
Transforming Audiences, Transforming Societies-Alternative Media and Participation
Upload
independentCategory
view
2download
0
PTTG Binding Factor – a New Gene in Breast Cancer
Rachel J Watkins, Martin L Read, Vicki E Smith, Neil Sharma, Gary M Reynolds*, LauraBuckley+, Craig Doig†, Moray J Campbell†, Greg Lewy, Margaret C Eggo, Laurence SLoubiere, Jayne A Franklyn, Kristien Boelaert, and Christopher J McCabeSchool of Clinical and Experimental Medicine, University of Birmingham, B15 2TH, UK.*Centre for Liver Research, Institute of Biomedical Research, University of Birmingham, B15 2TH,UK.+Cancer Research UK Clinical Trials Unit, School of Cancer Sciences, University of Birmingham,B15 2TT, UK.†Department of Pharmacology & Therapeutics, Roswell Park Cancer Institute, Buffalo, NY USA14263.
AbstractPTTG Binding Factor (PBF; PTTG1IP) is a relatively uncharacterized oncoprotein whose functionremains obscure. Because of the presence of putative oestrogen response elements (ERE) in itspromoter, we assessed PBF regulation by oestrogen. PBF mRNA and protein expression wereinduced by both diethylstilbestrol and 17ß-estradiol in oestrogen receptor alpha (ERα) positiveMCF-7 cells. Detailed analysis of the PBF promoter showed that the region −399 to −291 relativeto the translational start site contains variable repeats of an 18 bp sequence housing a putativeERE half-site (gcccctcGGTCAcgcctc). Sequencing the PBF promoter from 122 normal subjectsrevealed that subjects may be homozygous or heterozygous for between 1 and 6 repeats of theERE. ChIP and oligonucleotide pull down assays revealed ERα binding to the PBF promoter. PBFexpression was low or absent in normal breast tissue, but was highly expressed in breast cancers.Subjects with greater numbers of ERE repeats demonstrated higher PBF mRNA expression, andPBF protein expression positively correlated with ERα status. Cell invasion assays revealed thatPBF induces invasion through Matrigel, an action that could be abrogated both by siRNAtreatment and specific mutation. Further, PBF is a secreted protein, and loss of secretion preventsPBF inducing cell invasion. Given that PBF is a potent transforming gene, we propose thatoestrogen treatment in post-menopausal women may up-regulate PBF expression, leading to PBFsecretion and increased cell invasion. Further, the number of ERE half sites in the PBF promotermay significantly alter the response to oestrogen treatment in individual subjects.
KeywordsPBF; Breast; Oestrogen; Invasion; Secretion
INTRODUCTIONBreast carcinogenesis requires multiple genetic changes, including the altered expressionand function of tumour suppressor genes and oncogenes. Most human breast cancers evolve
Corresponding author: Dr CJ McCabe, Reader in Endocrine Cancer, School of Clinical and Experimental Medicine, Institute forBiomedical Research, University of Birmingham, Birmingham, B15 2TH, UK. Tel: 44 121 415 8713; Fax: 44 121 415 8712;[email protected].
Europe PMC Funders GroupAuthor ManuscriptCancer Res. Author manuscript; available in PMC 2010 November 01.
Published in final edited form as:Cancer Res. 2010 May 1; 70(9): 3739–3749. doi:10.1158/0008-5472.CAN-09-3531.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

from normal epithelial cells in terminal duct lobular units through a series of increasinglyabnormal stages over long periods of time. Key stages in this progression are hyperplasia,atypical hyperplasia, in situ carcinoma, invasive carcinoma and, finally, metastatic disease(1). Invasion into surrounding stroma defines the transition from in situ to invasivecarcinoma. However, most defects responsible for the development and progression ofmalignant disease remain unknown (1).
Described in only 9 publications (2-10), 4 of which are from our own group (2;3;6;8),Pituitary Tumor Transforming Gene binding factor (PBF) was identified through its abilityto interact with PTTG1, the human securin (4). First isolated in 1997 (11), PTTG1 is anoestrogen-regulated gene (12;13) previously implicated in breast cancer, with highestexpression in invasive and metastatic breast cancers (14). Its binding partner PBF has not,however, been studied in the context of breast cancer before.
Initially identified in 1998 (10), PBF comprises 6 exons spanning 24 Kb withinchromosomal region 21q22.3. The 180 amino acid peptide sequence of PBF shares nosignificant homology with other human proteins, but is highly conserved across a widediversity of animal species (73% homology to mouse, 67% frog, 60% chicken, 52% zebrafish), suggesting both unique functionality and significant evolutionary importance.
PBF is ubiquitously expressed (15), but a decade after its cloning, very little has beenreported concerning its function. We previously characterised PBF expression in thyroidcancers, and demonstrated it to be a transforming gene in vitro, and to be tumourigenic invivo (8). Further, high PBF expression was independently associated with poor prognosis inhuman thyroid cancer. Most recently, we showed that PBF repressed iodide uptake inthyroid cells, both through transcriptional regulation (3) and through altered subcellulartrafficking (6).
PBF is thus a relatively uncharacterised transforming gene that plays a part in multiplecellular processes, particularly in the setting of endocrine neoplasia. We now presentextensive data, outlined below, that suggest PBF represents an entirely novel gene of directrelevance to breast cancer.
MATERIALS AND METHODSTissues
Breast tumour paraffin-embedded samples arranged on tissue microarrays (TMAs) wereavailable from 146 patients (16). There were 6 cases of ductal carcinoma in situ, 4 cases ofmedullary carcinoma, 1 atypical medullary carcinoma, 7 cases of mucinious type, 2 cases oftubular carcinoma, 22 cases of lobular carcinoma, 1 lobular-papillary and 1 benign tumour.A further 101 cases were designated pathologically as being of no special type (breasttumours not fitting the histological categories above, but tending to be invasive ductalcarcinomas that could not be further characterised on morphological grounds) (16). Clinicalfollow-up, encompassing tumour grade, vascular invasion, lymph node stage (LNS),Nottingham Prognostic Index (NPI) and ER status was available for the breast cancer series.Normal breast paraffin-embedded samples were available from US Biomax (Rockville,USA) (n = 6).
Cell lines and hormonal treatmentsMCF-7 human Caucasian breast adenocarcinoma cells were obtained from the EuropeanCollection of Cell Cultures (ECACC) in April 2008. Low passage number cells obtainedfrom the original stock were maintained in RPMI 1640 medium (Gibco) supplemented with10 % fetal bovine serum (FBS). Cells were treated with diethylstilbestrol (DES) and 17β-
Watkins et al. Page 2
Cancer Res. Author manuscript; available in PMC 2010 November 01.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

estradiol (EST) at final concentrations of 10 nM and 20 nM, and with ICI 182780 (Faslodex/Fulvestrant) at 100 nM and 1 μM, in RPMI 1640 phenol red free medium (Gibco)supplemented with 10 % charcoal stripped serum.
RNA extraction, reverse transcription quantitative PCR and Western blot analysisTotal RNA was extracted from MCF-7 cells using the Sigma Trisol Kit, as previously (3).RNA was isolated from paraffin-embedded tissues on microscope slides using thePinpoint™ Slide RNA isolation System II kit (Zymo Research, USA). RNA was reversetranscribed using the Reverse Transcription System (Promega), as previously (3).Expression of specific mRNAs was determined using the 7500 real time PCR system(Applied Biosystems) (8). Western blot analyses were performed as we have describedpreviously (2;6;17;18).
Crosslinking Chromatin ImmunoprecipitationBriefly, MCF-7 cells were cross-linked by addition of 1 % final concentration formaldehydedirectly to 1.5 × 106 mid-exponential cells. After sonication samples wereimmunoprecipitated with Protein A Agarose beads (Upstate Biotech, Lake Placid, NewYork) with 5 μg ER antibody (Santa Cruz, CA), alongside an IgG sample as a backgroundcontrol (Abcam, Cambridge, UK). Immunocomplexes were then sequentially washed andcrosslinks removed before DNA extraction. PBF promoter primers of sequence 5′GCA-GCC-CTT-TAG-GAT-GGA-G and 5′GAG-GAA-AGG-AGC-CTG-GTA-GC were thenused with 5 μl of ChIP DNA material for analysis by semi-quantitative PCR.
DNA ExtractionGenomic DNA from normal and tumourous colorectal samples and normal thyroidspecimens had previously been extracted (17;19). Normal DNA was also obtained fromwhole blood from patients with normal thyroid function (20). Genomic DNA was isolatedfrom breast tumour and normal tissue which had been formalin fixed and paraffin embeddedusing the Pinpoint Slide DNA Isolation System™ kit (Zymo Research, USA).
PCRTwo primer sets were designed to amplify the region of the PBF promoter which contains arepeated 18bp sequence. Forward (F) and Reverse (R) sequences were: Primer Set 1(F): 5′-GCG-CTC-CCC-TAG-TCC-CCT-3′; Primer Set 1(R): 5′-GCG-AGG-AGA-GCG-GCT-GA-3′; Primer Set 2(F): 5′-GCA-GCC-CTT-TAG-GAT-GGA-G-3′; Primer Set 2(R): 5′-GAG-GAA-AGG-AGC-CTG-GTA-GC-3′. Product sizes were 220 bp and 286 bp,respectively. Sequencing was carried out to confirm ERE repeat numbers using the forwardprimer of Primer Set 1 or 2.
Luciferase assaysThe pGL3_PBFpromoter construct was created by inserting bases −510 to −211 relative tothe translational start site into the pGL3 basic vector (Promega). Cells were harvested inPassive Lysis Buffer (PLB: Promega). The Dual Luciferase Reporter Assay System(Promega) was used, and data were expressed as a ratio of Renilla luciferase activity.
Biotinylated oligonucleotide pull-down assayPull-down assays were essentially as described previously (21). Oligonucleotides of aconsensus ERα binding sequence (22), 5′-GTC-CAA-AGT-CAG-GTC-ACA-GTG-ACC-TGA-TCA-AAG-TT- 3′, PBF ERE sequence (half sites in bold), 5′-CTC-GCC-CCT-CGG-TCA-CGC-CTC-GCC-CCT-CGG-TCA-CGC-CTC-GCC-CCT-CGG-TCA-CGC-CTC-
Watkins et al. Page 3
Cancer Res. Author manuscript; available in PMC 2010 November 01.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

GCC-CCT C- 3′ and mutant PBF ERE sequence (base changes underlined), 5′-CTC-GCC-CCT-CAA-TTT-CGC-CTC-GCC-CCT-CAA-TTT-CGC-CTC-GCC-CCT-CAA-TTT-CGC-CTC-GCC-CCT-C-3′ were 5′end-labelled with biotin and incubated with recombinant ERαprotein (P2187; Invitrogen Ltd, Paisley, UK). In competition reactions, recombinant proteinwas additionally incubated with 1-2 nmoles of non-biotinylated oligonucleotide. DNA/protein complexes were captured with 0.1 mg of magnetic streptavidin beads (Promega,Madison, USA). Subsequently, bound proteins were probed with an anti-ERα antibody(sc-543; Santa Cruz, CA, USA).
ImmunohistochemistryNormal and tumourous breast specimens were immunostained using our specific rabbitpolyclonal antibody to PBF (1:200). For negative controls the primary antibody wasreplaced by 2 % normal serum. Sections were counterstained with Mayers Haematoxylin,and blinded scoring carried out according to the intensity (0 = not present, +1= least intense,+3 = most intense) and percentage (<25, 25-50, 50-75, >75) of PBF staining.
Invasion AssaysMCF-7 cells transfected with vector only (VO), haemagglutinin (HA)-tagged PBF (PBF-HA), or the PBF mutant Δ29-93 were seeded onto BD Falcon™ cell culture inserts (8 μMpore size). Subsequently, 800 μl of RPMI 1640 medium supplemented with 20 % FBS, orcharcoal stripped serum, was added to the well below the BD Falcon™ cell culture inserts.For knock-down, cells were treated with either 50 nM of Scrambled (negative control # 1,Ambion) or PBF specific siRNAs #14399 and #147350 (mixed in equal quantities;Ambion). After 24 or 48 hours cells were fixed and stained using Mayers Haematoxylin andEosin (Sigma).
MTT AssaysCells were transfected or treated with diethylstilbestrol and 17β-estradiol at finalconcentrations of 10 nM and 20 nM. 24 and 48 hours post-PBF transfection, or 48 hourspost-oestrogen treatment, 100 μg of MTT was added to each well, as previously (23).
Detection of PBF secretion by Western blottingCell lysates were harvested in RIPA buffer. Cell medium was removed from MCF-7 cellsand centrifuged to remove cellular debris. Supernatants were then added to 3 volumes of 100% ethanol, centrifuged, and the pellet resuspended in RIPA buffer.
Detection of PBF secretion by immunoprecipitating radiolabelled PBFCells were maintained in 1 ml of medium containing 2/3 standard medium and 1/3 leucine-free equivalent, along with 10 μCi L-leucine [3,4,5-3H] (MP Biomedicals). Media extractswere centrifuged to remove floating cells. To obtain cell lysate fractions, cells were lysed inRIPA buffer. 5 μl of rabbit polyclonal anti-PBF antibody (6) or 5 μl normal rabbit controlserum were added to cell medium and lysate fractions. Immunocomplexes were pelleted bycentrifugation at 13,000g, and disintegrations per minute measured. Western blot analysis ofPBF immunoprecipitation confirmed that PBF was specifically pulled down.
Immunofluorescent analysis of PBF vesicular localisationMCF-7 cells were transfected with 1 μg PBF-HA and 1 μg chromogranin A-GFP (24) oncoverslips. Cells were fixed and permeabilised before blocking. Rabbit polyclonal anti-HA(Y-11) antibody (Santa Cruz Biotechnology, Santa Cruz, CA, USA) was used as primaryantibody, and Alexa Fluor 594-conjugated goat anti-rabbit IgG (Invitrogen) as secondaryantibody. Hoechst stain was used to visualise nuclei (1:1000).
Watkins et al. Page 4
Cancer Res. Author manuscript; available in PMC 2010 November 01.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Statistical analysisData were analysed using Sigma Stat (SPSS Science Software UK Ltd). Normallydistributed data were analysed using a two sample Student's t-test. The Mann Whitney RankSum test was used for comparison between two groups of non-parametric data. Datacontaining categorical information were analysed using the Chi-squared test and Fisher'sExact test.
RESULTSPBF mRNA and protein expression are induced by oestrogen
The binding partner of PBF – PTTG1 – is an oestrogen regulated gene implicated in theaetiology of breast cancer (14). We therefore examined whether PBF might also beoestrogen regulated. PBF mRNA expression was induced 48 hours after treatment by 10 nMand 20 nM diethylstilbestrol (DES) and 10 and 20 nM 17ß-estradiol (EST) in ERα-positiveMCF-7 cells (Figure 1A). PBF protein expression levels were also significantly up-regulated2 to 3-fold compared with vehicle controls (Figure 1B). Further, treatment with the anti-oestrogen ICI 182780 at 100 nM and 1 μM concentrations repressed 17ß-estradiolstimulation of PBF expression in a dose-dependent manner (Figure 1C).
To investigate whether 17ß-estradiol stimulation of PBF expression occurred directlythrough the PBF promoter, crosslinking chromatin immunoprecipitation (ChIP) assays werecarried out as described in the Materials and Methods. In each case (N=4 experiments),oestrogen receptor alpha bound to the human PBF promoter, with most pronounced bindingoccurring between 24 and 48 hours post oestrogen treatment in MCF-7 cells (Figure 1D).
The human PBF promoter is polymorphic for ERE half sitesIn silico analysis of the human PBF promoter identified the region −399 to −292 relative tothe translational start site to be replete with putative EREs (Figure 2A). Preliminarysequencing of this region revealed that it contained variable repeat numbers of an 18 bpsequence housing a putative consensus ERE half-site (gcccctcGGTCAcgcctc) (Figure 2A).PCR and sequencing of the region was additionally examined with a separate set of primers,confirming the existence of polymorphic numbers of 18 bp repeats (Figure 2A).
A panel of 92 genomic DNA samples was available to us, DNA being prepared from normalthyroid and colon tissue, and from tumourous colon. PCR and sequencing revealed thatsubjects may be homozygous or heterozygous for between 1 and 6 repeats of the 18 bpregion housing the ERE (Figure 2B). We next examined ERE repeat number in formalin-fixed paraffin-embedded breast tumour microarray (TMA) samples. Of the ~60 tumoursassessed, 27 tumours gave informative sequencing data for ERE repeat number, and 3 out ofthe 6 normals yielded unambiguous sequence data. These studies revealed that breasttumours could either be homozygous (3/3 repeats) or heterozygous (3/5 repeats), whereasthe N=3 normal breast specimens were all homozygous for 3 repeats (Figure 2B). None ofthe other variants were detected in breast DNA. Overall, as each 18 bp repeat houses aputative consensus ERE half site, subjects therefore differ in the number of EREs present intheir PBF promoter.
The promoter region −399 to −292 is oestrogen responsiveHaving determined that PBF is regulated by oestrogen and that ERα binds to the PBFpromoter in ChIP assays, and having identified a polymorphic region in the human PBFpromoter containing variable numbers of putative ERE half sites, we next examined theoestrogen responsiveness of this fragment of the promoter. 20 nM DES and EST bothinduced significant luciferase activity compared to vehicle-treated cells after 24 hours
Watkins et al. Page 5
Cancer Res. Author manuscript; available in PMC 2010 November 01.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

(Figure 2 C). At 48 hours, the effect was more pronounced (DES: 1.8 ± 0.2-fold, p < 0.01compared with vehicle, N = 3; EST: 1.7 ± 0.3-fold, p<0.05 compared with vehicle, N = 3)(Figure 2 C).
Next, we investigated whether the polymorphic ERE half sites identified within thepromoter region −399 to −292 were capable of binding oestrogen receptor alpha (ERα)directly. We performed pull-down assays using oligonucleotides containing either a series of3 wild type or 3 mutated PBF ERE half site repeats. Recombinant human ERα boundspecifically to the biotinylated double-stranded PBF ERE oligonucleotide (Figure 2D),which could be competed out by incubation with increasing excesses of unlabelled PBFoligonucleotide. Further, recombinant ERα bound a biotinylated consensus double strandedERE oligonucleotide, and binding was repressed by competition with unlabelled wild typePBF ERE oligonucleotide, but not by the mutated PBF oligonucleotide in which the EREshad been abolished. As PBF mRNA was induced 1.5 to 3-fold at identical timepoints and atidentical doses of oestrogen (Figure 1), these data indicate that the short promoter region−510 to −211 is positively regulated by oestrogen and confers most, but not all, of PBF'sresponsiveness to DES and EST. In addition, the specific ERE half site region identifiedwithin the promoter region −399 to −292 is capable of binding ERα, suggesting that themain mechanism of oestrogen regulation of PBF is directly via the polymorphic EREs of theproximal promoter.
PBF expression and correlation in a breast tumour seriesWe next investigated whether PBF was expressed in human breast tumours. Initial mRNAinvestigations utilised 20 breast TMA specimens and 6 normal breast specimens. PBFmRNA expression was apparent in RNA extracted from 18/20 TMA tumour samples, with amean TaqMan RT-PCR ΔCt of approximately 8 to 10, suggesting robustly detectable levelsof expression (Figure 3A). Reverse transcriptase negative (RT-) controls confirmed thatamplification in the tumour samples examined was not an artefact of genomic DNAcontamination (data not shown). In contrast, PBF mRNA was not detected (ND) in any ofthe 6 normal breast specimens analysed (Figure 3A).
PBF protein expression was next quantified in a larger series of normal breast specimens(N=8) and TMA tumour samples (N=146) through immunohistochemistry, using our rabbitpolyclonal antibody (6). Examples of scoring intensities are provided in Figure 3B. PBFexpression was low or absent in normal breast tissue, whereas it was strongly expressed inepithelial cells of all tumour types and grades of breast tumour assessed (Figure 3C).Specificity of staining was confirmed in negative control experiments (data not shown).Importantly, ERα status positively and significantly correlated with the percentage of PBFprotein expression (p <0.001). However, the remaining phenotypic end-points examined(tumour grade, vascular invasion, lymph node stage or the Nottingham Prognostic Index)were not associated with PBF staining intensity or percentage expression.
Next, we investigated the relationship between ERE number and PBF mRNA expression. 8fixed tumour specimens and 3 normal breast samples yielded both mRNA and promotersequencing data. PBF expression was apparent in 86 % of 3/5 heterozygotes, but only 25 %of 3/3 homozygotes (p = 0.044). Thus, PBF is oestrogen-regulated, its expression is higherin ER-positive than ER-negative tumours, and a greater number of ERE repeats is associatedwith higher PBF expression.
Watkins et al. Page 6
Cancer Res. Author manuscript; available in PMC 2010 November 01.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

PBF and oestrogen both induce invasiveness in MCF-7 cellsThe exact function of PBF in cell transformation is not known (8). Given that PBF isinduced in breast cancer, and that invasion and metastasis are critical processes in breastcancer progression, we next assessed whether PBF might play a role in cell invasion.
When 1×105 cells/well were seeded in invasion assays, PBF over-expression was associatedwith a 2.5 ± 0.6-fold increased cell invasion compared to VO treatment at 24 hours (N=6,p<0.01), and 6.1 ± 2.9-fold increase at 48 hours (N=6, p<0.01; Figure 4A). At a density of2×105 cells/well, PBF induced a 2.5 ± 0.3-fold increase in cell invasion after 48 hours (N=6,p<0.01).
To determine whether the enhanced invasiveness of MCF-7 cells following PBF transfectionreflected an increase in cell proliferation, MCF-7 cells were transfected with VO or PBF,and cell number estimated after 24 and 48 hours using MTT assays (Figure 4B). PBF did notsignificantly increase the proliferation of MCF-7 cells after 24 hours, but did marginally(~10 %) increase cell number after 48 hours. These data suggest that increased proliferationdoes not explain PBF's influence upon invasion.
PBF confers oestrogen induction of MCF-7 cell invasionAs oestrogen induces PBF, and MCF-7 cells are well documented as an oestrogen-responsive cell line, we next examined the influence of treatment with 10 nM DES on cellinvasion. DES treatment induced MCF-7 cell invasion by approximately 2.3-fold comparedwith vehicle treatment (N = 3, p < 0.001) (Figure 5A). 10 nM and 20 nM diethylstilbestroland 17ß-estradiol treatment did not significantly alter MCF-7 cell proliferation (data notshown).
Subsequently, we validated transient knock-down of PBF in MCF-7 cells using siRNA. 50nM of a PBF-specific siRNA elicited ~80 to 90 % knock-down compared with a scrambledcontrol (Figure 5B). Cell invasion assays were then repeated after 24 hours in the presenceof 10 nM DES or vehicle. Critically, the increased cell invasion observed after DEStreatment in the presence of a scrambled control (103 ± 17 cells, p = 0.002, N = 2) wasabolished when PBF was simultaneously knocked down using a PBF specific siRNA (27 ±14 cells, P=NS, N=2) (Figure 5C). These data suggest that oestrogen induction of MCF-7cell invasion through Matrigel is mediated via PBF.
PBF is a secreted proteinAs the induction of invasion by PBF could not be explained by increased cell number, andbecause cell invasion is a process frequently associated with secretion, we assessed whetherPBF is a secreted protein. MCF-7 cells were grown in the presence of 3,4,5-3H-L-leucine,cell medium fractions harvested after 24 hours, and immunoprecipitations carried out withour PBF antibody (Figure 6). Immunoprecipitation of labelled PBF revealed that PBF isindeed a secreted protein, with vector-only transfected MCF-7 cells demonstrating thatapproximately 20 % of total cellular PBF is secreted over the experimental timeframe of 24hours (Figure 6A). This was significantly enhanced by transient over-expression of PBF (49± 3 % secretion, N = 3, p < 0.05 compared with VO). Further, a mutant of PBF lackingamino acids 29 to 93 (and hence a functional signal peptide and two potential putativeglycosylation sites (6)), showed reduced secretion into the cell medium compared with wildtype (28 ± 7 %, N = 3), but did not differ statistically from VO (Figure 6A).
To confirm that PBF is secreted into the cell medium, we further carried out Westernblotting for HA-tagged PBF. Cell lysates demonstrated successful transfection of HA-taggedPBF (Figure 6B). Wild type PBF is a putative glycoprotein (10), which runs as a doublet at
Watkins et al. Page 7
Cancer Res. Author manuscript; available in PMC 2010 November 01.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

around 25 to 30 kDa. Mutant Δ29-93 was, as anticipated, smaller, and ran at approximately20 kDa (Figure 6B). Whereas wild type PBF was easily detectable in cell media, confirmingthat PBF is a secreted protein, Mutant Δ29-93 was not apparent.
To investigate the mechanism by which PBF is secreted, MCF-7 cells were co-transfectedwith HA-tagged PBF and chromogranin A-GFP. Chromogranin A-GFP is a chimericphotoprotein that is transported via the regulated pathway for exocytosis (24), and hasrecently been used as a marker for secretory granules in MCF-7 cells (25). As demonstratedpreviously in other cell lines, PBF-HA was found within intracellular vesicles, oftendetected towards the periphery of cells (6), in keeping with a secretory phenotype (Figure6C). Although these vesicles were localised in a similar pattern to the chromogranin A-GFP-labelled secretory granules, no colocalisation with PBF was observed. This suggests thatPBF is secreted via the constitutive pathway of secretion rather than via regulated secretion.Overall then, PBF is secreted by MCF-7 cells, higher expression results in increasedsecretion, and deletion of the amino acid region 29-93 results in significantly attenuatedsecretion into the cell medium.
Induction of MCF-7 cell invasion is modulated by PBF secretionHaving shown that PBF is secreted by MCF-7 cells, we investigated the relationshipbetween secretion and invasion. As before, PBF induced significant cell invasion whenover-expressed (269 ± 52 invading cells, N = 10 experiments, p = 0.018 compared with VOcontrol; Figure 6C). However, Mutant Δ29-93 failed to induce cell invasion compared withVO (161 ± 45 invading cells, N = 10 experiments, p = 0.518 compared with VO). Thus,secretion of PBF contributes significantly to its induction of cell invasion.
In summary, PBF is a relatively uncharacterised proto-oncogene which is induced byoestrogen in MCF-7 cells, shows increased expression in breast cancer, and whichstimulates cell invasion, at least in part through secretion. On this basis, PBF warrantsfurther and intensive study in the context of breast cancer initiation and progression.
DISCUSSIONNumerous genetic changes governing the initiation, progression and metastasis of breastcancer have already been described, but new genetic markers and therapeutic targets arevital for continued progress in addressing the approximately half-million global deathsannually from the disease. Based on our investigations, we propose four principle lines ofevidence suggesting a critical role of PBF in breast cancer. First, PBF is highly expressed inbreast tumours, where its expression correlates with oestrogen receptor positivity. Second, itis regulated by diethylstilbestrol and 17β-estradiol, both at the mRNA and protein level.Third, PBF upregulation results in significant MCF-7 cell invasion through Matrigel, aphenomenon highly dependent upon PBF secretion. Fourth, repressing PBF expression inthe face of oestrogen stimulation prevents oestrogen mediated induction of cell invasion.Hence, oestrogen regulates PBF expression in MCF-7 cells, tumours show increased levelsof the protein, and its functional property in promoting invasion can be ameliorated boththrough mutation and knock-down.
Whilst highly conserved, the 180 amino acid peptide sequence of PBF shares no significanthomology with other human proteins. Its exact mechanisms of action have not been divined,but it has been shown to induce tumours in nude mice (8), to regulate expression andfunction of the sodium iodide symporter (3;6), and to interact with the human securin, PTTG(4). Our experiments are the first to report a role for PBF in the breast, and to elucidate afunction in cell invasion. We now propose that oestrogen regulates PBF mRNA and protein
Watkins et al. Page 8
Cancer Res. Author manuscript; available in PMC 2010 November 01.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

expression, and suggest that this is mediated predominantly by a cluster of ERE half sites~300 bp upstream of the translational start site.
One surprising finding of our preliminary sequence analysis was that the human PBFpromoter is polymorphic for an 18 bp repeat housing a putative ERE half site. Importantly, ahigher number of putative EREs was statistically associated with greater PBF mRNAexpression in the breast, oestrogen receptor positive tumours having significantly increasedPBF protein expression compared to receptor negative tumours. Although these findingswould need to be confirmed in a larger series of samples, a higher number of EREs in abreast tumour would therefore predict a greater response to circulating oestrogen and henceincreased expression of a known transforming gene.
Oestrogen has previously been shown to enhance expression of PTTG, the binding partnerof PBF (12;13), and PTTG upregulation has been described in breast cancer (14). Whilst itwas not the focus of the current investigation, it would be interesting to correlate PTTG andPBF expression in individual breast tumours. The functional implications of simultaneousover-expression are hard to gauge, given that both proteins are inherently multifunctionaland likely to have dependent and independent modes of action. However, future studies maydelineate the individual contributions of each gene to breast cancer initiation andprogression.
Our in vitro experiments were predominantly carried out in MCF-7 cells, which remain the‘gold standard’ ERα-positive and oestrogen-sensitive breast cell line (26). Further, MCF-7cells are weakly invasive (27), allowing us to perform physiologically relevant invasionassays.
Our wider clinical associations did not reveal striking associations between PBF expressionand tumour phenotype, which was unexpected. Clinical associations were hampered by alack of a matched normal:tumour cohort, which would allow a more detailed interrogationof the association between promoter polymorphism and clinical outcome. Further, freshtissue would have allowed us to perform Western blotting for PBF expression, which wouldhave provided more quantitative expression data than through immunohistochemistry onFFPE slides.
The mechanism by which PBF induces cell transformation is not known. Our current studysuggests that this might be via the induction of cell invasion. MCF-7 cells showed potentincreases in invasion through Matrigel in response to PBF transfection. A mutant PBF whichwas not secreted into the cell medium lost this phenotype. Further, a specific siRNA entirelyblocked oestrogen induction of cell invasion. Thus, we hypothesise that oestrogen inducesPBF which in turn drives breast cell invasion, at least in part through secretion. As oestrogenstimulation of PBF expression was abrogated by co-treatment with the anti-oestrogen ICI182780, oestrogen receptor antagonists might be useful in treating tumours with high PBFexpression, to block potential cell invasion mediated by PBF.
In summary, we present evidence that the poorly characterised proto-oncogene PBF hasparticular relevance to breast tumourigenesis, particularly with respect to progression. Basedon our in vitro and ex vivo findings we predict that oestrogen treatment in post-menopausalwomen may up-regulate PBF expression, leading to increased PBF secretion. Given thatindividuals have different numbers of ERE repeats, it is likely that oestrogen will stimulatePBF expression variably, with a potentially critical impact upon breast cell invasion.
Watkins et al. Page 9
Cancer Res. Author manuscript; available in PMC 2010 November 01.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

AcknowledgmentsWe thank Professor Richard Cheney (University of North Carolina at Chapel Hill, NC, USA) for the provision ofthe chromogranin A-GFP construct.
Financial support: This work was supported by grants from the IDA Cooper Foundation and the Medical ResearchCouncil.
References1. Allred DC, Medina D. The relevance of mouse models to understanding the development and
progression of human breast cancer. J Mammary Gland Biol Neoplasia. 2008; 13:279–88.[PubMed: 18704660]
2. Boelaert K, Tannahill LA, Bulmer JN, Kachilele S, Chan SY, Kim D, et al. A potential role forPTTG/securin in the developing human fetal brain. FASEB J. 2003; 17:1631–9. [PubMed:12958169]
3. Boelaert K, Smith VE, Stratford AL, Kogai T, Tannahill LA, Watkinson JC, et al. PTTG and PBFrepress the human sodium iodide symporter. Oncogene. 2007; 26:4344–56. [PubMed: 17297475]
4. Chien W, Pei L. A novel binding factor facilitates nuclear translocation and transcriptionalactivation function of the pituitary tumor-transforming gene product. J Biol Chem. 2000;275:19422–7. [PubMed: 10781616]
5. Mo Z, Zu X, Xie Z, Li W, Ning H, Jiang Y, et al. Antitumor effect of F-PBF(beta-TrCP)-inducedtargeted PTTG1 degradation in HeLa cells. J Biotechnol. 2009; 139:6–11. [PubMed: 18977400]
6. Smith VE, Read ML, Turnell AS, Watkins RJ, Watkinson JC, lewy gd, et al. A novel mechanism ofsodium iodide symporter repression in differentiated thyroid cancer. Journal of Cell Science. 2009;122:3393–402. [PubMed: 19706688]
7. Stock M, Schafer H, Fliegauf M, Otto F. Identification of novel genes of the bone-specifictranscription factor Runx2. J Bone Miner Res. 2004; 19:959–72. [PubMed: 15190888]
8. Stratford AL, Boelaert K, Tannahill LA, Kim DS, Warfield A, Eggo MC, et al. Pituitary tumortransforming gene binding factor: a novel transforming gene in thyroid tumorigenesis. J ClinEndocrinol Metab. 2005; 90:4341–9. [PubMed: 15886233]
9. Tfelt-Hansen J, Yano S, Bandyopadhyay S, Carroll R, Brown EM, Chattopadhyay N. Expression ofpituitary tumor transforming gene (PTTG) and its binding protein in human astrocytes andastrocytoma cells: function and regulation of PTTG in U87 astrocytoma cells. Endocrinology. 2004;145:4222–31. [PubMed: 15178645]
10. Yaspo ML, Aaltonen J, Horelli-Kuitunen N, Peltonen L, Lehrach H. Cloning of a novel humanputative type Ia integral membrane protein mapping to 21q22.3. Genomics. 1998; 49:133–6.[PubMed: 9570958]
11. Pei L, Melmed S. Isolation and characterization of a pituitary tumor-transforming gene (PTTG).Mol Endocrinol. 1997; 11:433–41. [PubMed: 9092795]
12. Heaney AP, Horwitz GA, Wang Z, Singson R, Melmed S. Early involvement of estrogen-inducedpituitary tumor transforming gene and fibroblast growth factor expression in prolactinomapathogenesis. Nat Med. 1999; 5:1317–21. [PubMed: 10546001]
13. Heaney AP, Fernando M, Melmed S. Functional role of estrogen in pituitary tumor pathogenesis. JClin Invest. 2002; 109:277–83. [PubMed: 11805140]
14. Ogbagabriel S, Fernando M, Waldman FM, Bose S, Heaney AP. Securin is overexpressed in breastcancer. Mod Pathol. 2005; 18:985–90. [PubMed: 15846392]
15. Chen L, Puri R, Lefkowitz EJ, Kakar SS. Identification of the human pituitary tumor transforminggene (hPTTG) family: molecular structure, expression, and chromosomal localization. Gene. 2000;248:41–50. [PubMed: 10806349]
16. Murray PG, Lissauer D, Junying J, Davies G, Moore S, Bell A, et al. Reactivity with A monoclonalantibody to Epstein-Barr virus (EBV) nuclear antigen 1 defines a subset of aggressive breastcancers in the absence of the EBV genome. Cancer Res. 2003; 63:2338–43. [PubMed: 12727860]
Watkins et al. Page 10
Cancer Res. Author manuscript; available in PMC 2010 November 01.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

17. Kim D, Pemberton H, Stratford AL, Buelaert K, Watkinson JC, Lopes V, et al. Pituitary tumourtransforming gene (PTTG) induces genetic instability in thyroid cells. Oncogene. 2005; 24:4861–6. [PubMed: 15897900]
18. Kim DS, Franklyn JA, Stratford AL, Boelaert K, Watkinson JC, Eggo MC, et al. Pituitary tumor-transforming gene regulates multiple downstream angiogenic genes in thyroid cancer. J ClinEndocrinol Metab. 2006; 91:1119–28. [PubMed: 16394085]
19. Kim DS, Franklyn JA, Smith VE, Stratford AL, Pemberton HN, Warfield A, et al. Securin inducesgenetic instability in colorectal cancer by inhibiting double-stranded DNA repair activity.Carcinogenesis. 2007; 28:749–59. [PubMed: 17071631]
20. Simmonds MJ, Heward JM, Carr-Smith J, Foxall H, Franklyn JA, Gough SC. Contribution ofsingle nucleotide polymorphisms within FCRL3 and MAP3K7IP2 to the pathogenesis of Graves'disease. J Clin Endocrinol Metab. 2006; 91:1056–61. [PubMed: 16384851]
21. Liu Y, Asch H, Kulesz-Martin MF. Functional quantification of DNA-binding proteins p53 andestrogen receptor in cells and tumor tissues by DNA affinity immunoblotting. Cancer Res. 2001;61:5402–6. [PubMed: 11454683]
22. Vyhlidal C, Samudio I, Kladde MP, Safe S. Transcriptional activation of transforming growthfactor alpha by estradiol: requirement for both a GC-rich site and an estrogen response elementhalf-site. J Mol Endocrinol. 2000; 24:329–38. [PubMed: 10828826]
23. James SR, Franklyn JA, Reaves BJ, Smith VE, Chan SY, Barrett TG, et al. Monocarboxylatetransporter 8 (MCT8) in neuronal cell growth. Endocrinology. 2008
24. Taupenot L, Harper KL, Mahapatra NR, Parmer RJ, Mahata SK, O'Connor DT. Identification of anovel sorting determinant for the regulated pathway in the secretory protein chromogranin A. JCell Sci. 2002; 115:4827–41. [PubMed: 12432071]
25. Jacobs DT, Weigert R, Grode KD, Donaldson JG, Cheney RE. Myosin Vc is a molecular motorthat functions in secretory granule trafficking. Mol Biol Cell. 2009; 20:4471–88. [PubMed:19741097]
26. Brooks SC, Skafar DF. From ligand structure to biological activity: modified estratrienes and theirestrogenic and antiestrogenic effects in MCF-7 cells. Steroids. 2004; 69:401–18. [PubMed:15219790]
27. Hazan RB, Phillips GR, Qiao RF, Norton L, Aaronson SA. Exogenous expression of N-cadherin inbreast cancer cells induces cell migration, invasion, and metastasis. J Cell Biol. 2000; 148:779–90.[PubMed: 10684258]
Watkins et al. Page 11
Cancer Res. Author manuscript; available in PMC 2010 November 01.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Figure 1.A. PBF mRNA expression was induced maximally at 48 hours by 20 nM diethylstilbestrol(DES) and 17ß-estradiol (EST) in ERα-positive MCF-7 cells compared to vehicle treatedcells. ΔCt values from TaqMan RT-PCR with associated standard errors of the mean (SEM)are given below. B. PBF protein expression levels were up-regulated by 20 nMdiethylstilbestrol and 17ß-estradiol. Scanning densitometry (N=3 experiments) was used tocalculate a ratio of expression compared to β-Actin. Vehicle treatment (V) = 1.0. C.Oestrogen (17ß-estradiol) and anti-oestrogen ICI 182780 treatment of MCF-7 cells at 48hours. V = vehicle. D. Crosslinking chromatin immunoprecipitation (ChIP) analysis of ERαbinding to the human PBF promoter, yielding an expected product of 286 bp. IgG =immunoglobin control; NTC = no template control. *p<0.05; **p<0.01; ***p<0.001.
Watkins et al. Page 12
Cancer Res. Author manuscript; available in PMC 2010 November 01.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
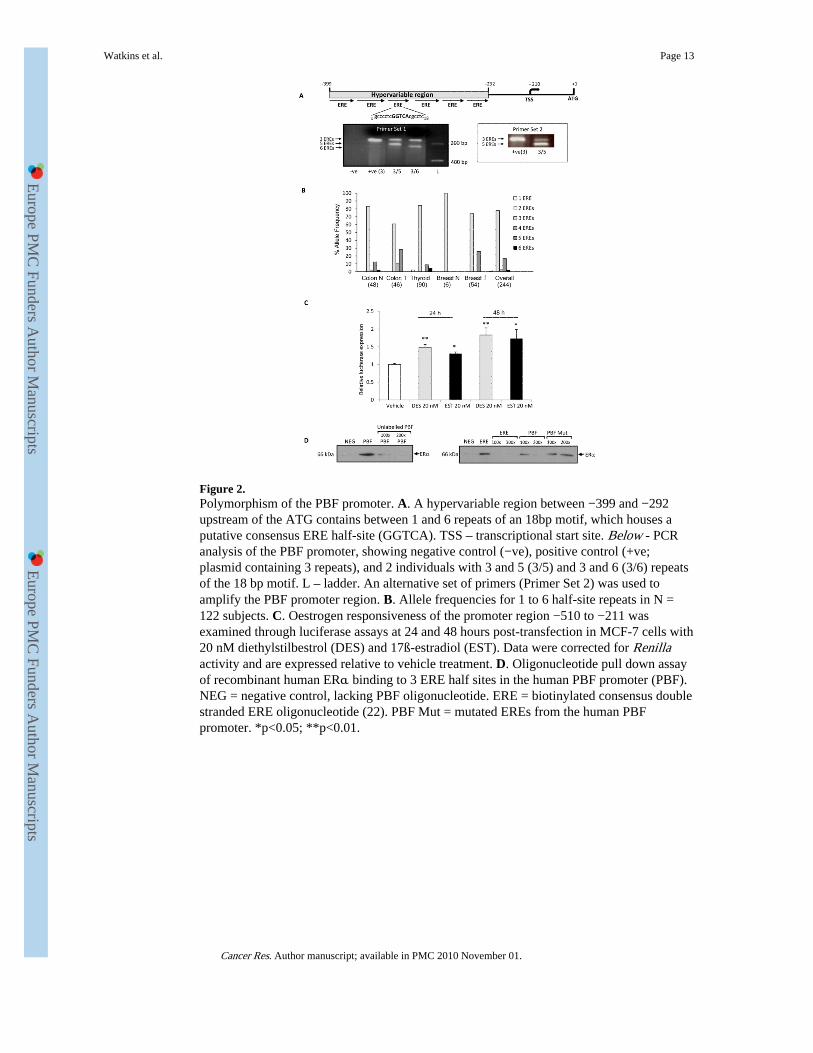
Figure 2.Polymorphism of the PBF promoter. A. A hypervariable region between −399 and −292upstream of the ATG contains between 1 and 6 repeats of an 18bp motif, which houses aputative consensus ERE half-site (GGTCA). TSS – transcriptional start site. Below - PCRanalysis of the PBF promoter, showing negative control (−ve), positive control (+ve;plasmid containing 3 repeats), and 2 individuals with 3 and 5 (3/5) and 3 and 6 (3/6) repeatsof the 18 bp motif. L – ladder. An alternative set of primers (Primer Set 2) was used toamplify the PBF promoter region. B. Allele frequencies for 1 to 6 half-site repeats in N =122 subjects. C. Oestrogen responsiveness of the promoter region −510 to −211 wasexamined through luciferase assays at 24 and 48 hours post-transfection in MCF-7 cells with20 nM diethylstilbestrol (DES) and 17ß-estradiol (EST). Data were corrected for Renillaactivity and are expressed relative to vehicle treatment. D. Oligonucleotide pull down assayof recombinant human ERα binding to 3 ERE half sites in the human PBF promoter (PBF).NEG = negative control, lacking PBF oligonucleotide. ERE = biotinylated consensus doublestranded ERE oligonucleotide (22). PBF Mut = mutated EREs from the human PBFpromoter. *p<0.05; **p<0.01.
Watkins et al. Page 13
Cancer Res. Author manuscript; available in PMC 2010 November 01.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Figure 3.A. Expression of PBF mRNA relative to 18s rRNA (ΔCt values) in 20 TMA samples ofbreast tumours compared with normal breast. ND – Not detected after 40 cycles of PCR. B.Representative immunohistochemical examination of PBF staining in 1 normal breastsample (US BioMax, Rockville, MD, USA) and 7 tumour samples from TMA sections.Columns 1 to 4 represent the different staining intensities observed, from 0 (absent) to +3(intense). Values in the bottom right hand corners indicate the percentage of PBF expressionobserved in the whole section, with original magnifications annotated next to tumour type.C. Representative immunohistochemical examination of PBF staining in 3 normal breastsamples (N1-N3; US BioMax, Rockville, MD, USA; 40× original magnification) and 3tumour samples from TMA sections. T1 - Grade I; T2 – Grade II; T3 – Grade III; all 40×original magnification.
Watkins et al. Page 14
Cancer Res. Author manuscript; available in PMC 2010 November 01.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Figure 4.A. PBF over-expression significantly increased the invasiveness of MCF-7 cells after both24 and 48 hours compared to vector-only (VO). Representative images of invading cells(arrowed) stained with Mayer's Haematoxylin (nuclear, blue) and eosin (cytoplasm, pink)are presented above. **p<0.01. B. MCF-7 cell number following transfection with vectoronly (VO) or PBF assessed through MTT assays. * p<0.05, N=4.
Watkins et al. Page 15
Cancer Res. Author manuscript; available in PMC 2010 November 01.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Figure 5.A. 10 nM diethylstilbesterol treatment of 1×105 native MCF-7 cells induced significantinvasion through Matrigel at 24 hours. B. siRNA knock-down of PBF. 10 nM DES inducedPBF in the presence of a scrambled siRNA control, compared to vehicle treatment.However, 50 nM PBF siRNA yielded 80 to 90 % knock-down of protein, which was notaltered by DES treatment. C. Invasion assays carried out in parallel to knock-downexperiments. DES = diethylstilbestrol, V = vehicle. Invading cells are arrowed.
Watkins et al. Page 16
Cancer Res. Author manuscript; available in PMC 2010 November 01.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts

Figure 6.Effect of wild type and mutant PBF overexpression on the secretion and invasiveness ofMCF-7 cells. A. The percentage of total PBF secreted into the medium extracted fromMCF-7 cells. Mutant Δ29-93 lacks amino acids 29 to 93. B. Western blot of cell mediumand whole cell lysate extracted from MCF-7 cells transfected with VO, wild type PBF andthe Δ29-93 PBF mutant. PBF constructs were HA-tagged. Whereas WT PBF is detectable inthe cell medium, Mutant Δ29-93 is not. C. Immunofluorescent subcellular analysis ofMCF-7 cells co-transfected with HA-tagged PBF and chromogranin A-GFP. (i) and (iv) –vesicular PBF-HA expression (red); (ii) and (v) – chromogranin A-GFP expression; (iii) and(vi) – merged image of PBF-HA and chromogranin A-GFP. D. Cell invasion assays. Incontrast to WT PBF, Mutant Δ29-93 failed to induce statistically significant cell invasion.Mean values ± SEM are shown. N = 10.
Watkins et al. Page 17
Cancer Res. Author manuscript; available in PMC 2010 November 01.
Europe PM
C Funders A
uthor Manuscripts
Europe PM
C Funders A
uthor Manuscripts
Copyright © 2022 FDOKUMEN




















