
yamaha dt100a, b, c - dt125a, b, c - dt175a, b, c (1974-1976)
Upload
independentCategory
view
6download
0
Physiological and Molecular Plant Pathology (1996) 49, 121–141
Phenolic biosynthesis inhibitors suppress adult plant
resistance to Erysiphe graminis in oat at 20 °C and
10 °C
T. L. W. C*", L. Z", R. J. Z# and M. P. R"
" Institute of Grassland and En�ironmental Research, Aberystwyth, Dyfed SY23 3EB, U.K.and #Department of Plant Pathology, Uni�ersity of Minnesota, St Paul MN 55108, U.S.A.
(Accepted for publication May 1996)
Seedling and adult plant leaves of three oat genotypes, Selma, Maldwyn and OM1387 (a hybridderived from a Selma¬Maldwyn cross), were excised and treated with water or with α-amino-β-phenylpropionic acid (AOPP) to inhibit phenylalanine ammonia lyase activity, or with [[(2-hydroxyphenly)amino] sulfinyl]acid 1,1-dimethylethyl ester (OH-PAS) to inhibit cinnamylalcohol dehydrogenase activity. None of these genotypes possess any known major gene resistanceto the isolate of Erysiphe graminis f.sp. a�enae with which they were inoculated, although Maldwynis known to possess quantitative adult plant resistance that has proved durable since 1947. Allgenotypes showed some adult plant resistance which limited the proportion of fungal appressoriathat formed haustoria in water-treated leaves. However, this resistance was expressed morestrongly in Maldwyn and OM1387 than in Selma. Treatment with AOPP or OH-PAS increasedsubstantially the proportion of fungal appressoria that formed haustoria in adult as well asseedling leaves, indicating that phenolic compounds, probably products of the lignin biosyntheticpathway, contributed to resistance. However, AOPP-treated adult leaves remained more resistantto infection than AOPP-treated seedling leaves, suggesting that additional unknown factor(s)contributed to adult plant resistance. In all genotypes and in seedling and adult leaves, failure ofappressoria to infect was correlated to the localized accumulation of autofluorescent componentsin host cell papillae and cell wall regions subtending appressoria. This localized autofluorescencewas suppressed by AOPP treatment, and to a lesser extent by OH-PAS treatment, indicating thatthe fluorogens were phenolic compounds. In Maldwyn and OM1387 a proportion of host cellsshowed whole-cell autofluorescence, probably indicative of cell death, following attack byappressoria. Whole-cell autofluorescence was almost totally suppressed by AOPP or OH-PAStreatment, suggesting that this response was also dependent upon active synthesis of phenoliccompounds. The contribution of phenolic compound synthesis to disease resistance appeared tobe independent of temperature; AOPP treatment had similar effects in suppressing resistance inplants acclimated and incubated at 10 °C and at 20 °C. # 1996 Academic Press Limited
INTRODUCTION
It is well recognized that adult plant resistance to cereal powdery mildew and rusts is
commonly expressed in certain cultivars under field conditions. With powdery mildew,
caused by Erysiphe graminis DC (Blumeria graminis (DC) Speer), adult plant resistance
*To whom correspondence should be addressed.Abbreviations used in text : AOPP, α-amino-β-phenylpropionic acid; CAD, cinnamyl alcohol dehydro-
genase ; OH-PAS, [[(2-hydroxyphenyl) amino] sulfinyl]acid, 1,1-dimethylethyl ester ; PAL, phenyl-alanine ammonia lyase ; PGT, primary germ tube; p.f.d., photon flux density.
0885–5765}96}08012121 $25.00}0 # 1996 Academic Press Limited

122 T. L. W. Carver et al.
is characterized by delay in latent period of infection, and restriction of fungal colony
development and sporulation compared to susceptible cultivars. Unlike single gene
controlled resistance to powdery mildew, which is generally race-specific and ephemeral
in effect, adult plant resistance is often (but not exclusively) under the control of
multiple Mendelian factors and is referred to as polygene resistance. It is generally race
non-specific and durable. The physiological bases of adult plant resistance are poorly
understood.
The adult plant resistance of Maldwyn oat to E. graminis f.sp. a�enae Marchal has
been the subject of interest since this cultivar’s release in 1947 [21, 24, 31]. Maldwyn’s
resistance has remained field-effective [30] despite changes in the virulence of E.
graminis f.sp. a�enae which overcame many single gene resistances deployed in oat since
1947. At least six additive Mendelian factors contribute to Maldwyn’s resistance [22].
In Maldwyn and in a derivative hybrid, OM1387, sporulating colonies form on adult
leaves but the percentage of leaf area affected is lower than on seedling leaves or on the
adult leaves of more susceptible oat genotypes [21, 23, 24, 31]. The present studies
considered the possibility that phenolic compounds synthesized in response to attack by
E. graminis contribute to the adult plant resistance of Maldwyn oat.
Even in nominally susceptible seedling leaves of oat, barley or wheat attacked by
virulent isolates of Erysiphe graminis DC, only a proportion of fungal germlings
successfully penetrate host epidermal cells to form haustoria and establish infection.
The failure of some germlings to penetrate from appressoria to penetrate is in part due
to the intrinsic weakness of germlings (due, for example, to exhaustion of reserves).
However, other attacks fail because even nominally susceptible host cells express a
certain degree of resistance to infection. This can be recognized when experimental
treatments, e.g. the use of metabolic inhibitors, are shown to increase leaf susceptibility.
This phenomenon can be detected by histological analyses if sufficiently large numbers
of host parasite contact sites are characterized [8, 12–17, 35, 36].
We have shown previously that the proportion of appressoria successfully penetrating
nominally susceptible leaves of oat, barley and wheat inoculated with virulent,
appropriate formae speciales of E. graminis increased greatly when leaves were treated
with inhibitors of enzymes involved in phenolic compound biosynthesis [12, 14–17].
However, these studies were restricted to seedling leaves in all cases. In the present
study we examine the effects of inhibiting phenolic compound biosynthesis on the adult
plant resistance in Maldwyn and in a derivative hybrid genotype OM1387, and in the
susceptible oat cultivar Selma.
Our earlier work examined the effects on E. graminis}host interaction of α-amino-
β-phenylpropionic acid (AOPP), a potent and specific competitive inhibitor of
phenylalanine ammonia lyase (PAL) [3], effective against oat and barley PAL in �itro
[12, 35]. However, PAL catalyses the first committed step in phenylpropanoid
biosynthesis and so affects the synthesis of many different phenolic compounds. Thus,
the effects of PAL inhibition can be interpreted only broadly. Later, we also used
[[(2-hydroxyphenyl) amino] sulfinyl]acid, 1,1-dimethylethyl ester (OH-PAS), a known
inhibitor of cinnamyl alcohol dehydrogenase (CAD) [1, 19]. CAD is directly involved
with the synthesis of lignin precursors, and we showed that OH-PAS inhibited CAD
from oat and barley in �itro [12, 35]. When applied to nominally susceptible seedling
leaves of oat and barley, the effects of OH-PAS and AOPP were indistinguishable in

Phenolic biosynthesis inhibitors 123
increasing susceptibility to attempted penetration by E. graminis appressoria. This
suggested that products synthesized as part of the lignin biosynthetic pathway,
synthesized de no�o in response to pathogen attack, contribute to resistance to attempted
penetration from appressoria.
Mayama and Shisiyama [27] first reported the accumulation of autofluorogenic
compounds in host cell papillae and surrounding cell wall regions. From spectral
characteristics, they suggested that the autofluorogens were probably phenolic
compounds involved in resistance. In our previous experiments, increased susceptibility
to attempted penetration from appressoria following treatment with either AOPP or
OH-PAS, was associated with decreased frequency and intensity of localized
autofluorescent responses in host epidermal cells at sites of contact with fungal primary
germ tubes (PGTs) and appressoria. This provided supporting evidence that the
autofluorogens are indeed phenolic compounds with a role in resistance to penetration.
We also found that AOPP and OH-PAS suppressed host cell death associated with
hypersensitivity in barley possessing the Mla1 allele for resistance [35]. When Mla1
barley is challenged by an avirulent pathogen isolate, hypersensitive cell death is
characterized by bright whole-cell autofluorescence [26, 35]. It is also known [15, 26]
that even in genetically compatible interactions a small proportion of host epidermal
cells die in response to attack by E. graminis. In Maldwyn (used here) an early study,
that preceded our investigations of the involvement of plant phenolics in resistance,
reported that after staining around 10–20% of attacked cells may show signs of
cytoplasmic disorganization indicative of death when attacked by an oat mildew isolate
capable of forming a fully compatible infection [11]. However, in later studies of oat
which used AOPP and OH-PAS [12, 14, 15, 17], we did not examine their effects on
whole-cell autofluorescence, because with the equipment we used the autofluorescence
of dead oat cells was relatively faint and difficult to observe with certainty. In the
present study, we used an improved incident fluorescence microscope which allowed us
to study whole-cell autofluorescence responses (epidermal cell death) in oat.
Finally, in all previous studies examining the effects of AOPP and OH-PAS on E.
graminis development and host responses, we used a single temperature of 20 °C, the
optimal for powdery mildew development but in the current study we compared events
occurring in host leaves acclimated and incubated at 20 °C and at 10 °C. This allowed
us to judge the efficacy of resistance involving phenolic compound synthesis at different
temperatures likely to be encountered under field conditions.
MATERIALS AND METHODS
Host and pathogen material
Spring oat (A�ena sati�a L.) cvs Maldwyn and Selma and a transgressive segregant
(OM1387) derived from a Selma¬Maldwyn cross were used. None has any known
major gene resistance to race 5 of Erysiphe graminis DC f.sp. a�enae Marchal (Blumeria
graminis (DC) Speer f.sp. a�enae). Maldwyn has quantitative adult plant resistance [23]
under the control of at least six additive genetic factors [22]. This resistance has
remained effective in the field since 1947 [22]. In the field, OM1387 displays a higher
level of resistance to powdery mildew than either parent [30]. Selma has little field
resistance to E. graminis.

124 T. L. W. Carver et al.
The E. graminis isolate was maintained on young Selma plants. One day before
inoculation, heavily sporulating leaves were shaken to displace older conidia, ensuring
that mainly young conidia were used for inoculation the following day.
Application of AOPP and OH-PAS to plant lea�es, and inoculation and incubation procedures
All experiments involved leaf uptake of enzyme inhibitors prior to and during E.
graminis attack. The phenylalanine ammonia lyase (PAL) inhibitor α-amino-β-
phenylpropionic acid (AOPP) [12] was purchased from Cambridge Research
Biochemicals Ltd. (Northwich, Cheshire, U.K.). The cinnamyl alcohol dehydrogenase
(CAD) inhibitor [[(2-hydroxyphenyl) amino] sulfinyl]acid, 1,1-dimethylethyl ester
(OH-PAS), was synthesized by D. A. Boyles [17]. Aqueous solutions of both inhibitors
were used at 1±0 m concentration [12, 17]. They were made by dissolution in distilled
water using continuous stirring for 60 min at room temperature. Distilled water was
the control treatment.
For application to first-formed leaves, the leaves were cut at soil level and their cut
ends trimmed beneath water to remove vascular tissue containing air embolisms to
prevent wilting [12, 14–17, 35]. The cut ends were then immersed in the required test
solution which was taken up in the transpiration stream. Where later-formed leaves
were needed, a 100 mm segment was cut from the centre of the leaf blade and the basal
end trimmed to remove vascular tissue containing air embolisms prior to immersing in
the required test solution. Leaves were allowed to take up test solutions for 24 h before
inoculation, at either 20 °C or 10 °C as appropriate (see experimental designs).
For inoculation, leaves were removed from test solutions, laid adaxial surface up in
a spore settling tower, and conidia from infected leaves were blown directly into the
tower [12, 14–17, 35]. A microscope slide placed among the leaves was used to monitor
inoculum density which was adjusted to 10–20 conidia mm−#. Inoculation took no
longer than 10 min, after which leaves were again trimmed to remove air embolisms,
replaced in the appropriate test solution, and returned to the appropriate controlled
environment for incubation.
Fixation, clearing and light and incident fluorescence microscopy
For microscopy, 30 mm leaf segments were cut from the centre of treated leaves, fixed
in acetic acid-ethanol and cleared in lactoglycerol by a previously described process
that avoids displacement of ungerminated conidia or loosely attached germlings [12].
For microscopy, cleared leaf segments were placed on a microscope slide without a
coverslip and observed by transmitted light microscopy using a ‘‘no coverslip ’’
40¬objective lens [12]. Germling structures including primary germ tubes (PGTs),
appressoria, haustoria and hyphae were clearly visible.
Our previous studies of autofluorescent responses of oat epidermal cells to attack by
E. graminis f.sp. a�enae [12, 14, 15, 17] used an Olympus BH-A microscope and blue light
excitation (Olympus Optical Co. Ltd., Tokyo, Japan). For the current studies we also
used an Olympus BH-2 microscope with an Olympus RFC reflected light fluorescence
attachment fitted with a blue-violet excitation filter (max. transmittance 440 nm) and
barrier filter with transmittance above 475 nm. The BH-2 system gave bright

Phenolic biosynthesis inhibitors 125
fluorescent images for visualization of blue-white autofluorescence associated with
localized responses of host epidermal cells and whole-cell autofluorescence indicative of
cell death [26].
In most of our previous studies of oat [12, 14, 17] we had not considered whole-cell
autofluorescence in oat because it is often faint and very difficult to see. However, with
the BH-2 system used here, we could discern whole-cell autofluorescence with relative
ease. In barley, Koga et al. [26] showed that epidermal cells with whole-cell
autofluorescence also failed to take up the vital dye neutral red, and to respond to
plasmolysing treatment. They concluded that such cells were dead. In preliminary tests
we followed the procedures of Koga et al. [26] using unfixed leaves of oat cv. Maldwyn
inoculated 36 h prior to testing. Our results were similar to those from the barley
system [26]. We found that unattacked epidermal cells, or cells with localized
autofluorescence associated with appressorial contact, almost invariably took up
neutral red and responded to plasmolysing treatment, indicating that they were alive.
By contrast, cells showing whole-cell autofluorescence almost always failed to take up
the vital dye neutral red and to respond to plasmolysing treatment, indicating that
these cells were dead. There were a few cells that showed faint whole-cell
autofluorescence but the protoplast also showed faint neutral red staining and
appeared to be slightly plasmolysed with an irregular outline. We, like Koga et al. [26],
concluded that such cells were alive but dying. We therefore consider that whole-cell
autofluorescence is a useful indication of cell death in oat as well as in barley epidermal
cells.
Data collection and statistical analyses
In previous studies, treatments with AOPP and OH-PAS did not affect spore
germination [12, 14–17, 35]. To confirm that this was true in adult as well as seedling
leaves, 100 conidia were examined on each leaf segment in Experiment 1, to determine
whether they had germinated i.e. formed at least a primary germ tube.
In all experiments, we determined penetration success (haustorium formation) from
fungal appressoria by examining 100 germlings with mature appressoria on each
leaf. Only those germlings were assessed if their PGT and appressorium were on host
cells that had no contact with other germlings. In addition, PGT and appressorium
contact sites on epidermal cells were examined by incident fluorescence microscopy for
localized autofluorescent responses, and host cells subtending appressoria were
examined for whole-cell autofluorescence.
Mean replicate data for each category of observation were calculated as a
percentage. Percentages were transformed to arcsine square roots (transformed value
¯ 180}π¬arcsine [o(percent}100)]) to normalize data and stabilize the variance
throughout the data range prior to standard analysis of variance using appropriately
written Genstat programs [12, 14–17, 35].
For appressorium contact sites, the intensity of localized autofluorescent host cell
responses was ranked visually on a 0–3 scale where: 0¯no visible fluorescence; 1¯faint fluorescence; 2¯moderate fluorescence; and 3¯ intense fluorescence [12].
Fluorescence intensity data were not analyzed statistically because distributions were
often heavily skewed and data often had a high proportion of zero values.

126 T. L. W. Carver et al.
The influence of AOPP on adult plant resistance in successi�ely-formed lea�es : Experiments 1
and 2
Experiments 1 and 2 were identical in design and were conducted in successive years.
We wished to produce plants reaching different growth stages, seedling to adult, at the
same time. This would allow comparison of fungal development and host responses in
leaves from different positions formed under the same environment. To produce such
plants, seeds were sown in succession over a 45 day period and plants were grown in
a spore-proof glasshouse under natural lighting conditionswith aminimum temperature
of 12 °C. For both experiments, pre-germinated seedlings of the susceptible genotype
Selma and the adult plant resistant genotype Maldwyn were planted separately, three
plants per 125 mm pot, in John Innes No. 3 potting compost. A pot of each oat
genotype was grown in each of three replicates of a randomized block. The first sowing
in each experiment was made in mid June, and freshly sown pots were incorporated
into the randomized block each week until the last week in July. Thus, successive
sowings produced plants at different growth stages with leaves at different ascending
positions on primary shoots reaching the same physiological age at the same time and
under the same environmental conditions.
In early August, the eighth-formed leaves on primary shoots were newly expanded
on plants from the earliest sowing, and seventh-formed leaves from these plants
provided segments (100 mm) for experimentation. At the same time, fifth-formed leaf
segments were cut from plants with newly expanded sixth leaves, third-formed leaf
segments were cut from plants with newly expanded fourth leaves, and first-formed
(seedling) leaves were cut from plants of the latest sowing where the second-formed leaf
was newly expanded.
In all cases, a leaf segment was taken from each of two phenotypically similar plants
grown in the same pot ; one leaf segment from each pot was used for AOPP treatment,
and the other as the water control. Thus, for each genotype and each leaf position,
three leaves (one from each replicate) were used for AOPP treatment, and three for
water controls. In total, therefore, there were six segments from leaves of the same
position (three replicate segments from each of the two oat genotypes) used for AOPP
treatment, and six for water control treatment. The cut ends of one set of six leaves from
each position were immersed in a vessel containing 20 ml of 1±0 m AOPP solution,
and their counterparts in a vessel containing 20 ml of distilled water.
All vessels were transferred to a growth cabinet at 20³0±5 °C, with light supplied by
warm white fluorescent tubes for 12 h day−" (9.00–21.00 h) at 150 µmol m−# s−" photon
flux density (p.f.d.) at leaf level. The first light period began immediately. After 24 h,
leaves were inoculated, returned to the appropriate solution and incubated in the
growth cabinet for a further 36 h before fixation.
Comparati�e effects of AOPP and OH-PAS on the resistance of fifth-formed lea�es : Experiment
3
In mid-April, pre-germinated seedlings of the susceptible genotype Selma and the
adult plant resistant genotype Maldwyn were planted separately, three plants per
125 mm pot, in John Innes No. 3 potting compost. Three pots of each genotype were

Phenolic biosynthesis inhibitors 127
included in each of three replicates of a randomized block. Plants were grown in a
spore-proof glasshouse as for Experiments 1 and 2. By late May plants had formed
newly expanded sixth-formed leaves on the primary shoot. To ensure that leaves
destined for different experimental treatments were comparable, one plant was selected
from every pot on the basis of its phenotypic similarity to equivalent plants. A 100 mm
segment was cut from the fifth-formed leaf of each selected plant. The basal cut ends
of sets of three leaf segments of each genotype (one from each of the three replicates)
were immersed in 20 ml 1±0 m AOPP, 20 ml 1±0 m OH-PAS, or 20 ml distilled
water. Pre- and post-inoculation conditions were the same as for Experiments 1 and 2.
The influence of AOPP on first- and fifth-formed lea�es of plants acclimated at 10 °C or 20 °C:
Experiments 4 and 5
Experiment 4: first-formed lea�es. Two sets of plants with first-formed leaves of similar
physiological age were grown under the same lighting environment but acclimated to
10 °C or 20 °C temperature conditions. For the 10 °C requirement, six pre-germinated
seedlings of Selma, Maldwyn and OM1387 oat were planted in separate 125 mm pots
of John Innes No. 3 compost, and grown in a spore-proof glasshouse. When the first-
formed leaves of these plants were approaching full expansion, they were transferred
to a growth room at 10³1±5 °C, with light supplied by warm white fluorescent tubes
for 12 h day−" (9.00–21.00 h) at 150 µmol m−# s−" p.f.d. at leaf level. At this time, a
second set of seedlings was planted in a growth room with the same lighting conditions
but at 20³1±5 °C. After 2 weeks plants of both sets had produced fully expanded
second-formed leaves and first leaves were taken for experimentation.
Six first-formed leaves of each oat genotype from each growth room were cut at
9.00 h (the beginning of the 12 h light period) and three were treated with 1±0 m AOPP
while three were treated with water. They were returned for 24 h to their original
growth room to allow chemical uptake, inoculated and then returned to the
appropriate growth room for incubation. Leaves held at 20³1±5 °C, were fixed after
36 h and leaves held at 10³1±5 °C were incubated for 72 h. Preliminary experiments
(Zhang, unpublished) showed that pathogen and germling development was greatly
delayed at 10³1±5 °C necessitating the longer 72 h incubation. It was impractical to
incubate leaves for longer because leaf tips began showing chlorosis at 72 h.
Experiment 5: fifth-formed leaves. Two sets of fifth-formed leaves of similar physiological
age from plants grown under the same lighting environment but acclimated to 10 °Cor 20 °C temperature conditions were required. Therefore, successive sowings of Selma,
Maldwyn and OM1387 were made to produce six plants of each genotype in separate
125 mm pots of John Innes No. 3 compost. All plants were grown in a spore-proof
glasshouse. For the 10 °C requirement, plants from the first sowing were taken when
their fifth-formed leaves were unrolling, and moved to the 10 °C growth room, with
light supplied by warm white fluorescent tubes for 12 h day−" (9.00–21.00 h) at
150 µmol m−# s−" p.f.d. at leaf level. At the same time, plants from the later sowing had
developed unrolling fourth-formed leaves. These plants were moved to a similar
growth room at 20 °C. After 2 weeks, plants of both sets had fully expanded sixth-
formed leaves and their fifth-formed leaves were taken for experimentation. Fifth-

128 T. L. W. Carver et al.
formed leaves were treated with AOPP or water in the same way as first-formed leaves
in Experiment 4.
RESULTS
Conidial germination
Germination rates in Experiment 1 were unaffected by inhibitor treatment, leaf
position or by host genotype. This confirmed previous findings [12, 14–17, 33], and
germination was not scored in subsequent experiments.
The influence of AOPP on adult plant resistance in successi�ely-formed lea�es : Experiments 1
and 2
These experiments compared success of attempted penetration and associated host cell
responses in leaves from ascending positions on the primary shoot of Selma, which has
little adult plant resistance, and of Maldwyn, which has marked adult plant resistance.
To examine the role of plant phenolics in these genotypes, we compared water-treated
with AOPP-treated leaves.
Results from Experiments 1 and 2 were very similar and so only those of Experiment 1
are described in detail. Experiment 2 results are described only when they varied from
Experiment 1.
Percentage penetration (haustorium formation) from appressoria. Overall, a higher percentage of
germlings with appressoria penetrated to form haustoria in the susceptible genotype
Selma than in the adult plant resistant genotype Maldwyn (mean percentages¯ 62±9and 29±2 respectively, P! 0±001, for data pooled over leaf positions and treatments).
Fewer appressoria penetrated (produced haustoria) in the upper leaves of both
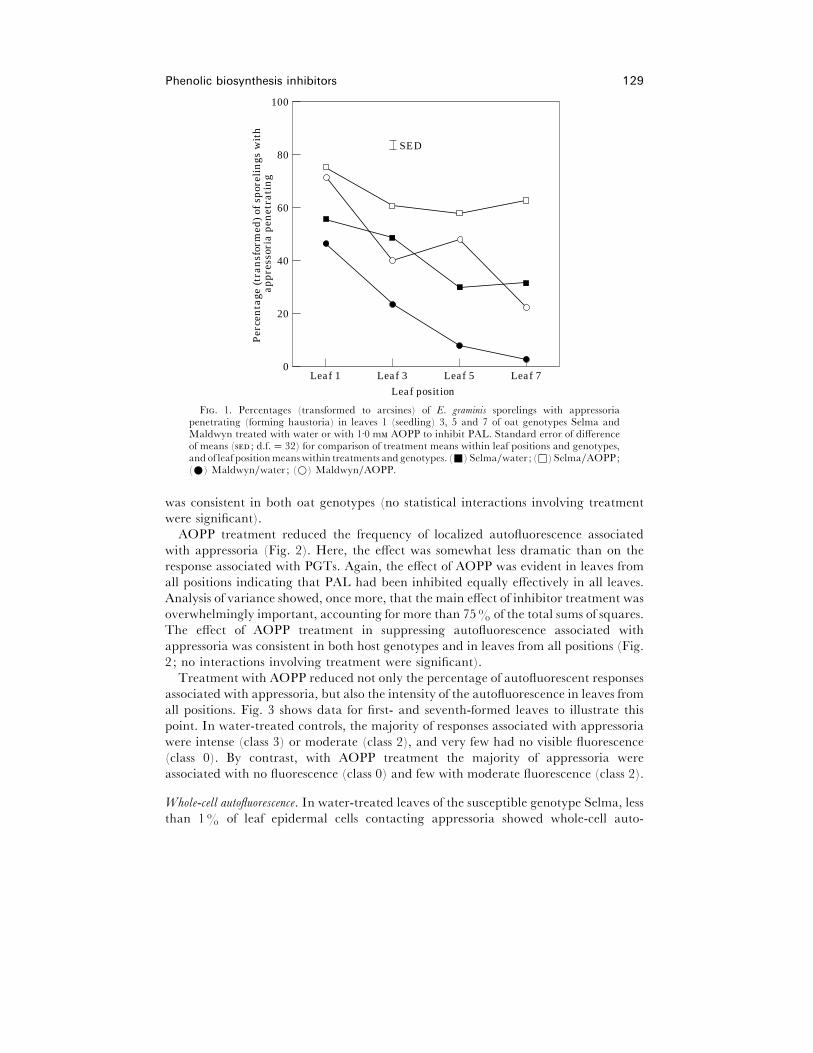
genotypes (the main effect of leaf position was significant; P! 0±001). Fig. 1 shows that
this effect was greater in the upper leaves of the adult plant resistant genotype
Maldwyn than in the more susceptible genotype Selma. This led to a significant (P!0±001) host genotype¬leaf position interaction.
Overall, a higher percentage of appressoria formed haustoria in AOPP- than in
water-treated leaves (mean percentages¯ 66±4 and 26±0 respectively, P! 0±001, for
data pooled over oat genotypes and leaf positions). The effect of AOPP was evident in
both oat genotypes (the genotype¬treatment interaction was not significant).
Furthermore, the effect of AOPP on penetration success was evident in leaves from all
positions (the leaf position¬treatment interaction was not significant). It is clear from
Fig. 1 that even with AOPP treatment, upper leaves remained more resistant to
penetration than first-formed leaves. This suggests that PAL activity was not the only
factor contributing to the penetration resistance of upper leaves.
Localized autofluorescent responses of epidermal cells in contact with fungal PGTs and appressoria.
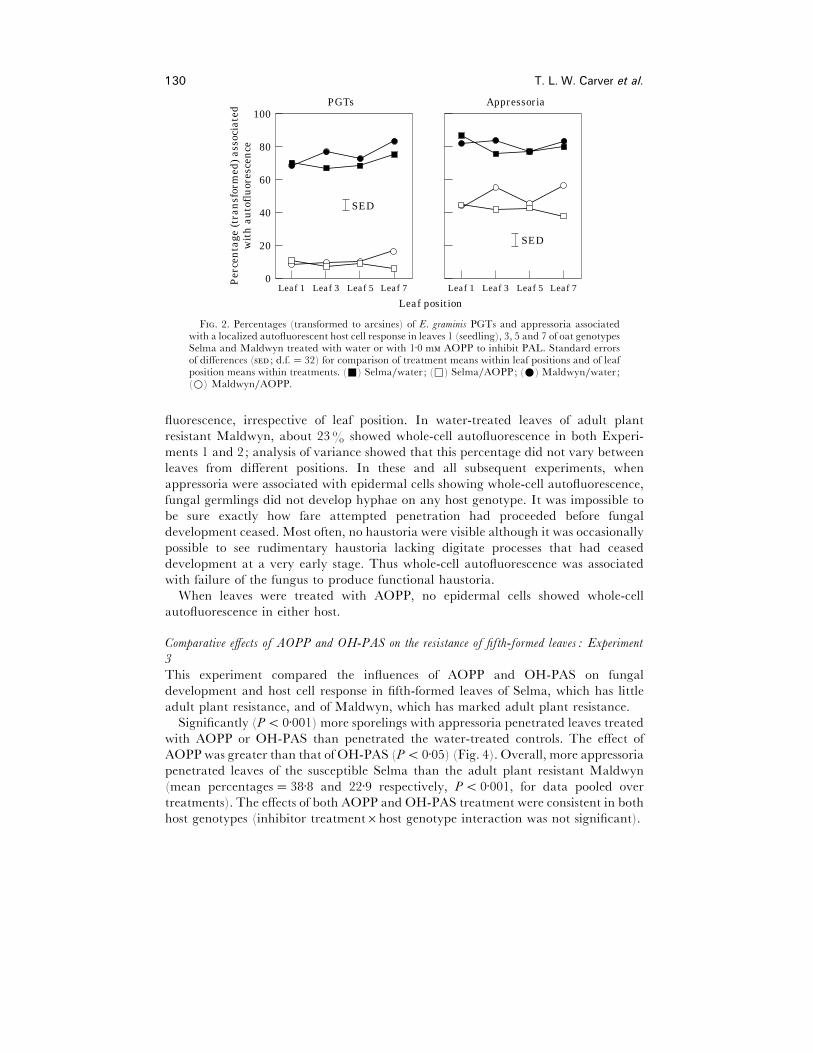
AOPP treatment greatly reduced the percentage of localized autofluorescent responses
to PGTs (Fig. 2). In analyses of variance, the main effect of AOPP treatment was
overwhelmingly important, accounting for more than 90% of the total sums of squares.
Very low percentages of autofluorescent responses to PGTs were noted in AOPP-
treated leaves from all positions. Thus the PAL inhibitor had suppressed PGT-
associated autofluorescence equally effectively in seedling and upper leaves, and this
Phenolic biosynthesis inhibitors 129
Leaf 7
100
0Leaf 1
Leaf position
Per
cen
tage
(tr
ansf
orm
ed)
of s
pore
lin
gs w
ith
app
ress
oria
pen
etra
tin
g
Leaf 3
80
60
40
20
SED
Leaf 5
F. 1. Percentages (transformed to arcsines) of E. graminis sporelings with appressoriapenetrating (forming haustoria) in leaves 1 (seedling) 3, 5 and 7 of oat genotypes Selma andMaldwyn treated with water or with 1±0 m AOPP to inhibit PAL. Standard error of differenceof means ( ; d.f.¯ 32) for comparison of treatment means within leaf positions and genotypes,and of leaf positionmeanswithin treatments and genotypes. (+) Selma}water ; (*) Selma}AOPP;(E) Maldwyn}water ; (D) Maldwyn}AOPP.
was consistent in both oat genotypes (no statistical interactions involving treatment
were significant).
AOPP treatment reduced the frequency of localized autofluorescence associated
with appressoria (Fig. 2). Here, the effect was somewhat less dramatic than on the
response associated with PGTs. Again, the effect of AOPP was evident in leaves from
all positions indicating that PAL had been inhibited equally effectively in all leaves.
Analysis of variance showed, once more, that the main effect of inhibitor treatment was
overwhelmingly important, accounting for more than 75% of the total sums of squares.
The effect of AOPP treatment in suppressing autofluorescence associated with
appressoria was consistent in both host genotypes and in leaves from all positions (Fig.
2 ; no interactions involving treatment were significant).
Treatment with AOPP reduced not only the percentage of autofluorescent responses
associated with appressoria, but also the intensity of the autofluorescence in leaves from
all positions. Fig. 3 shows data for first- and seventh-formed leaves to illustrate this
point. In water-treated controls, the majority of responses associated with appressoria
were intense (class 3) or moderate (class 2), and very few had no visible fluorescence
(class 0). By contrast, with AOPP treatment the majority of appressoria were
associated with no fluorescence (class 0) and few with moderate fluorescence (class 2).
Whole-cell autofluorescence. In water-treated leaves of the susceptible genotype Selma, less
than 1% of leaf epidermal cells contacting appressoria showed whole-cell auto-
130 T. L. W. Carver et al.
Leaf 7
100
0Leaf 1
Leaf position
Per
cen
tage
(tr
ansf
orm
ed)
asso
ciat
edw
ith
au
tofl
uor
esce
nce
Leaf 3
80
60
40
20
SED
Leaf 5 Leaf 7Leaf 1 Leaf 3
SED
Leaf 5
PGTs Appressoria
F. 2. Percentages (transformed to arcsines) of E. graminis PGTs and appressoria associatedwith a localized autofluorescent host cell response in leaves 1 (seedling), 3, 5 and 7 of oat genotypesSelma and Maldwyn treated with water or with 1±0 m AOPP to inhibit PAL. Standard errorsof differences ( ; d.f.¯ 32) for comparison of treatment means within leaf positions and of leafposition means within treatments. (+) Selma}water ; (*) Selma}AOPP; (E) Maldwyn}water ;(D) Maldwyn}AOPP.
fluorescence, irrespective of leaf position. In water-treated leaves of adult plant
resistant Maldwyn, about 23% showed whole-cell autofluorescence in both Experi-
ments 1 and 2; analysis of variance showed that this percentage did not vary between
leaves from different positions. In these and all subsequent experiments, when
appressoria were associated with epidermal cells showing whole-cell autofluorescence,
fungal germlings did not develop hyphae on any host genotype. It was impossible to
be sure exactly how fare attempted penetration had proceeded before fungal
development ceased. Most often, no haustoria were visible although it was occasionally
possible to see rudimentary haustoria lacking digitate processes that had ceased
development at a very early stage. Thus whole-cell autofluorescence was associated
with failure of the fungus to produce functional haustoria.
When leaves were treated with AOPP, no epidermal cells showed whole-cell
autofluorescence in either host.
Comparati�e effects of AOPP and OH-PAS on the resistance of fifth-formed lea�es : Experiment
3
This experiment compared the influences of AOPP and OH-PAS on fungal
development and host cell response in fifth-formed leaves of Selma, which has little
adult plant resistance, and of Maldwyn, which has marked adult plant resistance.
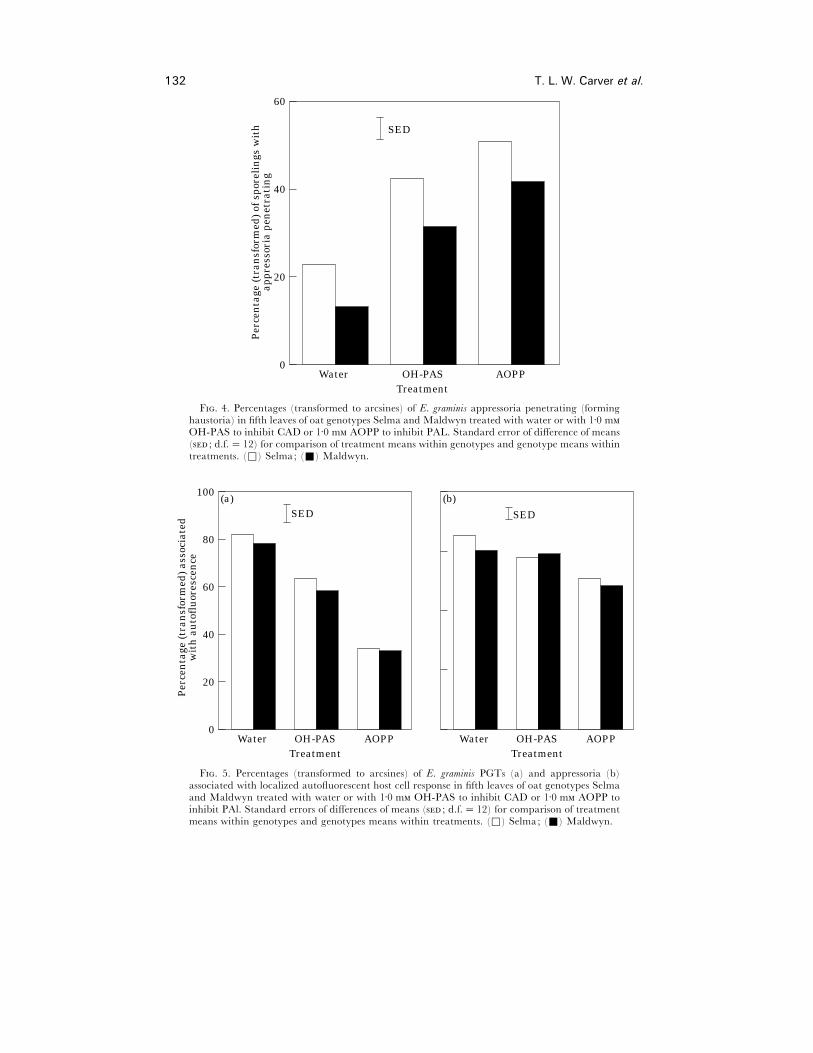
Significantly (P! 0±001) more sporelings with appressoria penetrated leaves treated
with AOPP or OH-PAS than penetrated the water-treated controls. The effect of
AOPP was greater than that of OH-PAS (P! 0±05) (Fig. 4). Overall, more appressoria
penetrated leaves of the susceptible Selma than the adult plant resistant Maldwyn
(mean percentages¯ 38±8 and 22±9 respectively, P! 0±001, for data pooled over
treatments). The effects of both AOPP and OH-PAS treatment were consistent in both
host genotypes (inhibitor treatment¬host genotype interaction was not significant).

Phenolic biosynthesis inhibitors 131
250
0
Leaf 1
Nu
mbe
rs o
f ap
pres
sori
a
200
150
100
50
Genotype: MaldwynWater (Control) AOPP (1 mM)
Inhibitor treatment
250
0
200
150
100
50
250
0
Leaf 1
Nu
mbe
rs o
f ap
pres
sori
a
200
150
100
50
Genotype: Selma
250
0
200
150
100
50
0 1 2 3 0 1 2 3Fluorescence intensity score
Leaf 7
Leaf 7
F. 3. Intensities of localized autofluorescent responses in host cells associated with E. graminisappressoria in first- and seventh-formed leaves of two oat genotypes treated with water (controls)or with the PAL inhibitor AOPP (1±0 m aqueous solution). O¯no fluorescence; 1, 2, 3¯ faint,moderate and intense fluorescence respectively. Data based on observation of 300 appressoria ineach case.
Both inhibitors significantly reduced (P! 0±001) the percentage of localized
autofluorescent host cell responses associated with PGTs (Fig. 5) and, again, the effect
of AOPP was greater than that of OH-PAS (P! 0±001). AOPP reduced significantly
(P! 0±001) the percentage of localized autofluorescent host cell responses associated
with appressoria, but OH-PAS had little or no effect. The effects of inhibitor
treatments were consistent in both host genotypes (inhibitor treatment¬host genotype
interactions were not significant).
132 T. L. W. Carver et al.
Per
cen
tage
(tr
ansf
orm
ed)
of s
pore
lin
gs w
ith
appr
esso
ria
pen
etra
tin
g
60
0
40
20
Water OH-PAS AOPPTreatment
SED
F. 4. Percentages (transformed to arcsines) of E. graminis appressoria penetrating (forminghaustoria) in fifth leaves of oat genotypes Selma and Maldwyn treated with water or with 1±0 mOH-PAS to inhibit CAD or 1±0 m AOPP to inhibit PAL. Standard error of difference of means( ; d.f.¯ 12) for comparison of treatment means within genotypes and genotype means withintreatments. (*) Selma; (+) Maldwyn.
Per
cen
tage
(tr
ansf
orm
ed)
asso
ciat
edw
ith
au
tofl
uor
esce
nce
100
0
40
20
Water OH-PAS AOPPTreatment
SED
60
80
Water OH-PAS AOPPTreatment
SED(a) (b)
F. 5. Percentages (transformed to arcsines) of E. graminis PGTs (a) and appressoria (b)associated with localized autofluorescent host cell response in fifth leaves of oat genotypes Selmaand Maldwyn treated with water or with 1±0 m OH-PAS to inhibit CAD or 1±0 m AOPP toinhibit PAl. Standard errors of differences of means ( ; d.f.¯ 12) for comparison of treatmentmeans within genotypes and genotypes means within treatments. (*) Selma; (+) Maldwyn.

Phenolic biosynthesis inhibitors 133
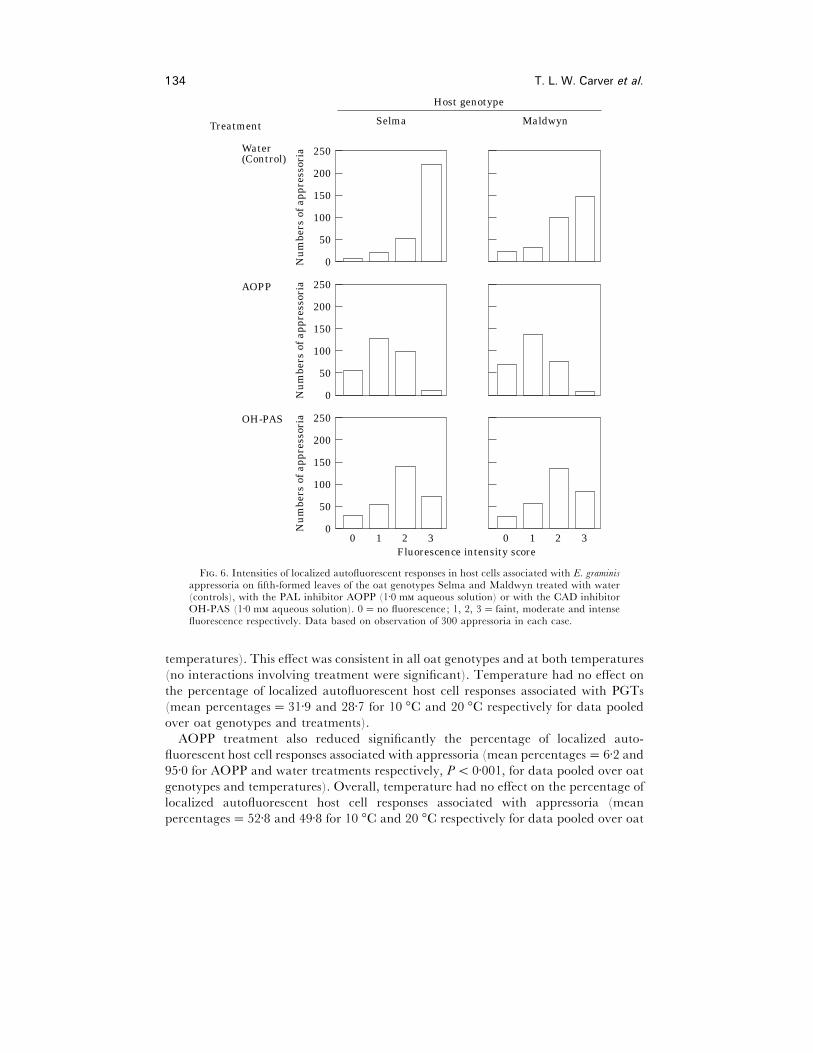
Treatment with either AOPP or OH-PAS decreased the intensity of localized
autofluorescent host cell responses associated with appressoria. Fig. 6 shows that in
both host genotypes treated with water the majority of appressoria were associated
with intense (class 3) or moderate (class 2) autofluorescent responses. In AOPP-treated
leaves, the majority were associated with faint (class 1) autofluorescence, and few with
intense autofluorescence. With OH-PAS treatment, autofluorescent responses were
generally less intense than in water treated leaves (moderate, class 2 autofluorescent
was most common) but the effect of OH-PAS was less than that of AOPP.
Whole-cell autofluorescence. In water-treated leaves, a mean of 5% and 13±7% respectively
of Selma and Maldwyn epidermal cells showed whole-cell autofluorescence when in
contact with appressoria. When Selma leaves were treated with either inhibitor, whole-
cell autofluorescence was totally prevented by both inhibitors. In Maldwyn, only 0±3%
and 1±7% of cells in leaves treated with AOPP or OH-PAS respectively showed whole-
cell autofluorescence as both inhibitors effectively suppressed this response in this adult
plant resistant genotype.
The influence of AOPP on first- and fifth-formed lea�es of plants acclimated at
10 °C or 20 °C: Experiments 4 and 5
In the previous experiments we examined the effects of adult plant resistance and
inhibitors of phenolic compound biosynthesis at the optimal temperature for E. graminis
development (20 °C). The current experiments were designed to test whether AOPP
treatment and leaf position had consistent effects at a lower temperature, as frequently
encountered in the field. The susceptible genotype Selma, the adult plant resistant
Maldwyn, and the Selma¬Maldwyn hybrid, OM1387, were used for these
experiments.
Experiment 4: first-formed leaves
Haustorium formation by appressoria. Overall, a higher percentage of appressoria penetrated
to form haustoria in AOPP- than in water-treated leaves (mean percentages¯ 69±3and 42±5 respectively,P! 0±001, for data pooled over host genotypes and temperatures).
This AOPP effect was consistent at both temperatures and in all oat genotypes (no
interactions involving treatment were significant).
Fewer appressoria penetrated at 10 °C than 20 °C (mean percentages¯ 46±3 and
65±8 respectively, P! 0±001, for data pooled over oat genotypes and treatments). This
temperature effect was consistent in all host genotypes (the incubation temperature¬host genotype interaction was not significant). Overall, a significantly (P! 0±001)
higher percentage of appressoria formed haustoria in Selma than in either Maldwyn
or OM1387 (mean percentages¯ 73±5, 46±0 and 48±0 respectively, for data pooled over
temperatures and treatments).
Localized autofluorescent host cell responses associated with PGTs and appressoria. As expected,
AOPP treatment reduced significantly (P! 0±001) the percentage of localized
autofluorescence associated with PGTs (mean percentages¯ 0±6 and 78±4 for AOPP
and water treatments respectively for data pooled over oat genotypes and
134 T. L. W. Carver et al.
250
0
Water(Control)
Nu
mbe
rs o
f ap
pres
sori
a
200
150
100
50
Treatment Selma Maldwyn
Host genotype
0 1 2 3 0 1 2 3Fluorescence intensity score
AOPP 250
0
200
150
100
50
250
0
200
150
100
50
OH-PAS
Nu
mbe
rs o
f ap
pres
sori
aN
um
bers
of
appr
esso
ria
F. 6. Intensities of localized autofluorescent responses in host cells associated with E. graminisappressoria on fifth-formed leaves of the oat genotypes Selma and Maldwyn treated with water(controls), with the PAL inhibitor AOPP (1±0 m aqueous solution) or with the CAD inhibitorOH-PAS (1±0 m aqueous solution). 0¯no fluorescence; 1, 2, 3¯ faint, moderate and intensefluorescence respectively. Data based on observation of 300 appressoria in each case.
temperatures). This effect was consistent in all oat genotypes and at both temperatures
(no interactions involving treatment were significant). Temperature had no effect on
the percentage of localized autofluorescent host cell responses associated with PGTs
(mean percentages¯ 31±9 and 28±7 for 10 °C and 20 °C respectively for data pooled
over oat genotypes and treatments).
AOPP treatment also reduced significantly the percentage of localized auto-
fluorescent host cell responses associated with appressoria (mean percentages¯ 6±2 and
95±0 for AOPP and water treatments respectively, P! 0±001, for data pooled over oat
genotypes and temperatures). Overall, temperature had no effect on the percentage of
localized autofluorescent host cell responses associated with appressoria (mean
percentages¯ 52±8 and 49±8 for 10 °C and 20 °C respectively for data pooled over oat

Phenolic biosynthesis inhibitors 135
genotypes and treatments). However, AOPP treatment had less effect in reducing
autofluorescence at 10 °C than at 20 °C. This led to a significant (P! 0±001)
treatment¬incubation temperature interaction. Nevertheless, autofluorescence inten-
sities were clearly reduced in AOPP-treated leaves at 10 °C (data not shown), so that
where localized autofluorescence was present, it was invariably faint (class 1)
compared to water-treated leaves that showed moderate or intense (class 2–3)
responses.
Whole-cell autofluorescence. In water-treated leaves of Selma from 20 °C treatment, only
1% of cells showed whole-cell autofluorescence and none did so in the 10 °C treated
plants. In both adult plant resistant genotypes, Maldwyn and OM1387, approximately
15% of cells contacted by appressoria showed whole-cell autofluorescence in water-
treated leaves from both temperatures. No whole-cell autofluorescence was found in
any host genotype following AOPP treatment at either temperature.
In summary, in first-formed leaves AOPP treatment was equally effective at 10 °Cand 20 °C in terms of its effects on penetration success (haustorium formation). Also in
terms of its effect on whole-cell autofluorescence and PGT-associated localized
autofluorescence, AOPP treatment was equally effective at both temperatures. AOPP
treatment was slightly more effective at 20 °C than at 10 °C in suppressing localized
epidermal cell autofluorescence associated with appressoria.
Experiment 5: Fifth-formed leaves
Haustorium formation by appressoria. Overall, a higher percentage of appressoria penetrated
to form haustoria in AOPP- than in water-treated leaves (mean percentages¯ 65±9and 24±8 respectively,P! 0±001, for data pooled over host genotypes and temperatures).
This effect was consistent at both 10 °C and 20 °C.
As in first-formed leaves, in fifth leaves fewer appressoria penetrated to form
haustoria at 10 °C than 20 °C (mean percentages¯ 29±8 and 60±5 respectively, P!0±001, for data pooled over host genotypes and treatments). Overall a significantly (P!0±001) higher percentage of appressoria formed haustoria in the susceptible genotype
Selma than in either of the adult plant resistant genotypes, Maldwyn or OM1387
(mean percentages¯ 68±3, 33±3 and 33±3 respectively, for data pooled over temperatures
and treatments). In all oat genotypes, more appressoria formed haustoria in AOPP-
than water-treated leaves. AOPP treatment had slightly less effect in increasing
penetration in fifth leaves of Selma than the other genotypes (leading to a significant
host genotype¬treatment interaction), but AOPP-treated Selma leaves remained
significantly (P! 0±001) more susceptible than AOPP-treated leaves of Maldwyn and
OM1387. Thus, even when PAL was inhibited, these adult leaves of Maldwyn and
OM1387 remained relatively more resistant to penetration than those of Selma.
Localized autofluorescent host cell responses associated with PGTs. Again, in fifth leaves, AOPP
treatment reduced significantly (P! 0±001) the percentage of localized auto-
fluorescence responses associated with PGTs (mean percentages¯ 10±1 and 84±2 for
AOPP and water treatments respectively for data pooled over oat genotypes and
temperatures). This effect was overwhelmingly important and accounted for
approximately 68% of the total sums of squares in the analysis of variance.

136 T. L. W. Carver et al.
Host genotype had a small but significant overall effect (P! 0±05) on the percentage
of autofluorescent host cell responses associated with PGTs (mean percentages¯ 40±3,
50±6 and 46±3 for Maldwyn, OM1387 and Selma respectively, for data pooled over
temperatures and treatments). However, AOPP treatment reduced significantly the
percentage of localized autofluorescence associated with PGTs in all host genotypes.
Temperature also had a small but significant overall effect and fewer PGTs were
associated with localized autofluorescence at 20 °C than 10 °C (mean percentages¯22±7 and 69±7 respectively, P! 0±001, for data pooled over host genotypes and
treatments). At 20 °C AOPP treatment almost totally suppressed autofluorescence
associated with PGTs (mean percentages¯ 0±1 and 67±3 for AOPP and water
treatments respectively for data pooled over host genotypes), while at 10 °C, AOPP
treatment had slightly less effect (mean percentages¯ 33±3 and 95±7 for AOPP and
water treatments respectively for data pooled over host genotypes). This caused a small
but significant treatment¬temperature interaction (accounting for ! 0±1% of total
sums of squares in analysis of variance).
Localized autofluorescent host cell responses associated with appressoria. AOPP treatment
reduced significantly (P! 0±001) the percentage of localized autofluorescent responses
associated with appressoria (mean percentages¯ 30±8 and 96±7 for AOPP and water
treatments respectively for data pooled over host genotype and temperature). This
effect was overwhelmingly important and accounted for approximately 63% of the
total sums of squares in the analysis of variance.
Host genotype had no effect on the percentage of localized autofluorescent host cell
response associated with appressoria (the main effect of host genotype was not signifi-
cant). AOPP was equally effective in reducing this percentage in all host genotypes.
Temperature had a small but significant overall effect and fewer appressoria were
associated with localized autofluorescence at 20 °C than 10 °C (mean percentages¯88±8 and 46±2 respectively, P! 0±001, for data pooled over host genotypes and
treatments). At 20 °C, AOPP treatment had more effect (mean percentages¯ 3±8 and
92±7 for AOPP and water treatments respectively for data pooled over host genotypes)
than at 10 °C (mean percentages¯ 69±1 and 99±1 for AOPP and water treatments
respectively for data pooled over host genotypes). This led to a significant
treatment¬incubation temperature interaction (accounting for approximately 9% of
total sums of squares in analysis of variance).
AOPP treatment also reduced the intensity of autofluorescence at appressorial
contact sites (data not shown). This was true in all genotypes at both temperatures. In
AOPP-treated leaves autofluorescent responses were almost always faint (class 1) and
there were few moderate (class 2) and no intense (class 3) responses at either incubation
temperature. This contrasted with water-treated leaves where the majority of responses
were moderate or intense (class 2 and 3).
Whole-cell autofluorescence. In water-treated leaves of the susceptible genotype Selma
there was no whole-cell autofluorescence at either incubation temperature. In the adult
plant resistant genotypes Maldwyn and OM1387 approximately 35% of cells in water-
treated leaves showed whole-cell autofluorescence at both temperatures. No whole-cell
autofluorescence was found in any host genotype following AOPP treatment.

Phenolic biosynthesis inhibitors 137
In summary, in fifth-formed leaves. AOPP treatment appeared to be equally
effective at 10 °C and 20 °C in terms of its effects on penetration success (haustorium
formation) and in suppressing whole-cell autofluorescence. In terms of its effects in
suppressing localized autofluorescent host cell responses associated with PGTs and
appressoria, AOPP treatment appeared slightly more effective at 20 °C than at 10 °C.
DISCUSSION
The adult plant resistance of Maldwyn oat has proved effective in the field for over four
decades [21, 24, 30, 31]. It has been argued that this durability is due to the fact that
at least six additive Mendelian factors contribute to this resistance [22]. Maldwyn’s
adult plant resistance is quantitative ; while sporulating colonies form on adult leaves,
the percentage of leaf area affected by disease is lower than on its seedling leaves or on
the adult leaves of more susceptible oat genotypes [23]. In part, this is because fewer
E. graminis germling appressoria penetrate successfully [9, 18]. The present findings
corroborate these earlier descriptive studies, showing progressively reduced success of
attempted penetration in successively formed leaves. The hybrid line OM1387 gave
reduced levels of penetration similar to those seen in the resistant progenitor Maldwyn
and significantly lower than in its other progenitor, the susceptible cultivar Selma (Fig.
1).
A main objective was to test the hypothesis that plant phenolic compounds are
involved in adult plant resistance. The finding that treatment with AOPP decreased
penetration resistance of adult leaves as measured by haustorium formation, confirms
phenolic involvement. This finding also indicates that, as in seedling leaves of oat and
barley [12, 14–17], the phenolic compounds are synthesized de no�o in response to E.
graminis attack.
Treatment with OH-PAS to inhibit CAD activity also significantly reduced the
penetration resistance of adult (fifth-formed) leaves. This indicates that cinnamyl
alcohols (the direct product of CAD activity), or later formed products of the lignin
biosynthetic pathway, are involved in adult plant resistance. However, in adult leaves
OH-PAS was slightly, but significantly, less effective in suppressing penetration
resistance than AOPP (Fig. 4). This finding was in contrast to results using nominally
susceptible seedling leaves where the effects of AOPP and OH-PAS on penetration
resistance were indistinguishable [16, 17].
As in previous studies using oat seedlings [12, 14–16], decreased resistance to
penetration following AOPP treatment of adult leaves was correlated to reduction in
the frequency and intensity of localized autofluorescent host cell responses associated
with fungal contact. This clearly suggests that in adult leaves also, autofluorogens
accumulating in contacted epidermal cell wall regions and subtending papillae are
phenolic in nature.
OH-PAS, which was slightly less effective than AOPP in reducing penetration
resistance, did not reduce the frequency of appressorium-associated localized
autofluorescence in adult plant resistant fifth leaves of Maldwyn. However OH-PAS
did reduce the intensity of localized autofluorescence in these fifth leaves (Fig. 3). The
results with Maldwyn contrast to those from the fifth leaves of the susceptible cv. Selma
(Figs 2 and 3) and to oat and barley seedling leaves where OH-PAS always decreased

138 T. L. W. Carver et al.
the frequency and intensity of localized autofluorescent host cell responses [16, 17]. It
may be that differences in leaf architecture prohibited OH-PAS from reaching the
epidermis of Maldwyn’s fifth leaves in sufficient concentration to have full effect.
Alternatively it may be that autofluorogenic compounds other than the cinnamyl
alcohols or later-formed products, also contribute to the adult plant resistance of
Maldwyn.
AOPP and OH-PAS treatments almost totally suppressed whole-cell auto-
fluorescence. In water-treated leaves, the frequency of whole-cell autofluorescence was
variable between experiments. However, it was always infrequent in the susceptible
genotype Selma (associated with ! 5% of appressoria), but was seen quite regularly
in the adult plant resistant Maldwyn (associated with between approx. 14% and 35%
of appressoria). In Experiments 1 and 2, whole-cell autofluorescence was seen at a
similar frequency in seedling and in adult leaves, and so alone cannot account for adult
plant resistance expressed in successively formed leaves.
Preliminary observations showed a negative correlation between occurrence of
whole-cell autofluorescence and the ability of epidermal cells both to take up the vital
dye neutral red and to respond to plasmolysing treatments. This result was comparable
to findings from the barley mildew system [26] which led Koga et al. to suggest that
whole-cell autofluorescence accompanies cell death. We cannot be sure, however, that
treatment with AOPP or OH-PAS, which almost totally prevented whole-cell
autofluorescence, also prevented cell death. It is possible that cell death occurred but
that treatment with inhibitors prevented the accumulation of autofluorogenic phenolics
in dead cells. Nevertheless, cell death was reported as a response to E. graminis in oat
and barley genotypes with no known race-specific resistance [4, 8, 11, 15]. It is
commonly associated with incompatibility conditioned by single gene controlled
resistance in barley true leaves [2, 35] and coleoptiles [2, 6, 7] and in oat, barley and
wheat leaves attacked by inappropriate formae speciales of E. graminis [15]. In all cases,
AOPP treatment reduced the proportion of cells that died in response to attack
[15, 35], and OH-PAS treatment had a similar effect in suppressing cell death when it
was tested in an incompatible E. graminis–barley interaction [35]. Similarly, in the
wheat stem rust-Sr-5 system, OH-PAS treatment reduced hypersensitive mesophyll cell
death due to incompatibility [28]. Taken together, these results strongly suggest that
the cell death response in cereals under attack by obligate biotrophic fungal pathogens
accompanies the engagement of responses which involve the synthesis of cinnamyl
alcohols or later-formed compounds derived from cinnamyl alcohols. Cinnamyl
alcohols provide a substrate for peroxidase in polymerization reactions. Increased
peroxidase activity is implicated in the responses of oat, barley and wheat to E. graminis
attack [5, 20, 25, 29, 34, 36] and peroxidation is associated with the production of free
radicals [20, 25]. Free radicals have been implicated in cell death associated with
several plant diseases [20, 25, 32, 33]. It may be that inhibition of cinnamyl alcohol
production by OH-PAS or AOPP treatment, deprives attacked cells of a substrate
necessary for peroxidase activity, and thus free radical production leading to cell death
does not occur.
Overall, our results indicate that phenolic compound synthesis, while important, was
not the only factor contributing to adult plant resistance in Maldwyn oat. Indeed, the
level of contribution made to resistance by newly synthesized phenolics in upper leaves

Phenolic biosynthesis inhibitors 139
appeared to be similar to that in seedling leaves. Thus, analysis of variance of
penetration data showed no significant interaction between leaf position and chemical
treatment in Experiments 1 or 2. If the contribution to resistance of phenolic
compounds varied between leaves, a significant interaction would have been expected.
This suggests that some other, unknown factor(s), either pre-formed or synthesized in
response to pathogen attack also contributed to adult plant resistance. A combination
of this unknown factor(s) together with the effects of phenolic compounds, apparently
conveys the relatively high level of penetration resistance expressed in the upper leaves
of Maldwyn.
All our previous studies using AOPP and OH-PAS [12, 14–17, 35], and the majority
of other studies of resistance to cereal powdery mildew, have used temperatures close
to optimal for powdery mildew development, i.e. 20 °C. Here we also studied plants
acclimated at, and incubated under, relatively low temperature conditions (10 °C).
AOPP treatment substantially increased penetration success at both temperatures
indicating the importance of phenolics at both temperatures. The lack of statistical
interaction between the effects of AOPP-treatment and temperature emphasizes the
similarity of the contribution of phenolics at both temperatures. This was true in
seedling and in adult (fifth) leaves. At both temperatures, AOPP treatment also
effectively reduced the frequency and intensity of localized autofluorescent responses,
and suppressed cell death.
The current results suggest that, overall, fewer haustoria were formed under the
10 °C than the 20 °C condition. Mildew development and epidermal host cell responses
are slower at 10 °C. To compensate for this, we allowed more incubation time for
penetration to be accomplished at 10 °C. However, we were unable to extend
incubation beyond 72 h under the 10 °C condition because the tips of excised leaves
started to show signs of physiological stress (yellowing). In other experiments (Zhang,
unpublished), powdery mildew development at 10 °C and 20 °C was studied on leaves
of intact plants. In these experiments leaves were fixed 96 h after inoculation, and there
was no difference between incubation temperatures in the final proportion of
appressoria that penetrated. Thus, in the current study 72 h may have been insufficient
for some late developing germlings to complete the processes of penetration.
From the present study it seems clear that while phenolic compound synthesis
contributes greatly to the high level of adult plant resistance expressed by Maldwyn,
and its hybrid derivative line OM1387, it is not the only component of adult plant
resistance to penetration. It seems that the processes of phenolic compound synthesis
in response to E. graminis attack have similar effects in adult and in seedling leaves
regardless of eventual adult plant resistance [12, 16, 17]. By plant breeding, it may
prove possible to enhance the production of phenolic compounds produced in response
to disease attack, and thus increase the effectiveness of disease resistance reliant upon
phenolic production. It should then prove feasible to combine this attribute with other,
physiologically independent, resistance mechanisms limiting the success of attempted
infection. These may include the unknown penetration resistance factor(s) in adult
plant resistance, silicon accumulation in papillae and cell walls beneath appressoria
[13] and other papilla or host cell wall characteristics. Such mechanisms may also be
combined with others that limit the growth rate and sporulation capacity of colonies
through limitation of haustorial efficiency and size [10]. The combination of these

140 T. L. W. Carver et al.
mechanisms could provide a complex series of barriers which are unlikely to be
overcome by simple changes in the pathogen genotype and prove extremely durable
in the field.
A cooperative investigation of the Institute of Grassland and Environmental Research,
Plas Gogerddan, Aberystwyth, U.K.; and the Department of Plant Pathology,
University of Minnesota, St Paul, MN, U.S.A. Research supported by MAFF (U.K.),
BBSRC (U.K.) BAGEC initiative, NATO Collaborative Research Grant 00441, and
by Project 22-70 of the Minnesota Agricultural Experiment Station. Published as
Paper u22,227 of the scientific journal series of the Minnesota Agricultural Experiment
Station. Mention of a trademark name or proprietary product does not constitute a
guarantee by the University of Minnesota or by IGER.
REFERENCES
1. Abeles RH, Maycock AL. 1976. Suicide enzyme inactivators. Accounts of Chemical Research 9 : 313–319.2. Aist JR, Bushnell WR. 1991. Invasion of plants by powdery mildew fungi, and cellular mechanisms
of resistance. In: Cole GT, Hoch HC, eds. The Fungal Spore and Disease Initiation in Plants and Animals.New York: Plenum Press 321–345.
3. Amrhein N, Godecke KH. 1977. a-Aminooxy-β-phenylpropionic acid—a potent inhibitor of L-phenylalanine ammonia lyase in �itro and in �i�o. Plant Science Letters 8 313–317.
4. Asher MJC, Thomas CE. 1983. The expression of partial resistance to Erysiphe graminis in spring barley.Plant Pathology 32 : 79–89.
5. Bryngelsson T, Collinge DB. 1992. Biochemical and molecular analysis of the response of barley toinfection by powdery mildew. In: Shewry PR, ed. Biotechnology in Agriculture Series, Vol. 5. Barley:Genetics, Biochemistry, Molecular Biology and Biotechnology. Wallingford: CAB International, 459–480.
6. Bushnell WR, Dueck J, Rowell JB. 1967. Living haustoria and hyphae of Erysiphe graminis f.sp. hordeiwith intact and partly dissected host cells of Hordeum �ulgare. Canadian Journal of Botany 45 : 1719–1732.
7. Bushnell WR, Liu Z. 1994. Incompatibility conditioned by the Mla gene in barley: Timing of the effectof cordycepin on the hypersensitive response. Physiological and Molecular Plant Pathology 44 : 389–402.
8. Carver TLW. 1986. Histology of infection by Erysiphe graminis f.sp. hordei in spring barley lines withvarious levels of partial resistance. Plant Pathology 35 : 232–240.
9. Carver TLW, Carr AJH. 1977. Race non-specific resistance of oats to primary infection by mildew.Annals of Applied Biology 86 : 29–36.
10. Carver TLW, Carr AJH. 1978. The early stages of mildew colony development on susceptible oats.Annals of Applied Biology, 89 : 201–209.
11. Carver TLW, Carr AJH. 1980. Some effects of host resistance on the development of oat mildew.Annals of Applied Biology, 94 : 290–293.
12. Carver TLW, Robbins MP, Zeyen RJ. 1991. Effects of two PAL inhibitors on the susceptibility andlocalized autofluorescent host cell responses of oat leaves attacked by Erysiphe graminis DC. Physiologicaland Molecular Plant Pathology 39 : 269–287.
13. Carver TLW, Zeyen RJ, Ahlstrand GG. 1987. The relationship between insoluble silicon and successor failure of attempted primary penetration by powdery mildew (Erysiphe graminis) germlings onbarley. Physiological and Molecular Plant Pathology 31 : 133–148.
14. Carver TLW, Robbins MP, Zeyen RJ, Dearne GA. 1992. Effects of PAL-specific inhibition onsuppression of activated defense, and on quantitative susceptibility of oats to Erysiphe graminis.Physiological and Molecular Plant Pathology 41 : 149–163.
15. Carver TLW, Zeyen RJ, Robbins MP, Dearne GA. 1992. Effects of PAL inhibition on oat, barley,and wheat cell responses to appropriate and inappropriate formae speciales of Erysiphe graminis DC.Physiological and Molecular Plant Pathology 41 : 397–409.
16. Carver TLW, Zeyen RJ, Bushnell WR, Robbins MP. 1994. Inhibition of phenylalanine ammonialyase and cinnamyl alcohol dehydrogenase increases quantitative susceptibility of barley to powderymildew (Erysiphe graminis DC). Physiological and Molecular Plant Pathology 44 : 261–272.
17. Carver TLW, Zeyen RJ, Robbins MP, Vance CP, Boyles DA. 1994. Suppression of host cinnamyl-alcohol dehydrogenase (CAD) increases oat epidermal cell susceptibility to powdery mildewpenetration. Physiological and Molecular Plant Pathology 44 : 243–259.

Phenolic biosynthesis inhibitors 141
18. Douglas SM, Sherwood RT, Lukezic FL. 1984. Effect of adult plant resistance on primarypenetration of oats by Erysiphe graminis f.sp. a�enae. Physiological Plant Pathology 25 : 219–228.
19. Grand C, Sarni F, Boudet AM. 1985. Inhibition of cinnamyl-alcohol-dehydrogenase activity andlignin synthesis in poplar (Populus¬euramericana Dode) tissues by two organic compounds. Planta 163 :232–237.
20. Hertig C, Rebmann G, Bull J, Mauch F, Dudler R. 1991. Sequence and tissue specific expressionof a putative peroxidase gene from wheat (Triticum aesti�um L). Plant Molecular Biology 16 : 171–174.
21. Jones IT. 1982. Transgressive segregation for enhanced level of adult plant resistance to mildew in theoat cross Mostyn¬Maldwyn. Euphytica 32 : 499–503.
22. Jones IT. 1986. Inheritance of adult plant resistance to mildew in oats. Annals of Applied Biology 109 :187–192.
23. Jones IT, Hayes JD. 1971. The effects of sowing date on adult plant resistance to Erysiphe graminis f.sp.a�enae in oats. Annals of Applied Biology 68 : 31–39.
24. Jones IT, Roderick HW. 1986. Sources of resistance in oats to mildew. Welsh Plant Breeding Station,Aberystwyth, Annual Report for 1985, 113–114.
25. Kerby K, Somerville SC. 1989. Enhancement of specific intercellular peroxidases following inoculationof barley with Erysiphe graminis f.sp. hordei. Physiological and Molecular Plant Pathology 35 : 323–337.
26. Koga H, Zeyen RJ, Bushnell WR, Ahlstrand GG. 1988. Hypersensitive cell death, autofluorescence,and insoluble silicon accumulation in barley leaf epidermal cells under attack by Erysiphe graminis f.sp.hordei. Physiological and Molecular Plant Pathology 32 : 395–409.
27. Mayama S, Shishiyama J. 1978. Localized accumulation of fluorescent and u.v.-absorbingcompounds at penetration sites in barley leaves infected with Erysiphe graminis hordei. Physiological PlantPathology 13 : 347–354.
28. Moerschbacher BM, Noll U, Gorrichon L, Reisner HJ. 1990. Specific inhibition of lignificationbreaks hypersensitive resistance of wheat to stem rust. Plant Physiology 93 : 465–470.
29. Rebmann G, Hertig C, Bull J, Mauch F, Dudler R. 1991. Cloning and sequencing of cDNAsencoding a pathogen-induced putative peroxidase of wheat (Triticum aesti�um L.). Plant MolecularBiology 16 : 329–331.
30. Roderick HW, Clifford BC. 1995. Variation in adult plant resistance to powdery mildew in spring oatsunder field and laboratory conditions. Plant Pathology 44 : 366–373.
31. Roderick HW, Jones IT. 1991. The evaluation of adult plant resistance to powdery mildew (Erysiphegraminis f.sp. a�enae) in transgressive lines of oats. Euphytica 53 : 143–149.
32. Slusarenko AJ, Milosevic N. 1995. Active oxygen metabolism in bean leaves in relation to bacteria-induced hypersensitive cell death. In: Walters DR, Scholes JD, Bryson RJ, Paul ND, McRoberts N,eds. Aspects of Applied Biology, Vol. 42. Physiological Responses of Plants to Pathogens. Warwick: Associationof Applied Biologists, 243–250.
33. Sutherland MW. 1991. The generation of oxygen radicals during host plant responses to infection.Physiological and Molecular Plant Pathology 39 : 70–93.
34. Thordal-Christensen H, Brandt J, Cho BH, Rasmussen SK, Gregersen PL. Smedegaard-Peterson V, Collinge DB. 1992. cDNA cloning and characterization of two barley peroxidasetranscripts induced differentially by the powdery mildew fungus Erysiphe graminis. Physiological andMolecular Plant Pathology 40 : 395–409.
35. Zeyen RJ, Bushnell WR, Carver TLW, Robbins MP, Clark TA, Boyles DA, Vance CP. 1995.Inhibiting phenylalanine ammonia lyase and cinnamyl-alcohol dehydrogenase suppresses Mla1 (HR)but not mlo5 (non-HR) barley powdery mildew resistances. Physiological and Molecular Plant Pathology 47 :119–140.
36. Zhang L, Robbins MP, Carver TLW. 1995. Oat powdery mildew resistance in relation totemperature. In: Walters DR, Scholes JD, Bryson RJ, Paul ND, McRoberts N, eds. Aspects of AppliedBiology, Vol. 42. Physiological Responses of Plants to Pathogens. Warwick: Association of Applied Biologists,345–348.
Copyright © 2022 FDOKUMEN




















