
Peroxynitrite and nitric oxide differ in their effects on pig coronary artery smooth muscle
31
Peroxynitrite and nitric oxide differ in their effects on pig coronary artery smooth muscle Mandeep Walia 2 , Sue E. Samson 1 , Tracey Schmidt 1 , Kelly Best 2 , Melinda Whittington 1 and Chui Yin Kwan 1 , Ashok K. Grover 1,2 Departments of Medicine 1 and Biology 2 , HSC 4N41 McMaster University, 1200 Main Street West Hamilton, Ontario, Canada L8N3Z5. Running Title: Peroxynitrite generators and coronary artery Corresponding Author: Dr. A.K. Grover Department of Medicine, HSC 4N41, McMaster University, 1200 Main Street West Hamilton, Ontario L8N 3Z5 Canada Phone: 905-525-9140 x22238 Fax: 905-522–3114 E-mail: [email protected] Copyright 2002 by the American Physiological Society. AJP-Cell Articles in PresS. Published on November 13, 2002 as DOI 10.1152/ajpcell.00405.2002
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Peroxynitrite and nitric oxide differ in their effects on pig coronary artery smooth muscle

Peroxynitrite and nitric oxide differ in their effects on pig coronary artery smooth muscle
Mandeep Walia2, Sue E. Samson1 , Tracey Schmidt1, Kelly Best2, Melinda Whittington1 and
Chui Yin Kwan1, Ashok K. Grover1,2
Departments of Medicine1 and Biology2, HSC 4N41 McMaster University, 1200 Main Street
West
Hamilton, Ontario, Canada L8N3Z5.
Running Title: Peroxynitrite generators and coronary artery
Corresponding Author:
Dr. A.K. Grover
Department of Medicine, HSC 4N41, McMaster University, 1200 Main Street West
Hamilton, Ontario L8N 3Z5 Canada
Phone: 905-525-9140 x22238
Fax: 905-522–3114
E-mail: [email protected]
Copyright 2002 by the American Physiological Society.
AJP-Cell Articles in PresS. Published on November 13, 2002 as DOI 10.1152/ajpcell.00405.2002

Abstract
Peroxynitrite generated in arteries from superoxide and NO may damage their function.
Here we compare the effects of peroxynitrite and peroxynitrite/NO generating agents SIN-1 (3-
morpholinosydnonimine hydrochloride), SNAP (S-nitroso-N-acetylypenicillamine), SNP
(sodium nitroprusside) and NONOate (Spermine NONOate) on pig coronary artery. De-
endoethelialized artery rings were pretreated with these agents and then washed before
examining their contractility. The pretreatment with all the agents (200 FM) results in a
decrease in the force of contraction in response to the sarco/endoplasmic Ca2+ pump (SERCA)
inhibitor cyclopiazonic acid (CPA): SNAP > NONOate $ peroxynitrite $SIN-1 > SNAP.
Pretreatment with SNAP, NONOate or SIN-1 also inhibits the force of contraction produced with
30 mM KCl, with SNAP being the most potent. Including catalase plus superoxide dismutase
during the preincubation has no effect. Including an NO-scavenger (2-(4-carboxyphenyl)-4,4,5,
5-tetramethylimidazoline-1-oxyl-3-oxide) or a guanylate cyclase inhibitor (1H-
[1,2,4]Oxadiazolo[4,3-a]quinoxalin-1-one) partially protects against SNAP. Pretreatment of
cultured cells with peroxynitrite but not with SNAP inhibits the Ca2+ transients produced in
response to CPA. Pretreating isolated membrane vesicles with peroxynitrite inhibits the Ca2+
uptake due to the SERCA pump, with all the other agents being less effective. Thus
peroxynitrite and NO both inhibit the CPA induced contractions in de-endothelialized artery
rings - peroxynitrite by damage to the SERCA pump and NO possibly by a step downstream
from the increase in cytosolic Ca2+.
Keywords: ATPase, free radicals, oxidative stress, vascular diseases, ischaemia-reperfusion.

INTRODUCTION
Reactive oxygen species (ROS) are pivotal as intermediates in mitochondrial function and
as signalling molecules in many physiologically important processes (5,14,22,25). Their rates of
formation and dissolution are rigorously controlled. The enhanced production of ROS during
ischaemia-reperfusion and in vascular pathologies such as atherogenesis and inflammation is
implicated in cardiovascular damage (1,9,22). The damage caused by ROS to transport processes
may disturb homeostasis in vascular smooth muscle and endothelium. Such damage is not
uniform to all transport processes. Ca2+-dependent K+-channels in rat cerebral artery smooth
muscle cells are more sensitive to damage by ROS than are voltage sensitive Ca2+ channels (6).
In pig coronary artery smooth muscle, the sarco/endoplasmic reticulum Ca2+ (SERCA) pump is
more sensitive to damage by peroxide and superoxide than the plasma membrane Ca2+ (PMCA)
pump (16,19). Consequently, contractions to angiotensin II and to the SERCA pump inhibitors
thapsigargin and cyclopiazonic acid (CPA) are more sensitive to such damage by pretreatment of
the arteries with peroxide and superoxide than are the contractions to membrane depolarization
by KCl or endothelins (12,20). The damage may also be tissue dependent, e.g., smooth muscle
may be more sensitive to damage by peroxide than endothelium (18). Individual physiological
processes within each tissue may also differ in their susceptibility to different species of ROS.
Peroxynitrite differs from ROS such as peroxide, superoxide and hydroxyl radicals
(3,24,33). It can modify proteins by nitrosylation of thiols and tyrosine residues in addition to the
protein and lipid oxidation pathways used by other ROS. The effects of peroxynitrite on the pig
coronary artery have not been reported. Peroxynitrite is produced in arteries from nitric oxide
and superoxide generated by macrophages during injury or atherogenic inflammatory responses.

It may also be produced in the arteries during ischaemia-reperfusion (1,3,24,30). In the isolated
perfused rat heart, it induces both vasodilation and impaired vascular relaxation (41). Here, we
set out to determine the effects of peroxynitrite on pig coronary artery smooth muscle at three
different levels of organization: contractility in artery rings, cytosolic Ca2+ ([Ca2+]i) transients in
cultured cells and oxalate-stimulated-azide insensitive Ca2+ uptake in the SERCA enriched
subcellular membrane fraction. The effects of peroxynitrite can only be determined by adding a
bolus of this agent since it is unstable at the physiological pH. There are no agents that can
slowly generate only peroxynitrite. Most peroxynitrite generating agents produce mixtures of
various ROS including NO. NO generating agents may also cause S- and O-nitrosylation of
proteins although it is not clear if they are all equally effective in doing so (5,24). Therefore, we
also compared the effects of peroxynitrite with different peroxynitrite/NO-generating agents.
EXPERIMENTAL METHODS
Contractility Experiments: Pig hearts were obtained from Maple Leaf Meats (Burlington,
Canada) and placed immediately in an ice cold physiological saline solution (20). The left anterior
descending coronary artery was dissected, de-endothelialized and placed in a Krebs’ solution
bubbled with 95% O2 - 5% CO2. The Krebs’ solution contained (in mM): 115 NaCl, 5 KCl, 22
NaHCO3, 1.7 CaCl2, 1.1 MgCl2, 1.1 KH2PO4, 0.03 EDTA, and 7.7 glucose. From the middle
region of each artery, 3 mm long rings were cut and hung under 3 g tension in 4 ml organ baths at
37 E C for contraction as previously described (20). The maximum force of contraction was
determined for each artery ring by adding 3 M KCl to a final concentration of 60 mM. The rings
were then washed five times with 5 ml Krebs’ solution over a total period of 20 min and then
allowed to equilibrate for another 20 min. The force of contraction with 30 mM KCl (20) was
examined and the tissues were washed again. Aliquots of peroxynitrite (Calbiochem) under

nitrogen were stored at -80EC and used within 3 months. The peroxynitrite concentration was
determined by its absorbance at 302 nm (extinction coefficient = 1670 M-1cm-1) in 0.1M NaOH
immediately before use. SNAP (S-nitroso-N-acetylypenicillamine) and SNP (sodium
nitroprusside) were prepared in chilled Krebs’ solution just before use. NONOate (Spermine
NONOate), CPTIO: 2-(4-carboxyphenyl)-4,4,5, 5-tetramethylimidazoline-1-oxyl-3-oxide and
SIN-1 (3-morpholinosydnonimine hydrochloride) were prepared as concentrated stock solutions
in water and stored at -80 EC. Concentrated stock solutions of CPA and ODQ (1H-
[1,2,4]Oxadiazolo[4,3-a]quinoxalin-1-one) were prepared in dimethylsulfoxide and stored at -80
EC. Aliquots of the frozen solutions were thawed only once. In all the instances aliquots of 4 Fl
were added to the organ baths containing 4 ml of Krebs’ solution. Peroxynitrite or other agents
were added and the tissues were incubated for another 30 min. They were then washed again five
times with 5 ml Krebs’ solution over a total period of 45 min and allowed to equilibrate for
another10 min. Their contraction to 30 mM KCl was examined for 20 min, the tissues washed
again over a 30 min period and then used for monitoring their contraction with 10 FM CPA.
[Ca2+] i measurements: Smooth muscle cells in passage 4 were cultured on cover slips and used
for the [Ca2+] i measurement experiments. Isolation and characteristics of these cells have been
previously described (15,20). These cells express smooth muscle actin, contract to A23187 and
show increase in [Ca2+] i in response to CPA, thapsigargin, angiotensin II or endothelin-1 but not
to membrane depolarization by KCl. The cover slips coated with cultured cells were removed
from the culture medium and rinsed with a solution containing (in mM): 115 NaCl, 5.8 KCl, 2
CaCl2, 0.6 MgCl2, 12 glucose, 25 HEPES-Na, pH 7.4 at 37EC. These were then incubated in this
buffer at 37EC for 30 min in the presence of 200 FM peroxynitrite or other agents, washed 3
times to remove these agents, loaded with FLUO 3/AM and then used for [Ca2+] i measurement.

The total time period between the end of the pretreatment with any agents and the [Ca2+] i
measurements was 90 min.
Ca2+ uptake in isolated membranes: Subcellular membrane fraction F3 enriched in SR
was isolated from smooth muscle. Isolation of these membranes and their characteristics have
been previously reported (16,19,21). The membranes were incubated with peroxynitrite or other
agents at 37EC and then 1 mM DTT was added to quench any remaining peroxynitrite or peroxide
produced from the decomposition. The samples were placed on ice until use. Ca2+ uptake was
carried out as described earlier (16,19). The final reaction mixture consisted of creatine kinase
(70 units/ml) and the following concentrations (in mM) of the other components 100 KCl, 5 Na-
azide, 5 MgCl2, 5 ATP, 10 creatine phosphate, 1 EGTA, 0.85 CaCl2 (plus trace amounts of
45Ca), 30 imidazole-HCl (pH 6.8 at 37EC), 0.33 DTT, 0 or 5 K-oxalate (with F3 fraction) and 0 or
5 K-phosphate (with F2 fraction, pH 6.8 at 37EC). This resulted in 5 µM free [Ca2+] as described
earlier (16,19).
Nitroblue tetrazolium reduction assay: Reduction of nitroblue tetrazolium was used to monitor
superoxide generation as described earlier (23). Briefly, 75 µl aliquots of each agent were
prepared and pipetted into a 96 well plate on ice and 175 µl of ice-cold nitroblue tetrazolium
solution was added with or without catalase and superoxide dismutase. The final reaction
mixtures contained: 200 µM of peroxynitrite or other specified agent, 1.5 mM nitroblue
tetrazolium, 0 or 200 units each of catalase and superoxide dismutase in the HEPES-Krebs’
solution described above for the contractility experiments. Absorbance of the samples at 595 nm
was monitored immediately, then the foil-wrapped samples were placed in a 37EC oven and
absorbance monitored again after 30, 60 and 90 min. The difference in the absorbance changes

with and without catalase plus superoxide dismutase after 30 min were used to express the
activity.
Data Analysis: Curve fitting was carried out by nonlinear regression using the FigP software
(BioSoft, Ancaster, Canada). Statistical significance was determined using the Student’s t-test.
All experiments were replicated 3-5 times.
RESULTS
Effects of pretreatment with peroxynitrite and NO-generating agents on contraction: De-
endothelialized arteries in Krebs’ solution contract when challenged with the SR Ca2+ pump
inhibitor CPA (31) or when membranes are depolarized with high concentrations of KCl. The
mean force of contraction produced by 30 mM KCl is 23.9 ± 1.2 mN (mean ± SEM, 100 tissues)
while that with 10 FM CPA, it is 4.9 ± 0.4 mN (100 tissues). In order to compare the damage to
the artery rings by various agents with that of peroxide and superoxide reported previously, we
used the same protocols as described earlier and given in the Experimental Methods (20). This
protocol includes extensive washing to remove from the organ baths any agents and their
decomposition products used during the pretreatment. This was done to ensure that these
substances would not directly interfere in the subsequent measurements. Fig. 1 shows the
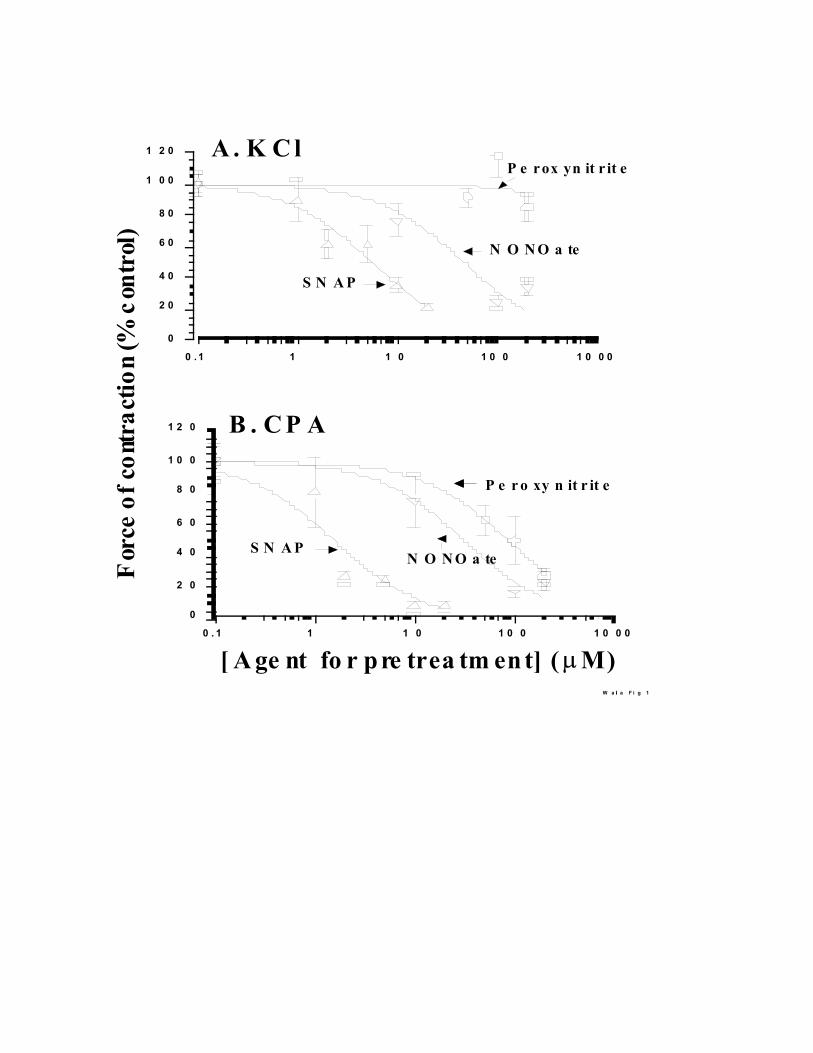
concentration-dependence of the effects of pretreatment with peroxynitrite, SNAP or NONOate.
The pretreatment with peroxynitrite does not alter the force of contraction produced with 30 mM
KCl but SNAP or NONOate inhibit the contraction with 30 mM KCl with SNAP (r2 = 0.9454, IC50
= 5.7 ± 0.9 FM) being more potent than NONOate ( r2 =0 .9366, IC50 = 47 ± 15 FM) (Fig. 1A).
Pretreatment with these agents decreases the force of contraction produced with CPA with the
following order of potency: SNAP ( r2= 0.9145, IC

50 = 1.6 ± 0.5 FM) > NONOate (r2 =0.9719, IC50 = 32 ± 8 FM) > peroxynitrite (r2 = 0.9935, IC50
= 87 ± 6 FM) (Fig. 1B). We also determined the effects of single concentrations of SNP and
SIN-1. Pretreatment with 200 FM SNP decreased the force of contraction with KCl and CPA to
76 ± 9 and 61 ± 16 % of the control, respectively (data not shown). The corresponding values with
SIN-1 were 62 ± 11 and 32 ± 10 %. The rank order of effectiveness of these agents (at 200 FM)
for the CPA contraction is SNAP > NONOate $ peroxynitrite $ SIN-1 > SNP. This order is
similar but not identical to that obtained for the contractions with KCl (SNAP $ NONOate > SIN-
1 > SNP = peroxynitrite). Since, peroxynitrite was added as a bolus in the presence of 5-fold
molar excess of NaOH, we also examined the effects of pretreatment with NaOH up to 1000 FM.
Preteatment with NaOH does not significantly affect the force of contraction (p > 0.05). For
example, in the tissues pretreated with 1000 FM NaOH, the force of contraction with KCl is 96 ±
8 % of the control and with CPA it is 86 ± 11 %.
Effects of pretreatment with various agents with catalase plus superoxide dimutase on contraction:
To ascertain which of these agents produced measurable amounts of superoxide, we used an assay
based on change in the absorbance of NBT upon reduction. We incubated these agents with NBT
with or without excess catalase plus SOD . SIN-1 shows the largest amount of SOD sensitive
reduction of NBT and peroxynitrite is next (Table 1). The change in the absorbance of NBT with
SNAP, NONOate and SNP was no different from the water used as a control.
Pretreatment with peroxide and superoxide is known to inhibit the contractions to CPA
(20). Therefore, we considered the possibility that the hydrogen peroxide or superoxide produced
by decomposition of these agents may cause the inhibition of the contractions observed in Fig. 1.
We have shown previously that catalase plus SOD can overcome the effect of superoxide, and that
catalase alone can abolish the effect of hydrogen peroxide in this system (11,16). Hence, we

examined the effects of preincubation with SIN-1, peroxynitrite and SNAP with or without excess
catalase plus SOD. Fig. 2 shows that peroxynitrite inhibited only the CPA induced contractions
(p > 0.05) and all the other agents significantly decreased the force of contractions with KCl and
CPA (p < 0.05). Catalase plus SOD had no effect (p > 0.05) indicating that the observed effect is
primarily due to peroxynitrite and not due to its decomposition products.
Further experiments on the effects of SNAP on force of contraction: Since SNAP has the highest
potency, we considered the possibility that a stable decomposition product of SNAP was
responsible for the inhibition. To test this hypothesis, we incubated a stock solution of SNAP in
Krebs’s overnight at 37 E C and then pretreated the artery rings with it. Whereas a precincubation
with 10 FM freshly prepared SNAP decreases the force of contraction with KCl to 35 % of the
control and with CPA to 7 % of the control, in 3 tissues pretreated with the decomposed SNAP, the
force of contraction is 95 % of the control with KCl and 103 % with CPA. Thus the decomposed
SNAP is ineffective. This loss of ability of SNAP to inhibit the force of contraction indicates that a
metastable product of SNAP (such as NO) is responsible for the inhibition. Since SNAP is known
to generate NO we next conducted an experiment in which CPTIO (2-(4-carboxyphenyl)-4,4,5, 5-
tetramethylimidazoline-1-oxyl-3-oxide) was used to quench the NO generated by SNAP (32). In
preliminary experiments we established that CPTIO was active in that it inhibited the endothelium
dependent relaxation, thereby showing that CPTIO can effectively quench the NO produced by the
endothelium. We next examined if CPTIO could quench the NO produced by SNAP and thus
overcome its inhibition of the CPA induced contraction in de-endothelialized artery rings. We
preincubated the de-endothelialized artery rings with 10 FM SNAP with and without 50 FM
CPTIO (data not shown). Including this concentration of CPTIO in the preincubation mixture does
not overcome the effect of 10 FM SNAP on the force of contraction produced by 30 mM KCl or

by CPA. We then preincubated the de-endothelialized artery rings with 10 FM SNAP with and
without 200 FM CPTIO. It alleviates the inhibition by 10 FM SNAP significantly (p < .05) but
only partially on the force of contraction produced with KCl or CPA (Fig. 3).
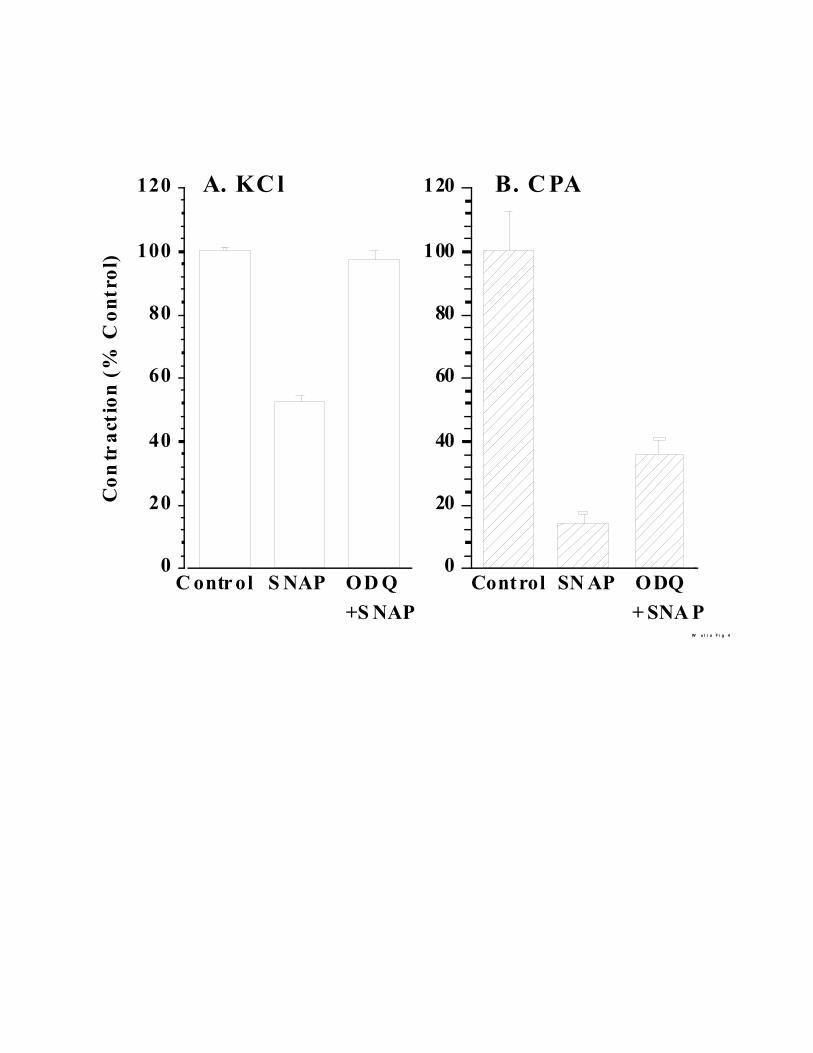
The established mode of action of NO on smooth muscle is stimulation of guanylate
cyclase (24) and hence we next examined the effects of the guanylate cyclase inhibitor 1H-
[1,2,4]Oxadiazolo[4,3-a]quinoxalin-1-one (ODQ) (38). In a preliminary experiment we
determined that ODQ at 10 FM completely inhibits the endothelium dependent relaxation in this
tissue but preincubating de-endothelialzed artery rings with ODQ does not affect the force of
contraction with KCl or CPA (data not shown). We next preincubated the de-endothelialized
artery rings with 10 FM SNAP with and without 10 FM ODQ (Fig. 4A). Including ODQ in the
preincubation mixture completely overcomes the effect of 10 FM SNAP on the force of
contraction produced by 30 mM KCl. However, its effect is only partial when the CPA induced
contraction is examined (Fig. 4B).
Effects of peroxynitrite pretreatment on [Ca2+] i transients: Smooth muscle cells cultured from the
coronary artery show an increase in [Ca2+] i transients when challenged with 10 FM CPA. When
cells are pretreated with 200 FM peroxynitrite and then washed as described in the Experimental
Methods, a challenge with CPA gives a smaller increase in [Ca2+] i transients (p < 0.05, Fig. 5).
Including catalase plus SOD with peroxynitrite does not alter its effects (data not shown).
However, a similar pretreatment with 200 FM SNAP does not significantly diminish the CPA
induced [Ca2+] i transients (p > 0.05, Fig. 5) suggesting that peroxynitrite and SNAP act via
different mechanisms. This result is in contrast to the contractility experiments where pretreatment
with this concentration of SNAP decreases the force of contraction with CPA by > 90 % (Fig. 1).

Effects of pretreatment with various agents on Ca2+-uptake in isolated membrane vesicles: We next
examined the possibility that the pretreatment with these agents may damage the SERCA pump in
the the SR-enriched fraction F3 isolated from pig coronary artery smooth muscle. The subcellular
fractionation and Ca2+ uptake properties of F3 have been characterized previously (17,21). With
these membranes, one can use an ATP-regenerating system to obtain Ca2+ uptake which is linear
with time for up to 2 h. The uptake due to the SR in this fraction is ATP-dependent, azide
insensitive, stimulated by oxalate that prevents back flux of Ca2+, and is inhibited by CPA. In
preliminary experiment we observed that pretreating the membranes with 100 FM peroxynitrite
gave similar values for inhibition of the Ca2+ uptake as those observed for the CPA-induced
contractions with 200 FM peroxynitrite. Therefore, we compared the effects of pretreating the
membranes with 100 FM of each of these agents on the Ca2+ uptake. Fig. 6 shows that: (a)
pretreating the F3 membrane vesicles with 100 FM peroxynitrite results in an 80 % inhibition of
the Ca2+ uptake, (b) a similar pretreatment with 100 FM SNAP or SNP leads to marginal inhibition
of the uptake and 100 FM NONOate or SIN-1 do not inhibit the uptake significantly (Fig. 6), and
(c) the inhibition of the uptake by the pretreatment with any of the agents is not overcome by
including catalase plus SOD during the pretreatment. Thus the rank order for inhibition appears to
be peroxynitrite >> SNP $SNAP > NONOate > SIN-1.
DISCUSSION
The Results show that: (a) pretreating the artery rings with peroxynitrite and NO-
generating agents decreases the force of contraction in the following decreasing order of
effectiveness:
SNAP > NONOate $ peroxynitrite $ SIN-1 > SNP with CPA and SNAP $ NONOate > SIN-1
with KCl (no inhibition with SNP and peroxynitrite), (b) the force of contraction with none of the

agents is significantly altered when catalase + SOD are included during the pretreatment, (c) the
effectiveness of SNAP is decreased with CPTIO or ODQ, (d) pretreatment of cultured cells with
peroxynitrite but not with SNAP decreases the CPA induced Ca2+i -transients, and (e) in isolated
membranes, the Ca2+ uptake due to the SERCA pump is inhibited predominantly by pretreatment
with peroxynitrite with the order of effectiveness of the different agents being peroxynitrite >>
SNP $SNAP $ NONOate = SIN-1. The Discussion focuses on the methods used in this study,
comparison of these results with the literature, a proposed model to explain the results and possible
biological implications of these findings.
The agents used during the preincubation or their decomposition products may react with
the reagents used subsequently in the assays. To avoid this possibility, we washed the tissues after
the treatment with the agents used for the pretreatment and before adding any other reagents. We
have shown earlier that including catalase plus SOD prevents the effects of superoxide and
peroxide on artery rings, cultured cells and isolated membranes (11,23). Thus protocols used in
the study would monitor the damage produced by the agents during pretreatment on contractility
or other properties rather than their effects when present during the course of the assays. This
distinction is important in comparing this work with the literature. Since one can not obtain pure
PM or SR membranes from the coronary artery smooth muscle, we used the effect of oxalate on
the inhibition of back flux of Ca2+ to identify the activity of the SERCA pump (16,17,21). The
membrane fractions used in this study and the characteristics of the oxalate-stimulated Ca2+ uptake
have been established previously (21). Thus the methods for monitoring contractility, [Ca2+] i and
SR Ca2+ pump activity in isolated membranes have been previously validated using pig coronary
artery (15,20).

We first conducted the work on artery rings with peroxynitrite. However, the damage
produced in this study required high micromolar concentrations of this agent. Since in these
experiments the peroxynitrite is extremely short lived but in vivo peroxynitrite may be generated
constantly during ischaemia-reperfusion or inflammation like responses during atherosclerosis, we
then used the peroxynitrite generating agent SIN-1 (23,24). Since all peroxynitrite generating
agents also produce NO, we also used several NO generating agents. None of the NO generating
agents produce only one species of NO and even NO gas is converted to other species and/or
oxidized to nitrites and nitrates under aerobic conditions (34). The various agents not only produce
NO at different rates but may also produce different species of NO (2,23,24,35). Predominant
species generated by some of the agents are NONOate and SNP (NO.), SNAP (NO+) and Angeli’s
salt (NO-) (24). We tested SNAP and NONOate and they were both able to produce relaxation in
the de-endothelialized arteries which had been precontracted with KCl . However, in inhibiting
the contractility SNAP was the most potent, even more potent than SIN-1 and peroxynitrite. In the
NBT reduction assay, only SIN-1 and peroxynitrite showed any production of superoxide
indicating that SNAP, NONOate and SNP generated predominantly various species of NO.
Thus we verified under our experimental conditions that SIN-1 produces significantly more
superoxide than SNAP. SNAP is in fact a very potent NO release agent. It produces NO at a rate
several hundred times faster than SIN-1 (23,24). SNAP activates guanylate cyclase at lower
concentrations than spermine-NONOate and the rate of NO release from SNAP is much faster,
than from the NONOate (2,24). The various agents not only produce NO at different rates but may
also produce different species of NO (2,23,24,35). Even when NO generated as a gas may form
different species depending not only on the aqueous environment but also in a tissue dependent
manner (10,34). In addition S-nitrosothiols like SNAP decomposes spontaneously in solution to

their disulfide forms while generating NO. However, decomposed SNAP by itself was not
effective though one can not rule out that the ability of SNAP to generate simultaneously NO and
disulfide groups that would allow it to act differently than other agents.
We have shown earlier that the pretreatment with peroxide or superoxide causes a decrease
in: (a) the SR Ca2+ pump activity in the coronary artery, (b) Ca2+ -transients produced by CPA in
cultured smooth muscle cells, and (c) tissue contractility to CPA and to agents such as angiotensin
II which depend on the mobilization of the SR Ca2+ (18,20). Peroxide and superoxide act simply
in oxidation reactions presumably involving thiol groups (3,24). However, the action of
peroxynitrite or SIN-1 did not occur via decomposition into superoxide and peroxide as their
effects were insensitive to the inclusion of catalase plus SOD during the pretreatment.
Peroxynitrite has been shown to cause nitrosylation of thiol groups and tyrosine residues
(27,36,40). Such nitrosylation of SERCA2a in slow twitch skeletal muscle decreases its enzymatic
activity (42). Therefore, the most likely mechanism appears to be inactivation of the SR Ca2+
pump by peroxynitrite directly or through SIN-1. A direct effect of peroxynitrite on the contractile
machinery or on voltage dependent Ca2+ channels is thus ruled out since contraction with 30 mM
KCl was not affected. However, one cannot rule out that pathways such as store depletion
dependent Ca2+ channels are also affected by the treatment.
Several effects of NO have been reported on vascular smooth muscle. An NO mediated
increase in store depletion dependent Ca2+ entry has been reported in some studies and decrease in
others (8,13). NO binding to heme group of guanylate cyclase activates it thus producing an
increase in cellular cGMP levels (2,24). NO mediated decrease in vascular reactivity has been
proposed to occur by various mechanisms (7,26,28,37). Our results show that pretreatment with
SNAP inhibits the contractions produced by KCl and CPA. The inhibition is not affected by

CPTIO at 50 FM but it can be overcome partially with 200 FM CPTIO indicating the involvement
of NO. However, even the high concentration of CPTIO used here does not completely overcome
the effects of SNAP possibly because SNAP produces NO very rapidly (2). SNAP produces
NO+. NO+ and NOC are both quenched with CPTIO although NO- produced by Angeli’s salt is
not quenched effectively with this agent (10,29). Since these experiments required pretreatment at
high concentrations of NO for a prolonged period under aerobic conditions, one can not rule out
the conversion of the initially generated NO into other products including the nitroxyl ion.
Gaseous NO under aerobic conditions can be converted to products which no longer react with
CPTIO (34). The effect of SNAP is also overcome in part by the guanylate cyclase inhibitor ODQ
suggesting that cGMP plays a key role in this pathway. However, 10 FM ODQ does not
completely overcome the effect of SNAP even though IC50 for guanylate inhibition of ODQ is 84
nM (38). This may be because ODQ may not as readily cross the cell membranes as NO, NO has
been converted into other products or another pathway is involved in the observed effects of
SNAP. Finally, the pretreatment of cultured cells with SNAP does not produce a decrease in Ca2+
transients in response to CPA. Together, these results suggest that SNAP acts by a step
downstream of Ca2+ mobilization.
Physiological Implications: The effects of peroxynitrite and NO may be important in aging,
ischaemia-reperfusion, inflammation, atherosclerosis and spesis (5,14,22,25). SERCA2a in rat
twitch skeletal muscle shows that nitrosylated tyrosine residues increase with aging (42). Such
data are not currently available for the coronary artery. Damage to the SERCA pump in this tissue
would mean a loss of reactivity to agents such as angiotensin and α-adrenergic agents that depend
on mobilization of the SR Ca2+ pools (4,20). NO signalling pathways are of central importance in
both the maintenance of vascular homeostasis and the progression of vascular disease. Multiple

interactions of NO with other pathways can lead to either vascular protection or potentiation of
inflammatory vascular injury (33). During sepsis, an increase in [Ca2+ ]i may be an early event
responsible for cellular injury. This would normally lead to an increased contractile force in the
affected areas (39). A decrease in the Ca2+ sensitivity of the contractile apparatus would thus
provide a useful protection.
We conclude that peroxynitrite and NO both inhibit the CPA induced contractions in de-
endothelialized artery rings - peroxynitrite by via damage to the SERCA pump and NO at a step
downstream from a change in [Ca2+ ]i as one of the possibilities. Both types of interactions have
important pathophysiological ramifications.
Abbreviations: CPA: cyclopiazonic acid, CPTIO: 2-(4-carboxyphenyl)-4,4,5, 5-
tetramethylimidazoline-1-oxyl-3-oxide, DTT: dithiothreitol, EGTA: ethylene glycol-bis-(β-
aminoethyl ether)-N,N,N’,N’ tetraacetate, NONOate (Spermine NONOate, diazeniumdiolates),
ODQ: 1H-[1,2,4]Oxadiazolo[4,3-a]quinoxalin-1-one, PM: plasma membrane, PMCA: PM Ca2+
pump, ROS: reactive oxygen species, SIN-1: 3-morpholinosydnonimine hydrochloride, SNAP: S-
nitroso-N-acetylypenicillamine, SNP: sodium nitroprusside, SR: sarco/endoplasmic reticulum,
SERCA: SR Ca2+ pump, SOD: superoxide dismutase.
Acknowledgements: The authors thank Dr. N. Narayanan of University of Western Ontario for
critical comments on the manuscript. This work was funded by a grant -in-aid from Heart &
Stroke Foundation of Ontario (Grant # T-4815). AKG received a Career Investigator Award from
the Heart & Stroke Foundation of Ontario.

Figure Legends
Fig. 1. Inhibition of contractility by preatment of de-endothelialized artery rings with different
concentrations of peroxynitrite, SNAP and NONOate. In control tissues, force of contraction
produced with 30 mM KCl is 23.9 ± 1.2 mN (mean ± SEM, 100 tissues). A. Force of contraction
produced with KCl by tissues pretreated the agents specified in the figure, was expressed as
percent of this value. B. The force of contraction with 10 FM CPA, in control tissues is 4.9 ± 0.4
mN (100 tissues) and values for the CPA induced contraction in treated samples are expressed as a
percent of this control. The values are mean ± SEM of several tissues for each concentration of an
agent. The total number of tissues for the curves were: 113 for peroxynitrite, 107 for SNAP and
60 for NONOate. The data were fitted with a simple inhibition model. In A and B, at each
concentration tested the order of inhibition produced by the pretreatment was SNAP > NONOate >
peroxynitrite.
Fig. 2. Contractility experiments with catalase + SOD included during pretreatment. Artery rings
were preincubated with 10 FM SNAP, 200 FM of peroxynitrite (peroxyn), 200 FM NONOate or
200 FM SIN-1, and tested as in Fig. 1. Catalase + SOD was included during preincubation where
shown. A total of 243 tissues were used in all the groups in this experiment. All the values are
percent of control carried out in parallel without any agents added during the preincubation. Mean
± SEM percent values are given.
Fig. 3. Contractility experiments with 200 FM CPTIO included during pretreatment. Artery rings
were preincubated with 10 FM SNAP and tested as in Fig. 1. 200 FM CPTIO was included
during preincubation where shown. The number of tissues used were: 22 (control), 44 (SNAP) and
21 (SNAP + CPTIO). All the values are percent of control carried out in parallel without any
agents added during the preicubation. Mean ± SEM percent values are given.

Fig. 4. Contractility experiments with ODQ included during pretreatment. Artery rings were
preincubated with 10 FM SNAP and tested as in Fig. 1. 10 FM ODQ was included during
preincubation where shown. The number of tissues used were: 25 (control), 19 (SNAP) and 17
(SNAP + ODQ). All the values are percent of control carried out in parallel without any agents
added during the preincubation. Mean ± SEM percent values are given.
Fig. 5. Effects of pretreatment with 200 FM peroxynitrite (peroxyn) or 200 FM SNAP on CPA
induced increase in [Ca2+] i. The values are mean ± SEM from 34 replicates for control, 13 for
peroxynitrite and 9 for SNAP. ‘*’ indicates that the peroxynitrite treatment inhibits the increase in
[Ca2+]i significantly (p < 0.05). SNAP does not produce a significant inhibition ( p > 0.05).
Fig. 6. Ca2+ uptake by SR enriched membranes pretreated with different agents. Membranes
treated without any additives showed an uptake of 804 ± 18 nmol/mg protein. Values for the Ca2+
uptake for the samples treated with the various additives are expressed as percent of this control
value and are mean ± SEM of 6 replicates. Treatment with peroxynitrite significantly (p < 0.05)
inhibited the uptake.

Table 1. SOD sensitive nitroblue tetrazolium reduction
Additive Change in absorbancy
Water 0.029 ± 0.009 (11)
Peroxynitrite (200 FM) 0.127 ± 0.015 (6)
SIN-1 (200 FM) 0.400 ± 0.063 (3)
SNAP (200 FM) 0.015 ± 0.003 (6)
SNP (200 FM) -0.007 ± 0.007 (6)
Nonoate (200 FM) -.018 ± 0.007 (6)
NaOH (200 FM) -0.014 ± 0.008 (6)
Note: The values are mean ± SEM of the number of replicates shown in parentheses.

References
1. Abel, F. L. Myocardial function in sepsis and endotoxin shock. Am.J.Physiol 257[6 Pt 2],
R1265-R1281. 1989.
2. Artz, J. D., Toader, V., Zavorin, S. I., Bennett, B. M., and Thatcher, G. R. In vitro
activation of soluble guanylyl cyclase and nitric oxide release: a comparison of NO donors and
NO mimetics. Biochemistry 40[31], 9256-9264. 8-7-2001.
3. Beckman, J. S. and Koppenol, W. H. Nitric oxide, superoxide, and peroxynitrite: the
good, the bad, and ugly. Am.J.Physiol 271[5 Pt 1], C1424-C1437. 1996.
4. Benkusky, N. A., Lewis, S. J., and Kooy, N. W. Peroxynitrite-mediated attenuation of
alpha- and beta-adrenoceptor agonist-induced vascular responses in vivo. Eur.J.Pharmacol.
364[2-3], 151-158. 1-8-1999.
5. Bergendi, L., Benes, L., Durackova, Z., and Ferencik, M. Chemistry, physiology and
pathology of free radicals. Life Sci. 65[18-19], 1865-1874. 1999.
6. Brzezinska, A. K., Gebremedhin, D., Chilian, W. M., Kalyanaraman, B., and Elliott,
S. J. Peroxynitrite reversibly inhibits Ca(2+)-activated K(+) channels in rat cerebral artery smooth
muscle cells. Am.J.Physiol Heart Circ.Physiol 278[6], H1883-H1890. 2000.
7. Chuprun, J. K. and Rapoport, R. M. Nitroglycerin-induced desensitization of vascular
smooth muscle may be mediated through cyclic GMP-disinhibition of phosphatidylinositol
hydrolysis. Experientia 43[3], 316-318. 3-15-1987.
8. Cohen, R. A., Weisbrod, R. M., Gericke, M., Yaghoubi, M., Bierl, C., and Bolotina,
V. M. Mechanism of nitric oxide-induced vasodilatation: refilling of intracellular stores by

sarcoplasmic reticulum Ca2+ ATPase and inhibition of store-operated Ca2+ influx. Circ.Res.
84[2], 210-219. 2-5-1999.
9. Dhalla, N. S., Elmoselhi, A. B., Hata, T., and Makino, N. Status of myocardial
antioxidants in ischemia-reperfusion injury. Cardiovasc.Res. 47[3], 446-456. 8-18-2000.
10. Ellis, A., Lu, H., Li, C. G., and Rand, M. J. Effects of agents that inactivate free radical
NO (NO*) on nitroxyl anion-mediated relaxations, and on the detection of NO* released from the
nitroxyl anion donor Angeli's salt. Br.J.Pharmacol. 134[3], 521-528. 2001.
11. Elmoselhi, A. B., Butcher, A., Samson, S. E., and Grover, A. K. Free radicals uncouple
the sodium pump in pig coronary artery. Am.J.Physiol 266[3 Pt 1], C720-C728. 1994.
12. Elmoselhi, A. B. and Grover, A. K. Peroxide sensitivity of endothelin responses in
coronary artery smooth muscle: ET(A) vs. ET(B) pathways. Mol.Cell Biochem. 202[1-2], 47-52.
1999.
13. Favre, C. J., Ufret-Vincenty, C. A., Stone, M. R., Ma, H. T., and Gill, D. L. Ca2+ pool
emptying stimulates Ca2+ entry activated by S-nitrosylation. J.Biol.Chem. 273[47], 30855-30858.
11-20-1998.
14. Griendling, K. K. and Ushio-Fukai, M. Reactive oxygen species as mediators of
angiotensin II signaling. Regul.Pept. 91[1-3], 21-27. 7-28-2000.
15. Grover, A. K., Fomin, V. P., and Samson, S. E. Angiotensin II contractions in coronary
artery. Nature of receptors and calcium pools. Mol.Cell Biochem. 135[1], 11-19. 6-15-1994.
16. Grover, A. K. and Samson, S. E. Effect of superoxide radical on Ca2+ pumps of
coronary artery. Am.J.Physiol 255[3 Pt 1], C297-C303. 1988.

17. Grover, A. K. and Samson, S. E. Protection of Ca pump of coronary artery against
inactivation by superoxide radical. Am.J.Physiol 256[3 Pt 1], C666-C673. 1989.
18. Grover, A. K. and Samson, S. E. Peroxide resistance of ER Ca2+ pump in endothelium:
implications to coronary artery function. Am.J.Physiol 273[4 Pt 1], C1250-C1258. 1997.
19. Grover, A. K., Samson, S. E., and Fomin, V. P. Peroxide inactivates calcium pumps in
pig coronary artery. Am.J.Physiol 263[2 Pt 2], H537-H543. 1992.
20. Grover, A. K., Samson, S. E., Fomin, V. P., and Werstiuk, E. S. Effects of peroxide
and superoxide on coronary artery: ANG II response and sarcoplasmic reticulum Ca2+ pump.
Am.J.Physiol 269[3 Pt 1], C546-C553. 1995.
21. Grover, A. K., Samson, S. E., and Lee, R. M. Subcellular fractionation of pig coronary
artery smooth muscle. Biochim.Biophys.Acta 818[2], 191-199. 8-27-1985.
22. Halliwell, B. Antioxidants and human disease: a general introduction. Nutr.Rev. 55[1 Pt
2], S44-S49. 1997.
23. Holm, P., Kankaanranta, H., Metsa-Ketela, T., and Moilanen, E. Radical releasing
properties of nitric oxide donors GEA 3162, SIN-1 and S-nitroso-N-acetylpenicillamine.
Eur.J.Pharmacol. 346[1], 97-102. 4-3-1998.
24. Ignarro, L. J., Napoli, C., and Loscalzo, J. Nitric oxide donors and cardiovascular
agents modulating the bioactivity of nitric oxide: an overview. Circ.Res. 90[1], 21-28. 1-11-2002.
25. Irani, K. Oxidant signaling in vascular cell growth, death, and survival : a review of the
roles of reactive oxygen species in smooth muscle and endothelial cell mitogenic and apoptotic
signaling. Circ.Res. 87[3], 179-183. 8-4-2000.

26. Knapp, L. T., Kanterewicz, B. I., Hayes, E. L., and Klann, E. Peroxynitrite-induced
tyrosine nitration and inhibition of protein kinase C. Biochem.Biophys.Res.Commun. 286[4], 764-
770. 8-31-2001.
27. Kuhn, D. M., Aretha, C. W., and Geddes, T. J. Peroxynitrite inactivation of tyrosine
hydroxylase: mediation by sulfhydryl oxidation, not tyrosine nitration. J.Neurosci. 19[23], 10289-
10294. 12-1-1999.
28. Lee, M. R., Li, L., and Kitazawa, T. Cyclic GMP causes Ca2+ desensitization in
vascular smooth muscle by activating the myosin light chain phosphatase. J.Biol.Chem. 272[8],
5063-5068. 2-21-1997.
29. Li, C. G., Karagiannis, J., and Rand, M. J. Comparison of the redox forms of nitrogen
monoxide with the nitrergic transmitter in the rat anococcygeus muscle. Br.J.Pharmacol. 127[4],
826-834. 1999.
30. Liaudet, L., Soriano, F. G., and Szabo, C. Biology of nitric oxide signaling. Crit Care
Med. 28[4 Suppl], N37-N52. 2000.
31. Lytton, J., Westlin, M., and Hanley, M. R. Thapsigargin inhibits the sarcoplasmic or
endoplasmic reticulum Ca- ATPase family of calcium pumps. J.Biol.Chem. 266[26], 17067-
17071. 9-15-1991.
32. Margulis, A. and Sitaramayya, A. Rate of deactivation of nitric oxide-stimulated soluble
guanylate cyclase: influence of nitric oxide scavengers and calcium. Biochemistry 39[5], 1034-
1039. 2-8-2000.
33. O'Donnell, V. B. and Freeman, B. A. Interactions between nitric oxide and lipid
oxidation pathways: implications for vascular disease. Circ.Res. 88[1], 12-21. 1-19-2001.

34. Ohkawa, T., Hiramoto, K., and Kikugawa, K. Standardization of nitric oxide aqueous
solutions by modified Saltzman method. Nitric.Oxide. 5[6], 515-524. 2001.
35. Pryor, W. A. and Squadrito, G. L. The chemistry of peroxynitrite: a product from the
reaction of nitric oxide with superoxide. Am.J.Physiol 268[5 Pt 1], L699-L722. 1995.
36. Rowan, W. H., III, Sun, P., and Liu, L. Nitration of annexin II tetramer. Biochemistry
41[4], 1409-1420. 1-29-2002.
37. Savineau, J. P. and Marthan, R. Modulation of the calcium sensitivity of the smooth
muscle contractile apparatus: molecular mechanisms, pharmacological and pathophysiological
implications. Fundam.Clin.Pharmacol. 11[4], 289-299. 1997.
38. Schrammel, A., Behrends, S., Schmidt, K., Koesling, D., and Mayer, B.
Characterization of 1H-[1,2,4]oxadiazolo[4,3-a]quinoxalin-1-one as a heme-site inhibitor of nitric
oxide-sensitive guanylyl cyclase. Mol.Pharmacol. 50[1], 1-5. 1996.
39. Song, S. K., Karl, I. E., Ackerman, J. J., and Hotchkiss, R. S. Increased intracellular
Ca2+: a critical link in the pathophysiology of sepsis? Proc.Natl.Acad.Sci.U.S.A 90[9], 3933-
3937. 5-1-1993.
40. Souza, J. M., Giasson, B. I., Chen, Q., Lee, V. M., and Ischiropoulos, H. Dityrosine
cross-linking promotes formation of stable alpha -synuclein polymers. Implication of nitrative and
oxidative stress in the pathogenesis of neurodegenerative synucleinopathies. J.Biol.Chem.
275[24], 18344-18349. 6-16-2000.
41. Villa, L. M., Salas, E., Darley-Usmar, V. M., Radomski, M. W., and Moncada, S.
Peroxynitrite induces both vasodilatation and impaired vascular relaxation in the isolated perfused
rat heart. Proc.Natl.Acad.Sci.U.S.A 91[26], 12383-12387. 12-20-1994.

42. Viner, R. I., Huhmer, A. F., Bigelow, D. J., and Schoneich, C. The oxidative
inactivation of sarcoplasmic reticulum Ca(2+)-ATPase by peroxynitrite. Free Radic.Res. 24[4],
243-259. 1996.
0 . 1 1 1 0 1 0 0 1 0 0 0
[ A ge nt fo r pre trea tm ent] ( µM)
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
For
ce o
f con
trac
tion
(% c
ontr
ol)
0 . 1 1 1 0 1 0 0 1 0 0 0
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
A . K C l
B . C P A
P e rox yn it r it e
P e r o xy n it r it e
N O N O a te
N O N O a te
S N A P
S N A P
W a li a F i g 1

0
2 0
4 0
6 0
8 0
1 0 0
1 2 0F
orce
of c
ontr
actio
n (%
Con
trol)
0
2 0
4 0
6 0
8 0
1 0 0
1 2 0
A . K C l
B . C PA
P e ro x y n SI N -1SN A P N ON O a te
W ith o ut c a ta l a se + S OD
Wi th c a ta l a se + SO D
W a l ia F i g 2

0
20
40
60
80
100
120
Con
trac
tion
(%
Con
trol
)
0
20
40
60
80
100
120A. KCl B. CPA
Control ControlSNAP SNAP+SNAPCPTIO
+SNAPCPTIO
W a lia F ig . 3
0
20
40
60
80
100
120
Con
trac
tion
(%
Con
trol
)
0
20
40
60
80
100
120A. KC l B. C PA
C ontr ol ControlS NAP SN AP+S NAPOD Q
+ SNA PODQ
W a l i a F i g 4

0
2 0
4 0
6 0
8 0
1 0 0
CPA
-indu
ced
incr
ease
in [
Ca2
+] i
(nM
)
C o ntr o l SN A PP e ro x y n
*
W a l i a et a l F i g . 5

0
20
40
60
80
100
120C
a2+ u
ptak
e (%
Con
trol
)
Control Peroxyn SIN-1 SNAP SNP NONOate
Without catalase + SOD
With catal ase + SOD
W a li a Fi g 6




















