
Comparative Biochemistry of Chondroitin Sulphate-Proteins of ...

System. Appl. Microbiol. 21, 557-568 (1998) SYSTEI\t1ATIC AND _©_G_us_ta_v F_is_ch_er_V_erl_ag ________________ APPLIED MICROBIOLOGY
Phylogenetic and Physiological Diversity of Sulphate-Reducing Bacteria Isolated from a Salt Marsh Sediment
JULIETIE N. ROONEy-VARGA *, BARBARA R. SHARAK GENTHNER!, RICHARD DEVEREUX2, STEPHANIE G. WULIS2,
STEPHANIE D. FRIEDMAN2, and MARK E. HINES!,3
lInstitute for the Study of Earth, Oceans, and Space, University of New Hampshire, Durham 2Gulf Ecology Division, U.S. Environmental Protection Agency, National Health and Environmental Effects Research Laboratory, Gulf Breeze
3Present address: Department of Biology Sciences, University of Alaska Anchorage, Anchorage
Received September 1, 1998
Summary
The phylogenetic and physiological diversity of sulphate-reducing bacteria inhabiting a salt marsh rhizosphere were investigated. Sulphate-reducing bacteria were isolated from a salt marsh rhizosphere using enrichment cultures with electron donors thought to be prevalent in the rhizosphere of Spartina alterniflora. The relationship between phylogeny and nutritional characteristics of 10 strains was investigated. None of the isolates had 16S rRNA sequences identical to other delta subclass sulphate-reducers, sharing 85.3 to 98.1 % sequence similarity with 16S rRNA sequences of their respective closest relatives. Phylogenetic analysis placed two isolates, obtained with ethanol as an electron donor, within the Desulfovibrionaceae. Seven isolates, obtained with acetate, butyrate, propionate, or benzoate, were placed within the Desulfobacteriaceae. One isolate, obtained with butyrate, fell within the Desulfobulbus assemblage, which is currently considered part of the Desulfobacteriaceae family. However, due to the phylogenetic breadth and physiological traits of this group, we propose that it be considered a new family, the "Desulfobulbusaceae." The isolates utilised an array of electron donors similar to their closest relatives with a few exceptions. As a whole, the phylogenetic and physiological data indicate isolation of several sulphate-reducing bacteria which might be considered as new species and representative of new genera. Comparison of the Desulfobacteriaceae isolates' 16S rRNA sequences to environmental clones originating from the same study site revealed that none shared more than 86% sequence similarity. The results provide further insight into the diversity of sulphate-reducing bacteria inhabiting the salt marsh ecosystem, as well as supporting general trends in the phylogenetic coherence of physiological traits of delta Proteobacteria sulphate reducers.
Key words: sulphate-reducing bacteria - SRB - 16S rRNA phylogeny - salt-marsh - rhizosphere -Desulfobacteriaceae - Desulfovibrionaceae
Introduction
Until the 1970's, the dissimilatory sulphate-reducing bacteria (SRB) were thought to be comprised of a few species in two genera that were capable of utilising only lactate or pyruvate as energy sources (BARTON and TOMEI, 1995). However, SRB are now known to be both physiologically and phylogenetic ally diverse. Close to 100 electron donors for sulphate reduction have been described
'f Present address of corresponding author: Department of Biological Sciences, University of Massachusetts Lowell, 1 University Ave., Lowell, MA 01854, U.S.A. Telephone: (978) 934-4715; Fax: (978) 934-3044; e-mail: [email protected].
(HANSEN, 1993) and the capacity for dissimilatory sulphate reduction has been demonstrated in bacteria from three major eubacterial lines of descent and one genus within the Archaea domain (STACKEBRANDT et aI., 1995). To date, the majority of characterised SRB are members of the Gram-negative non-sporeforming mesophilic SRB within the delta subclass of the Proteobacteria, and it appears that this group is the most widely distributed in nature (WIDDEL and BAK, 1992). In addition, the delta subclass SRB have been extensively studied using comparative 16S rRNA sequence analysis and were found to form phylogenetic groups that are consistent with various physiological traits, particularly in the utilisation of electron

558 ]. N. ROONEy-VARGA et al.
donors. Due to this correlation, 165 rRNA-based analyses provide potentially powerful tools for inferring physiological traits of both cultivated and uncultivated SRB.
5RB are ubiquitous and are known to play ecologically important roles in such diverse habitats as freshwater ponds, oil production facilities, animal intestines, rice paddies, and estuarine and marine sediments (GIBSON, 1990; SMITH, 1993; WIDDEL and BAK, 1992). In particular, SRB have been found to exhibit high sulphate-reducing activity and to play an extremely important role in organic carbon remineralisation, below-ground geochemistry, and plant-microbe interactions in salt marsh environments (HINES et a!., 1989; HOWARTH, 1993). As the dominant terminal electron accepting process, it has been estimated that sulphate reduction accounts for up to 50% of organic carbon remineralisation in these ecosystems (HOWARTH and Hobbie, 1982).
Sulphate reduction rates in organic-rich salt marsh sediments are among the highest of any natural system (HOWARTH, 1993). Since plant root exudates and decomposing plant litter may provide a complex mixture of electron donors in salt marsh sediments, it is likely that diverse populations of 5RB exist therein. As discussed by ROONEy-VARGA et a!. (1997) and HINES et a!. (submitted), the application of 165 rRNA probes and sequence comparisons to the study of 5RB community structure in this ecosystem has suggested that as yet undescribed species are present in significant numbers. In the current study, physiological and phylogenetic analyses of 5RB isolated from a salt marsh sediment were undertaken in order to: (i) compare the phylogenetic diversity of cultivated isolates to 'molecular isolates' (i.e., 16S rRNA of uncultivated organisms) from the same study site (ROONEY-VARGA et a!. 1997); (ii) investigate the phylogenetic and nutritional diversity of cultivable SRB in a salt marsh sediment and rhizosphere of Spartina alterniflora, and (iii) further investigate the correlation between 165 rRNA phylogeny and nutritional characteristics of delta Proteobacteria SRB and the application of the currently available probes for 5RB (Devereux et a!., 1992).
Materials and methods
Source of inocula and techniques used to obtain isolates: Isolates were obtained from dilution series prepared with rhizosphere cores taken from Chapman's Marsh in the Great Bray region of New Hampshire (HINES et aI., 1989) in August and October 1993 and April 1994 (HINES et aI., submitted). The medium described by WIDDEL and BAK (1992) was used, except that the gas phase was 90% N2 with 10% CO2 and the medium was prepared in serum-stoppered bottles or tubes using anaerobic techniques as previously described (SHARAK GENTHNER et aI., 1989). To prevent precipitation during autoclaving, concentrated stocks of CaCI2 . 2 H20, MgCI2 . 6 H20, and NaCI were prepared as separate, sterile anaerobic solutions under 100% N2 and were added to individual bottles or tubes of sterile, cooled solution containing the remaining salts, trace metals, and sodium bicarbonate. The salinity of the medium was adjusted to that of the overlying marsh water at the time of sampling (August 1993, 29 ppt; October 1993,26 ppt; April 1994, 8 ppt) by modifying the amount of NaCI and MgCI . 6 H20 added in
appropriate ratios (WIDDEL and PFENNIG, 1981). Before inoculation, the electron donor and filter-sterilised vitamin solution were added, and the medium was reduced with sodium sulphide. Each dilution set contained one of the following electron donors: acetate, 20 mM; ethanol, 20 mM; benzoate, 5 mM; butyrate, 10 mM; malate, 10 mM; and propionate, 10 mM. All incubations were at 20 DC.
A most probable number (MPN) dilution series (RODINA, 1972) (10-2 to 10-9 ) was prepared from each sample with each electron donor. Tubes were observed for growth, and active dilutions were considered those that showed both visible growth and reduction of sulphate as measured by ion chromatography (SHARAK GENTHNER et aI., 1994). The 10-5 through 10-7 dilutions were inoculated into triplicate anaerobic roll tubes of the same composition as the initial MPN (SHARAK GENTHNER et aI., 1981) and were incubated at 20 DC until well-formed colonies became visible. Colonies were picked with a sterile needle (1.5", 20 g) attached to a 1 ml glass syringe that had been flushed with NzlC02 (90:10). The colony was immediately transferred into a bottle of fresh liquid medium. Cultures positive for growth, i.e., increase in optimal density at 600 nm and reduction of sulphate, were examined by phase-contrast microscopy. Pure cultures were maintained in liquid medium of the same composition as the MPN medium.
Restriction fragment length polymorphism (RFLP) analysis: Prior to characterisation of the isolates, comparison of restriction digest patterns of their 16S rRNA gene (16S rDNA) and those of previously described pure cultures was undertaken. One ml of turbid anaerobic culture was collected, centrifuged 5 min at 8,000xg, decanted and resuspended in 500 pi of PBS (145 mM NaCI, 100 mM NaP04, pH 7.4). This wash step was repeated and the cells were resuspended in 500 pi of O.lxTE (1 mM Tris, 0.1 mM EDTA, pH 8.0). Cells were subjected to three freeze-thaw cycles at -70 DC and 65 DC.
Primers fD1 and rP2 (WEISBURG et aI., 1991) were used to amplify near-complete 16S rRNA gene (16S rDNA) sequences from celllysates. PCR mixtures contained 50 mM KCI, 10 mM Tris-CI pH 8.3, 3 mM MgCI2, 200 pM each dNTP (dATP, dCTP, dGTP, dTTP), 0.2 pM each primer, and 3-5 pi cell lysate in a total volume of 100 pI. The 'hot-start' method was used by heating the PCR mixture to 94 DC for 2 min, and then adding 2 U Taq DNA polymerase to each reaction mixture. The reaction mixtures were then subjected to 35 cycles, each consisting of 1 min at 94 DC, 0.5 min at 55 DC, and 2 min at 72 DC, followed by 5 min at 72 DC, using a Perkin Elmer Thermal Cycler (Perkin Elmer-Cetus, Norwalk, CT). 5 pi of each PCR product was analysed by electrophoresis in 1 % agarose using standard methods (SAMBROOK et aI., 1989). The remaining 95 pi of PCR product was ethanol precipitated (SAMBROOK et ai, 1989), washed with an Ultra free MC filter (10,000 NMWL; Millipore Inc., Bedford, MA), and resuspended in 20 pi distilled H 20. 10 pi of each PCR product was digested separately with the tetrameric endonucleases Mspl, Hhal, and HinfI (SAMBROOK et aI., 1989). The restriction fragments were resolved by gel electrophoresis in 2.5% MetaPhor agarose (FMC Bioproducts, Rockland ME) at 12.7 V!cm; 2% SynergeJTM-0.8% agarose at 4.5 V!cm, or 1.7% agarose in 1xTBE (90 mM Tris-borate, 20 mM EDTA, pH 8.0). pGem and PhI X 174 HinfI molecular weight markers (Promega Corp., Madison, WI) were included on each gel. The 0.5xTBE running buffer was chilled to 14 DC with a Bio-Rad mini chilled and recirculated using a Bio-Rad recirculating pump. The gels were stained with ethidium bromide and photographed under UV transillumination (SAMBROOK et aI., 1989). Fragment sizes were estimated using the DIGIGEL software availabe with the DNASTAR programs (DNASTAR, Inc., Madison, WI). The numbers and sizes of the bands obtained were used to place isolates into RFLP categories.

16S rDNA analysis: Sequences of near-complete 16S rDNA from representatives of RFLP categories were obtained directly from PCR products or from inserts cloned into plasmid pNoTAff7 (Five Prime Three Prime, Inc., Boulder, CO) using a PRISM Ready Reaction Dye Deoxy Terminator Cycle Sequencing Kit (Perkin-Elmer Cetus) and an ABI 373A automated sequencer (Applied Biosystems, Foster City, CAl as previously described (ROONEY-VARGA et al., 1997). Primers that anneal to conserved regions of 16S rDNA genes were used in sequencing reactions. These included primers fDl (WEISBURG et al., 1991), 926F (BRITSCHGI and FALLON, 1994), 1115F (DORSCH and STACKEBRANDT, 1992), 357R (DORSCH and STACKEBRANDT, 1992), 536R (DEVEREUX et al., 1989), and rP2 (WEISBURG et al., 1991). In addition, primers M13-20, and M13 reverse (Stratagene, Inc., La Jolla, CAl were used for sequencing cloned 16S rDNA (i .e., BG6, BG8, BG18, BG23, BG33, and BG74). The sequences were deposited in GenBank under accession numbers U85468 through U85477.
The newly determined sequences were aligned with 16S rRNA sequences of other delta subclass Proteobacteria sulphate-reducing bacteria and E. coli. Unambiguously aligned base positions were used to calculate Jukes-Cantor distances and construct phylogenetic trees with the maximum parsimony, neighbour-joining, and least-squares methods available in the phylogenetic analysis package PHYLIP 3.57 (FELSENSTEIN, 1989). For maximum parsimony and neighbour-joining trees, 100 bootstrapped data sets were analysed.
Electron donor utilisation by new SRB strains: The ability of the isolated SRB strains to use hydrogen and eleven different organic compounds as electron donors for sulphate reduction was determined in the medium described above, modified by the addition of 0.02 % yeast extract to provide additional nutritional factors. Electron donors were added from concentrated (100x), anoxic stock solutions (pH 7.0, 100% N2) to final concentrations of 20 mM acetate, ethanol, fumarate, lactate, malate, pyruvate or succinate; 10 mM butyrate or propionate; 80 mM formate; or 2.5 mM benzoate. Medium containing 2x bicarbonate (final concentration 26 mM) and a gas phase of 80:20 N/C02 was used with H2 as an electron donor. The gas phase was flushed with Hz/C02 (90:10) for 2 min and brought to 2 atmospheres before reducing and inoculating this medium. Cultures containing H2 were shaken during incubation. Growth was scored visually and sulphate reduction was determined by ion chromatography as previously described (SHARAK GENTHNER et aI., 1994).
Results and discussion
Isolation and initial screening of SRB strains
Sixty-one isolates were obtained from the salt marsh sediment samples following the dilution series enrichments and colony purifications with roll tubes. Electron donors used for isolations represented compounds that are present in the S. alterni{lora rhizosphere (i.e., malate, ethanol, and acetate) (HINES et al., 1994; MENDELSSOHN and McKEE, 1987; NEDWELL and ABRAM, 1979; SMITH and AP REES, 1979), as well as common products of fermentation that are know to be utilised by SRB (WIDDEL and BAK, 1992). RFLP analyses provided a rapid means to assess the purity of each culture by comparing the sum of the lengths of the restriction fragments generated from each culture to the expected length of 16S rDNA (i.e., about 1,500 bp). The potential limitations of this ap-
Salt Marsh SRB Diversity 559
Table 1. Sampling dates, electron donors, and salinities used for enrichment and isolation of sulphate-reducing bacteria.
Isolate Sampling Electron Salinity Number of strains date donor (ppt) in OTU group
BG6 Oct. 1993 ethanol 26 2 BG8 Oct. 1993 propionate 26 6 BG14 Oct. 1993 butyrate 26 2 BG18 Aug. 1993 butyrate 28 8 BG23 Aug. 1993 butyrate 28 7 BG25 Aug. 1993 butyrate 28 1 BG33 Oct. 1993 benzoate 26 2 BG45 April 1994 malate 8 9 BG50 April 1994 ethanol 8 16 BG72 April 1994 acetate 8 5 BG74 April 1994 benzoate 8 3
proach include difficulty in detecting contamination with a closely related strain and difficulty in distinguishing between different 16S rRNA sequences from the same strain. However, we found that RFLP analysis with the three enzymes used here is sufficient to distinguish between 16S rRNA genes that differ by more than 5% and, after analysing about 80 isolates with this approach, we have not observed more than one RFLP pattern per isolate. In addition, comparison of 16S rDNA RFLP patterns was useful for sorting strains into operational taxonomic units (OTUs). The RFLP analysis revealed four unique OTUs among strains isolated on butyrate, 2 OTUs each among strains isolated on benzoate and ethanol, and one OTU each among strains isolated on propionate, malate, and acetate (Table 1). Comparison of RFLPs from the isolates to those of eighteen SRB reference strains showed that all but one of the isolate OTUs differed from the analysed SRB reference strains (Table 2). Strain BG45, obtained from a culture enriched with malate, had an RFLP pattern identical to that of Desulfovibrio desulfuricans subsp. aesturii.
Correlation between phylogenetic and physiological characteristics
Analysis of nearly complete 16S rRNA gene sequences from the ten new SRB isolates revealed that they were all members of the delta subclass of the Proteobacteria and were closely related to other known SRB within this group (Fig. 1 and 2). As mentioned above, two distinct lineages of SRB are known to exist within this group, the first being defined by the family Desulfovibrionaceae (DEVEREUX et al., 1990) and the second by the proposed family Desulfobacteriaceae (WIDDEL and BAK, 1992). All the SRB isolates in the current study appear to be members of these two lineages (Fig. 1 and 2). This result lends support to the hypothesis that the Gram-negative, nonsporeforming, mesophilic SRB are a phylogenetically coherent group and is consistent with the general observation that most SRB isolated to date are members of this group (STACKEBRANDT et al., 1995).

Tab
le 2
. F
ragm
ent s
izes
(ba
se p
airs
) ob
tain
ed b
y en
donu
clea
se d
iges
tion
of 1
6S r
DN
A.
Spec
ies
or s
trai
n
Des
ulfo
mic
robi
um n
orve
gicu
m
Des
ulfo
vibr
io g
igas
D
esul
fom
icro
bium
bac
ulat
us
Des
ulfo
vibr
io v
ulga
ris
subs
p. v
ulga
ris
Des
ulfo
vibr
io v
ulga
ris
Mar
burg
D
esul
fovi
brio
des
ulfu
rica
ns
Des
ulfo
vibr
io d
esul
furi
cans
sub
sp.
aest
uari
i D
esul
vofi
brio
pig
er
Des
ulfo
vibr
io s
alex
igen
s D
esul
fom
onil
e ti
edje
i D
esul
foar
culu
s ba
arsi
i D
esul
fobu
lbus
pro
pion
icus
D
esul
fobo
tulu
s sa
povo
rans
D
esul
foco
ccus
mu
ltiv
ora
ns
Des
ulfo
bact
eriu
m a
uto
tro
ph
icu
m
Des
ulfo
bact
eriu
m v
acu
ola
tum
D
esul
foba
cter
ium
nia
cini
D
esul
foba
cter
sp.
str
ain
3ac1
0 D
esul
foba
cter
sp.
str
ain
4ac1
1 D
esul
foba
cter
lat
us
BG
6 B
G8
BG
14
BG
15
BG
23
BG
25
BG
33
BG
45
BG
50
BG
72
BG
74
Res
tric
tion
Enz
yme
Use
d (a
)
Hin
fI
36
0,3
15
,24
5,1
85
, 14
5 (c
) 7
35
,24
5,2
20
,18
5
36
0,3
15
,24
5,1
85
, 14
5 (c
) 6
36
,24
5,2
22
,18
5,1
10
,10
5
63
6,2
45
,22
2,1
85
,11
0,1
05
6
36
,31
5,2
45
,18
5,1
10
7
18
,24
0,2
32
,19
0,1
10
6
36
,24
5,2
22
,18
5,1
10
,10
5
32
1,3
15
,24
9,2
32
,18
5
46
5,3
60
,24
9,2
15
,19
0
56
5,3
00
,29
0,1
85
4
65
,37
0,2
70
,22
5
81
0,2
60
,18
5,1
10
6
68
,22
6,1
85
,16
0,1
45
,14
0
66
8,2
62
,18
5,1
50
,10
0
66
8,3
11
,25
5,1
85
,11
0
66
8,3
11
,25
5,1
85
,11
0
80
0,3
65
,19
5,1
90
,16
0
80
0,3
65
,19
0
80
0,3
65
,21
0,2
00
, (1
90),
160
(d)
3
40
,33
6,2
49
,22
2,1
85
,11
0
66
0,3
65
,26
5,
110
56
0,2
25
,21
5,1
90
,17
0,1
50
560,311,255,19~ 1
10
80
0,3
60
,19
0,1
38
8
00
,36
0,1
90
,13
8
66
0,3
11
,22
5,1
85
,11
0
718,240,232,19~ 1
10
62
0,2
55
,22
2,1
90
,11
0,1
00
8
00
,19
0,1
60
(c)
51
0,3
00
,18
5,
155,
114
(c)
Ha
e II
I
25
8,2
10
,19
0,1
75
,14
5,1
40
,11
8 (
c)
ND
(b)
25
8,2
10
,19
0,1
75
,14
5,1
40
,11
8 (
c)
51
7,2
58
,22
5,2
10
,17
9,9
2,8
9
51
7,2
58
,22
5,2
10
,17
9,9
2,8
9
40
0,2
58
, 18
5, 1
75,
145
(c)
56
0,2
80
,20
5,1
79
,14
0
ND
410,258,215,140~)
45
0,2
65
,25
0,8
0 (c
) 305,250,170,168,138~)
51
7,2
65
,22
2,
150
(c)
25
0,1
68
, 16
1, 1
79,
118
(c)
51
7,2
65
,22
2,1
97
,15
0,1
45
5
20
,26
8,2
62
,17
9,1
50
5
10
,27
3,2
62
,17
9,1
50
5
10
,27
0,2
62
,17
9,1
50
5
10
,26
6,2
62
,17
5,1
46
,14
0
51
0,2
66
,26
2,1
75
,14
6,1
40
5
10
,26
6,2
62
,17
9,1
51
,14
8
27
0,2
60
,20
0,1
85
,17
9,1
35
,12
8
51
0,2
70
,22
2,1
44
,12
8 (
c)
44
0,3
15
,27
0,1
85
,17
9,1
40
5
10
,29
0,2
70
,17
9,1
51
5
10
,29
0,2
70
,17
9,1
51
,14
0
51
0,2
85
,27
0,1
79
,15
1
51
0,2
90
,27
0,1
79
,15
1
56
0,2
80
,20
5,1
79
,14
0
31
0,3
80
,22
2,1
85
,17
9,1
38
5
10
,29
0,2
70
,17
9,1
51
,14
0
67
6,2
80
,17
0,1
38
(c)
(a)
Pare
nthe
ses
indi
cate
ban
ds t
hat
stai
ned
wit
h et
hidi
um b
rom
ide
to a
less
er i
nten
sity
tha
n th
e ot
her
band
s.
(b)
Not
det
erm
ined
. (c
) L
ow m
olec
ular
wei
ght
band
s w
ere
obse
rved
but
thei
r si
zes
coul
d no
t be
acc
urat
ely
dete
rmin
ed.
(d)
Ban
ds w
ith
less
er i
nten
sity
wer
e ca
used
by
part
ial
dige
stio
n of
16S
rD
NA
fra
gmen
t.
Msp
I
43
5,3
45
,22
5,1
75
,14
0
ND
4
35
,34
5,2
25
,17
5,1
40
3
11
,25
5,1
98
,17
0,1
60
, 14
7 (c
) 3
11
,25
5,
198,
170
, 16
0, 1
47 (c
) 4
15
,34
5,1
75
,16
0,1
49
,13
8
50
0,4
40
,22
0,1
47
,13
2
ND
3
45
,32
0,2
25
,17
9,1
49
,13
8
190,
166
, 14
5, 1
18 (
c)
517,
(43
0),
32
5,1
75
,14
0,1
26
,12
6 (d
) 3
60
,29
0,1
81
,17
5,1
70
,14
5,1
30
51
7, (
430)
, 36
0, 2
22
,18
1,1
46
,12
5 (
d)
36
0,3
00
,22
2,1
81
,14
5,1
40
,12
0,1
10
3
60
,24
0,
151,
145
, 14
0 (c
) 3
60
,34
0,
181,
138
, 11
0 (c
) 3
60
,34
0,
181,
138
, 11
0(c)
3
60
,34
0,
(179
), 1
75 (
c)
36
0,3
40
,22
2,
(179
), 1
75
36
0,2
22
, (1
79),
175
, (1
60),
148
, 13
8, 1
10
33
8,3
20
,25
8,
175
(c)
66
0,3
38
,17
7,1
38
3
55
,33
8,3
00
,18
7,1
49
,11
2
34
5,3
11
,18
1,1
38
,11
2 (
c)
36
5,3
45
,30
0,1
81
,14
6,1
32
,11
0
36
5,3
45
,30
0,1
81
,14
7,1
32
,11
0
34
5,3
11
,23
0,1
81
,13
8,1
12
5
00
,44
0,2
20
,14
7,1
32
5
17
,42
7,1
45
,13
2,1
20
3
65
,34
0,
(18)
, 17
5, (
145)
, 13
5, 1
12
51
7,3
15
,17
5,1
40
, 12
6 (c
)
V,
G\
o ':"-' ~
:::0
o o Z
m ~ ;:: ;:
0 ~ ~ ~
69
2S
98
100
87
43
20 31
36
21
100
0.02
Myx
ococ
cuS)
Des
u
Des
uljo
vibr
io g
Des
uljo
vib1
Des
uljo
vibr
io
Des
u/ft
Des
uljo
vibr
.
Des
uljo
v
Des
ljov
ibl
BG
50
Des
uljo
vibr
io s
c
BG
6
Des
ulfn
v
D.
De,
Des
'anl
hus
Ijoh
alob
ium
rel
baen
se
bone
nsis
o gi
gas
fair
fiel
dens
is
vibr
io p
iger
o de
su/f
uric
ans
brio
vul
gari
s
o sp
, st
r. P
T-2
o lo
ngre
achi
i
Des
uljo
vibr
io s
p. s
tr.
MIT
87-
599
cale
doni
ensi
s
'exi
gens
brio
hal
ophi
lus
'ong
us
'esu
ljuri
cans
str.
EI
Agh
eila
Z
'ric
anus
'um
nor
vegi
cum
um b
acul
alus
ium
esc
ambi
ense
Esc
heri
chia
col
i
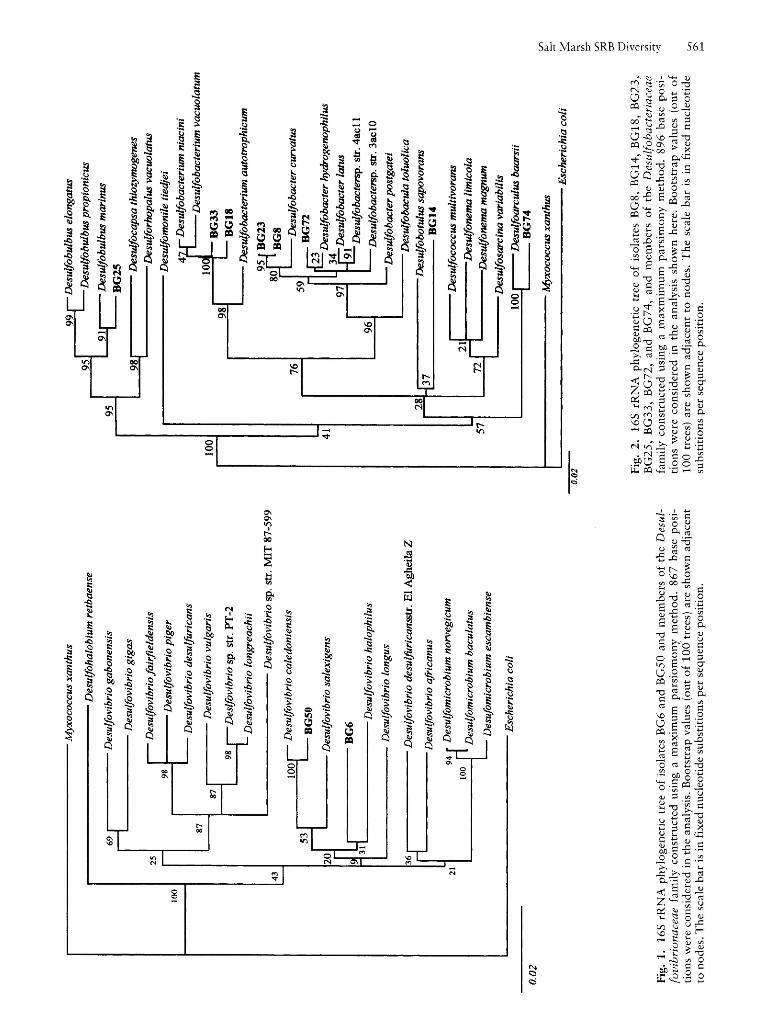
Fig
. 1.
1
65
rR
NA
ph
ylo
gen
etic
tre
e o
f is
olat
es B
G6
and
BG
50
an
d m
emb
ers
of
the
Des
ul
fovi
brio
nace
ae f
amil
y co
nst
ruct
ed u
sing
a m
axim
um
par
sio
mo
ny
met
ho
d.
86
7 b
ase
po
si
tio
ns
wer
e co
nsi
der
ed i
n th
e an
alys
is.
Bo
ots
trap
val
ues
(ou
t o
f 1
00
tree
s) a
re s
ho
wn
ad
jace
nt
to n
od
es.
Th
e sc
ale
bar
is i
n fi
xed
nu
cleo
tid
e su
bst
itio
ns
per
seq
uen
ce p
osi
tio
n.
100 41
95
95
Des
uljo
bulb
us e
long
atus
Des
uljo
bulb
us p
ropi
onic
us
981
Des
uljo
caps
a th
iozy
mog
enes
D
esul
jorh
opal
us v
acuo
laoo
.-------------
Des
u(fo
mon
ile
tieq
iei
28
98
76
59
97
Des
uljo
bact
eriu
m v
acuo
latu
m
BG
18
Des
ulfo
bact
eriu
m a
utot
roph
icum
23
Des
uljo
bact
er c
urva
tus
BG
72
-D
esul
foba
cter
hyd
roge
noph
ilus
34
'
91 D
esul
joba
cter
latu
s D
esul
foba
cter
sp.
str.
4ac
ll
96
Des
ulfo
bact
ersp
. st
r. 3
acl0
Des
ulfo
bact
er p
ostg
atei
'-
----
-Desu
lfo
ba
cu
la to
luol
ica
Jr- 3-7-------
Des
ulfo
botu
Jus
sapo
vora
ns
.... ---------
BG
14
21.
Des
ulfo
nem
a li
mic
ola
57
r D
esu(
foco
ccus
mul
tivo
rans
L-.
....
.:7 .;;:2.
... D
esul
fone
ma
mag
num
1
-_
__
_ D
esul
fosa
rcin
a va
riab
ilis
L _
__
__
__
-.:1.
;::O
.::,0
tr==
_
_ -=
Des
ulfo
arcu
lus
boar
sii
'----
BG
74
1--
----
----
----
Myx
ococ
cus
xant
hus
0.02
E
sche
rich
ia c
oli
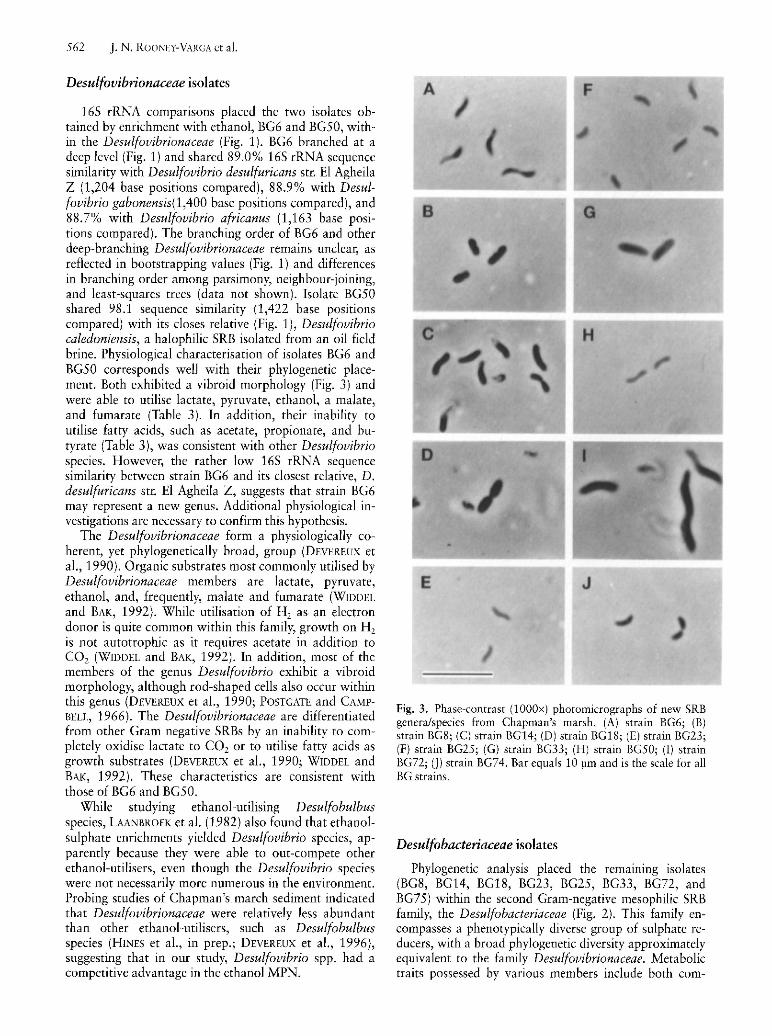
Fig
. 2
. 1
65
rR
NA
ph
ylo
gen
etic
tre
e o
f is
ola
tes
BG
8,
BG
14
, B
G1
8,
BG
23
, B
G2
5,
BG
33
, B
G7
2,
and
BG
74
, an
d m
emb
ers
of
the
Des
ulfo
bact
eria
ceae
fa
mil
y c
on
stru
cted
usi
ng
a m
axm
imu
m p
arsi
mo
ny
met
ho
d.
89
6 b
ase
po
si
tio
ns
wer
e co
nsi
der
ed i
n th
e an
aly
sis
sho
wn
her
e. B
oo
tstr
ap v
alu
es (
ou
t o
f 1
00
tre
es)
are
sho
wn
ad
jace
nt
to n
od
es.
Th
e sc
ale
bar
is
in f
ixed
nu
cleo
tid
e su
bst
itio
ns
per
seq
uen
ce p
osi
tio
n.
Vl ~
2:::
~ C/O ::r
Vl
:;0
O:l o :;: (1) '"' ~.
'<
Vo
~
,....
562 J. N. ROONEy-VARGA et al.
Desulfovibrionaceae isolates
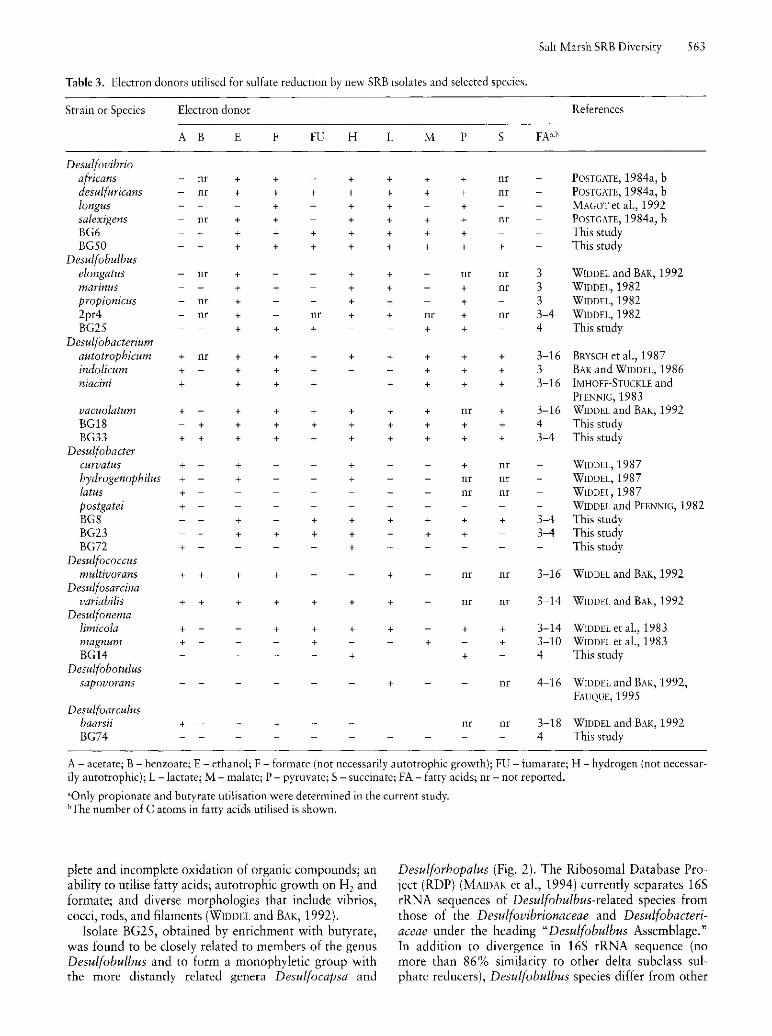
165 rRNA comparisons placed the two isolates obtained by enrichment with ethanol, BG6 and BG50, within the Desulfovibrionaceae (Fig. 1). BG6 branched at a deep level (Fig. 1) and shared 89.0% 165 rRNA sequence similarity with Desulfovibrio desulfuricans str. El Agheila Z (1,204 base positions compared), 88.9% with Desulfovibrio gabonensis(1,400 base positions compared), and 88.7% with Desulfovibrio africanus (1,163 base positions compared). The branching order of BG6 and other deep-branching Desulfovibrionaceae remains unclear, as reflected in bootstrapping values (Fig. 1) and differences in branching order among parsimony, neighbour-joining, and least-squares trees (data not shown). Isolate BG50 shared 98.1 sequence similarity (1,422 base positions compared) with its closes relative (Fig. 1), Desulfovibrio caledoniensis, a halophilic 5RB isolated from an oil field brine. Physiological characterisation of isolates BG6 and BG50 corresponds well with their phylogenetic placement. Both exhibited a vibroid morphology (Fig. 3) and were able to utilise lactate, pyruvate, ethanol, a malate, and fumarate (Table 3). In addition, their inability to utilise fatty acids, such as acetate, propionate, and butyrate (Table 3), was consistent with other Desulfovibrio species. However, the rather low 165 rRNA sequence similarity between strain BG6 and its closest relative, D. desulfuricans str. El Agheila Z, suggests that strain BG6 may represent a new genus. Additional physiological investigations are necessary to confirm this hypothesis.
The Desulfovibrionaceae form a physiologically coherent, yet phylogenetic ally broad, group (DEVEREUX et aI., 1990). Organic substrates most commonly utilised by Desulfovibrionaceae members are lactate, pyruvate, ethanol, and, frequently, malate and fumarate (WIDDEL and BAK, 1992). While utilisation of H2 as an electron donor is quite common within this family, growth on H2 is not autotrophic as it requires acetate in addition to CO2 (WIDDEL and BAK, 1992). In addition, most of the members of the genus Desulfovibrio exhibit a vibroid morphology, although rod-shaped cells also occur within this genus (DEVEREUX et aI., 1990; POSTGATE and CAMPBELL, 1966). The Desulfovibrionaceae are differentiated from other Gram negative 5RBs by an inability to completely oxidise lactate to CO2 or to utilise fatty acids as growth substrates (DEVEREUX et aI., 1990; WIDDEL and BAK, 1992). These characteristics are consistent with those of BG6 and BG50.
While studying ethanol-utilising Desulfobulbus species, LAANBROEK et al. (1982) also found that ethanolsulphate enrichments yielded Desulfovibrio species, apparently because they were able to out-compete other ethanol-utilisers, even though the Desulfovibrio species were not necessarily more numerous in the environment. Probing studies of Chapman's march sediment indicated that Desulfovibrionaceae were relatively less abundant than other ethanol-utilisers, such as Desulfobulbus species (HINES et aI., in prep.; DEVEREUX et aI., 1996), suggesting that in our study, Desulfovibrio spp. had a competitive advantage in the ethanol MPN.
A
B G
H
o I
E J
Fig. 3. Phase-contrast (lOOOx) photomicrographs of new SRB genera/species from Chapman's marsh. (A) strain BG6; (B) strain BG8; (C) strain BG14; (D) strain BG18; (E) strain BG23; (F) strain BG25; (G) strain BG33; (H) strain BG50; (I) strain BG72; (J) strain BG74. Bar equals 10 ]lm and is the scale for all BG strains.
Desulfobacteriaceae isolates
Phylogenetic analysis placed the remammg isolates (BG8, BG14, BG18, BG23, BG25, BG33, BG72, and BG75) within the second Gram-negative mesophilic 5RB family, the Desulfobacteriaceae (Fig. 2). This family encompasses a phenotypically diverse group of sulphate reducers, with a broad phylogenetic diversity approximately equivalent to the family Desulfovibrionaceae. Metabolic traits possessed by various members include both com-
Salt Marsh SRB Diversity 563
Table 3. Electron donors utilised for sulfate reduction by new SRB isolates and selected species.
Strain or Species Electron donor References
A B E F FU H L M P S FN,b
Desulfovibrio africans nr + + + + + + nr POSTGATE, 1984a, b desulfuricans nr + + + + + + + nr POSTGATE, 1984a, b longus + + + + MAGOT et aI" 1992 salexigens nr + + + + + + nr POSTGATE, 1984a, b BG6 + + + + + + + This study BG50 + + + + + + + + This study
Desulfobulbus elongatus nr + + + nr nr 3 WIDDEL and BAK, 1992 marinus + + + + + nr 3 WIDDEL, 1982 propionicus nr + + + + 3 WIDDEL, 1982 2pr4 nr + nr + + nr + nr 3-4 WIDDEL, 1982 BG25 + + + + + 4 This study
Desulfobacterium autotrophicum + nr + + + + + + + + 3-16 BRYSCH et aI" 1987 indolicum + + + + + + + 3 BAK and WIDDEL, 1986 niacini + + + + + + + + 3-16 IMHOFF-STUCKLE and
PFENNIG, 1983 vacuolatum + + + + + + + nr + 3-16 WIDDEL and BAK, 1992 BG18 + + + + + + + + + 4 This study BG33 + + + + + + + + + + 3-4 This study
Desulfobacter curvatus + + + + nr WIDDEL, 1987 hydrogenophilus + + + nr nr WIDDEL, 1987 latus + nr nr WIDDEL, 1987 postgatei + WIDDEL and PFENNIG, 1982 BG8 + + + + + + + 3-4 This study BG23 + + + + + + 3-4 This study BG72 + + This study
Desulfococcus multivorans + + + + + nr nr 3-16 WIDDEL and BAK, 1992
Desulfosarcina variabilis + + + + + + + nr nr 3-14 WIDDEL and BAK, 1992
Desulfonema limicola + + + + + + + 3-14 WID DEL et aI" 1983 magnum + + + + 3-10 WIDDEL et aI" 1983 BG14 + + 4 This study
Desulfobotulus sapovorans + nr 4-16 WIDDEL and BAK, 1992,
FAUQUE, 1995 Desulfoarculus
baarsii + + nr nr 3-18 WIDDEL and BAK, 1992 BG74 4 This study
A - acetate; B - benzoate; E - ethanol; F - formate (not necessarily autotrophic growth); FU - fumarate; H - hydrogen (not necessar-ily autotrophic); L -lactate; M - malate; P - pyruvate; S - succinate; FA - fatty acids; nr - not reported,
'Only propionate and butyrate utilisation were determined in the current study, bThe number of C atoms in fatty acids utilised is shown,
plete and incomplete oxidation of organic compounds; an ability to utilise fatty acids; autotrophic growth on H2 and formate; and diverse morphologies that include vibrios, cocci, rods, and filaments (WIDDEL and BAK, 1992).
Isolate BG25, obtained by enrichment with butyrate, was found to be closely related to members of the genus Desulfobulbus and to form a monophyletic group with the more distantly related genera Desulfocapsa and
Desulforhopalus (Fig. 2). The Ribosomal Database Project (RDP) (MAIDAK et aL, 1994) currently separates 16S rRNA sequences of Desulfobulbus-related species from those of the Desulfovibrionaceae and Desulfobacteriaceae under the heading "Desulfobulbus Assemblage." In addition to divergence in 16S rRNA sequence (no more than 86% similarity to other delta subclass sulphate reducers), Desulfobulbus species differ from other

564 J. N. ROONEy-VARGA et al.
sulphate reducers in the major menaquinone (COLLINS and WIDDEL, 1986), G+C content of genomic DNA (WIDDEL and Bak, 1992), and physiological traits (WIDDEL and BAK, 1992) described below. This assemblage is phylogenetic ally diverse as demonstrated by the deep branches shown in Fig. 2 and additional deep branches (not shown) of sequences cloned from environmental samples taken in northwest Florida (DEVEREUX and MUNDFROM, 1994) and a Danish fjord (TESKE et aI., 1996). Recent description of the new genera Desulfocapsa (JANSSEN et aI., 1996) and Desulforhopalus (ISAKSEN and TESKE, 1996) and characterisation of the isolate BG25 in this study lead to the emergence of this assemblage as a distinct group of delta subclass sulphate reducers which should be considered for recognition as a new bacterial family, the "Desulfobulbusaceae."
BG25 and its closest relative, Desulfobulbus marinus, shared 95.6% sequence similarity (1,208 base positions compared). These sequence data and the physiological characteristics of the strain suggest that BG25 should be considered a separate species within the genus Desulfobulbus. BG25 was similar to all Desulfobulbus species in an ability to utilise ethanol and pyruvate and an inability to oxidise acetate. It was able to utilise both butyrate and formate, which may be used by Desulfobulbus sp. strain 2pr4 and Desulfobulbus marinus, respectively (Table 3). However, BG25 does differ from other members of the genus Desulfobulbus in several phenotypic traits that were heretofore considered characteristic of the genus (Table 3). In particular, its inability to utilise propionate (Table 3) and its vibroid sigmoidal shape (Fig. 3) are not consistent with characteristic traits of the genus. In addition, BG25 is incapable of growth on two electron donors, hydrogen and lactate, that other characterised Desulfobulbus species utilise and, conversely, is capable of growth on two electron donors, fumarate and malate, that are not utilised by other Desulfobulbus species (Table 3) (WIDDEL, 1982).
Phylogenetic analysis of isolates BG18 and BG33 revealed that both fell within the bifurcation of Desulfobacterium autotrophicum and Desulfobacterium vacuolatum (Fig. 2). BG18 shared a 16S rRNA sequence similarity with its two closest relatives of 96.4% (1,102 base positions compared) with Desulfobacterium niacini and 98.0% (1,525 base positions compared) with BG33. Similarly, BG33 shared 96.6% (over 1,102 base positions) sequence similarity with D. niacini. The level of sequence divergence observed suggests that each isolate represents a novel species within the genus Desulfobacterium.
The genus Desulfobacterium was defined by BAK and WIDDEL (1986) to consist of non-sporeforming, completely oxidising sulphate-reducers that utilise a number of fatty acids, may grow autotrophically, and are widespread in marine sediments. Many members of the genus are nutritionally versatile. Cellular morphologies within this genus include the ovoid shape of the type strain (D. autotrophicum) (BRYSCH et aI., 1987), the spherical shape of the bacterium D. niacini (IMHOFFSTICKLE and PFENNIG, 1983), and curved cells. The morphologies of both BG18 and BG33 were ovoid (Fig. 3)
and were therefore consistent with those of other members of the genus. "Nicks and gaps" that probably represented gas vacuoles were also present in BGI8, making it the second species in this genus, after D. vacuolatum, that possesses gas vacuoles.
Electron donors that are commonly utilised by members of this genus include acetate, ethanol, formate, fumarate, malate, succinate, and fatty acids (Table 3). Autotrophic growth on hydrogen is also frequently observed within the genus (BRYSCH et aI., 1987; WIDDEL and BAK, 1992). Like its closest relatives, BG33 was quite nutritionally versatile and was capable of utilising acetate, ethanol, formate, fumarate, malate, and succinate, as well as benzoate, butyrate, propionate, and pyruvate (Table 3). Benzoate utilisation is not common among Desulfobacterium species, but other members are known to utilise aromatic compounds such as phenol and indole (BAK and WIDDEL, 1986; WIDDEL and BAK, 1992). BG18 only differed from BG33 in its inability to utilise acetate and propionate (Table 3). While its inability to utilize acetate was unusual among Desulfobacterium species, it should be noted that other members of the genus are capable of only slight growth on acetate (WIDDEL and BAK, 1992). Thus, 16S rRNA analysis, nutritional analysis, and morphology of isolates BG18 and BG33 permit placement of both strains within the genus Desulfobacterium.
Isolates BG8, BG23, and BG72 clustered within the genus Desulfobacter (Fig. 2), with isolates BG8 and BG23 most closely related to Desulfobacter curvatus, sharing sequence identifies of 96.6% and 96.4%, respectively (over 1,337 base positions compared). BG8 and BG23 were extremely closely related to each other, with 99.8% sequence similarity (over 1,337 base positions). BG72 was most closely related to Desulfobacter postgatei and Desulfobacter hydrogenophilus, sharing 97.1 % (over 1,371 base positions) and 97.0% (over 1,349 base positions) sequence similarity, respectively.
Morphologies exhibited by the three Desulfobacter isolates were straight thick rod-shaped cells for BG8 and vibroid to sigmoidal cells for BG23 and BG72 (Fig. 3). While these morphologies were consistent with other Desulfobacter species (which include oval-, rod-, and vibrio-shaped cells) (WIDDEL, 1987), preliminary nutritional characterisation revealed some marked differences between the isolates BG8 and BG23 and their Desulfobacter relatives. The characteristic traits of Desulfobacter species include their ability to utilise acetate (which is commonly used to enrich Desulfobacter species) more effectively than other SRB and their lack of nutritional versatility (Table 3) (WIDDEL and BAK, 1992). BG8 and BG23, however, did not utilise acetate, and did utilize several other electron donors such as fumarate, malate, and propionate that are not commonly used by Desulfobacter species (Table 3). BG23 also utilised butyrate and formate, which are not utilised by other characterised Desulfobacter species. BG72's substrate utilisation patterns were more similar to those of other Desulfobacter species and consisted of utilisation of acetate and hydrogen, but not any other electron donors tested

thus far (Table 3). Thus, based on 16S rRNA analysis and physiological traits, it appears that BG8 and BG23 represent two strain of a species, while BG72 represents a second species, all within the genus Desulfobacter.
Interestingly, BG8 and BG23 were isolated on propionate and butyrate, respectively, while other Desulfobacter species, including BG72, have been isolated on acetate (WIDDEL, 1987; WIDDEL and PFENNIG, 1982). While butyrate and propionate utilisation has been reported among Desulfobacterium, Desulfococcus, Desulfosarcina, Desulfonema, Desulfobotulus, Desulfoarculus, Desulfobulbus, and Desulforhabdus species (BAK and WIDDEL, 1986; BEEDER et aI., 1995; BRYSCH et aI., 1987; FAUQUE, 1995; IMHOFF-STUCKLE and PFENNIG, 1983; WIDDEL 1982; WIDDEL and BAK, 1992; WIDDEL et aI., 1983) (Table 3), butyrate has not been commonly used for enrichmentlisolation of SRB and propionate has only been used for isolation of Desulfobulbus species (WIDDEL, 1982; WID DEL and BAK, 1992). Perhaps it was use of these substrates that enabled the isolation of these novel phenotypes within the Desulfobacter genus.
Phylogenetic analysis of BG74 revealed that its closest relative was Desulfoarculus baarsii (formerly Desulfovibrio baarsii) (Fig. 2). BG74 and D. baarsii shared 95 .3% sequence similarity (1,350 base positions compared). BG74 likely represents the second species obtained in the genus Desulfoarculus. At the time of writing D. baarsii was the only known member of the genus Desulfoarculus and was considered a separate lineage within the Desulfobacteriaceae family (DEVEREUX et aI., 1989; STACKEBRANDT et aI., 1995). Desulfoarculus is characterised by its ability to carry out complete oxidation of organic carbon, its vibroid morphology, and its ability to utilise C1-C18 fatty acids but few other electron donors (WIDDEL and BAK, 1992).
D. baarsii and BG73 appear to be nutritionally limited - both are incapable of utilising benzoate, ethanol, fumarate, hydrogen, lactate, and malate (Table 3). In fact, of all the substrates tested so far, BG74 was only capable of growth on butyrate, the electron donor used for its isolation (Table 1). This trait is consistent with D. baarsii's ability to utilise fatty acids. Further characterisation of BG74 should include tests of its abilities to completely oxidise organic carbon and to utilise higher fatty acids which, as mentioned above, are considered characteristic of the genus Desulfoarculus.
Phylogenetic analysis of isolate BG 14 separated it from other members of the Desulfobacteriaceae family at what can be considered the genus level (Fig. 2). It shared 85.3% sequence similarity with its closest relative, Desulfobotulus sapovorans (1,215 base positions compared). The branching order of BG14, D. sapovorans, Desulfomonile tiedjei, D. baarsii, and the DesulfobacterDesulfobacterium and Desulfococcus-Desulfosarcina lineages is unclear, as reflected in bootstrap values for phylogenetic trees constructed by maximum parsimony (Fig. 2) and neighbour-joining methods (data not shown). Phenotypic traits of BG14 include its capacity to utilise butyrate, hydrogen, and pyruvate, but no other electron donors tested (Table 3).
Salt Marsh SRB Diversity 565
Comparison of isolate sequences to environmental clones
Comparison of the Desulfobacteriaceae isolates' 16S rRNA sequences to 'molecular isolates' A01, 2B14, and 4D19 originating from the same study site (ROONEYVARGA et aI., 1997) revealed that none of the isolates shared more than about 86% (over 754 base positions compared) sequence similarity with the environmental clones. All three clones fell within the DesulfococcusDesulfosarcina-Desulfonema assemblage (ROONEYVARGA et aI., 1997); a group which did not include the isolates described here. The results from the two methods of surveying SRB phylogenetic diversity (i.e., direct retrieval of 16S rRNA gene fragments and 16S rRNA analysis of novel isolates) provide further evidence of the great diversity of SRB inhabiting the salt marsh sediment. The fact that phylotype A01, which was shown to account for 7.5% of the total bacterial rRNA (ROONEYVARGA et aI., 1997), was not isolated is likely indicative of biases in isolation techniques. By now, discrepancies between molecular and cultivation based investigations of microbial diversity have been well-documented (e.g., HEAD et aI., 1998; PACE et aI., 1986) and there are many potential sources of bias in cultivation techniques. For example, an organism's nutrient or cofactor requirements, syntrophic relationships with other microorganism, or specific temperature, pH, or redox potential requirements may not be provided in the culture media used. In the current study, an additional source of cultivation bias may have been that the medium used did not provide a solid sub-stratum for microorganisms to adhere to. A01 was closely related to members of the genus Desulfonema (ROONEY-VARGA et aI., 1997), which is comprised of filamentous SRB that require a solid surface for growth (WIDDEL et aI., 1983). Therefore, if A01 is also a filamentous bacterium, it may also require a solid surface for healthy growth.
Evaluation of SRB probes
A series of oligonucleotide probes intended to specifically target various groups or genera within the Gramnegative mesophilic SRB were designed by DEVEREUX et a1. (1992) and are re-evaluated here against 16S rRNA sequences of the new SRB isolates. Several probes were found to contain a single mismatch with isolates that are members of the target group or genus. Probe 129 was designed to target the genus Desulfobacter and while it has no mismatches with sequences of Desulfobacter isolates BG8 and BG23, it does have a single mismatch with the sequence of Desulfobacter isolate BG72. This mismatch is fairly centrally located within the target site (Fig. 4) and sufficient to significantly destabilise the probe-target hybrid (STACKEBRANDT et aI., 1995; WARD et aI., 1992). A mismatch with probe 129 was also reported in the target sequence of Desulfobacter-related strain WN by VAN DER MAAREL et a1. (1996) . Probe 221, for the genus Desulfobacterium, has one mismatch with Desulfobacterium isolates BG18 and BG33. This mismatch, howev-

566 J. N. ROONEy-VARGA et al.
er, is very close to the 3' end of the target site and may not have a significant effect on probe-target hybridisation. In both cases, adding a mixed based position to the probe sequence allows for inclusion of the new isolates in
the probe target groups, while maintaining specificity for the group in question. Specifically, changing probe 129 to 5'-CAGGCTTGAAGSCAGATT-3' (where S is C or G) results in a probe that targets both previously charac-
Probe 687 (Desulfovibrionaceae)
Target AGGAGUGAAAUCCGUA
BG6 AGGAGUGAAAUCCGUA
Probe 804 (mixed Desulfobacteriaceae)
UCCACGCAGUAAACGUUG UCCACGCYGUAAACG!UG
terised Desulfobacter species and isolates BGS, BG23, and BGn. By using the RDP Check Probe utility (MAIDAK et aI., 1994), the modified probe was checked against all 16S rRNA sequences
BG8 AGAGGUGAAAUYCGUA UCCACGCAGUAAACGUUG in the unaligned RDP database BG14 AGCGGUGAAAU~GUA
BG18 AGAGGUGAAAUYCGUA
BG23 AGAGGUGAAAUYCGUA
BG25 AGAGGUGAAAUYCGUA
BG33 AGAGGUGAAAUYCGUA
BG50 AGGAGUGAAAUCCGUA
BG72 AGAGGUGAAAUYCGUA
BG74 AGAGGUGAAAUYCGUA
Probe 660 (Desulfobulbus)
Target CAGAGGGGAAAGUGGAAUUC
BG6 UGGAG!GGGUGG~GGAAUNC
BG8 GGGAG!GGA~AG!GGAAUUC
BG14 GGUAG!GGAAAG~GGAAUUC
BG18 GGGAG!GGAAAG~GAAUUC
BG23 GGGAG!GGA~AG!GGAAUUC
BG25 CAGAGGGGAAAGUGGAAUUC BG33 GGGAG!GGAAAG~GAAUUC
BG50 CgGAG!GGUUGG~GGAAUUC
BG72 GGGAG!GGAAAG~AAUUC
BG74 UGGAG!GGA~AGUGGAAUUC
Probe 221 (Desulfobacterium)
Target UUUGAAGAUGAGUCCGCGCA BG6 UUUCCAGAUGAGUCCGCGUC
BG8 UUUGGGGAUGAGUUUGCGYA BG14 !U~G~AG~UGAGCUUGCGUC
BG18 UUUGAAGAUGAGUCCGCG~A
BG23 UUUGGGGAUGAGUUUGCGYA BG25 CAUG~AGA~~UCYGCGYA
BG33 UUUGAAGAUGAGUCCGCG~A
BG50 ~UUUUGGAUGAGUCCGCGUC
BG72 UUUGGGGAUGAGUUUGCGYA BG74 CCCUUAGA~GAG~CCGCGUC
UCCAYGCAGUAAACGUUG UCCACGCAGUAAACGUUG UCCACGCAGUAAACGUUG UCCACGCCGUAAACG!UG UCCACGCAGUAAACGUUG UCCACGCYGUAAACG!UG UCCACGCAGUAAACGUUG UCCACGCCGUAAACG~UG
Probe 129 (Desulfobacter)
AAUCUGCCUUCAAGCCUG AAUCUCCCUGGAAAUUCG AAUCUGCCUUCAAGCCUG AAUCUGYCU~C~AAUC~G
AAUCU!CCUUCAAAUCgG AAUCUGCCUUCAAGCCUG AA~CU!CCU~CAYGUUUG
AAUCU!CCUUCAAAUCgG AAUCUGCCCUGAAGAUCG AAUCUG~UUCAAGCCUG
AAUCU!CCUAAA~UACG
Probe 814 (Desulfococcus-Desulfosarana-
Desulfobotulus) AAACGUUGAUCACUAGGU AAACG!UGGAUGCUAGGU AAACGUUGUACACU~GGU
AAACGUUGAUYACUAGGU AAACGUUGUAUGCUAGGU AAACGUUGUACACU~GGU
AAACG!UGUCAACUAG!U AAACGUUGUAUGCUAGGU AAACG!UGGAUAYUAGGU AAACGUUGUACACU~GGU
AAACG~UGUCCACUAGGU
Fig. 4. Comparison of probe 687,804,660,129,221, and 814 target sites with aligned 16S rRNA sequences of SRB isolates. Mismatches with the probe target site are shown in boldtype and underlined.
and found to be specific for Desulfobacter species. Similarly, probe 221 could be modified to 5'-TSCGCGGACTCATCTTCAAA-3', and once again, the CheckProbe utility was used to show that this modified probe targeted characterised Desulfobacterium and new Desulfobac-terium isolates to the exclusion of other 16S rRNA molecules.
Probe S14, designed to target the Desulfococcus-Desulfosarcina-Desulfobotulus assemblage contains one centrally located mismatch with BG14. However, while this assemblage constitutes BG14's closest relatives, as discussed above, BG14 appeared to represent a separate lineage not included in the group originally intended to be targeted by probe S14. Probe S04 was designed to target all members of the Desulfobacteriaceae family except Desulfobulbus and Desulfoarculus. Isolates BGS, BG18, BG23, BG33, and BGn, all members of this group, contained the target sequence. Isolate BG14 and Desulfoarculus isolate BG74 each had single mismatches with 804 that were centrally located. The former finding may be lend support to the placement of BG 14 in a separate lineage, while the latter is consistent with expected specificity of probe S04. As previously described (DEVEREUX et aI., 1992), the target site for probe S04 is more conserved than sites targeted by probes 129 and 221. Probe 804 is therefore expected to detect the rRNAs of bacteria with mismatches to the more specific probes.
The target sequence for the Desulfobulbus probe 660 was found in Desulfobulbus isolate BG25, as was the target sequence for the

Desulfovibrionaceae probe 687 in Desulfovibrionaceae isolates BG6 and BG50 (Fig. 4). It should be noted that the latter probe was recently found to also target members of the Geobacteraceae family (LONERGAN et al., 1996) and is no longer considered specific for its originally intended targets.
Conclusions
Using various electron donors thought to be prevalent in their native habitat, sixty-one SRB strains were isolated from a salt marsh sediment. Ten of these were chosen for further analysis based on unique RFLPs generated by digesting their 16S rRNA genes with tetrameric restriction enzymes. Analysis of near-complete 16S rRNA gene sequences from these ten isolates placed all the isolates within the delta Proteobacteria SRB. Isolates BG6 and BG50 were both members of the family Desulfovibrionaceae, with BG6 possibly representing a novel genus and BG50 representing a novel species within the genus Desulfovibrio. Isolates that were members of the Desulfobacteriaceae family were distributed among the genera Desulfobacter (BG8, BG23, and BG72), Desulfobacterium (BG18 and BG33), Desulfoarculus (BG74) and a novel genus represented by isolate BG14. Based on 16S rRNA analysis, all isolates were distinct from characterised Desulfobacteriaceae at the species or genus levels. One isolate (BG25) was placed within the Desulfobulbus assemblage, a group that should be considered for recognition as a new family, the "Desulfobulbusaceae." With the exception of several traits of BG25, BG8, and BG23, physiological characteristics of the isolates corresponded well with those of their closest relatives. None of the isolates shared more than 86% sequence similarity with environmental clones obtained from the same study site (ROONEY-VARGA et al., 1997), thereby providing further evidence of the diversity of SRB inhabiting salt marsh sediments. In addition, evaluation of currently available probes for SRB (DEVEREUX et al., 1992) against the isolates' 16S rRNA sequences revealed that, with the addition of mixed base positions to
probes 129 and 221, SRB-directed probes could still be used against their originally intended targets.
Acknowledgements This work was supported by the u.s. Environmental Protec
tion Agency Cooperative Agreement #CR-820062 and the National Science Foundation Ecology Program grant #DEB-9520272. Use of trade names does not imply endorsement by the U.S. E.P.A.
References
BAK, F. and WIDDEL, F.: Anaerobic degradation of indolic compounds by sulfate-reducing enrichment cultures and description of Desulfobacterium indolicum gen. nov., sp. nov. Arch. Microbio!. 146, 170-176 (1986).
BARTON, L. L. and TOMEI, F. A.: Characteristics and Activities of Sulfate-Reducing Bacteria, pp. 1-32. In: Sulfate-Reducing Bacteria (L. L. BARTON, ed.). Plenum Press, New York (1995).
BEEDER, J., TORSVIK, T and LIEN, T: Thermodesulforhabdus norvegicus gen. nov., sp. nov., a novel thermophilic sulfate-
Salt Marsh SRB Diversity 567
reducing bacterium from oil field water. Arch. Microbiol. 164,331-336 (1995).
BRITSCHGI, T. B. and FALLON, R. D.: PCR-Amplification of mixed 16S rRNA genes from an anaerobic, cyanide-degrading consortium. FEMS Microbiol. Ecol. 13,225-231 (1994).
BRYSCH, K., SCHNEIDER, c., FUCHS, G. and WIDDEL, F.: Lithoautotrophic growth of sulfate-reducing bacteria, and description of Desulfobacterium autotrophicum gen. nov., sp. nov. Arch. Microbiol. 148,264-278 (1987).
COLLINS, M. D. and WIDDEL, F.: Respiratory qui nones of sulphate-reducing and sulphur-reducing bacteria: a systematic study. Syst. Appl. Microbiol. 8, 8-18 (1986).
DEVEREUS, R., DELANEY, M., WIDDEL, F. and STAHL, D. A.: Natural relationships among sulfate-reducing eubacteria . ]. Bacteriol. 171,6689-6695 (1989).
DEVEREUX, R., HE, S.-H., DOYLE, C. L., ORKLAND, S., STAHL, D. A., LEGALL, J. and WHITMAN, W. B.: Diversity and origin of Desulfovibrio species: Phylogenetic definition of a family. J. Bacteriol. 172,3609-3619 (1990).
DEVEREUX, R., HINES, M. E. and STAHL, D. A.: S cycling: Characterization of natural communities of sulfate-reducing bacteria by 16S rRNA sequence comparisons. Microb. Ecol. 32, 283-292 (1996).
DEVEREUX, R. , KANE, M. D., WINFREY, J. and STAHL, D. A.: Genus- and group-specific hybridization probes for determinative and environmental studies of sulfate-reducing bacteria. System. Appl. Microbiol. 15,601-609 (1992).
DEVEREUX, R. and MUNDFROM, G. W.: A phylogenetic tree of 16S rRNA sequences from sulfate-reducing bacteria in a sandy marine sediment. Appl. Environ. Microbiol. 60, 3437-3439 (1994).
DORSCH, M. and STACKEBRANDT, E.: Some modifications in the procedure of direct sequencing of PCR amplified 16S rDNA. J. Microbiol. Methods 16,271-279 (1992).
FAUQUE, G. D.: Ecology of Sulfate-Reducing Bacteria, pp. 217-242. In : Sulfate-Reducing Bacteria (L. L. BARTON, eds) New York, Plenum Press 1995.
FELSENSTEIN, J. : PHYLIP - phylogeny inference package. Cladistics 5, 164-166 (1989).
GIBSON, G. R.: Physiology and ecology of sulphate-reducing bacteria. J. Appl. Bacteriol. 69, 769-797 (1990).
HANSEN, T. A.: Carbon Metabolism of Sulfate-Reducing Bacteria, pp. 21-40. In: The Sulfate-Reducing Bacteria: Contemporary Perspectives (J. M. ODOM and R. SINGLETON, Jr., eds.) Springer-Verlag, New York (1993) .
HEAD, I. M., SAUNDERS, J. R. and PICKUP, R. W.: Microbial evolution, diversity, and ecology: A decade of ribosomal RNA analysis of uncultivated microorganisms. Microb. Eco!. 35; 1-21 (1998).
HINES, M. E., BANTA, G. T, GIBLIN, A. E., HOBBIE, ]. E. and TUGEL, ]. T.: Acetate concentrations and oxidation in salt marsh sediments. Limnol. Oceanogr. 39, 140-148 (1994).
HINES, M. E., EVANS, R. S., SHARAK GENTHNER, B. R., WILLIS, S. G., FRIEDMAN, S. and DEVEREUX, R.: Molecular phylogenetic and biogeochemical studies of sulfate-reducing bacteria in the rhizosphere of Spartina alterniflora. (submitted to App!. Environ. Microbiol.) .
HINES, M. E., KNOLLMEYER, S. L. and TUGEL, J. B.: Sulfate reduction and other sedimentary biogeochemistry in a northern New England salt marsh. Limnol. Oceanogr. 34, 578-590 (1989).
HOWARTH, R. W.: Microbial processes in salt-marsh sediments, pp. 239-260. In: Aquatic Microbiology: an Ecological Approach (T. E. FORD, ed.) Blackwell Scientific Publications, Boston (1993).
HOWARTH, R. W. and HOBBIE, J. E.: The regulation of decomposition and heterotrophic microbial activity in salt marh soils: a review, pp. 103-127. In: Estuarine Comparisons (Y. S. KENNEDY, ed.) Academic Press, New York (1982).

568 J. N. ROONEy-VARGA et a!.
IMHOFF-STUCKLE, D. and PFENNIG, N.: Isolation and characterization of a nicotinic acid-degrading sulfate-reducing bacterium, Desulfococcus niacini sp. nov. Arch. Microbio!. 136, 194-198 (1983).
ISAAKSEN, M. E and TESKE, A.: Desulforhopalus vacuolatus gen. nov., sp., nov., a new moderately psychrophilic sulfate-reducing bacterium with gas vacuoles isolated from a temperate estuary. Arch. Microbio!. 166, 160-168 (1996).
JANSSEN, P. H., SCHUHMANN, A., BAK, E, and LiESACK, W.: Disproportion of inorganic sulfur compounds by the sulfate-reducing bacterium Desulfocapsa thiozymogenes gen. nov., sp. nov. Arch. Microbio!. 166, 184-192 (1996).
LAANBROEK, H. J., ABEE, T. and VOOGD, I. L.: Alcohol conversions by Desulfobulbus propionicus Lindhorst in the presence and absence of sulfate and hydrogen. Arch. Microbio!. 133,178-184 (1982).
LANE, D., PACE, B., OLSEN, G. J., STAHL, D. A., SOGIN, M. L. and PACE, N. R.: Rapid determination of 16S ribosomal RNA sequences for phylogenetic analyses. Proc. Nat!' Acad. Sci. USA 82, 6955-6959 (1985).
LONERGAN, D. J., JENTER, H. L., COATES, J. D., PHILLIPS, E. J. P., SCHMIDT, T. M. and LOVLEY, D. R.: Phylogenetic analysis of dissimilatory Fe(III)-reducing bacteria. J. Bacterio!' 178, 2402-2408 (1996).
MAGOT, M., CAUMEITE, P., DESPERRIER, J. M. and MATHERON, R.: Desulfovibrio longus sp. nov., a sulfate-reducing bacterium isolated from an oil-producing wei!. Int. j. Syst. Bacterio!. 42, 398-403 (1992).
MAIDAK, B. L., LARSEN, N., MCCAUGHEY, M. J., OVERBEEK, R., OLSEN, G. J., FOGEL, K., BLANDY, J. and WOESE, e. R.: The Ribosomal Database Project. Nucleic Acids Res. 22, 3485-3487 (1994).
MENDELSSOHN, I. A. and McKEE, K. L.: Root metabolic response of Spartina alternif/ora to hypoxia, pp. 239-353. In: Plant Life in Aquatic and Amphibious Habitats (R. M. M. CRAWFORD, ed.), British Eco!. Soc. Spec. Pub!. No.5 (1987).
NEDWELL, D. B. and ABRAM, J. W.: Relative influence of temperature and electron donor and electron acceptor concentrations on bacterial sulfate reduction in salt marsh sediment. Microb. Eco!. 5, 67-72 (1979).
PACE, N. R., STAHL, D. A., LANE, D. J. and OLSEN, G. J.: The analysis of natural microbial populations by ribosomal RNA sequences, pp. 1-55. In: Advances in Microbial Ecology (K. e. MARSHALL, ed.) Plenum Press, New York (1986).
POSTGATE, J. R.: Genus Desulfovibrio, pp. 666-672. In: Bergey's manual of systematic bacteriology (N. R. KRIEG and j. G. HOLT, eds.) Williams and Wilkins, Baltimore (1984).
POSTGATE, J. R.: The Sulphate-Reducing Bacteria, 2nd ed., Cambridge University Press, Cambridge, UK (1984).
POSTGATE, J. R. and CAMPBELL, L. L.: Classification of Desulfovibrio species, the nonsporulating sulfate-reducing bacteria. Bacterio!' Rev. 30, 732-738 (1966).
RODINA, A. G.: pp. 177-180. In: Methods in Aquatic Microbiology. University Park Press Baltimore (1982).
ROONEy-VARGA, j. N., DEVEREUX, R., EVANS, R. A. and HINES, M. E.: Seasonal changes in the relative abundances of rRNA from uncultivated sufi ate-reducing bacteria in the rhizosphere of Spartina alternif/ora. App!. Environ. Microbio!. 63, 3895-3901 (1997).
SAMBROOK, J., FRITSCH, E. E and MANIATIS, T.: Molecular Cloning: A Laboratory Manual, 2 ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor (1989).
SHARAK GENTHNER, B. R., DAVIS, e. L. and BRYANT, M. P.: Features of rumen and sewage sludge strains of Eubacterium limosum, a methanol- and Hz-C02-utilizing species. App!. Environ. Microbio!. 42,12-19 (1981).
SHARAK GENTHNER, B. R., MUNDFROM, G. and DEVEREUX, R.: Characterization of Desulfomicrobium escambium sp. nov.
and proposal to assign Desulfovibrio desulfuricans strain Norway 4 to the genus Desulfomicrobium. Arch. Microbiol. 161,215-219 (1994).
SHARAK GENTHNER, B. R., PRICE, W. A. and PRITCHARD, P. H.: Anaerobic degradation of chI oro aromatic compounds in aquatic sediments under a variety of enrichment conditions. Appl. Environ. Microbiol. 55,1466-1471 (1989).
SMITH, A. M. and AP REES, T.: Pathways of carbohydrate fermentation in the roots of marsh plants. Planta 146,327-334 (1979).
SMITH, D. W.: Ecological actions of sulfate-reducing bacteria, pp. 161-188. In: The Sulfate-Reducing Bacteria: Contemporary Perspectives (J. M. ODOM and R. SINGLETON, Jr., eds.) Springer-Verlag, New York (1993).
STACKEBRANDT, E., STAHL, D. A. and DEVEREUX, R.: Taxonomic Relationships, pp. 49-87. in: Sulfate-Reducing Bacteria (L. L. BARTON, ed.) Plenum Press, New York (1995).
TESKE, A., WAWER, e., MUYZER, G. and RAMSING, N.: Distribution of sulfate-reducing bacteria in a stratified fjord (Mariager Fjord, Denmark) as evaluated by most-probablenumber counts and denaturing gradient gel electrophoresis of PCR-amplified ribosomal DNA fragments. Appl. Environ. Microbiol. 62, 1405-1415 (1996).
VAN DER MAAREL, M. J. E. e., JANSEN, M., HAANSTRA, R., MEIJER, W. G. and HANSEN, T. A.: Demethylation of dimethyl sulfonioproprionate to 3-s-methylmercaptopropionate by marine sulfate-reducing bacteria. App!. Environ. Microbiol. 62, 3978-3984 (1996).
WARD, D., BATESON, M., WELLER, R. and RUFF-ROBERTS, A.: Ribosomal RNA analysis of microorganisms as they occur in nature, pp. 219-286. In: Advances in Microbial Ecology (K. MARSHALL, eds.) Plenum Press, New York (1992).
WEISBURG, W. G., BARNS, S. M., PELLEITIER, D. A. and LANE, D. J.: 16S ribosomal DNA amplification for phylogenetic study. J. Bacteriol. 173,697-703 (1991).
WIDDEL, E: Studies on dissimilatory sulfate-reducing bacteria that decompose fatty acids. II. Incomplete oxidation of propionate by Desulfobulbus propionicus gen. nov., sp. nov. Arch. Microbio!. 131,360-365 (1982).
WID DEL, E: New types of acetate-oxidizing, sulfate-reducing Desulfabaeter species, D. hydrogenaphilus sp. nov., D. latus sp. nov., and D. eurvatus sp. nov. Arch. Microbio!. 148,286-291 (1987).
WIDDEL, E and BAK, E: Gram-Negative Mesophilic Sulfate-Reducing Bacteria, pp. 3352-3378. In: The Prokaryotes (A. BALOWS, H. G. TRUPER, M. DWORKIN, W. HARDER and K.-H. SCHLEIFER, eds.) 2nd, New York, Springer-Verlag 1992.
WIDDEL, E, KOHRING, G.-W. and MAYER, E: Studies on dissimilatory sulfate-reducing bacteria that decompose fatty acids III. Characterization of the filamentous Desulfonema limicola gen. nov. sp. nov. and Desulfonema magnum sp. nov. Arch. Microbio!. 134,286-294 (1983).
WID DEL, E and PFENNIG, N.: Studies on dissimilatory sulfate-reducing bacteria that decompose fatty acids. I. Isolation of new sulfate-reducing bacteria enriched with acetate from saline environments. Description of Desulfobacter postgatei gen. nov. sp. nov. Arch. Microbio!. 129,395-400 (1981).
WIDDEL, E and PFENNIG, N.: Studies on dissimilatory sulfate-reduzing bacteria that decompose fatty acids. I. Isolation of new sulfate-reducing bacteria enriched with acetate from saline environments. Description of Desulfobacter postgatei gen. nov. sp. nov. Arch. Microbio!. 129,395-400 (1982).
Corresponding author: J. N. ROONEy-VARGA, Department of Biological Sciences, University of MAssachusetts Lowell, 1 University Ave., Lowell, MA 01854,. U.S.A. Telephone: (978) 934-4715; Fax: (978) 934-3044; e-mail: juliette_RooneyVarga@um!.edu.
Copyright © 2022 FDOKUMEN




















