
Phosphosite conservation in single domain orthologs versus paralogs: A way to combine differential...
7
Hypothesis Phosphosite conservation in single domain orthologs versus paralogs: A way to combine differential regulation with redundant core functions Lynn Huyck, Marleen Van Troys, Christophe Ampe ⇑ Department of Biochemistry, Faculty of Medicine and Health Sciences, Ghent University, Ghent, Belgium article info Article history: Received 15 December 2011 Revised 4 January 2012 Accepted 9 January 2012 Available online 18 January 2012 Edited by Miguel De la Rosa Keywords: Single domain protein Sequence conservation Actin binding protein Isoform specific regulation Phosphorylation abstract Evolutionary conservation for structure function relations is commonly accepted. Here we hypoth- esize that closely related single domain paralogous proteins, having similar expression profiles and redundant biochemical core functions, additionally evolved to allow and maintain isoform specific differential regulation by single conserved amino acid substitutions. To substantiate this, we consid- ered two families of closely related actin binding proteins combined with data mining of phosphor- ylated residues in human and mouse proteins. We show that such residues are identical in other orthologs whereas paralogs have a different, but also conserved, non-phosphorylatable residue at the equivalent positions. Ó 2012 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. 1. Introduction Evolutionary conservation of protein structure–function rela- tionships is reflected in similarity in primary structure. This para- digm is well-accepted for orthologous proteins but also applies to closely related paralogs that have distinct sequence features. Regulation in nature is complex, involving many pathways includ- ing kinase signaling. Recently it was highlighted in several ana- lyzes of phosphoproteomic data that evolutionary information is useful to identify functional regulatory features in eukaryotic pro- teomes, especially in vertebrates [1–3]. Whereas these studies fo- cused on orthologous proteins they did not compare paralogs. Although it has been argued that tissue specific expression is an underlying necessity for having two closely related proteins in an organism, such isoforms with very similar biochemical functions are often co-expressed. One emerging concept is that the possibil- ity for differential spatio-temporal regulation of these functions re- quires the presence of paralogs in the same cell. Larger multimodular proteins can achieve this by varying their regulatory properties by differential function of their modules; single structural domain proteins, however, are confronted with a prob- lem since the number of (surface) residues potentially participat- ing in regulation is more restricted. Here we hypothesize that differential regulation of closely related single domain paralogs is brought about by distinct but conserved local sequence features, including single amino acid substitutions, in a context of maintain- ing core function and a stable structure. We illustrate this hypoth- esis by an in silico analysis of two case examples of biochemically well-characterized families of single domain proteins from the ac- tin system. The actin cytoskeleton is important in many cellular processes including cell migration, cytokinesis, endocytosis and cell contrac- tion. It functions in all tissues or organs of multicellular organisms. The very conserved actin molecule is the central protein in this sys- tem [4]. Its main property is to form filaments of which the assem- bly, disassembly or stabilization is governed by a multitude of actin binding proteins [5,6]. To generate orchestrated productive func- tions, the actin system in cells is tightly regulated in space and time by a wide array of regulators like kinases, controlling the activity of the actin binding proteins [7]. Many actin binding proteins come in small families of closely related isoforms encoded by different genes. It is not always under- stood why this is. Because the system is so important for viability, some redundancy may be expected. Yet, mouse genetics has shown in several cases that the system is not entirely redundant [8–11]. Moreover, many of the isoforms of actin binding proteins are ex- pressed in the same tissue or cells (see also results). Another 0014-5793/$36.00 Ó 2012 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved. doi:10.1016/j.febslet.2012.01.018 Abbreviations: EST, expressed sequence tag; ADF, actin depolymerizing factor; EVH, Ena/VASP homology; PtdIns(4,5)P 2 , 1-(3-sn-Phosphatidyl)-D-myo-inositol 4,5- bis(phosphate); SH2, src homology 2; SH3, src homology 3 ⇑ Corresponding author. Address: Albert Baertsoenkaai 3, B-9000 Ghent, Belgium. Fax: +32 92649490. E-mail address: [email protected] (C. Ampe). FEBS Letters 586 (2012) 296–302 journal homepage: www.FEBSLetters.org
Transcript of Phosphosite conservation in single domain orthologs versus paralogs: A way to combine differential...

FEBS Letters 586 (2012) 296–302
journal homepage: www.FEBSLetters .org
Hypothesis
Phosphosite conservation in single domain orthologs versus paralogs: A wayto combine differential regulation with redundant core functions
Lynn Huyck, Marleen Van Troys, Christophe Ampe ⇑Department of Biochemistry, Faculty of Medicine and Health Sciences, Ghent University, Ghent, Belgium
a r t i c l e i n f o
Article history:Received 15 December 2011Revised 4 January 2012Accepted 9 January 2012Available online 18 January 2012
Edited by Miguel De la Rosa
Keywords:Single domain proteinSequence conservationActin binding proteinIsoform specific regulationPhosphorylation
0014-5793/$36.00 � 2012 Federation of European Biodoi:10.1016/j.febslet.2012.01.018
Abbreviations: EST, expressed sequence tag; ADF,EVH, Ena/VASP homology; PtdIns(4,5)P2, 1-(3-sn-Phosbis(phosphate); SH2, src homology 2; SH3, src homol⇑ Corresponding author. Address: Albert Baertsoenk
Fax: +32 92649490.E-mail address: [email protected] (C. Am
a b s t r a c t
Evolutionary conservation for structure function relations is commonly accepted. Here we hypoth-esize that closely related single domain paralogous proteins, having similar expression profiles andredundant biochemical core functions, additionally evolved to allow and maintain isoform specificdifferential regulation by single conserved amino acid substitutions. To substantiate this, we consid-ered two families of closely related actin binding proteins combined with data mining of phosphor-ylated residues in human and mouse proteins. We show that such residues are identical in otherorthologs whereas paralogs have a different, but also conserved, non-phosphorylatable residue atthe equivalent positions.� 2012 Federation of European Biochemical Societies. Published by Elsevier B.V. All rights reserved.
1. Introduction
Evolutionary conservation of protein structure–function rela-tionships is reflected in similarity in primary structure. This para-digm is well-accepted for orthologous proteins but also appliesto closely related paralogs that have distinct sequence features.Regulation in nature is complex, involving many pathways includ-ing kinase signaling. Recently it was highlighted in several ana-lyzes of phosphoproteomic data that evolutionary information isuseful to identify functional regulatory features in eukaryotic pro-teomes, especially in vertebrates [1–3]. Whereas these studies fo-cused on orthologous proteins they did not compare paralogs.Although it has been argued that tissue specific expression is anunderlying necessity for having two closely related proteins in anorganism, such isoforms with very similar biochemical functionsare often co-expressed. One emerging concept is that the possibil-ity for differential spatio-temporal regulation of these functions re-quires the presence of paralogs in the same cell. Largermultimodular proteins can achieve this by varying their regulatoryproperties by differential function of their modules; single
chemical Societies. Published by E
actin depolymerizing factor;phatidyl)-D-myo-inositol 4,5-ogy 3aai 3, B-9000 Ghent, Belgium.
pe).
structural domain proteins, however, are confronted with a prob-lem since the number of (surface) residues potentially participat-ing in regulation is more restricted. Here we hypothesize thatdifferential regulation of closely related single domain paralogs isbrought about by distinct but conserved local sequence features,including single amino acid substitutions, in a context of maintain-ing core function and a stable structure. We illustrate this hypoth-esis by an in silico analysis of two case examples of biochemicallywell-characterized families of single domain proteins from the ac-tin system.
The actin cytoskeleton is important in many cellular processesincluding cell migration, cytokinesis, endocytosis and cell contrac-tion. It functions in all tissues or organs of multicellular organisms.The very conserved actin molecule is the central protein in this sys-tem [4]. Its main property is to form filaments of which the assem-bly, disassembly or stabilization is governed by a multitude of actinbinding proteins [5,6]. To generate orchestrated productive func-tions, the actin system in cells is tightly regulated in space andtime by a wide array of regulators like kinases, controlling theactivity of the actin binding proteins [7].
Many actin binding proteins come in small families of closelyrelated isoforms encoded by different genes. It is not always under-stood why this is. Because the system is so important for viability,some redundancy may be expected. Yet, mouse genetics has shownin several cases that the system is not entirely redundant [8–11].Moreover, many of the isoforms of actin binding proteins are ex-pressed in the same tissue or cells (see also results). Another
lsevier B.V. All rights reserved.

1 In the phosphosite repository many other residues, including threonines orserines, of ADF/cofilins are found as phosphorylated but these did not meet the
L. Huyck et al. / FEBS Letters 586 (2012) 296–302 297
reason for having closely related actin binding isoforms could berelated to a differential interaction with beta- or gamma-actin, thatonly differ in four residues at the N-terminus [12]. However, theknown interfaces of many actin regulatory proteins with actinsdo not map to this N-terminal region. A scenario which has beenlittle explored is that of differential regulation. In multimodular ac-tin binding proteins, this may be brought about by the non-actin-interacting domains. A well known example are the Ena/VASP pro-teins that harbor actin binding activity in their C-terminal part butare targeted to specific cellular sites by their N-terminal EVH (Ena/VASP homology) domains and differentially regulated via interac-tion of their central polyproline-rich region with WW or SH3 (srchomology 3) domain containing proteins [13]. However, closely re-lated single domain actin binding proteins require high sequenceconservation for maintaining their fold and their core business: ac-tin binding and regulation of actin dynamics. Together this putsstructural and thus evolutionary constraints on differential regula-tion. Two important families of single domain modulators of actindynamics are ADF/cofilins (cofilin 1, cofilin 2 and actin depolymer-izing factor or destrin) and profilins (profilin I and IIa). ADF/cofilinsdynamize actin turnover [14] and profilins are either actin seques-tering proteins or assist in filament elongation [15]. Within onefamily, the actin binding properties are very similar, if not identical[13,16–18]. For these actin binding proteins regulatory ligandbinding with the polyphosphoinositide PtdIns(4,5)P2 (1-(3-sn-Phosphatidyl)-D-myo-inositol 4,5-bis(phosphate)) has also beendescribed [19,20]. Multiple positively charged amino acids partici-pate in multivalent interaction of which the interface, at leastpartly, overlaps with the one for actin. This makes this a less likelyavenue for isoform specific regulation, although for profilins (butnot for ADF/cofilins) differences in affinities have been observed[21–24]. Cofilin and destrin members also share regulation byphosphorylation of Ser3 that inactivate these proteins with respectto actin binding [25–27].
Starting from the initial observation that members of either theADF/cofilin or profilin family are co-expressed, we here explore thepossibility that paralogs within these two families of small actinbinding proteins are differentially regulated by (conserved)changes in phosphorylatable key residues. The overall high se-quence conservation serves to impose biochemical conservationof actin or ligand binding, whereas local but specific sequence var-iation allows differential regulation. We base this hypothesis oncomparing primary structures, mapping differences on 3D-struc-tures and database mining of (often functionally uncharacterized)phosphorylation events. Similar mechanisms of differential regula-tion may not only apply to the studied proteins but also to othersingle domain paralogs with high sequence conservation.
2. Materials and methods
We only use human nomenclature for protein symbols althoughwe often refer to proteins from different species. Sequence num-bering starts at the initiator methionine. For alignment of ADF/cofi-lins, the protein sequences in Table S1 were retrieved at NCBI(RefSeq release 50) or Ensembl (release 64). These were used toconstruct a tree and position poorly annotated sequences of lowervertebrate ADF/cofilins (Fig. S1). For profilins, we relied on previ-ously published alignments [28]. Expression data for human wereretrieved from http://ist.genesapiens.org (human) and for mouseESTs (expressed sequence tag) are available at Unigene http://www.ncbi.nlm.nih.gov/unigene. Mus musculus and human profilinIIa (NCBI RefSeq IDs: NP_062283.1, AAG09754.1) and IIb (NCBI Ref-Seq IDs: AAH95444.1, AAH43646.1) were used to determine thenumber of human or mouse ESTs using tblastn at http://blas-t.ncbi.nlm.nih.gov. The information on phosphorylation was re-
trieved from http://www.phosphosite.org [29]. Initial statisticalanalysis on the global dataset indicates that phosphorylated pep-tides are on average detected approximately 5 times over all hu-man and mouse proteomics experiments (Table 1). We withheldonly phosphorylations with a frequency of detection above thisaverage in a particular isoform and observed for both mouse andhuman homologs. Phosphosites known to be phosphorylated byparticular kinase studies (Table 1) were also included. Isoform-spe-cific assignment of sequences containing a phosphosite was not al-ways possible (see Table 1). Coordinates for the 3D-structures ofhuman cofilin 1 (PDB ID: 1Q8X), the bovine profilin I-actin complex(PDB ID: 2BTF) and human profilin IIa (PDB ID: 1d1j), were re-trieved at http://www.pdb.org/pdb/home/home.do and visualizedwith Viewerlite.
3. Results and discussion
3.1. Cofilin/ADF members have differentially phosphorylatedconserved residues
Cofilin 1 and ADF are co-expressed in comparable levels in awide variety of tissues (with the exception of blood, bone marrowand the lymphatic system mainly expressing cofilin 1) (human(Fig. 1), for mouse see NCBI Unigene IDs: Mm.329655,Mm.276826 and Mm.28919). Cofilin 2 has a more restrictedexpression pattern and is typically present in muscle tissues andthe central nervous system (Fig. S2). The co-occurrence of cofilin1 and destrin, with very similar actin binding properties, in varioustissues potentially renders redundancy but may additionally en-able fine tuning of cytoskeletal function or regulation in subcellularspace and time. We investigated the hypothesis that isoform spe-cific regulation is based on differential phosphorylation at evolu-tionary conserved key residues within paralogs.
In function of this, we evaluated sequence conservation be-tween vertebrate ADF/cofilin members from selected (model)organisms. Most of these organisms contain the three family mem-bers. A remarkable exception is bird species: chicken and zebrafinch lack cofilin 1. Although the inferred tree is not truly phyloge-netic (Fig. S1), the three expected groups, a cofilin 1, a cofilin 2 anda destrin group, can be distinguished albeit that some sequences oflower vertebrates are less reliably assigned to a particular group(see Supplementary results). Sequence conservation between ver-tebrate homologs is generally high and in particular residues in-volved in actin binding are at most of the positions identical (forbird and mammalian para- and orthologs) or very conserved (forXenopus destrin and Danio rerio cofilin 1, Fig. S3). This is consis-tent with the conserved biochemical properties of ADF/cofilins[18]. In view of availability of phosphoproteomics data, we furtherfocused on mammalian ADF/cofilin members.
We searched the http://www.phosphosite.org repository forphosphorylation events in these isoforms. Based on a thresholdfor detection frequency (see Section 2), we consider the tyrosineand serine residues listed in Table 1 as being phosphorylated1
(http://www.phosphosite.org). The well characterized phosphoryla-tion on S3 [25,26] is well sampled in all three isoforms. S3 is at theinterface with actin and its phosphorylation results in inhibition ofactin binding (Fig. 2C, Fig. S3) [25–27]. A second site in cofilin 1,known to be phosphorylated by Src kinase, is Y68 which is, witha single exception, conserved in all vertebrate reference sequences(see Fig. S3). This residue is also present in cofilin 2 (Table 1), how-ever, in contrast to cofilin 1, there is no mass spectrometry evi-
imposed threshold, see also supplementary results section.

Table 1Frequently observed phosphorylations for human and mouse ADF/cofilin members and for profilin I and II retrieved from http://www.phosphosite.org.
aAmino acid residue (AA) in the protein, numbering starts at the initiator Met.bThe number of records in which this modification was assigned using only proteomic discovery-mode mass spectrometry (MS) (human/mouse), ‘no’ indicates not observedfor that species, ‘na’ not applicable.cReferences to published studies on human/mouse.dReference based on kinase experiments.eSequence of the tryptic peptide in which the phosphorylated amino acid (lower case) is found. Residues in bold allow discriminating the peptides of the paralogs in massspectrometric experiments. Only one sequence is given when it is identical in human (h) and mouse (m).fThe exact number could not be retrieved. For statistics this number was set to 25. The calculated averages are thus under estimations.
298 L. Huyck et al. / FEBS Letters 586 (2012) 296–302
dence for phosphorylation of Y68 in cofilin 2. Interestingly, in des-trin this residue is phenylalanine in all NCBI vertebrate referencesequences (Fig. S3) and therefore destrins cannot be phosphory-lated by Src at this position. The tryptic peptide containing bothY85 and Y89 does not discriminate between cofilin 1/2 but doesso for destrin. The latter contains a phenylalanine at position 89which, obviously, cannot be posttranslationally modified but Y85of destrin is phosphorylated (Table 1). Similar to residue 68,residue 89 is always Y in cofilins and F in destrins (with exceptionof Xenopus tropicalis destrin, Fig S3). Y117 is located in a peptide
that does not allow deriving the paralog it originates from. Phos-phorylation of Y140 is specific for cofilin 1 since both cofilin 2and destrin do not contain a phosphorylatable residue at this posi-tion. Again Y140 is conserved in cofilin 1 with a single exception(for Xenopus tropicalis). S156 is conserved in cofilin 1 and destrin,but not in cofilin 2, and the tryptic peptide containing this serineallows making a distinction between cofilin 1 and destrin. Phos-phorylation of destrin S156 is not reported, but is detected in cofi-lin 1 (Table 1). Collectively this indicates that the two moreabundant and co-expressed forms cofilin 1 and destrin are differ-

Fig. 1. Gene expression data of cofilin1, destrin, profilin I and II. Cofilin and destrin are often co-expressed and a similar observation can be made for profilin I and IIa. For dataon cofilin 2 see Fig. S2. Data are derived from micro-array experiments at http://www.genesapiens.org and filtered on healthy tissues. The Y-axis is relative expression level,the X-axis numbers refer to a tissue (right).
L. Huyck et al. / FEBS Letters 586 (2012) 296–302 299
entially phosphorylated at residues 68, 89, 140 and perhaps S156and may form avenues for isoform specific regulation within thisfamily.
Mapping the phosphosites on the 3D-structure of cofilin 1 re-veals that 3 out of 5 of the considered tyrosines locate at the inte-rior of the protein (Fig. 2A). In view of phosphorylation, the buriednature of Y68, Y85 and Y117 and the resulting expected limitedaccessibility is surprising. However, phosphorylation of Y68 by
Src has a role in ubiquitination and subsequent degradation of cofi-lin 1 [30]. This suggests that (partly) unfolded cofilin 1 is a Src sub-strate and it is tempting to speculate that phosphorylation of Y85and Y117 also occurs in such a manner (not necessarily by Src).Nevertheless, based on our analysis above, the Src induced phos-phorylation at Y68 appears specific for the cofilin 1 paralog. Thehydroxyl groups of S156 and Y89 (in cofilin 1/2) are at the surfaceof cofilin 1 (Fig 2A). Phosphorylation of S156 is observed in cofilin 1
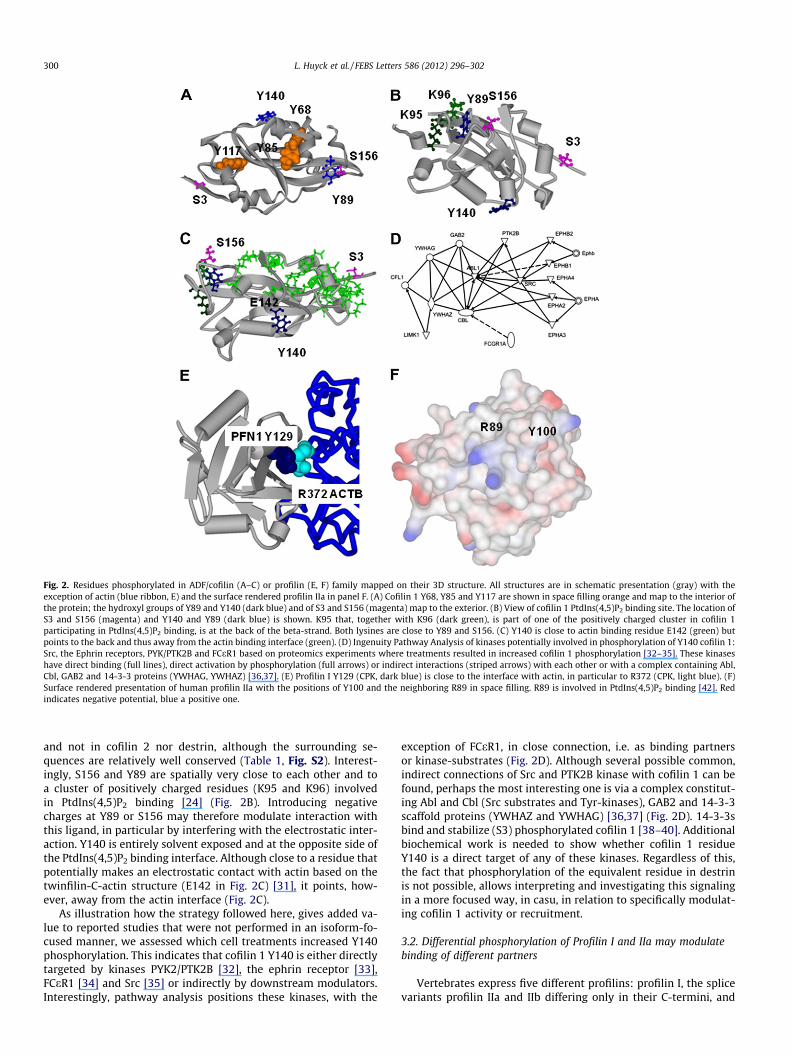
Fig. 2. Residues phosphorylated in ADF/cofilin (A–C) or profilin (E, F) family mapped on their 3D structure. All structures are in schematic presentation (gray) with theexception of actin (blue ribbon, E) and the surface rendered profilin IIa in panel F. (A) Cofilin 1 Y68, Y85 and Y117 are shown in space filling orange and map to the interior ofthe protein; the hydroxyl groups of Y89 and Y140 (dark blue) and of S3 and S156 (magenta) map to the exterior. (B) View of cofilin 1 PtdIns(4,5)P2 binding site. The location ofS3 and S156 (magenta) and Y140 and Y89 (dark blue) is shown. K95 that, together with K96 (dark green), is part of one of the positively charged cluster in cofilin 1participating in PtdIns(4,5)P2 binding, is at the back of the beta-strand. Both lysines are close to Y89 and S156. (C) Y140 is close to actin binding residue E142 (green) butpoints to the back and thus away from the actin binding interface (green). (D) Ingenuity Pathway Analysis of kinases potentially involved in phosphorylation of Y140 cofilin 1:Src, the Ephrin receptors, PYK/PTK2B and FCeR1 based on proteomics experiments where treatments resulted in increased cofilin 1 phosphorylation [32–35]. These kinaseshave direct binding (full lines), direct activation by phosphorylation (full arrows) or indirect interactions (striped arrows) with each other or with a complex containing Abl,Cbl, GAB2 and 14-3-3 proteins (YWHAG, YWHAZ) [36,37]. (E) Profilin I Y129 (CPK, dark blue) is close to the interface with actin, in particular to R372 (CPK, light blue). (F)Surface rendered presentation of human profilin IIa with the positions of Y100 and the neighboring R89 in space filling. R89 is involved in PtdIns(4,5)P2 binding [42]. Redindicates negative potential, blue a positive one.
300 L. Huyck et al. / FEBS Letters 586 (2012) 296–302
and not in cofilin 2 nor destrin, although the surrounding se-quences are relatively well conserved (Table 1, Fig. S2). Interest-ingly, S156 and Y89 are spatially very close to each other and toa cluster of positively charged residues (K95 and K96) involvedin PtdIns(4,5)P2 binding [24] (Fig. 2B). Introducing negativecharges at Y89 or S156 may therefore modulate interaction withthis ligand, in particular by interfering with the electrostatic inter-action. Y140 is entirely solvent exposed and at the opposite side ofthe PtdIns(4,5)P2 binding interface. Although close to a residue thatpotentially makes an electrostatic contact with actin based on thetwinfilin-C-actin structure (E142 in Fig. 2C) [31], it points, how-ever, away from the actin interface (Fig. 2C).
As illustration how the strategy followed here, gives added va-lue to reported studies that were not performed in an isoform-fo-cused manner, we assessed which cell treatments increased Y140phosphorylation. This indicates that cofilin 1 Y140 is either directlytargeted by kinases PYK2/PTK2B [32], the ephrin receptor [33],FCeR1 [34] and Src [35] or indirectly by downstream modulators.Interestingly, pathway analysis positions these kinases, with the
exception of FCeR1, in close connection, i.e. as binding partnersor kinase-substrates (Fig. 2D). Although several possible common,indirect connections of Src and PTK2B kinase with cofilin 1 can befound, perhaps the most interesting one is via a complex constitut-ing Abl and Cbl (Src substrates and Tyr-kinases), GAB2 and 14-3-3scaffold proteins (YWHAZ and YWHAG) [36,37] (Fig. 2D). 14-3-3sbind and stabilize (S3) phosphorylated cofilin 1 [38–40]. Additionalbiochemical work is needed to show whether cofilin 1 residueY140 is a direct target of any of these kinases. Regardless of this,the fact that phosphorylation of the equivalent residue in destrinis not possible, allows interpreting and investigating this signalingin a more focused way, in casu, in relation to specifically modulat-ing cofilin 1 activity or recruitment.
3.2. Differential phosphorylation of Profilin I and IIa may modulatebinding of different partners
Vertebrates express five different profilins: profilin I, the splicevariants profilin IIa and IIb differing only in their C-termini, and

L. Huyck et al. / FEBS Letters 586 (2012) 296–302 301
profilin III and IV [28]. The two last forms are low abundant withrestricted tissue expression. We only consider profilin I and IIa thathave identified phosphorylated peptides in phosphoproteomicsexperiments (Table 1). Similar to cofilin 1 and destrin, profilin Iand II2 are often present at similar levels in various tissues(Fig. 1, NCBI Unigene ID: Mm.2647, Mm.271744). Profilin I andIIA each have one tyrosine residue that has been found in manyproteomics studies. Both are located in tryptic peptides that allowdiscriminating these isoforms. Note that S138 in profilin I, phos-phorylated by ROCK [41], is not sampled in the phosphoproteomicsdatabase. Comparing the phosphoproteome data for profilin I andIIa reveals an intriguing result. In profilin I Y129 is phosphorylatedwhereas the equivalent tyrosine in profilin IIa has not been re-ported to be phosphorylated. By contrast, Y100 in profilin IIa islisted as being phosphorylated and the equivalent residue in profi-lin I is F. With three exceptions of the (predicted) proteins fromMonodelphis domestica, Oncorhynchus mykiss and Xenopus trop-icalis, all complete vertebrate profilin I reference sequences havethis phenylalanine.
Mapping these residues on the 3D-structure further demon-strates differences (Fig. 2E and F). Y129 is not considered as an inter-face residue with actin but is in close proximity of actin R372(Fig. 2E). Phospho-Y129 could therefore create an additional saltbridge strengthening the profilin I-actin interaction. Alternativelymutation of profilin R136, which is in the same C-terminal helix asY129, is known to negatively affect both PtdIns(4,5)P2 and polypro-line binding [42]. Therefore the possibility that Y129 phosphoryla-tion additionally affects the interaction with these ligands cannotbe excluded. Y100 in profilin IIa points away from the actin interface(based on the position of F100 in the profilin I actin complex). Theresidue spatially closest with a known functional importance isR89 which is involved in PtdIns(4,5)P2 binding [42,43]. Althoughthe polyphosphoinositide interface of profilins remains poorly de-fined, phosphorylation of Y100 would increase the negative surfacepotential (Fig. 2F) and thereby reduce the affinity for this ligand.
In summary, although, with exception of Src kinase targetingcofilin Y68, no tyrosine kinases that phosphorylate cofilin or profi-lin family members have been identified, it is evident from ourdata mining that these proteins are tyrosine phosphorylated. It isalso clear that within each family isoforms undergo, at least in part,differential phosphorylation and that this possibility is evolution-ary conserved. This potentially allows differential regulation whichmay include but is not necessarily limited to: targeting to the deg-radation machinery (Y68 in cofilin), recognition by SH2-domains(src homology 2) in scaffolding proteins or modulating interactionwith actin (predicted for profilin I Y129) or with polyphosphoinos-itides (predicted for cofilin Y89 and profilin IIa Y100). This must beviewed in the context of high structural conservation of single do-main proteins required for exerting a very similar core business(here actin binding). Indeed the extended interface with actin im-plies that multiple residues of profilins or ADF/cofilins need to beconserved to make the appropriate contacts. Posttranslationalmodifications like phosphorylation often require a more limitedrecognition site that can be changed either by mutation of sur-rounding residues (such as for ADF/cofilin S156 and profilinY129) or by single mutations of the phosphorylatable residues(all other discussed cases). Nature likely exploits both scenariosfor differential regulation of conserved single domain proteins.The first scenario is affinity-based and depends on recognition bykinases, the second precludes a catalytic event. Additional phos-phoproteomics (i.e. from species other than human and mouse)
2 The Genesapiens and unigene data on profilin II do not allow discriminatingbetween profilin IIa and IIb. Based on an EST analysis of specific splice variants aNCBI, profilin IIa is the predominant form (human IIa: >250 validated ESTs, humanIIb: 85 ESTs, mouse IIa >60 ESTs, mouse IIb 0 Ests).
t
and identification of conserved signaling pathways for regulationof orthologous proteins are needed to validate this hypothesis inconjunction with elaborate in cellulo experiments measuring theimpact of site directed mutagenesis of the identified key residueson spatio-temporal localization and activity of the (mutated) iso-forms downstream of identified signaling pathways. It is likely thatthe second scenario also applies to other paralogs with stronglyconserved functions. Indeed, in at least three other protein fami-lies: the high mobility group box family of proteins, the GRB2and the clathrin family, such local conservative changes in paralogsleading to altered capacity to become phosphorylated occur. Innearly all species the (un)phosphorylatable residue is conservedin a given ortholog of these three protein families. The rare excep-tions are reference protein sequences from fish or Xenopus specieswhere the assigned annotation of a particular isoform is uncertainbased on a preliminary phylogenetic analysis (data not shown).The high mobility group box family of proteins always contains Yin the B1 group (excluding B1-like) at position 109 whereas thisis a non-phosphorylatable residue in the B2 group (usually H). Sim-ilarly, vertebrate clathrin A light chains contain a conserved tyro-sine residue (Y84, two exceptions in predicted proteins in fish)that is phosphorylated in human whereas in the related B lightchain orthologs this residue is invariably F. Also, human GRB2 isphosphorylated on the conserved residue Y160 while this residueis always F in the GRB2-related adaptor protein reference se-quences. Since this residue resides in the second SH3 domain ofGRB2 this may also suggest that the proposed hypothesis also ap-plies to single domains within multidomain proteins.
Acknowledgements
This work was supported by fund G.0441.10N from the FWOVlaanderen and fund 01601207 from the GOA (GeconcerteerdeOnderzoeksactie) granted to C.A. L.H. was funded by the BOF (Bij-zonder Onderzoeksfonds) 01J04806.
Appendix A. Supplementary data
Supplementary data associated with this article can be found, inthe online version, at doi:10.1016/j.febslet.2012.01.018.
References
[1] Gnad, F., Forner, F., Zielinska, D.F., Birney, E., Gunawardena, J. and Mann, M.(2010) Evolutionary constraints of phosphorylation in eukaryotes,prokaryotes, and mitochondria. Mol. Cell Proteomics 9, 2642–2653.
[2] Landry, C.R., Levy, E.D. and Michnick, S.W. (2009) Weak functional constraintson phosphoproteomes. Trends Genet. 25, 193–197.
[3] Wang, Z., Ding, G., Geistlinger, L., Li, H., Liu, L., Zeng, R., Tateno, Y. and Li, Y.(2011) Evolution of protein phosphorylation for distinct functional modules invertebrate genomes. Mol. Biol. Evol. 28, 1131–1140.
[4] Tondeleir, D., Vandamme, D., Vandekerckhove, J., Ampe, C. and Lambrechts, A.(2009) Actin isoform expression patterns during mammalian development andin pathology: insights from mouse models. Cell Motil Cytoskeleton 66, 798–815.
[5] Ridley, A.J. (2011) Life at the leading edge. Cell 145, 1012–1022.[6] Parsons, J.T., Horwitz, A.R. and Schwartz, M.A. (2010) Cell adhesion:
integrating cytoskeletal dynamics and cellular tension. Nat. Rev. Mol. CellBiol. 11, 633–643.
[7] Winograd-Katz, S.E., Brunner, M.C., Mirlas, N. and Geiger, B. (2011) Analysis ofthe signaling pathways regulating Src-dependent remodeling of the actincytoskeleton. Eur. J. Cell Biol. 90, 143–156.
[8] Kwiatkowski, A.V. et al. (2007) Ena/VASP Is Required for neuritogenesis in thedeveloping cortex. Neuron 56, 441–455.
[9] Witke, W., Sutherland, J.D., Sharpe, A., Arai, M. and Kwiatkowski, D.J. (2001)Profilin I is essential for cell survival and cell division in early mousedevelopment. Proc. Natl. Acad. Sci. U.S.A. 98, 3832–3836.
[10] Gurniak, C.B., Perlas, E. and Witke, W. (2005) The actin depolymerizing factorn-cofilin is essential for neural tube morphogenesis and neural crest cellmigration. Dev. Biol. 278, 231–241.
[11] Zhang, W., Zhao, J., Chen, L., Urbanowicz, M.M. and Nagasaki, T. (2008)Abnormal epithelial homeostasis in the cornea of mice with a destrin deletion.Mol. Vis. 14, 1929–1939.

302 L. Huyck et al. / FEBS Letters 586 (2012) 296–302
[12] Vandekerckhove, J. and Weber, K. (1978) Mammalian cytoplasmic actins arethe products of at least two genes and differ in primary structure in at least 25identified positions from skeletal muscle actins. Proc. Natl. Acad. Sci. U.S.A. 75,1106–1110.
[13] Lambrechts, A., Kwiatkowski, A.V., Lanier, L.M., Bear, J.E., Vandekerckhove, J.,Ampe, C. and Gertler, F.B. (2000) CAMP-dependent protein kinasephosphorylation of EVL, a Mena/VASP relative, regulates its interaction withactin and SH3 domains. J. Biol. Chem. 275, 36143–36151.
[14] Carlier, M.F. et al. (1997) Actin depolymerizing factor (ADF/cofilin) enhancesthe rate of filament turnover: implication in actin-based motility. J. Cell. Biol.136, 1307–1322.
[15] Pantaloni, D. and Carlier, M.F. (1993) How profilin promotes actin filamentassembly in the presence of thymosin beta 4. Cell 75, 1007–1014.
[16] Van Troys, M., Huyck, L., Leyman, S., Dhaese, S., Vandekerkhove, J. and Ampe,C. (2008) Ins and outs of ADF/cofilin activity and regulation. Eur. J. Cell Biol. 87,649–667.
[17] Vartiainen, M.K., Mustonen, T., Mattila, P.K., Ojala, P.J., Thesleff, I., Partanen, J.and Lappalainen, P. (2002) The three mouse actin-depolymerizing factor/cofilins evolved to fulfill cell-type-specific requirements for actin dynamics.Mol. Biol. Cell 13, 183–194.
[18] Yeoh, S., Pope, B., Mannherz, H.G. and Weeds, A. (2002) Determining thedifferences in actin binding by human ADF and cofilin. J. Mol. Biol. 315, 911–925.
[19] Goldschmidt-Clermont, P.J., Furman, M.I., Wachsstock, D., Safer, D., Nachmias,V.T. and Pollard, T.D. (1992) The control of actin nucleotide exchange bythymosin beta 4 and profilin. A potential regulatory mechanism for actinpolymerization in cells. Mol. Biol. Cell 3, 1015–1024.
[20] Leyman, S. et al. (2009) Unbalancing the phosphatidylinositol-4,5-bisphosphate-cofilin interaction impairs cell steering. Mol. Biol. Cell 20,4509–4523.
[21] Lambrechts, A. et al. (2000) Profilin II is alternatively spliced, resulting inprofilin isoforms that are differentially expressed and have distinctbiochemical properties. Mol. Cell Biol. 20, 8209–8219.
[22] Van Troys, M., Dewitte, D., Verschelde, J.L., Goethals, M., Vandekerckhove, J.and Ampe, C. (2000) The competitive interaction of actin and PIP2 withactophorin is based on overlapping target sites: design of a gain-of-functionmutant. Biochemistry 39, 12181–12189.
[23] Gorbatyuk, V.Y., Nosworthy, N.J., Robson, S.A., Bains, N.P., Maciejewski, M.W.,Dos Remedios, C.G. and King, G.F. (2006) Mapping the phosphoinositide-binding site on chick cofilin explains how PIP2 regulates the cofilin-actininteraction. Mol. Cell 24, 511–522.
[24] Zhao, H., Hakala, M. and Lappalainen, P. (2010) ADF/cofilin bindsphosphoinositides in a multivalent manner to act as a PIP(2)-density sensor.Biophys. J. 98, 2327–2336.
[25] Yang, N., Higuchi, O., Ohashi, K., Nagata, K., Wada, A., Kangawa, K., Nishida, E.and Mizuno, K. (1998) Cofilin phosphorylation by LIM-kinase 1 and its role inRac-mediated actin reorganization. Nature 393, 809–812.
[26] Amano, T., Tanabe, K., Eto, T., Narumiya, S. and Mizuno, K. (2001) LIM-kinase 2induces formation of stress fibres, focal adhesions and membrane blebs,dependent on its activation by Rho-associated kinase-catalysedphosphorylation at threonine-505. Biochem. J. 354, 149–159.
[27] Toshima, J., Toshima, J.Y., Takeuchi, K., Mori, R. and Mizuno, K. (2001) Cofilinphosphorylation and actin reorganization activities of testicular protein kinase2 and its predominant expression in testicular Sertoli cells. J. Biol. Chem. 276,31449–31458.
[28] Polet, D., Lambrechts, A., Vandepoele, K., Vandekerckhove, J. and Ampe, C.(2007) On the origin and evolution of vertebrate and viral profilins. FEBS Lett.581, 211–217.
[29] Hornbeck, P.V., Kornhauser, J.M., Tkachev, S., Zhang, B., Skrzypek, E., Murray,B., Latham, V. and Sullivan, M. (2012) PhosphoSitePlus: a comprehensiveresource for investigating the structure and function of experimentallydetermined post-translational modifications in man and mouse. NucleicAcids Res. 40, D261–D270.
[30] Yoo, Y., Ho, H.J., Wang, C. and Guan, J.L. (2010) Tyrosine phosphorylation ofcofilin at Y68 by v-Src leads to its degradation through ubiquitin-proteasomepathway. Oncogene 29, 263–272.
[31] Paavilainen, V.O., Oksanen, E., Goldman, A. and Lappalainen, P. (2008)Structure of the actin-depolymerizing factor homology domain in complexwith actin. J. Cell Biol. 182, 51–59.
[32] Bonnette, P.C., Robinson, B.S., Silva, J.C., Stokes, M.P., Brosius, A.D., Baumann, A.and Buckbinder, L. (2010) Phosphoproteomic characterization of PYK2signaling pathways involved in osteogenesis. J. Proteomics 73, 1306–1320.
[33] Jorgensen, C. et al. (2009) Cell-specific information processing in segregatingpopulations of Eph receptor ephrin-expressing cells. Science 326, 1502–1509.
[34] Cao, L. et al. (2007) Quantitative time-resolved phosphoproteomic analysis ofmast cell signaling. J. Immunol. 179, 5864–5876.
[35] Luo, W. et al. (2008) Global impact of oncogenic Src on a phosphotyrosineproteome. J. Proteome Res. 7, 3447–3460.
[36] Yoshida, K., Yamaguchi, T., Natsume, T., Kufe, D. and Miki, Y. (2005) JNKphosphorylation of 14–3-3 proteins regulates nuclear targeting of c-Abl in theapoptotic response to DNA damage. Nat. Cell Biol. 7, 278–285.
[37] Brummer, T. et al. (2008) Phosphorylation-dependent binding of 14–3-3terminates signalling by the Gab2 docking protein. EMBO J. 27, 2305–2316.
[38] Birkenfeld, J., Betz, H. and Roth, D. (2003) Identification of cofilin and LIM-domain-containing protein kinase 1 as novel interaction partners of 14–3-3zeta. Biochem. J. 369, 45–54.
[39] Dreiza, C.M. et al. (2005) Transducible heat shock protein 20 (HSP20)phosphopeptide alters cytoskeletal dynamics. FASEB J. 19, 261–263.
[40] Gohla, A. and Bokoch, G.M. (2002) 14–3-3 regulates actin dynamics bystabilizing phosphorylated cofilin. Curr. Biol. 12, 1704–1710.
[41] Shao, J., Welch, W.J., Diprospero, N.A. and Diamond, M.I. (2008)Phosphorylation of profilin by ROCK1 regulates polyglutamine aggregation.Mol. Cell Biol. 28, 5196–5208.
[42] Lambrechts, A., Jonckheere, V., Dewitte, D., Vandekerckhove, J. and Ampe, C.(2002) Mutational analysis of human profilin I reveals a second PI(4,5)-P2binding site neighbouring the poly(L-proline) binding site. BMC Biochem. 3,12.
[43] Sohn, R.H., Chen, J., Koblan, K.S., Bray, P.F. and Goldschmidt-Clermont, P.J.(1995) Localization of a binding site for phosphatidylinositol 4,5-bisphosphateon human profilin. J. Biol. Chem. 270, 21114–21120.




















