
Historic, archived document Do not assume content reflects current ...

This article appeared in a journal published by Elsevier. The attachedcopy is furnished to the author for internal non-commercial researchand education use, including for instruction at the authors institution
and sharing with colleagues.
Other uses, including reproduction and distribution, or selling orlicensing copies, or posting to personal, institutional or third party
websites are prohibited.
In most cases authors are permitted to post their version of thearticle (e.g. in Word or Tex form) to their personal website orinstitutional repository. Authors requiring further information
regarding Elsevier’s archiving and manuscript policies areencouraged to visit:
http://www.elsevier.com/copyright

Author's personal copy
Journal of Biotechnology 142 (2009) 21–30
Contents lists available at ScienceDirect
Journal of Biotechnology
journa l homepage: www.e lsev ier .com/ locate / jb io tec
Phenotypic diversity of hydrogen production in chlorophycean algae reflectsdistinct anaerobic metabolisms
Jonathan E. Meusera, Gennady Ananyevb, Lauren E. Wittiga, Sergey Kosourovc, Maria L. Ghirardi c,Michael Seibert c, G. Charles Dismukesb, Matthew C. Posewitzd,∗
a Division of Environmental Science and Engineering, Colorado School of Mines, 1500 Illinois St., Golden, CO 80401, United Statesb Department of Chemistry & Environmental Institute, Princeton University, Princeton, NJ 08544, United Statesc Chemical and Biosciences Center, National Renewable Energy Laboratory, 1617 Cole Blvd., Golden, CO 80401-3393, United Statesd Department of Chemistry and Geochemistry, Colorado School of Mines, 1500 Illinois St., Golden, CO 80401, United States
a r t i c l e i n f o
Article history:Received 23 September 2008Received in revised form 1 January 2009Accepted 15 January 2009
Keywords:AlgaeAnoxiaBiofuelChlamydomonasHydrogenasePhotosynthesis
a b s t r a c t
Several species of green algae use [FeFe]-hydrogenases to oxidize and/or produce H2 during anoxia. Tofurther define unique aspects of algal hydrogenase activity, the well-studied anaerobic metabolismsof Chlamydomonas reinhardtii were compared with four strains of Chlamydomonas moewusii and aLobochlamys culleus strain. In vivo and in vitro hydrogenase activity, starch accumulation/degradation, andanaerobic end product secretion were analyzed. The C. moewusii strains showed the most rapid inductionof hydrogenase activity, congruent with high rates of starch catabolism, and anoxic metabolite accumu-lation. Intriguingly, we observed significant differences in morphology and hydrogenase activity in the C.moewusii strains examined, likely the result of long-term adaptation and/or genetic drift during culturemaintenance. Of the C. moewusii strains examined, SAG 24.91 showed the highest in vitro hydrogenaseactivity. However, SAG 24.91 produced little H2 under conditions of sulfur limitation, which is likely aconsequence of its inability to utilize exogenous acetate. In L. culleus, hydrogenase activity was minimalunless pulsed light was used to induce significant H2 photoproduction. Overall, our results demonstratethat unique anaerobic acclimation strategies have evolved in distinct green algae, resulting in differ-ential levels of hydrogenase activity and species-specific patterns of NADH reoxidation during anoxia.
© 2009 Elsevier B.V. All rights reserved.
1. Introduction
Water-oxidizing, phototrophic microorganisms have servedas exceptional model systems to examine photosynthetic path-ways and processes (Grossman, 2000; Gutman and Niyogi, 2004;Rochaix, 2002). Although these organisms are typically associ-ated with photosynthesis and oxic metabolism, it is becoming
Abbreviations: ACK, acetate kinase; ADH, alcohol dehydrogenase; ADHE,alcohol/aldehyde dehydrogenase; AIB, anaerobic induction buffer; DHAP, dihydrox-yacetone phosphate; FDX, ferredoxin; G3P, glycerol 3-phosphate; GAP, glyceralde-hyde 3-phosphate; GPD, glycerol 3-phosphate dehydrogenase/dihydroxyacetonephosphate reductase; GPP, glycerol 3-phosphatase; HYDA1/HYDA2, [FeFe]-hydrogenases; LDH, lactate dehydrogenase; LED, light emitting diode; mt, matingtype; MOPS, 3-morpholinopropanesulfonic acid; MV, methyl viologen; PAT, phos-photransacetylase; PDC, pyruvate decarboxylase; PFL, pyruvate-formate lyase; PFR,pyruvate-ferredoxin oxidoreductase; SSU rRNA, small subunit ribosomal RNA; TAP,tris-acetate-phosphate.
∗ Corresponding author. Tel.: +1 303 919 1605; fax: +1 303 273 3629.E-mail address: [email protected] (M.C. Posewitz).
increasingly evident that many species are able to withstandanoxia in ecosystems where O2 availability is limited due tomicrobial respiration, particularly at night (Tsygankov et al., 2002;Hemschemeier and Happe, 2005; Atteia et al., 2006; Steunou etal., 2006; Grossman et al., 2007; Mus et al., 2007). Consequently,many algae and cyanobacteria possess anaerobic metabolisms andenzymes, including hydrogenases, which reversibly reduce pro-tons to H2 gas (Kosourov et al., 2003; Boichenko et al., 2004;Kruse et al., 2005a,b; Rupprecht et al., 2006; Ghirardi et al.,2007; Seibert et al., 2008). The hydrogenase enzymes, which aretypically associated with strict anaerobes (Vignais et al., 2001;Vignais and Billoud, 2007), are able to interact with the photosyn-thetic electron transport chain under the appropriate physiologicalconditions, providing the means to couple photosynthesis to H2production (Weaver et al., 1980; Kruse et al., 2005a; Ghirardiet al., 2007). Currently, the prospect of renewable H2 produc-tion via water oxidation is the subject of extensive researchefforts in a variety of organisms (Melis et al., 2000; Ghirardiet al., 2005; Rupprecht et al., 2006; Melis, 2007; Seibert et al.,2008).
0168-1656/$ – see front matter © 2009 Elsevier B.V. All rights reserved.doi:10.1016/j.jbiotec.2009.01.015

Author's personal copy
22 J.E. Meuser et al. / Journal of Biotechnology 142 (2009) 21–30
Early investigations of hydrogenase activity in eukaryoticalgae provided intriguing insights into the diversity of algal H2metabolism and demonstrated variability in (a) dark and photosyn-thetic H2 production, (b) in vivo hydrogenase activity, and (c) theperiod of anaerobic adaptation required to induce maximal activ-ity amongst diverse chlorophycean algae (Frenkel and Rieger, 1951;Healey, 1970; Stuart and Gaffron, 1972; Ohta et al., 1987; Brand etal., 1989; Winkler et al., 2002). Although these studies surveyedsome aspects of H2 production, additional research is required tounderstand the metabolic and enzymatic context of the observeddifferences. It is also necessary to compare the H2-production activ-ity of phylogenetically distinct species under similar culture andassay conditions.
The majority of H2-production research in Chlorophyteshas focused primarily on the model organism, Chlamydomonasreinhardtii, despite the availability of at least 15,000 additional can-didate strains for comparative study (Andersen, 1992; Rupprechtet al., 2006). C. reinhardtii has emerged as a model organism due touseful genetic and physiological traits, and it is the most intensivelystudied and best understood algal system (Grossman et al., 2007;Merchant et al., 2007). In C. reinhardtii, O2 levels and/or cellularredox status contribute to the complex regulation of photosynthe-sis, mitorespiration, glycolysis, and chlororespiration, all of whichcan affect hydrogenase activity (Wykoff et al., 1998; Cournac etal., 2002; Antal et al., 2003; Doebbe et al., 2007; Posewitz et al.,2009). However, similar insights regarding the metabolic contextof hydrogenase activity and evaluation of H2-production capac-ity in other Chlorophyta are lacking. A comparative approach wastherefore undertaken to examine the unique aspects of anoxicmetabolism and hydrogenase activity in other green algae. Animproved understanding of the metabolic factors mediating robustH2 production and examination of the unique characteristics of thehydrogenase enzymes in these organisms are necessary to eluci-date the critical elements required for the rational engineering ofphototrophs optimized for H2 production. This will likely requirethe expression of recombinant hydrogenases in an appropriate hostdesigned to direct significant portions of metabolic electron fluxto H2 production. In this study, four strains of C. moewusii anda L. culleus strain were analyzed concomitantly with the C. rein-hardtii CC-124 control strain under identical conditions. These algaewere selected because C. moewusii and L. culleus were previouslyreported to have robust H2-production rates in surveys of eukary-otic algae compared to C. reinhardtii (Brand et al., 1989; Winkler etal., 2002). Several C. moewusii strains were studied as we observedthe occurrence of a palmelloid growth phenotype (unorganizedaggregates of non-motile cells often surrounded by extracellularpolysaccharide) and relatively low hydrogenase activities in thethree UTEX strains characterized. Under our experimental condi-tions, SAG 24.91 was the only C. moewusii strain that producedstrictly motile zoospores in liquid culture. Selected strains of C.moewusii, from two continents and over 60 years of separationbetween isolation, were examined to resolve whether robust H2production is the result of phylogenetically conserved or strain-specific traits. To further assess the H2-production capacity andanaerobic pathways used in these strains, we examined culturesunder identical conditions for the accumulation and degradationof intracellular starch, secretion of anaerobic end products; H2-photoproduction capacity; dark, anaerobic H2-production yields;and in vitro hydrogenase activity. We demonstrate significantlydifferent hydrogenase activities amongst the algae studied andassociated species-specific anaerobic metabolisms when culturedunder standardized conditions. Moreover, we observed significantdifferences in the metabolic activity amongst C. moewusii strainsobtained from different culture collections, which are likely theresult of long-term adaptation and/or genetic drift during culturemaintenance.
2. Materials and methods
2.1. Algae strains and growth conditions
C. reinhardtii, CC-124 (mt−, nit−), was obtained from the Chlamy-domonas Center at Duke University. Chlamydomonas moewusiistrains UTEX 10 (mt−), UTEX 97 (mt+) and UTEX 2018 (mt−);and Lobochlamys culleus (Pröschold et al., 2001) UTEX 1060 (mt−)were obtained from the UTEX Culture Collection at the Uni-versity of Texas at Austin, USA. C. moewusii SAG 24.91 (mt−)was obtained from the Culture Collection of Algae (SAG) at theUniversity of Göttingen, Germany. Sixty milliters algal cultureswere grown photoheterotrophically in TAP medium (Harris, 1989),pH 7.2, to mid-logarithmic phase at 25 ◦C, and were shaken at120 rpm in 250 ml Erlenmeyer flasks under constant cool-whitefluorescent illumination (80 �mol photons m−2 s−1), unless indi-cated otherwise. For sulfur deprivation, 1-l cultures were grownusing magnetic stirring in flat glass bottles (5-cm optical pathlength, Roux-type) under one-sided continuous cool-white fluores-cent illumination (120 �mol photons m−2 s−1) and bubbled with 3%CO2 in air that was filter-sterilized though 0.2 �m pore filters (Acro37 TF, Gelman Sciences, Inc. Ann Arbor, MI).
2.2. Molecular phylogeny
DNA was isolated from algal colonies on agar using the Ultr-aClean Soil DNA Isolation Kit (MoBio Laboratories, Carlsbad, CA,USA). Partial 18S SSU rRNA genes (>1350 bp), GenBank accessionnos. EU925395 (UTEX 97), EU925396 (SAG 24.91), EU925397 (CC-124), FJ211063 (UTEX 10), FJ211064 (UTEX 1060), were amplifiedfrom each alga using the eucaryote-specific primers 360FE and1492RE (Dawson and Pace, 2002). Sequences were aligned usingClustalW (Thompson et al., 1994) and further aligned manually inARB (Ludwig et al., 2004). The RAxML program (Stamatakis, 2006)was used to determine a best maximum likelihood tree (not shown)with bootstrap values greater than 80%.
2.3. Chlorophyll determination
Total Chl a and b was spectrophotometrically determined byextraction in 95% ethanol (Harris, 1989). Total chlorophyll wasselected as the standard for comparison to correlate hydrogenaseactivity with the light-absorbing capacity of the cultures. Under thegiven culturing conditions, total chlorophyll per cell was similaramongst the strains examined.
2.4. Anaerobic induction
Liquid cell cultures (50 ml) were concentrated by centrifugationat 5000 rpm for 5 min and resuspended in 0.1 volume of AIB (50 mMpotassium phosphate, pH 7.2, and 3 mM MgCl2). Concentrated cellswere transferred to 13-ml glass serum vials covered with aluminumfoil to exclude light. Vials were sealed with butyl rubber septaand purged with argon for 30 min to establish anaerobiosis. Cellswere incubated at 25 ◦C in a shaking (160 cycles min−1) water bath(Boekel Grant ORS200). All transfers of cells, solutions, and gaseswere made using argon-flushed, gas-tight syringes (Hamilton Com-pany, Reno, NV, USA).
2.5. H2-photoproduction measurements
2.5.1. Initial rates of H2 photoproductionCalibration of electrodes and H2-evolution measurements with
Clark electrodes were done as previously described (Ghirardi et al.,1997; Seibert et al., 1998). Briefly, H2- and O2-production rates weremeasured simultaneously with two electrodes (YSI Incorporated,

Author's personal copy
J.E. Meuser et al. / Journal of Biotechnology 142 (2009) 21–30 23
Yellow Springs, OH, USA) in a water-jacketed (25 ◦C) assay cham-ber. Anaerobically induced, concentrated cell suspensions (0.2 ml)were added to 1.8 ml of argon-purged MOPS buffer (50 mM, pH6.8). Initial rates of H2 photoproduction were determined by expo-sure of the cell suspension to 1000 �mol photons m−2 s−1 of heatfiltered (1%, w/v CuSO4; path length, 9.5 cm) actinic light (Nolan-Jenner 170-D). The H2 signal was amplified and analyzed usingDTVee software. Hydrogen-photoproduction rates were calculatedfrom the initial slope of the H2-dependent current increase afterillumination.
2.5.2. Pulse-illuminated H2-photoproduction kineticsAn ultra-sensitive Clark electrode for measuring dissolved H2
concentrations was custom built to measure algal H2-productionrates under flash illumination. The H2 electrode (smooth Pt/Ir(80%:20%) with a polarization voltage of +400 mV against aAg/AgCl reference electrode) can measure concentrations as low as1 × 10−8 M in a 7.5 �l sample. For pulse-illumination, we fabricateda LED (Luxeon Star, Inc., red 3 W LED) flasher. Hydrogen productionfrom cells grown as patches on TAP agar (1.25%) plates was observedwith 10-s light (200 �mol photons m−2 s−1) and 90-s dark cycles.Pulse-illumination data was acquired using a multifunctional dataacquisition card (PCI-6036E, National Instruments, Austin, TX, USA).Hydrogen-production rates were calculated from the current signalusing Faraday’s law. Gas chromatography was used for calibrationof the H2 electrode.
2.5.3. Chemochromic Pd/WO3 sensor measurement of relative H2photoproduction
Chemochromic sensors were prepared as described previously(Seibert et al., 1998, 2001a,b). Colonies of C. reinhardtii CC-124, C.moewusii, SAG 24.91, UTEX 10, and L. culleus UTEX 1060 were platedin triplicate on TAP agar plates and grown for 3 weeks under con-stant fluorescent illumination (∼15 �mol photons m−2 s−1, 25 ◦C).The agar plates were anaerobically adapted in a sealed jar (OxoidLtd., Hampshire, UK) by filling and vacuum evacuating nine times.Plates were stored under a slightly positively pressurized N2 atmo-sphere for 4 h. The anaerobic jar was transferred to an anaerobicchamber (COY Laboratory Products Inc., Grass Lake, MI, USA), wherethe cells were exposed to ∼380 �mol photons m−2 s−1 of diffused,incandescent light for 3 min. H2 photoproduction was detected byplacement of a chemochromic sensor in contact with cell coloniescovered by #2 filter paper (Whatmann Inc., Florham Park, NJ,USA).
2.6. Sulfur-deprived H2 photoproduction
Cells were harvested by centrifugation as described above,washed twice with TAP-S medium (sulfate anions replaced withchloride), and resuspended in TAP-S medium to a final concen-tration of approximately 15 �g Chl ml−1. Cultures resuspended inTAP-S were placed, unsealed, under continuous two-sided, cool-white fluorescent illumination (∼120 �mol photons m−2 s−1 oneach side) for the first 24 h. After 24 h and an initial period of O2evolution, the cultures were sealed to initiate anaerobiosis and H2collected in inverted graduated cylinders by water displacement.Biophysical and electrochemical changes during sulfur depriva-tion of identically treated C. reinhardtii CC-124 and C. moewusiiSAG 24.91 cultures were examined using a custom built bioreac-tor system fitted with appropriate sensors to monitor dissolved O2and redox potential (Kosourov et al., 2002). Analogous to methodspreviously described for establishing anaerobiosis in photoau-totrophically grown C. reinhardtii (Tsygankov et al., 2006), light wasreduced (60 �mol photons m−2 s−1 on each side) after 24 h of sulfurdeprivation where indicated.
2.7. In vitro hydrogenase activity
Methyl viologen assays were performed by transferring 200 �lof anaerobically adapted cells to 1 ml of MV solutions (10 mM MV,50 mM potassium phosphate, pH 6.9, and 0.2% Triton X-100) insealed, argon-purged, 13 ml vials. Hydrogen evolution was initi-ated by the addition of 100 �l of freshly prepared reduced sodiumdithionite (100 mM in 30 mM NaOH) solution and measured by gaschromatography (Hewlett Packard Series II 5890 fitted with a Restek5 Å Molecular Sieve 80/100 6’ 1/8” column and a thermal con-ductivity detector; argon was the carrier gas). The resulting signalwas integrated using ChemStation software, and H2 was quantifiedusing a standard curve generated using known quantities of H2. Thereaction was incubated in a shaking water bath at 25 ◦C.
2.8. Dark H2 production
During dark, anaerobic acclimation, H2 production was mea-sured by withdrawing 0.2 ml of headspace gas and analyzing by gaschromatography as above. Following each headspace gas analysis,samples were argon-purged for 5 min.
2.9. Dark, anaerobic end product quantitation
One milliter aliquots of anaerobically induced, concentrated cellsuspensions were collected for end product measurements andstarch analysis. At the indicated times cultures were centrifugedat 10,000 rpm for 5 min, and the supernatant and cell pelletsfrozen separately in liquid nitrogen. Secreted anaerobic end prod-ucts were quantified using HPLC analyses (Hewlett Packard Series1050 HPLC equipped with an Aminex HPX-87H [300 mm × 7.8 mm]ion exchange column). One-hundred microliters of thawed/filteredsupernatant was injected and then eluted using an isocratic mobilephase of 8 mM sulfuric acid at a flow rate of 0.5 ml min−1 at 45 ◦C.Products were quantified using an HPLC refractive index detector(Hewlett Packard Model 1047A), with the resulting retention peaksintegrated using ChemStation software and compared to calibra-tion curves generated using known standards of each metabolite(Sigma-Aldrich, St. Louis, MO, USA).
2.10. Starch analysis
Starch was extracted from the frozen cell pellets and quantitatedas glucose using a Sigma Glucose (HK) Assay Kit according to themanufacturer’s instructions, as described previously (Gfeller andGibbs, 1984; Kosourov et al., 2003). In brief, starch was extractedtwice in methanol. The insoluble starch was separated by centrifu-gation, resuspended in 100 mM sodium acetate (pH 4.5), sonicated,autoclaved, and digested overnight at 55 ◦C with 4 units ml−1 ofamylglucosidase (Sigma–Aldrich, St. Louis, MO, USA) to liberatemonomeric glucose. Glucose was measured by spectrophotomet-rically monitoring the enzymatic reduction of NAD to NADH (ε340
nm = 6.22 mmolar−1 cm−1).
3. Results
3.1. Anaerobic induction of H2 photoproduction
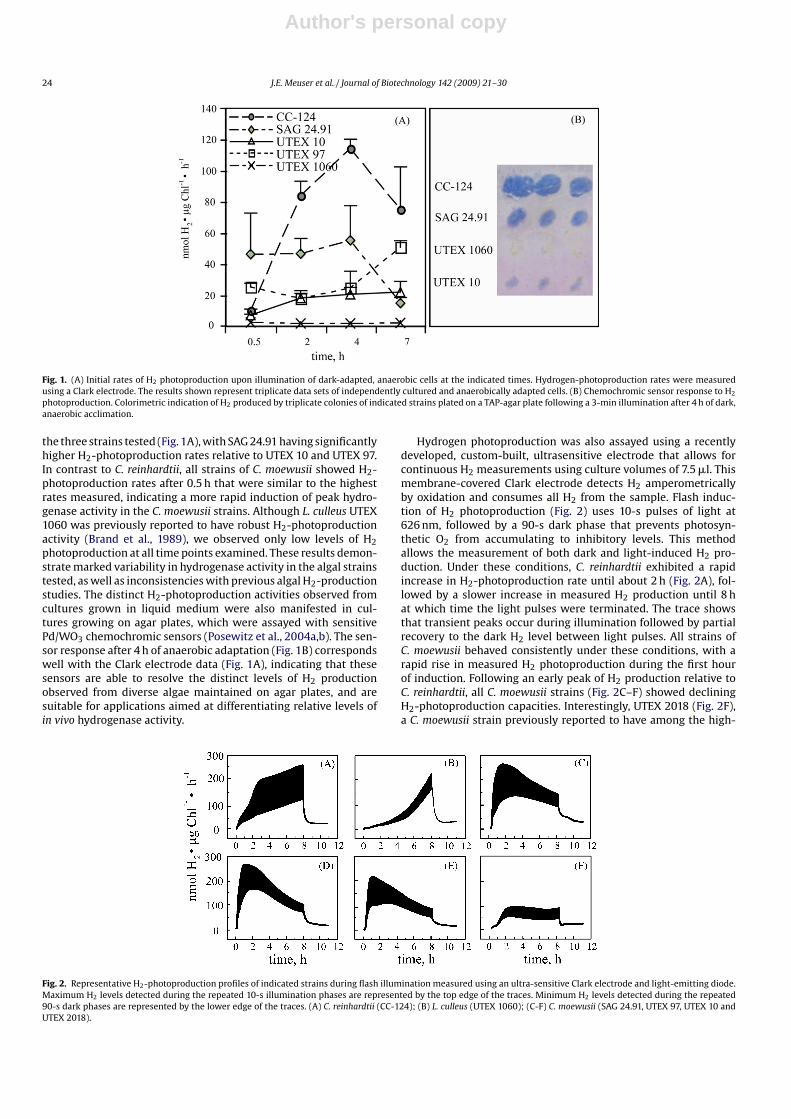
The induction of hydrogenase activity in C. reinhardtii has beenoptimized under conditions in which cultures are grown photo-heterotrophically in the presence of acetate (Ghirardi et al., 1997;Forestier et al., 2003). In agreement with previous studies, Clarkelectrode measurements show that C. reinhardtii has the highestrates of H2 photoproduction (Fig. 1A) after 2–4 h of dark, anaerobicacclimation (Forestier et al., 2003; Roessler and Lien, 1984). In C.moewusii, the maximal rates of H2 photoproduction varied among
Author's personal copy
24 J.E. Meuser et al. / Journal of Biotechnology 142 (2009) 21–30
Fig. 1. (A) Initial rates of H2 photoproduction upon illumination of dark-adapted, anaerobic cells at the indicated times. Hydrogen-photoproduction rates were measuredusing a Clark electrode. The results shown represent triplicate data sets of independently cultured and anaerobically adapted cells. (B) Chemochromic sensor response to H2
photoproduction. Colorimetric indication of H2 produced by triplicate colonies of indicated strains plated on a TAP-agar plate following a 3-min illumination after 4 h of dark,anaerobic acclimation.
the three strains tested (Fig. 1A), with SAG 24.91 having significantlyhigher H2-photoproduction rates relative to UTEX 10 and UTEX 97.In contrast to C. reinhardtii, all strains of C. moewusii showed H2-photoproduction rates after 0.5 h that were similar to the highestrates measured, indicating a more rapid induction of peak hydro-genase activity in the C. moewusii strains. Although L. culleus UTEX1060 was previously reported to have robust H2-photoproductionactivity (Brand et al., 1989), we observed only low levels of H2photoproduction at all time points examined. These results demon-strate marked variability in hydrogenase activity in the algal strainstested, as well as inconsistencies with previous algal H2-productionstudies. The distinct H2-photoproduction activities observed fromcultures grown in liquid medium were also manifested in cul-tures growing on agar plates, which were assayed with sensitivePd/WO3 chemochromic sensors (Posewitz et al., 2004a,b). The sen-sor response after 4 h of anaerobic adaptation (Fig. 1B) correspondswell with the Clark electrode data (Fig. 1A), indicating that thesesensors are able to resolve the distinct levels of H2 productionobserved from diverse algae maintained on agar plates, and aresuitable for applications aimed at differentiating relative levels ofin vivo hydrogenase activity.
Hydrogen photoproduction was also assayed using a recentlydeveloped, custom-built, ultrasensitive electrode that allows forcontinuous H2 measurements using culture volumes of 7.5 �l. Thismembrane-covered Clark electrode detects H2 amperometricallyby oxidation and consumes all H2 from the sample. Flash induc-tion of H2 photoproduction (Fig. 2) uses 10-s pulses of light at626 nm, followed by a 90-s dark phase that prevents photosyn-thetic O2 from accumulating to inhibitory levels. This methodallows the measurement of both dark and light-induced H2 pro-duction. Under these conditions, C. reinhardtii exhibited a rapidincrease in H2-photoproduction rate until about 2 h (Fig. 2A), fol-lowed by a slower increase in measured H2 production until 8 hat which time the light pulses were terminated. The trace showsthat transient peaks occur during illumination followed by partialrecovery to the dark H2 level between light pulses. All strains ofC. moewusii behaved consistently under these conditions, with arapid rise in measured H2 photoproduction during the first hourof induction. Following an early peak of H2 production relative toC. reinhardtii, all C. moewusii strains (Fig. 2C–F) showed decliningH2-photoproduction capacities. Interestingly, UTEX 2018 (Fig. 2F),a C. moewusii strain previously reported to have among the high-
Fig. 2. Representative H2-photoproduction profiles of indicated strains during flash illumination measured using an ultra-sensitive Clark electrode and light-emitting diode.Maximum H2 levels detected during the repeated 10-s illumination phases are represented by the top edge of the traces. Minimum H2 levels detected during the repeated90-s dark phases are represented by the lower edge of the traces. (A) C. reinhardtii (CC-124); (B) L. culleus (UTEX 1060); (C-F) C. moewusii (SAG 24.91, UTEX 97, UTEX 10 andUTEX 2018).

Author's personal copy
J.E. Meuser et al. / Journal of Biotechnology 142 (2009) 21–30 25
Table 1Volumetric H2 production under sulfur-deprivation conditions.
Alga Light regime Acetate uptake
Low/high Low/high/low
Lobochlamys culleus (UTEX 1060) 0 10 ml NoChlamydomonas moewusii (SAG 24.91) 0 10 ml NoChlamydomonas reinhardtii (CC-124) 120 ml 60 ml Yes
Comparison of two light regimes on the volumetric yield of H2 in indicated algal strains. Both regimes consisted of sulfur-replete growth under low light(120 �mol photons m−2 s−1 on one side) followed by high light (120–140 �mol photons m−2 s−1 on both sides) during the first 24 h following resuspension in TAP-S medium.After the first 24 h of sulfur deprivation, one light regime involved decreasing the light intensity (60 �mol photons m−2 s−1) to assist in the establishment of anaerobiosis.
est H2-photoproduction rates in a survey of diverse algae (Brand etal., 1989), underperformed other C. moewusii strains and showedextremely poor growth following axenic isolation from a rhizobialbacterium that was present in the sample obtained. L. culleus exhib-ited (Fig. 2B) substantial H2 accumulation only after long periods(>6 h) of flash acclimation. Both the dark H2 and light-induced H2production in L. culleus increased slowly until about 6 h, followedby a more rapid increase in H2 production. Possible sources for theobserved unique kinetic response in L. culleus are (a) a decreasingcapacity for H2 uptake, (b) increasing levels of reduced ferredoxin,or (c) increased hydrogenase expression and activity under condi-tions where reduction of the plastoquinone pool is enhanced byphotosynthetic electron transport during anaerobiosis.
3.2. Sulfur deprivation
Currently, sulfur deprivation in C. reinhardtii is the only reportedmechanism to obtain sustained H2 photoproduction in eukaryoticalgae (Melis et al., 2000). Therefore, the capacity of the other algaein this study to sustain H2 photoproduction under sulfur depri-vation was assayed. Using protocols typical of sulfur-deprived H2photoproduction in C. reinhardtii, CC-124 produced the expectedvolumetric yields of H2 (Table 1). However, none of the otheralgae evolved significant amounts of H2 under the sulfur-depletionconditions described. Interestingly, acetate has been shown to beimportant in establishing anaerobiosis and facilitating maximalH2 yields in C. reinhardtii (Kosourov et al., 2003, 2007; Fouchardet al., 2005; Kamp et al., 2008). Analysis of acetate levels in thegrowth medium following aerobic growth to mid-log phase indi-cated that L. culleus and all C. moewusii strains examined lackedthe capacity for acetate uptake. To further probe the absence ofsulfur-deprived H2 photoproduction, O2 and redox sensors wereused to compare the C. reinhardtii and C. moewusii phenotypes.C. reinhardtii resuspended in TAP-S medium achieved anaerobio-sis after approximately 30 h, followed by H2 photoproduction for26 h (data not shown). Concomitant O2 measurements of sulfur-deprived C. moewusii SAG24.91 showed only slowly declining O2levels, even after 80 h in TAP-S medium (data not shown). This datawas corroborated by concomitant redox potential measurementsin the same cultures.
Given these results, light conditions previously found to induceH2 photoproduction in autotrophically grown (lacking acetate)C. reinhardtii (Tsygankov et al., 2006) were utilized to achieveH2 photoproduction in cultures of C. moewusii SAG 24.91 and L.culleus in TAP-S medium (Table 1). This method of establishinganaerobiosis in photoautotrophic cultures involves exposing cellsresuspended in sulfur-deplete medium first to high levels of light(110–120 �mol photons m−2 s−1 on both sides), causing starch over-accumulation and reduced O2 evolution (Tsygankov et al., 2006),potentially by the inability of the algae to repair light-damagedPSII (Wykoff et al., 1998; Melis et al., 2000; Kosourov et al., 2007).A reduction in light intensity is then used to decrease photosyn-thetic O2 production and allow faster establishment of anaerobiosis.Although volumetric levels of sulfur-deprived H2 production was
observed in similarly treated C. moewusii (Table 1) in TAP-S medium,the levels were below those reported for similarly treated photoau-totrophic cultures of C. reinhardtii (Tsygankov et al., 2006).
3.3. In vitro hydrogenase activity
To evaluate the catalytic capacity of the selected strains to pro-duce H2 after dark, anoxic acclimation, in vitro hydrogenase activitywas measured using MV reduced with dithionite (Fig. 3). This assaycompares the capacity of the strains examined to evolve H2 anddoes not require in vivo metabolic pathways to provide reductantto hydrogenase via photosynthesis and/or fermentation. In contrastto the in vivo measurements, C. moewusii SAG 24.91 cell lysatesshowed significantly higher MV-catalyzed H2 evolution, exhibitinggreater than 3-fold hydrogenase activity than similarly treated C.reinhardtii cells. This is in accordance with previous studies show-ing greater in vitro hydrogenase activity in C. moewusii relative toC. reinhardtii (Kamp et al., 2008; Winkler et al., 2002). C. moewusiiUTEX 97 showed similar rates of MV-catalyzed H2 evolution fromcell lysates relative to C. reinhardtii despite significantly lower in vivorates of H2 photoproduction. Similar to the in vivo measurements, L.culleus exhibited low rates of in vitro hydrogenase activity followingdark anaerobic adaptation. It should be noted that comparative datafor all strains shown represent MV assays of hydrogenase activity at25 ◦C. The MV assay temperature significantly affects hydrogenaseactivity and substantially enhanced rates were recorded up to 52 ◦Cfrom C. reinhardtii cells (data not shown). Assay temperature is acritical consideration when comparing in vitro rates of hydrogenaseactivity to in vivo rates, and to other reported in vitro activities.
3.4. Dark H2 production and starch catabolism
As shown in Fig. 4, the rates of dark H2 production also var-ied greatly among the algae tested. All strains examined, with the
Fig. 3. In vitro hydrogenase activity of strains shown at the indicated times of dark,anaerobic acclimation. The results shown represent triplicate data sets of indepen-dently cultured and anaerobically adapted cells.

Author's personal copy
26 J.E. Meuser et al. / Journal of Biotechnology 142 (2009) 21–30
Fig. 4. Rates of dark, anoxic H2 production for indicated strains. The total H2 pro-duced between time points was used to calculate indicated rates. The results shownrepresent triplicate data sets of independently cultured and anaerobically adaptedcells.
exception of L. culleus, exhibited significant dark H2 productionimmediately after the onset of anaerobiosis. Similar to the inductionof H2-photoproduction activity using pulsed light, the C. moewusiistrains, particularly SAG 24.91, showed a rapid induction of hydro-genase activity followed by a decline in H2-production rates priorto 4 h of anaerobic acclimation. In contrast, C. reinhardtii was ableto sustain H2 production at higher rates for more extended peri-ods. Starch levels were quantified in each of the strains (data notshown) since starch catabolism is a likely source of reductant fordark H2 production. Relative to the three C. moewusii strains exam-ined, C. reinhardtii degraded starch more gradually; whereas starchcatabolism was extremely rapid in each of the C. moewusii strainsexamined. After 4 h of dark anoxia, C. reinhardtii had only catab-olized about half of the measured intracellular starch. In contrast,the C. moewusii strains exhibited rapid catabolism of starch reservesunder the same conditions, with less than one third of the initialstarch reserves remaining at the 0.5 h timepoint. Starch degrada-
tion, induction of hydrogenase activity, and anaerobic metabolicend product accumulation data are well correlated in C. reinhardtiiand C. moewusii. However, it should be noted that the end productsmeasured often slightly exceeded the stoichiometry of measuredstarch, indicating that the catabolism of protein or soluble sugarsmay also contribute to product yields. Overall, C. moewusii appearsto induce dark, anaerobic metabolism faster than C. reinhardtii;however, the rates of end product formation decline relative toC. reinhardtii after approximately 4 h of anoxia. Despite low lev-els of in vivo H2 production, L. culleus also consumed significantstarch reserves by the 0.5 h time point. L. culleus was unique inthat approximately half of the measured starch remained, evenafter 20 h of dark anoxia. In the case of L. culleus, it is presentlyunclear why significant levels of starch (measured as glucose) aredetected after prolonged anaerobiosis, or whether this accuratelyrepresents unmetabolized intracellular starch. It is conceivable thatthe origin of measured glucose is from unmetabolized extracellu-lar polysaccharide in this strain, which accumulates in significantquantities.
3.5. Anaerobic end products and anaerobic glycolytic pathways
In C. reinhardtii, the genome (Merchant et al., 2007) and exper-imental data (Grossman et al., 2007; Mus et al., 2007) indicatethe presence of diverse anaerobic pathways. For comparative pur-poses, secreted anaerobic metabolites were quantified from mid-to late-log cultures to elucidate distinct anaerobic acclimationpathways in the species examined (Fig. 5). As shown in earlierstudies, formate, acetate and ethanol are the primary anaerobicmetabolites observed in C. reinhardtii (Gfeller and Gibbs, 1984;Ohta et al., 1987; Kosourov et al., 2003; Mus et al., 2007). The for-mate:acetate:ethanol ratio observed for C. reinhardtii CC-124 wasapproximately 2:1:1 (Fig. 5A), consistent with previous reports(Gfeller and Gibbs, 1984; Kreuzberg, 1984). Ohta et al. (1987) andMus et al. (2007) have reported ratios of approximately 2:2:1for the respective metabolites, which likely reflect differences instrains, culturing, and/or assay conditions. Although the ratios ofmetabolites appear somewhat dynamic, the primary C. reinhardtii
Fig. 5. Products secreted from the indicated strains during anoxic acclimation after the specified periods of dark anaerobiosis. Data were collected concomitant with themeasurement of the dark, anaerobic H2-production rates (Fig. 4) and starch measurements. The results shown represent triplicate data sets of independently cultured andanaerobically adapted cells.

Author's personal copy
J.E. Meuser et al. / Journal of Biotechnology 142 (2009) 21–30 27
anaerobic metabolism end products are consistent in the litera-ture. In this study, cells were grown to a relatively high densityin air, but were not bubbled with an air/CO2 gas mixture. More-over, the acetate in the culture medium was completely consumed,and anaerobically adapted cells were vigorously agitated through-out the anaerobic induction protocol. Lastly H2 was purged fromthe headspace after each assay point to determine the rate of H2production between points (see Fig. 4), with the consequence thatthe partial pressure of H2 in the incubation vial was reduced. Asin an earlier report (Klein and Betz, 1978), the C. moewusii strainssecreted no formate. All C. moewusii strains clearly exhibited endproduct formation divergent from C. reinhardtii under the sameculturing and induction conditions. Acetate, glycerol, and ethanolwere the major dark, anaerobic end products, and were produced inan approximately 2:1:1 ratio after 4 h of anaerobiosis. Acetate andethanol appear earlier than glycerol during anoxia, possibly reflect-ing shifting metabolic needs or differential rates of secretion fromthe cell. Malate is also observed; however, significant accumula-tion is not observed until later times of anoxic acclimation (20 h).L. culleus also exhibited mixed alcohol and organic acid formation,with acetate, ethanol and formate being the predominant productsaccumulated. Lactate was secreted slowly over 20 h of anaerobiosisin L. culleus, a product that was observed in only trace amounts inC. reinhardtii and was not observed in C. moewusii. Low levels ofmalate, succinate and glycerol were also secreted.
4. Discussion
Hydrogen production in green algae is likely an ancientmetabolism with diverse phenotypes evolving across variousspecies over the last billion years or more of algal evolution. Thoughfew diversity studies exist, reports of hydrogenase activity (Brandet al., 1989; Winkler et al., 2002; Boichenko et al., 2004; Skjåneset al., 2008) combined with molecular-based phylogeny (Lewis etal., 1992; Pröschold et al., 2001; Lewis and McCourt, 2004) pro-vide strong evidence that hydrogenase activity is an ancestral traitin the Chlorophyta with secondary losses in some taxa. For exam-ple, our own examination of 6 strains of Dunaliella verified earlierreports (Winkler et al., 2002) of the absence of hydrogenase activ-ity in this genus (data not shown). Morphological and moleculardata place the algae under investigation in three distinct cladesspanning the CW lineage of Chlorophyceae (Lewis et al., 1992;Pröschold et al., 2001; Lewis et al., 2004). Our own analysis ofpartial (>1350 bp) SSU (18S) rRNA gene amplicons from each algais consistent with the suggested taxonomy. C. moewusii strainsUTEX 10, UTEX 97 and SAG 24.91 formed a distinct clade in amaximum likelihood tree (bootstrap >80%) with >99% sequenceidentity, while greater than species-level variations of 18S rRNAgenes (<95% identity) was found among C. moewusii, C. reinhardtiiand L. culleus. Morphologically, C. moewusii (Chlamydella clade)share a lateral position of two pyrenoids compared to the basal posi-tion of a single pyrenoid in C. reinhardtii (Euchlamydomonas clade)(Gerloff, 1940; Pröschold et al., 2001; Harris, personal communi-cation). Though isogamous, L. culleus, on the basis of chloroplastperforations/incisions and molecular evidence, has been groupedclose to oogamous taxa (Oogamochlamys clade) (Pröschold et al.,2001).
Reports of high hydrogenase activity in C. moewusii (Brand et al.,1989; Winkler et al., 2002) provided the incentive for examining thebiological H2-production phenotype and concomitant anaerobicpathways relative to C. reinhardtii. In this study, we examined fourC. moewusii isolates under identical culture and assay conditions.C. moewusii UTEX 10, earlier known as C. eugametos, was isolatedby Moewus in Germany in 1929 (Gowans, 1960; Harris, 1989).Almost two decades later (1948), Provasoli isolated C. moewusii
UTEX 97, an isolate infertile with UTEX 10, in the United States(Harris, 1989). In our hands, these earlier isolates exhibited palmel-loid morphology and far slower growth rates on both Brand andWright and TAP media than C. moewusii SAG 24.91. The mainte-nance of two of the three C. moewusii strains for over 60 yearsunder conditions favoring slow growth has a likely relationship tothe observed phenotypes. Of the four C. moewusii examined, onlyC. moewusii SAG 24.91, isolated by Schlösser in Germany in 1997,exhibited robust growth and exemplary autotrophic H2-productionphenotypes. Despite differential growth and variable hydrogenaseactivity, the similar anaerobic metabolisms shown provide newevidence that all three taxa are true C. moewusii. This data lendssupport for examination of other recent isolates of C. moewusii andfor continued bioprospecting directly from nature, or the use ofrecently accessed strains to avoid strain adaptation and/or geneticdrift during long-term culturing.
Despite significant hydrogenase activity in SAG 24.91, our resultsprovide further evidence that acetate uptake is important in estab-lishing anaerobiosis during sulfur deprivation. In C. reinhardtii, thefirst 40 h of sulfur deprivation is marked by rapid acetate uptake(Kosourov et al., 2003; Fouchard et al., 2005), which likely sup-port the levels of respiration necessary to cause a transition toanaerobiosis in the light (Melis et al., 2000; Fouchard et al., 2005;Grossman et al., 2007). Thus, under standard sulfur deprivationconditions, an anaerobic transition was not observed in the obli-gate autotroph C. moewusii. In a recent survey of green algae,only those strains with both hydrogenase activity and the abil-ity to utilize acetate achieved anaerobiosis rapidly and evolvedvolumetric amounts of H2 during sulfur deprivation (Skjånes etal., 2008), paralleling our results with C. moewusii and L. culleus.However, in our hands C. moewusii could be induced to pro-duce volumetric quantities of H2 using conditions developed forachieving anaerobiosis in photoautotrophically grown C. reinhardtii(Tsygankov et al., 2006). Fouchard et al. (2005) have shown thatacetate is not requisite for starch accumulation during sulfur depri-vation when bicarbonate is provided. However, even followingstarch accumulation and degradation under these conditions, onlyacetate or DCMU could stimulate sustained anaerobiosis. Thus, itis likely that improved H2 photoproduction yields in C. moewusiiduring sulfur deprivation will require unique physiological orgenetic approaches for establishing and sustaining anaerobio-sis.
Both C. moewusii and L. culleus produced species-specific anaer-obic end products that are distinct from those observed in C.reinhardtii. Although C. reinhardtii appears to encode an extensivediversity of potential anaerobic pathways (Grossman et al., 2007;Mus et al., 2007; Posewitz et al., 2009), analysis of secreted productssuggests that some of these are active only under certain acclima-tion conditions, as demonstrated at distinct pHs (Kreuzberg, 1984;Kosourov et al., 2003). L. culleus showed minimal levels of dark H2production (Fig. 4) and elevated production of ethanol and organicacids (Fig. 5) relative to the other strains tested, indicating alter-native NADH reoxidation pathways. High levels of ethanol wereobserved at the earliest timepoint (0.5 h) without further signifi-cant accumulation. In contrast, formate and acetate accumulatedat similar rates throughout the course of dark anaerobiosis, evi-dence for formate formation via PFL followed by acetate productionfrom acetyl-coA. The observed secretion of lactate, malate, suc-cinate, and glycerol, products more reduced than pyruvate andnot produced by C. reinhardtii under the same conditions, likelyallows NADH reoxidation for redox balance during anaerobiosis inL. culleus.
The dark, anaerobic metabolisms observed in C. moewusii alsodiverged from those observed in C. reinhardtii. The production ofglycerol by C. moewusii likely provides a substantial redox sink dur-ing dark anaerobiosis, as is observed in yeast (Wang et al., 2001),

Author's personal copy
28 J.E. Meuser et al. / Journal of Biotechnology 142 (2009) 21–30
Fig. 6. Endogenous anoxic pathways available to Chlamydomonas based on the C. reinhardtii genome and observed product secretion (modified from Posewitz et al., 2009).Partitioning of flux from pyruvate to the indicated anaerobic pathways is based on the putative redox and ATP requirements of the cell. Two anaerobic pathways to acetyl-CoAfrom pyruvate have been established. The decarboxylation of pyruvate and reduction of FDX by PFR leads to electron donation to the [FeFe]-hydrogenase(s) (HYDA1 and/orHYDA2) and yields acetyl-CoA. PFL can cleave pyruvate yielding acetyl-CoA and formate. The reoxidation of NADH below pyruvate can occur via two distinct paths to ethanol,the PDC/ADH pathway and the two-step ADHE pathway from acetyl-CoA. The production of lactate may also occur, catalyzed by LDH. The production of glycerol in C. reinhardtiiis a minor path compared to C. moewusii under the same conditions, though glycerol production by GPD and GPP provides an additional mechanism for anaerobic NADHreoxidation. The stoichiometry shown is based on pyruvate equivalents.
while in C. reinhardtii this represents only a minimally active anaer-obic pathway. Orthologs of GPD and GPP, providing a putativepathway for glycerol production conserved in chlorophycean algae(Fig. 6), have been identified in Dunaliella salina (He et al., 2007) andthe C. reinhardtii genome. Although ratios of anaerobic end productsin C. moewusii differ from those in C. reinhardtii, theoretical stoicho-metric redox balance is still achieved. The metabolism of pyruvateto acetate via ACK and PTA yields ATP but does not reoxidize NADH,requiring other pathways to reoxidize one NADH per acetate to sus-tain glycolysis. Ethanol production via ADHE oxidizes two NADHmolecules, and glycerol production oxidizes one NADH. In the for-mation of glycerol, glycolytic NADH is not formed as GAP is notoxidized. Thus, the 2:1:1 ratio of acetate:glycerol:ethanol (Fig. 5)observed in C. moewusii could yield a balanced pyridine pool usingestablished metabolic pathways.
C. moewusii does not secrete formate under the conditions used,suggesting a lack of PFL activity. An unlikely alternative possibilityis that formate is further metabolized by a formate dehydroge-nase. While the absence of a PFL pathway might be expected toimprove dark H2 production by routing more reductant through thePFR pathway, this was not observed in C. moewusii. Interestingly,formate can reversibly inhibit O2 evolution in isolated thylakoids(Shevela et al., 2007), potentially by PSII donation or inhibition.Thus, intracellular formate may contribute some enhancement ofH2 photoproduction under conditions of sulfur deprivation in C.
reinhardtii by suppressing O2 evolution and/or providing supple-mental reductant.
In summary, our results demonstrate significant differences inanoxic metabolism in selected chlorophycean algae. When culturedunder standardized conditions, distinct hydrogenase activities andmetabolite secretion profiles are observed, showing that eachspecies has evolved unique strategies to maintain redox bal-ance during anoxia. Hydrogen, ethanol, acetate, glycerol, lactate,malate and formate each have relevant uses in the areas ofbioenergy and biomaterials. The ability of some phototrophicorganisms to perform photosynthesis, as well as the anaerobiccatabolism of endogenous substrates such as starch, presents anattractive opportunity to modulate cellular metabolism for the effi-cient conversion of sunlight into a variety of renewable energycarriers. Moreover, the distinct enzymatic features observed insome organisms could be leveraged to engineer phototrophs forthe improved production of targeted metabolites using recom-binant techniques that take advantage of the unique catalyticproperties of native or evolved enzymes (Nagy et al., 2007).The extraordinary metabolic and enzymatic features observed inwater-oxidizing photosynthetic microorganisms is just now beingappreciated in detail, and new insights into the effective useof these organisms in renewable energy strategies will continueto emerge as this large class of organisms is examined morethoroughly.

Author's personal copy
J.E. Meuser et al. / Journal of Biotechnology 142 (2009) 21–30 29
Acknowledgements
This work was supported by the United States Air Force Officeof Scientific Research under AFOSR grant FA9550-05-1-0365 (toJEM, GA, LEW, GCD, MCP), the United States National Air andSpace Administration (NASA) under the Graduate Student ResearchProgram (GSRP) grant #NNG05GL52H (to JEM, MCP), and theU.S. Department of Energy Hydrogen Program under Contract No.DEAC36-99GO10337 with the National Renewable Energy Labora-tory (MLG, MS).
The authors acknowledge thoughtful discussions with Dr. JohnSpear and Pin Ching Maness. The authors would also like to thankDr. John Cushman for providing 6 strains of Dunaliella and Dr. Eliz-abeth Harris for bibliographic assistance and helpful discussions inthe preparation of this manuscript. Appreciation is also extendedto Alex Trujillo for assistance in the preparation of Fig. 6.
References
Andersen, R.A., 1992. Diversity of eukaryotic algae. Biodivers. Conserv. 1, 267–292.Antal, T.K., Krendeleva, T.E., Laurinavichene, T.V., Makarova, V.V., Ghirardi, M.L.,
Rubin, A.B., Tsygankov, A.A., Seibert, M., 2003. The relationship between the pho-tosystem 2 activity and hydrogen production in sulfur deprived Chlamydomonasreinhardtii cells. Biochim. Biophys. Acta 1607, 153–160.
Atteia, A., van Lis, R., Gelius-Dietrich, G., Adrait, A., Garin, J., Joyard, J., Rolland, N.,Martin, W., 2006. Pyruvate formate-lyase and a novel route of eukaryotic ATPsynthesis in Chlamydomonas mitochondria. J. Biol. Chem. 281, 9909–9918.
Boichenko, V.A., Greenbaum, E., Seibert, M., 2004. Hydrogen production by photo-synthetic microorganisms. In: Archer, M.D., Barber, J. (Eds.), Molecular to GlobalPhotosynthesis: Photoconversion of Solar Energy. Imperial College Press, Lon-don, pp. 397–452.
Brand, J.J., Wright, J.N., Lien, S., 1989. Hydrogen production by eukaryotic algae.Biotechnol. Bioeng. 33, 1482–1488.
Cournac, L., Latouche, G., Cerovic, Z., Redding, K., Ravenel, J., Peltier, G., 2002. In vitrointeractions between photosynthesis, mitorespiration, and chlororespiration inChlamydomonas reinhardtii. Plant Physiol. 129, 1921–1928.
Dawson, S.C., Pace, N.R., 2002. Novel kingdom-level eukaryotic diversity in anoxicenvironments. Proc. Natl. Acad. Sci. U.S.A. 99, 8324–8329.
Doebbe, A., Rupprecht, J., Beckmann, J., Mussgnug, J.H., Hallmann, A., Hankamer, B.,Kruse, O., 2007. Functional integration of the HUP1 hexose symporter gene intothe genome of C. reinhardtii: impacts on biological H2 production. J. Biotechnol.131, 27–33.
Forestier, M., King, P., Zhang, L., Posewitz, M., Schwarzer, S., Happe, T., Ghirardi,M.L., Seibert, M., 2003. Expression of two [Fe]-hydrogenases in Chlamydomonasreinhardtii under anaerobic conditions. Eur. J. Biochem. 270, 2750–2758.
Fouchard, S., Hemschemeier, A., Caruana, A., Pruvost, J., Legrand, J., Happe, T., Peltier,G., Cournac, L., 2005. Autotrophic and mixotrophic hydrogen photoproduction insulfur-deprived Chlamydomonas cells. Appl. Environ. Microbiol. 71, 6199–6205.
Frenkel, A.W., Rieger, C., 1951. Photoreduction in algae. Nature 167, 1030.Gerloff, J., 1940. Beiträge zur kenntnis der variabilität und systematik der gattung
Chlamydomonas. In: Schaudinn, F., Hartmann, M., Pascher, A. (Eds.), Archiv fürProtistenkunde Bd. XCIV, pp. 311–502.
Gfeller, R.P., Gibbs, M., 1984. Fermentative metabolism of Chlamydomonas reinhardtii.I. Analysis of fermentative products from starch in dark and light. Plant Physiol.75, 212–218.
Ghirardi, M.L., King, P.W., Posewitz, M.C., Maness, P.C., Fedorov, A., Kim, K., Cohen,J., Schulten, K., Seibert, M., 2005. Approaches to developing biological H(2)-photoproducing organisms and processes. Biochem. Soc. Trans. 33, 70–72.
Ghirardi, M.L., Posewitz, M.C., Maness, P.-C., Dubini, A., Yu, J., Seibert, M., 2007. Hydro-genases and hydrogen photoproduction in oxygenic photosynthetic organisms.Annu. Rev. Plant Biol. 58, 71–91.
Ghirardi, M.L., Togasaki, R.K., Seibert, M., 1997. Oxygen sensitivity of algal hydrogenproduction. Appl. Biochem. Biotechnol. 63 (65), 141–151.
Gowans, C.S., 1960. Some genetic investigations on Chlamydomonas eugametos. Z.Vererb. 91, 63–73.
Grossman, A.R., 2000. Chlamydomonas reinhardtii and photosynthesis: genetics togenomics. Curr Opin. Plant Biol. 3, 132–137.
Grossman, A.R., Croft, M., Gladyshev, V.N., Merchant, S.S., Posewitz, M.C., Prochnik,S., Spalding, M.H., 2007. Novel metabolism in Chlamydomonas through the lensof genomics. Curr. Opin. Plant Biol. 10, 190–198.
Gutman, B.L., Niyogi, K.K., 2004. Chlamydomonas and Arabidopsis: a dynamic duo.Plant Physiol. 135, 607–610.
Harris, E.H., 1989. The Chlamydomonas Sourcebook: A Comprehensive Guide toBiology and Laboratory Use. Academic Press, San Diego.
He, Q., Qiao, D., Bai, L., Zhang, Q., Yang, W., Li, Q., Cao, Y., 2007. Cloning and character-ization of a plastidic glycerol 3-phosphate dehydrogenase cDNA from Dunaliellasalina. J. Plant Physiol. 164, 214–220.
Healey, F.P., 1970. Hydrogen evolution by several algae. Planta 91, 220–226.Hemschemeier, A., Happe, T., 2005. The exceptional photofermentative hydrogen
metabolism of the green alga Chlamydomonas reinhardtii. Biochem. Soc. Trans.33, 39–41.
Kamp, C., Silakov, A., Winkler, M., Reijerse, E.J., Lubitz, W., Happe, T., 2008. Isola-tion and first EPR characterization of the [FeFe]-hydrogenases from green algae.Biochim. Biophys. Acta 1777, 410–416.
Klein, U., Betz, A., 1978. Fermentative metabolism of hydrogen-evolving Chlamy-domonas moewusii. Plant Physiol. 61, 953–956.
Kosourov, S., Patrusheva, E., Ghirardi, M.L., Seibert, M., Tsygankov, A., 2007.A comparison of hydrogen photoproduction by sulfur-deprived Chlamy-domonas reinhardtii under different growth conditions. J. Biotechnol. 128,776–787.
Kosourov, S., Seibert, M., Ghirardi, M.L., 2003. Effects of extracellular pH on themetabolic pathways in sulfur-deprived H2-producing Chlamydomonas rein-hardtii cultures. Plant Cell Physiol. 44, 146–155.
Kosourov, S., Tsygankov, A., Seibert, M., Ghirardi, M.L., 2002. Sustained hydrogenphotoproduction by Chlamydomonas reinhardtii: effects of culture parameters.Biotechnol. Bioeng. 78, 731–740.
Kreuzberg, K., 1984. Starch fermentation via a formate producing pathway in Chlamy-domonas reinhardii, Chlorogonium elongatum and Chlorella fusca. Physiol. Plant.61, 87–94.
Kruse, O., Rupprecht, J., Mussgnug, J.H., Dismukes, G.C., Hankamer, B., 2005a. Pho-tosynthesis: a blueprint for solar energy capture and biohydrogen productiontechnologies. Photochem. Photobiol. Sci. 4, 957–970.
Kruse, O., Rupprecht, J., Bader, K.-P., Thomas-Hall, S., Schenk, P.M., Finazzi, G., Han-kamer, B., 2005b. Improved photobiological H2 production in engineered greenalgal cells. J. Biol. Chem. 280, 34170–34177.
Lewis, L.A., McCourt, R.M., 2004. Green algae and the origin of land plants. Am. J. Bot.91, 1535–1556.
Lewis, L.A., Wilcox, L.W., Fuerst, P.A., Floyd, G.L., 1992. Concordance of molecularand ultrastructural data in the study of zoosporic chlorococcalean green algae.J. Phycol. 28, 375–380.
Ludwig, W., Strunk, O., Westram, R., Richter, L., Meier, H., Yadhukumar, Buch-ner, A., Lai, T., Steppi, S., Jobb, G., Forster, W., Brettske, I., Gerber, S., Ginhart,A.W., Gross, O., Grumann, S., Hermann, S., Jost, R., Konig, A., Liss, T., Lussmann,R., May, M., Nonhoff, B., Reichel, B., Strehlow, R., Stamatakis, A., Stuckmann,N., Vilbig, A., Lenke, M., Ludwig, T., Bode, A., Schleifer, K.-H., 2004. ARB:a software environment for sequence data. Nucleic Acids Res. 32, 1363–1371.
Melis, A., 2007. Photosynthetic H2 metabolism in Chlamydomonas reinhardtii (uni-cellular green algae). Planta 226, 1075–1086.
Melis, A., Zhang, L., Forestier, M., Ghirardi, M.L., Seibert, M., 2000. Sustained pho-tobiological hydrogen gas production upon reversible inactivation of oxygenevolution in the green alga Chlamydomonas reinhardtii. Plant Physiol. 122,127–136.
Merchant, S.S., Prochnik, S.E., Vallon, O., Harris, E.H., Karpowicz, S.J., Witman, G.B.,Terry, A., Salamov, A., Fritz-Laylin, L.K., Marechal-Drouard, L., Marshall, W.F., Qu,L.-H., Nelson, D.R., Sanderfoot, A.A., Spalding, M.H., Kapitonov, V.V., Ren, Q., Fer-ris, P., Lindquist, E., Shapiro, H., Lucas, S.M., Grimwood, J., Schmutz, J., Cardol, P.,Cerutti, H., Chanfreau, G., Chen, C.-L., Cognat, V., Croft, M.T., Dent, R., Dutcher, S.,Fernandez, E., Fukuzawa, H., Gonzalez-Ballester, D., Gonzalez-Halphen, D., Hall-mann, A., Hanikenne, M., Hippler, M., Inwood, W., Jabbari, K., Kalanon, M., Kuras,R., Lefebvre, P.A., Lemaire, S.D., Lobanov, A.V., Lohr, M., Manuell, A., Meier, I.,Mets, L., Mittag, M., Mittelmeier, T., Moroney, J.V., Moseley, J., Napoli, C., Nedelcu,A.M., Niyogi, K., Novoselov, S.V., Paulsen, I.T., Pazour, G., Purton, S., Ral, J.-P.,Riano-Pachon, D.M., Riekhof, W., Rymarquis, L., Schroda, M., Stern, D., Umen,J., Willows, R., Wilson, N., Zimmer, S.L., Allmer, J., Balk, J., Bisova, K., Chen, C.-J.,Elias, M., Gendler, K., Hauser, C., Lamb, M.R., Ledford, H., Long, J.C., Minagawa, J.,Page, M.D., Pan, J., Pootakham, W., Roje, S., Rose, A., Stahlberg, E., Terauchi, A.M.,Yang, P., Ball, S., Bowler, C., Dieckmann, C.L., Gladyshev, V.N., Green, P., Jorgensen,R., Mayfield, S., Mueller-Roeber, B., Rajamani, S., Sayre, R.T., Brokstein, P., et al.,2007. The Chlamydomonas genome reveals the evolution of key animal and plantfunctions. Science 318, 245–250.
Mus, F., Dubini, A., Seibert, M., Posewitz, M.C., Grossman, A.R., 2007. Anaerobicacclimation in Chlamydomonas reinhardtii: anoxic gene expression, hydrogenaseinduction, and metabolic pathways. J. Biol. Chem. 282, 25475–25486.
Nagy, L.E., Meuser, J.E., Plummer, S., Seibert, M., Ghirardi, M.L., King, P.W., Ahmann,D., Posewitz, M.C., 2007. Application of gene-shuffling for the rapid generationof novel [FeFe]-hydrogenase libraries. Biotechnol. Lett. 29, 421–430.
Ohta, S., Miyamoto, K., Miura, Y., 1987. Hydrogen evolution as a consumption mode ofreducing equivalents in green algal fermentation. Plant Physiol. 83, 1022–1026.
Posewitz, M.C., King, P.W., Smolinski, S.L., Zhang, L., Seibert, M., Ghirardi, M.L., 2004a.Discovery of two novel radical s-adenosylmethionine proteins required for theassembly of an active [Fe] hydrogenase. J. Biol. Chem. 279, 25711–25720.
Posewitz, M.C., Smolinski, S.L., Kanakagiri, S., Melis, A., Seibert, M., Ghirardi, M.L.,2004b. Hydrogen photoproduction is attenuated by disruption of an isoamylasegene in Chlamydomonas reinhardtii. Plant Cell 16, 2151–2163.
Posewitz, M.C., Dubini, A., Meuser, J.E., Seibert, M., Ghirardi, M.L., 2009. Hydroge-nases, Hydrogen Production and Anoxia. In: Harris, E.H., Stern, D., Witman, G.(Eds.), The Chlamydomonas Sourcebook: Organellar and Metabolic Processes 2.Academic Press, San Diego, pp. 217–246.
Pröschold, T., Marin, B., Schlosser, U.G., Melkonian, M., 2001. Molecular phy-logeny and taxonomic revision of Chlamydomonas (Chlorophyta). I. emendationof Chlamydomonas ehrenberg and Chloromonas gobi, and description of Oog-amochlamys gen. nov. and Lobochlamys gen. nov. Protist 152, 265–300.
Rochaix, J.D., 2002. Chlamydomonas, a model system for studying the assembly anddynamics of photosynthetic complexes. FEBS Lett. 529, 34–38.
Roessler, P.G., Lien, S., 1984. Activation and de novo synthesis of hydrogenase inChlamydomonas. Plant Physiol. 76, 1086–1089.

Author's personal copy
30 J.E. Meuser et al. / Journal of Biotechnology 142 (2009) 21–30
Rupprecht, J., Hankamer, B., Mussgnug, J.H., Ananyev, G., Dismukes, C., Kruse, O.,2006. Perspectives and advances of biological H2 production in microorganisms.Appl. Microbiol. Biotechnol. 72, 442–449.
Seibert, M., Benson, D.K., Flynn, T., Inventors, 2001a. Method and Apparatus for RapidBiohydrogen Phenotypic Screening of Microorganisms Using a ChemochromicSensor. August 21. US Patent 6,277,589.
Seibert, M., King, P.W., Posewitz, M.C., Melis, A., Ghirardi, M.L., 2008. Photosyntheticwater-splitting for hydrogen production. In: Wall, J., Harwood, C.S., Demain, A.(Eds.), Bioenergy. ASM Press, Washington, DC, pp. 273–291.
Seibert, M., Flynn, T., Benson, D., Tracy, E., Ghirardi, M.L., 1998. Development ofselection and screening procedures for rapid identification of H2-producingalgal mutants with increased O2 tolerance. In: Zaborsky, O.R. (Ed.), Biohydrogen.Plenum Press, NY, pp. 227–234.
Seibert, M., Flynn, T., Ghirardi, M.L., 2001b. Strategies for improving oxygen toleranceof algal hydrogen production. In: Miyake, J., Matsunaga, T., San Pietro, A. (Eds.),Biohydrogen II. Elsevier, The Netherlands, pp. 67–77.
Shevela, D., Klimov, V., Messinger, J., 2007. Interactions of photosystem II with bicar-bonate, formate and acetate. Photosynth. Res. 94, 247–264.
Skjånes, K., Knutsen, G., Källqvist, T., Lindblad, P., 2008. H2 production from marineand freshwater species of green algae during sulfur deprivation and considera-tions for bioreactor design. Int. J. Hydrogen Energy 33, 511–521.
Stamatakis, A., 2006. RAxML-VI-HPC: maximum likelihood-based phylogeneticanalyses with thousands of taxa and mixed models. Bioinformatics 22,2688–2690.
Steunou, A.-S., Bhaya, D., Bateson, M.M., Melendrez, M.C., Ward, D.M., Brecht, E.,Peters, J.W., Kuhl, M., Grossman, A.R., 2006. In Situ analysis of nitrogen fixationand metabolic switching in unicellular thermophilic cyanobacteria inhabitinghot spring microbial mats. Proc. Natl. Acad. Sci. U.S.A. 103, 2398–2504.
Stuart, T.S., Gaffron, H., 1972. The mechanism of hydrogen photoproduction by sev-eral algae. Planta 106, 91–100.
Thompson, J.D., Higgins, D.G., Gibson, T.J., 1994. CLUSTAL W: improving the sensitiv-ity of progressive multiple sequence alignments through sequence weighting,position specific gap penalties and weight matrix choice. Nucleic Acids Res. 22,4673–4680.
Tsygankov, A., Kosourov, S., Seibert, M., Ghirardi, M.L., 2002. Hydrogen photo-production under continuous illumination by sulfur-deprived, synchronousChlamydomonas reinhardtii cultures. Int. J. Hydrogen Energy 27, 1239–1244.
Tsygankov, A.A., Kosourov, S.N., Tolstygina, I.V., Ghirardi, M.L., Seibert, M., 2006.Hydrogen production by sulfur-deprived Chlamydomonas reinhardtii under pho-toautotrophic conditions. Int. J. Hydrogen Energy 31, 1574–1584.
Vignais, P.M., Billoud, B., 2007. Occurrence, classification, and biological function ofhydrogenases: an overview. Chem. Rev. 107, 4206–4272.
Vignais, P.M., Billoud, B., Meyer, J., 2001. Classification and phylogeny of hydroge-nases. FEMS Microbiol. Rev. 25, 455–501.
Wang, Z.X., Zhuge, J., Fang, H., Prior, B.A., 2001. Glycerol production by microbialfermentation: a review. Biotechnol. Adv. 19, 201–223.
Weaver, P.F., Lien, S., Seibert, M., 1980. Photobiological production of hydrogen. Sol.Energy 24, 3–45.
Winkler, M., Hemschemeier, A., Gotor, C., Melis, A., Happe, T., 2002. [Fe]-hydrogenases in green algae: photo-fermentation and hydrogen evolution undersulfur deprivation. Int. J. Hydrogen Energy 27, 1431–1439.
Wykoff, D.D., Davies, J.P., Melis, A., Grossman, A.R., 1998. The regulation of pho-tosynthetic electron-transport during nutrient deprivation in Chlamydomonasreinhardtii. Plant Physiol. 117, 129–139.
Copyright © 2022 FDOKUMEN




















