
Parasitism, immune response and reproductive success in the house martin Delichonurbica
6
F. de Lope Æ A.P. Møller Æ C. de la Cruz Parasitism, immune response and reproductive success in the house martin Delichon urbica Received: 25 August 1997 / Accepted: 3 November 1997 Abstract Parasites often exert strong selection pressures on their hosts that have evolved anti-parasite defences to counter the negative eects of parasites. We studied the relationship between intensity of parasitism, one aspect of host immune response, and host reproductive success, using the house martin bug Oeciacus hirundinis and its house martin Delichon urbica host as a model system. Experimental manipulation of parasite load of nests during laying of the first clutch altered the intensity of parasitism. Parasites reduced the reproductive success of their hosts measured in terms of body condition and survival of nestlings. Host immune response, measured as the concentration of gammaglobulins and total plas- ma proteins, was positively associated with parasite reproduction, estimated as the number of juvenile par- asites, but was only weakly related to the intensity of adult parasites. The concentration of gammaglobulins was negatively related to nestling body mass, implying a trade-o between immune function and body condition. Parasite reproduction thus exerts a cost on hosts by increasing the immune response. Key words Cost of immune response Æ Gammaglobulins Æ Host-parasite interactions Æ Immunoglobulins Æ Oeciacus hirundinis Introduction Parasites by definition extract resources from their hosts for their own maintenance and reproduction and thereby cause some damage to their hosts (Price 1980). The vir- ulence of parasites varies considerably, presumably de- pending on factors such as the frequency of multiple infections by parasites of dierent genetic strains and the frequency of horizontal transmission among unrelated hosts versus vertical transmission from parents to their ospring (Bull 1994). Numerous studies have recently demonstrated consistent severe eects of parasites on the reproductive success and survival of their hosts (reviews in Lehmann 1993; Møller 1997). Although these negative eects of parasitism are well documented, it is at present not clear to what extent host fitness is reduced by par- asites removing host resources, hosts compensating for parasite removal of host resources, or hosts spending resources on anti-parasite defences. Given the ubiquity and diversity of host-parasite interactions, the diversity of anti-parasite defences by hosts is not surprising. Such defences can be either be- havioural, physiological or immunological. Behavioural defences include avoidance of sites harboring parasites, such as the avoidance of parasitized nests by cli swal- lows Hirundo pyrrhonota (Emlen 1986; Loye and Carroll 1991), and compensation for parasite removal of host resources by increased parental eort (Johnson and Albrecht 1993; Christe et al. 1996). Physiological re- sponses include defences like elevated body temperatures that are detrimental to many micro-parasites (Banet 1986; Blatteis 1986; Kluger 1991). Finally, hosts may use immune responses as a direct weapon designed to deter and eliminate parasite attacks (Klein 1990; Wakelin 1996). The immune system has evolved as a means of distinguishing self from non-self, and thus preventing parasites from having free access to limiting host re- sources. The great level of sophistication of the immune system of hosts is not surprising given the multitude of ways in which a diverse array of parasites are able to exploit their hosts. If host immune responses are ecient ways of man- aging the threats of parasite attacks, we might expect hosts to continuously have elevated responses perpetu- ally on guard for parasite attacks. However, this is not the case. Hosts generally have low circulating titres of Oecologia (1998) 114:188–193 Ó Springer-Verlag 1998 F. de Lope Æ A.P. Møller (&) Æ C. de la Cruz Departamento de Ciencias Morfologicas y Biologia Celular y Animal, Universidad de Extremadura, E-06071 Badajoz, Spain Laboratoire d’Ecologie, CNRS URA 258, Universite´ Pierre et Marie Curie, Baˆt. A, 7e`me e´tage, 7 quai St. Bernard, Case 237, F-75252 Paris Cedex 05, France Fax: (+33) 1 44 27 35 16; e-mail: [email protected]
Transcript of Parasitism, immune response and reproductive success in the house martin Delichonurbica

F. de Lope á A.P. Mùller á C. de la Cruz
Parasitism, immune response and reproductive successin the house martin Delichon urbica
Received: 25 August 1997 /Accepted: 3 November 1997
Abstract Parasites often exert strong selection pressureson their hosts that have evolved anti-parasite defences tocounter the negative e�ects of parasites. We studied therelationship between intensity of parasitism, one aspectof host immune response, and host reproductive success,using the house martin bug Oeciacus hirundinis and itshouse martin Delichon urbica host as a model system.Experimental manipulation of parasite load of nestsduring laying of the ®rst clutch altered the intensity ofparasitism. Parasites reduced the reproductive success oftheir hosts measured in terms of body condition andsurvival of nestlings. Host immune response, measuredas the concentration of gammaglobulins and total plas-ma proteins, was positively associated with parasitereproduction, estimated as the number of juvenile par-asites, but was only weakly related to the intensity ofadult parasites. The concentration of gammaglobulinswas negatively related to nestling body mass, implying atrade-o� between immune function and body condition.Parasite reproduction thus exerts a cost on hosts byincreasing the immune response.
Key words Cost of immune response áGammaglobulins á Host-parasite interactions áImmunoglobulins á Oeciacus hirundinis
Introduction
Parasites by de®nition extract resources from their hostsfor their own maintenance and reproduction and therebycause some damage to their hosts (Price 1980). The vir-
ulence of parasites varies considerably, presumably de-pending on factors such as the frequency of multipleinfections by parasites of di�erent genetic strains and thefrequency of horizontal transmission among unrelatedhosts versus vertical transmission from parents to theiro�spring (Bull 1994). Numerous studies have recentlydemonstrated consistent severe e�ects of parasites on thereproductive success and survival of their hosts (reviewsin Lehmann 1993; Mùller 1997). Although these negativee�ects of parasitism are well documented, it is at presentnot clear to what extent host ®tness is reduced by par-asites removing host resources, hosts compensating forparasite removal of host resources, or hosts spendingresources on anti-parasite defences.
Given the ubiquity and diversity of host-parasiteinteractions, the diversity of anti-parasite defences byhosts is not surprising. Such defences can be either be-havioural, physiological or immunological. Behaviouraldefences include avoidance of sites harboring parasites,such as the avoidance of parasitized nests by cli� swal-lows Hirundo pyrrhonota (Emlen 1986; Loye and Carroll1991), and compensation for parasite removal of hostresources by increased parental e�ort (Johnson andAlbrecht 1993; Christe et al. 1996). Physiological re-sponses include defences like elevated body temperaturesthat are detrimental to many micro-parasites (Banet1986; Blatteis 1986; Kluger 1991). Finally, hosts may useimmune responses as a direct weapon designed to deterand eliminate parasite attacks (Klein 1990; Wakelin1996). The immune system has evolved as a means ofdistinguishing self from non-self, and thus preventingparasites from having free access to limiting host re-sources. The great level of sophistication of the immunesystem of hosts is not surprising given the multitude ofways in which a diverse array of parasites are able toexploit their hosts.
If host immune responses are e�cient ways of man-aging the threats of parasite attacks, we might expecthosts to continuously have elevated responses perpetu-ally on guard for parasite attacks. However, this is notthe case. Hosts generally have low circulating titres of
Oecologia (1998) 114:188±193 Ó Springer-Verlag 1998
F. de Lope á A.P. Mùller (&) á C. de la CruzDepartamento de Ciencias Morfologicas y Biologia Celular yAnimal, Universidad de Extremadura, E-06071 Badajoz, SpainLaboratoire d'Ecologie, CNRS URA 258,Universite Pierre et Marie Curie, Baà t. A, 7eÁ me e tage, 7 quai St.Bernard, Case 237, F-75252 Paris Cedex 05, FranceFax: (+33) 1 44 27 35 16;e-mail: [email protected]

immune substances such as leukocytes, immunoglobu-lins and complement (Klein 1990). The main reason forsuch low levels is the mere magnitude of the immunesystem and the cost of running these sophisticated de-fences at full speed. Evidence of costs of immune func-tion comes from ®eld studies demonstrating a trade-o�between foraging and immune responsiveness (KoÈ nigand Schmid-Hempel 1995) and between investment inexpression of secondary sexual characters and immuneresponsiveness (Saino and Mùller 1996). A selectionexperiment has also demonstrated a trade-o� betweenparasitoid encapsulation e�ciency and ®tness in Dros-ophila melanogaster (Kraaijeveld and Godfray 1997). Weknow very little about the magnitude and the costs ofhost immune responses, and how they are related toparasite loads, under natural conditions. Since resourcesare generally limiting under natural conditions, hostimmune responses may be a severe cost of parasitism.
The aims of this study were to (1) test experimentallyfor costs of parasitism to hosts in terms of reproductivesuccess, and (2) assess the relationship between hostimmune response and parasite virulence measured as thereproductive rate of parasites. If parasites reproduce ata high rate, they should in¯ict more damage on theirhosts and be more costly than parasites that reproduceat a low rate. These aspects of host-parasite interactionswere investigated using the house martin Delichon ur-bica and the house martin bug Oeciacus hirundinis as amodel system. The house martin is a c. 17 g insectivo-rous colonial passerine that feeds on the wing, mainlyon small insects (Cramp 1988). House martins returnfrom Africa to our Spanish study site in February. Amud nest is built by male and female, and nests areusually re-used for subsequent broods. In Badajoz58% of 583 pairs raised two broods and 6% threebroods per season (Pajuelo et al. 1993). The housemartin bug (Hemiptera, Cimicidae) is a generalist ha-ematophagous parasite that has been recorded from alarge number of bird hosts and some mammals as well(Orsza gh et al. 1990). It is particularly common in thenests of hirundines, where it feeds on the blood ofnestlings (Pe ricart 1972). Experimental manipulation ofintensities of house martin bugs revealed strong negativee�ects on several measures of reproductive success suchas number and condition of o�spring (de Lope et al.1993), partly because nestlings have increased metabo-lism in the presence of parasites (Mùller et al. 1994), butpotentially also because of reduced feeding rates byparents having infested nests.
Materials and methods
Study area
The study took place at Badajoz (38°50¢N, 6°59¢W), Spain, wherewe have been conducting studies of the house martin for severalyears. Colonies of house martins occur throughout the city, with atotal population of 3000 pairs (de Lope 1983), and most of ourstudies have used several very large colonies.
Field procedures
Nests were regularly visited from the start of the breeding season(every 2nd day) 1995, and the start of laying, clutch size, brood sizeat hatching and brood size at ¯edging were determined. Secondbroods were followed in a similar way. When nestlings were 15 daysold, their tarsus length was measured with a digital caliper to thenearest 0.01 mm and their body mass recorded on a Pesola springbalance to the nearest 0.5 g. A blood sample of 100 ll was collectedin two capillaries and stored in a cooling bag. Body mass was againrecorded 24 h later.
Experimental manipulations
Nests were randomly assigned to one of three experimental treat-ments:
1. The nest was fumigated with a 0.91% solution of pyrethrinduring the late egg laying period to remove any ectoparasites.Bugs were actually killed by this pesticide, while adult andnestling birds were unharmed (we did not ®nd any dead or sickhouse martins).
2. The nest was fumigated during the late egg-laying period and 2days later inoculated with 10 adult house martin bugs.
3. The nest was fumigated during the late egg-laying period and2 days later inoculated with 100 adult house martin bugs.House martin bugs were obtained from old house martin nestscollected in a nearby colony. The nests were searched forhouse martin bugs which therefore originated from di�erentnests. Bugs were kept in plastic vials at 20°C before inocula-tion.
After second broods had ¯edged, nests were collected andcarefully searched for house martin bugs, that were classi®ed aseither adults or juveniles depending on clear morphological dif-ferences in morphology (Pe ricart 1972).
Estimating immunoglobulins
After blood sampling, capillaries were centrifuged for 15 min at12,500 rpm and the plasma separated from the blood cells. Weestimated the concentration of immunoglobulins and total proteinsin the plasma using an Elphor and Tractoscan from Mercx. Theplasma (50 ll) was subjected to electrophoresis at 120 V, 20 mA for13 min. The gels were subsequently dyed and the concentrationmeasured in a photospectrometer at a wavelength of 525 nm.
Statistical procedures
We determined the e�ect of the experimental manipulation onparasite intensities, host reproductive success, nestling condition,and nestling immune response using analysis of variance. Somevariables were log-transformed in order to obtain approximatelynormally distributed frequency distributions. The relationship be-tween parasite intensity and gamma-globulins was determinedusing linear and multiple linear regression analysis. Values reportedare means (SE). The level of signi®cance is 5% and two-tailed testsare used throughout.
Results
E�ects of parasites on host reproductive success
Experimental manipulation of parasite intensity suc-cessfully a�ected the number of parasites found in nests
189

at the end of the breeding season (Table 1). While therewere on average 16 house martins bugs in fumigatednests, there were 73 in nests inoculated with 10 bugs atthe start of the breeding season, and 178 in nests in-oculated with 100 bugs at the start of the breedingseason. The prevalence of parasites was 100% in alltreatments (Table 1). The abundance of both juvenileand adult life stages of house martin bugs was similarlya�ected by the manipulation (Table 1). The abundanceof adult and juvenile house martin bugs was stronglypositively correlated (Pearson r � 0.82, n � 93, P <0.001).
We found no e�ect of parasite treatment on repro-ductive success measured as total production of ¯edg-lings (Table 1). However, ¯edging success was lower inthe two treatment groups with parasite inoculation,although only signi®cantly so in the second brood(Table 1). The same pattern applied to body mass, withnestlings being signi®cantly lighter in nests with manyparasites added (Table 1). Hirundine nestlings decreasein body mass during the last part of their nestling periodmainly due to water loss (e.g. Ricklefs 1968), and massreduction by nestlings over 1 day was also larger inparasite-inoculated nests (Table 1). There was no e�ectof parasites on nestling tarsus length (Table 1), which is ameasure of skeletal body size. Hence nestling body masscorrected for structural body size, which is a measure ofbody condition, from parasite-inoculated nests was lowerthan that of nestlings from the fumigation group.
Immune responses of nestlings
We measured the immune response of nestlings from theconcentration of gammaglobulins in the plasma. Therewere signi®cantly more gammaglobulins in the bloodof nestlings from parasite-inoculated nests than fromfumigated nests (Table 2). The total amount of bloodproteins showed a similar pattern (Table 2). The pro-portion of gammaglobulins was similar across treatmentgroups (Table 2).
Nestlings di�ered in condition between ®rst and sec-ond broods. While tarsus length was similar in the twobroods (paired t-test, t � 0.50, df � 17, P � 0.63),body mass was signi®cantly depressed in second broods[®rst brood: 19.1 g (0.5); second brood: 16.5 g (0.7),paired t-test, t � 3.05, df � 17, P � 0.007]. The con-centration of gammaglobulins was larger in second thanin ®rst brood nestlings [®rst brood: 0.27 (0.03); secondbrood: 0.32 (0.03), paired t-test, t � 2.05, df � 17,P � 0.03]. There was no signi®cant di�erence in totalprotein concentration between broods [®rst brood: 5.69(0.34); second brood: 5.54 (0.30), paired t-test, t � 0.31,df � 17, P � 0.76], and therefore a di�erence in thepercentage of gammaglobulins between broods [®rstbrood: 4.6% (0.4); second brood: 5.9% (0.6), pairedt-test, t � 1.98, df � 17, P � 0.047]. Hence nestlingswere in poorer condition and had stronger immune re-sponses in the second as compared with the ®rst brood.There was no signi®cant di�erence in the number of
Table 1 Parasite load and reproductive parameters of house martins in relation to experimental treatment. ANOVAs were based on log-transformed parasite loads. Values are means (SE). Mean values with di�erent superscripts di�er signi®cantly in Sche�e a posteriori tests
Treatment F df P
Fumigated 10 Bugs added 100 Bugs added
Adult house martin bugs 8.93 (1.29)a 24.72 (2.55)b 91.15 (8.17)c 203.65 2,91 0.0001Juvenile house martin bugs 7.40 (1.53)a 47.94 (11.02)b 87.07 (9.19)c 47.93 2,91 0.0001Total house martin bugs 16.33 (2.19)a 72.67 (16.32)b 178.22 (14.64)c 128.80 2,91 0.0001No. of ¯edglings 4.54 (0.43) 4.67 (0.49) 4.33 (0.45) 0.11 2,91 0.90Fledging success (%)1st brood 95.8 (4.2) 71.2 (6.7) 74.3 (7.3) 2.21 2,91 0.132nd brood 58.3 (14.4)a 20.8 (6.7)b 30.4 (6.9)b 3.28 2,26 0.05
Body mass (g) 19.3 (0.4)a 17.2 (0.8)b 17.8 (0.4)b 3.98 2,91 0.022Change in mass (g d)1) )0.76 (0.07)a )0.87 (0.10)ab )1.06 (0.06)b 5.82 2,91 0.004Tarsus length (mm) 10.69 (0.49) 10.48 (0.64) 10.65 (0.43) 1.04 2,91 0.36n 30 28 36
Table 2 Immune response of house martin nestlings in relation to experimental parasite treatment. Values are means (SE). Mean valueswith di�erent superscripts di�er signi®cantly in Sche�e a posteriori tests
Treatment F df P
Fumigated 10 Bugs added 100 Bugs added
Gammaglobulins 0.249 (0.02)a 0.268 (0.044)ab 0.325 (0.021)b 6.92 2,91 0.01Total proteins 5.05 (0.30)a 5.19 (0.23)ab 5.94 (0.19)b 11.03 2,91 0.0001% Gammaglobulins 5.2 (0.5) 5.1 (0.8) 5.5 (0.4) 0.14 2,91 0.87n 30 28 36
190
house martin bugs in nests with only a ®rst brood andnests with ®rst and second broods (log-transformed in-tensities of adult, juvenile and total parasites, t � 1.19±1.66, df � 92, P � 0.10±0.24).
If immune function is costly, we should expect to ®nda negative relationship between the concentration ofimmunoglobulins and nestling growth, for example es-timated as body mass or body condition. The concen-tration of gammaglobulins was negatively related tonestling body mass [F � 3.47, df � 1,92, r2 � 0.05,P � 0.037; slope (SE) � )0.72 (0.22)], and also to thechange in nestling body mass from one day to another[F � 13.40, df � 1,92, r2 � 0.13, P < 0.001; slope(SE) � )0.98 (0.27)]. Hence immune function appearsto be traded against nestling body mass.
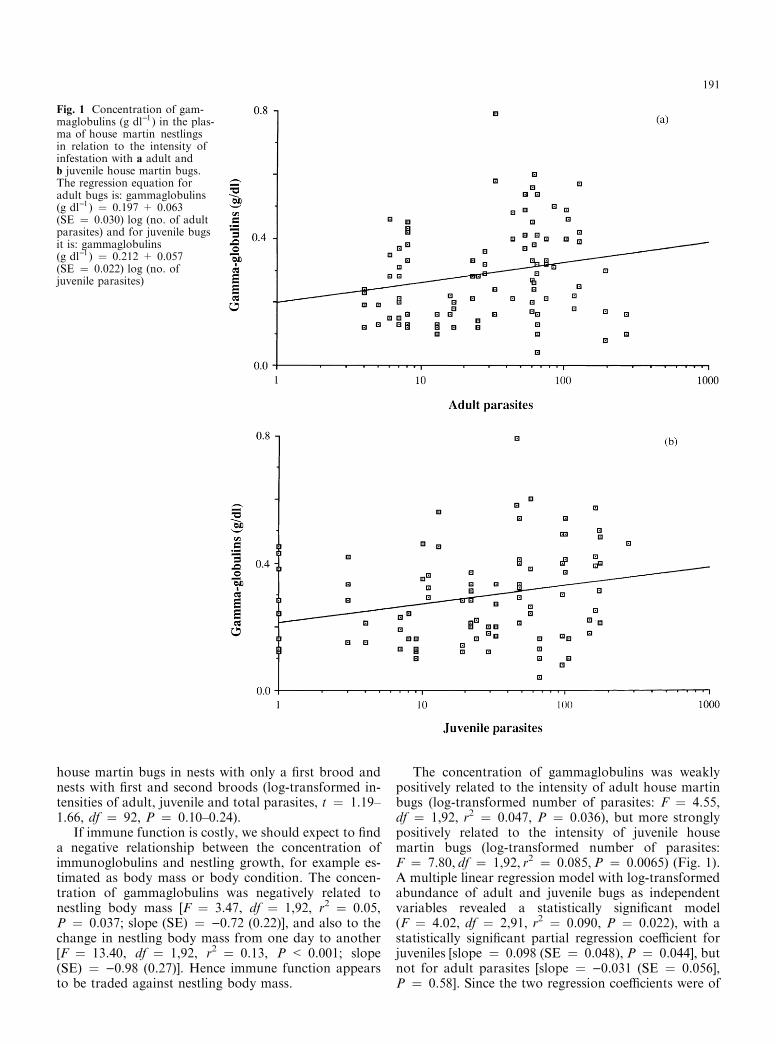
The concentration of gammaglobulins was weaklypositively related to the intensity of adult house martinbugs (log-transformed number of parasites: F � 4.55,df � 1,92, r2 � 0.047, P � 0.036), but more stronglypositively related to the intensity of juvenile housemartin bugs (log-transformed number of parasites:F � 7.80, df � 1,92, r2 � 0.085,P � 0.0065) (Fig. 1).A multiple linear regression model with log-transformedabundance of adult and juvenile bugs as independentvariables revealed a statistically signi®cant model(F � 4.02, df � 2,91, r2 � 0.090, P � 0.022), with astatistically signi®cant partial regression coe�cient forjuveniles [slope � 0.098 (SE � 0.048), P � 0.044], butnot for adult parasites [slope � )0.031 (SE � 0.056],P � 0.58]. Since the two regression coe�cients were of
Fig. 1 Concentration of gam-maglobulins (g dl)l ) in the plas-ma of house martin nestlingsin relation to the intensity ofinfestation with a adult andb juvenile house martin bugs.The regression equation foradult bugs is: gammaglobulins(g dl)l ) � 0.197 + 0.063(SE � 0.030) log (no. of adultparasites) and for juvenile bugsit is: gammaglobulins(g dl)l ) � 0.212 + 0.057(SE � 0.022) log (no. ofjuvenile parasites)
191

opposite signs, they also di�ered signi®cantly from eachother. The proportion of total protein being gamma-globulins did not increase with the number of adultparasites (F � 0.46, df � 1,92, r2 � 0.005, P � 0.50),but increased signi®cantly with the intensity of juvenilebugs (F � 4.75, df � 1,92, r2 � 0.046, P � 0.032).The relative number of juvenile parasites calculated asthe residuals from a regression of log-transformed num-ber of juvenile parasites on log-transformed adult para-sites (F � 168.82, df � 1,92, r2 � 0.67, P < 0.001)was signi®cantly positively related to the concentrationof gammaglobulins (F � 4.07, df � 1,92, r2 � 0.046,P � 0.047).
Discussion
Experimental manipulation of the intensity of infesta-tion of house martin nests with the house martin bugresulted in a reduction in ¯edging success, but also bodycondition of nestlings (Table 1). Large parasite loadsdepressed nestling body mass while e�ects on tarsuslength were much less obvious (Table 1). Previousstudies of the house martin bug have revealed signi®cantreductions in brood size and tarsus length, but also anincrease in the interval between ®rst and second broodsand thus in the timing of reproduction (de Lope et al.1993; Mùller et al. 1994). The e�ects of parasites on hostreproductive success are similar to those reported for alarge number of other avian hosts with potentially highfrequencies of multiple infestations of nests and frequenthorizontal transmission of parasites (Mùller and Err-itzùe 1996; Mùller 1997). The exact mechanisms gener-ating the negative e�ects of parasites on host ®tness arenot well understood. House martin bugs remove bloodfrom their hosts, although the total amount removed isrelatively small compared to that available. If bloodaccounts for 10% of the body mass of nestlings, anaverage ®rst brood of 3.47 nestlings should have 6.63 gblood which constitutes an enormous amount even forthe average number of 178 bugs present in nests inocu-lated with 100 parasites. Other mechanisms are thereforeclearly needed to explain the negative e�ects of parasiteson host reproductive success.
Given the mere size of the immune system (Klein1990), we may consider the possibility that hosts payindirect costs of parasitism through the activity ofimmune defence. There is some evidence for a cost ofimmune function as demonstrated by negative rela-tionships between immune responses and host activity inthe bumble bee Bombus terrestris and sexual display inthe barn swallow Hirundo rustica (KoÈ nig and Schmid-Hempel 1995; Saino and Mùller 1996), and a negativerelationship between parasitoid encapsulation e�ciencyand ®tness in Drosophila melanogaster (Kraaijeveld andGodfray 1997). Virtually nothing is known about thee�ects of parasites on host immune function under ®eldconditions. The study of house martin bugs demon-
strated two dimensions of costs of immune responses.First, we found a positive relationship between parasitereproduction measured as the number of juvenile bugsand concentration of gammaglobulins. Immunoglobu-lins act as cell surface receptors produced in excessivequantities and released into the circulation. They rec-ognize and combine with parasite antigens and thus playan important role in host immunity (Wakelin 1996).Parasite virulence can be de®ned as the rate of repro-duction by parasites (Bull 1994), since a high rate impliesremoval of large amounts of host resources and thusreductions in host ®tness. Although the intensity ofparasitism by both adult and juvenile bugs was associ-ated with the concentration of gammaglobulins, thee�ect of juveniles was stronger than that of adult para-sites. A multiple regression analysis revealed a statisti-cally signi®cant regression coe�cient only for juveniles,but not for adult, parasites. Hence hosts seem to pay acost of increased parasite virulence not only in terms ofresources removed for parasite reproduction, but also interms of increased immune responses caused by thelarger number of parasites.
Second, we found a negative relationship between theconcentration of gammaglobulins and nestling bodymass and change in body mass, respectively. Since housemartin bugs had a negative impact on nestling bodymass (Table 1), the negative correlation between gam-maglobulins and nestling body mass implies that nes-tlings trade body mass against immune responses.Hirundine nestlings reach a peak in body mass duringthe late half of the nestling period, with a subsequentdecrease mainly due to water loss (Ricklefs 1968). Nes-tlings being raised in parasitized nests are generallylighter than nestlings from parasite-free nests (review inMùller 1997), and the probability of recruitment isgenerally positively related to nestling body mass in hi-rundines (Mùller 1994) and in birds in general (Linde net al. 1992). Therefore, the negative correlation betweengammaglobulins and nestling body mass under experi-mentally manipulated parasite loads implies a trade-o�between immune function and an important componentof ®tness. Defences against parasites thus do not comefor free, and hosts must therefore optimize their immuneresponses relative to the magnitude of the damagein¯icted by parasites.
Di�erent components of the immune system varyinterspeci®cally, and the determinants of such variationare unknown. However, it is possible that parasites thatmainly attack nestlings, such as the house martin bug,particularly select for investment in components of theimmune system that are active at a very early age, whileother kinds of parasites in the house martin (or in otherhosts) may select for the main investment in immunefunction taking place at a di�erent time of the life cycle.Future comparative and/or experimental studies may beable to determine the relationship between the ontogenyand the size of the immune system in relation to timingof parasitism.
192

Acknowledgements F. de Sande kindly made the immunoglobulinanalyses and L. Sanabria helped with the ®eld work. S. Merinokindly provided constructive comments. The research was sup-ported by grants from the Danish Natural Science ResearchCouncil to A.P.M. and DGICYT (PB-95-0020) to F. de L. and C.de la C.
References
Banet M (1986) Fever in mammals: is it bene®cial? Yale J Biol Med59:117±124
Blatteis CM (1986) Fever. Is it bene®cial? Yale J Biol Med 59:107±116
Bull JJ (1994) Perspective: virulence. Evolution 48:1423±1437Christe P, Richner H, Oppliger A (1996) Begging, food provi-
sioning and nestling competition in great tit broods infestedwith ectoparasites. Behav Ecol 7:127±131
Cramp S (ed) (1988) The birds of the Western Palearctic, vol 5.Oxford University Press, Oxford
Emlen JT (1986) Responses of breeding cli� swallows to nidicolousparasite infestations. Condor 88:110±111
Johnson LS, Albrecht DJ (1993) E�ects of haematophagous ecto-parasites on nestling house wrens, Troglodytes aedon: who paysthe cost of parasitism? Oikos 66:255±262
Klein J (1990) Immunology. Blackwell, OxfordKluger M (1991) The adaptive value of fever. In: Mackowiak P (ed)
Fever: basic mechanisms and management. Raven, New York,pp 105±124
KoÈ nig C, Schmid-Hempel P (1995) Foraging activity and immu-nocompetence in workers of the bumblebee, Bombus terrestrisL. Proc R Soc Lond B 260:225±227
Kraaijeveld AR, Godfray HCJ (1997) Trade-o� between parasitoidresistance and larval competitive ability in Drosophila melano-gaster. Nature 389:278±280
Lehmann T (1993) Ectoparasites: Direct impact on host ®tness.Parasitol Today 9:8±13
Linde n M, Gustafsson L, PaÈ rt T (1992) Selection on ¯edging massin the collared ¯ycatcher and the great tit. Ecology 73:336±343
Lope F de (1983) La avifauna de las Vegas Bajas del Guadiana.Do~nana Acta Vert 10:91±121
Lope de F, Gonza lez G, Pe rez JJ, Mùller AP (1993) Increaseddetrimental e�ects of ectoparasites on their bird hosts duringadverse environmental conditions. Oecologia 95:234±240
Loye JE, Carroll SP (1991) Nest ectoparasite abundance and cli�swallow colony site selection, nestling development, and de-parture time. In: Loye JE, Zuk M (eds) Bird-parasite interac-tions: ecology, evolution, and behavior. Oxford UniversityPress, New York, pp 222±241
Mùller AP (1994) Sexual selection and the barn swallow. OxfordUniversity Press, Oxford
Mùller AP (1997) Parasites and the evolution of host life history.In: Clayton D, Moore J (eds) Host-parasite evolution: generalprinciples and avian models. Oxford University Press, Oxford,pp 105±127
Mùller AP, Erritzùe J (1996) Parasite virulence and host immunedefense: host immune response is related to nest re-use in birds.Evolution 50:2066±2072
Mùller AP, Lope F de, Moreno J, Gonza lez G, Pe rez JJ (1994)Ectoparasites and host energetics: house martin bugs and housemartin nestlings. Oecologia 98:263±268
Orsza gh I, Krumpal MV, Cyprich D (1990) Contribution to theknowledge of the martin bug Oeciacus hirundinis (Heteroptera,Cimicidae) in Czechoslovakia. Zb Slov Na r Mu z Prir Vedy36:43±60
Pajuelo L, Lope F, de Silva da E (1993) Biologõ a de la reproduccio ndel avio n comu n (Delichon urbica) en Badajoz, EspanÄ a. Ardeola39:15±23
Pe ricart J (1972) He mipteres: Anthocoricidae, Cimicidae, Micro-physidae de l'Ouest-Pale arctique. Masson, Paris
Price PW (1980) Evolutionary biology of parasites. PrincetonUniversity Press, Princeton
Ricklefs RE (1968) Weight recession in birds. Auk 85:30±35Saino N, Mùller AP (1996) Sexual ornamentation and immuno-
competence in the barn swallow. Behav Ecol 7:227±232Wakelin D (1996) Immunity to parasites, 2nd edn. Cambridge
University Press, Cambridge
193




















