons the sequence of firing is clockwise, thus: 1 2 3 1 2 3 1 2 .... The firing rate of eachneuron is 40/3 Hz. (b) In this case the three neurons fire simultaneously (in an ‘epileptic’ manner) at 40 Hz because of the coincidentactivation of two inputs in all neurons. (c) In a combination of the above cases, there are two more connections in each pyramidal neuron.There is also a chandelier cell (with an arbitrary threshold ¼ 2 input neurons). When it receives two or more excitations it inhibits allpyramidal neurons, avoiding coincident activation over them. Here there are two possible sequences of firing: clockwise as in (a), orcounterclockwise 3 2 1 3 2 1 3 2 ... (which is performed through broken lines). The system is hooked in one of the sequences dependingon which organization synchronizes first with the 40 Hz synchronizing input. However, it could be forced into one sequence by meansof an external activation. (d) Three basket neurons are added. Basket neurons only regulate the threshold of pyramidal neurons, insteadof cutting off their firing. They are inhibiting according to the surrounding level of pyramidal activation in a control loop manner.Subsequently the pyramidal neurons’ threshold increases when it is more inhibited. Depending on the pyramidal neurons’ threshold,the sequences of firing vary from an ‘epileptic’ behaviour (threshold ¼ 0 when at the very beginning the network was in a steady state) toa wide sequence of three neurons as in cases (a) or (c) (threshold ¼ 2 at the end) through three oscillatory possibilities 1 2 1 2 1 ...,1 3 1 3 1 ... or 2 3 2 3 ... (when threshold ¼ 2 in a transitory state). ln these cases the threshold is proportional to the number of activebasket neurons.

pyramidal inputs and a sequential decomposition ofinput patterns in the thalamus, are why thalamocorticalinputs to the cortex might be ordered according to timein dendrites.
3. IDEALS AS AXES IN PATTERNVECTOR CODING
In the Myth of the Cave Plato describes the ‘world ofideas’ in which eternal and immutable patterns give riseto various phenomena in nature. Similarly, there exist inthe brain specific locations that store very abstract
codifications of the world, as in the retina which decom-poses the colour into the three basic types, or in thetonotopic and visuotopic map in the cerebral cortex.According to several authors (DeValois & DeValois,1988), these visuotopic maps, in which cortical areasrespond differently to different spatial frequencies ofpatterns, are the result of Fourier decomposition, whichis a kind of mathematical process known as ortho-gonalization. One way to think of this is that all theinformation entering the cortex must previously undergosome kind of orthogonalization.
Figure 8 is an example of orthogonalization
1277Plato’s Theory of Ideas Revisited
FIGURE 7. Three different theoretical situations in layer IV signal processing. (a) Two trains of bursts coming through two different axonsare represented at both sides of the neuron. Bursts b 1 and b 19 are produced at the same time as b 2 and b 29. Since the first couple of bursts,b 1 and b 19, arrive synchronously at the spines p 4 and p 3 but not at the same instant as the ‘bearer wave’ generated by b O, synapses inthese spines are not reinforced (see also Figure 4e). Synapses in p 1 and p 2 are not reinforced because they have not already beenstimulated. (b) At time t 1, b 1and b 19 arrive at the synapses in p 2 and p 1 respectively synchronously with the bearer wave produced by b 0.At t 2 the same occurs by means of bursts b 2 and b 29 over synapses in p 4 and p 3. The result is a reinforcement R of all these synapses (seealso Figure 4f). (c) This is the same case as (b) but with an extra pair of spines. Since these spines p 5 and p 6 are not properly placed, thebearer wave is not coincident with the bursts and reinforcement is not produced. Synaptic depression in the spines of both pyramidaland sparsely spinous interneurons will hypothetically cause these spines to be disconnected. In this way spatial and temporal synchro-nization can be produced during development. It should be noted that in the above cases, in which spines act as coincident detectors, aretrograde potential is not necessary for reinforcing an appropiate spatial order for placing incoming inputs over the dendritic shaft, asseen in Figure 5.

and vectorial coding adapted from Churchland andSejnowsky, 1992. It shows that an infinite number offaces can be defined by merely varying the intensityof the three basic characteristics, or ideals, that aredepicted in the figure. To representn ideals, ann-dimen-sional space would be required.
Another fact that leads us to believe that there is somekind of orthogonalization process taking place in thebrain is that the orthogonalization of input infor-mation is a necessary condition to achieve ahebbianlearning in pattern associator models (McClelland et al.,1986). It seems that the brain utilizes this kind ofhebbian learning instead of complicated neural networkalgorithms.
Events in the brain occur by means of impulse rates,making the use of statistics in the present research seemconvenient. But if orthogonalization is consideredinside statistics it is necessary to redefine some importantstatistical notions, so that statistics can be seen as abranch of euclidean algebra. This topic is treated at theend of the paper to provide the reader with a generalbackground without diving into abstract mathematics.Several results from that section will be utilized in thecourse of the paper.
On the basis of that section, nervous informationwill be treated as statistical events. For example, ifthe input layer is a matrix ofm neurons, any of the 2m
subsets that could be accomplished with them neuronswill be considered an event. Every input would berelated to the others in an auto-associative manner, as
if all the inputs constituted a unique pattern thatcould be completed coherently if some of the inputswere missing.
Every input is defined as a vector ofn components,nbeing the number of orthogonal neurons in the reticularneuron layer. Orthogonality can be considered asfollows.
Two patterns,A andB, are orthogonal when the scalarproduct, defined as the number of coincidences of thetwo of them, is zero (see Section 9):
n(~Ad ~B) ¼ 0
In other words, two patterns are orthogonal if the prob-ability of their coincidence is zero. All the neuronsyi
in the reticular layer could be considered orthogonal ormutually exclusive because there exists a competitiveinhibition between them (when one neuron fires theothers do not). Orthogonality in reticular neuronsinteractions allows these neurons to be considered asaxes in a coordinate system.
According to these ideas, inputb is defined as:
~b¼ k1~y1 þ k2~y2 þ :::þ kn~yn
In this pattern ~b, all the components are mutuallyexclusive and define a partition of the whole space.Every factorki is a flag that exhibits the partial contribu-tion of the reticular activation~yi , to the pattern~b; in otherwords, k1 ¼ P(~b=~yi). It is also possible to compute theprobability of pattern~b performing its scalar product
1278 J. Ropero Pelaez
FIGURE 8. Vector coding. Three axes define an infinite number of faces.

by the normalized diagonal vector~I (see Section 9):
P(~B) ¼ n(~Bd~I )
through which the usual expression of a vector in termsof its components is obtained:
P(~b) ¼ P(~b=~y1)P(~y1) þ P(~b=~y2)P(~y2) þ :::þ P(~bn=~yn)P(~yn)
4. HOW IDEALS ARE OBTAINED FROMINPUT PATTERNS
According to the author’s hypothesis, every newpattern that arrives at the thalamus determines, in part,the processing of subsequent patterns, and, conversely,is processed according to the organization defined byprevious patterns. Similarly, in the Gram–Schmidtorthogonalization process, the first pattern will definethe first axis of a coordinate system, with successivepatterns helping to find the orthogonal basis over whichall the vectors of a certain euclidean space can bedefined.
If the generic scalar product in the Gram–Schmidtequation is substituted by the previously defined scalarproduct, then the next version of Gram–Schmidt isobtained (see Section 9):
~y1 ¼ ~b1
~yr þ 1 ¼ ~br þ 1 ¹∑r
i ¼ 1P(~br þ 1=~yi)~yi
Calculating the probabilities of the obtained axes, theresult is:
P(~y1) ¼ P(~b1)
P(~yr þ 1) ¼ P(~br þ 1) ¹∑r
i ¼ 1P(~br þ 1=~yi)P(~yi)
Orthogonalization allows all the input patterns to berepresented in a coordinate system. During the ortho-gonalization process thal-cor neurons and RE neuronshypothetically discharge in the tonic mode. This iscorroborated by the fact that the presentation ofnew information is normally done in the awakenedstate in which the tonic mode is also more frequentlydetected.
In the orthogonalization process, thalamocorticalneurons serve as inputs and reticular neurons serve asthe axes over which the information is projected. Forthe sake of simplicity, Figure 9 depicts only the connec-tion between these two types of neurons.
In this very simple example, an analysis is performedof the computation produced when the first two patternsare input. In the first patternb1 (Figure 9a.1 and 9a.2), thefirst three thalamic neuronst1, t2 and t3 are activatedwith a certain probability that is considered to be thesame in all of them. The second pattern presentation(Figure 9b.1) corresponds to the setb2 that, in thisexample, is characterized by the activation of thalamicneuronst2, t3, t4 and t5. Inhibitory connections fromreticular neurons are not modifiable but behave as ifthey were, according to the description of the glomerulusgiven in Section 2.3. The weight in the glomerulus iscalculated as the probability of an input neuronb being
1279Plato’s Theory of Ideas Revisited
FIGURE 9. This figure illustrates the Gram–Schmidt (G–S) pro-cess of axis generation. Only active connections are shown. (a.1)Pattern b 1 is presented. Reticular neuron y 1 is selected througha competitive process of inhibitory neurons. Connectionsbetween thalamocortical ( t) and reticular neurons ( y) are rein-forced. The first iteration of G–S is computed: P(y1) ¼ P(b1).(a.2) Feedback connections are trained using a process that issimilar to a delta rule. (b.1) The second pattern b 2 is input.Candidate reticular neurons are selected. (b.2) Reinforced feed-back connections of y 1 inhibit inputs B2
2 and b 32 . The remaining
inputs activate y 2. The probability of y 2 is calculated as follows:P(y2) ¼ P (b2) ¹ P(b2=y1)P (y1), which is the second iteration ofG–S. The activity of y 1 decreases because the activity of inputsalso diminishes. This situation prepares y 1 to start firing in aburst mode if an excitation arrives from the paralaminar nucleus.(b.3) A little later, y 2 also diminishes its activity. (b.4) y 1 and y 2
commence firing in a burst mode due to an excitation from theparalaminar nucleus.

conditioned by a reticular neurony, P(b=y), so that theinhibition by y over the thalamic neurons isP(b=y)P(y),as was discussed in Section 2.3.
When the first pattern~b1 is presented, Figure 9a.1, acertain reticular neurony1 in the competitive reticularneuron layer wins the competition to represent theinput. During presentation, the conditional probabilitiesP(y1=tj) for each of the relay neuronst1j are reinforced.After reinforcement, every time~b1 is presented it willgenerate the activation ofy1, such thatP(y1) ¼ P(b1),which is the first iteration in the Gram–Schmidt algo-rithm. The weights of the inhibitory backward projec-tions P(~b
uj =~yi) are also reinforced (u being each of the
input units of the input pattern~bj) (Figure a.2), and finallythe thalamic neurons are silenced.
When the second input pattern~b2, Figure 9b.1, ispresented, thalamic neuronst2 and t3, that were activeduring ~b1 presentation, project towardsy1 through thepreviously reinforced synapses. Subsequently, the rein-forced backward inhibition will causet2 and t3 to rest,Figure 9b.2.
During the presentation of the second pattern~b2
another reticular neurony2 wins the competition insidethe RNT. It could be argued that the previously selectedreticular neuron,y1, should win the competition to repre-sent the second input pattern because of the previouslyreinforced forward connections. However, reticularneurons possess threshold accommodation, a uniqueproperty among thalamic neurons (Mulle et al., 1986,p. 2140). In that paper it is shown thattonic firing isalways associated with a marked increase of the spikethreshold, as is demonstrated in an experiment in whichthe first spike was fired at¹50 mV while the tenthspike reached threshold at¹38 mV. This means that acontinuously increasing input voltage is required formaintaining the spiking rate of reticular neurons. In thepresent case (during the presentation of the second inputpatternb2), y1 diminishes its firing rate because inputst2
and t3 go rapidly to rest. If the slope of the net inputactivation overy2 is bigger than that overy1, y2 winsthe competition in the reticular layer. The probabilityof reticular neurony2 being activated, which correspondsto the activation of inputs~b
42 and ~b
52, is calculated as
the probability of pattern~b2 after the inhibition ofinputs~b
22 and~b
32 from y1:
P(~y2) ¼ P(~b2) ¹ P(~b2=~y1)P(~y1)
which is the second iteration of the Gram–Schmidt equation. Inhibitory connectionsP(~b
22=~y2)
and P(~b32=~y2) from y2 over t2 and t3 would also be
reinforced if there were no inhibition byy1 (activatedby b22 and b23) over y2, which counterbalances thistendency.
y2 now starts firing, and, as a result, the backwardinhibitory connections to the thalamus are reinforced.This inhibition causest4 and t5 to be at rest again;
consequently, having no active inputs,y2 decreases itsactivation, as occurred previously in the case ofy1 (seethe upper part of Figure 9b.2). The sudden drop in inputsignals, toy1 and subsequently toy2, inactivates themgradually (Figure 9b.3), and the reticular neuronprepares to start firing in a bursting mode after a slightdepolarization arriving from the paralaminar nucleus(the paralaminar neurons seem to burst when, beinginhibited by reticular neurons, they receive excitationfrom 40 Hz intralaminar neurons, filtering in thismanner the activity of the latter according to the stateinside the thalamus). Although thalamocortical neuronsshould also be in the burst mode, brain stem reticularformation, through its cholinergic inputs, preventsthalamocortical neurons from bursting (McCormick &Prince, 1987a, b). On the contrary, these samecholinergic inputs facilitate the bursting mode in reticu-lar neurons. After depolarization, the bursting modecommences earlier and with a stronger frequency inthose neurons that were inactivated earlier or whichhad been further inactivated. In this case, in whichy1
and y2 suffer a similar inactivation,y1 changes intoburst mode first because it was the first to be inactivated,Figure 9b.4. This bursting mode in reticular neuronsrepresents the beginning of a new process for bindingthe different inputs and features that characterize acertain input pattern. This binding ensures that a certainpattern will be remembered as a whole, with innercoherence between the different constituent ideals andbetween the inputs that constitute every ideal. Thistopic will be discussed in Section 5. However, it is stillnecessary to remark further on the process explainedabove.
Once the selection of axes is achieved in a competitiveneural network-like manner, the process of correlatinginputs and outputs is a typical example of a hebbiandelta rule (a good review of this work can be found inMcClelland & Rumelhart, 1988, pp. 84–95) in which
the input patterns are coded in the reticular neuron layerand the target patterns are those input to the thalamus.According to the delta rule, the weights in the networkare proportional to the correlation between the inputsand a certain error computed in the output neurons.This error is equal to the difference between the targetpattern and the actual network output. In the thalamus,this error is computed by thalamocortical neurons. Inthem the pattern of inhibitions by the reticular neuronsover thalamocortical neurons corresponds to the outputin the delta learning rule, while inputs from senses tothe thalamus correspond to the target pattern. Accordingto the delta rule, when the input patterns used in trainingare independent, the correct association with the targetpatterns is obtained, thereby avoiding contaminationbetween the desired output and the output to otherlearned input patterns. In the present case, during thepresentation of pattern~b1, vector (1,0) that correspondsto reticular neurony1 activation was obtained. During
1280 J. Ropero Pelaez

the presentation of~b2, reticular neuronsy1 andy2 weresequentially activated, giving rise to vector (1,1). Bothvectors (1,0) and (1,1) constitute an independentinput set. During each learning pattern presentation anew axis is obtained, but former axes are also recalled,giving rise to a new independent vector. These vectorsare associated with each target pattern~bj through thedelta rule. Moreover, when a testing pattern in RNTis a linear combination of the independent vectorsobtained during learning, the same linear combination(in this case between the outputs) explains the outputto the testing pattern. A certain independent vector inthe reticular nucleus of the thalamus codifies a linearcombination of the different features of an incomingpattern. Each component of this combination is stored,as will be explained in Section 5, in the connections frompyramidal neurons to reticular neurons.
5. BINDING
Input patterns to the thalamus are split into characteristicfeatures or ideals, each one associated through a deltarule with an independent vector in the RNT. One pre-condition to preserve the identity of each pattern is tobind the activation of different neurons inside a certainfeature and to bind the different features inside apattern. Pyramidal neurons in the inferior temporalcortex provide an example of this type of binding.Lesions in this region produce prosopagnosia, in whichpatients are able to identify the parts of a face and evenspecific emotions expressed on it, but are incapableof identifying a person’s identity. Identity can bedefined as the concurrent activation of certain inputsand ideals at a specific instant. The concurrent acti-vation of two input neuronsb1 andb2, or in general oftwo parts of an image, can be measured asP(~b2 ∩ ~b1).In this section it will be suggested that a pyramidalneuron can produce this computation, firing accordingto a certain combination of ideals which constitute theidentity of a pattern.
Binding any two inputsb1 and b2 also means thatthe conditional probabilitiesP(b1=b2) or P(b2=b1) couldbe computed and stored in memory. In general, everyinput can share different ideals or, in other words,different ideals can be partially represented by a certaininput. The only way by which the above computationcan be accomplished is by projecting every patternover the orthogonal axes previously found. The generalexpression for a conditional probability is:
P(~b2=~b1) ¼
P(~b2 ∩ ~b1)P(~b1)
And, expressing every projection ofbj over theyi axesP(~bj =~yi)~yi in the more compact~bji , the conditionalprobability expression of the two eventsb2 and b1 interms of theirn orthogonal components is derived (see
Section 9):
P(~b2=~b1) ¼
P(~b1 ∩ ~b2)P(~b1)
¼
∑n
i ¼ 1P(~b1i ∩ ~b2i)
P(~b1)
The computation of this algorithm inside the thalamuswill now be described using Figure 10. For this purpose,the explanation will first describe the learning process(having all the inputs) followed by the test processin which omitted inputs will be obtained through thepresentation of the remaining inputs.
Figure 10 does not aim to be completely exhaustivebut rather to facilitate the reader’s understanding.Near some thalamic synapses the value of their weightis written, while the equation near the spine correspondsto the value of the projection of an incoming input overa certain axis.
5.1. Learning Process
Incoming information promotes the activation of certainreticular neurons that constitute the axes over which theinformation is going to be projected. During activation,these axes inhibit thalamocortical neurons, completelycounterbalancing their excitation through a sculpturingprocess explained in Section 4. For example, the prob-ability of the excitation overt1 from inputb1 in Figure 10is exactly equal to the probability of the inhibition fromy1 andy2:
P(~b1) ¼ P(~b1=~y1)P(~y1) þ P(~b1=~y2)P(~y2)
which is the theorem of total probability. After thisinhibition, thalamocortical neurons return to the restinglevel.
Reticular neurons are awakened through the Gram–Schmidt orthogonalization process. When all the selectedreticular neurons simultaneously receive excitation fromparalaminar neurons, the first inactivated is the first tochange its tonic firing mode into burst firing (Llinas &Jahnsen, 1982). The remaining reticular neurons willproduce burst firing in the order in which they wereinactivated. In the present case of two reticular neurons,y2 fires first (instant 4 in Figure 10), inhibiting deeplyand abruptly all the thalamocortical neurons that, as weconcluded, were at a resting level. This deep inhibitionis necessary to counterbalance projections from thebrain-stem reticular formation that try to keep thalamo-cortical neurons in the tonic mode. In this situation ofdeep hyperpolarization, if an input volley arrives tot1
and t2, they start bursting (instant 5). The bursts int1
and t2 are then conditioned by the burst iny2 (see alsoSection 2.4.1 and Figure 3b). Bursts int1 andt2 due toy2
correspond, respectively, to the projections ofb1 andb2
overy2, b12 andb22 (Figure 10, top). Therefore burstb12
(which occurs with probabilityP(b12)) and burstb22 passbeside spinesp3 andp4, respectively (instant 6), without
1281Plato’s Theory of Ideas Revisited

1282 J. Ropero Pelaez
FIGURE 10. This figure explains the process of selection of ‘ideals’ (axes) and the binding process of two inputs. These tables areintended to be read column by column from instant 1 to 12. Every column is designed to form a sentence: the first column will say ‘‘Atinstant 0 the neurons t 1 and t 2 that receive excitation/inhibition [in this case excitation] from b 1 and b 2 and a net input VO, V1 respec-tively...’’. Dashes must be skipped when reading. Other abbreviations: f.e. ¼ for example; res. ¼ respectively. Data that is squared in bothtables must be compared carefully to understand the difference between the learning and the testing process.

producing any effect, as explained in Figure 7a andFigure 4e; but, when arriving at spinesp1 andp2 (instant7), their simultaneous coincidence (their intersection)boosts the 40 Hz response that has travelled up to thispoint through the dendritic shaft (see also Figure 7b). Atthis moment burstsb12 and b22 also reinforce thesynapses in the spine (the tonic mode firing of the thala-mocortical neuron was not strong enough to allow rein-forcement during the previous process of axis selectionas seen in Figure 4b). While the consequences of theburst ofy2 reticular neuron evolve, the burst mode ofy1
produces its first barrage, inhibiting strongly the samethalamocortical neuronst1 andt2 (instant 8) and therebyproducing the projections ofb1 andb2 over axisy1 (thatare led through the path depicted as broken lines in thefigure). These projections correspond to the second pairof travelling burstsb11 andb21. The intersection of thesetwo bursts, when arriving at spinesp3 and p4, booststhe same bearer wave that was previously boosted bythe intersection ofb12 andb22 (see also Figure 7c). Thebearer wave then conveys the information of the sum ofthe intersection of projections overy1 andy2. This sumdefines the intersections of the inputsb1 andb2:
P(~a1) ¼ P(~b1 ∩ ~b2) ¼ P[(~b11 ∩ ~b21) þ (~b12 ∩ ~b22)]
The concurrent activation of both inputs is what definesthe identity of the input pattern as a whole. This meansthat a pyramidal neuron only responds to a certain com-bination of inputs, firing concurrently each of them witha specific probability. The activity of each pyramidalneuron expresses the inner coherence that defines acertain input pattern, as would a signature. The activationof the pyramidal neuron represents the input patternitself, or the coherent characteristics that define it,without considering other circumstantial details. Forexample, in the identification of a face neither the lengthof the hair nor the type of ear rings or the luminositywill be taken into consideration. In this sense, the outputof the pyramidal neuron can be understood as a transfor-mation of the input pattern through a coherence functionover patternb that will be in the notationb*:
a¼ bp
5.2. Testing Process
During the testing process onlyb1 is presented. Asb1 isconsidered to share axesy1 andy2, both are alternatelyactivated. Wheny1 andy2 burst over thalamocorticalt2,burst firing does not result sincet2 is without the excita-tory input that is necessary to deactivatet2 and startbursting. Only the projections of the unique inputb1
over y2 and y1, b11 and b12 respectively, approach thespines sequentially. When arriving at the spines, andmultiplied by the weights, they produce the same resultas ifb22 andb21 had also arrived at the spines. In the caseof spinep3, if the probability of the input to the spine
P(~b11) ¼ P(~b1=~y1)P(~y1) is multiplied by the weightP(~b11 ∩ ~b21)=P(~b11) (that was computed in the spineduring the learning process) the result isP(~b11 ∩ ~b21),as if the other inputb21 had just arrived at spinep4. Inthe case of spinep1, the result isP(~b12 ∩ ~b22), as if therewere a stimulus in spinep2. These two fictitious inter-sections produce, however, the firing of the pyramidalneuron that encodes the intersection of inputsb1 andb2:
P(~a1) ¼ P(~b1 ∩ ~b2) ¼ P[(~b11 ∩ ~b21) þ (~b12 ∩ ~b22)]
This result is the same as that of applying a certain con-ditional probability P(~b2=
~b1) over the input pattern~b1
which occurs with probabilityP(~b1)) so that
P(~b1)P(~b2=~b1) ¼
∑n
i ¼ 1P(~b2i ∩ ~b1i) ¼ P(~b2 ∩ ~b1)
In this way the expression of a conditional probabilityis identified in the computation between thalamus andcortex, which was the purpose of this section.
When the probability of pyramidal neurona1 is multi-plied by the weight between thalamocortical neuront2
and pyramidal neurona1, which is P(~b2=~a1) ¼ 1, theresult is the excitation overt2:
P(~a1)·P(~b2=~a1) ¼ P(~a1) ¼ P(~b1 ∩ ~b2)
1283Plato’s Theory of Ideas Revisited
FIGURE 11. The second barrage of the burst in reticular cellsseems to play a specific role in the reinforcement of backwardconnections from the cortex. (a) Bursts of y 1 hyperpolarize t 2
and t 3, creating in them the conditions for burst firing thatcommences after an excitation from the paralaminar nucleus.(b) y 2 also facilitates the burst mode in t 4 and t 5. (c) Corticotha-lamic connections are reinforced, silencing incoming inputs.(d) Weights from pyramidal to thal-cor are calculated asP(b 2p
2 =a) and from pyramidal to local circuit cell l 1 as P(b 2p21 =b
p).

This is the same result as that of multiplying~b1 by aconditioned probabilityP(~b2=
~b1) stored in the network.In this case, the pattern~b2 is reconstructed, but only tothe extent of spiking with a probabilityP(~b1 ∩ ~b2) whichis not the original probabilityP(~b2). However, this prob-ability defines the contribution oft2 to the identificationof the input pattern.
Pyramidal neurona1 also fires in a burst mode overy1
and y2. When doing this, the conditional probabilitiesP(y1=a1) and P(y2=a1) are stored in its synapses. Theseprobabilities represent the extent to which a certainfeature or ideal contributes to the identification of aninput pattern. As mentioned at the end of Section 4,one of the characteristics of a neural network trainedthrough a delta rule is that it is possible to obtain anoutput pattern that is a blend of several features, eachof which is associated with an orthogonal input, if thissame blend of orthogonal inputs is introduced to the net-work. In this case the orthogonal inputsyi are combinedas follows:
~bp¼ P(~b
p=~y1)~y1 þ P(~b
p=~y2)~y2
This combination of activations of reticular neuronsproduces an output pattern of inhibitions over thalamo-cortical neurons (through the weights stored duringthe Gram–Schmidt process,P(bj =yi)) that reproducespattern ~b
p. Therefore the sculpturing inhibition of
reticular neurons over thalamocortical ones is accom-plished through an ideal image of the input pattern~b.
Some remarks on the adjustment of backward connec-tions from pyramidal neurons to thalamic neurons arestill necessary. For this purpose the example given inSection 4 will be continued. In the upper part ofFigure 11c, the typical response of a reticular neuron toan input consisting of a depolarization, a hyperpolari-zation and another depolarization is shown. The responseto a slight excitation after hyperpolarization is composedof a burst with a rebound around 40 ms later. Whilethe first reticular burst produces the hyperpolarizationof thalamic neurons so that the projections of the inputpatterns over the reticular axesy1 and y2 can be trans-ferred to the cortex (Figure 11a and 11b), the secondburst seems to play a special role over the reinforcementof the connections from the pyramidal neuron to thereticular and thalamocortical neurons (Figure 11c). Itseems as if the rebound waited until the pyramidalneuron sent its message back to the thalamus so thatboth stimuli could be related through hebbian rein-forcement. The process of this reinforcement is thefollowing: the bursting reticular neurony1 inhibitingcell l 2 (Figure 11d) allows forl 1 to inhibit thalamo-cortical cell t with a probabilityP(~b
22=~y1)·P(~y1) (see also
Figure 2). If the influence of all the inhibitions over thethalamocortical neuron (in the example fromy1 andy2) iscomputed, then the probability of the whole inhibition
equals that from the stimulus~b2p
2 :
P(~b2p
2 ) ¼ P(~b22=~y1)P(~y1) þ P(~b
22=~y2)P(~y2)
Depending on the strength of this inhibition overt2, aproportional bursting frequency is produced whithinit. For example, the postsynaptic burst int2 with prob-ability P(bp
22), combined with a presynaptic burst frompyramidal neuron ‘a’ with frequency P(bp), producesthe synaptic weightP(~b
2p
2 =~b
p) in their synapse. The
synaptic weight betweena and l 1 is P(~b2p
21=~b
p). This
second type of connection ensures that when there isno inhibition over t2 from reticular neurons, the same
1284 J. Ropero Pelaez
FIGURE 12. (a) Two pyramidal neurons a1 and a19, each onerepresenting a certain coherent input pattern, are related bymeans of a third pyramidal neuron a2. The reverberant activityof the two pyramidal neuron layers in the figure ensures themaintenance of corticothalamic projections that complete(lined areas) partial input patterns (arrows). These loops of rever-berant activity tend to vanish because pyramidal neurons recallsurrounding basket neurons that inhibit them. This fading iscounterbalanced if pyramidal neurons of higher layers arerecruited (b) so that these loops become wider. Connectionsbetween layers are the same as between the neurons depictedin Figure 6a. their activity being regulated by the same rules ofsequentiality. Higher layer neurons are able to participate inseveral reverberant loops (comprising lower layer neurons),correlating them. The highest layer neurons cannot be correlatedby upper neurons but form a large loop that is hypothesized tobe the hippocampal formation.

inhibition is produced overt2 to allow it to enter into thebursting mode, in this case promoted by a pyramidalneuron (the reticular inhibition is absent overt2 if ~b
22 is
not present). Another interesting effect is that when thethalamocortical loop is reinforced, pyramidal neuronsmute sensorial inputs hypothetically throughl 1 (Sillitoet al., 1993); see Figure 11c, in which the input arrowsare omitted.
6. TRANSIENT MEMORIES MAINTAINEDTHROUGH THE ACTIVATION OF
CORTICAL LOOPS
In Section 5 was proposed a mechanism by whichpyramidal neurons that represent a certain coherentpattern can reconstruct a partial one by means of thefeedback stimulation of thalamocortical neurons. It wasalso explained that pyramidal neurons can mute sensorialinputs so that, transiently, the input to the thalamuswould beb* (from cortex) instead ofb (from senses).
Figure 12a shows that two pyramidal neuronsa1 anda19, each of which represents a certain unitary contentor identity, can also be related through a thirdpyramidal neurona2 if both contents are simultaneous.In the example, pyramidal neuronsa1 and a19 wererecalled, respectively, by a fragmented presentation oftwo patterns depicted as continuous and dotted arrowsrespectively. The activation of pyramidal neuronsa1 anda19 complete the patterns 1 and 2 over thalamocorticalneurons, muting actual inputs. At the same time, theconcurrent activation ofa1 and a19 over a2 reinforcessynapses ina2, as explained in Section 2.4.3 andFigure 4c. Reciprocally pyramidal neurona2, firingover the previously activateda1 and a19, activates thebackward path. If only the inputs represented by the con-tinuous arrows were present, both input patterns wouldbe reconstructed, because only one input toa2 producesthe same response as if both inputs were present (seeSection 5.2). These reconstructed patterns would beonly evanescent images if the inputs completely dis-appeared. However, these memories are maintainedactive in the reinforced loop of pyramidal neuronswhena1 anda19 produce the shot ofa2 and this neuronfires again overa1 anda19. Despite this, these reverberantloops tend to desappear because of basket cell inhibitionunless pyramidal neurons recruit higher level neurons(Figure 12b).
In this latter case neurona3 is recruited and is involvedin a circuit in which the three layers of neurons fire in acycle. This cycle is the same as that previously shownin Figure 6a. The activity of pyramidal neurona3 notonly contributes to the maintainance of this loop butalso to the engagement of other loops. The pyramidalneurons of higher layers engage wider loops. It isassumed here that the hippocampus is hypotheticallycapable of coordinating inside other loop the activity ofneurons in the highest layer.
7. EXPERIMENTAL RESULTS
A simplified version of the model described above hasbeen implemented in a computer program. It comprisesthirty modules with a thalamocortical, a reticular, apyramidal and a basket neuron in each module. A twohemispheres version with callosal neurons connectingboth of them was also accomplished. The followingconclusions should be noted.
1. The model learns non-orthogonal patterns.2. Eliminating or modifying connections confirms the
previously described properties of different kind ofneurons.
3. After learning, when inputs are omitted and theneurons’ threshold is decreased, the model continuesits inner dynamic mixing of previously learnedpatterns in a continuous way.
8. DISCUSSION
The hypothesis of thalamus and cortex computation justpresented allows incoming patterns to be considered asvectors that are unequivocally characterized by theirprojections over a coordinate system. The axes in thisbasis (the ‘ideals’ in Plato’s theory) are represented bythe firing in each reticular thalamic neuron. These idealsare extracted from input patterns through a Gram–Schmidt orthogonalization process that is accomplishedin the thalamus. The association between an activereticular neuron and the feature or ideal that it definesis achieved through a delta-rule-like process in whichdendritic glomeruli have a special role. Binding thedifferent ideals that are present in a certain input patternis achieved in pyramidal neurons. Since Gram–Schmidtselection of axes is an iterative process accomplishedover time, the exact temporal sequence of axes or idealsfor a determined pattern is crucial, and this sequenceis stored like a certain sequence of inputs in layer IVpyramidal apical dendrites.
This process also allows the binding of the inputneurons that give rise to a certain ideal, and of the differ-ent parts of an input pattern. When a certain pyramidalneuron shoots, it produces an ideal image of the inputpattern over thalamocortical neurons. If only a part ofthe input pattern is presented, it is fully reconstructedthrough the projections from pyramidal to thalamicneurons. The firing of pyramidal neurons also representsa measure of the coherence of a certain pattern. Severalcoherent patterns represented by different pyramidalneurons are related through the activation of pyra-midal neurons of higher layers. Cycling loops of neuronsin different cortical layers maintains the integrity ofmemories, and higher layer neurons engage lowerlayer cycling loops. Hypothetically, the hippocampuswould engage the activity of pyramidal neurons in thehighest layers, giving a unified representation of currentsensorial events.
1285Plato’s Theory of Ideas Revisited

9. MATHEMATICAL FOUNDATIONS
This section is devoted to reviewing the mathematicalbackground of the model. Each of the 2m subsets thatcan be formed with them input neurons defines a certainpattern. Patterns can be represented as elements of alinear n-dimensional space that will be referred to asY,with addition and multiplication defined in the usualway. In this linear space of patternsY, a factork multi-plied by a certain patterny indicates the extent to whichthis pattern collaborates in the identification of anotherpatternb (b¼ kyþ :::). In Y, a negative pattern is thatwhich, when added to its opposite, annihilates it (excita-tion and inhibition are an example in neural systems).Taking this into account,Y accomplishes all the proper-ties that define a linear or euclidean space.
In Y an inner product(~A, ~B) can be defined as:
n(~A ∩ ~B)
which represents how many times the coincidence oftwo patterns takes place. This inner product shares theproperties of every inner product:
1. Commutative:n(~A ∩ ~B) ¼ n(~B ∩ ~A);2. Distributive:n(~A ∩ (~Bþ ~C)) ¼ n(~A ∩ ~B) þ n(~A ∩ ~C);3. Associative:n(~A ∩ k~B) ¼ k·n(~A ∩ ~B) (for k , 0 this
scalar product is a measure of opposition rather thancoincidence);
4. Positivity:n(~A ∩ ~A) $ 0.
This scalar product makes it possible to define ortho-gonality between two patterns as:
n(~A ∩ ~B) ¼ 0
Y becomes aeuclidean spacebecause it possesses theabove inner product. In a generic euclidean space, avector B is written in terms of its projections over anorthogonal basis as follows:
~B¼∑n
i ¼ 1
(~B, ~yi)(~yi , ~yi)
~yi
Substituting the new scalar productn(~A ∩ ~B) in the aboveexpression yields
~B¼∑n
i ¼ 1
n ~B ∩ ~yi
� �
n ~yi ∩ ~yi
ÿ �
2
4
3
5·~yi
As all axesyi are orthogonal they can be consideredas mutally exclusive events from a statistical pointof view becausen(~yi ∩ ~yj) ¼ 0 ⇒ P(~yi ∩ ~yj) ¼ 0, ;i Þ j.
It can also be considered that the sum of the prob-abilities of all the axes inY is equal to 1, thereby defininga partition of the sample space.
Taking this into account, the above expression ofa pattern B in terms of its components can berewritten as shown below, taking into account that thepart within brackets is equivalent to a conditional
probability
~B¼∑n
i ¼ 1P(~B=~yi)h i
~y
Let us define a diagonal vector~i ¼ (~y1, ~y2, ::: ~yn).Dividing ~i by the sum norm, ~i
2¼ n(~i ∩ ~i) ¼ n(~y1) þ
n(~y2) þ , ::: n(~yn), a normalised vector~I is obtained:
~I ¼~i
~i2
The probability of event~B in terms of its componentscan be defined as the scalar productP(~B) ¼ n(~B ∩ ~I ):
P(~B) ¼ n(~B ∩ ~I ) ¼∑n
i ¼ 1P(~B=~yi)·
n(~yi ∩ ~yi)i2
� �
¼∑n
i ¼ 1P(~B=~yi)·
n(~yi)n(~y1) þ n(~y2) þ , ::: n(~yn)
� �
¼∑n
i ¼ 1P(~B=~yi)P(~yi)h i
making it possible to apply the probability as if it were alinear operator over pattern~B and its~yi components:
P(~B) ¼ P∑n
i ¼ 1P ~B=~yi
� �·(~yi)
!
¼∑n
i ¼ 1P ~B=~yi
� �·P(~yi)
The above expression helps to clarify the meaning ofthe components in the euclidean spaceY. Every factorcan be understood as a conditional probability: it showsthe probability of ‘~B’ having the additional informationof ‘~yi ’. This is consistent with our previous interpretationof a factor that is multiplied by a pattern as a measureof the extent to which a certain pattern,~yi , collaboratesor prejudices (if the factor were negative) in the identi-fication of another one (in this case~B).
It was suggested that the thalamic model orthogona-lizes the afferent information. A basis of orthogonal pat-terns~yi can be obtained from other non-orthogonal onesby substituting in the Gram–Schmidt equation thepreviously defined inner product for the generic innerproducts:
~y1 ¼ ~b1
~yr þ 1 ¼ ~br þ 1 ¹∑r
i ¼ 1
n(~br þ 1 ∩ ~yi)n(~yi ∩ ~yi)
~yi
The probability of each axis is obtained by applyingthe above new definition of probability over eachmember of the Gram–Schmidt equations:
P(~y1) ¼ P(~b1)
P(~yr þ 1) ¼ P(~br þ 1) ¹∑r
i ¼ 1P(~br þ 1=~yi)P(~yi)
It is also possible to calculate the intersection of two
1286 J. Ropero Pelaez

patterns~b1 and ~b2 by substituting for each of them thesum of their projections over the previously obtainedorthogonal axes:
~b1 ∩ ~b2 ¼∑n
i ¼ 1P(~b1=~yi)·~yi
h i∩
∑n
j ¼ 1P(~b2=~yj)h i
·~yj
When~b1 and~b2 are projected over a generic axis~yi theresulting projections~b1i and~b2i can be expressed as:
~b1i ¼ P(~b1=~yi)·~yi
~b2i ¼ P(~b2=~yi)·~yi
Thus the above scalar product can be written as:
~b1 ∩ ~b2 ¼∑n
i ¼ 1
~b1i ∩∑n
j ¼ 1
~b2 j
but as there is no intersection,;i Þ j
~b1 ∩ ~b2 ¼∑n
i ¼ 1(~b1i ∩ ~b2i)
The probability of the intersection of~b1 and~b2 is calcu-lated through the scalar product of this intersection andvector~I :
P(~b1 ∩ ~b2) ¼ n (~b1 ∩ ~b2) ∩ ~Ih i
¼ n∑n
i ¼ 1(~b1i ∩ ~b2i) ∩ ~I
" #
¼ ~n (~b11 ∩ ~b21) ∩ ~y1
i2
� �þ n (~b12 ∩ ~b22) ∩ ~y2
i2
� �
þ ::: þ n (~b1n ∩ ~b2n) ∩ ~yn
i2
� �
¼ n P(~b1=~y1)~y1 ∩ P(~b2=~y1)� �
∩ ~y1
n(~i)
" #
þ:::þ n P(~b1=~yn)~yn ∩ P(~b2=~yn)~yn
� �∩ ~yn
n(~i)
" #
¼∑n
i ¼ 1P(~b1i ∩ ~b2i)
This expression yields either
P(~b1 ∩ ~b2) ¼∑n
i ¼ 1P(~b1i ∩ ~b2i)
or
P(~b1 ∩ ~b2) ¼∑n
i ¼ 1P (~b1 ∩ ~b2)=~yi
� �·P(~yi)
¼∑n
i ¼ 1P(~b1=~yi)·P(~b2=yi)·P(~yi)
so that
P (~b1 ∩ ~b2)=~yi
� �¼ P(~b1=~yi)·P(~b2=~yi)
A useful expression of the conditional probability of acertain pattern~b2 due to another one~b1 is also derivedfrom those above:
P(~b2=~b1) ¼
P(~b1 ∩ ~b2)P(~b1)
¼
∑n
i ¼ 1P(~b1i ∩ ~b2i)
P(~b1)
REFERENCES
Artola, A., Brocher, S., & Singer, W. (1990). Different voltage-dependent threshold for inducing long-term depression andlong-term potentiation in slices of rat visual cortex.Nature, 347,69–72.
Churchland, P. S. & Sejnowsky, T. J. (1992).The computational brain.Cambridge, MA: MIT Press.
Contreras, D., Destexhe, A., Sejnowski, T.J., & Steriade, M. (1995).Synchronization of thalamic spindle oscillation is enhanced bycortical inputs.Society for Neuroscience Abstracts, 21, 1187.
DeFelipe, J., & Farin˜as, I. (1992). The pyramidal neuron of the cerebralcortex. Morphological and chemical characteristics of the synapticinputs.Progress in Neurobiology, 39, 563–607.
Desmond, N.L., & Levy, W.B. (1983). Synaptic correlates of associa-tive potentiation/depression: an ultrastructural study in thehippocampus.Brain Research, 265,21–30.
DeValois, L. R. & DeValois, K. K. (1988).Spatial vision. OxfordUniversity Press.
Huntley, G.W., Vickers, J.C., & Morrison, J.H. (1994). Cellularand synaptic localizaton of NMDA and non-NMDA receptor sub-units in neocortex: organizational features related to corticalcircuitry, function and disease.Trends in Neuroscience, 17(12),536–542.
Konig, P., Engel, A.K., & Singer, W. (1996). Integrator or coincidencedetector? The role of the cortical neuron revised.Trends inNeuroscience, 19(4), 130–137.
Lester, R.A.J., Clements, J.D., Westbrook, G.L., & Jahr, C.E. (1990).Channel kinetics determine the time course of NMDA receptor-mediated synaptic currents.Nature, 346,565–566.
Llinas, R.R. (1988). The intrinsic electrophysiological properties ofmammalian neurons: insights into central nervous systemfunction.Science, 242,1654–1664.
Llinas, R., & Jahnsen, H. (1982). Electrophysiology of mammalianthalamic neurons in vitro.Nature, 297,406–408.
Llinas, R.R., Grace, A.A., & Yarom, Y. (1991). In vitro neurons inmammalian cortical layer 4 exhibit intrinsic oscillatory activity inthe 10- to 50-Hz frequency range.Proceedings of the NationalAcademy of Sciences, 88, 897–901.
Llinas, R., Ribary, U., Joliot, M., & Wang, X. J. (1994). Content andcontext in temporal thalamocortical binding. In G. Buzsaki et al.(Eds.), Temporal coding in the brain(pp. 151–172). Berlin:Springer-Verlag.
McClelland, J. L. & Rumelhart, D. E. (1988).Explorations in paralleldistributed processing. Cambridge, MA: MIT Press.
McClelland, J. L., Rumelhart, D. E. and PDP Research Group (1986).Parallel distributed processing: Exploration in the microstructureof cognition. Cambridge, MA: MIT Press.
McCormick, D. A., & Prince, D. A. (1987a). Actions of acetylcholine inthe guinea-pig and cat medial and lateral geniculate nuclei, in vitro.Journal of Physiology (London), 392, 147–165.
McCormick D. A., & Prince, D. A. (1987b). Acetylcholine causes rapidnicotinic excitation in the medial habenular nucleus of guinea-pig,in vitro. Journal of Neurosciences, 7, 742–752.
Mulle, C., Madariaga, A.M., & Deschenes, M. (1986). Morphology andelectrophysiological properties of reticularis thalami neurons in cat:
1287Plato’s Theory of Ideas Revisited

in vivo study of a thalamic pacemaker.Journal of Neurosciences,6(8), 2134–2145.
Pinault, D., & Deschenes, M. (1992). Voltage dependent 40-Hz oscilla-tions in rat reticular thalamic neurons in vitro.Neuroscience, 51,245–258.
Ropero Pelaez, J. (1996a). A formal representation of thalamus andcortex computation. In R. Moreno-Diaz & J. Mira-Mira (Eds.),Brain processes, theories, and models: an international conferencein honor of W. S. McCulloch 25 years after his death(pp. 154–163).Cambridge, MA: MIT Press.
Ropero Pelaez, J. (1996b). A model of information processing andlearning through thalamus and cortex neural connectivity andtheir neuron specific properties. In Institute of Electrical Engineersof Japan and Muroran Institute of Technology (Eds.),Proceedingsof International Workshop on Soft Computing in Industry ’96(pp. 139–147). Japan: Muroran.
Sillito, A., Cudeiro, J., & Murphy, P. (1993). Orientation sensitiveelements in the corticofugal system from visual cortex to lateralgeniculate nucleus in the cat (with an appendix on geniculo-cortical
monosynaptic connections).Experimental Brain Research, 93,6–16.
Steriade, M., & Llinas, R. R. (1988). The functional state of thethalamus and the associated neuronal interplay.PhysiologyReview, 68(3), 649–739.
Steriade, M., Curro´ Dossy, R., & Contreras, F. (1993). Electrophysio-logical properties of intralaminar thalamocortical cells dischargingrhythmic (40 Hz) spike-bursts at 1000 Hz during waking and rapideye movement sleep.Neuroscience, 59, 21–31.
Stuart, G. J., & Sakmann, B. (1994). Active propagation of somaticaction potential into neocortical pyramidal cells dendrite.Nature,367,69–72.
Thomson, A. M., & Deuchars, J. (1994). Temporal and spatial proper-ties of local circuits in neocortex.Trends in Neuroscience, 17(3),119–126.
Yuste, R., & Denk, W. (1995). Dendritic spines as basic functional unitsof neuronal integration.Nature, 375,682–684.
Yuste, R., & Tank, D. W. (1996). Dendritic integration in mammalianneurons, a century after Cajal.Neuron, 16, 701–716
1288 J. Ropero Pelaez

1997 SPECIAL ISSUE
Metastable Associative Network Models of Dream Sleep
M. Nakao,1 I. Honda,1 M. Musila2 and M. Yamamoto1
1Tohoku University and2Third Medical School of Charles University
(Received10 July 1996;accepted13 May 1997)
Abstract—Up to the present day, simulations using a neural network model have been carried out under the globalinhibition and the uncorrelated noise in order to simulate the dynamics transition of neuronal activities from the flat(slow wave sleep) to the 1/f (dream sleep) power spectral density profile during the sleep cycle in cats. In particular, themetastability of the network attractor is shown to be essential in generating the 1/f fluctuations. Here, the dynamics ofneuronal and network activities are analyzed under the correlated noises mimicking a cholinergic drive. Regardlessof the network structure, symmetry and asymmetry, the behavior of network activity and the escape time distributionsshow that the long-lasting autocorrelation of the noise prolongs its prescence in the metastable states. These results andthe newly estimated network attractor show that the interplay between the metastability of the network attractor and thenoise statistics determines the dynamics of network activity. Our results may be able to provide the novel frameworkto investigate the function of dreaming in the higher-order brain function.q 1997 Elsevier Science Ltd.
Keywords—Correlated noise, Dream sleep, Dynamics transition, Escape time, 1/f Fluctuations, Metastability, Networkattractor, Slow wave sleep.
1. INTRODUCTION
Sleep state is one of the substantial aspects of conscious-ness. Concerning the function of sleep in memory andlearning, much physiological and psychological datahave been accumulated (e.g. Smith, 1995; Antrobus,1997). Many ideas have been proposed to elucidate themechanisms underlying them (e.g. Crick and Mitchison,1983). However, none of these ideas has been estab-lished. It is essential to construct a model which canprovide an insight into the mechanisms of sleep andenables its computational interpretation. We have madeefforts following this line from the physiological andmodel-based points of view.
In a cat’s central nervous system, we have found thefollowing phenomena concerning dynamics of singleneuronal activities during sleep cycle. (i) During rapid
eye movement sleep (REM or dream sleep), neuronalactivities showed slow fluctuations, and their powerspectral densities (PSD) were approximately inverselyproportional to frequency in the frequency range of0.01–1.0 Hz (simply abbreviated as ‘‘1/f’’). (ii) Duringthe steady state of slow wave sleep (SWS), neuronsdemonstrated the almost flat spectral profiles in thesame frequency range. These phenomena have beenfound in various regions of the cat’s brain, such as themesencephalic reticular formation (Yamamoto et al.,1986), the hippocampus, the thalamus, and the cortex(Mushiake et al., 1988; Kodama et al., 1989; Yamamotoet al., 1995).
Based on neurophysiological knowledge, we success-fully simulated the dynamics transition using a symmetryneural network model including a globally appliedinhibitory input and a random noise. That is, the neuronaldynamics during SWS and REM was reproduced by thenetwork model under the strong and the weak inhibitoryinputs, respectively. We suggested the monotonousstructure of the network attractor, where the ‘‘0’’ stateis highly attractive under the strong inhibition, andthe metastability of the attractor is dominant under theweak. Thus, the structural change in the network attractorassociated with an increase in the global inhibitoryinput could underlie the neuronal dynamics transition.We also showed that the variance of the noise could
Neural Networks, Vol. 10, No. 7, pp. 1289–1302, 1997q 1997 Elsevier Science Ltd. All rights reserved
Printed in Great Britain0893–6080/97 $17.00+.00
PII: S0893-6080(97)00061-0
Pergamon
1289
Acknowledgements: This work was partly supported by a Grant-in-Aid for Scientific Research from the Ministry of Education, Science andCulture of Japan, no. 07459003. M. Musila wished to thank the JapanSociety for Promotion of Science, which sponsored his postdoctoralresearch fellowship in Japan. He passed away last year. The authorswould like to thank Professor Risaburo Sato and Hiroaki Sato for theirsimulations and useful comments.
Requests for reprints should be sent to M. Nakao, Graduate Schoolof Information Sciences, Tohoku University, Sendai 980-77, Japan;Tel and Fax: +81-22-217-7178; E-mail: [email protected].

differentiate the network state behavior in the metastableattractor. We assumed the serotonergic system as thephysiological substrate providing the global inhibi-tion from our pharmacological experimental results(Mushiake et al., 1988; Kodama et al., 1989). Moregenerally, from the results of recent studies aminergicsystems such as the noradrenergic should be includedin the substrate (McCarley et al., 1995). On the otherhand, the noise might consist of a membrane noise of aneuron, external inputs from other neural networks, etc.Our experimental results also suggest the necessityof cholinergic drive for generating 1/f fluctuations(Mushiake et al., 1988). In addition, this is shown to bepossibly mediated by the muscarinic pathway ratherthan the nicotinic (Yamamoto et al., 1993). Furthermore,acetylcholine is released in high concentration duringREM (Kodama et al., 1990). Therefore, the cholinergicsystem which originates from the brainstem and projectsdirectly/indirectly its fibers to the whole brain area couldbe a possible candidate of a noise source. Actually, ourprevious results showed how the dynamics of the net-work state evolution depends on the magnitude of theinhibition and the variance of a Gaussian noise (Nakaoet al., 1992). However, it is not yet been known whetheror not those results could be applied for dynamics undermore generalized conditions. Furthermore, a possiblerationale and functional interpretations of the 1/f fluctua-tion during dream sleep remain unsolved.
In this paper, the dynamics of neuronal activities andnetwork state evolution are investigated under a corre-lated noise which is a first-order autoregressive process.In addition, escape time distributions in metastablestates are derived for varied correlations of the noise inorder to understand the interaction between the geo-metrical structure of network attractor and the correlatednoise. Through these studies, the temporal correlation ofnoise is shown to distinctly change the network dynamicscooperating with the metastable structure of networkattractor. So far, we have been regarding the globalinhibition and the noise variance as control variables toinduce the neuronal dynamics transition. However, theymay not be the only possibilities. Here, by showing thatdiluting connections between neurons in the networkcould play a similar role to them, we propose a generalpossible mechanism for generating the 1/f fluctuationsduring dream sleep and the white PSD during SWS.We will discuss the neurophysiological mechanismsunderlying the dynamics transition as well as the rela-tionship between the dynamics of neuronal activities anddreaming.
2. NEURAL NETWORK MODEL
Here, the model structure is reviewed briefly (Nakaoet al., 1990). The neural network model we used consistsof fully interconnected neuron-like elements (abbre-viated as ‘‘neuron’’). For theith neuron, the state
evolution rule is defined as follows:
ui(t þ 1) ¼∑Nj ¼ 1
wij xj(t) ¹ hþ yi(t þ 1) (1)
xi(t þ 1) ¼ g(ui(t þ 1)) (2)
g(x) ¼1 x $ 0
0 x , 0
(
(3)
yi(t þ 1) ¼ayi(t) þ «i(t), i ¼ 1,2, …, N (4)
whereN denotes the number of neurons contained in thenetwork, andt is a discrete time.« i(t) denotes randomnoises which are assumed as mutually independent, zero-mean white gaussian noises with an identical variancej2(1 ¹ a2), in which a variance of autoregressive processis kept constant regardless ofa. h( . 0) is the inhibitoryinput which is fixed here independent of neurons for thesake of simplicity, anda is an autoregressive parameter.In this case, the autocorrelation functionr y(k) is given by
ry(k) ¼ j2a k , k¼ …, ¹ 2, ¹ 1, 0, 1, 2, … (5)
As described in Section 1, an acetylcholine-containingneuron, which is called a cholinergic neuron, could beone of possible sources for this noise. Although theirstate-dependent activities are heterogeneous, mostcholinergic neurons raise their activities during REMand reduce them during SWS (Kodama et al., 1990;Koyama and Kayama, 1995). Furthermore, a muscarinicreceptor in the cat brain is known to respond as fast as onthe order of several seconds (McCormick and Price,1987), and facilitatory effects are exerted on mostpyramidal neurons in the hippocampus (Stewart andFox, 1990) and cortex (Sato et al., 1987; McKennaet al., 1988), and a thalamic relay neuron (McCormickand Price, 1987). Considering the responsiveness of amuscarinic receptor and temporal variabilities in thedischarge pattern of cholinergic neurons during REM(Sei et al., 1993; Koyama and Kayama, 1995), thecholinergic input is supposed to be a band-limitednoise with a d.c. component in the frequency range0.01–1.0 Hz, which we are interested in. Taking accountof supposition, the correlated noise is assumed tocome from various sources, such as the cholinergic/noncholinergic input from external environment, andan intrinsic membrane noise.
Synaptic weightswij are defined following the case ofan associative memory:
wij ¼
1N
∑Mm¼ 1
(2x(m)i ¹ 1)(2x(m)
j ¹ 1) i Þ j
0 i ¼ j
8
><
>:
(6)
1290 M. Nakao et al.

where x(m)i indicates theith neuron’s state of themth
memorized pattern, andM is the number of memorizedpatterns. This possibly enables the parametric control ofthe fundamental structure of the network attractor. In thiscase, the symmetry condition
wij ¼ wji (7)
is satisfied.In addition, the asymmetry network is subject to simu-
lations, which can be regarded as more physiologicalthan the symmetry. In the symmetry network, eachneuron is allowed to exert its inhibitory and excitatorysynaptic effects on target neurons simultaneously. Onthe other hand, in the asymmetry neural network, a per-mitted sign of synapse depends on the attribute of aneuron, i.e. excitatory or inhibitory (Dale’s rule: Gray,1987). Here, synapses of the symmetry network whichdo not obey Dale’s rule are cut in order to obtain theasymmetry neural network (Shinomoto, 1987). The attri-bute of a neuron is assigned at random on each neuronwith proportionsr (excitatory) and 1¹ r (inhibitory).With synchronous state evolution, the constructed asym-metry network was shown to preserve the same retrievingproperty of the embedded memory patterns as the origi-nal symmetry network except for the limit cycle, whichsome initial states induce (Shinomoto, 1987). Our simu-lations with asynchronous state evolution find no limitcycles and no chaotic behavior.
For both types of networks, state evolution innetworks is performed in the asynchronous (cyclic)manner (Hopfield, 1982). The memorized patterns andthe initial states are given as equiprobable binary randomsequences. Unless otherwise stated, simulations arecarried out for 11 000 Monte Carlo steps (MCS), andthe initial 1000 MCS is not analyzed to exclude thestate sequence dependent on the initial pattern. Since,in our case, a PSD of a state sequence is almost invariantagainst the temporal translation of the sequence, thestarting time of the analysis scarcely affects the resultedPSD.
The data length, 10 000 MCS, is selected to estimatePSD in the frequency band width of three decades withsufficient statistical reliability. The PSDs of actualneuronal activities referred here were given in the similarfrequency band width (Yamamoto et al., 1986). Further-more, the data length of the neuronal spike train analyzedwas at most several hundred seconds. Comparing thisactual data length with 10 000 MCS, 1MCS could corres-pond to several tens of milliseconds. This could beregarded as a time unit during which a neuron keepsits state active (1) or inactive (0). The neuronal statemay be determined to be responsible for the numberof spikes during this time unit. This time resolution ispresumably sufficient, considering that the firing rates ofactual neurons under study were at most 30–40 spikes s\MN1,and the concerned frequency range is lower than 1 Hz(Yamamoto et al., 1986).
3. SPECTRAL ANALYSIS OF NEURONALACTIVITIES IN NEURAL NETWORK MODEL
Typical PSD profiles of single neuronal activities in thenetwork model are shown in Figure 1 for varied inhibi-tions anda values. Unless otherwise stated, the numberof neuronsN ¼ 100 and the number of memorizedpatternsM ¼ 30. In this figure, activity of a singleneuron is picked up from 100 neurons included in thenetwork. The raster plot ofxi(t) is shown together withthe corresponding PSD, where a dot indicatesxi(t) ¼ 1.As one can see for the case ofa ¼ 0 (i.e. white noise),PSD changes its profile from the 1/f to the flat as theinhibitory input increases. Here, the parameter valueshandj are selected regardless of the connection type, sothat most of neurons in the network show the 1/f PSDprofiles under the weak inhibition and the flat PSD pro-files under the strong inhibition. The time seriesxi(t)responsible for the 1/f PSD shows larger and slowervariations than that for the flat PSD. Naturally, the activ-ity is reduced as the inhibitory input increases. Asdescribed previously (Nakao et al., 1990), the strongand the weak inhibitory inputs are responsible for SWSand REM in our framework, respectively. Qualitatively,the PSD profiles and the temporal characteristics ofactivities are well reproduced in our simulations.Through simulations witha ¼ 0 presented here, an inhi-bitory input is manipulated so that an activity underthe strong inhibition is reduced to about 10% of thatunder the weak inhibition. This proportion is selectedas the limit of inhibition, because under the strongerinhibition the number of excitatory states could not besufficient to statistically validate the estimated PSD.Regardless of inhibition level, finely fragmented activ-ities tend to be suppressed asa increases, which is moreobvious in the strong inhibition case. Slopes of PSDscommonly become steeper, and are associated with anincrease ina. Since an increase ina results in longerlasting autocorrelation, the above results can be attribu-ted to a change in the correlation structure of the noise.The neuronal activities appear to more closely follow thedynamics of the noise asa increases.
The simulation results for the asymmetry network(M ¼ 30,r ¼ 0.5) are presented in Figure 2. In contrastwith the symmetry, clusters of activities still survive evenunder the strong inhibition. However, the above resultssummarized for the symmetry network qualitativelyapply to the asymmetry network.
4. DYNAMICS OF NEURAL NETWORK INSTATE SPACE
Up to now, we have been studying the dynamics ofneural network models which receive the global inhibi-tion and the uncorrelated random noise (Nakao et al.,1990), where the structural change of the network attrac-tor associated with the inhibition has been investigated
1291Metastable Network Models of Dream Sleep

through the state space analysis. We suggested that thisstructural change could underlie the dynamics transitionof neuronal activities during sleep cycle. Therefore, themetastable properties of the attractor could be a key issuein understanding the physiological mechanism whichcontrols the dynamics of neuronal activities during thesleep cycle. Here, we analyze how the correlation ofthe random noise modifies the network dynamics in thestate space.
For the same symmetry network in Figure 1, activitiesof all neurons (network activity) are briefly presented inFigure 3(A) under the weak inhibition, and (B) underthe strong inhibition. In each figure, the autoregressive
parameter a differentiates the pattern of networkactivities.
As shown in Figure 3(A), under the weak inhibition,the network activity explicitly indicates that the regularand irregular patterns appear alternatively with varieddurations. In the regular states, several different stripepatterns can clearly be seen. In contrast, only the irregu-lar state becomes dominant under the strong inhibition.It can be shown that these stripe patterns correspond tothe vicinities of equilibrium states under this condition,while the irregular pattern corresponds to the vicinity ofthe ‘‘0’’ state where all neurons are silent. Naturally,these equilibrium states except for the ‘‘0’’ state are
1292 M. Nakao et al.
FIGURE 1. Simulation results on the dynamics transition of neuronal activities in the symmetry neural network for varied a under weakand strong inhibitions, h, where M ¼ 30 and j ¼ 0.26. Raster plots of single activities and the corresponding PSDs for the picked-upneuron are shown together. The upper panels (A) show the results for h ¼ 0.40 and the lower (B) for h ¼ 0.50. In PSD, the frequency axis isnormalized by f 0 which denotes an arbitrary standard frequency.

different from those memorized by eqn (6) due to theglobal inhibition. The closest reference equilibriumstate to the current network state is determined everyMCS in terms of a direction cosine (DC). DC here repre-sents the ‘‘agreement’’ between a current network statex(t) and a certain reference statex*, which is defined by
DC¼(2xp ¹ 1)9(2x(t) ¹ 1)l2xp ¹ 1ll2x(t) ¹ 1l
,
x9(t) ¼ [x1(t), x2(t), …, xN(t)], x9p ¼ [xp1, xp
2, …, xpN],
19 ¼ [1, 1, …, 1], ð8Þ
wherex9 denotes a transpose of a vectorx, andx a length
of x. For the networks appearing in Figure 1, referenceequilibrium states including the ‘‘0’’ state are reachedfrom 4000 statistically independent initial states underno noise, i.e.j ¼ 0. Here, the ‘‘0’’ state is denoted byx0 ¼ [0, 0,…, 0]. Sixty-three and two equilibriumstates are found for the networks in Figure 1(A) and(B), respectively; 115 and 18 for the networks inFigure 2(A) and (B), respectively. For all references,the closest reference to the current state is determinedstep by step by comparing the magnitude of the corres-ponding DCs.
Under the weak inhibition, the network state is driftingamong the vicinities of the equilibrium states. Here, the
1293Metastable Network Models of Dream Sleep
FIGURE 2. Simulation results on the dynamics transition of neuronal activities in the asymmetry neural network for varied a under weakand strong inhibitions, h, where r ¼ 0.5, M ¼ 30 and j ¼ 0.21. Raster plots of single activities and the corresponding PSDs for the picked-up neuron are shown together. The upper panels show the results for h ¼ 0.40 and the lower for h ¼ 0.57. In PSD, the frequency axis isnormalized by f 0 which denotes an arbitrary standard frequency.

equilibrium states are not absolutely stable, becauseintermittent transitions among them driven by the noiseare observed. In this sense, they are denoted here as‘‘metastable equilibrium states’’ or simply metastablestates following the terminology of statistical physics(Amit, 1989). In spite of a constant drive by the noise,the network state is trapped in metastable states for a
certain period. In the irregular states, the network ispossibly drifting around the vicinity of the ‘‘0’’ state.While in the vicinity of the ‘‘0’’ state, each neuronalstate is expected to be determined by an instantaneousvalue of the noise rather than inputs from the otherneurons. This is presumably the reason why the spatio-temporal activity pattern appears random. Here, the
1294 M. Nakao et al.
FIGURE 3. Dynamics of network state evolution for the network shown in Figure 1. This shows the brief sequences of all neuronalactivities (network activity), where the numbers on the left end denote neuron numbers, and the number on the top indicates the numberof steps from the beginning of evolution.
FIGURE 4. Dynamics of network state evolution for the network shown in Figure 2. This shows the brief sequences of all neuronalactivities, where the numbers on the left end denote neuron numbers, and the number on the top indicates the number of steps from thebeginning of evolution.

‘‘metastability’’ represents the structural properties ofthe network attractor in which such metastable equi-librium states dominantly exist. Therefore, the followingdescription is possible concerning the dynamics transi-tion of single neuronal activities. The globally appliedinhibitory input modifies the structure of the networkattractor. In the weakly inhibited case, the metastabilityof the network attractor becomes dominant so as torealize the 1/f fluctuations of single neuronal activities,and in the strongly inhibited case, the ‘‘0’’ state becomesthe global attractor which underlies low and randomactivities. In other words, we suggest that these behaviorreflect the geometrical structure of the network attractor:a ‘‘bumpy’’ structure becomes dominant but rathermonotonous under the strong inhibition.
For a ¼ 0.5, one may not be able to recognize thedifference between the behavior of the network stateand the above results in the case of the white Gaussiannoise, i.e.a ¼ 0. However, finely fragmented patternssuch as snow noises become suppressed fora ¼ 0.9 inboth the strongly and weakly inhibited cases. For theweakly inhibited case, more types of regular patternscould be recognized than the case with the smallera.Similarly, in the strongly inhibited case, the distinctregular patterns are clearly raised asa increases.
For the same asymmetry network withr ¼ 0.5 inFigure 2, the network state behavior almost resemblesthe dynamical features observed in the symmetry case(Figure 4). We suggested that several metastable states
still survive even under the strong inhibition, which isdistinct from the symmetry case (Nakao et al., 1992).This is reflected on the cluster activities of a singleneuron in Figure 2(B). As seen in Figure 3(B), thelarge a distinctly raises the regular patterns under thestronger inhibition than the symmetry network.
5. METASTABLE NETWORK ATTRACTOR ANDCORRELATED NOISE
In this section, we investigate a mechanism underlyingthe behavior of the network activity controlled by thecorrelation properties of the noise. Asa increases, wehave observed that the irregular patterns are suppressedfor the weakly inhibited case, and the regular patternsare raised even under the strong inhibition. These obser-vations suggest that the correlation properties of thenoise could control an elapsed time during whichthe network state is trapped in a metastable state.
5.1. Escape Time Distribution in a MetastableEquilibrium State
In order to confirm the above implication morequantitatively, distribution of escape time in respectivemetastable state is obtained for varieda by a Monte-Carlo simulation with 10,000 trials. Here, the escapetime is defined as the time required for the networkstate, which is initially located in a metastable
1295Metastable Network Models of Dream Sleep
FIGURE 5. Escape time distributions in metastable states (on the definition of an escape time, see the text). Part (A) shows the semi-logarithmic plots of the escape time distribution in metastable states, state 0 and state 29, for the network shown in Figure 1(A), where h ¼
0.43 and j ¼ 0.26. Part (B) shows the semi-logarithmic plots of the escape time distribution in metastable states, state 0 and state 1 for thenetwork shown in Figure 1(B), where h ¼ 0.50 and j ¼ 0.26. Here, the abscissa denotes the escape time in Monte Carlo steps, the ordinatethe value of a, and the vertical the frequency.

equilibrium state, to first cross a boundary distant fromthe corresponding equilibrium state by 0.2 in terms of theDC under the correlated noise with the parametera.Figure 5 and Figure 6 show the escape time distributionsfor the symmetry and the asymmetry networks, respec-tively. For both types of networks, an escape time isdistributed in a monophasic manner peaking in theshort range. As shown in the distributions of states 29and 1 in Figure 5 and those of states 8 and 1 in Figure 6,the escape time for a metastable state (except for the ‘‘0’’state) usually tends to be prolonged and associated withan increase ina, while for the ‘‘0’’ state a consistentrelationship between the distribution and the value ofa
could not be found; in some cases, it is rather shortened.This might be attributed to the peculiar landscape ofpotential energy around the ‘‘0’’ state described inSection 5.2. Nevertheless, since the prolongation of theescape time is commonly observed in all other meta-stable states, the above results support our implication.
5.2. Potential Walls Surrounding Metastable States
Stochastic properties of network activity is roughly char-acterized by staying probability in a metastable state aswell as transition probability from one metastable state tothe other, where a higher-order Markovian nature isassumed to be negligible. Both probabilities possiblyreflect the height of the potential wall between meta-stable states. However, since the network potential
function is a multi-dimensional function, it is difficultto estimate the height of the potential wall. Here, thefollowing computational procedure is employed to esti-mate the height of the potential wall between metastablestates. We now show how to estimate the height ofthe wall between equilibrium statessi and sj. Firstly,the maximum network potential is obtained during theprocess approachingsi to sj by flipping the different bits(neuronal states) one by one. Then, the potential maximaare collected by repeating the same procedure, changingthe flipping order 100 times. The minimum in the set ofthe maxima is selected as an potential wall heightbetweensi andsj. In addition, the transition probabilityfrom si to sj is estimated by 10 runs of 10,000 MCSnetwork state evolutions, where the transition probabilityfor i ¼ j indicates the staying probability in statei. Thewall height and the transition probability are presentedfor the symmetry network in Figure 7. Since a networkpotential function is not known for the asymmetrynetwork studied here, the analysis is confined to thesymmetry network. For the weakly inhibited case,the wall heights and the transition probabilities fromthe ‘‘0’’ state and state 29 to all other metastable statesare presented. Note that the wall heights in the objectivestates are ‘‘0’’. Characteristically, there are high poten-tial walls between the ‘‘0’’ state and any other meta-stable states. In contrast, there are several low wallsaround state 29, e.g. to states 4, 26 and 55. This structuralproperty of the network potential around state 29 is
1296 M. Nakao et al.
FIGURE 6. Escape-time distributions in metastable states. Part (A) shows the semi-logarithmic plots of the escape time distribution inmetastable states, state 0 and state 8, for the network shown in Figure 2(A), where h ¼ 0.40 and j ¼ 0.21. Part (B) shows the semi-logarithmic plots of the escape time distribution in metastable states, state 0 and state 1 for the network shown in Figure 2(B), where h ¼
0.57 and j ¼ 0.21. Here, the abscissa denotes the escape time in Monte Carlo steps, the ordinate the value of a, and the vertical thefrequency.

1297Metastable Network Models of Dream Sleep
FIGURE 7. Estimated height of network potential walls between metastable states and the transition probability (for the procedure onestimating the potential wall height, see the text). (A) Height of network potential walls around the ‘‘0’’ state and the transition probabilityfrom the ‘‘0’’ state. (B) Height of network potential walls around state 29 and the transition probability from the state 29. Parts (A) and (B)for the symmetry network under the weak inhibition appeared in Figure 1(A). (C) Height of network potential walls around the ‘‘0’’ stateand the transition probability from the ‘‘0’’ state. (D) Height of network potential walls around state 1 and the transition probability fromthe state 1. Parts (C) and (D) for the symmetry network under the strong inhibition appeared in Figure 1(B).

shared by the other metastable states except for the ‘‘0’’state. In this sense, the potential landscape around the‘‘0’’ state is special. Regarding the transition probability,transition from the ‘‘0’’ state to the other metastablestates is rare in comparison with staying, which couldbe well understood from the above special potentiallandscape. On the other hand, the transition probabilityfrom state 29 is high in itself and the other metastablestates with low potential walls. This agreement betweenthe height of the potential wall and the transition prob-ability demonstrates the validity of the procedure forderiving the wall height. A largera is shown to makethe transition to the other states less frequently and toincrease a staying probability. Although, under thestrongly inhibited condition, a few metastable statescould only be analyzed, the results in Figure 7(C) and(D) show similar features to the weakly inhibited case inFigure 7(A) and (B). Under this condition, the potentialwall from the ‘‘0’’ state to state 1 is much higher thanfrom state 1 to the ‘‘0’’ state, which is thought to makethe staying probability close to 1. For state 1, the stayingprobability increases and the transition decreases, asa
moves closer to 1.Escape time distribution is expected to depend on the
local landscape of network potential around an equi-librium state. A symmetry neural network is known tobe a multi-dimensional discrete gradient system (seefor example Hopfield, 1982). However, there is nogeneral theory describing the metastability of such amulti-dimensional system. On the other hand, for a one-dimensional continuous gradient system with a two-wellpotential, escape time under a small Gaussian noiseobeys an exponential distribution whose parameterdepends on the height of the potential wall betweentwo wells (see Bulsara and Ross, 1991). That is, a stayingprobability in a shallow potential well has a faster decay-ing profile than in a deep well. Although under thecorrelated noise, the theoretical results are derived onlyunder limited conditions even for one-dimensionalpotential case, from some numerical experiments theescape time distribution is expected to depend onthe local geometry of the attractor as well as the correla-tion structure of the noise (Moss and McClintock, 1989).Our result obtained for the neural network qualitativelycoincides with those of one-dimensional system.
These interpretations are based on the gradientproperty of the system. As far as the asymmetry networkused here is concerned, it remains unclear whetherthis network is a gradient system or not, because itsLyapunov function appears difficult to obtain (Kobuchi,1991). However, according to the basic property of theasymmetry network dynamics described in Section 2and its a-dependency of the escape time distributionssimilar to the symmetry one, it could be regarded as agradient system. Therefore, its behavior is expected tobe understood, at least qualitatively, within the sameframework as the symmetry.
In short, the behavior of network activity in the statespace consists of stochastic transitions among metastableequilibrium states. The stochastic features of transitionsare determined by the height of the potential wallsaround metastable states and the correlation structureof the noise. As far as the inhibition induced dynamicstransition is concerned, the global inhibition reduces theheight of the potential walls and the number of meta-stable states so that a PSD of a neuronal activity changesits profile from the 1/f to the white.
6. NEURONAL DYNAMICS TRANSITIONINDUCED BY DILUTING CONNECTIONS
In the previous sections, the global inhibition is shownto change the structure of the network attractor so as toinduce the neuronal dynamics transition. From anotherpoint of view, an increase in the global inhibition and/orin the noise variance implies that external inputs becomedominant in comparison with the inputs from the otherneurons interconnected (network input). Here, ‘‘exter-nal’’ means that neurons under consideration are notinvolved in its dynamics. According to this context,the previously proposed mechanism for the neuronaldynamics transition could be reinterpreted as follows.Under the weakly inhibited condition, interactionbetween the external inputs and the network inputs isprevailing, while under the strongly inhibited condition,the external inputs dominates the network inputs.That is, the balance between the external and thenetwork inputs is supposed to play an essential role ininducing the neuronal dynamics transition. In order torealize the same situation in a different manner fromthe global inhibition, we investigate how randomlydiluting connections between neurons affect the structureof the network attractor and the metastable behavior ofthe network.
Figure 8 shows briefly the network activities in thestate space, as the connections are diluted with variedratios, where the connection is symmetry,N ¼ 196,M ¼ 59, j ¼ 0.28 andh ¼ 0.53. The noise is a whiteGaussian, and the values ofj andh are set so that mostof the neurons exhibit the 1/f fluctuations. The dilutionis carried out at random and in a symmetrical manner.With no dilution, the metastable behavior is clearlyshown to be similar to that in Figure 3(A) andFigure 4(A). As the dilution ratio increases, irregularpatterns such as a snow noise become distinct in thenetwork activities, where most of the neurons exhibitflat PSDs in the frequency range less thanf,10¹1. Inorder to understand the structural change of the networkattractor associated with the dilution, the number of equi-librium states are derived by the same proceduredescribed in Section 4. Figure 9 shows the number ofmetastable equilibrium states as a function of the dilutionratio together with that as a function of an inhibitoryinput in the fully interconnected network. Both curves
1298 M. Nakao et al.

monotonically decline with an increase in the dilutionratio or the inhibitory input. Furthermore, a similarresult is obtained for the other trials of random dilution.Naturally, full dilution isolates each neuron, whichresults in purely random neuronal activities elicitedonly by the noise. Within the numerical range of theinhibition and the dilution ratio used here, this resultsuggests that the similar structural change is causedby both of the dilution and the inhibitory input, alsotaking account of the network activities in Figure 8.Although, of course, there are many possible ways ofreducing effective network inputs, the balance betweenthe external inputs and the network inputs is suggestedto be essential for inducing the neuronal dynamicstransition.
7. DISCUSSION
In this paper, the neuronal activities and the networkactivities in the state space have been analyzed for thesymmetry as well as the asymmetry neural networkmodels under a drive by the correlated noise. Thiscorrelation represents the supposed dynamics ofcholinergic inputs. The noise correlation was observedto suppress the finely fragmented activity so that theslope of the 1/f and the flat PSDs became steeper. Thisresult indicates augmentation of frequency power inthe lower frequency range. In order to demonstratethe relationship between the network attractor and thenoise statistics, the escape time distributions forthe metastable states have been derived for the variedcorrelations. Our main finding is that the escape timetends to be prolonged and associated with an increasein the correlation time of the noise regardless ofthe connection type, symmetry and asymmetry. Here
1299Metastable Network Models of Dream Sleep
FIGURE 8. Dynamics of network state evolution for the networkwhose connections are partly diluted, where N ¼ 196, M ¼ 59, h ¼
0.53 and j ¼ 0.28. The dilution ratios are (A) 3.6%, (B) 5.5%, (C)7.6% and (D) 9.4%. This shows the brief sequences of all neuro-nal activities (network activity), where the numbers on the leftend denote neuron numbers, and the number on the top indi-cates the number of steps from the beginning of evolution.
FIGURE 9. Number of metastable equilibrium states as functionsof the dilution ratio (A) and the global inhibition (B). They areestimated for the network shown in Figure 8.

the correlation time indicates the time during whichsignificant autocorrelation exists.
We have already suggested that the structural changeof the network attractor associated with the global inhi-bitory input could be the underlying mechanism for theneuronal dynamics transition we observed physiologi-cally (Nakao et al., 1993, 1994). In particular, the meta-stable properties of the network attractor are found toplay a main role for generating the 1/f fluctuations ofneuronal activities. We also found that the correlationproperties of the noise could change the dynamics ofthe network activities, and the staying time in the meta-stable state was prolonged due to an increase in thecorrelation time of the noise (Nakao et al., 1994). Here,those results have been confirmed in terms of the escapetime distribution in each metastable state, and the meta-stable structure of the network attractor has been roughlyvisualized by estimating the wall height of the networkpotential energy between metastable states. In addition,diluting the connections in the network has been shownto modify the structure of the network attractor so that thedynamics transition of neuronal activities took place,which was similar to that induced by the global inhibi-tion. This result generalizes the conditions for generatingthe 1/f fluctuations and the dynamics transition of neuro-nal activities. According to this result, one of the essen-tial factors for inducing the dynamics transition issuggested to be the balance between the external inputssuch as the global inhibition and the noise, and theinputs from the other interconnected neurons, i.e. net-work inputs. In other words, when the network inputsand the external inputs are of a comparable order, themetastability of the network attractor distinctly appears:the 1/f fluctuation of neuronal activities are shown. Incontrast, when the external inputs exceed the networkinput, the ‘‘0’’ state is highly attractive: the neuronalactivities exhibit the flat PSD. According to our simula-tion results, mechanisms underlying the actual neuronaldynamics transition may be anticipated as follows.During SWS, neurons receive stronger inhibitory inputsand/or less input magnitude from interconnectedneurons in comparison with REM. In contrast, duringREM, neurons are released from inhibitory inputs and/or receives comparable input magnitude from inter-connected neurons with the inhibition and the noise.
We suggest the serotonergic system as a possiblecandidate responsible for the globally working inhibitorysystem. This is based on our neuropharmacologicalresults (Mushiake et al., 1988; Kodama et al., 1989).However, the noradrenergic system, which is known tohave a similar state-dependent activity to the serotoner-gic system (Jacobs, 1985), is qualified as another sub-strate exerting a biasing effect. In addition, thecholinergic input, mediated mainly by the muscarinicpathway, possibly has a d.c. component in addition tothe temporally variable component as a source of thenoise. Furthermore, there seems to exist a negative
feedback loop between serotonergic (noradrenergic)and cholinergic neurons (McCarley et al., 1995). There-fore, to be physiologically precise, the common inhibi-tory postsynaptic input,h, represents overall influencesof the ‘‘modulatory systems’’ (Maeda, 1995)" such asthe serotonergic, noradrenergic and cholinergic systems.So, from the physiological point of view, our results aremeaningful with respect to the possible function of themodulatory systems being deeply involved in controllingand maintaining consciousness. In order to confirm ourhypothesis and to clarify the contribution of respectivesystems, we are planning to set up an experiment wherethe relationship between extracellular concentrationsof amine/acetylcholine and the dynamics of neuronalactivities will be investigated. For this purpose, wehave developed the special technique combining amicrodialysis and a unit recording (Nakamura, 1996).
Rapid eye movement sleep, where the single neuronalactivity shows the 1/f fluctuation, is well known as adream sleep. Crick and Mitchison (1983) postulatedthat dream sleep functions acts to remove certainundesirable modes of interaction in neural networks ofthe cerebral cortex by nonspecific random excitation(PGO waves), which are known to be generated in thebrainstem and delivered to the cerebral cortex duringdream sleep. Their idea was concurrent with the‘‘unlearning’’ algorithm (Hopfield et al., 1983). How-ever, its physiological reality has not yet been known.Concerning the relationship between the 1/f fluctuationof a single neuronal activity and PGO waves, we havethe preliminary physiological evidence suggesting thatthere is no correlation between them (Yamamoto et al.,1993). Nevertheless, the metastable behavior of thenetwork activity appears to be suitable for unlearning,because the depth of a potential well where a metastablestate is located is reflected on the corresponding stayingtime. Appropriate unlearning could be performed byreducing synaptic weights every time the networkremains in the vicinity of a metastable state. Based ona physiology-based concept similar to ours, Sutton et al.(1992) studied the state-dependent behavior of theassociative sequencing asymmetry network whichmemorized limit cycles (Peretto, 1992). Although theimplementation of the aminergic and cholinergic effectswas different from ours, the network was assumed tobe less excitable in the non-REM state (SWS in ourcase), and more excitable and randomly perturbed inREM. Therefore, from our previous and current results(Nakao et al., 1993; Yamamoto et al., 1995), the meta-stable property is expected to be pronounced in thesimulated REM, which could underlie the diversedynamics which they observed. These assumptions andthe observed phenomena qualitatively coincide withours. Although their network attractor is considered tohave a more complex structure, the mechanism under-lying the state-dependent behavior of their network couldbe understood within the same framework as ours.
1300 M. Nakao et al.

Dreaming might be regarded as a random recallingprocess of memorized patterns without logical context.In this respect, the metastable behavior of the artificialneural network could be analogous to dreaming. There-fore, our model-based approach could provide novelinformation for investigating the functions of dreamingand REM through the 1/f fluctuations of neuronal activ-ities. From the cognitive point of view, computationalnetwork models on dreaming have been proposed(Hinton et al., 1995; Antrobus, 1997). In addition, therehave been many physiological and psychological studieswhich suggest an important role of a dream sleep inmemory and learning processes such as Wilson andMcNaughton (1994), Karni et al. (1994) and Smith(1995). The relationship between these ideas and the1/f fluctuations merits further study.
Some researchers have reported dreamings duringnon-REM (e.g. Vogel et al., 1972). Currently, we donot have any appropriate explanations for this phenom-ena. According to our simulation results, metastablestates with shallow potential wells and peculiar periodicevents during non-REM such as spindling and slow waveactivity may be a possibility. This will be the subject of afuture paper.
REFERENCES
Amit, D. J. (1989).Modeling Brain Function. Cambridge: CambridgeUniversity Press.
Antrobus, J. (1997). Dream theory 1997: toward a computationalneurocognitive model.Sleep Research Society Bulletin, 3, 5–10.
Bulsara, A. R. & Ross, F. E. (1991). Cooperative stochastic processesin reduced neuron models.Proceedings of the InternationalConference on Noise in Physical Systems and 1/f Fluctuations,pp. 621–627.
Crick, F., & Mitchison, G. (1983). The function of dream sleep.Nature,304,111–114.
Gray, E. G. (1987). Synapse, morphology. In G. Adelman (ed.)Encyclopedia of neuroscience(pp. 1158–1162). Boston, MA:Birkhauser.
Hinton, G. E., Dayan, P., Frey, B. J., & Neal, R. M. (1995). The ‘‘wake-sleep’’ algorithm for unsupervised neural networks.Sciences, 268,1158–1161.
Hopfield, J. J. (1982). Neural networks and physical systems with emer-gent collective computational abilities.Proceedings of the NationalAcademy of Science USA, 79, 2254–2258.
Hopfield, J. J., Feinstein, D. I., & Palmer, R. G. (1983). ‘‘Unlearning’’has a stabilizing effect in collective memories.Nature, 304,158–159.
Jacobs, B. L. (1985). Overview of the activity of brain monoaminergicneurons across the sleep-wake cycle. In A. Wauquier et al. (Eds.)Sleep: neurotransmitters and neuromodulators(pp. 1–14). NewYork: Raven.
Karni, A., Tanne, D., Rubenstein, B. S., Askenasy, J. J. M., & Sagi, D.(1994). Dependence on REM sleep of overnight improvement of aperceptual skill.Science, 265,679–682.
Kobuchi, Y. (1991). State evolution functions and Lyapunov functionsfor neural networks.Neural Network, 4, 505–510.
Kodama, T., Mushiake, H., Shima, K., Nakahama, H., & Yamamoto,M. (1989). Slow fluctuations of single unit activities of hippocampaland thalamic neurons in cats. I. Relation to natural sleep and alertstates.Brain Research, 487,26–34.
Kodama, T., Takahashi, Y., & Honda, Y. (1990). Enhancement ofacetylcholine release during paradoxical sleep in the dorsal teg-mental field of the cat brain stem.Neuroscience Letters, 114,277–282.
Koyama, Y., & Kayama, Y. (1995). Properties of neurons participatingin regulation of sleep and wakefulness.Advances in NeurologicalSciences, 39(1), 29–40.
Maeda, T. (1995). Neural mechanisms of sleep.Advances in Neuro-logical Science, 39(1), 11–19.
McCarley, R. W., Greene, R. W., Rainnie, D., & Portas, C. M. (1995).Brainstem neuromodulation and REM sleep.Neurosciences, 7,341–354.
McCormick, D. A., & Price, D. A. (1987). Actions of acetylcholine inthe guinea-pig and cat medial and lateral geniculate nuclei, in vitro.Journal of Physiology, 392,147–165.
McKenna, T. M., Ashe, J.H., Hui, G. K., & Weinberger, N.W. (1988).Muscarinic agonists modulate spontaneous and evoked unit dis-charge in auditory cortex.Synapse, 2(1), 54–68.
Moss, F. & McClintock, P. V. E. (Eds.) (1989).Noise in nonlineardynamical systems, Vols 1–3. Cambridge: Cambridge UniversityPress.
Mushiake, H., Kodama, T., Shima, K., Yamamoto, M., & Nakahama,H. (1988). Fluctuations in spontaneous discharge of hippocampaltheta cells during sleep-waking states and PCPA-induced insomnia.Journal of Neurophysiology, 60, 925–939.
Nakamura, K. (1996).Development of neuropharmacological tech-nique and its application to neuronal unit recording. MasterThesis, Graduate School of Information Sciences, TohokuUniversity.
Nakao, M., Takahashi, T., Mizutani, Y., & Yamamoto, M. (1990).Simulation study on dynamics transition in neuronal activityduring sleep cycle by using asynchronous and symmetry neuralnetwork model.Biological Cybernetics, 63, 243–250.
Nakao, M., Watanabe, K., Takahashi, T., Mizutani, Y. & Yamamoto, M.(1992). Structural properties of network attractor associatedwith neuronal dynamics transition.Proceedings of the IJCNN,Baltimore, MD, Vol. 3, pp. 529–534.
Nakao, M., Watanabe, K., Mizutani, Y., & Yamamoto, M. (1993).Metastability of network attractor and dream sleep.Proceedingsof the ICANN, Amsterdam, 27–30.
Nakao, M., Honda, I., Musila, M., & Yamamoto, M. (1994). Metastablebehavior of neural network under correlated random perturbations.Proceedings of the ICONIP, Seoul, pp. 1692–1697.
Peretto, P. (1992).An introduction to the modeling of neural networks.Cambridge: Cambridge University Press.
Sato, H., Hata, Y., Masui, H., & Tsumoto, T. (1987). A functional roleof cholinergic innervation to neurons in the cat visual cortex.Journal of Neurophysiology, 58(4), 765–780.
Sei, H., Sakai, K., Yamamoto, M., & Jouvet, M. (1993). Spectralanalyses of PGO-on neurons during paradoxical sleep in freelymoving cats.Brain Research, 612,351–353.
Shinomoto, S. (1987). A cognitive and associative memory.BiologicalCybernetics, 56, 1–10.
Smith, C. (1995). Sleep states and memory processes.Behavioral BrainResearch, 69, 137–145.
Stewart, M., & Fox, S. E. (1990). Do septal neurons pace the hippo-campal theta rhythm?.Trends in Neuroscience, 13, 163–168.
Sutton, J. P., Mamelak, A. N., Hobson, J. A. (1992). Network model ofstate-dependent sequencing. In R. P. Lippmann, J. E. Moody, and S.J. Hanson (Eds.),Advances in neural information processingsystems4, Proceedings of NIPS’91 (pp. 283–290). San Meteo,CA: Morgan Kauffmann.
Vogel, G. W., Barrowclough, B., & Giesler, D. D. (1972). Limiteddiscriminability of REM and sleep onset reports and its psychiatricimplications.Archives of General Psychiatry, 26, 449–455.
Wilson, M. A., & McNaughton, B. L. (1994). Reactivation ofhippocampal ensemble memories during sleep.Science, 265,676–679.
1301Metastable Network Models of Dream Sleep

Yamamoto, M., Nakahama, H., Shima, K., Kodama, T., & Mushiake,H. (1986). Markov-dependency and spectral analyses on spikecounts in mesencephalic reticular neurons during sleep and attentivestates.Brain Research, 366,279–289.
Yamamoto, M., Arai, H., Takahashi, T., Sasaki, N., Nakao, M.,Mizutani, Y., & Kodama, T. M. (1993). Pharmacological basis of1/f fluctuations of neuronal activities during REM sleep.SleepResearch, 22, 458.
Yamamoto, M., Nakao, M., Mizutani, Y., & Kodama, T. (1995).Dynamic properties in time series of single neuronal activityduring sleep.Advances in Neurological Science, 39, 29–40
NOMENCLATURE
a parameter of an autoregressive process« i(t) random noise mutually independent, zero-
mean white Gaussian noise with an identicalvariancej2(1 ¹ a2)
j2 variance of the noise {y i(t)}
y i(t) first-order autoregressive processui(t) internal state of theith neuron at timetwij synaptic weight from theith to jth neuronsxj(t) output of j ¹ the neuron at timeth global inhibitory inputN the number of neurons contained in the
networkM the number of the memorized patternsr y(k) autocorrelation function of the autoregres-
sive processx(m)
i the ith neuron’s state of themth memorizedpattern
x(t) current state vectorx* reference equilibrium state vectorx0 the ‘‘0’’ state vectorPSD power spectral densityDC direction cosineMCS Monte Carlo step
1302 M. Nakao et al.

1997 SPECIAL ISSUE
Consciousness and Neural Cognizers:A Review of Some Recent Approaches
Chris Browne, Richard Evans, Nick Sales and Igor Aleksander
Imperial College of Science, Technology and Medicine
(Received26 July 1996;accepted28 June1997)
Abstract—This paper synthesises three diverse approaches to the study of consciousness in a description of an existingprogram of work in Artificial Neuroconsciousness. The three approaches are drawn from automata theory (Aleksander,1995, 1996), psychology (Karmiloff-Smith, 1992; Clark & Karmiloff-Smith, 1993) and philosophy (Searle, 1992).Previous work on bottom-level sensory-motor tasks from the program is described as a background to the currentwork on generating higher-level, abstract concepts which are an essential part of mental life. The entire program of workpostulates automata theory as an appropriate framework for the study of cognition. It is demonstrated how both thebottom-level sensory-motor tasks and abstract knowledge representations can be tackled by a single neural statemachine architecture. The resulting state space representations are then reconciled with both the psychological andphilosophical theories, suggesting the appropriateness of taking an automata theory approach to consciousness.q 1997Elsevier Science Ltd.
1. INTRODUCTION
This paper brings together three recent approachesin the study of consciousness. A neurally based theoryof artificial consciousness (Aleksander, 1995, 1996), aphilosophy of mind based on cognitive acts (Searle,1992) and a theory of human development based on aredescription of learned competences (Clark &Karmiloff-Smith, 1993). The purpose of this paper is topresent a neural automata theory approach to conscious-ness, rather than describe particular machine architec-tures, which are liable to undergo refinement as thework progresses. Particular details of the design of theexperimental work described can be found in the appro-priate references.
For continuity and completeness, the paper will com-mence with a brief recapitulation of the salient proposi-tions of the Artificial Consciousness Theory which arelater compared with Searle’s philosophy of mind (Searle,1992). Searle insists that the study of cognition is thestudy of consciousness, just as the study of biology isthe study of life.
[The brain’s] special feature, as far as the mind is concerned,the feature in which it differs remarkably from other bio-logical organs, is its capacity to produce and sustain all ofthe enormous variety of our conscious life. By conscious-ness I do not mean the passive subjectivity of the Cartesiantradition, but all of the forms of our conscious life-from the
famous ‘‘four f’s’’ of fighting, fleeing, feeding and fornicat-ing, to driving cars, writing books and scratching our itches.All of the processes that we think of as especially mental—whether perception, learning, inference, decision making,problem solving, the emotions, etc.— are in one way oranother crucially related to consciousness. Furthermore,all of those great features that philosophers have thoughtof as special to the mind are similarly dependent on con-sciousness: subjectivity, intentionality, rationality, free will(if there is such a thing), and mental causation. More thananything else, it is the neglect of consciousness thataccounts for so much barrenness and sterility in psychology,the philosophy of mind, and cognitive science.
The study of the mind is the study of consciousness, in muchthe same sense that biology is the study of life. Of course,biologists do not need to be constantly thinking about life,and indeed, most writings on biology need not even makeuse of the concept of life (Searle, 1992).
So, taking Searle’s argument that the study of mind isthe study of consciousness, this paper sets out to examineneural network models of mind. Having defined the posi-tion on consciousness adhered to and claimed that thestudy of mind is the study of consciousness, all thatremains is to define the stance taken on mind in thispaper. The mind is assumed to be directly responsiblefor cognition. Clark & Karmiloff-Smith (1993) make a
Neural Networks, Vol. 10, No. 7, pp. 1303–1316, 1997q 1997 Elsevier Science Ltd. All rights reserved
Printed in Great Britain0893–6080/97 $17.00+.00
PII: S0893-6080(97)00063-4
Pergamon
1303

powerful distinction between complex informationprocessors, which includes computers, and genuinecognizers.
The sea slug and the VAX mainframe are both effectiveprocessors of information. Yet it is only human beings,and perhaps some higher animals, who are credited withgenuine thoughts. Is this mere prejudice, or have we some-how latched on to a genuine joint in the natural order? If it isa genuine joint in nature, what feature of set of features markit?
The hypothesis to be considered is that there is indeed a jointin the natural order such that humans fall on one side andmany other systems (including some quite sophisticatedinformation processors) fall on the other. The joint, weargue, marks a pivotal difference in internal organisation.The representational redescription model embodies specifichypothesis about the nature of this joint (Karmiloff-Smith,1979a, 1979b, 1986, 1990, 1992). For genuine thinkers, wesubmit, are endowed with an internal organisation which isgeared to the repeated redescription of its own stored knowl-edge. This organisation is one in which information alreadystored in an organism’s special-purpose responses to theenvironment is subsequently made available, by the RRprocess, to serve a much wider variety of ends. Thus knowl-edge that is initially embedded in special purpose effectiveprocedures subsequently becomes a data structure availableto other parts of the system (Clark & Karmiloff-Smith,1993).
There has been some discussion about the boundariesbetween congizers and non-cognizers (Aleksander,1996) which suggests that animals and even machinescould redescribe their knowledge. Apart from notingthis slight objection, the general principle that redescrip-tion is an essential feature of cognition is not at stake,there is only a slight difference in opinion over the posi-tion and abruptness of the divide.
In the search for artificial consciousness, the chal-lenge, therefore, is to bestow a neural network with theability to re-represent its own internal states. Previouswork (Browne & Parfitt, 1997) suggests that such asystem might well add further weight to the numerousrefutations (Smolensky, 1987; Pollack, 1990; VanGelder, 1990; Aleksander & Morton, 1993) of Fodorand Pylyshyn’s attack on connectionist models of cogni-tion (Fodor & Pylyshyn, 1988). The previous workfurther suggests that recursive redescription of systemrepresentations might provide a possible mechanism bywhich a pseudo-symbolic system, which many agree isthe basis of cognitive function, could arise in a connec-tionist network. The general model of emergent hierarch-ical data structures presented here also has much incommon with Harnad’s proposals of the recursivegrounding of language (Harnad, 1992).
Before the redescriptive process can be discussed indetail, the type of neural architecture and represen-tational form being proposed must be described.Karmiloff-Smith’s representational redescription model
(Karmiloff-Smith, 1992) is then presented in detail,moving to a suggested neural architecture with the capa-city for spontaneous re-representation of its own internalstates. The paper concludes with a discussion of the rela-tionship between Searle’s philosophy, the FundamentalPostulate (Section 2.2) of the Artificial ConsciousnessTheory (Aleksander, 1996) and the proposed cognitiveneural architecture which emphasises the importance ofexamining the mental processes occurring in the systemas conscious in some form—the authors would stress anartificial, rather than biological form.
2. NEURAL STATE MACHINE MODELSOF COGNITION
For over 20 years, one of the authors (I.A.) has beensuggesting that the capacity for thought can be encapsu-lated in a machine with an adaptable state structure(Aleksander & Hanna, 1975; Aleksander, 1996). Anadaptable state machine can be implemented in amachine with some form of learning capacity. The con-temporary versions of such machines have been dubbedas ‘‘Neural State Machine Models (NSMMs)’’(Aleksander & Morton, 1993); the Multi-AutomataGeneral Neural Unit System (MAGNUS) (Aleksanderet al., 1993) being the most widely used. Since theNSMM is about as far as one can get from much ofthe philosophical discussion concerning consciousness,it is worth pausing a moment, before pressing on with adescription of the Artificial Consciousness Theory toplace this proposition in relation to the philosophicaldiscourse. A more detailed account can be found inAleksander (1996).
2.1. Relationship to a Genealogy
The position taken in the program of work relating to theArtificial Consciousness Theory is that the term con-sciousness was first used by John Locke when in 1690he wrote:
First, I shall enquire into the origin of those ideas, notions, orwhatever else you please to call them, which a man observesand is conscious to himself he has in his mind; and the wayin which the understanding comes to be furnished with them(Locke, 1690, Ch. i. 4. Reproduced in Locke, 1975).
This can be distinguished from three millennia of con-cerns with the physical makeup of mind and its relation-ship to body culminating with Cartesian dualism(DesCartes, 1637) about which Locke and his successorsfor the next 300 expressed considerable skepticism.Locke’s motivation for his study of knowledge is reputedto have come from his view that metaphysics was ulti-mately futile. He is reported to have turned to friendsafter an inconclusive debate on metaphysics and asked;‘‘ Shouldn’t we first determine whether we are capable ofanswering such questions?’’ (Soloman, 1993).
1304 C. Browne et al.

Philosophy from Locke in the 17th century toWittgenstein at the turn of the 20th attempted to refinethe taxonomy of different ‘‘ideas, notions, ...the under-standing’’. This in itself is not a matter for mathe-matical analysis but one of philosophical discourse.However, when suggesting that consciousness could beanalysed in an artificial neural model it is preciselythis taxonomy which has to be borne in mind, as themodel must palpably be able to represent ‘‘notions andtheir understanding’’ which are as different as, say, feel-ings, objective, knowledge, actions, beliefs, etc. It isfor this reason that a list of corollaries has been createdin the work. Some of the corollaries are described inlater sections of this paper; a complete list can befound in the Appendix.
The curious nature of contemporary analyses ofconsciousness is that they spread over a very widemethodological front. At one extreme is the naturalphilosophical descent of the lineage mentioned above,where it is the nature of consciousness and not itsphysical basis which is at stake. Many such discussions(Nagel, 1974; Lucas, 1994, for example) take the posi-tion that only the inexplicable subjective nature of con-sciousness is worth discussing. Giving it a neural basisis therefore at best uninteresting and at worst irrelevantto philosophical concerns. Others (Penrose, 1994, forexample) believe that consciousness is beyond thatwhich can be achieved through current mathematicalanalyses, particularly neuronal computations. He advo-cates a solution based on the function of microtubles inthe brain cells causing effects which could only bedescribed by a non-computable extension to quantumtheory which includes the gravitational force. Mostanalyses, however, which actually attempt to find aneural basis of consciousness operate in a domainwhere effects observed in the human brain are correlatedto conscious experience reported by or inferred in theowner of that brain. Greenfield (1995) and Crick(1994) are major contributors to this methodology.Even further away from classical philosophy is thework of computationalists such as Dennett (1991) andSloman (1994) who argue that consciousness is a productof the architectural character of the brain which, forDennett, can be expressed in computational terms suchas virtual machines and interacting agents.
The approach taken in this paper, described in detail in(Aleksander, 1996), is founded on iconic representationand its consequences, which suggests that consciousnesscould be found in neuronal models which are neitherstructured like the brain, nor are they computationalistin the sense that they represent a programmer’s idea ofwhat the elements of consciousness are. The approachis constructivist only in the sense that it looks foremergent properties of a very large structure, but relatessuch properties to the global parameters of that structure.The only construction is the fundamental postulate, therest is consequential or a matter of refinement. This is as
far removed from the philosophical lineage as one canget, as a philosopher might immediately argue that suchstructures are disqualified for not being the flesh andblood for which consciousness makes sense. But theaim of the rest of this paper is to illustrate that this con-structivist approachdoesprovide a general perspectiveon the synthesis of consciousness, while respecting thegenealogy of the subject. A synthesised consciousnessmight in itself, if Searle is correct, be at the root ofbuilding truly cognitive machines.
2.2. The Fundamental Postulate
The Fundamental Postulate (Aleksander, 1996) states thefollowing:
The personal sensations which lead to the consciousness ofan organism are due to the firing patterns of some neurons,such neurons being part of a larger number which form thestate variable of a neural state machine, the firing patternshaving been learned through a transfer of activity betweensensory input neurons and the state neurons.
The Fundamental Postulate posits, as an appro-priate framework in which to examine cognitive pro-cesses, an automata theory in which the state variableshave neural learning properties, and also that neuralstates which contribute to consciousness must be createdthrough a process which has been described asiconiclearning.
2.2.1. Iconic Learning. Iconic learning is a represen-tational scheme proposed in previous work by one ofthe authors (Aleksander & Morton, 1993) and others(Harnad, 1987b). The representational scheme has twosalient properties which are; (1) preservation of the struc-tural form of the environment through the direct transferof activity from sensory neurons to the state neuronsresponsible for consciousness and (2) the representationsare concatinatively combinatorial.
Harnad proposes iconic representations for discrimi-nation, which he defines as, ‘‘[T]he ability to judgewhether two inputs are the same or different, and, ifdifferent, how different they are’’ (Harnad, 1992).Identification, on the other hand, is concerned withcategorisation and requires a different representationalform. As opposed to discrimination, which is a relativejudgement between objects, identification is an absolutejudgement as to which category a particular perceptionbelongs to. Contrary to some theorists (Paivio, 1986, forexample), Harnad maintains that iconic representationsare insufficiently distinguishable for reliable identifica-tion (Harnad, 1982, 1987b). He proposes a system of‘‘categorical perception’’ for reliable identification(Harnad, 1987a, b) which utilise categorical represen-tations. Categorical representations are the product ofcategory specific feature detectors which filter outinformation1 in the iconic representations which are not
1305Consciousness and Neural Cognizers

directly involved in the differentiation between membersof a category and nonmenmbers (Harnad, 1990).
Harnad (1987b) proposes that the categorical represen-tations only lose information which is irrelevant to thedisambiguation between category members and non-menbers within the existing context of alternatives. Theissue here is that the features which differentiate a catfrom say a dog are different from those which distinguishbetween a cat and a tiger, which are again differentfrom the set of features which between a cat and anelephant. In other words, the category features varydepending on the particular features of the class of non-members with which the object for categorisation iscompared. In order to accommodate a large range ofconfusable alternatives the category representationsmust maintain a great deal of the structural informationabout the environment, but in a compressed form. Sincethe structural form of the environment is to largely bemaintained, then the resulting representational form isbetter dubbed asreduced iconic representationswithinthe current discourse to highlight their compatibilitywith the Fundamental Postulate (Section 2.2). Harnad(1987b) describes the proposition as follows:
Note that [the conversion from iconic to categorical repre-sentation] is still analogue, for it continues to preserve someof the physical ‘‘shape’’ of the input, albeit in a highlyreduced and abstracted form. Iconic/Categorical conversionis really a transformation from macro-icons to micro-icons,the former preserving the context independent, holistic,configural properties of the proximal stimulus, the latterselectively reduced to only those properties that areinvariant in a specific context of confusable alternatives(Harnad, 1987b).
2.3. State Space Representations of Knowledge
State spaces are the currency of automata theory. Two ofthe authors (Sales, 1996; Evans, 1996) have examinedthe properties of learned state spaces in neural statemachines as models of cognition. The work was under-taken using a simulated, virtual, environment calledkitchenworld.
2.3.1.The Kitchenworld Environment. Thekitchenworldarrangement (Figure 1) consists of MAGNUS (Multi-Automata General Neural Unit System) exploring andnaming objects in a virtual environment calledkitchen-world. The kitchenworld consists of objects such asglasses, plates, cutlery, cups, bottles and the odd apple.The digitised image, available to the MAGNUS simula-tion, was created from a real image taken from a videocamera. Since both MAGNUS and thekitchenworldare
simulated, it is important to distinguish between them.Everything within the large dotted frame (Figure 1) isthe MAGNUS program. Everything else, including thepicture, is another program—thekitchenworldprogram.
The two interact as follows. At its output, MAGNUSissues signals tokitchenworldwhich controls the positionand size of a window placed over the picture. In theexample (Figure 1) the window is shown to be over anapple. Another dotted position for the window is shownover a plate. So MAGNUS is to derive how muchto move in the two directions X and Y, and how big tomake the size (Z) of its window on thekitchenworld.The output of the MAGNUS (Figure 1) consists ofthree bars, one each for X, Y and Z. These can be thoughtof as "strengths of firing of three separate regions of theneural state machine which determine the position ofthe window.
Given the position and size of a window, thekitchen-world delivers to MAGNUS the content of the window asa thresholded binary pattern. This pattern forms one partof the neural state machine input. It can be seen as avisual input—MAGNUS current view of thekitchen-world. The second part of the system input can be seenas linguistic. It consists (Figure 1) of a binary pattern inthe form of a word. This pattern is selected by an instruc-tor and is considered to be a name for the object currentlyin view. The system is, therefore, exposed to two sensoryevents in different modalities, visual and linguistic,which occur in close time proximity; the word input isdeliberately chosen and presented to coincide with thevisual stimulus.
1306 C. Browne et al.
1 Harnad (1990) actually uses the word features, but followingcorrespondence between one of the authors (C.B.) and Harnad, theauthor believed that information is a more apt description.
FIGURE 1. MAGNUS in a kitchen world.

2.3.2.Simple Object Naming.In previous work (Sales,1996), a model of simple object naming was constructed,which verified Corollary 8 (Aleksander, 1996).
2.3.2.1. Corollary 8: Representation of meaning.
When sensory events occur simultaneously or in close timeproximity in different sensory modalities, iconic learningand generalisation of the neural state machine ensures thatone can be recalled from the other (Aleksander, 1996).
The MAGNUS visual input was centred on an objectin the kitchenworld, the framed apple (Figure 1) forexample, and simultaneously presented patterns corres-ponding to the written name for the object to the linguis-tic input. MAGNUS, a generalising neural state machine,was then iconically trained on the pair of inputs. Theneural state machine learned attractor states which con-sisted of both the visual and linguistic inputs (Figure 1).It is known that in common with any neural model, localgeneralisation in the logical neurons used in MAGNUSleads to the property of state reconstruction (Aleksander& Morton, 1991). This means that either the name inputor the object input will lead to the reconstruction ofthe state representing both (Figure 2). In other words,an input from a single modality, with the other set torandom noise, will lead to the recall of a state consistingof the iconic representations of the sensory experienceof both modalities. The experience of each modalitycan be reconstructed from the other, as proposed inCorollary 8.
The learning of simple object names discussed here isopen to the criticisms raised by philosopher’s such asWittgenstein (1953) and Quine (1960). The problem, inshort, concerns the underdeterminance of language. Theissue at stake is how a child learning a language is todetermine which part of the visual scene relates to the
object being named. The refutations of these criticismslie in the formation of group nouns for objects, which isa more complex extension of the work described in thissection and is discussed in the section relating to that(Section 3.1.1).
2.3.3. Locating Objects.Previous work (Evans, 1996)used the MAGNUS system in thekitchenworldenviron-ment to examine the capacity of an iconically trainedneural state machine to locate objects. Object locationis crucial if it is to be understood how a consciousorganism might partition its sensory world into objectsthat need naming. It has been demonstrated (Evans,1996) that it is possible to train a window to locate ona particular object such as an apple (Figure 1). Locationon an object is achieved by displacing a window, whichis perfectly positioned over an object, in both locationand size, and training the network to output a movetowards the correct position. The interesting part of thiswork is that having done this for the apple, as shown, notonly the apple becomes an attractor in the image space,but so does any other object. A close look shows thatthe system is interpretingany disturbance from the back-ground in the visual input as a pattern which causes anoutput that moves the movable input window towardsthe object. So when the input window covers a part ofan object, it will tend to be attracted towards that object.Curiously, this creates not only attractors where theobjects are, but also ‘‘repellers’’, such as R (Figure 3)in areas where there are no objects.
The experiments demonstrate that it comes naturallyfor a neural state machine with a visual input and a motoroutput to develop a general notion of finding an objectby treating it as a disturbance in a background afterminimal instruction.
1307Consciousness and Neural Cognizers
FIGURE 2. State characteristics for two named objects.FIGURE 3. Attractor objects and a repeller background in thekitchen world.

3. REPRESENTATIONAL REDESCRIPTION
The tasks presented so far have all concerned bottom-level, sensory-motor processing. Earlier, we suggestedthat cognition only arose through a process of redescrip-tion of lower-level representations into higher-level ones.That process is now described in detail.
The representational redescription (RR) model hasbeen developed by Karmiloff-Smith over a period oftime. A comprehensive description is given in Karmiloff-Smith (1992). At the heart of the RR model is an internalprocess of representational change by which the mindenriches itself.
The RR model posits a number of levels. It should bestressed that RR is not a Piagetian stage model. RR ispresented as a domain general process, but importantlyit is posited to act on domain specific representations.Three levels of the RR model are of interest to the con-nectionist modeller.
The first level of implicit, or level-I, representationsis the level achieved by connectionist models of beha-viour. At this level stored representations are activatedby external stimuli, but are not available to the systemin an explicit re-usable format (Karmiloff-Smith, 1992;Browne & Parfitt, 1997). Beyond level-I is the firstlevel of explicit representation of knowledge, level-E1.At level E1 representations are not available to con-sciousness or verbal report, but they are available withoutthe need for external stimuli and to other domains.At higher levels of redescription, levels E2/E3 [alsodenoted by Eþ (Clark & Karmiloff-Smith, 1993)] theknowledge is available to conscious access and verbalreport.
Karmiloff-Smith has undertaken a number of develop-mental studies in order to investigate changes inchildren’s representations (Karmiloff-Smith, 1992).These studies bear out the predictions of the RR model.A particularly salient study is one concerning drawings(Karmiloff-Smith, 1990).
Six drawings were produced by four- to 11-year-olds.The children were told the subjects of the drawings. Firsta house was requested, then a house which does notexist. Similar pairs of drawings for a man and an animalwere asked for. Only the drawings of men (Figure 4) areconsidered here.
The motivation behind asking the children to draw abasic object, succeeded by a version of that object thatdid not exist was as follows. The standard object couldbe produced by the simple execution of an actionschema. To produce the fictitious drawing, however,the children would be forced to break down their actionschema for drawing the regular objects in order to resize,reshape, omit or swap components. The hypothesis,which was borne out, was that young children withonly level-I representations would be unable to drawthe fictional manifestations of the objects. Their implicitonly representations constrained these children tomerely reproducing the entire schema, despite verbalreport that they were about to draw ‘‘a man who doesnot exist’’. The components of the schema were notavailable as explicit information available to the childrenfor manipulation.
Of greater interest was the difference in drawingsproduced by four- to six-year-olds and eight- to10-year-olds. The imaginary drawings from the formerwere created by altering the size and shape of compo-nents and some deletions of components. The deletedcomponents were often the last one to be drawn in theundistorted pictures. The later group’s fictitious drawingsshowed a much greater variety of manipulations fromthe original. The components of the original imageswere manipulated as independent entities, leading toinsertion of additional elements from the same concep-tual category, change of position and/or orientation ofwhole elements, or even the insertion of componentsfrom a different conceptual category.
The children’s drawing study demonstrates theunderlying thesis of RR, which is that knowledgewhich is implicit in the original action schema, in thiscase the action schemas for drawing body compo-nents, gradually becomes more explicit through aninternally generated process of representationalredescription.
Another important feature of the RR process is thatit is recursive. The redescribed representations mustthemselves be capable of undergoing further redescrip-tion. As more levels of redescription occur, the knowl-edge becomes more abstract and more accessible. Asstated above, previous work suggests that it is the recur-sive nature of RR which may lead to an insight as to how
1308 C. Browne et al.
FIGURE 4. Example drawings inspired by children’s drawings ofa man who does not exist. After (Clark & Karmiloff-Smith, 1993).Taking the drawings in rows from the top left and denoting theages of the artists as years; months, then the ages are as follows(a) 4;8 (b) 5;9 (c) 5;3 (d) 9;0 (e) 9;8 (f) 9;6 (g) 9;8.

a pseudo-symbolic system of cognitive representationis constructed from low-level processing (Browne &Parfitt, 1997).
3.1. Redescription in State Spaces: Complex States
Previous work (Aleksander, 1996) suggests that thebottom-level sensory-motor, simple states learned intasks such as those studied above (Section 2.3) [Evans,1996; Sales, 1996] are equivalent to Karmiloff-Smith’sbottom-level, implicit (level-I) representations(Karmiloff-Smith, 1992). He further suggests thathigher-level, complex states can be formed which areequivalent to the first level of explicit, abstract knowl-edge; level-E1.
3.1.1. First Level of Redescription.To illustrate thenotion of complex state representations of abstract con-cepts, a particular example which makes reference tothe kitchenworldenvironment will be used. Within thekitchenworldthere are a number of repeated instancesof various objects (Figure 1). There are two apples,four glasses, two cups and two plates. When the objectsare named like objects will receive identical names;both cups will be labelled as ‘‘cup’’, each glass as‘‘glass’’, etc. Since the object naming function of thesystem allows the visual input to be recalled fromthe linguistic label, the multiple visual input for eachlinguistic label might now appear to cause a conundrum.Which visual input is to be recalled in association withthe linguistic label? The dilemma can be resolved byadding a probabilistic element to the system (Browne& Aleksander, 1996). This probabilistic element allowsthe system to explore regions of state space where theinput does not explicitly define a particular state. Inthe case where the visual input is undefined and thelinguistic input is a recognised label, the probabilisticstate space exploration allows all the states whichmatch the linguistic input to be explored (Figure 5).During the exploration of state space the states corres-ponding to several, or even all, the visual inputs arevisited. The linguistic labels have now become classnouns, representing the commonality between a clusterof bottom-level, learned states. These clusters of statecan now be represented as single, complex states[Figure 6(a)].
There is some controversy as to whether the complexstates described are adequate representations of classnouns. The deficiencies of this approach are discussedin a later section. For the current argument, however,which is based on the linguistic construction of classnouns the described complex states are arguably suffi-cient, if recourse is made to Wittgenstein’s proposition ofthe non-essentialism of language (Wittgenstein, 1953).Wittgenstein argued that there are no hidden, or ultimate,universals in language. He suggests that a word such as‘‘dog’’ derives its meaning from a mental image, the
visual modality region of the described complex states,and that the image must be derived from only the set ofdogs which the person in question has experienced. Thesimple states which together form the complex state areall derived directly sensory experience. This is ensuredby the insistence on iconic learning. It is in this retreatfrom the realism of word meaning, which is arguablywhat prompted Wittgenstein to write ‘‘PhilosophicalInvestigations’’ (Wittgenstein, 1953), that the concernsof Quine (1960) and particularly Wittgenstein (1953),noted above (Section 2.3.2) are refuted. The argumentstated here is expanded in detail by both Harnad (1995)and Searle (1992). Searle argues in particular thatQuine’s theory (Quine, 1960) is only appropriate for azombie robot, which conscious human beings areclearly not.
1309Consciousness and Neural Cognizers
FIGURE 5. Concepts in state space. The first level ofredescription.
FIGURE 6. State grouping to form concepts (a) and higher-levelconcepts (b).

3.1.2. Second Level of Redescription.Karmiloff-Smith(1992) demands the redescribed states must themselvesbe available for redescription. Continuing with the pre-vious example wherekitchenworldstates are redescribedthrough the addition of linguistic tags, the second level ofredescription can be driven by providing new, equallyvalid but broader domain, linguistic tags for knownobjects. Moving slightly away from the actualkitchen-world, assume that a system is exposed to an environ-ment consisting of a number of fruits; strawberries,pears and grapes for instance. Assume also that theneural state machine has learned the names of the indi-vidual fruits, so it has complex states representingthe class nouns strawberry, pear and grapes. Now saythe system’s visual input is centred on one of the straw-berries and the word fruit is input to the linguisticmodality. The visual input of the strawberry will causea region of the complex state representing the nounstrawberry to be reconstructed and learning will combinethe new linguistic tag, fruit into that state. A new com-plex state with two linguistic tags, strawberry andfruit, for the associated visual stimuli will be created. Ifthe word ‘‘fruit’’ were then associated, in an identicalmanner, with the complex states representing pearand grapes, then a set of states which are unified by acommon linguistic tag [Figure 6(b)] is created, whichis identical in form to the set of simple states whichcombined to create the initial complex states[Figure 6(a)] except that the lower-level states are them-selves complex states, rather than the bottom-level,sensory-motor states of the previous example. In otherwords, the more general group noun ‘‘fruit’’ has allowedthe first level complex states representing the groupnouns for individual fruits to be recursively redescribedat a higher level. This recursive redescription can beseen to be constructing a general, hierarchical informa-tion structure.
3.2. Inadequacy of Language Based Redescription
The proposed method of representational redescriptiongives an excellent demonstration of the applicability ofcomplex states to the task. The redescription process, ascurrently described, however, requires external linguisticinput. Karmiloff-Smith is quite prepared to accept thatexplicit knowledge can arise through verbal instruction,but she insists that this is not true representationalredescription. That, she insists is driven by an internal,spontaneous process.2
The possibility of deriving internally driven grouprepresentations, their form and the implications on thesystem are the topics of the next section.
4. INTERNALLY DRIVEN REDESCRIPTION IN ASINGLE NEURAL ARCHITECTURE
The task of building a single neural architecture with aninternally driven redescriptive mechanism can now besplit into two questions. (1) Given the explanatory powerof the suggested complex state approach to redescription,what form of internal representation might replace theexternally provided class nouns? (2) How could a singleneural architecture derive the appropriate representations?
4.1. Representational Replacement for Class Nouns
Since representational redescription is posited as a recur-sive process, the appropriate representational form forthe abstracted concepts must be highly related to thatproposed for the bottom-level; i.e. iconic learning. FodorandPylyshynargue,correctly, that the token3 representingan higher level concept should consist of the systematiccombination of the items from which it is comprised(Fodor & Pylyshyn, 1988).What is at stake is whethersystematic combination of tokens can only be achievedin computational data structures, or whether a neural net-work is capable of such functionality. Pollack has clearlydemonstrated that a connectionist system is capable ofsystematic combination (Pollack, 1990). The suggestionpresented here is an extension and refinement ofPollack’s recursive auto-associative memory (RAAM).
The concept of glass has already been established to begrounded in the sensory experience of all the glasses aperson has seen. It would seem appropriate to constructthe representation of the concept from this sensory experi-ence. Doing so would satisfy both the criteria for systema-tic combination of the tokens from which the conceptis constructed and the demand for iconic representation.
The most obvious technique of combining the sensoryiconic representations of the four glasses (Figure 5)which have been experienced would be to superimposethem. This technique has two serious inadequacies; (1)the resulting representation is liable to be amorphous and(2) the amorphous nature of the representation makes iteven more open to Harnad’s criticism that it is insuffi-cient for identification (Harnad, 1990).
4.2. Internally Generating the AppropriateRepresentations
Since class nouns represent categories, the most obviouscandidate for the appropriate representational form is
1310 C. Browne et al.
2 Interestingly, Harnad has recently shown the use of language inconveying explicit knowledge to a group which creates an evolutionaryadvantage over groups with no language where each member mustderive all their explicit knowledge for themselves (Harnad, 1996).
3 Token, rather than icon, is used in this context as reference is beingmade to Fodor and Pylyshyn’s work which is symbol based. The authorwould strenuously argue that arbitrary symbols are insufficient forcognition and that iconic representations are appropriate (Aleksander& Morton, 1993; Aleksander, 1996). Throughout this section, if theword token is used in relation to a connectionist system it should beread as icon. Token is used for consistency and chosen for its generality.Icons are a subset of tokens, thus token is universally applicable.

Harnad’s categorical representations (Harnad, 1987b)described above (Section 2.2.1) and dubbed as reducediconic representations for clarity. Such reduced descrip-tions are entirely commensurate with Hinton’s proposals(Hinton, 1988).
4.2.1.Learning Rules.Two learning rules, Bienenstock,Cooper and Munro (BCM) [Bienenstock et al., 1982],and Local Error-driven and Associative BiologicallyRealistic Algorithm (LEABRA) [O’Reilly, 1996] overtlydisplay representational reduction whilst maintainingstructural integrity. Previous work (Browne & Parfitt,1997) describes the features of both these learningrules and posits them as ideal candidates for the learningrules underlying a neural system capable of repre-sentational redescription. O’Reilly gives an excellentaccount of the inadequacy of backpropagation in main-taining the structural form of the environment (O’Reilly,1996) and so it is not considered further as a learningrule.
Intrator (Intrator, 1992) has applied exploratory pro-jection pursuit methods in order to derive a modificationto the original BCM learning rule (Bienenstock et al.,1982). The technique results in an unsupervised learningrule for input to output dimensionality reduction whichseeks projections that emphasise distinguishing features.Such a learning rule is ideally suited to creating reducediconic representations.
4.2.2.Approximationism.It is important to note that thefeatures extracted by Intrator’s (Intrator, 1992) learningrule emphasise distinguishing features in only the datapresented. It is therefore possible that a feature whichdistinguishes two categories for a portion of the data islater proven to be inadequate for the task and thereforerequires refinement. Harnad (1987b) frames the problemas follows:
Consider a simple problem in machine vision: Suppose allthat a visual scene categoriser had to do was to tell aparttrees from animals, that is, to categorise all instances of treesas trees and all instances of animals as animals. Suppose, byway of further simplification, that trees and animals werethe only patterns the analyser ever encountered, and suppose(to simplify still further) that its input patters were alreadysuitably smoothed and parsed so that they only appeared instandard positions, with figure and ground, parts and whole,already appropriately sorted.
Now it is evident that if there were nothing to worry aboutbut sorting trees and animals under these canonical con-ditions, a very simple rule would work quite well, forexample, counting the number of legs, L, and calling aninstance a tree if L was less than or equal to 1 and an animalotherwise. Obviously such a rule could only work with wellsmoothed and parsed inputs, free of anomalous instances(such as storks standing on one leg, trees with split trunks,or tables). As an approximation, however, the rule wouldsort the standard cases described. As anomalies were
introduced, the rule could be revised and elaborated so asto tighten the approximation in accordance with the newcontingencies (Harnad, 1987b).
Harnad (1987b) refers to this effect as ‘‘Approxima-tionism’’. The suggestion being that a categorical repre-sentation is not a fixed absolute, but an ever improvingapproximation driven by the nature of the context; beingthe noncategory members the category members have tobe differentiated from.
4.2.3.Construction of Category Representations OccursOver Time.The implication of approximationism forthe internally generated category representation is thatthis must occur over time. The feature extractionsystem will develop features which distinguish categorymembers from previously encountered nonmembers, butas anomalies or new items which from other nonmembercategories are experienced the set of reduced featureswhich defines the category representation will requirerefinement or addition.
4.2.4.Neural Architecture Capable of Building Appro-priate Category Representations.Previous work(Browne & Parfitt, 1997) has suggested a two unitneural system (Figure 7) for producing internallydriven category representations. The first networkextracts the category features from sensory stimuliusing Intrator’s modification (Intrator, 1992) to theBCM rule (Bienenstock et al., 1982). The second net-work then recombines these extracted features as theyare extracted over time by means of its feedback loopin much the same manner as Elman’s Simple RecurrentNetwork (Elman, 1990) constructs temporal relations inthe context units. If, in contrast to Elman (1990), whouses error backpropagation (Rummelhart et al., 1986), astructure preserving algorithm, such as the two suggestedabove (Section 4.2.1) is used in the second network,then the evolved category representation maintains itsiconic form as desired.
5. COMPARISON WITH OTHER APPROACHES
It is certainly not intended to claim in this paper thatautomata theory is the only approach to understandingconsciousness. The aim of this discussion is to argue thatthe described automata based theory does provide auseful theoretical framework for exploring conscious-ness, or at least the particular aspects focussed on here;redescription and abstraction. It is, therefore, importantto note the essential features of iconically trained neuralstate machines that differentiates them from otherapproaches. The authors are perfectly willing to acceptthat other approaches may satisfy the fundamentalpostulate and corollaries. The theory is, however,couched in terms of state spaces which fits naturallywith neural state machines. The appropriateness of
1311Consciousness and Neural Cognizers

automata theory to the study of consciousness is of littlesurprise considering the fact that it was developedagainst a background of the Macy Conferences on Feed-back Mechanisms and Circular Causal Systems inBiology and the Social Sciences, at which the questionof mechanistic consciousness was raised at an early stagein the development of computing (Asprey, 1990).
It is first important to point out that the iconic trainingis an essential feature. Iconic training ensures that thesystem is not subject to the symbol grounding problem.The symbol grounding problem relates to the absence ofintrinsic meaning in a system of arbitrary symbols,the currency of classical Artificial Intelligence. Searle(1980) provided one of the original demonstrations ofthe issue in what he called ‘‘The Chinese Room’’ thoughtexperiment. Harnad has reformulated the idea in a clearerform by describing the never ending search for meaningby a non speaker attempting to translate a Chinese textfrom a Chinese-to-Chinese dictionary (Harnad, 1990).Both these arguments are considered to be powerfulcriticisms of any claim computationalism has to model-ling cognition or consciousness. Harnad takes the ideasa stage further using them to refute the claim that cog-nition is computation (Harnad, 1994). The authors, there-fore, suggest that symbolic Artificial Intelligence, asit currently stands, is not an appropriate model ofconsciousness.
Neural networks, on the other hand, derive their repre-sentations directly from the environment and are there-fore less susceptible to the symbol grounding problem.It is not true to say that neural networks are immune
from the problem. If the neural network uses a localistrepresentation at any stage of processing, then theenvironmental structure present at the input is lost andthe symbol grounding problem re-emerges. However,there are a large number of neural network modelswhich employ distributed representations throughout.Further discussion is limited to these systems.
Above (Section 1), it was stated that Fodor andPylyshyn’s (Fodor & Pylyshyn, 1988) objections to con-nectionism had been refuted. Although this is true, it hasnot as yet been fully demonstrated how a connectionistnetwork might generate flexible representations whichare available throughout a system (Clark & Karmiloff-Smith, 1993). Current neural networks are capable ofgenerating concept trees (Pollack, 1990), but this stilldoes not describe the full capacity for internally driven,recursive concept abstraction demanded by Clark &Karmiloff-Smith (1993).
The failure of both traditional Artificial Intelligenceand connectionist approaches in cognitive modellinghas lead, in an attempt to find a successful compromise,to the development of a number of hybrid systems.Implicit in hybrid systems is the hope that the juxtaposi-tion of the two approaches will alleviate the inadequaciesof both. Unfortunately, what is equally as likely is thatthe resultant system will suffer from the deficienciesof both connectionism and computationalism. Symbolgrounding demands that there is a clear, consistantprocess by which higher-level, more abstract symbolsare generated from lower-level ones. The break in therepresentational scheme between the symbolic and
1312 C. Browne et al.
FIGURE 7. A schematic of a suggested system to learn the concept of rotation through the redescription of iconically learned states. Thefirst network extracts features from the currently presented views (in this case a bar down the left in the upper image and a bar down theright in the lower image). An intermediate state is formed from the combination of the features extracted from the current trainingexample. These sets of features extracted from a number of individual pairs of training examples are combined, over time, in the second(right hand) network, to form a representation of the overall concept. This representation consists of the combination of all the featuresextracted from individual instances of the concept.

connectionist regions of a hybrid system means that theyare still susceptible to the symbol grounding problem, ifat a higher-level. Also, the tokens utilised by symbolprocessing region of the system are constrained by theconnectionist system, so may well be far from ideal forthe required computation.
What is required is for either a symbol system touneqivocably overcome the symbol grounding problem,or the development of a connectionist system with thecapacity to flexibly manipulate its representations. It issuggested that the automata theory approach describedhere does provide a clear view as to how the latter typeof system might be constructed. That is not to say thatother approaches are incapable of some, or even all, ofthe functions outlined by the corollaries. The automatatheory framework taken as an overall approach, however,does stand out, as it does provide useful insight intocognitive functioning which other approaches have sofar failed to clearly reveal.
What the automata theory approach described in thispaper offers can be summarised as a clear view as to howa neural system might build up a set of internally gener-ated, hierarchical, flexible representations in a number ofdomains. Such representations have been claimed to be atthe heart of cognition (Clark & Karmiloff-Smith, 1993)and are later argued to be commensurate with a contem-porary philosophy of mind (Section 6).
6. CONCLUSIONS: WHERE DOESCONSCIOUSNESS COME IN?
A fair critique of the above would be that all that has beensuggested is method for learning to represent worlds con-taining nested concepts, so why have a fanciful referenceto the difficult and woolly concept of consciousness? Infact, why refer to a program of work which goes underthe heading of Artificial Consciousness at all? Theanswer lies in the fundamental postulate and thereforeits associated corollaries. A neural state machine hasbeen shown to satisfy the requirements of corollaries 4,iconic learning and 8, the representation of meaning.Corollary 4 relates to consciousness by suggesting thaticonic transfer creates ‘‘sentience’’ by making neuralfiring patterns meaningful in terms of the organism’ssensory world. It is this corollary which gives the propo-sals immunity from the symbol grounding problem, dis-cussed above.
Corollary 8 is discussed as being the key to the repre-sentation of abstractions such as place and categorywithin a neural state machine.
Of course, any attempt at a neural construction of theproperties of consciousness can be dismissed purely onthe grounds that some definition of consciousness canbe found which excludes the activity altogether eitherfor not being biologically relevant or for being a philo-sophical error of category as is the belief of some of theauthors discussed above (Section 2.1). It is arguable,
however, that the model developed in this paper istrue to the style of enquiry advocated by Locke [asquoted above (Section 2.1)]. The principle of an iconictransfer comes from the ‘‘consciousness to one’s self’’ of‘‘ideas, notions or whatever else you please to callthem...’’
Of all the contemporary philosopher’s, Searle ispossibly the closest to a continuation of the line ofenquiry commenced by Locke. Searle’s position is welldefended in Searle (1992). It can be summarised asfollows. Consciousness is an emergent property ofthe brain. We do not understand how this occurs as yet,but advances in science will unravel the mystery intime. For the present we should just accept that one ofthe functions of the brain is to cause consciousness.Consciousness is inextricably bound to mental life. Thecontents of consciousness are mental phenomena.Unconscious mental phenomena are only describablein terms of their similarity to conscious ones. Althoughunconscious mental phenomena may never become thecontent of consciousness they must have an identicalform. Attributes of the brain which can never enterconsciousness are nonconscious. Searle differentiatesunconscious from nonconscious mental phenomena asfollows:
Think of the difference, for example, between my belief(when I am not thinking about it) that the Eiffel Tower isin Paris, and the myelination of the axons in my centralnervous system. There is a sense in which both are uncon-scious. But there is a big difference between them in that thestructural states of my axons couldn’t themselves be con-scious states, because there isn’t anything mental aboutthem. I assume for the sake of this argument that myelina-tion functions essentially in the production of my mentalstates, but even if myelinated axons were themselves objectsof experiences, even if I could feel inwardly the state of themyelin sheaths, still the actual structures are not themselvesmental states. Not every unconscious feature of my brainthat (like myelination) functions essentially in my mentallife is itself a mental feature. But the belief that the Eiffeltower is in Paris is a genuine mental state, even though ithappens to be a mental state that is most of the time notpresent to consciousness. So here are two states in me, mybelief and my axon myelination: both have something to dowith my brain and neither is conscious. But only one ismental, and we need to get clear about what makes it mentaland the connection between that feature—whatever it is—and consciousness. Just to keep this distinction clear, I pro-pose in this chapter to call phenomena like myelination,which are not in the mental line of business at all, ‘‘non-conscious’’ and phenomena like mental states that I am notthinking about or have repressed ‘‘unconscious’’ (Searle,1992).
In the neural state machine approach to consciousness,the fundamental postulate defines the current systemstate as the existing conscious state. Other iconic statesin the learned state structure are possible candidates forbecoming the system state, so in Searle’s terminology are
1313Consciousness and Neural Cognizers

unconscious mental phenomena. Since the neural statemachine approach to consciousness is defined in termsof state spaces, little reference need be made to the actualstructure of the system, although it is worth mentioningthat the MAGNUS simulation (Section 2.3) describedabove is extremely adept for the task, which compriseSearle’s nonconscious mental phenomena need not bestudied in detail.
The insistence on iconic learning provides the systemwith both symbol grounding (Searle, 1980; Harnad,1990) at the bottom-level and a maintenance of theaspectural shape (Searle, 1992) of the higher-levelmental states.
In this light, the items discussed above are well withinthe realms of the study of consciousness and might proveto be a fruitful approach to a complex problem.
REFERENCES
Aleksander, I. (1995). Artificial neuroconsciousness: An update. InJ. Mira & F. Sandoval (Eds.),From natural to artificial neuralcomputation. (pp. 566–583). Heidelberg: Springer.
Aleksander, I. (1996).Impossible minds: my neurons, my conscious-ness. IC Press.
Aleksander, I., Evans, R. & Penny, W. (1993). MAGNUS: An iconicneural state machine. InProceedings of the Weightless NeuralNetworks Workshop.
Aleksander, I. & Hanna, F. K. (1975).Automata theory: An engineeringapproach. New York: Crane Russak.
Aleksander, I., & Morton, H. (1991). General neural unit: retrievalperformance.Electronic Letters, 27, 1776–1777.
Aleksander, I. & Morton, H. (1993).Neurons and symbols. Chapmanand Hall.
Asprey, W. (1990).John von Neumann and the origins of moderncomputing. MIT Press.
Bienenstock, E. L., Cooper, L. N., & Munro, P. W. (1982). Theory forthe development of neuron selectivity: Orientation specifity andbinocular interaction in the visual cortex.Journal of Neuro-science, 2, 32–48.
Browne, C., & Aleksander, I. (1996). Digital general neural units withcontrolled transition probabilities.Electronics Letters, 32(9),824–825.
Browne, C. J. & Parfitt, S. (1997). Iconic learning and epistemology. InDoes representation need reality? Proceedings of the InternationalConference New Trends in Cognitive Science. Vienna: AustrianSociety of Cognitive Science Report 97-01.
Clark, A., & Karmiloff-Smith, A. (1993). The cognizer’s innards.Mindand Language, 8(4), 487–519.
Crick, F. (1994).The astonishing hypothesis. New York: Scribner’s.Dennett, D. C. (1991).Consciousness explained. Penguin Books.DesCartes, R. (1637).Discourse on method.Elman, J. (1990). Finding structure in time.Cognitive Science, 14,
179–211.Evans, R. G. (1996).A neural architecture for a visual exploratory
system. PhD thesis, Imperial College of Science, Technology andMedicine.
Fodor, Z. W., & Pylyshyn, J. (1988). Connectionism and cognitivearchitecture: A critical analysis.Cognition, 28, 3–71.
Greenfield, S. (1995).Journeys to the centres of the mind. New York:Freeman.
Harnad, S. (1982).Metaphor and mental duality. Hillsdale, NJ:Lawrence Earlbaum Associates.
Harnad, S. (1987a).Categorial perception: a critical overview. NewYork: Cambridge University Press.
Harnad, S. (1987b).Category induction and representation. New York:Cambridge University Press.
Harnad, S. (1990). The symbol grounding problem.Physica D, 42,335–346.
Harnad, S. (1992).Connecting object to symbol in modelling cognition.Springer.
Harnad, S. (1994). Computation is just interpretable symbol manipula-tion: Cognition isn’t. Minds and Machines, 4(Special issue on‘‘What is Computation?’’), 379–390.
Harnad, S. (1995).The origins of words: A psychological hypothesis.Muenster: Nodus Publishers.
Harnad, S. (1996). On the virtues of theft over honest toil: Groundinglanguage and thought in sensorimotor categories. InProceedings ofthe Hang Seng Centre Conference on Language and Thought.
Hinton, G. E. (1988). Representing part-whole hierarchies in connec-tionist networks. InProceedings Tenth Annual Conference of theCognitive Science Society(pp. 48–54). Montreal, Quebec.
Intrator, N. (1992). Feature extraction using an unsupervised neuralnetwork.Neural Computation, 4, 98–107.
Karmiloff-Smith, A. (1990). Constraints on representational change:Evidence from children’s drawing.Cognition, 34, 57–83.
Karmiloff-Smith, A. (1992).Beyond modularity. MIT Press.Locke, J. (1975).Essay concerning human understanding. Oxford:
Clarendon Press.Lucas, J. R. (1994). A view of one’s own.Philosophical Transactions of
the Royal Society, London. A, 349,147–152.Nagel, T. (1974). What is it like to be a bat?Philosophical Review, 83,
435–450.O’Reilly, R. C. (1996).The LEABRA model of neural interactions and
learning in the neocortex. PhD thesis, Carnegie Mellon University,Pittsburgh.
Paivio, A. (1986).Mental representation: A dual coding approach.Oxford, New York.
Penrose, R. (1994).Shadows of the mind. Oxford University Press.Pollack, J. (1990). Recursive distributed representations.Artificial
Intelligence, 46(1-2), 77–105.Quine, W.V.O. (1960).Word and object. MIT Press.Rummelhart, D.E., Hinton, G.E., & Williams, R. (1986).Learning
internal representations by error propagation(pp. 45–76). Cam-bridge, MA: MIT Press.
Sales, N. J. (1996).Aspects of lexical development in artificial neuralnetworks. PhD thesis, Imperial College of Science, Technology andMedicine.
Searle, J. R. (1980). Minds, brains and programs.Behavioural andBrain Sciences, 3, 417–424.
Searle, J. R. (1992).The rediscovery of the mind. The MIT Press.Sloman, A. (1994). Semantics in an intelligent control system.Philo-
sophical Transactions of the Royal Society, London A, 349,43–58.Smolensky, P. (1987). The constituent structure of connectionist mental
states: A reply to Fodor and Pylyshyn.The Southern Jnl ofPhilosophy, XXVI(Supplement), 137–161.
Soloman, R. (1993).Introducing philosophy. Harcourt Brace CollegePublishers.
Van Gelder, T. (1990). Compositionality: a connectionist variation on aclassical theme.Cognitive Science, 14, 355–384.
Wittgenstein, L. (1953).Philosophical investigations. New York:Macmillan.
APPENDIX A: A SUMMARY OF THEARTIFICIAL CONSCIOUSNESS FRAMEWORK
A.1. The Fundamental Postulate
The personal sensations which lead to the consciousness ofan organism are due to the firing patterns of some neurons,such neurons being part of a larger number which form the
1314 C. Browne et al.

state variables of a neural state machine, the firing patternshaving been learned through a transfer of activity betweensensory input neurons and the state neurons.
A.1.1. Corollary 1: The Brain is a State Machine
The brain of a conscious organism is a state machine whosestate variables are the outputs of neurons. This implies that adefinition of consciousness be developed in terms of theelements of automata theory.
A.1.2. Corollary 2: Inner Neuron Partitioning
The inner neurons of a conscious organism are partitionedinto at least three sets:
Perceptual Inner Neurons: responsible for percep-tion and perceptual memory;Auxiliary Inner Neurons: responsible for inner‘‘labelling’’ of perceptual events;Autonomous Inner Neurons: responsible for ‘‘life-support’’ functions—not involved in consciousness.
A.1.3. Corollary 3: Conscious and UnconsciousStates
The contribution to consciousness of the inner neurons andthe sensory neurons has three major modes:
Perceptual:which is active during perception—whensensory neurons are active;Mental, conscious:which is an act of thinking in thesame neurons even when sensory neurons are inactiveor otherwise engaged;Mental, unconscious:which is activity generated bythe neurons involved in conscious activity, but whichdoes not cause sensations of consciousness.
A.1.4. Corollary 4: Iconic Learning
To qualify for a contribution to consciousness, firingpatterns in the inner perceptual/conscious neurons need tobe created through dominant neural inputs which samplethe activity of outer sensory neurons and influence thefunction of the inner neurons. This has been dubbed ‘‘iconiclearning’’.
A.1.5. Corollary 5: Prediction
Relationships between world states are mirrored in the statestructure of the conscious organism enabling the organismto predict events.
A.1.6. Corollary 6: The Awareness of Self
As a result of iconic learning and feedback betweenphysical output and the senses, the internal state structureof a conscious organism carries a representation of its ownoutput and the effect that such an output can have on worldstates.
A.1.7. Corollary 7: Will
The organism, in its mental mode, can enter state trajec-tories according to need, desire or in an arbitrary mannernot related to need. This gives it its powers of acting in botha seemingly free and purposeful manner.
A.1.8. Corollary 8: Representation of Meaning
When sensory events occur simultaneously or in close timeproximity in different sensory modalities, iconic learningand generalisation of the neural state machine ensures thatone can be recalled from the other.
A.1.9. Corollary 9: Learning Utterances
The feedback loop responsible for the creation of ‘‘self’’representations is also responsible for the creation of staterepresentations of the basic utterances of the organismwhich are retrieved in response to the utterances of other,‘‘adult’’, organisms and may be used by the adult to teachthe organism more complete utterances such as the words ofa language.
A.1.10. Corollary 10: Learning Language
Language is a result of the growth process of a social repo-sitory from which it can be learned by a conscious organism,given the availability of knowledgeable ‘‘instructors’’. Thestructure of such language is a process of the social evolu-tion of a best-match to the development of state structure inthe organism.
A.1.11. Corollary 11: Iconic Transfer ofQualitative Sensory Properties (Qualia?)
Iconic transfer operates on all aspects of sensoryperception discriminated by receptors. Therefore qualitativeproperties of objects such as colour become candidatesfor iconic transfer and representation in recallable statestructure.
1315Consciousness and Neural Cognizers

A.1.12. Corollary 12: Instinct
To enhance survival, an organism needs a substrate of out-put actions that are linked to inputs or inner autonomicneurons and which are or become independent of the stateof inner neurons. These may be inborn or become inde-pendent of state variables as a result of development.
A.1.13. Corollary 13: Emotions
Emotion in a neural state machine is an iconic encodingof instinctive sensations. It leads to behaviour whichenhances the avoidance of danger and the protection ofsurvival.
1316 C. Browne et al.

1997 SPECIAL ISSUE
Learning, Action and Consciousness: A HybridApproach Toward Modelling Consciousness
Ron Sun
Department of Computer Science, The University of Alabama
(Received2 July 1996;accepted4 December1996)
Abstract—This paper is an attempt at understanding the issue of consciousness through investigating its functionalrole, especially in learning, and through devising hybrid neural network models that (in a qualitative manner) approxi-mate characteristics of human consciousness. In doing so, the paper examines explicit and implicit learning in a varietyof psychological experiments and delineates the conscious/unconscious distinction in terms of the two types of learningand their respective products. The distinctions are captured in a two-level action-based model Clarion. Some funda-mental theoretical issues are also clarified with the help of the model. Comparisons with existing models of conscious-ness are made to accentuate the present approach.q 1997 Elsevier Science Ltd.
Keywords—Neural networks, Hybrid systems, Consciousness, Implicit learning, Reinforcement learning, Proceduralknowledge, Rule extraction, Dual representation.
1. INTRODUCTION
Amidst the widespread enthusiasm of recent years con-cerning the scientific study of consciousness, there are alarge number of models being proposed (including com-putational models, which in turn include neural networkmodels), and various claims have been made about them.These models capture to various extents experimentalfindings and pretheoretical intuitions about conscious-ness (see, e.g. Taylor, 1994; Schacter, 1990; Jackendoff,1987; Shallice, 1988; Baars, 1988; Dennett & Kins-bourne, 1992; Penrose, 1994). Unfortunately, however,some of these models (such as Dennett & Kinsbourne,1992; Shallice, 1988; Jackendoff, 1987) are aimed at avery high and gross level of explanation (e.g. overallarchitectures) and thus unable to provide more detailedpredictions and explanations. On the other hand, existingcomputational, especially neural network, models tend torush directly into complex neural physiological thickets(Taylor, 1994; Edelman, 1989) and thus may lose sight offorests. In addition, most existing models do not dealadequately with one crucial aspect of human conscious-ness: learning. In contrast to these approaches, we intend
to stay at an intermediate and functional level; investi-gating the detailed functional roles of consciousnessand determining how various aspects of the consciousand the unconscious should figure into the architectureof the mind (in terms of learning as well as performance).In other words, we posit a middle level between phenom-enology and physiology/neurobiology, which might bemore apt at capturing fundamental characteristics of con-sciousness. We will also link computational models ofconsciousness to parts of (phenomenological) philo-sophy that are concerned with consciousness.
As we will focus mainly on the learning aspect inconsciousness, let us briefly describe the learning settingsthat we examined (from Sun et al., 1995). Thesesettings are more complex than simple categorisation/classification and though action-based, involve morethan just simple stimulus–response pairing. Psychologi-cal experiments involving dynamic decision making orartificial grammars will be discussed. In dynamic deci-sion making (Berry & Broadbent, 1988), subjects wererequired to control the levels of an output variable bymanipulating levels of an input variable. In one instance,subjects were to manage a simulated sugar productionfactory and the goal was to reach and maintain a parti-cular level of productivity by manipulating the size of theworkforce. In another instance, subjects were to interactwith a computer simulated ‘‘person’’ and to maintain thebehaviour of the person at ‘‘very friendly’’ by manipu-lating his/her own behaviour. In artificial grammar
Neural Networks, Vol. 10, No. 7, pp. 1317–1331, 1997q 1997 Elsevier Science Ltd. All rights reserved
Printed in Great Britain0893–6080/97 $17.00+.00
PII: S0893-6080(97)00050-6
Pergamon
1317
Acknowledgements: This work was supported in part by Office ofNaval Research grant N00014-95-1-0440. Thanks to Ed Merril andDiana Gordon for their comments.
Requests for reprints should be sent to Ron Sun, Department ofComputer Science, The University of Alabama, Tuscaloosa, AL35487, USA; Tel.: (205) 348-1667; e-mail: [email protected].

learning (Reber, 1989), subjects were presented with astring of letters that were generated in accordance witha simple grammar. Although subjects were unaware ofthe underlying grammars, they were asked to judge thegrammaticality of novel strings. In addition, two naviga-tion tasks were used for both psychological experimentsand computational modelling in our lab (see Sun et al.,1995, 1996a, b). One is maze running, in which froma starting location, a subject has to find a goal location,using only local sensory information. The other isnavigation through minefields, in which a subjectis required to go from a starting location to a goal loca-tion within a given short period of time by navigatingthrough a densely packed field of mines (which willexplode if the subject gets too close). I will later showthe relevance of these learning tasks to the study ofconsciousness.
In the rest of this paper, I will first present a cognitivearchitecture Clarion1 for accounting for the distinctionof the conscious and the unconscious (in Section 2). I willthen show how it accounts for a number of phenomenarelated to the conscious/unconscious distinction in learn-ing (which may also be referred to as the explicit andimplicit distinction) (in Section 3). A discussion of somefundamental theoretical issues will take place after that(Section 4). A comparison to existing models of con-sciousness such as Baars (1988), Schacter (1990), andDamasio (1994) will follow, which will show the com-monalities shared by some of these models and Clarionand the unique features of the present model (in Section5). Some concluding remarks (Section 6) will completethe paper.
2. A HYBRID NEURAL NETWORK MODEL
A computational model that can tackle the learning tasksmentioned above is needed. It needs to satisfy some basicrequirements as follows. It must be able to learn fromscratch on its own (as human subjects often do in thelearning tasks outlined earlier; Berry & Broadbent,1988; Reber, 1989; and also Sun et al., 1996a). Themodel also has to perform concurrent, on-line learning.That is, it has to learn continuously from on-goingexperience in the world; for, as indicated by Medinet al. (1987), Nosofsky et al. (1994) and others, humanlearning is often gradual, on-going and concurrent, whichis true of all the aforementioned tasks. As suggested byAnderson (1983) and many others, there are clearly twotypes of knowledge involved in human learning — pro-cedural and declarative: while one is generic and easilyaccessible, the other is embodied and specific. Moreover,different types of learning processes are involved inacquiring different types of knowledge (Anderson,
1983; Keil, 1989; Smolensky, 1988; Stanley et al.,1989). Humans are able to learn procedural knowledgethrough trial and error (without a priori knowledge) in theaforementioned tasks. On top of low-level proceduralskills, declarative knowledge can be acquired alsothrough on-going experience in the world (see Stanleyet al., 1989). Furthermore, it is important for declarativeknowledge to be learned through the meditation of lowlevel skills (i.e. bottom-up learning; see Sun et al.,1996a).
Procedural knowledge (skills) can be captured by sub-symbolic distributed representation, such as that pro-vided by a backpropagation network. Because of theimplicit nature of procedural skills, details of such skillsare in general inaccessible to consciousness (Anderson,1983; Reber, 1989). A distributed representation natu-rally captures this property with representational unitsthat are capable of accomplishing tasks but are in generaluninterpretable and subsymbolic (Sun, 1994, 1995). (Asymbolic representation may be used, but then this wouldrequire an artificial assumption that these representationsare not accessible, while other similar representations areaccessible — such a distinction is arbitrary.)
Procedural knowledge can be learned in a couple ofdifferent ways. In the case where correct input/outputmappings are provided, straight backpropagation canbe used on a neural network. Otherwise, reinforcementlearning can be used (Sutton, 1990; Watkins, 1989). Thisis preferred because there is often no uniquely correctaction in the aforementioned tasks, although feedback isusually available. Using reinforcement learning in neuralnetworks, we can measure the goodness of an actionthrough a payoff-reinforcement signal. An adjustmentcan be made to weights to increase the chance of select-ing the actions that receive positive reinforcement andto reduce the chance of selecting the actions that receivenegative reinforcement.
This level can be modular; that is, a number of smallnetworks can co-exist each of which is adapted tospecific modalities, tasks, or groups of input stimuli.This coincides with the well known modularity claim(Fodor, 1983; Karmiloff-Smith, 1986; Cosmides &Tooby, 1994), in that much processing in the humanmind is done by limited, encapsulated (to some extent),specialized processors that are highly efficient. It is alsosimilar to the idea of Shallice (1988) that a multitude of‘‘action systems’’ compete with each other. There alsohas been some work in neural network and machinelearning communities in developing modular systems,which are equally relevant.
On the other hand, declarative knowledge can becaptured by a symbolic or a ‘‘localist’’ representation(Clark & Karmiloff-Smith, 1993), in which each unithas a clear conceptual meaning or interpretation. Thisallows declarative knowledge to be highly accessibleand inferences to be performed explicitly (Smolensky,1988; Sun, 1994, 1995).
1318 R. Sun
1 It was originally developed for modelling human skill learning; seeSun et al. (1995).

Declarative knowledge can be learned in a variety ofways. In this work, because of the dynamic on-goingnature of the learning tasks, we need to be able to dyna-mically acquire a representation and to modify the repre-sentation subsequently if necessary, in an efficient oreven one-shot fashion.
The difference in representing procedural and declara-tive knowledge revealed by the above discussion leadsnaturally to a two-level architecture, in which one level isprocedural and the other declarative. This structuring canbe argued on both psychological and philosophicalgrounds. Anderson (1983) put forward the dichotomyof separate yet interconnected declarative and proceduralknowledge bases to account for a variety of learning data.Smolensky (1988) suggested that the separation of con-ceptual-level and subconceptual-level processing. Theconceptual level possesses three characteristics: (1)public access; (2) reliability; and (3) formality. It canthus be modelled by symbolic processing. In contrast,skills, intuition, and the like are not expressible in lin-guistic forms and do not conform to the three criteriaprescribed. Hence, skills and intuition constitute a differ-ent type of capacity, reflecting the ‘‘subsymbolic’’ pro-cessing at the subconceptual level (see also Shiffrin &Schneider, 1977). In a similar vein, Dreyfus and Dreyfus(1987) contrasted analytical and intuitive thinking, froma phenomenological analysis of human cognitive skilllearning in which the fluent, holistic and situation sensi-tive way of solving problems (intuition) as observed inmaster level performers is in sharp contrast with theslow, deliberate thinking that often occurs in the novices(analytical thinking). Models have been proposed toaccount for such two-tiered structures, which oftenposit the existence of at least two separate components,each of which responds to one side of a dichotomy (e.g.Posner & Snyder, 1975; Schacter, 1990; Murphy &
Medin, 1985; Keil, 1989; Sun, 1992a, 1994; Sun &Bookman, 1994). The dual representation hypothesisput forth in Sun (1994) stated that:
It is assumed in this work that cognitive processes arecarried out in two distinct levels with qualitatively differentprocessing mechanisms. Each level encodes a fairly com-plete set of knowledge for its processing, and the coverageof the two sets of knowledge encoded by the two levelsoverlaps substantially.
Based on the above considerations, we developedClarion: Connectionist Learning with Adaptive RuleInduction ON-line. It consists of two main components:the top level encodes explicit declarative knowledge, andthe bottom level encodes implicit procedural knowledge.In addition, there is an episodic memory, which storesrecent experiences in the form of ‘‘input, output, result’’(i.e. stimulus, response, and consequence) that arerecently-filtered (episodic memory will not be used inthis paper and therefore will not be further discussedhere) (see Figure 1).
An overall pseudo-code algorithm that describes theoperation of Clarion is as follows:
1. Observe the current statex (in a proper representa-tion).
2. Compute in the bottom level theQ-values ofx asso-ciated with each of the possible actionsai ’s:Q(x,a2), :::,Q(x, an). Select one action or a fewbased onQ-values.
3. Find out all the possible actions(b1,b2, :::,bm) at thetop level, based on the inputx (sent up from thebottom level) and the rules in place.
4. Compare the values of the selectedai ’s with those ofthebj ’s (sent down from the top level), and choose anappropriate actionb.
1319Learning, Action and Consciousness: A Hybrid Approach Toward Modelling Consciousness
FIGURE 1. The CLARION architecture.

5. Perform the actionb, and observe the next statey and(possibly) the reinforcementr.
6. UpdateQ-values at the bottom level in accordancewith the Q-Learning-Backpropagationalgorithm.
7. Update the rule network at the top level using theRule-Extraction-Refinementalgorithm.
8. Go back to step 1.
In the bottom level, aQ-value is an evaluation of the‘‘quality’’ of an action in a given state:Q(x, a) indicateshow desirable actiona is in statex (which consists ofsome sensory input). We can choose an action based onQ-values, e.g. by choosing the one that has the maximumQ-value in the current state or by choosing an actionprobabilistically based onQ-values. To acquire theQ-values, one option is to use theQ-learningalgorithm(Watkins, 1989), a reinforcement learning algorithm.2 Inthe algorithm,Q(x, a) estimates the maximum discountedcumulative reinforcement that the agent will receivefrom the current statex on:
max∑i ¼ 0
gi ri
!(1)
whereg is a discount factor that favours reinforcementreceived sooner relative to that received later, andri isthe reinforcement received at stepi (which may be 0).The updating ofQ(x,a) is based on minimising
r þ ge(y) ¹ Q(x,a) (2)
where g is a discount factor ande(y) ¼ maxaQ(y,a).Thus, the updating is based on thetemporal differencein evaluating the current state and the action chosen. Inthe above formula,Q(x,a) estimates, before actiona isperformed, the (discounted) cumulative reinforcement tobe received if actiona is performed, andr þge(y) esti-mates, after actiona is performed, the (discounted)cumulative reinforcement that the agent will receive; sotheir difference (the temporal difference in evaluating anaction) enables the learning ofQ-values that approximatethe (discounted) cumulative reinforcement. UsingQ-learning allows sequential behaviour to emerge. Throughsuccessive updates of theQ-function, the agent can learnto take into account future steps in longer and longersequences.3
We chose to use a four-layered network for implemen-tation (see Figure 2), in which the first three layers form a(either recurrent or feedforward) backpropagation net-work for computing Q-values and the fourth layer(with only one node) performs stochastic decisionmaking. The network is internally subsymbolic andimplicit in representation (in accordance with our
previous considerations). The output of the third layer(i.e. the output layer of the backpropagation network)indicates theQ-value of each action (represented by anindividual node), and the node in the fourth layer deter-mined probabilistically the action to be performed basedon the Boltzmann Distribution (i.e. Luce’s choice axiom;Watkins, 1989):
p(alx) ¼e1=aQ(x, a)∑
ie1=aQ(x, ai )
(3)
Here,a controls the degree of randomness (temperature)of the decision making process.4 The combination ofQ-learning and backpropagation facilitates the develop-ment of procedural skills in the bottom level, which canpotentially be done solely on the basis of acting andexploring in the real world. This learning processperforms both structural credit assignment and temporalcredit assignment.
In the top level, declarative knowledge is captured in asimple propositional rule form. To facilitate correspon-dence with the bottom level and to encourage uniformityand integration (Clark & Karmiloff-Smith, 1993), wechose to use a localist network model for representingthese rules. Basically, we connect the nodes representingconditions of a rule to the node representing the conclu-sion. However, we need to determine how we wire upa rule involving conjunctive conditions. There are anumber of previous attempts (e.g. Sun, 1992b; Towel& Shavlik, 1993) that we can draw upon. For eachrule, a set of links can be established, each of whichconnects to a concept in the condition of a rule to theconclusion of the rule. So the number of incoming linksto the conclusion of a rule is equal to the number ofconditions of the rule. If the concept in the condition is
1320 R. Sun
2 Supervised learning methods can also be applied, when correctmappings of an input and output are available.
3 In terms of both simplicity and performance,Q-learning is bestamong similar reinforcement learning methods (Lin, 1992; Sun et al.,1995).
FIGURE 2. The Q-Learning method.
4 The training of the backpropagation network is based on minimis-ing the following:
erri ¼r þge(y) ¹ Q(x,a) if ai ¼ a
0 otherwise
((4)
where i is the index for an output node representing the actionai .Backpropagation is then applied as usual to adjust the weights. Or,when correct mappings are available for each step, backpropagationcan be directly applied.

in a positive form, the link carries a positive weightw;otherwise, it carries a negative weight —w. Sigmoidalfunctions are used for node activation (as an obviouschoice; other functions are also possible):
1
1þ e
∑i
i iwi ¹ t(5)
The thresholdt of a node is set to ben timesw¹ v, wheren is the number of incoming links (the number of condi-tions leading to the conclusion represented by this node),andv is a parameter, selected along withw to make surethat the node has activation above 0.9 when all of itsconditions are satisfied, and has activation below 0.1when some of its conditions are not met. (Activationsabove 0.9 are considered 1, and activations below 0.1are considered 0; so rules are crisp/binary.) In addition,if there is more than one rule that leads to the sameconclusion, an intermediate node is created for eachsuch rule: all of the conditions of a rule are linked tothe same intermediate node, and then all the intermediatenodes are linked to the node representing the conclusion(see Figure 3). (For more complex rule forms includingpredicate rules and variable binding, see Sun, 1992b).
To fully capture bottom-up learning processes, wedevised an algorithm for learning declarative knowledge(rules) using information in the bottom level. The basicidea is as follows: if an action decided by the bottomlevel is successful (here, being successful could mean anumber of different things, including the differencebetween theQ-value of the state before an action is per-formed and that after the action is performed, whichcomes from the bottom level; the details are specifiedin Sun et al., 1995), then the agent extracts a rule thatcorresponds to the action selected by the bottom leveland adds the rule to the network. Then, in subsequentinteractions with the world, the agent verifies theextracted rule by considering the outcome of applyingthe rule: if the outcome is not successful, then the ruleshould be made more specific and exclusive of the cur-rent case; if the outcome is successful, the agent may tryto generalise the rule to make it more universal (Mitchell,1982). (The detail of the algorithm can be found in Sun etal., 1995.)
At the top level, after rules have been learned,backward and forward chaining reasoning, means-ends
analysis, counterfactual reasoning and explicit hypoth-esis testing can be used. These rules, expressed in the‘‘state → action result’’ form (which constitutes aschema; cf. Waltz, 1991). allows powerful operationsto be performed. Backward chaining means-ends analy-sis is accomplished at the top level through backwardtracing of rule links from the ‘‘result’’, which is thenew state entered after the ‘‘action’’ is performed, tothe state, which is the state before the action is per-formed. This process is successively applied, with thederived ‘‘state’’ as the new state (the result) to be backtraced, until reaching an initial state. All of the actionsinvolved in the derivation are collected, which form aplan for accomplishing the desired final result. Counter-factual reasoning can also be applied (because we haveinformation concerning conditions, actions, and resultsreadily available): one can thus hypothetically altereither the conditions or the action of a rule, and see thechange in (immediate or final) results. Such counter-factual reasoning can be used to justify (or explain) achosen action, or a sequence of such actions (a plan).Explicit hypothesis testing, similar to techniques dis-cussed by e.g. Bruner et al. (1956), Nosofsky et al.(1994), and Michalski (1983), can also be applied atthis level of Clarion.
The algorithm (at step 4) makes the final decision onwhich action to take by incorporating influences fromboth levels (as has been shown by Willingham et al.,1989, in humans, declarative knowledge can influenceprocedural performance). It allows different operationalmodes: (1) relying only on the top level; (2) relying onlyon the bottom level; or (3) combining the outcomes fromboth levels weighing them differently. The weights canchange over time and in different situations. (Thesedifferent operational modes roughly correspond to the
1321Learning, Action and Consciousness: A Hybrid Approach Toward Modelling Consciousness
FIGURE 3. A network for representing rules: (1) b c → a; (2)¬f g → b; (3) ¬h i → b; (4) j → c .
FIGURE 4. The implementation of C LARION.

folk psychological notions of the intuitive mode, thedeliberative mode, and the various mixtures of the twowith different percentages of each; more later; see Sunet al., 1995.) Figure 4 shows the details of the two levelsof the model.
The necessity of a two level architecture that incorpo-rates two types of processes can be summed up asfollows:
• In terms of representation, without the bottom level,the model will not be able to represent proceduralskills properly. Such skills may involve graded, uncer-tain and inconsistent knowledge and autonomousstochastic exploration (with numeric calculation andprobabilistic firing).
• In terms of learning, without learning in the bottomlevel, the model will not be able to learn from experi-ence dynamically to acquire procedural skill. Thebottom level capturesgradual learning of skills,which is different from one-shot rule learning at thetop level.
• Without the top level, the model will not be able to (1)represent generic, easily accessible, and crisp knowl-edge and (2) explicitly access and communicate thatknowledge. When precision, consistency, and cer-tainty are needed, declarative knowledge is preferred.
• Without rule learning, the model will not be able toacquire quickly and dynamically explicit knowledgefor the top level from experience, and therefore haveto resort to externally given declarative knowledge orto procedural knowledge exclusively.
There is ample biological evidence that indicates theexistence of multiple pathways (in visual, linguistic, andother processing modes) some of which lead to consciousawareness, while others do not (e.g. one type is corticalwhile the other is subcortical), as described in Damasio(1994) and LeDoux (1992). For example, LeDoux (1992)described a cortical pathway from stimulus to thalamusto cortex, which produces conscious thoughts, and a sub-cortical pathway from stimulus to thalamus then toamygdala, which can lead directly to brain stem andeffect actions without any explicit process. A two-levelmodel such as Clarion approximates the separation ofthe twokindsofpathways toacertain degree,andsuggests,in a concrete and tangible way, how subcortical pro-cesses can play a fundamental role in supporting and initi-ating consciousness in a bottom-up direction (more later).
3. PSYCHOLOGICAL DATA AND THE MODEL.
The crucial link between this model of procedural/declarative knowledge and the conscious/unconsciousdistinction in humans is in the psychological work onimplicit learning (by e.g. Reber, 1989; Lewicki et al.,1992; Berry & Broadbent, 1988; Stanley et al., 1989;Willingham et al., 1989). Such work shows the disso-ciation between conscious and unconscious learning.
Human knowledge, and its acquisition process, couldbe partially or completely unconscious. The connectionfrom such illustrative data to our model lies in the abilityof the model to account for some of the most importantcharacteristics of human implicit/explicit learning, aswill be sketched below. (More detailed comparisonsand data can be found in Sun et al., 1996a, b)
3.1. The Difference of Conscious/UnconsciousLearning
In the psychological data, there is a clear demonstrationof the difference between conscious and unconscious (or,explicit and implicit) learning. Berry & Broadbent(1988) demonstrated this through an experiment usingtwo similar dynamic decision tasks differing in thedegree of saliency of the input/output relation. Humansubjects were required to maintain the behaviour of acomputer person at a ‘‘very friendly’’ level throughtheir inputs. In the salient version, the computerresponded in accordance with the subjects immediatelypreceding input. In the non-salient version, the computerresponded in accordance with the input prior to that.Results suggested that subjects in the two conditionslearned the tasks in very different ways: subjects inthe non-salient condition learned the task implicitlywhile subjects in the salient condition learned the taskexplicitly, as demonstrated by tests of their explicitknowledge. Reber (1989) described a similar situationin artificial grammar learning. When complex hierarch-ical relations were needed to judge grammaticality, sub-jects tended to use implicit, unconscious learning; forexample, when a sequence consisted of pairings ofadjacent symbols that were ambiguous pair-wise butunambiguous when the entire sequence was viewedthrough hierarchical relations, such as in the case of101110, implicit learning was preferred by the subjects.When only pair-wise relations were needed, such as inthe case of 101010, subjects were more likely to useexplicit, conscious learning by inducing an explicitrule. In other tasks, Cohen et al. (1990) also expresslydemonstrated a dissociation between learning simple(pairwise) relations and learning complex hierarchicalrelations. A pattern emerging from the human data isthat, if the to-be-learned relationships are simple, usuallyexplicit/conscious learning prevails, while, when morecomplex relationships are involved, implicit/unconsciouslearning becomes more prominent. The implicit learningmechanism appears to be more structurally sophisticatedand able to handle more difficult situations (Lewickiet al., 1992). It is important to note the inability ofhuman subjects to articulate their implicitly learnedknowledge, no matter how hard they tried (this is espe-cially true in Lewicki et al., 1992). The subjects wereoften not even aware that they were learning. Never-theless their performance improved over time, whichdemonstrated that their knowledge was unconscious.
1322 R. Sun

This accords well with the Clarion model. In themodel, one can freely move from one type of processto another, by engaging or disengaging the top leveland its associated learning mechanisms (explicit pro-cesses that are consciously accessible, as discussedbefore), or the bottom level and its associated mechan-isms (implicit processes that are not consciously acces-sible). Furthermore, in general, the bottom level is usedto tackle more complex relations while the top level takeson simpler and crisper relations (cf. Reber, 1989; Seger,1994). This is because the top level does not lend itselfeasily to the learning of complex structures due to itscrisp, individuated, and symbolic representation andrigorous learning process. The bottom level, with itsdistributed network representation that incorporatesgradedness and temporal information, handles complexrelations better.
A specific example of this complexity difference is asfollows. Implicit learning of sequences (e.g. artificialgrammar sequences) is biased towards sequences witha high level of statistical structure with much correlation(Stadler, 1992). As has been demonstrated by Elman(1990) and by Cleeremans and McClelland (1991), recur-rent backpropagation networks, as used in the bottomlevel of Clarion (in conjunction withQ-learning), canhandle sequences with complex statistical structures,given proper training procedures. Dienes (1992) reportedsimilar results, in which a simple network model out-performed other models in capturing sequence learningdata. The rule learning mechanism, as used in the toplevel of Clarion, clearly has trouble handling suchsequences. Therefore, in the circumstances in which ahigh level of statistical structure is involved in sequences,the bottom level prevails.
Note that there has been other work that demonstratedthe distinction and dissociation of the two types ofknowledge and proposed models based on that (e.g.Schacter, 1990; Shallice, 1988). However, some of theempirical work on which these models are based is con-cerned with abnormal subjects, most typically patientswith brain damages. For example, Schacter (1990) dis-cussed the following types of patients: amnesia (a selec-tive inability to remember recent experience and to learnnew information, typically due to lesions in the medialtemporal lobe), blindsight (the inability to make certainresponses in the absence of conscious perceptual aware-ness due to damages in the visual cortex), aphasia(impairment in processing syntactic or semantic informa-tion due to damages to a particular brain region), hemi-neglect (an impaired ability to attend to the sidecontralateral to the damaged hemisphere), and so on,all of which were characterised by dissociation of differ-ent types of information/knowledge somewhat similar tosituations discussed above. Schacter (1990) proposed amodel for accounting for the dissociation (see Section 5for details). Although the model was, I believe, on theright track, the support for it was not as strong as it could
have been, because in brain damaged patients, it waspossible that certain reconfiguration and reallocationmight have taken place (Shallice, 1988) and thus ren-dered the findings less applicable to normal human sub-jects. In this work, I only examine experimental findingsfrom normal human subjects and thus results obtainedmay be generalised to a wider range of settings.
3.2. Delayed Explication of Unconscious Processes
In the implicit learning literature, implicit performancetypically improves earlier than explicit knowledge thatcan be verbalised by the subject (Stanley et al., 1989).For example, in dynamic decision tasks, although perfor-mance quickly rises to a high level, subjects’ verbalknowledge improves far slower; the subjects cannot pro-vide usable verbal knowledge until near the end of theirtraining (Stanley et al., 1989). Bowers et al. (1990) alsoshowed delayed explication of implicit processes. Whensubjects were given patterns to complete, they showedimplicit recognition of what a proper completion mightbe even though they did not have explicit recognition of acorrect completion. The implicit recognition improvedover time and eventually, an explicit recognition wasachieved. In all of these cases, as suggested by Stanleyet al. (1989) and Seger (1994), we may hypothesise that,due to the fact that explicit knowledge lags behind butimproves along with implicit knowledge, explicit knowl-edge is in a way extracted from implicit knowledge.Cleeremans and McClelland (1991) also pointed outthis possibility in discussing their data and models.
Several developmental theorists have considered asimilar process in child development. Karmiloff-Smith(1986) suggested that developmental changes involverepresentational redescription. In young children, firstlow level implicit representations of stimuli wereformed and used, then, when more knowledge was accu-mulated and stable behaviours developed, through aredescription process, more abstract representationswere formed that transformed low-level representationsand made them more explicit and usable. Based on dataon perceptual analysis and categorization in infancy,Mandler (1992) proposed that relatively abstract‘‘image-schemas’’ were extracted from perceptual sti-muli, which coded several basic types of movements.On top of such image schemas, concepts were formedusing information therein. She suggested that it waslikely that an infant gradually formed ‘‘theories’’ ofhow his/her sensorimotor procedures work and therebygradually made such processes explicit and accessible.Finally, Keil (1989) suggested that conceptual represen-tations were composed of an associative component(with frequency and correlational information; Hasher& Zacks, 1979) and a ‘‘theory’’ component (with explicitknowledge; Murphy & Medin, 1985). Developmentally,there was a clear shift from associative to theory basedrepresentations in children. In data concerning learning
1323Learning, Action and Consciousness: A Hybrid Approach Toward Modelling Consciousness

concepts of both natural and nominal kinds, simplesimilarity-based or prototype representations dominatedat first, but gradually more explicit and focused theoriesdeveloped and became more prominent. Keil (1989)pointed out that it was unlikely that theories developedindependently, but rather they developed somehow fromassociative information that was already available. Thesefindings further testify to the ubiquity of an implicit-to-explicit transition (Clark & Karmiloff-Smith, 1993).
Clarion readily captures this kind of bottom-up pro-cess. The bottom level develops implicit, embodied skillson its own (Section 2, eqn (2)), while the top levelextracts explicit rules using algorithmRule-Extraction-Refinement(Section 2). Thus, the delayed bottom-uplearning naturally falls out of the model.5
3.3. Differences in Conscious/Unconscious Processes:Flexibility, Generalizability, and Robustness.
It has been shown that implicit learning produces lessflexible knowledge than explicit knowledge (Seger,1994; Berry & Broadbent, 1988; Stanley et al., 1989;Karmiloff-Smith, 1986). Seger (1994) argued that impli-cit learning results in knowledge that was more tied to thespecific stimulus modality of the learning environmentand less manipulable. Based on psycholinguistic data,Karmiloff-Smith (1986) observed that with the growthof explicit representations, more and more flexibility wasshown by subject children. Clarion can account for thehigher degree of flexibility of explicit, conscious knowl-edge relative to implicit, unconscious knowledge. Due tothe explicit (i.e. localist) representation used at the toplevel of Clarion (which stores explicit knowledge), avariety of explicit manipulations can be performed thatare not available to the bottom level. For example, back-ward and forward chaining reasoning, counterfactualreasoning, explicit hypothesis testing learning, and soon can be used individually or in combination. Thesecapacities lead to heightened flexibility in the top level.The bottom level employs only backpropagation net-works and thus cannot have the same flexibility.
As observed in many experiments, following explicitlearning, subjects are able to handle novel stimuli in asimilar way (or in other words, to generalise). In artificialgrammar learning, Reber (1967, 1976) found goodtransfer to strings using different letters but based onthe same grammar. Berry and Broadbent (1988) showedthat subjects trained on a dynamic decision task couldtransfer to another task with a similar cover story andidentical underlying relations. Generalisation has been
demonstrated in neural network models by e.g. Elman(1990) and many others. Elman (1990) reported goodgeneralisation of sequences by recurrent backpropaga-tion networks in grammar learning. Pollack (1991)found generalisation of such networks to arbitrarilylong sequences. As in human learning, generalizationin neural networks is based in part on similarity of oldand new sequences but also in part on certain structuresexhibited by the sequences. Thus, the bottom level ofClarion, which incorporates a backpropagation net-work, has the capability to capture the generalizationexhibited in human implicit learning. (Explicit processes,as in the top level of Clarion, can also generalise, albeitin a different way as discussed in Sun et al., 1995.)
It has also been observed that implicit processes aremore robust than explicit processes (Reber, 1989) in theface of internal disorder and malfunctioning. For exam-ple, Hasher and Zacks (1979) found that encoding offrequency information (an implicit process) was cor-rectly performed by clinically depressed patients, eventhough they could not perform explicit tasks consciously.Warrington and Weiskrantz (1982) found that amnesicswere more successful in performing implicit rather thanexplicit memory tasks. This effect is consistent with thedual representation framework of Clarion: while the toplevel employs localist representation and is thus morevulnerable to malfunctioning, the bottom level utilisesa distributed representation that is more resistant todamages and faults, as demonstrated amply in neuralnetwork models.
3.4. Unconscious Initiation of Action
Existing evidence indicates that unconscious processesoften (if not always) initiate actions in skilled perfor-mance in advance of conscious awareness. Libet (1985)reported that electrophysiological ‘‘readiness poten-tials’’ (RPs) always precede conscious initiation of anact that is fully endogenous and voluntary. After a con-scious intention to act appears, whether the action actu-ally takes place or not can still be decided consciouslyby a subject within a time period of somewhere between100 and 200 ms. As suggested by Libet (1985), the roleof the conscious mind is not to initiate a specific courseof action, but to control and influence (implicitly selectedand initiated) actions.
This view is consistent with that of Willingham et al.(1989) that the role of explicit processes is to influencethe implicit process but not to directly take control ofskill learning or performance. Willingham et al. (1989)posited that the effects from the two processes are‘‘superimposed’’ on each other, so that each type com-plement each other. Kelley and Jacoby (1993) alsoinsisted that an important function of the explicit mindis to oppose, or counterbalance, the influence of theimplicit mind.
The aforementioned view is also consistent with
1324 R. Sun
5 There is also evidence that explicit knowledge may developindependently. Willingham et al. (1989) reported such data. Thesedata rule out the possibility that one type of knowledge isalwayspreceded by the other type, at least under their experimental conditions.To account for this phenomenon, in Clarion, explicit hypothesistesting can be employed in the top level for learning rules, inde-pendently of the bottom level, as mentioned before.

voluminous data on the ever-present role of unconsciousprocesses in all kinds of tasks: lexical priming, semanticprocessing, visual attention, unconscious perception andso on (as discussed in Velmans, 1991; Marcel, 1983).Velmans (1991) summarized evidence for the existenceof implicit (preconscious) analysis of input stimuli,implicit processing of semantic content of word pairsin ‘‘shadowing’’ experiments, and implicit processingof bilingual messages in similar experimental settings.Most of these findings support the possibility that uncon-scious processes start before conscious processes takehold.
Clarion can readily accommodate this phenomenon,in that the bottom level, which captures unconsciousprocesses, can work independently and initiate proces-sing without the involvement of the top level. However,after the initiation of action and, consequently, the acti-vation of the relevant nodes, the corresponding represen-tations at the top level can then be activated by thebottom-up information flow (see Section 2). The acti-vated explicit representations and their associated pro-cesses at the top level will in turn influence the implicitprocessing at the bottom level, in way of modifying andrectifying its outcomes and decisions (through the com-bination mechanism; see Section 2). Thus the implicitprocesses, which directly control actions in skilled per-formance, incorporate the results of explicit processesfrom the top level.
3.5. Synergy Between the Conscious/UnconsciousProcesses
Why are there two separate (although interacting) sys-tems, one conscious and the other unconscious? Based onearlier discussions, we may hypothesize that each systemserves a unique function and the two are complementaryto each other; that is, there may be a synergy between theconscious and the unconscious. Such a synergy mayshow up by speeding up learning, improving learnedperformance, and facilitating transfer of learned skills.
In terms of speeding up learning, Stanley et al. (1989)reported that in a dynamic decision task (the sugar fac-tory task), subjects’ learning improved if they were askedto generate verbal instructions for other subjects alongthe way during learning. Willingham et al. (1989) foundthat those subjects who acquired full explicit knowledgeappeared to learn faster.
In terms of learned performance, Willingham et al.(1989) found that subjects who verbalized while per-forming were able to attain a higher level of perfor-mance, because the requirement that they verbalisedtheir knowledge prompted the formation and utilizationof explicit knowledge. In high-level skill acquisition,Gick and Holyoak (1980) found that good problemsolvers could better state rules that described theiractions in problem solving. This phenomenon may berelated to the self-explanation effect (Chi et al., 1989):
subjects who explained the examples in textbooks morecompletely did better in solving new problems. In allthese cases, it may well be the explication and the useof explicit knowledge that helped the performance.
In terms of facilitating transfer of skills, Willinghamet al. (1989) obtained some suggestive evidence thatexplicit declarative knowledge facilitated transfer ofskilled performance. It was reported that: (1) subjectswho acquired explicit knowledge in a training taskstended to have faster response times in a transfer task;(2) these subjects were also more likely to acquire expli-cit knowledge in the transfer tasks. In high-leveldomains, Ahlum-Heath and DiVesta (1986) also foundthat the subjects who were required to verbalize whilesolving the Tower of Hanoi problem performed better ona transfer task after training.
Sun et al. (1995) reported some simulation experi-ments that demonstrated Clarion was able to exhibitanalogous synergy effects in learning, performance,and transfer through the interaction of the two levels.The simulation experiments were conducted in twodomains: maze running and navigation through mine-fields. The details of the experiments and completedata can be found in Sun et al. (1995). Briefly, in themaze task, a subject/agent was to find a unknowntarget in the maze and had only rudimentary sensoryinputs regarding its immediate left, front and right side,indicating whether there was a wall, an opening, or thegoal; the agent could move forward, turn to the left, orturn to the right, until it found the target (see Figure 5). Interms of speeding up learning, the differences in learningspeeds betweenQ-learning (which in Clarion capturesunconscious learning at the bottom level) and Clarion(which includes both unconscious and conscious learn-ing) were very significant. In terms of trained perfor-mance (measured by the average number of stepsneeded to reach the target in one episode), Clarion out-performed pureQ-learning by large margins again. Wealso compared the trained performance of the bottomlevel of Clarion alone (after the training of the entiresystem together, includingQ-learning) with the perfor-mance of pureQ-learning, and discovered that the expli-cation of skills not only improved the performance of thewhole system, but it also improved theQ-learning partwhen included as part of Clarion. We also assessed theperformance of trained models in a new and larger maze.Clarion transferred much better thanQ-learning alone
1325Learning, Action and Consciousness: A Hybrid Approach Toward Modelling Consciousness
FIGURE 5. The Maze. The starting position is marked by ‘‘S’’ inwhich the agent faces upward to the upper wall. The goal ismarked by ‘‘G’’.

(in terms of number of steps to reach the goal in oneepisode). Furthermore, by comparing the correspondingperformance of the top level, the bottom level and thewhole Clarion model, it was clear that often learnedrules alone (the top level) performed better in transferthan the bottom level, as well as than the whole Clarionmodel, which showed that explicit knowledge facilitatedtransfer.
In the simulated navigation task shown in Figure 6, thesubject/agent had to navigate an underwater vesselthrough a minefield to reach a target location. Theagent received only local information from a number ofinstruments, as shown in Figure 7. Using only this infor-mation, the agent decided (1) how to turn and (2) howfast to move, and within an allotted time period, couldeither (a) reach a target (which is a success), (b) hit amine (a failure), or (c) run out of fuel (a failure). In termsof learning speeds, the superiority of Clarion over Q-learning was statistically significant. To access transfer,after training models on 10-mine minefields, we assessedperformance of these models in new minefields that con-tained 30 mines. Clarion outperformedQ-learning. Thedifference between the best transfer of Q-learning andthe best transfer of Clarion was statistically significant.In sum, Clarion is able to replicate similar findings inhuman conscious/unconscious learning.
4. THEORETICAL ISSUES
Some theoretical issues concerning consciousness will bediscussed below in relation to the Clarion model.
4.1. Casual Efficacy of Consciousness
Is consciousness epiphenomenal as some have claimed?To see the casual role of consciousness, it is useful toexamine the available cognitive. The following has beenobserved from patients who suffered the loss of some ofthe capacities of their consciousness due to braindamages (Marcel, 1988):
• They lost the ability to act on parts of the environmentthat were not accessible to their explicit/consciousmind (as in the case of blindsight patients who couldnot grasp objects in the blind field).
• They lost the ability to form an integrated self-concept(as in the case of amnesiac patients).
• They lost the ability to learn new complex tasks thatrequired explicit, verbal instructions.
• They lost the ability to form explicit plans of actionsbefore acting on them.
These effects indicate certain casual efficacy of con-sciousness, and are consistent with Clarion. In themodel, these effects follow from the loss of the mechan-isms in the top level of Clarion, such as backwardchaining reasoning (planning), verbal instruction taking,and the use of explicit knowledge (regarding self andobjects in the world).
Through contrasting ‘‘aware’’ vs ‘‘unaware’’ condi-tions on their experiments with human subjects, Kelley &Jacoby (1993) showed that conscious awareness per sehad a distinct effect in subsequent behaviour. The twodifferent conditions produced two different causal attri-butions in the subjects: one to true causes (in the‘‘aware’’ condition) and the other to spurious causes(in the ‘‘unaware’’ condition; see Nisbett & Wilson,1977 for causal attribution as error-prone, post hoc inter-pretation); consequently, different causal attributionsled to different actions on the part of the subjectsmaking the attributions. This kind of causal role in con-sciousness is consistent with that of the top level ofClarion, which shares the responsibility of controllingactions using explicit knowledge. A second mechanismon top of unconscious processes can offer counter-balance, and thus can have clear survival values to theagent possessing it.
4.2. Human Intuition
While the top level of Clarion captures conscious pro-cesses, the bottom level may captureintuition to someextent (as a form of skill): This level has the character-istics of being implicit, inaccessible, and holistic, whichare also characteristics of human intuition (James, 1890;Dreyfus & Dreyfus, 1987). According to Smolensky
1326 R. Sun
FIGURE 6. Navigating through mines.
FIGURE 7. The navigation input. The display at the upper leftcorner is the fuel gauge; the vertical one it the upper right corneris the range gauge; the round one in the middle is the bearinggauge; the 7 sonar gauges are at the bottom.

(1988), as mentioned before, intuition and skill are notexpressible in linguistic forms and constitute a differentkind of capacity, reflecting ‘‘subsymbolic’’ processing.Dreyfus & Dreyfus (1987) suggested that intuition ismanifested in the fluent, holistic and situation sensitiveway of dealing with the world, unique to humans and notcaptured by conventional symbolic computation. Theseidentified characteristics can be found in the bottom levelof Clarion to some extent.
It was hypothesised by Reber (1989) that human intui-tion may be the direct result of implicit, unconsciouslearning: Through the gradual process of implicit learn-ing, ‘‘tacit’’ (implicit) representations emerge that cap-ture environmental regularities and are used in directcoping with the world (without the involvement of anyintrospective process). Intuition is the end product of thisprocess of unconscious and bottom-up learning (Reber,1989). Bowers et al. (1990) also suggested that intuitionis the outcome of an unconscious, implicit process(which later becomes explicit due to the emergence ofa coherent pattern of activation) in the context of discov-ery. Clarion indeed uses implicit learning to developtacit (implicit) representations in the bottom level andthus acquires intuition in the sense identified above.
4.3. Two Types of Consciousness
We can also examine the two levels of Clarion usingthe perspective of phenomenological philosophy.Heidegger (1927) emphasised a basic mode of existence,that is, the immediate comportment with the world.Normally, when going about its daily business, anagent is notthematicallyconscious of routine activities.Everyday routine activities are mostly made up of non-deliberate ‘‘primordial’’ coping. For example, in normalperception, we are usually not having thematic experi-ence of the world (Dreyfus, 1992). An agent’s ‘‘opennessonto the world’’ is fundamental and makes possible thesecondary experience of deliberate looking or trying tosee (Dreyfus, 1992). Comportment is prior to any(explicit) belief, (explicit) knowledge, or (explicit)representation; it is a direct connection between anagent and its existential context. It is comportment withthe world that is in fact a more fundamental kind ofconsciousness, according to Heidegger, and in thisview, consciousness is non-representational (i.e. withoutexplicit representation).
This is in contrast with the notion of ‘‘thematic’’ con-sciousness, which involves a focused, meditated aware-ness of the object of consciousness (akin to the commonnotion of consciousness). Thematic consciousness isrepresentational because it treats awareness itself as anobject (see also Clark & Karmiloff-Smith, 1993). As hasbeen argued extensively by Heidegger (1927), thematicconsciousness can indeed arise, but it presupposes anonthematic, nondeliberate, direct, and on-going wayof dealing with the world (i.e. comportment); for direct
comportment with the world is anecessarymeansfor coping with a complex world that exhibits complexregularities. Explicit representations are derived fromdirect comportment with the world. Derived representa-tion and thematic consciousness come into play, e.g.during breakdown in which established routines getdisrupted and thus alternative ways are necessary (seealso Sun, 1994, Chapter 8). In the light of the above,the distinction between the two levels in Clarion canbe corresponded roughly to the distinction between com-portment and thematic consciousness. Specifically, thebottom level captures the implicit and routine activitiesand embodies ‘‘comportment’’. This is because thebottom level embodies skills resulting from and usedfor directly coping with the world and involves distrib-uted representation, which is hence unable to presentexplicit traces of its processes (Smolensky, 1988). How-ever, some events (e.g. certain implicit processes orexplicit verbal inputs; Velmans, 1991) may lead to acti-vation of corresponding explicit representations and pro-cesses at the top level and therefore lead to (thematic)conscious awareness. This is because at the top levelexplicit/localist representation is used, which makes itpossible to articulate the content that is present andtrace the processes as they are occurring (Sun, 1994,Chapter 2). Furthermore, due to the fact that the toplevel of Clarion is derived from, mediated by, andgrounded in the bottom level, which has direct inter-actions with the external world and develops bottom-upfrom such interactions, (thematic) consciousness isclearly grounded, in this model as in humans, in theinteraction between the agent and the external world.
4.4. Qualia
But what about qualia? Qualia refer to the phenomenalquality of conscious experience. Block (1994) distin-guishes access consciousness and phenomenal con-sciousness, whereby access consciousness refers to theutilization of the content of consciousness while phe-nomenal consciousness refers to the subjective feel ofconscious experience. Although it has been a major dif-ficulty to understand phenomenal consciousness/qualia(‘‘the hard problem’’; Chalmers, 1992), some specula-tions may be made here in relation to Clarion: qualia(phenomenal consciousness) may be accounted for bythe totality of a multi-modal (see the next section regard-ing modalities), multi-level organization and its totalcollective states, which are of extremely high complexityinvolving external perception (of many modalities),internal perception, action decision making, explicitconcepts, etc. The complexity of this organisation mayexplain the difficulty (or impossibility) of describingphenomenal qualities (qualia) of consciousness, whichis the most striking feature of phenomenal consciousness(as argued by Nagel, 1974). In this approach, a particularkind of phenomenal quality may be accounted for by a
1327Learning, Action and Consciousness: A Hybrid Approach Toward Modelling Consciousness

particular region of total-state space (involving thetotality of all the aforementioned aspects) or the manifoldas termed by Van Gulick (1993), which gives rise to thesense of what something is like (Nagel, 1974). Clearly,such regions depend on particularfunctional organi-zations of modules and levels (Chalmers, 1992) thatsupport such a space of total-states. Qualia are thus(partially) the result of functional organisations (archi-tectures) of cognitive apparatuses. In Clarion, qualiaare (partially) the result of the two-level organization,on top of all the detailed, intricate structures involvedin various fine-grained modalities (the detail of whichis not covered here). Equally important, such regionsarise from the interaction of the agent and the world, asthe consciousness is grounded in learning, action, and theworld in which the agent exists. Thus, phenomenal con-sciousness is in general derived from a complex integra-tive organisation of cognitive apparatuses that develop asa result of the interaction between the agent and theworld. This is true both in an ontological and in an onto-genetic sense. Note that, though access consciousnesshas a clear causal role in behaviour (as discussed earlier),phenomenal consciousness is less clear in this regard(Block, 1994).
4.5. Functionalism
An important question regarding consciousness iswhether there is a physical basis for consciousnessand what it is. It is pretty much agreed upon amongcontemporary philosophers that there is indeed aphysical basis but they disagree on what constitutesthat physical basis. In the Clarion model, we basicallystipulate that the physical basis of consciousness is madeup of the detailed architecture, the fine-grained func-tional organisation (e.g. the two-level modular frame-work with detailed sensory modality structures at avery fine level), of one’s cognitive apparatus, in inter-action with one’s world. The distance between the physi-cal and the phenomenological/psychological is so greatthat intermediate levels (functional levels) are necessaryfor studying cognition, and this is especially true in thecase of studying consciousness. The present approachis a form of fine grained functionalism (Chalmers,1992), which states that consciousness is invariantacross systems with the same functional organization ata sufficiently fine-grained level (i.e. the principle oforganizational invariance), as argued for elegantly byChalmers (1992). It is a weak form of functionalism, inthat it is not just causal connections between functionalstates that are important, but also the level at which weidentify functional states. This approach is also inter-actionalism in the sense that the interaction of internaland external systems (the agent and its world), on thebasis of internal (developing) fine-grained functionalorganizations, is crucial in giving rise to consciousexperience (Heidegger, 1927).
5. COMPARISONS
The following comparisons will further explicate andaccentuate the Clarion model in accounting for issuesof consciousness. Clarion captures important featuresof existing models. The comparisons reveal that Clar-ion has the potential to account for the integration ofsensory modalities, global consistency, and the unity ofconsciousness.
First we can compare Clarion with the model ofBaars (1988) (see Figure 8), in which a large numberof specialist processors perform unconscious processingand a global workspace coordinates their activitiesthrough global broadcasting to achieve consistency andthus conscious experience. The model bears some resem-blance to Clarion, in that unconscious specialist proces-sors in that model can be roughly equated to modulesin the bottom level of Clarion, and the global workplacemay be roughly captured by the top level, which‘‘synthesizes’’ the bottom level modules and is essentialin conscious processing. One difference is that Clariondoes not emphasise as much internal consistency(Marcel, 1983): it is believed to be limited as a phenom-enon in consciousness and may have only limited roles inthe emergence of consciousness. Global broadcastingin Baars’ model (Baars, 1988; Revonsuo, 1993) can beviewed as the integration of the two levels of representa-tions (with the bottom-level representations dispersedwithin multiple modules) in Clarion, which does pro-duce somewhat consistent outcomes (which lead to theunity of consciousness; Baars, 1988; Marcel, 1983; moreon this later).
We can also compare Clarion with the model ofSchacter (1990) (see Figure 9), which is based onneurophysiological findings of the dissociation of differ-ent types of knowledge in brain damaged patients asmentioned earlier. It is similar to Clarion, in that itincludes a number of ‘‘knowledge modules’’ that per-form specialized and unconscious processing (analogousto bottom-level modules in Clarion) and send their
1328 R. Sun
FIGURE 8. Baars’ model of consciousness.

outcomes to a ‘‘conscious awareness system’’ (analo-gous to the top level in Clarion), which gives rise toconscious awareness. Schacter’s explanation of somedisorders (e.g. the loss of short-term memory or explicitlearning abilities, as mentioned earlier) is that certainbrain damages result in the disconnection of some ofthe modules from the conscious awareness system,which leads to their inaccessibility to consciousness.An alternative explanation offered by Clarion is thatdisorders may not be due to disconnected modules, butthe loss of some explicit learning and performancemechanisms at the top level (resulting from braindamages etc.).
Finally, we can examine Damasio’s neuroanatomi-cally motivated model (Damasio, 1994; Revonsuo,1993). The model (see Figure 10) hypothesised theexistence of many ‘‘sensory convergence zones’’ thatintegrated information from individual sensory modal-ities through forward and backward synaptic connectionsand the resulting reverberations of activations, withoutthe need for a central location for information storageand comparisons; it also hypothesised the global‘‘multi-modal convergence zone’’, which integratedinformation across modalities also through reverberation
via recurrent connections. In Clarion, different sensoryconvergence zones may be roughly captured by bottom-level modules, each of which takes care of sensory inputsof one modality (at a properly fine level), and the role ofthe global multi-modal convergence zone (similar to the‘‘global workspace’’ in a way) may be played by the toplevel of Clarion, which has the ultimate responsibilityfor integrating information (and serves as ‘‘consciousawareness system’’). The widely recognised role ofreverberation (Damasio, 1994; Taylor, 1994) may becaptured in Clarion through using recurrent connec-tions within modules at the bottom level and throughmultiple top-down and bottom-up information flowsacross the two levels, which lead to the unity of con-sciousness that is the synthesis and integration of allthe information present (Marcel, 1983; Baars, 1988).
6. CONCLUDING REMARKS
This paper presented a hybrid neural network model forlearning that incorporated the distinction of declarativeand procedural knowledge, and succeeded to some extentin accounting for the distinction of the conscious andthe unconscious (or the explicit and the implicit). Morespecifically, the Clarion model applied neural networkand machine learning techniques to explain complexhuman learning and consciousness in normal humansubjects. It accounted for phenomena in psychologicalliterature on learning and development in terms of thetwo levels in the model and their associated mechanisms.The model readily accommodated important features ofexisting models of consciousness. We also had some-thing to say about theoretical issues such as qualia andintuition on the basis of the model, which helped clarifycomplex issues in a tangible way. The key issue forfuture research is scaling up the model by incorporatinga variety of sensory information and dealing with com-plex environments, in order to allow rudimentary formsof consciousness to emerge, because, I believe, complex-ity is a necessary condition for consciousness.
1329Learning, Action and Consciousness: A Hybrid Approach Toward Modelling Consciousness
FIGURE 9. Schacter’s model of consciousness.
FIGURE 10. Damasio’s model of consciousness.

REFERENCES
Ahlum-Heath, M., & DiVesta, F. (1986). The effect of conscious con-trolled verbalization of a cognitive strategy on transfer in problemsolving.Memory and Cognition, 14, 281–285.
Anderson, J. R. (1983).The architecture of cognition. Cambridge, MA:Harvard University Press.
Baars, B. (1988).A cognitive theory of consciousness. CambridgeUniversity Press.
Berry, D., & Broadbent, D. (1984). On the relationship between taskperformance and associated verbalizable knowledge.QuarterlyJournal of Experimental Psychology, 36A,209–231.
Block, N. (1994). On a confusion about a function of consciousness.Brain and Behavioral Sciences.
Bowers, K., Regehr, G., Balthazard, C., & Parker, K. (1990). Intuitionin the context of discovery.Cognitive Psychology, 22, 72–110.
Bruner, J., Goodnow, J., & Austin, J. (1956).A study of thinking. NewYork: Wiley.
Chalmers, D. (1993).Towards a theory of consciousness. Ph.D Thesis,Indiana University.
Chi, M., Bassok, M., Lewis, M., Reimann, P., & Glaser, P. (1989). Self-explanation: how students study and use examples in learning tosolve problems.Cognitive Science, 13, 145–182.
Clark, A., & Karmiloff-Smith, A. (1993). The cognizer’s innards: apsychological and philosophical perspective on the developmentof thought.Mind and Language, 8(4), 487–519.
Cleeremans, A., & McClelland, J. (1991). Learning the structure ofevent sequences.Journal of Experimental Psychology: General,120,235–253.
Cohen, A., Ivry, R., & Keele, S. (1990). Attention and structure insequence learning.Journal of Experimental Psychology: Learning,Memory, and Cognition, 16, 17–30.
Cosmides, L., & Tooby, J. (1994). Beyond intuition and instinctblindness: toward an evolutionarily rigorous cognitive science.Cognition, 50, 41–77.
Damasio, A. (1994).Decartes’ error. New York: Grosset/Putnam.Dennett, D., & Kinsbourne, M. (1992). Time and the observer.
Behavioral and Brain Science, 15, 183–200.Dienes, Z. (1992). Connectionist and memory-array models of artificial
grammar learning.Cognitive Science, 16, 41–79.Dreyfus, H., & Dreyfus, S. (1987).Mind over machine: the power of
human in tuition. New York, NY: The Free Press.Dreyfus, H. (1992).Being-in-the-world. Cambridge, MA: MIT Press.Edelman, G. (1989).The remembered present: a biological theory of
consciousness. New York: Basic Book.Elman, J. (1990). Finding structures in time.Cognitive Science, 14,
179–211.Fodor, J. (1983).The modularity of mind. Cambridge, MA: MIT Press.Jackendoff, R. (1987).Consciousness and the computational mind. MIT
Press.Gick, M., & Holyoak, K. (1980). Analogical problem solving.Cogni-
tive Psychology, 12, 306–355.Hasher, J., & Zacks, J. (1979). Automatic and effortful processes in
memory. Journal of Experimental Psychology: General, 108,356–358.
Heidegger, M. (1927).Being and time(English translation, 1962). NewYork: Harper and Row.
James, W. (1890).The principles of psychology. New York: Dover.Karmiloff-Smith, A. (1986). From meta-processes to conscious access:
evidence from children’s metalinguistic and repair data.Cognition,23, 95–147.
Keil, F. (1989). Concepts, kinds, and cognitive development.Cambridge, MA: MIT Press.
Kelley, C., & Jacoby, L. (1993). The construction of subjective experi-ence: memory attribution. In M. Davies & G. Humphries (Eds.),Consciousness. Oxford, UK: Blackwell.
LeDoux, J. (1992). Brain mechanisms of emotion and emotional learn-ing. In Current opinion in neurobiology, 2(2) (pp. 191–197).
Lewicki, P., Hill, T., & Czyzewska, M. (1992). Nonconscious acquisi-tion of information.American Psychologist, 47, 796–801.
Libet, B. (1985). Unconscious cerebral initiative and the role ofconscious will in voluntary action.Behavioral and BrainSciences, 8, 529–566.
Lin, L. (1992). Self-improving reactive agents based on reinforcementlearning, planning, and teaching.Machine Learning, 8, 293–321.
Mandler, J. (1992). How to build a baby.Psychology Review, 99(4),587–604.
Marcel, A. (1983). Conscious and unconscious perception: an approachto the relations between phenomenal experience and perceptualprocesses.Cognitive Psychology, 15, 238–300.
Marcel, A. (1988). Phenomenal experience and functionalism. InA. Marcel & E. Bisiach (Eds.),Consciousness in contemporaryscience. Oxford, UK: Oxford University Press.
Medin, D., Wattenmaker, W., & Michalski, R. (1987). Constraints andpreferences in inductive learning: an experimental study of humanand machine performance.Cognitive Science, 11, 299–339.
Michalski, R. (1983). A theory and methodology of inductive learning.Artificial Intelligence, 20, 111–161.
Mitchell, T.(1982). Generalization as search.Artificial Intelligence, 18,203–226.
Murphy, G., & Medin, D. (1985). The role of theories in conceptualcoherence.Psychological Review, 92, 289–316.
Nisbett, R., & Wilson, T. (1977). Lelling more than we can know:verbal reports on mental processes.Psychological Review, 84(3).
Nagel, T. (1974). What is it like to be a bat?Philosophical Review, 4,435–450.
Nosofsky, R., Palmeri, T., & McKinley, S. (1994). Rule-plus-exceptionmodel of classification learning.Psychological Review, 101(1),53–79.
Penrose, R. (1994).Shadows of the mind. Oxford, UK: OxfordUniversity Press.
Posner, M., & Snyder, C. (1975). Facilitation and inhibition. In P. Rabbit& S. Dornick (Eds.),Attention and performance. Academic Press.
Pollack, J. (1991). The induction of dynamic recognizers.MachineLearning, 7(2/3), 227–252.
Reber, A. (1967). Implicit learning of artificial grammars.Journal ofVerbal Learning and Verbal Behavior, 6, 855–863.
Reber, A. (1976). Implicit learning of synthetic languages: the role ofinstructional set.Journal of Experimental Psychology. HumanLearning and Memory, 2, 88–94.
Reber, A. (1989). Implicit learning and tacit knowledge.Journal ofExperimental psychology: General, 118(3), 219–235.
Revonsuo, A. (1993). Cognitive models of consciousness. In M. Kamp-pinen (Ed.),Consciousness, cognitive schemata and relativism(pp.27–130). Dordrecht: Kluwer.
Schacter, D. (1990). Toward a cognitive neuropsychology of aware-ness: implicit knowledge and anosagnosia.Journal of Clinicaland Experimental Neuropsychology, 12(1), 155–178.
Seger, C. (1994). Implicit learning, Psychological Bulletin,155(2),163–196.
Shallice, T. (1988).From neuropsychology to mental structure.Cambridge University Press.
Shiffrin, R., & Schneider, W. (1977). Controlled and Automatic humaninformation processing II.Psychological Review, 84, 127–190.
Smolensky, P. (1988). On the proper treatment of connectionism.Behavioral and Brain Sciences, 11(1), 1–74.
Stadler, M. (1992). Statistical structure and implicit serial learning.Journal of Experimental Psychology: Learning, Memory, andCognition, 18, 318–327.
Stanley, W., Mathews, R., Buss, R., & Kotler-Cope, S. (1989). Insightwithout awareness: on the interaction of verbalization, instructionand practice in a simulated process control task.Quarterly Journalof Experimental Psychology, 41A(3), 553–577.
Sun, R. (1992a). A connectionist model for commonsense reasoningincorporating rules and similarities.Knowledge Acquisition, 4,293–321.
1330 R. Sun

Sun, R. (1992b). On variable binding in connectionist networks.Connection Science, 4(2), 93–124.
Sun, R. (1994). Integrating rules and connectionism for robustcommonsense reasoning. New York, NY: John Wiley and Sons.
Sun, R. (1995). Robust reasoning: integrating rule-based and similarity-based reasoning.Artificial Intelligence, 75(2), 241–296.
Sun, R., & Bookman, L. (Eds.). (1994).Computational architecturesintegrating neural and symbolic processes. Norwell, MA: KluwerAcademic Publishers.
Sun, R., Peterson, T., & Merrill, E. (1995).Hybrid architecture andsituated learning(Technical Report TR-CS-96-0019). University ofAlabama. submitted to a journal.
Sun, R., Peterson, T., & Merrill, E. (1996a).Bottom-up skill learning(Technical Report TR-CS-96-0021). University of Alabama. sub-mitted to a journal.
Sun, R., Peterson, T., & Merrill, E. (1996b). Bottom-up skill learningin reactive sequential decision tasks.Proc. of 18th CognitiveScience Society Conference. Hillsdale, NJ: Lauwrence ErlbaumAssociates.
Sutton, R. (1990). Integrated architectures for learning, planning, andreacting based on approximating dynamic programming.Proc. of
7th International Conference on Machine learning. San Mateo, CA:Morgan Kaufmann.
Taylor, J. (1994). Goal, drives and consciousness.Neural Networks,7(6/7), 1181–1190.
Towel, G., & Shavlik, J. (1993). Extracting Refined Rules from Knowl-edge-Based Neural Networks.Machine Learning. 7.
Van Gulick, R. (1993). Understanding the phenominal mind. InM. Davies & G. Humphries (Eds.),Consciousness. Oxford, UK:Blackwell.
Velmans, M. (1991). Is human information processing conscious?Behavioral and Brain Sciences, 14, 651–726.
Waltz, D. (1991). How to build a robot. In S. Wilson (Ed.),Proc. onConf. on Simulation of Adaptive Behaviors. Cambridge, MA: MITPress.
Warrington, E., & Weiskrantz, L. (1982). Amnesia: a disconectionsyndrome?.Neuropsychologica, 20, 233–248.
Watkins, C. (1989).Learning with delayed rewards. Ph.D Thesis.Cambridge, UK: Cambridge University.
Willingham, D., Nissen, M., & Bullemer, P. (1989). On the develop-ment of procedural knowledge.Journal of Experimental Psychol-ogy: Learning, Memory, and Cognition, 15, 1047–1060
1331Learning, Action and Consciousness: A Hybrid Approach Toward Modelling Consciousness

1997 SPECIAL ISSUE
Consciousness in a Self-Learning, Memory-Controlled,Compound Machine
Robert Alan Brown
8 Foster Street, Mattapoisett, Massachusetts
(Received6 July 1996;accepted15 April 1997)
Abstract—A memory-controlled, sensor/actuator machine senses conditions in its environment at given moments, andattempts to produce an action based upon its memory. However, a sensor/actuator machine will stop producing newbehavior if its environment is removed. A sensor/sensor unit can be added to the sensor/actuator machine, forming acompound machine. The sensor/sensor unit produces a stream of internally created sensed conditions, which can replacethe sensed conditions from the environment. This illusion of an environment is similar to consciousness. In addition,actuator/sensor and actuator/actuator units can be added to this compound machine to further enhance its ability tofunction without an environment. Predetermined and empirical memory cells can be distributed throughout the controlunits of this compound machine to provide instinctive and learned behavior. The internal and exterior behavior of thiscompound machine can be modified greatly by changing the cycle start and ramp signals that activate these differentkinds of memory cells. These signals are similar in form to brain waves.q 1997 Elsevier Science Ltd.
Keywords—Memory-controlled machines, Conditional memory cells, Logarithmic changes in memory, Trinaryencoders and decoders, Interrogation signals, Brain waves, Anesthesia, Coma.
1. MEMORY-CONTROLLED, SENSOR/ACTUATOR MACHINE
The diagram in Figure 1 can represent a typical machine.A memory-controlled, sensor/actuator machine sensesconditions in its environment at given moments, andconverts these sensed conditions into memory addresses.It then attempts to produce an action based upon thecontent of these memory addresses, completing a transi-tion cycle (Ashby, 1960).
The behavior of a memory-controlled, sensor/actuatormachine can be predetermined (programmed or setbeforehand), or it can be established over time by anempirical (self-learning) process. The output of thecontroller of a memory-controlled machine also canbe absolute (determined by individual memory cells),or conditional (determined by the relative values ofmany memory cells).
1.1. Absolute-Predetermined, Memory-ControlledMachine
An absolute-predetermined, memory-controlled machinecan be programmed to produce a specific action for eachspecific sensed condition. This is done by placing thetoggle switch in each of its absolute-predetermined
(AP) memory cells in a ‘‘1’’ (conducting) or ‘‘0’’(non-conducting) position, as shown in the scalar matrixin Figure 2.
At the beginning of a transition cycle, the value ofthe sensor variable (A) is identified and held by a latchedconnect. This energizes the input disconnect, whichprohibits any other value of the sensor variable (A)from being identified in that transition cycle. The voltagefrom the output enable appears at the output terminalof the AP cell with a closed toggle switch and the ener-gized latched connect. This energizes an output connectbelonging to the actuator variable (Z). The actuator delaytimer (td) in the cycle timing circuit delays the movementof the actuator to the position of the energized outputconnect, and the cycle end timer (te) terminates thetransition cycle, releasing the latches and timers. Abrake holds the actuator in a fixed position betweentransition cycles.
1.2. Conditional-Predetermined, Memory-ControlledMachine
Complex behavior may be difficult to program into anabsolute memory-controlled machine. A cumulativeprogramming process can be achieved by changing thestored memory values logarithmically between zero and
Neural Networks, Vol. 10, No. 7, pp. 1333–1343, 1997q 1997 Elsevier Science Ltd. All rights reserved
Printed in Great Britain0893–6080/97 $17.00+.00
PII: S0893-6080(97)00057-9
Pergamon
1333

1 in steps. This is done by means of the logarithmicsubtraction mechanism (LSM) (Brown, 1994) in eachconditional-predetermined (CP) memory cell in thecontroller of the conditional-predetermined, memory-controlled machine shown in Figure 3.
The CP memory cells are interrogated by the rampsignal (R). The ramp signal (R) causes a CP cell tobecome conductive when the ramp signal reaches thesensitivity value of that CP cell. The output fromthe matrix is terminated by the output disconnect whenan output latch is energized. This prohibits any otherless sensitive CP cells from selecting another value ofthe actuator variable (Z).
1.3. Logarithmic Subtraction Mechanism
The logarithmic response curves in Figure 4 show howthe sensitivity of a single conditional-predetermined
(CP) memory cell approaches its maximum or minimumvalue asymptotically.
An equal input of 1’s and 0’s leaves the sensitivity ofa CP cell near the mid-position after a sufficient numberof programming steps.
2. SELF-LEARNING, SENSOR/ACTUATORMACHINE
A self-learning, memory-controlled machine measuresthe action that is actually carried out in each transitioncycle. It then adjusts its memory to increase the likeli-hood of selecting the actual action when the sameconditions reoccur (Brown, 1994), as shown in Figure 5.
The behavior of a self-learning machine will changeuntil it is no longer interfered with by outsideinfluences. Thus, the behavior of a self-learning,memory-controlled machine will come to reflect theintentions of its operators, and the constraints of itsenvironment.
2.1. Absolute-Empirical, Memory-ControlledMachine
A circuit can be added to the absolute-predeterminedcontroller shown in Figure 2 that measures the actualaction that takes place in a given transition cycle. Thisactuator feedback circuit can be used to change the stateof absolute-empirical (AE) memory cells, forming theabsolute-empirical memory controlled machine shownin Figure 6.
The memory state of each absolute-empirical (AE)memory cell is set by changing its value to 0 when itselects an output, and changing its state to 1 if theoutput its selects actually takes place in a given transi-tion cycle.
1334 R. A. Brown
FIGURE 1. A memory-controlled, sensor/actuator machineproduces an ongoing line of behavior when each sensedcondition produces a new action, and each action creates anew sensed condition through the environment in eachtransition cycle.
FIGURE 2. The binary memory matrix of an absolute-predetermined, memory-controlled machine is similar to a computer memory matrix.

2.2. Absolute-Empirical Algorithm
The operation of the absolute-empirical, memory-controlled machine in Figure 6 is shown in theabsolute-empirical algorithm in Figure 7.
An absolute-empirical, memory-controlled machineloses all traces of its previous experience when itencounters new learning.
2.3. Conditional-Empirical, Memory-ControlledMachine
The actuator feedback circuit shown in the absolute-empirical (AE) cell memory-controlled machine inFigure 6 can be added to the conditional-predetermined
memory-controlled machine shown in Figure 3, formingthe monolithic conditional-empirical memory-controlledmachine shown in Figure 8.
The sensitivity of the selecting CE memory cell isfractionally decreased. Then, the actuator feedbackcircuit measures the actual action that takes place in agiven transition cycle. It then fractionally increases thesensitivity of a feedback-selected CE cell, which is atthe intersection of the actual value of the actuatorco-variable (Z9) and the value of the input variable (A)in that transition cycle. This makes the feedback-selectedCE cell more likely to select the value of the actuatorvariable (Z) that takes place with the value of the sensorvariable (A) in that transition cycle, as explained in thenext section.
2.4. Logarithmic Response Curves of a CE Cell
The logarithmic subtraction mechanism (LSM) in each
1335Consciousness in a Self-Learning, Memory-Controlled, Compound Machine
FIGURE 3. The analog sensitivity of each conditional-predetermined (CP) memory cell in the controller of a conditional-predeterminedmemory-controlled machine is determined by the position of the wiper arm on its variable resistor.
FIGURE 4. A falling logarithmic response curve is producedwhen the LSM subtracts a fixed percentage of the sensitivityeach time the conditional-predetermined (CP) memory cell isprogrammed with a ‘‘0’’. A rising logarithmic response curveis produced when the LSM adds the same fixed percentageof the difference between the sensitivity and the maximumsensitivity each time the conditional-predetermined (CP)memory cell is programmed with a ‘‘1’’.
FIGURE 5. A self-learning, sensor/actuator machine learns toproduce the action that is most likely to be carried out witheach sensed condition in a given transition cycle.

conditional-empirical (CE) memory cell decreases itssensitivity by a fixed percentage of its existing sensitivityvalue when it selects an action. It then increases itssensitivity by a fixed percentage of its difference from1 if that action actually takes place in that transitioncycle, creating the logarithmic response curves shownin Figure 9.
If more selection events occur than feedback events,the sensitivity of a CE cell will reach a value closer tothe minimum sensitivity. If more feedback events occurthan selection events, the sensitivity of the CE cell willreach a position closer to the maximum sensitivity. Ifan equal number of selection and feedback eventsoccur for a sufficient number of times, indicatingthat the selected action always occurs, the sensitivityof the CE cell will approach the mid-position. Usually,there are more selection events than feedbackevents, because not every selected action is carriedout successfully. Thus, a successful selection/feedbackevent increases the sensitivity of a CE cell toward themid-position asymptotically, which is the maximumsensitivity a cell can have if every selected action iscarried out.
2.5. The Conditional-Empirical Algorithm
The conditional-empirical algorithm shown in Figure 10is similar to the accounting system used in financialmarkets.
The conditional-empirical algorithm causes the mostsensitive memory CE cell connected to a sensed condi-tion in a given transition cycle to select an action, but atthe price of losing some of its sensitivity. If its actiontakes place, it regains more sensitivity than it lost. Thus,successful CE cells become more likely to select actions.However, the memory of an unsuccessful CE cell isreduced only slightly, leaving a permanent record ofprevious successes.
2.6. Duplex Control Memory
The control memory of a self-learning, memory-controlled machine works best if it is divided into two
1336 R. A. Brown
FIGURE 6. An absolute-empirical, memory-controlled machine learns the last action that is actually carried out with each sensedcondition.
FIGURE 7. The absolute-empirical algorithm is typical of a binarycomputer memory, which leaves no trace of a recorded valueonce it is erased.

memories. An input decoder unit is connected to anoutput encoder/decoder unit by an intermediate variable(I) and intermediate co-variable (I9). The two unitsform the duplex control memory shown in Figure 11.This configuration allows a value of the intermediatevariable (I) to represent a specific actuator state, andgreatly reduces the number of required memory cellswhen a limited number of sensor/actuator relations arerequired.
The self-learning duplex sensor/actuator machinemeasures the actual values of its actuator variables,and uses this feedback signal to decode a back-selected value of its intermediate co-variable (I9).This back-selected value of the intermediate co-variable(I9) is then used to increase the likelihood that theprogrammable input decoder will select the value ofthe intermediate variable (I) that is most likely to
produce the actual values of the actuator variable thattake place with the sensed conditions in that transitioncycle.
1337Consciousness in a Self-Learning, Memory-Controlled, Compound Machine
FIGURE 8. In a conditional-empirical, memory-controlled machine, a ramp signal causes the most sensitive CE cell connected to theexisting value of the sensor variable ( A) to select its value of the actuator variable ( Z ).
FIGURE 9. The logarithmic response curves show that thesensitivity stored in a conditional-empirical (CE) cell approachesthe mid-position when there are an almost equal number ofselection and feedback events.
FIGURE 10. According to the conditional-empirical alogrithm, abuyer’s liquid assets are reduced when a investment is made. Ifthe investment is successful, the assets are increased more thanthey were decreased, when the investment is liquidated.

2.7. Conditional-Empirical, Bidirectional ActuatorUnit
The CE cell matrix shown in Figure 8 can be used as aself-learning input decoder. However, it cannot be usedas the output encoder/decoder in a self-learning duplexmachine because it cannot feed back the informationabout the actual behavior of its actuators to its sensorunit. Thus, the conditional-empirical, bidirectionalactuator unit shown in Figure 12 is required in a self-learning duplex network.
Each bidirectional (BD3) memory cell uses the sameLSM and variable resistor (memory element) in theforward-selection and back-selection process. Theforward-selecting ramp (F-R) is isolated from the back-selecting (B-R) ramp by the diodes shown. A bidirec-tional actuator matrix is not used as a stand-alonemachine like the monolithic machine in Figure 8, ratheras a unit in an empirical network.
2.8. Conditional-Empirical, Duplex Network
Two or more of the scalar conditional-empirical matricesin Figure 8 can be connected to form a multivariablesensor matrix. Two or more of the scalar bidirectionalmatrices in Figure 12 can be connected to form a multi-variable actuator matrix. The multivariable sensor andactuator matrices can be connected to form the multi-variable, conditional-empirical, duplex network shownin Figure 13.
The conditional input matrix forward-selects a valueof the intermediate variable (I) where, in the set of CEcells connected to the latched terminal of each inputvariable (A and B) and the forward-selected value ofthe intermediate variable (I), the least sensitive CE cellin the set is more sensitive than the least sensitive CE
cell in any other set connected to the latched inputterminals and an unselected value of the intermediatevariable (I). The bidirectional actuator matrix thenforward-selects the value of the actuator variables (Yand Z) connected to the most sensitive bidirectionalmemory (BD3) cells connected to the forward-selectedvalue of the intermediate variable (I). The sensitivitiesof the forward-selecting memory cells in both matricesare reduced according to the conditional-empiricalalgorithm (Figure 10).
The bidirectional actuator matrix then back-selects avalue of the intermediate co-variable (I9) where, in the setof BD3 cells connected to the actual latched terminalof each actuator co-variable (Y9 and Z9) and the back-selected value of the intermediate co-variable (I9), theleast sensitive BD3 cell in the set is more sensitivethan the least sensitive BD3 cell in any other set con-nected to the latched actuator co-terminals and anun-back-selected value of the intermediate co-variable(I9). The sensitivities of the back-selecting BD3 cellsare increased logarithmically. Then the sensitivities ofthe conditional-empirical (CE) memory cells in the
1338 R. A. Brown
FIGURE 11. A duplex control memory must be programmed todecode the values of its sensor variables into a forward-selectedvalueof its intermediatevariable ( I ). Also, it must be programmedto encode this forward-selected value ( I ) into forward-selectedvalues of its actuator variables. The duplex network is similarto a computer memory system. The sensor matrix acts as aprogrammable address decoder, and the actuator matrix actsas a set of data (memory) registers.
FIGURE 12. A conditional-empirical, bidirectional actuator unitforward-selects a value of its output variable ( Z ) based upon thevalue of its intermediate variable ( I ) and the sensitivity of itsbidirectional (BD3) memory cells. It then back-selects a valueof its intermediate co-variable ( I9) based upon the value of itsactuator co-variable ( Z9) and the sensitivity of its bidirectional(BD3) memory cells.

input matrix connected to the latched values of the sensorvariables (A and B) and the back-selected value of theintermediate co-variable (I9) are increased logarithmi-cally according to the conditional-empirical algorithm(Figure 10).
2.9. Network Algorithm
The network algorithm shown in Figure 14 describes theoperation of a duplex network of empirical memorymatrices.
2.10. Digitized Self-Learning Machine
The empirical machine shown in Figure 13 has to learnbehavior for each value of the sensed conditions encoun-tered, even though the same output behavior may applyto many values of these sensed conditions. This problemcan be overcome by connecting an encoder to the sensorvariable and the sensor matrix through additional aggre-gate variablesA(a) and A(b). A decoder and feedbackencoder can be connected to additional aggregate actua-tor variablesZ(a) andZ(b) and the bidirectional actuatormatrix, as shown in Figure 15.
A generally high or a generally low value of the sensorvariable (A) can select a unique value of the intermediatevariable (I) through the most significant aggregatevariable (A(b)) when high sensitivity values appear inboth values of the least significant aggregate variable(A(a)). Alternatively, each specific value of the sensorvariable (A) can each select a unique value of the inter-mediate variable (I) through each combination of valuesof the most significant and least significant aggregatevariables (A(b) andA(a)).
1339Consciousness in a Self-Learning, Memory-Controlled, Compound Machine
FIGURE 13. The conditional-empirical (CE) and bidirectional (BD3) cells in a multivariable, conditional-empirical, duplex networkcontribute to the forward-selection and back-selection of a value of the intermediate variable ( I) that represents a sensor/actuator relationwith the highest historical probability.
FIGURE 14. A duplex network uses the network algorithm toforward-select values of its actuators according to the forward-selected value of its intermediate variable. It then back-selects avalue of the intermediate co-variable ( I9) that represents the valueof the intermediate variable ( I) most likely to forward-select theactual values of the actuator variables that occurs in thattransition cycle. The back-selected value of the intermediateco-variable ( I9) is then used to increase the sensitivity of theconditional-empirical (CE) cells in the sensor matrix.

A binary sensor encoder, actuator decoder, and feed-back encoder operating upon two aggregate variables isshown for the sake of simplicity. Additional aggregatevariables must be added if the number of values of asensor or actuator variable exceeds four. Trinary (three-value) encoders and decoders provide higher resolutionwith fewer memory cells than binary or quadraryencoders and decoders (Brown, 1994). The trinary digi-tized duplex network is the most effective configurationof the empirical machines presented.
3. COMPOUND MACHINE
If the environment of a sensor/actuator machine isremoved, it will stop running. However, a sensor/sensorunit can supply new sensed conditions even if the realsensed conditions are suspended. The sensor/sensorunit can be connected to the sensor variables of a
sensor/actuator machine, forming the compoundmachine (Brown, 1994) shown in Figure 16.
The four units of a compound machine make a uniquecontribution to its overall success. The sensor/sensor unitcan provide the illusion of an environment by producinga stream of new sensations. The actuator/sensor unitmay help the machine anticipate certain events bysupplying new sensed conditions based upon the actualbehavior of its actuators. The actuator/actuator unit cansupply routine actuator behavior, which may contributeto the physical skills of the machine without the need toinvolve the sensor systems.
3.1. Cycle Control Signal
Since these memory-controlled machines operate indiscrete steps at distinct moments, they may be calleddiscontinuous machines, in contrast to continuousmachines, such as analog computers. These discontinu-ous, memory-controlled machines require a cycle control
1340 R. A. Brown
FIGURE 15. A digitized self-learning machine can learn to provide general or specific representations of its sensor and actuator variables(self-partitioning), and can do so with a minimum number of memory cells and a minimum learning time.
FIGURE 16. A compound machine can keep running even if itsenvironment is removed.
FIGURE 17. The cycle control signal determines the periodbetween each input/output relation (transition).

signal to start and end each sense/action (transition)cycle, as shown in Figure 17.
Suspending the cycle control signal stops the behaviorof a discontinuous machine.
3.2. Ramp Signal
Conditional-predetermined and conditional-empiricalcontrol memories require the additional ramp (search)signal shown in Figure 18 to find the action that occursmost likely with a sensed condition.
Loss of the ramp signal results in the suspension ofall behavior produced by the conditional (CP and CE)memory cells, but does not influence the behavior ofthe absolute (AP and AE) memory cells shown inFigs 2, and 6.
3.3. Mixed Memory Matrix
Absolute-predetermined (AP) cells in Figure 2 andabsolute-empirical (AE) cells in Figure 6 can be modifiedto operate with conditional-predetermined (CP) cells inFigure 3 and conditional-empirical (CE) cells in Figure 8,as shown in the mixed memory matrix in Figure 19.
The values of the mixed absolute-predetermined(MAP) cells and conditional-predetermined (CP) cellsmay be determined beforehand, and be programmedinto mixed matrices to take care of self-regulation(homeostatic) functions of the machine. These predeter-mined memory cells may provide for innate behavioralmechanisms such as ‘‘ducking a brick’’ (Dennett, 1991).Behavior that must be learned can be established inconditional-empirical (CE) memory cells and in mixedabsolute-empirical (MAE) memory cells in an inputmatrix, and bidirectional empirical (BD3) cells in theremaining matrices after the machine is put into use.
The behavior of the empirical cells can override andsupersede the behavior of the predetermined cells ifconsistent behavior is found and learned. This is becausethe maximum sensitivity of the empirical cells can bedesigned to be greater than the sensitivity of the pre-determined cells. The predetermined cells can alsoprovide fallback behavior in cases where consistentbehavior cannot be found and learned. All of thesecells can be distributed throughout mixed memoriesof sensor/actuator, sensor/sensor, actuator/sensor, andactuator/actuator units of the compound machineshown in Figure 16.
1341Consciousness in a Self-Learning, Memory-Controlled, Compound Machine
FIGURE 18. The voltage ramp signal is used to find the CP cell orCE cell with the highest sensitivity in the set of conditional cellsconnected to the sensed value at the start of a transition cycle.
FIGURE 19. In a mixed memory matrix, the mixed absolute-predetermined (MAP) cells and conditional-predetermined (CP) cells canproduce instinctive behavior, while the mixed absolute-empirical (MAE) cells and conditional-empirical (CE) cells can produce learnedbehavior.

4. CONSCIOUSNESS
The sensor/sensor unit of a compound machine mayserve the function of consciousness. That is, it mayprovide an ongoing stream of internally-generatedsensations that carries the machine through states thatlack meaningful or useful external stimulation. Thisprocess may be extremely valuable in situations whereinterruptions occur in the normal stream of sensed con-ditions. For example, too much light may saturate thevisual sensors of an autonomous guided vehicle (AGV)if it looks directly at the Sun. Alternatively, the visualsensors may be unable to see in an environment withtoo little light, say when it enters a dark room. In thesecases, the sensor/sensor system of this AGV may be ableto imagine the normal view that would be expectedunder the circumstances that preceded the loss of theimage, and it could proceed as if it could see. Withoutthe contribution of the sensor/sensor unit (consciousnessunit) the AGV would have to be designed to stop whenits image sensor fails. Otherwise, it may produce uncon-trolled and/or inappropriate behavior.
Most of the memory cells in a sensor/sensor controlmemory of a compound machine may be conditional-empirical (CE and BD3) cells. This is because it is diffi-cult or impossible to determine the sequence of sensorvalues that may occur in a given task beforehand. How-ever, it is conceivable that predetermined memory cellsmay be used in a sensor/sensor controller to provideinnate illusions (Dennett, 1991) that are useful to theoperation of the machine.
4.1. Changing Levels of Consciousness
Changing the distribution of the ramp signals throughouta compound machine will result in major changes in itsbehavior. If the ramp signals are suspended to all theconditional memory cells in a compound machine witha mixed memory, the machine will go into what appearsto be a coma. In this case, the memory cells involvedin learning will not operate. Only the absolute-predetermined (AP) cells used for self-regulation willcontinue to operate, producing a vegetative state. If theramp signal is discontinued to the conditional memorycells of its sensor/sensor controller only, the compoundmachine will appear to be in a hypnotic state. It willappear to react to all outside suggestions, having lostits internal sense of intentionality. Other physical andmental states are produced when the cycle start andramp signals are discontinued to the other controllersin a compound machine. For example, a dreaming,sleep-like state can be produced if the ramp signal isdiscontinued to all of the controllers except the sensor/sensor controller. In this case, the machine can think,but cannot move. The trajectory of the sensor/sensorcontroller would correspond to a dream. If the ramp
signals were suspended to all of the controllers exceptthe actuator/actuator controller of a compound machine,the machine might go into sleep walking line-of-behaviorinvolving routine movements, such as getting out ofbed. If the cycle start signal were to be suspended to allof the controllers of a compound machine, a non-livingmachine would go into a state of suspended animation.Its memories would not change, and it would start rightup where it left off when the cycle start signals arereintroduced. However, a living being would die if itscycle start signals were discontinued. This is becauseits self-regulation functions would cease.
4.2. Tests of Consciousness
Physicians test the degree of consciousness of patients byasking them if they know what day it is, or what city theyare in. Since there are no obvious clues in the patients’room from which they can react responsively, they mustdig into their conscious mind to find the answer to thesetypes of questions. Thus, their answer provides a beha-vioral clue of consciousness (Herbert, 1993). Likewise, acompound machine can be placed in a task that involvessensor input. Then the sensors can be disconnectedintermittently. If the compound machine is able to copewith the task successfully, a strong case is made forthe machine having an internal representation (con-sciousness) of the external environment normally seenby its sensors. This internal representation is not amodel, but a dynamic line-of-behavior that is similar toour intuitive understanding of our conscious thoughtprocess. We seem to live in (act through) our consciousthoughts.
4.3. A Cycle and Ramp Signal Mode Controller
Since there may be many memory units in a complexcompound machine, there may be many overlappingcycle start signals and ramp signals. Each may operatein phase and out of phase with other signals. Somesignals may be present at certain times, and be suspendedat other times. The changing pattern of these signalsgreatly alters the type of behavior produced by a com-pound machine. These changes in behavior occurwithout changes in the basic memory state of themachine. This is similar to the changes in behavior thatoccur with changing patterns of brain waves in livingbeings. For example, patterns associated with sleepcreate different behaviors than patterns associated withwakefulness. Since these different types of behaviormay be useful in different situations, a separate modecontroller may be required. The mode controller can beused to control when and where the cycle start and rampsignals operate. Since each matrix is influenced by the
1342 R. A. Brown

cycle timing circuit, the mode controller can controleach cycle timing circuit, thus controlling the mode ofbehavior of a compound machine. For example, themode controller can determine the level of physicalaction required for a given task, and set the sensor/actuator and actuator/actuator ramps to produce theappropriate level of arousal. Likewise, the modecontroller can shut down the cycle start signals to anAGV used in a space voyage, thus immobilizing theAGV. It can then restart the cycle start signals whenthe AGV is ready to be deployed.
5. CONCLUSION
The local determinations of success rate by the logarith-mic subtraction mechanism (LSM) in each conditionalmemory cell allow the digitized, self-learning, com-pound machine described in this paper to learn thebehavior allowed by its operators and its environment.A useful enhancement of this compound machine is asensor/sensor unit that produces internal signals similarto our conscious or internal sensing process. ThisNeural activity (Freemam, 1995) can occur withoutthe machine sensing or acting in a real environment.Altering the cycle control and/or ramp signals of thiscompound machine can further enhance its behavior.This non-computational memory system may corres-pond to the living brain to a greater degree than con-ventional neural networks, which must sum currents,or fuzzy systems that depend upon global softwarecalculations.
5.1. Questions Raised by the Design of Memory-Controlled, Compound Machines
X Does neural activity cause brain waves, as iscommonly believed, or do brain waves cause theactivity of neural memory cells in the same waythat ramp signals cause conditional memory cellsto operate, as described above? If neural activitycauses brain waves, why are there no brain wavesin the spinal cord, where we know there is muchneural activity, but few if any memory cells?
X Do the cycle start signals and ramp signals, withtheir overlapping phase and different periods,correspond to brain waves? If so, the source ofthese signals in living beings may be found. Ifchemicals can be found that change the phase and/or relative strength of these signals, it may bepossible to suspend and re-establish consciousnessin a living being in a controlled manner. This mayprovide new ways of creating anesthesia and/orreviving individuals from coma.
REFERENCES
Ashby, W. Ross (1960).Design for a brain. New York: John Wiley andSons.
Brown, Robert Alan (1994).Machines that learn. New York: OxfordUniversity Press.
Dennett, Daniel C. (1991).Consciousness explained. New York: Little,Brown.
Freemam, Walter J. (1995).Society of brains. New Jersey: LawrenceErlbaum Associates.
Herbert, Nick (1993).Elemental mind: Human consciousness and thenew physics. New York: Dutton.
1343Consciousness in a Self-Learning, Memory-Controlled, Compound Machine




















