Cancer mortality in German male workers exposed to phenoxy herbicides and dioxins

Photosynthetic Reaction Centers Embedded in Polyelectrolyte Multilayer as a Tool in the Determination of PSII Herbicides
Francesco Lopez1, Mauro Mennuni2, Mauro Giustini2, Marcello Giomini2,
Manuela Dezi3, Giovanni Venturoli3, Antonia Mallardi4, Gerardo Palazzo1
1 CSGI and Dipartimento di Chimica, Università di Bari, via Orabona 4, I-70126, Bari, Italy
2 Dipartimento di Chimica, Università “La Sapienza”, Roma, Italy
3 Dipartimento di Biologia and CNISM, Università di Bologna, Bologna, Italy
4 Istituto per i Processi Chimico-Fisici, CNR, Bari, Italy
AUTHOR EMAIL ADDRESS [email protected]
Abstract
The photosynthetic bacterial reaction center (RC) has been adsorbed onto a glass surface by alternating deposition with the cationic polimer poly(dimethyldiallylammonium chloride) (PDDA) obtaining as an end result an ordinate polyelectrolyte multilayer (PEM) where the protein retains its integrity and photoactivity over a period of several months. This polymer/RC assembly revealed to be extremely sensitive to the presence of herbicides belonging to the so called “PSII family”. The RC-herbicides interaction can be easily monitored since it alters the protein photoactivity, such variation resulting in an absorbance decrement at 870 nm. To this purpose, an instrumental setup has been developed based on a monochromatic light source and on a solid state detector connected to a DAQ acquisition board for the measurement storage onto a PC.
The system PEM/instrument has been tested on atrazine based herbicides. The bound herbicide can be easily removed from PEM, thus allowing the reuse of the same sample. Moreover, the system has been tested for sensitivity, reproducibility and reliability of the measured data. The results of these tests were so promising to encourage us to propose the PEM as an optical biosensor for herbicides.
1 Introduction Herbicides are widely used on a variety of crops for the
control of broadleaf weeds but can be highly toxic for human and animal health. Their wide use in agriculture has resulted often in the herbicide pollution of water and the level of herbicides allowed in drinking water is subject to regulation, at least in the industrialized countries. Different attempts have been made to introduce biological detection systems in order
to overcome the high cost of time consuming HPLC analysis [1, 2, 3]. Several immunoassays have been proposed, mainly ELISA and immunosensors [4, 5, 6]. However, antibodies can detect only one or few cross-reacting substances. To overcome this limit, the analytical use of Photosystem II (PSII) as a receptor protein in bioassays has been investigated [7], allowing the simultaneous detection of all PSII herbicides present in the water sample. The practical application of herbicide biosensors based on PSII preparations is, however, seriously hampered by their instability (in particular when illuminated) [7].
In this paper we propose a different approach, based on the use of purified photosynthetic bacterial reaction center (RC) of Rhodobacter Sphaeroides. This protein shows a high homology to PSII from plants and is inhibited by many of PSII herbicides active in plants [8]. The main advantage in the use of this protein is in its high stability against denaturation that makes it suitable for biosensing application.
The RC has been immobilized in multilayered ordered films obtained by alternate adsorption of layers made of poly(dimethyldiallylammonium chloride) (PDDA) and of protein on glass substrates. The herbicide binding to RC in this system has been investigated and an assay for herbicides, based on the response of the PDDA-RC multilayer to continuous illumination, has been tested. Finally, its employment in a simple optical herbicide biosensor has been proposed.
2 Structure and Function of RC The RC is a pigment-protein complex which in vivo spans
the bacterial intracytoplasmic membrane. It promotes the primary events of photosynthetic energy transduction by
1-4244-1245-5/07/$25.00 ©2007 IEEE
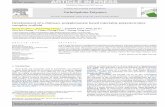
catalyzing a light-induced charge separation across the protein through a sequence of electron transfer events, involving cofactors held at fixed distances by the polypeptide scaffolding [9]. These cofactors are arranged along two almost symmetrical branches. However, the electron transfer is restricted to only one branch because of the structural engineering and specific environmental effects (see fig. 1).
Figure 1: RC cofactors and electron transfer processes between them after light excitation. See text for details. (Image from pdb structure 1pss).
Upon photon absorption, a bacteriochlorophyll special
pair (P), facing the periplasmic side of the intracytoplasmic membrane, acts as the primary electron donor by delivering, through a molecule of bacteriochlorophyll and a molecule of bacteriopheophytin, an electron to the primary acceptor (QA), a ubiquinone molecule bound at a site close to the opposite, cytoplasmic side of the complex. This primary charge separation, accomplished in about 200 ps, is stabilized by electron transfer from QA
- to a second ubiquinone molecule, bound at the QB site of the RC, which is located symmetrically to the QA site on the same cytoplasmic side of the membrane [9]. In vivo, the photoxidized donor, P+, is rapidly re-reduced by a soluble c-type cytochrome, so that a second charge separation can take place across the RC, leading to the double reduction and protonation of QB, which leaves the RC in its quinol state, QH2. In this reaction two protons are taken up from the cytoplasmic side of the membrane so that the RC catalyzes the first step in the generation of the transmembrane electrochemical potential for protons which is the driving force for ATP synthesis [10]. In vitro, when no physiological or artificial electron donor is available to re-reduce flash generated P+, the electron on QB
- recombines with the hole on P+, restoring the initial ground state of the RC. A striking functional homology exists between the electron acceptors of the bacterial RC and of photosystem II in plants [8]. In both systems, the two acceptor quinones (QA and QB), act in series and function as a two electron gate which passes reducing equivalents out of the RC only in pairs, i.e. when QH2 is formed at the QB site. This homology is reflected by similar inhibitor sensitivities: many economically important herbicides, largely used in agriculture, known to act at PS II, are effective in the bacterial RC, where they compete with quinone for binding at the QB site. In view of these properties and of the crystallographic information available, the bacterial RC has become over the
last decades the reference model in studying several features of the more complex PS II system.
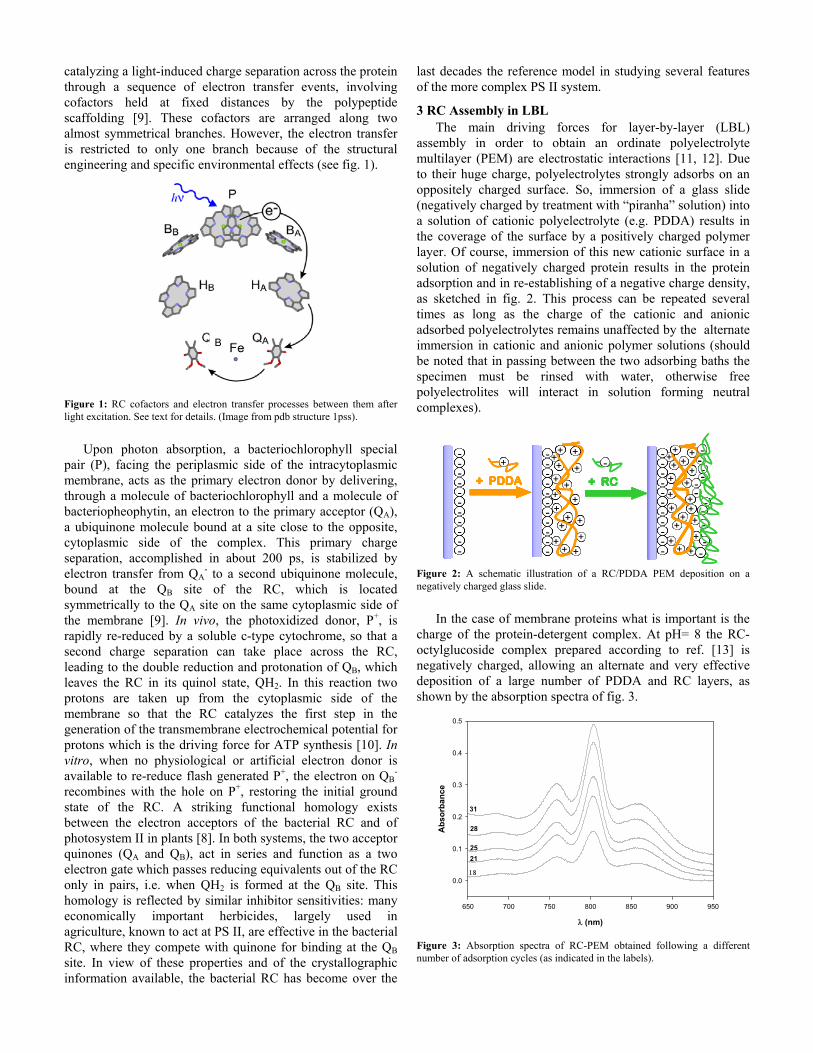
3 RC Assembly in LBL The main driving forces for layer-by-layer (LBL)
assembly in order to obtain an ordinate polyelectrolyte multilayer (PEM) are electrostatic interactions [11, 12]. Due to their huge charge, polyelectrolytes strongly adsorbs on an oppositely charged surface. So, immersion of a glass slide (negatively charged by treatment with “piranha” solution) into a solution of cationic polyelectrolyte (e.g. PDDA) results in the coverage of the surface by a positively charged polymer layer. Of course, immersion of this new cationic surface in a solution of negatively charged protein results in the protein adsorption and in re-establishing of a negative charge density, as sketched in fig. 2. This process can be repeated several times as long as the charge of the cationic and anionic adsorbed polyelectrolytes remains unaffected by the alternate immersion in cationic and anionic polymer solutions (should be noted that in passing between the two adsorbing baths the specimen must be rinsed with water, otherwise free polyelectrolites will interact in solution forming neutral complexes).
- +--
-
-
-
-
-
-
-
-
+
+ +++
++
+++
+
+
+
+
---
-
-
-
-
-
-
-
-
+ PDDA
+
+
+ RC
+ --- ++----
--
--
--
--
--
--
--
--
++
++ ++++++
++++
++++++
++
++
++
++
------
--
--
--
--
--
--
--
--
+ PDDA
++
++
+ RC
++ -- +
+
+ +++
+
+
+++
+
+
+
+
---
-
-
-
-
--
-
-
+
+
-
-
-
-
-
-
-+
-
+++
++
++ ++++++
++
++
++++++
++
++
++
++
------
--
--
--
--
----
--
--
++
++
--
-
--
--
--
--
-
---
--++
--
++- +--
-
-
-
-
-
-
-
-
+
+ +++
++
+++
+
+
+
+
---
-
-
-
-
-
-
-
-
+ PDDA
+
+
+ RC
+ --- ++----
--
--
--
--
--
--
--
--
++
++ ++++++
++++
++++++
++
++
++
++
------
--
--
--
--
--
--
--
--
+ PDDA
++
++
+ RC
++ -- +
+
+ +++
+
+
+++
+
+
+
+
---
-
-
-
-
--
-
-
+
+
-
-
-
-
-
-
-+
-
+++
++
++ ++++++
++
++
++++++
++
++
++
++
------
--
--
--
--
----
--
--
++
++
--
-
--
--
--
--
-
---
--++
--
++
Figure 2: A schematic illustration of a RC/PDDA PEM deposition on a negatively charged glass slide.
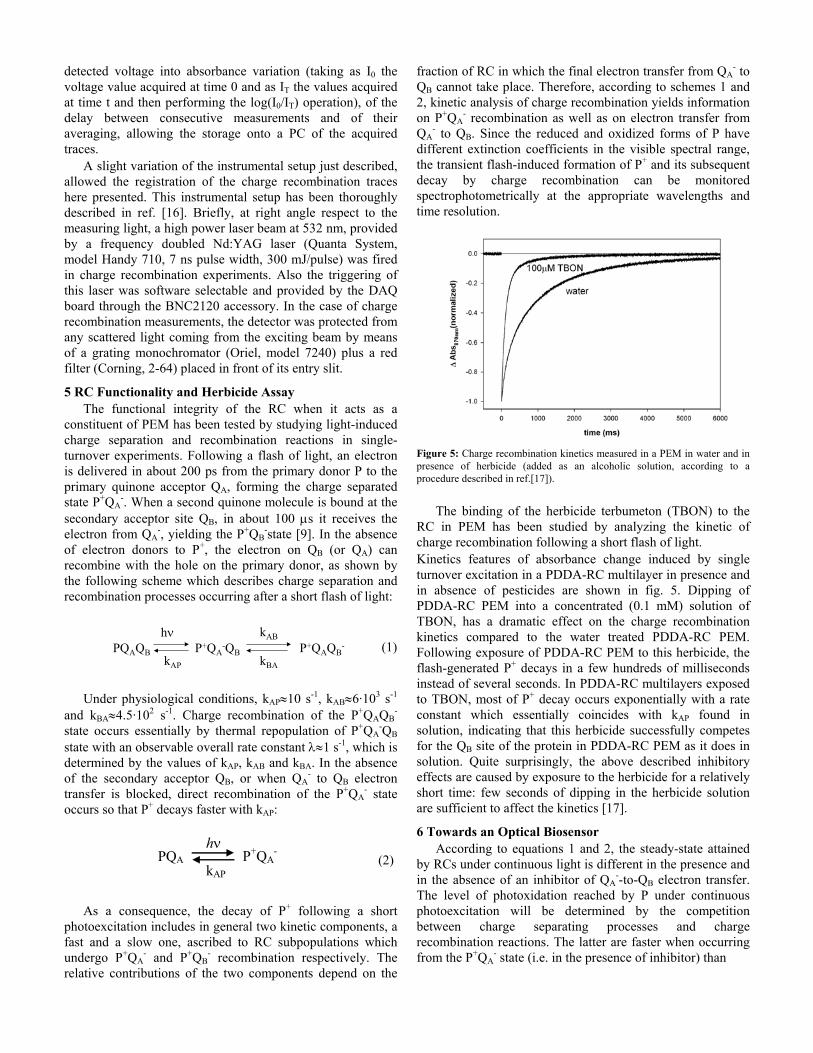
In the case of membrane proteins what is important is the
charge of the protein-detergent complex. At pH= 8 the RC-octylglucoside complex prepared according to ref. [13] is negatively charged, allowing an alternate and very effective deposition of a large number of PDDA and RC layers, as shown by the absorption spectra of fig. 3.
λ (nm)
650 700 750 800 850 900 950
Abs
orba
nce
0.0
0.1
0.2
0.3
0.4
0.5
18
2125
28
31
Figure 3: Absorption spectra of RC-PEM obtained following a different number of adsorption cycles (as indicated in the labels).
BB

PDDA does not absorb in the spectral region considered, while the RC cofactors give rise to three intense absorbance bands in the region between 700 and 950 nm [14, 15]. The band at 760 nm is ascribable to the Qy transition of bacteriopheophytin molecules. The band at 802 nm is due to the Qy transitions of the monomeric bacteriochlorophylls, together with a contribution from the high energy exciton component of the Qy transition of the bacteriochlorophyll molecules forming the special pair P. The long-wavelength band (at 865 nm in solution) is identified as the low energy exciton component of the Qy transition of P.
The PDDA-RC PEMs prepared with the above described procedure, and stored in distilled water at 4°C in the dark, display a striking stability, maintaining their spectral features over a period of several months.
4 Instrument Setup The criteria at the basis of the instrumental setup adopted
were the following. Since what is needed is just the value of the absorbance variation at 870 nm, the use of a monochromatic, coherent light source is recommended since it ensures stability in terms both of wavelength and intensity, in the mean time avoiding the use of a monochromator, thus simplifying the optical scheme of the instrument. In order to have an appreciable bleaching of the protein due to the impinging laser beam, its intensity should be high enough, thus allowing the use of a solid state detector, such as a silicon photodiode. This kind of detector, compared with photomultipliers, is less demanding in terms of electronic circuitry, it is easier to drive and has a maximum of sensitivity just in the region where we probed our PEM system. With these constrains in mind, a simple instrument has been built, whose block scheme is reported in fig. 4 together with the electrical scheme of the current to voltage converter.
For the laser source, a 10 mW index guided single transverse mode infrared laser diode (Roithner Lasertechnik, RLT8710MG) driven by a CAC board from the same manufacturer, has been used. The diode, operating at an average power of 2 mW, has been mounted on a passive thermal dissipator for cooling.
As it can be seen from the figure, the silicon photodiode (PD) is driven in photovoltaic mode since for our measurements high response speed are not required and the probed signal is not very intense. Because the incident beam is unfocused (to probe a large surface of the intrinsic heterogeneous PEM), a large active area PD has been used. For the current-to-voltage conversion, a di-FET OPAmp from Texas Intruments has been chosen. It works with practically unitary gain and with a feedback capacitor of 10 pF to limit the frequency response and to avoid gain peeking. As a whole, the electronic dead time of the detection unit is of few microseconds, therefore ensuring reliable measurements when the time resolution is of the order of thousands of microseconds. To acquire and digitalize the signal coming from the detection unit, a 16 bit resolution/200 ksample/s acquisition board (DAQ) has been used (National Instruments PCI 6013 in connection with the BNC 2120 multifunction board). This DAQ not only performs the A/D conversion of the input signal, but manage the triggering of the laser diode used to excite and probe the PEM. This is possible since the PCI 6013 not only can manage up to 16 not referenced single ended analog input signals, but it allows the generation of two analog pulses (used to trigger on and off the acquisition) and the managing of 8 independent digital I/O ports. To drive the hardware of the instrument, a suitable software program has been developed by us under LabView environment. This acquisition software took care of the gating of the exciting/measuring laser beam, of the conversion of the
Figure 4: Block scheme of the instrumentation used in this work with the details of the current-to-voltage conversion circuitry
detected voltage into absorbance variation (taking as I0 the voltage value acquired at time 0 and as IT the values acquired at time t and then performing the log(I0/IT) operation), of the delay between consecutive measurements and of their averaging, allowing the storage onto a PC of the acquired traces.
A slight variation of the instrumental setup just described, allowed the registration of the charge recombination traces here presented. This instrumental setup has been thoroughly described in ref. [16]. Briefly, at right angle respect to the measuring light, a high power laser beam at 532 nm, provided by a frequency doubled Nd:YAG laser (Quanta System, model Handy 710, 7 ns pulse width, 300 mJ/pulse) was fired in charge recombination experiments. Also the triggering of this laser was software selectable and provided by the DAQ board through the BNC2120 accessory. In the case of charge recombination measurements, the detector was protected from any scattered light coming from the exciting beam by means of a grating monochromator (Oriel, model 7240) plus a red filter (Corning, 2-64) placed in front of its entry slit.
5 RC Functionality and Herbicide Assay The functional integrity of the RC when it acts as a
constituent of PEM has been tested by studying light-induced charge separation and recombination reactions in single-turnover experiments. Following a flash of light, an electron is delivered in about 200 ps from the primary donor P to the primary quinone acceptor QA, forming the charge separated state P+QA
-. When a second quinone molecule is bound at the secondary acceptor site QB, in about 100 μs it receives the electron from QA
-, yielding the P+QB-state [9]. In the absence
of electron donors to P+, the electron on QB (or QA) can recombine with the hole on the primary donor, as shown by the following scheme which describes charge separation and recombination processes occurring after a short flash of light:
(1) Under physiological conditions, kAP≈10 s-1, kAB≈6·103 s-1
and kBA≈4.5·102 s-1. Charge recombination of the P+QAQB-
state occurs essentially by thermal repopulation of P+QA-QB
state with an observable overall rate constant λ≈1 s-1, which is determined by the values of kAP, kAB and kBA. In the absence of the secondary acceptor QB, or when QA
- to QB electron transfer is blocked, direct recombination of the P+QA
- state occurs so that P+ decays faster with kAP:
(2) As a consequence, the decay of P+ following a short
photoexcitation includes in general two kinetic components, a fast and a slow one, ascribed to RC subpopulations which undergo P+QA
- and P+QB- recombination respectively. The
relative contributions of the two components depend on the
fraction of RC in which the final electron transfer from QA- to
QB cannot take place. Therefore, according to schemes 1 and 2, kinetic analysis of charge recombination yields information on P+QA
- recombination as well as on electron transfer from QA
- to QB. Since the reduced and oxidized forms of P have different extinction coefficients in the visible spectral range, the transient flash-induced formation of P+ and its subsequent decay by charge recombination can be monitored spectrophotometrically at the appropriate wavelengths and time resolution.
Figure 5: Charge recombination kinetics measured in a PEM in water and in presence of herbicide (added as an alcoholic solution, according to a procedure described in ref.[17]).
The binding of the herbicide terbumeton (TBON) to the
RC in PEM has been studied by analyzing the kinetic of charge recombination following a short flash of light. Kinetics features of absorbance change induced by single turnover excitation in a PDDA-RC multilayer in presence and in absence of pesticides are shown in fig. 5. Dipping of PDDA-RC PEM into a concentrated (0.1 mM) solution of TBON, has a dramatic effect on the charge recombination kinetics compared to the water treated PDDA-RC PEM. Following exposure of PDDA-RC PEM to this herbicide, the flash-generated P+ decays in a few hundreds of milliseconds instead of several seconds. In PDDA-RC multilayers exposed to TBON, most of P+ decay occurs exponentially with a rate constant which essentially coincides with kAP found in solution, indicating that this herbicide successfully competes for the QB site of the protein in PDDA-RC PEM as it does in solution. Quite surprisingly, the above described inhibitory effects are caused by exposure to the herbicide for a relatively short time: few seconds of dipping in the herbicide solution are sufficient to affect the kinetics [17].
6 Towards an Optical Biosensor According to equations 1 and 2, the steady-state attained
by RCs under continuous light is different in the presence and in the absence of an inhibitor of QA
--to-QB electron transfer. The level of photoxidation reached by P under continuous photoexcitation will be determined by the competition between charge separating processes and charge recombination reactions. The latter are faster when occurring from the P+QA
- state (i.e. in the presence of inhibitor) than
PQAQB P+QA-QB P+QAQB
-
kAP
kAB
kBA
hν
PQA P+QA-
kAP
hν
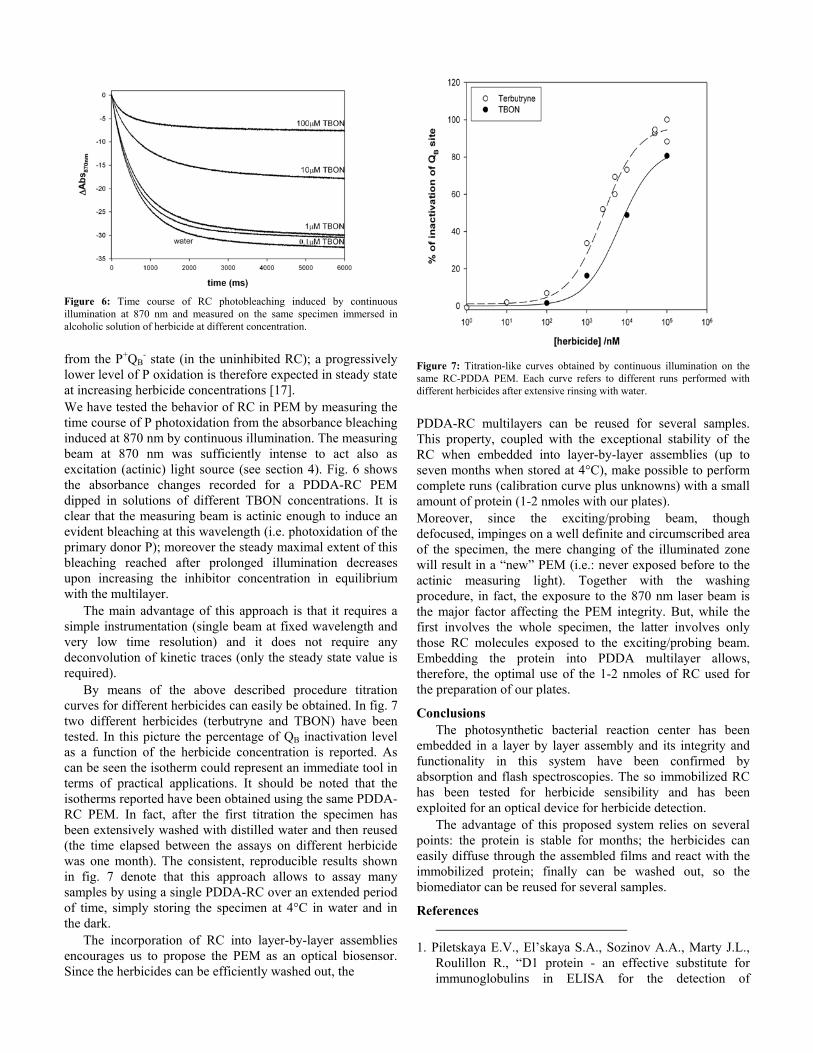
Figure 6: Time course of RC photobleaching induced by continuous illumination at 870 nm and measured on the same specimen immersed in alcoholic solution of herbicide at different concentration.
from the P+QB
- state (in the uninhibited RC); a progressively lower level of P oxidation is therefore expected in steady state at increasing herbicide concentrations [17]. We have tested the behavior of RC in PEM by measuring the time course of P photoxidation from the absorbance bleaching induced at 870 nm by continuous illumination. The measuring beam at 870 nm was sufficiently intense to act also as excitation (actinic) light source (see section 4). Fig. 6 shows the absorbance changes recorded for a PDDA-RC PEM dipped in solutions of different TBON concentrations. It is clear that the measuring beam is actinic enough to induce an evident bleaching at this wavelength (i.e. photoxidation of the primary donor P); moreover the steady maximal extent of this bleaching reached after prolonged illumination decreases upon increasing the inhibitor concentration in equilibrium with the multilayer.
The main advantage of this approach is that it requires a simple instrumentation (single beam at fixed wavelength and very low time resolution) and it does not require any deconvolution of kinetic traces (only the steady state value is required).
By means of the above described procedure titration curves for different herbicides can easily be obtained. In fig. 7 two different herbicides (terbutryne and TBON) have been tested. In this picture the percentage of QB inactivation level as a function of the herbicide concentration is reported. As can be seen the isotherm could represent an immediate tool in terms of practical applications. It should be noted that the isotherms reported have been obtained using the same PDDA-RC PEM. In fact, after the first titration the specimen has been extensively washed with distilled water and then reused (the time elapsed between the assays on different herbicide was one month). The consistent, reproducible results shown in fig. 7 denote that this approach allows to assay many samples by using a single PDDA-RC over an extended period of time, simply storing the specimen at 4°C in water and in the dark.
The incorporation of RC into layer-by-layer assemblies encourages us to propose the PEM as an optical biosensor. Since the herbicides can be efficiently washed out, the
Figure 7: Titration-like curves obtained by continuous illumination on the same RC-PDDA PEM. Each curve refers to different runs performed with different herbicides after extensive rinsing with water.
PDDA-RC multilayers can be reused for several samples. This property, coupled with the exceptional stability of the RC when embedded into layer-by-layer assemblies (up to seven months when stored at 4°C), make possible to perform complete runs (calibration curve plus unknowns) with a small amount of protein (1-2 nmoles with our plates). Moreover, since the exciting/probing beam, though defocused, impinges on a well definite and circumscribed area of the specimen, the mere changing of the illuminated zone will result in a “new” PEM (i.e.: never exposed before to the actinic measuring light). Together with the washing procedure, in fact, the exposure to the 870 nm laser beam is the major factor affecting the PEM integrity. But, while the first involves the whole specimen, the latter involves only those RC molecules exposed to the exciting/probing beam. Embedding the protein into PDDA multilayer allows, therefore, the optimal use of the 1-2 nmoles of RC used for the preparation of our plates.
Conclusions The photosynthetic bacterial reaction center has been
embedded in a layer by layer assembly and its integrity and functionality in this system have been confirmed by absorption and flash spectroscopies. The so immobilized RC has been tested for herbicide sensibility and has been exploited for an optical device for herbicide detection.
The advantage of this proposed system relies on several points: the protein is stable for months; the herbicides can easily diffuse through the assembled films and react with the immobilized protein; finally can be washed out, so the biomediator can be reused for several samples.
References
1. Piletskaya E.V., El’skaya S.A., Sozinov A.A., Marty J.L., Roulillon R., “D1 protein - an effective substitute for immunoglobulins in ELISA for the detection of

photosynthesis inhibiting herbicides”, Anal. Chim. Acta, Vol. 398, (1999), No. 1, pp. 49-56.
2. Banks K.E., Hernandez S., “Evaluation and validation of commercially available enzyme-linked immunosorbent assays (ELISAs) specific for atrazine, chlorpyrifos, and diazinon in aqueous phase”, Talanta, Vol. 61, (2003), No. 3, pp. 257-265.
3. Schneider P., Goodrow M.H., Gee S.J., Hammock B.D., “A highly sensitive and rapid Elisa for the arylurea herbicides diuron, monuron, and linuron”, J. Agric. Food Chem., Vol. 42, (1994), No. 2, pp. 413-422.
4. Harris R.D., Luff B.J, Wilkinson J.S., Piehler J., Brecht A., Gauglitz G., Abuknesha R.A., “Integrated optical surface plasmon resonance immunoprobe for simazine detection”, Biosens. Bioelectron., Vol. 14, (1999), No. 4, pp: 377-386.
5. Brecht A., Piehler J., Lang G., Gauglitz G., “A direct optical immunosensor for atrazine detection”, Anal. Chim. Acta, Vol. 311, (1995), No. 3, pp. 289-299.
6. Skl´adal P., “Effect of methanol on the interaction of monoclonal antibody with free and immobilized atrazine studied using the resonant mirror-based biosensor”, Biosens. Bioelectron., Vol. 14, (1999), No. 3, pp. 257-263.
7. Giardi M.T., Pace E., “Photosynthetic proteins for technological applications”, TRENDS Biotec., Vol. 23, (2005), No. 5, 257-263.
8. Stein R. R., Castellvi A. L., Bogacz J. P. and Wraight C. A., “Herbicide-Quinone Competition in the Acceptor Complex of Photosynthetic Reaction Center from Rhodopseudomonas Sphaeroides: a Bacterial Model for PS-II-Herbicide Activity in Plants”, Journal of Cellular Biochemistry, Vol. 24, (1984), pp. 243-259.
9. Feher G., Allen J.P., Okamura M.Y., Rees D.C., “Structure and function of bacterial photosynthetic reaction centres”, Nature, Vol. 33, (1989), pp. 111-116.
10. Ort D.R., Melandri B.A., Photosynthetsis. Energy Conversion by Plants and Bacteria, Academic press (New York, 1982), pp. 537-555.
11. Decher G., Schlenoff J.B., Multilayer Thin Films: Sequential Assembly of Nanocomposite Materials; Wiley-VCH (Weinheim, Germany, 2003).
12. Schonhoff M., “Self-assembled polyelectrolyte multilayers”, Curr. Opin. Colloid Interface Sci., Vol. 8, (2003), No. 1, pp. 86-95.
13. Gray K.A., Wachtveitl J., Breton J., Oestherhelt D., “Initial characterization of site-directed mutants of tyrosine M210 in the reaction centre of Rhodobacter sphaeroides”, EMBO J., Vol. 9, (1990), No. 7, pp. 2061-2070.
14. Cherepy N. J., Shreve A. P., Moore L. J., Boxer S. G., Mathies R. A., “Electronic and Nuclear Dynamics of the Accessory Bacteriochlorophylls in Bacterial Photosynthetic Reaction Centers from Resonance Raman Intensities”, J. Phys. Chem. B, Vol. 101, (1997), pp.3250-3260.
15. Steffen M. A., Lao K., Boxer S. G., “Dielectric asimmetry
in the photosynthetic reaction-center”, Science, Vol. 264, (1994), No. 5160, pp. 810-816.
16. Giustini M., Castelli F., Husu I., Giomini M., Mallardi A., Palazzo G., “Influence of cardiolipin on the functionality of the Q(A) site of the photosynthetic bacterial reaction center” J. Phys. Chem. B, Vol. 109, (2005), No. 44, pp. 21187-21196.
17. Mallardi A., Giustini M., Lopez F., Dezi M., Venturoli G., and Palazzo G., “Reaction centers in polyelectrolyte multilayers: towards an herbicide biosensor” J. Phys. Chem. B, Vol. 111, (2007), No. 12, pp. 3304-3314.
Copyright © 2022 FDOKUMEN