
?Phytoantibodies?: a general vector for the expression of immunoglobulin domains in transgenic...
10
Plant Molecular Biology 17: 865-874, 1991. © 1991 Kluwer Academic Publishers. Printed in Belgium. 865 'Phytoantibodies': a general vector for the expression of immunoglobulin domains in transgenic plants Eugenio Benvenuto 1, Ricardo J. Orders 2, Raffaela Tavazza 1, Giorgio Ancora 1, Silvia Biocca 3, Antonino Cattaneo 3 and Patrizia Galeffi 1 1ENEA Dipartimento Ricera e S viluppo Agroindustrali, Divisione Ingegneria Genetica C.P.2400, 00100 Roma A.D., Italy; 2Laboratorio de Fisiolog[a Vegetal, Facultad de Biologia, c~ Jesgts Arias de Velasco, 33005 Oviedo, Spain; 3CNR Istituto di Neurobiologia, Viale Marx 15, 00156 Roma, Italy Received 31 December 1990; accepted in revised form 21 June 1991 Key words: isolated VH domain (IVD), 'single domain antibody' (dAb), transgenic plants Abstract Sequences encoding the immunoglobulin heavy-chain variable (VH) domains were engineered in a new general purpose vector to transform plants via Agrobacterium. The expression of an isolated VH domain (IVD) after introduction into the plant genome has been monitored by northern, western and immuno- histochemical analysis. Immunoblotting showed that the polypeptide was stably expressed and ac- counted for up to 1 ~o of the soluble protein fraction. It is therefore proposed that single immunoglob- ulin domains of suitable specificity expressed in plants may constitute an effective system to inhibit the activity of molecules involved in plant pathology or plant development. Introduction The virtually unlimited repertoire of immunoglo- bulins is widely used as a source of specific re- agents for biochemical or physiological studies in which specific gene products can be inhibited or modulated. Gene transfer techniques can be used to achieve expression of specific immunoglobulin in different cell types. Efficient assembly of anti- body molecules has been shown to occur both in the secretory [6] or other intracellular compart- ments [2] of transfected non-lymphoid cells of different origin. This has led to the suggestion that the expression of genes for specific monoclonal antibodies in transgenic organisms can be used to perturb or interfere with the activity of selected cellular functions [ 34, 48 ]. In the light of the extensive use of Agrobacte- rium to transfer exogenous genes into the plant cell nuclear genome [47, 50] it should be possible to engineer the expression of appropriate specific antibody genes in plants and exploit the potential of the antibody repertoire for plant physiology and pathology studies. Production of complete antibody molecules in transgenic plants has re- cently been obtained either by crossing of plants expressing single y- or x-immunoglobulin chains [14, 15] or utilizing a single transformation step with an expression vector containing two differ- ent active promoters [ 12]. Simpler forms of the antibody molecules would probably be more apt for expression in plants, thus overcoming possi- ble protein assembly problems such as those aris- ing in the expression of intracellular cytosolic an-
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of ?Phytoantibodies?: a general vector for the expression of immunoglobulin domains in transgenic...

Plant Molecular Biology 17: 865-874, 1991. © 1991 Kluwer Academic Publishers. Printed in Belgium. 865
'Phytoantibodies': a general vector for the expression of immunoglobulin domains in transgenic plants
Eugenio Benvenuto 1, Ricardo J. Orders 2, Raffaela Tavazza 1, Giorgio Ancora 1, Silvia Biocca 3, Antonino Cattaneo 3 and Patrizia Galeffi 1 1ENEA Dipartimento Ricera e S viluppo Agroindustrali, Divisione Ingegneria Genetica C.P.2400, 00100 Roma A.D., Italy; 2Laboratorio de Fisiolog[a Vegetal, Facultad de Biologia, c~ Jesgts Arias de Velasco, 33005 Oviedo, Spain; 3CNR Istituto di Neurobiologia, Viale Marx 15, 00156 Roma, Italy
Received 31 December 1990; accepted in revised form 21 June 1991
Key words: isolated VH domain (IVD), 'single domain antibody' (dAb), transgenic plants
Abstract
Sequences encoding the immunoglobulin heavy-chain variable (VH) domains were engineered in a new general purpose vector to transform plants via Agrobacterium. The expression of an isolated VH domain (IVD) after introduction into the plant genome has been monitored by northern, western and immuno- histochemical analysis. Immunoblotting showed that the polypeptide was stably expressed and ac- counted for up to 1 ~o of the soluble protein fraction. It is therefore proposed that single immunoglob- ulin domains of suitable specificity expressed in plants may constitute an effective system to inhibit the activity of molecules involved in plant pathology or plant development.
Introduction
The virtually unlimited repertoire of immunoglo- bulins is widely used as a source of specific re- agents for biochemical or physiological studies in which specific gene products can be inhibited or modulated. Gene transfer techniques can be used to achieve expression of specific immunoglobulin in different cell types. Efficient assembly of anti- body molecules has been shown to occur both in the secretory [6] or other intracellular compart- ments [2] of transfected non-lymphoid cells of different origin. This has led to the suggestion that the expression of genes for specific monoclonal antibodies in transgenic organisms can be used to perturb or interfere with the activity of selected cellular functions [ 34, 48 ].
In the light of the extensive use of Agrobacte- rium to transfer exogenous genes into the plant cell nuclear genome [47, 50] it should be possible to engineer the expression of appropriate specific antibody genes in plants and exploit the potential of the antibody repertoire for plant physiology and pathology studies. Production of complete antibody molecules in transgenic plants has re- cently been obtained either by crossing of plants expressing single y- or x-immunoglobulin chains [14, 15] or utilizing a single transformation step with an expression vector containing two differ- ent active promoters [ 12]. Simpler forms of the antibody molecules would probably be more apt for expression in plants, thus overcoming possi- ble protein assembly problems such as those aris- ing in the expression of intracellular cytosolic an-
866
a)
(pUC 19)
(pGEM 9zf<->)
-X- -X--X I 4)<- -~ -~
XbXh E KSm Cs C (pOEM 7zf<+>)
Xb
(pBI 121,1)
H p
Biiiiiiiiiiii~it pel B
H
H
b)
, li~!i!iiiiiiii!#i!il
Right Npt II pel B Border 35 S CaMV
Bs E
m tag
E S I '
S m '
p S W I - V H N C 1 - T A G I
pBG - a 1
pBG - a2
m
pBG-dAb
tag NOS Left Border
T e r m i n a t o r
p B G - d A b - B I N
Xb
Xba I H ind III I I I I
TCTAGA CTCG AAGCT
Pet B Ps t I Leader '
A I I
ANY VH DOMAIN
S
I BstE II
, I
I GGTnACC
A I
Tag 1 EcoRI Sal I Sac I peptide J , I I I I
. . . . ~ . , I G A A T T C GTCOAC 6AGCTC ..
pBG-dAb

867
tibodies [14] and facilitating access of the molecule to the desired target structure.
The basic building block of any antibody-based antigen-binding structure is represented by the heavy-chain (VH) and light-chain (VL) variable domains [ 24 ]. Recently the successful cloning and expression of rearranged immunoglobulin heavy- chain variable domains in Escherichia coli has been reported [46]. The most intriguing aspect of this work was the discovery that the VH domain alone (without contribution from the VL domain) was able to bind antigen with good afl~ity. Con- sidering that 'single-domain antibodies' (dAbs) may offer a more versatile alternative to whole antibody molecules for regulated expression in different tissues or cellular compartments of transgenic plants, we have addressed the question as to whether such a 'handy' polypeptide can be efficiently expressed in plants.
We report in this paper the construction of a general purpose vector for the cloning of any VH variable region of interest in a form suitable for the subsequent transformation of plants. We uti- lized this vector system to drive the expression in transgenic plants of a selected VH domain from an anti-substance P (neuropeptide) antibody [9] which is unlikely to interact with endogenous plant antigens. We demonstrated that the trans- genic plants produced high levels of correctly pro- cessed isolated VH domains (IVD). The cellular distribution was studied by immunohistochemis- try, confirming that IVDs can indeed be expressed efficiently in plants.
Materials and methods
Plant material, bacterial strains and culture condi- tions
Nicotiana benthamiana plants were aseptically grown on half-strength Murashige and Skoog (MS) medium [27] (Flow) supplemented with 10 g/l sucrose, 8 g/l agar at 26 °C with 16 h day length under 6 W / m z light intensity.
E. coli strains DH5~ and BMH 71-18 were grown in LB [38] and in 2 x TY [23] medium respectively. Agrobacterium tumefaciens strain LBA4404 [16] was grown in AB minimal me- dium [ 22 ]. Antibiotic concentrations, when used, were for E. coli: kanamycin 50 #g/ml, ampicillin 100#g/ml, and for A. tumefaciens: kanamycin 100 #g/ml, rifampicin 10 #g/ml.
Recombinant DNA techniques
Unless otherwise specified, all standard tech- niques were performed as described [38]. All ma- nipulations were conducted under P1 contain- ment according to NIH Recombinant DNA guidelines.
Plasmid and constructs
The immunoglobulin cloning vector pSWl-VH- POLY-TAG 1 (generous gift of G. Winter, MRC, Cambridge) described by Ward et al. [46] is uti- lized for the inducible expression in bacteria of isolated VH domains and also contains a peptide
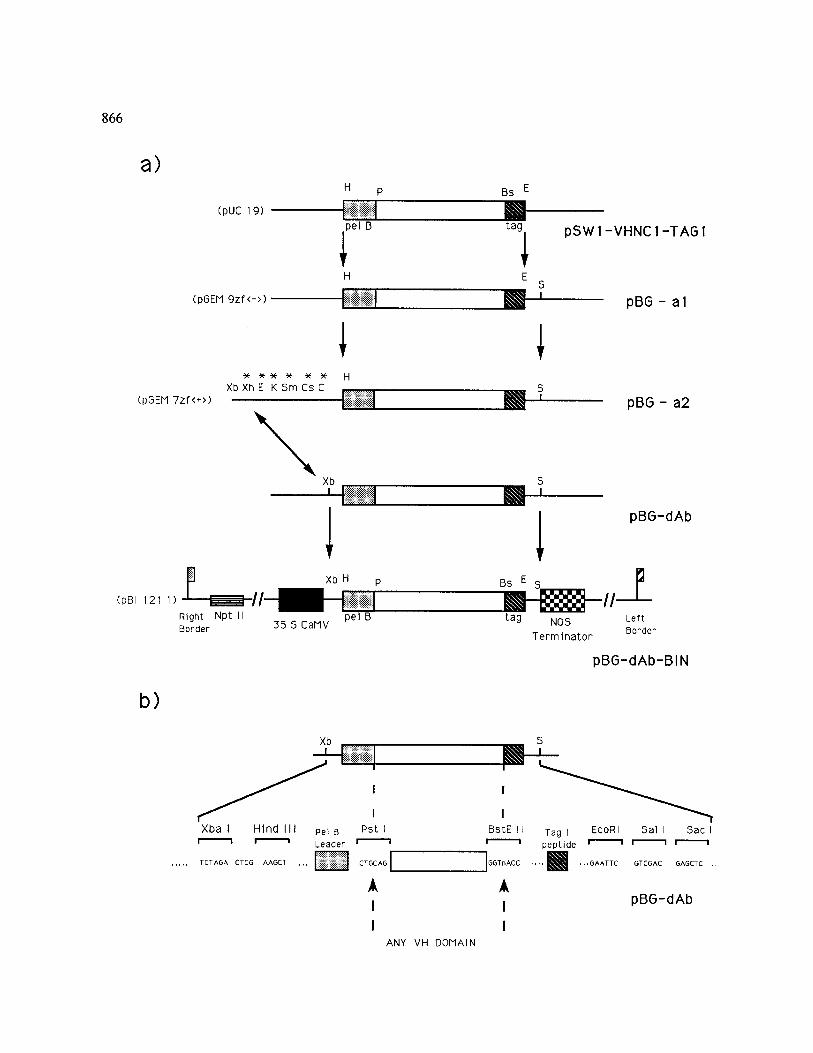
Fig. 1. a) Scheme for the construction of the VH immunoglobulin domain chimaeric gene introduced into plant cells. The plasmids and cloning steps are described in the 'Materials and methods' and 'Results' sections, respectively. Arrows indicate the direction of cloning. Sites marked with an * have been deleted. Bs, Bst EII; C, Cla I; Cs, Csp45 I; E, Eco RI; H, Hind III; K, Kpn I; P, Pst I; S, Sac I; Sm, Sma I; Xb, Xba I; Xh, Xho I. Right and left border indicate the 25 bp imperfect direct repeat sequences of T-DNA which define the vector (Ti-plasmid derived) portion destined for transfer [50]. The dotted box refers to the pel B leader sequence for secretion, the empty box to the VH variable region, the hatched box to the c-myc Tag (Tag 1) peptide sequence, the filled box to the 35S CaMV promoter and the chequered box to the NOS transcription terminator. The Npt II marker gene is indicated by the slatted box. Plasmid names in brackets indicate the original cloning vectors, b) Detailed map of the general purpose
vector (pBG-dAb) for the stepwise cloning of any variable region of interest to be expressed in plant cells.

868
tag (Tag 1; 11 amino acids from the human c-myc protein) recognized by the monoclonal antibody 9El0 [26]. The pSW1-VHNC1-TAG1 plasmid [34], derived by inserting the VH domain from the monoclonal antibody NC1/34 HL, was the starting material for our constructs (see Fig. la). The pBG-al plasmid was obtained by the sub- cloning of the VH expression cassette as a Hind III-Eco RI fragment into pGEM 9zf( - ). Subsequently, the pBG-a2 plasmid was obtained by subcloning of the insert into a pGEM 7zf( + ) as a Hind Ill-Sac I fragment. Unnecessary se- quences between the Xba I and Hind III sites of this plasmid were removed by restriction diges- tion and fill-in reaction to yield the general vec- tor pBG-dAb (see Fig. lb). The vector pBI 121.1 [18] (kindly provided by R.A. Jefferson, PBI Cambridge) was utilized for plant transformation after substitution of the fl-glucuronidase gene by the XBa I-Sac I cassette from pBG-dAB. All pGEM vectors were purchased from Promega.
Antibodies
All antibodies and immunological chemicals were obtained from Amersham. The anti-tag polypep- tide 9El0 monoclonal antibody [26] was a kind gift from H. Pelham (MRC, Cambridge).
Transformation of plants and NPT H assay
Mobilization of binary vector pBG-dAB- BIN(NC1) from E. coli into A. tumefaciens was performed by triparental mating as described by Ditta et al. [ 11 ]. Integrity of the construct was checked by restriction enzyme analysis. Leaf discs of four-week-old N. benthamiana plants were in- fected with Agrobacterium, basically following the procedure described by Tavazza et al. [42]. After 3-4 weeks regenerants appeared on selective ka- namycin (100/~g/ml) shoot-promoting medium (MS with 1 mg/l 3-indolebutyric acid (IBA), 1 mg/1 6-benzylaminopurine (BAP) and 30 g/1 su- crose), at a frequency of ca. 95~. Each leaf ex- plant usually gave rise to 13.8 + 1.8 shoots. Re-
generated shoots were rooted on selective kanamycin (25#g/ml) medium (1/2 MS with 0.05 mg/1 IBA and 30 g/1 sucrose) at a frequency of 51 + 3 ~o. Kanamycin-resistant plants were as- sayed for N P T I I (neomycin phospho-trans- ferase II) activity as described [40].
Nucleic acid analysis
Total DNA was extracted from leaves according to the method of Della Porta et al. [ 10] and was further purified on CsCI gradients. The procedure of Southern [41] was used to detect the presence of the IVD chimaeric gene. Total DNA (15 #g) was digested with Eco RI and Hind III and sub- jected to agarose gel electrophoresis (1~o) and transferred to nitrocellulose filter (Hybond-C, Amersham). Total RNA was extracted from leaves essentially using the acid guanidinium thiocyanate-phenol-chloroform procedure [ 8 ] uti- lizing the RNAgents kit (Promega). RNA (20/ag) was separated in 1.2~o formaldehyde-agarose gel and vacuum-blotted onto nylon membrane (Hy- bond-N, Amersham). Southern and northern blots were hybridized as recommended by the manufacturer with the 544 bp Eco RI-Hind III fragment from pBG-dAb-GEN(NC 1), purified by Geneclean kit (Bio 101) and labelled by the ran- dom priming procedure (Multiprime Labeling System, Amersham) to a specific activity of 5 × 10 9 cpm/#g DNA. Molecular weight markers were 1 kb DNA and 0.24-9.5 kb RNA ladder (BRL).
Protein extraction and analysis
Total proteins were extracted by grinding the whole plant to a fine powder in liquid nitrogen and extracted at 4 ° C in dounce grinder in 50 mM Tris-HC1, 0.5 M NaC1 and 0.25~/o Nonidet P-40 pH 7.6 in the presence of protease inhibitors (le- upeptin 15 #g/ml and crystals of PMSF) and cen- trifuged at 10000 x g for 30min at 4 °C (Frac- tion I). The resulting pellet was resuspended in 50mM Tris-HC1, 0.15 M NaC1, 0.5~o sodium

deoxycholate, 1~o Nonidet P-40, 0.1~o SDS pH 7.6 with protease inhibitors, homogenized and centrifuged as previously (Fraction II). The resid- ual pellet was treated with cold acetone for 2 h at -20 °C, centrifuged and dried in Speed-vac (Fraction III). Fractions I and II were subjected to affinity chromatography with activated Sepharose (Pharmacia) coupled to 9El0 mono- clonal antibody [26]. In all fractions the proteins were quantitated by the Bradford method [5] prior to electrophoresis in a 13 ~o polyacrylamide- SDS gel [20] and transferred electrophoreticaUy to a nitrocellulose (Hybond-C, Amersham) [44]. Western blots were incubated with 9El0 mono- clonal antibody followed by a rabbit anti-mouse serum coupled to horseradish peroxidase. Perox- idase activity was determined by development in 25 mM Tris-HC1, 150 mM NaCI pH 8.2, 0.6 mg/ ml 4-chloro-l-naphthol and 0.015 ~o H202. Rela- tive molecular masses of proteins were estimated by high- and low-range Rainbow markers (Amer- sham).
Immunohistochemical assay
Different organs of transgenic and control plants were sectioned (10 #m) as described by Cettomai [ 7 ] and prepared for indirect immunofluorescence essentially following the protocol of Langdale et al. [21]. Slides were incubated with 9El0 an- tibody followed by goat anti-mouse IgG biotiny- lated antibodies and fluorescence was developed by fluorescein isothiocyanate-labelled streptavi- din. Sections were observed and photomicro- graphed with a fluorescence microscope (Axio- phot Zeiss, Germany). All photomicrographs were recorded on Kodak Technical Ektachrome P800/1600. The following controls were included: (a) omission of the primary antibody; (b) normal goat serum, (c) omission of the fluorescent re- agent.
Isolation of extracellular solution
Infiltration techniques for the isolation of extra- cellular fluid from plant tissues were used. Stem and petioles were carefully sectioned and pro-
869
cessed essentially as described by Terry and Bon- ner [43] with the exception of the extraction buffer (50 mM MES, 0.3 M NaC1 pH 5.5, correspond- ing to 600 mOsm/kg H20). The recovered extra- cellular protein fraction was quantitated by Brad- ford's method [5]. Contamination of the extracellular solution by the cytosol from dam- aged cells was evaluated by comparing G6PD activity (monitored spectrophotometrically at 340 nm in 50 mM Tris-HC1, 5 mM MgCI2 pH 7.6, 60/~g/ml NADP, 3 mM G6P) in the secreted and the cytosolic fractions.
Results
Construction of vectors for IVDs and plant transfor- mation
Complementary DNA clones for the heavy-chain variable region (VH) of the monoclonal antibody NC1/34 HL [9], which recognizes a neuropep- tide, the tachykynin substance P, have recently been derived [34]. The VH domain, cloned into antibody expression vectors [ 32] and transfected in mammalian cells, together with its cognate light chain, retains the antigen-binding activity of the parental antibody [34]. The NC1 VH was sub- cloned into the pSW1-VHPOLY-TAG1 vector for expression of isolated VH domains in E. coli cells [46], yielding the plasmid pSWl-VHNC1- TAG 1 [ 34]. Expression and secretion of the cor- responding polypeptide from transformed E. coli can be demonstrated by western blotting (Fig. 3) with the 9El0 antibody directed against the C- terminal tag peptide (11 amino acids) from the human c-myc protein [26].
The plasmid pSW1-VHNC1-TAG1 was then adapted through a series of intermediate steps (Fig. la) to create the general-purpose vector pBG-dAb. In this vector (Fig. lb) any variable region of interest can be cloned utilizing the Pst I/ Bst EII restriction sites present in the PCR prim- ers described by Orlandi et al. [32] and pulled out as a complete VH cassette (Xba I-Sac I fragment) to be directionally cloned into the Agrobacterium binary vector, to yield pBG-dAb-BIN. The Xba I site in pBG-dAb is sufficiently close to the initial

870
ATG to ensure a short 5' untranslated region, as has been recommended [ 19]. The regulatory el- ements 35S CaMV (cauliflower mosaic virus) promoter [ 31 ] and NOS (nopaline synthase) ter- minator [4], widely used to drive high-level ex- pression of foreign genes in transgenic plants were chosen for these experiments. N. benthamiana plants were transformed (as described in Mate- rials and methods) with Agrobacterium cells har- bouring the plasmid pBG-dAb-BIN(NC 1). Forty independent putative transgenic plants rooted on kanamycin-selective medium were assayed for NPTII activity. More than 65 ~o of the transgenic plants were positive in this assay (data not shown). The putative transgenic plants multiplied in vitro appeared normal and did not reveal mor- phological changes from the control. Independent transformants were subjected to a Southern blot analysis (probing with the VH cassette contained in the Hind III-Eco RI fragment of pBG-dAb) (data not shown). The intensity of the bands hy- bridizing the probe together with evaluation by FACS (fluorescence-activated cell sorting) of the cellular DNA content (2 pg per haploid genome) allowed us to estimate that the copy number of the transgene in the different clones ranged be- tween 1 and 5.
Expression of the transgenic IVDs: RNA and pro- tein analysis
Total cytoplasmic RNA was extracted from leaves of fifteen NPTIIposi t ive independent
transformants at the same physiological stage, and the expression ofVH m R N A was assayed by northern blot analysis. Five of the plants were positive in this assay, including the clones 1A16, 1B2 and 2A3 (Fig. 2). A dramatic difference in the levels of the VH m R N A expression was found between the clones. This difference cannot be ac- counted for either by a difference in copy number or by degradation and may be due to position effeets in the plant genome [31, 39]. Initiation of transcription at the start site of 35S CaMV pro- moter and termination at the polyadenylation site in NOS terminator predicts a mRNA size of about 700 bases, consistent with that of the hy- bridizing bands (Fig. 2). The size of the VH RNA from the recombinant bacterial cells was about 100 bases smaller, as expected.
Soluble protein extracts were prepared from leaves of transgenic and control plants and the expression of the IVD protein was investigated by western blot analysis with the monoclonal anti- body 9El0 which recognizes the C-terminal tag peptide (see Fig. 1). All three positive clones showed a band of a relative molecular mass of about 15000 (Fig. 3, lanes E, F and G), which was absent in extracts from a control plant (Fig. 3, lane D). It is worth noting that the molecular weight of the plant IVD molecules was slightly lower than that of the bacterial IVD (compare lanes E, F, G with lane I in Fig. 3). It is well known that in bacterial cells the signal sequence for secretion is proteolytically cleaved from the mature protein [25, 36, 37]. Since the molecular weight of the IVD expressed in plants was ap-
Fig. 2. Northern blot analysis. Lanes A, C, D, E, F: RNA extracted from NPT II-positive plant clones 1B2, 2A3, 1A16, 1A5, 4B5. Lane B: untransformed control. Lane G: RNA from E. coli 71-18 strain 12 h after induction [46].
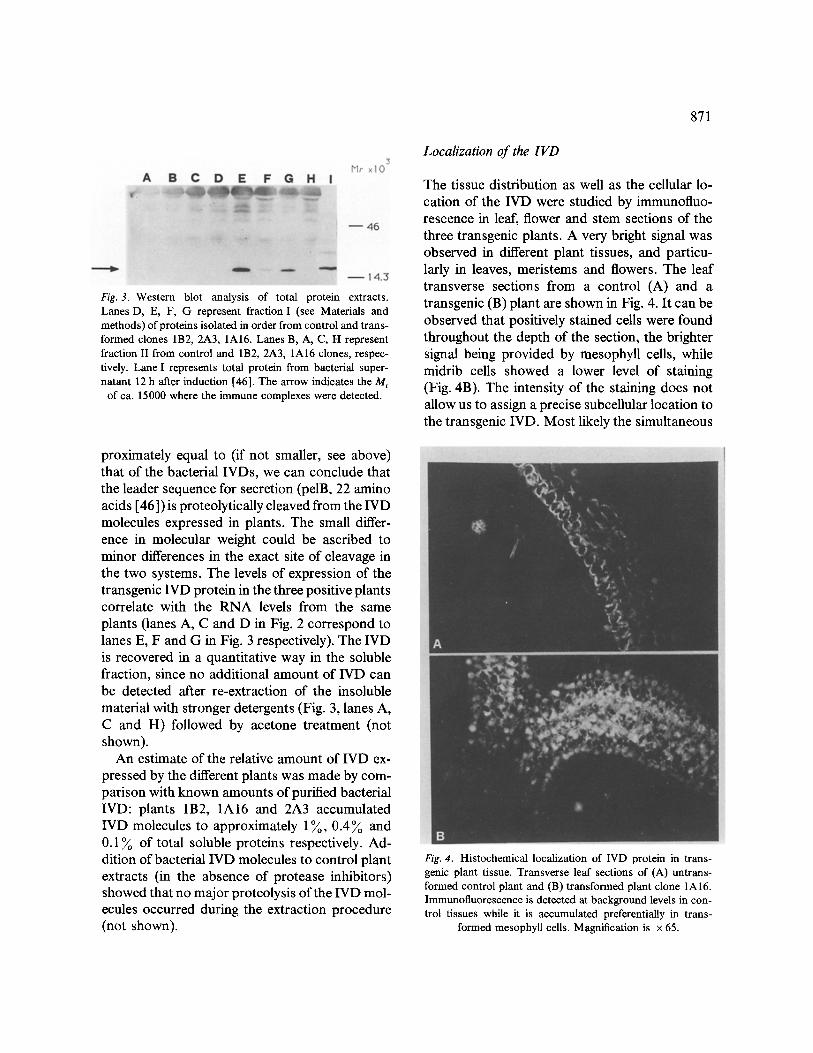
Fig. 3. Western blot analysis of total protein extracts. Lanes D, E, F, G represent fraction I (see Materials and methods) of proteins isolated in order from control and trans- formed clones 1B2, 2A3, 1A16. Lanes B, A, C, H represent fraction II from control and 1B2, 2A3, 1A16 clones, respec- tively. Lane I represents total protein from bacterial super- natant 12 h after induction [46]. The arrow indicates the M r
of ca. 15000 where the immune complexes were detected.
proximately equal to (if not smaller, see above) that of the bacterial IVDs, we can conclude that the leader sequence for secretion (pelB, 22 amino acids [46]) is proteolytically cleaved from the IVD molecules expressed in plants. The small differ- ence in molecular weight could be ascribed to minor differences in the exact site of cleavage in the two systems. The levels of expression of the transgenic IVD protein in the three positive plants correlate with the RNA levels from the same plants (lanes A, C and D in Fig. 2 correspond to lanes E, F and G in Fig. 3 respectively). The IVD is recovered in a quantitative way in the soluble fraction, since no additional amount of IVD can be detected after re-extraction of the insoluble material with stronger detergents (Fig. 3, lanes A, C and H) followed by acetone treatment (not shown).
An estimate of the relative amount of IVD ex- pressed by the different plants was made by com- parison with known amounts of purified bacterial IVD: plants 1B2, 1A16 and 2A3 accumulated IVD molecules to approximately 1~o, 0.4~o and 0.1 ~ of total soluble proteins respectively. Ad- dition of bacterial IVD molecules to control plant extracts (in the absence of protease inhibitors) showed that no major proteolysis of the IVD mol- ecules occurred during the extraction procedure (not shown).
871
Localization of the IVD
The tissue distribution as well as the cellular lo- cation of the IVD were studied by immunofluo- rescence in leaf, flower and stem sections of the three transgenic plants. A very bright signal was observed in different plant tissues, and particu- larly in leaves, meristems and flowers. The leaf transverse sections from a control (A) and a transgenic (B) plant are shown in Fig. 4. It can be observed that positively stained cells were found throughout the depth of the section, the brighter signal being provided by mesophyU cells, while midrib cells showed a lower level of staining (Fig. 4B). The intensity of the staining does not allow us to assign a precise subcellular location to the transgenic IVD. Most likely the simultaneous
Fig. 4. Histochemical localization of IVD protein in trans- genic plant tissue. Transverse leaf sections of (A) untrans- formed control plant and (B) transformed plant clone 1A16. Immunofluorescence is detected at background levels in con- trol tissues while it is accumulated preferentially in trans-
formed mesophyll cells. Magnification is × 65.

872
contribution to the fluorescence signal from dif- ferent focal plains of the section contributed to a blurring of the image. Immunoelectron and con- focal microscopy (work in progress) should allow more accurate determination of the intracellular distribution of the IVD molecules. The high lev- els of expression are consistent with the nomi- nally constitutive 35S CaMV promoter activity [3, 49, 28, 18].
As demonstrated above (Fig. 3), the signal peptide for secretion of the IVD molecule was cleaved from the IVD protein expressed in plants. While this suggests that the IVD had entered the endoplasmic reticulum, it does not necessarily imply that the protein was secreted. Therefore we attempted to follow the fate of the IVD polypep- tide by preparing an extracellular fraction (see Materials and methods). This fraction was esti- mated to contain about 0.05~o of total soluble proteins. No significant contamination from cy- tosol was present, as demonstrated by measuring G6PD activity (data not shown); moreover, the protein profiles of the intracellular and extracel- lular fractions differed significantly (data not shown). A western analysis of the extracellular fraction, however, indicated that small amounts of IVD molecules were indeed present in extra- cellular fraction (Fig. 5, lane H), although IVD molecules were much more easily revealed in cor- responding amounts of cell extracts (Fig. 3, lanes E, F, G and Fig. 5, lane D) (discussed below).
Discussion
In recent years techniques for gene transfer in different higher organisms have been applied to the immunoglobulin genes [33, 29]. Cloning of antibody genes of known specificity is greatly fa- cilitated by the polymerase chain reaction (PCR) and by the availability of suitable cloning vectors [32]. Therefore the virtually unlimited repertoire of the immune system may be exploited to achieve expression of antibodies in different cellular con- texts, where the effects of expressing the antibod- ies themselves could be studied [6]. The antibody molecules can be modified, either to acquire new
Fig. 5. Western blot analysis of extracellular protein fraction. Lane A: protein extraction from bacterial supematant. Lanes B, C: stem homogenate from clone 1A16 and negative control. Lanes D, E: protein fraction I of leaf from clone 1A16 and negative control. Lanes F, G: extracellular fraction tri- chloroacetic acid-precipitated from clone 1A16 and negative control. Lanes H, I: extracellular fraction from clone 1A16 and negative control. With the exception of samples in lanes F and G, all fractions were subjected to affinity chromatography
(9E10-coupled Sepharose) prior to electrophoresis.
effector functions [30] or to change their intra- cellular targeting [2]. The application of such an experimental strategy would be particularly use- ful in plant biology, where strategies to interfere with the activity of plant [45, 13] or pathogen [35] molecules are being actively pursued.
The results obtained confirm that high levels of IVD expression can be achieved with the vector described in this work. The IVD molecules in plants were proved to be correctly processed and quite stable, as demonstrated by their very large accumulation in leaf tissues. The proteolytic cleavage of the leader sequence for secretion sug- gests that this accumulation may occur in the secretory compartments of the cell in accordance with the results obtained by During et al. [12]. The amount of IVD molecules found in the ex- tracellular fraction is however low (Fig. 5). This may have different causes: (1) the relative amount of secreted protein might be low, due to the hy- drophobic nature of the isolated VH domain [46]; (2) the tag peptide of the IVD molecules may be proteolytically cleaved during the process of se- cretion, or outside the cell.
The efficient expression of IVDs with antigen- binding activity (dAbs) in plants may be partic- ularly suitable for the targeting of these molecules

to intraceUular compartments (cytoplasm, nu- cleus or organelles) [1 ]. This may allow to over- come assembly problems such as those reported for the expression of leaderless heavy and light immunoglobulin chains in the plant cell cytoplasm [ 14]. It is noteworthy that the general purpose vector described can be utilized not only for the expression in plants of any dAb of interest, but also for the expression of other forms of antibody domains (work in progress) such as single-chain antibodies (scAbs) [17].
Isolated VH domains have been shown to fold correctly in E. coli cells where, often but not al- ways, they retain the antigen-binding activity. In our case, however, the bacterial anti-substance P isolated VH domain is not able to bind the anti- gen with the same affinity as the parental NC1 monoclonal antibody [34]. Therefore, antigen- binding activity by plant-produced anti-substance P IVD per se could not be addressed in our con- ditions.
The results reported in this paper allow us to conclude that the expression of 'single VH do- mains' with antigen-binding activity may repre- sent an alternative strategy to interfere with the function of specific proteins involved in plant pathogenesis and/or development. The success of the approach will depend on other factors, such as the availability of specific antibodies, their antigen-binding capacity as 'single-domain anti- bodies', their functionality in different cell com- partments, the effectiveness of the immune com- plex in terms of stability and interactions with the normal physiological processes of the engineered plant.
Acknowledgements
The authors are grateful to the many colleagues at ENEA and CNR (Roma) for their useful suggestions and encouragement. Special thanks are due to Patrizia Piccioli for providing the pSWl-VHNC1-TAGI plasmid, and to Sergio Lucretti for FACS measurements. Finally, we acknowledge Riccardo Cettomai and Flavio Coppola for help in the histological sec- tions and photographic work. Part of this work was supported by a CNR grant Progetto Finalizzato Biotecnologie e Bios- trumentazioni to A.C.; S.B. is on leave of absence from the Dipartimento di Biochimica, Universit/t La Sapienza, Roma.
873
R.J.O. was supported by a fellowship of Ministerio de Educaci6n y Ciencia (Spain).
References
1. Biocca S, Cattaneo A: Expression ofintracellular Fv and Fab antibody fragments in mammalian ceils. Cell Biol Int Rep 14:217 (1990).
2. Biocca S, Neuberger MS, Cattaneo A: Expression and targeting of intracellular antibodies in mammalian cells. EMBO J 9 :101-108 (1990).
3. Benfey PN, Chua NH: Regulated genes in transgenic plants. Science 244:174-181 (1989).
4. Bevan MW, Barnes WM, Chilton MD: Structure and transcription of the nopaline synthase gene region of T- DNA. Nucl Acids Res 11:36%385 (1982).
5. Bradford M: A rapid and sensitive method for the quan- titation of microgram quantities of protein utilizing the principe of protein-dye binding. Anal Biochem 72: 248- 254 (1976).
6. Cattaneo A, Neuberger MS: Polymeric immunoglobulin M is secreted by transfectants of non lymphoid cells in the absence of immunoglobulin J chain. EMBO J 6: 2753- 2758 (1987).
7. Cettomai R: Tecnica per sezioni istologiche di apici veg- etativi in piante arboree. Inf Bot Ital 2:56-58 (1970).
8. Chomczynski P, Sacchi N: Single step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloro- form extraction. Anal Biochem 162:156-159 (1987).
9. Cuello AC, Galfre G, Milstein C: Detection of substance P in the central nervous system by a monoclonal anti- body. Proc Natl Acad Sci USA 76:3532-3536 (1979).
10. Della Porta SL, Wood J, Hicks JB: A plant DNA mini~ preparation: version II. Plant Mol Biol Rep 1(4): 19-21 (1983).
11. Ditta G, Stanfield S, Corbin D, Helsinki DR: Broad host range DNA cloning system for Gram negative bacteria: construction of a gene bank of Rhizobium melilott'. Proc Natl Acad Sci USA 77:7347-7351 (1980).
12. During K, Hippe S, Kreuzaler F, Schell J: Synthesis and self-assembly of a functional monoclonal antibody in transgenic Nicotiana tabacum. Plant Mol Biol 15:281-293 (1990).
13. Hamilton AJ, Lycett GW, Grierson D: Anti-sense gene that inhibits synthesis of the hormone ethylene in trans- genic plants. Nature 346:284-287 (1990).
14. Hiatt A, Cafferkey R, Bowdish K: Production of anti- bodies in transgenic plants. Nature 342:76-78 (1989).
15. Hiatt A: Antibodies produced in plants. Nature 344: 469- 470 (1990).
16. Hoekema A, Hirsch PR, Hooykaas PJJ, Schilperoort RA: A binary plant vector strategy based on separation of Vir- and T-region of Agrobacteriurn tumefaciens Ti plasmid. Nature 303:179-180 (1983).
17. Huston JS, Levinson D, Mudgett-Hunter M, Tai M-S, Novotny J, Margolies MN, Ridge RJ, Bruccoleri RE, Haber E, Crea R, Hoppermann H: Protein engineering of antibody binding sites: Recovery of specific activity in an anti-digoxin single-chain Fv analogue produced in E. coli. Proc Natl Acad Sci USA 85:5879-5883 (1988).

874
18. Jefferson RA, Kavanagh TA, Bevan MW: GUS fusions: betaglucuronidase as a sensitive and versatile gene fusion marker in higher plants. EMBO J 6:3901-3907 (1987).
19. Kozak M: Leader length and secondary structure mod- ulate mRNA functions under conditions of stress. Mol Cell Biol 8:2737-2744 (1988).
20. Laemmli UK: Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227: 680-685 (1970).
21. Langdale JA, Metzler MC, Nelson T: The Argentia mu- tation delays normal development of photosynthetic cell types in Zea mays. Dev Biol 122:243-245 (1987).
22. Lichtenstein C, Draper J: DNA Cloning: A practical Ap- proach, vol 2. IRL Press, Oxford (1986).
23. Miller JH: Experiments in Molecular Genetics. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1972).
24. Milstein C: From the structure of antibodies to the di- versification of the immune response. EMBO J 4: 1083- 1092 (1985).
25. Model P, Russel M: Procaryotic secretion. Cell 61: 739- 741 (1990).
26. Munro S, Pelham H: An Hsp70-1ike protein in the ER: identity with the 78 kd glucose-regulated protein and im- munoglobulin heavy chain binding protein. Cell 46: 291- 30 (1986).
27. Murashige T, Skoog F: A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol Plant 15:473-497 (1962).
28. Nagata T, Okada K, Kawazu T, Tabeke I: Cauliflower mosaic virus 35S promoter directs S phase specific ex- pression in plant cells. Mol Gen genet 207:242-244 (1987).
29. Neuberger MS, Caskey HM, Pettersson S, Williams GT, Surani MA: lsotype exclusion and transgene down- regulation in immunoglobulinlamba transgenic mice. Na- ture 338:350-352 (1989).
30. Neuberger MS, Williams GT, Fox RO: Recombinant an- tibodies possessing novel effector functions. Nature 312: 604-608 (1984).
31. Odell JT, Nagy F, Chua NH: Identification of sequences required for activity of the cauliflower mosaic virus 35S promoter. Nature 313:810-812 (1985).
32. Orlandi R, Gussow DH, Jones PT, Winter G: Cloning immunoglobulin variable domains for expression by the polymerase chain reaction. Proc Natl Acad Sci USA 86: 3833-3837 (1989).
33. Palmiter RD, Brinster RL: Germ-line transformation of mice. Annu Rev Genet 20:465-499 (1986).
34. Piccioli P, Ruberti F, Biocca S, Di Luzio A, Werge TM, Bradbury A, Cattaneo A: Neuroantibodies: molecular cloning of a monoclonal antibody against substance P for expression in the central nervous system. Proc Natl Acad Sci USA (in press) (1991).
35. Powell PA, Stark DM, Sanders PR, Beachy RN: Pro- tection against tobacco mosaic virus in transgenic plants that express tobacco mosaic virus antisense RNA. Proc Natl Acad Sci USA 86:6949-6952 (1989).
36. Randall LL, Hardy SJS: Unity in function in the absence of consensus in sequence: role of leader peptides in ex- port. Science 243:1156-1159 (1989).
37. Randall LL, Topping TB, Hardy SJS: No specific recog- nition of leader peptide by SecB, a chaperone involved in protein export. Science 248:860-863 (1990).
38. Sambrook J, Fritsch EF, Maniatis T: Molecular Cloning. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1989).
39. Sanders PR, Winter JA, Zarnason AR, Rogers SG, Fra- ley RT: Comparison of cauliflower mosaic virus 35S and nopaline synthase promoters in transgenic plants. Nucl Acid Res 4:1543-1558 (1987).
40. Scott R, Draper J, Jefferson R, Dury G, Jacob L: Anal- ysis of gene organization and expression in plants. In: Draper Je t al. (eds) Plant Genetic Transformation and Gene Expression, pp. 301-310. Blackwell Scientific Pub- lications, Oxford (1988).
41. Southern EM: Detection of specific sequences among DNA fragments separated by gel electrophoresis. J Mol Biol 98:503-517 (1975).
42. Tavazza R, Ordas R J, Tavazza M, Ancora G, Benvenuto E: Genetic transformation of Nicotiana clevelandii using a Ti plasmid derived vector. J Plant Physiol 133:640-644 (1988).
43. Terry ME, Bonner BA: Examination of centrifugation as a method of extracting an extracellular solution from peas, and its use for the study of indoleacetic acid-induced growth. Plant Physiol 66:321-325 (1980).
44. Towbin H, Staehelin T, Gordon J: Electrophoretic trans- fer of proteins from polyacrylamide gels to nitrocellulase sheets: procedure and some applications. Proc Natl Acad Sci USA 67:4350-4354 (1979).
45. van der Krol AR, Lenting PE, Veenstra J, van der Meer IM, Koes RE, Gerats AGM, Mol JNM, Stuitje AR: anti-sense chalcone synthase gene in transgenic plants inhibits flower pigmentation. Nature 333:866-869 (1988).
46. Ward ES, Gussow D, Griffiths AD, Jones PT, Winter G: Binding activities of a repertoire of single immunoglobu- lin variable domains secreted from Escherichia coli. Na- ture 341:544-546 (1989).
47. Weising K, Schell J, Kahl G: Foreign genes in plants: transfer structure, expression, and application. Annu Rev Genet 22:421--477 (1988).
48. Werge TM, Biocca S, Cattaneo A: Intracellular immuni- zation: cloning and intracellular expression of a mono- clonal antibody to the p21-ras protein. FEBS Lett 274: 193-198 (1990).
49. Williamson JD, Hirsch ME, Larkins BA, Gelvin SB: Differential accumulation of a transcript driven by the CaMV 35S promoter in transgenic tobacco. Plant Physiol 90:1570-1576 (1989).
50. Zambryski P, Temp6 J, Schell J: Transfer and function of T-DNA genes from Agrobacterium Ti and Ri plasmids in plants. Cell 56:193-201 (1989).




















