
Diversity of ‘benzenetriol dioxygenase’ involved in p -nitrophenol degradation in soil bacteria
Upload
srikrishnadevarayaCategory
view
1download
0
Environmental Toxicology and Chemistry, Vol. 31, No. 9, pp. 1980–1988, 2012# 2012 SETAC
Printed in the USADOI: 10.1002/etc.1931
p-NITROPHENOL TOXICITY TO AND ITS REMOVAL BY THREE SELECT SOIL
ISOLATES OF MICROALGAE: THE ROLE OF ANTIOXIDANTSSURESH R. SUBASHCHANDRABOSE,y MALLAVARAPU MEGHARAJ,*y KADIYALA VENKATESWARLU,z and RAVI NAIDUyyCentre for Environmental Risk Assessment and Remediation, University of South Australia, Mawson Lakes, and Cooperative Research Centre for
Contamination Assessment and Remediation of Environment, Salisbury South, Australia
zDepartment of Microbiology, Vikrama Simhapuri University, Nellore, India
(Submitted 25 March 2012; Returned for Revision 3 May 2012; Accepted 28 May 2012)
* T(megha
Pub(wileyo
Abstract—The nontarget effects, in terms of biochemical changes induced by p-nitrophenol (PNP) in three soil microalgae, Chlorellasp., Chlorococcum sp., and Heterochlamydomonas sp., and the PNP removal efficiency of these isolates, were determined. On exposureto 20mgL�1 PNP, Chlorella sp. showed greater activity of peroxidase, superoxide dismutase, and glutathione reductase as well as highcontents of proline and carotenoids. While Heterochlamydomonas sp. exhibited higher levels of catalase and protein, Chlorococcum sp.produced greater amounts of malondialdehyde, a measure of lipid peroxidation, in the presence of PNP. Chlorella sp. tolerated PNPby producing large quantities of antioxidants coupled with less lipid peroxidation, whileChlorococcum sp. was susceptible, as evidencedby low antioxidant production and high lipid peroxidation. During 7-d exposure, Chlorella sp., Heterochlamydomonas sp., andChlorococcum sp. were able to remove 39, 18, and 4% of 20mgL�1 PNP, respectively. The present results indicate that proline,carotenoids and malondialdehyde are the potential biomarkers for assessing PNP toxicity toward microalgae, and their response could beconsidered for differentiating tolerant and susceptible strains. Moreover, there is a clear correlation between PNP removal andantioxidant synthesis in microalgae on exposure to the pollutant. Environ. Toxicol. Chem. 2012;31:1980–1988. # 2012 SETAC
Keywords—Microalgae Antioxidant p-Nitrophenol Biochemical change Oxidative damage
INTRODUCTION
p-Nitrophenol (PNP), a slightly yellowish, crystalline, solid,nitroaromatic phenolic compound, is widely used in the syn-thesis of pesticides, drugs, and dyes [1]. Approximately 84% ofPNP produced is used for the synthesis of organophosphorouspesticides (ethyl and methyl parathions) and 8% for the syn-thesis of acetaminophen (paracetamol), while the remaining 8%is used in leather processing, dyestuffs manufacturing, and soon (National Center for Biotechnology Information, PubChemCompound Database; http://pubchem.ncbi.nlm.nih.gov/sum-mary/summary.cgi?cid¼980#x321). p-Nitrophenol is formedin soil and aquatic systems as a hydrolytic product from lesspersistent organophosphate insecticides, parathion, and methylparathion [2]. Several enzymatic assays involving PNP-basedsubstrates also release this mononitrophenol [3]. Exposure ofgeneral populations to PNP could occur by drinking PNP-contaminated water or through inhalation of air containing thischemical. It was found to cause several health-related problemsin humans such as methemoglobinemia and may also causemutation [1]. Acute exposure leads to liver and kidney damageas well as systemic poisoning [4]. In humans, parathion inges-tion and its toxic kinetics are monitored by the release ofPNP through urine [5]. Therefore, PNP is considered one ofthe 126 priority pollutants by the U.S. Environmental ProtectionAgency (U.S. EPA; Priority Pollutants; http://water.epa.gov/scitech/methods/cwa/pollutants.cfm).
Microalgae are ubiquitous in the soil and water environmentand very frequently exposed to toxic substances released fromindustrial and domestic wastes. As microalgae are at the bottom
o whom correspondence may be [email protected]).lished online 3 July 2012 in Wiley Online Librarynlinelibrary.com).
1980
of the ecological pyramid, any adverse effect on these beneficialorganisms will be carried over to other levels. Because PNPposes a significant threat to both soil and freshwater microalgae,its toxic effects on microalgae have been studied extensively[6,7]. A few studies have investigated the efficiency of micro-algae at removing PNP [8]. Although it is difficult for manymicroalgae to grow in the presence of PNP, some strains aretolerant and efficient at removing PNP. It is essential to studythe factors for efficient degradation of PNP by microalgae. Thefirst step toward this goal is to understand the toxic nature ofPNP and the way through which it affects microalgae byaltering the levels of biochemicals and antioxidant systems.In fact, biochemical assays offer great advantages over othermolecular techniques as they give instantaneous results andmost of the assays require simple analytical equipment and onlymodest skill and experience. However, information about thebiochemical and molecular changes induced in microalgae bypollutants is very limited [9].
Biochemicals such as antioxidants are produced in excessfor scavenging free radicals produced during biotic and abioticstress [10]. Numerous studies have established that stress-related changes in the antioxidant parameters of microalgaeare induced by ultraviolet (UV) radiation [11], heavy metals[12], and organic compounds like pyrene [13]. However, infor-mation on the biochemical response of microalgae to PNPexposure is very limited [7]. In the present study, the impactof different concentrations of PNP for varying exposure-timeintervals was assessed on several antioxidant parameters suchas peroxidase (POX), catalase (CAT), superoxide dismutase(SOD), glutathione reductase (GR), lipid peroxidation (LPO),proline, carotenoids, as well as total proteins in three soilisolates of microalgae, Chlorella sp., Chlorococcum sp., andHeterochlamydomonas sp. The final objective was to under-stand the relationship between the antioxidant parameters andPNP removal by microalgae.

p-Nitrophenol toxicity to microalgae Environ. Toxicol. Chem. 31, 2012 1981
MATERIALS AND METHODS
Microalgal cultures
Axenic cultures of green unicellular microalgae, Chlorellasp., Chlorococcum sp., and Heterochlamydomonas sp., isolatedfrom soil were used for the present study. Cultures werechecked for contamination from time to time by microscopicanalysis and by streaking on Bold’s basal medium (BBM)and nutrient agar plates. All three isolates are deposited atthe Phycology Laboratory, Centre for Environmental RiskAssessment and Remediation of the Environment (CERAR),University of South Australia. Stock cultures were maintainedin 250-ml flasks containing BBM kept under continuous coolfluorescent light at 24� 28C in an orbital shaker set at 120 rpmas described by Megharaj et al. [14].
Experimental design
For a particular biochemical parameter involving all threemicroalgal species, a total of 75 sterile 40-ml Agilent tubesfilled with 10ml of autoclaved BBM were used. p-Nitrophenolwas added to the medium at final concentrations of 5, 10, and20mgL�1. For inoculation, 7-d-old exponentially growing cellswere used. The initial cell density of the culture was maintainedat 6� 105 cells ml�1. Cultures were incubated under the sameconditions as mentioned above. The growth of algal cellswas monitored microscopically by direct counting using ahemocytometer. Sampling for determination of levels of PNPdegradation and other biochemical parameters was done after24 and 96 h of incubation. Total soluble protein and totalcarotenoids were quantitated only after 96 h. Cell growth andPNP removal were analyzed in cells exposed to 20mgL�1 PNPafter 1, 4, and 7 d of incubation. For all of the biochemicalparameters, the cell density maintained was approximately thesame.
Preparation of cell-free enzyme extract
The entire 10ml of culture was centrifuged at 4,600 g for15min, and the cell pellet was washed twice with sterileultrapure water. The pellet was resuspended in the respectiveenzyme assay buffer and lysed using an ultrasonicator (Bransondigital sonifier). Cell lysates were then centrifuged at 9,167 g for5min, and the supernatant was transferred to fresh microfugetubes and stored on ice or at 48C until the enzyme assays weredone.
Peroxidase
Total POX activity was assayed according to the method ofChance and Maehly [15]. An aliquot of 100ml of cell extractwas added to 3ml of assay mixture containing 14mM potas-sium phosphate buffer (pH 7.0), 0.027% H2O2, and 0.5%pyrogallol. Change in absorbance at 420 nm was monitoredin a spectrophotometer every 20 s for 5min. Activity of POXwas expressed as units per milligram protein.
Catalase
Activity of CAT was measured following the method ofBeers and Sizer [16]. The reaction mixture was comprised of50mM potassium phosphate buffer (pH 7.0), 0.035% H2O2,and 100ml of cell extract. The time required for a decreasein absorbance from 0.45 to 0.40 was read at 240 nm. Activity ofCAT was expressed as units per milligram protein.
Glutathione reductase
Activity of GR was measured according to the method ofMavis and Stellwagen [17]. The assay mixture contained75mM potassium phosphate, 2.6mM EDTA, 1mM gluta-thione, 0.09mM reduced b-nicotinamide adenine dinucleotidephosphate, and 0.13% bovine serum albumin mixed with 100mlof crude enzyme extract. The decrease in absorbance was readfor 5min. Activity of GR was expressed as units per milligramprotein.
Superoxide dismutase
The enzyme SOD in microalgae was assayed per the methodof McCord and Fridovich [18]. The ability of superoxide radicalto inhibit the reduction of cytochrome cwas used for measuringSOD activity. The reaction mixture contained 50mM of potas-sium phosphate buffer (pH 7.8), 0.1mM EDTA, 0.01mMcytochrome c, 0.05mM xanthine, and 0.005U of xanthineoxidase mixed with the crude enzyme extract. The increasein absorbance at 550 nm was recorded for 5min. Activity ofSOD was expressed in units per milligram protein.
Lipid peroxidation
An aliquot (10ml) of algal cultures was pelleted by cen-trifugation and washed with sterile ultrapure water. Cells weresuspended in 0.1% trichloroacetic acid and lysed by ultra-sonication. Malondialdehyde (MDA) content, as a measureof LPO, was estimated based on the method described by Heathand Packer [19]. The final concentration of MDA was deter-mined by means of the extinction coefficient 155 mM�1 cm�1
and expressed in nanomoles per milligram protein.
Proline determination
Microalgal cells pelleted from 10-ml cultures by centrifu-gation were washed with sterile ultrapure water, suspended in3% sulfosalicylic acid, and lysed by ultrasonication. The lysatewas centrifuged and 2ml of clear supernatant was used for thedetermination of proline content as described by Bates et al.[20].
Total soluble protein
Washed cells of microalgae from 10ml of cultures weresuspended in 50mM Tris (pH 7.50) containing 0.5mM phenyl-methylsulfonyl fluoride and lysed in an ultrasonicator. Aftercentrifugation, the clear supernatant containing soluble proteinwas used for protein analysis following the method of Bradford[21].
Total carotenoids
Total carotenoids were extracted from 10ml of microalgalcultures using methanol as the solvent and estimated per themethod of Wellburn [22]. Total carotenoids were calculatedfollowing the formula Cxþc¼ (1,000A470 – 1.63Ca – 104.96Cb)/221, where Cxþc is total carotenoids, Ca is chlorophyll a, and Cb
is chlorophyll b (all in micrograms per milliliter fresh wt).
p-Nitrophenol estimation
One milliliter of culture was centrifuged and the supernatantfiltered through a 0.45-mm filter and directly used for analysisusing an HPLC system (Agilent Technologies 1200 series)equipped with a Zorbax eclipse column XDB-C18, particlesize 5mm, 4.6� 150mm. A sample of 10ml was separated inthe column by methanol and water as mobile phase in a gradientprogram, starting with the mobile phase of 20% methanol

Fig. 1. Changes in catalase (CAT) activity in microalgae exposed top-nitrophenol (PNP). Data are means�SD (n¼ 3). Letters on top of the barsfor each sampling indicate significant difference between different PNPconcentrations at p� 0.005 based on Tukey’s test. (a) Chlorella sp., (b)Chlorococcum sp., and (c) Heterochlamydomonas sp.
1982 Environ. Toxicol. Chem. 31, 2012 S.R. Subashchandrabose et al.
initially for 10min followed by 90% methanol for the next10min and a 5-min postrun. Detection of PNP was based onits UV absorbance at 254 nm, and the amount remaining in theculture medium was calculated using a calibration graph.
Statistical analyses
When necessary, the data were log-transferred to meet theanalysis of variance (ANOVA) assumptions of normality. Allbiochemical assays were carried out in triplicate of a singleexperiment, and data were expressed as mean values� standarddeviation. One-way ANOVA was carried out for significantdifferences, if any, in the biochemical parameters among themicroalgae grown with and without 20mgL�1 PNP. Tukey’smultiple comparison tests were used to group the microalgalspecies based on the level of significance at p� 0.05. One-wayANOVA was also used to test the level of significant variationbetween various biochemicals produced by the microalgae atdifferent PNP concentrations. A multifactorial analysis wasdone to test the effect of interactions between the biochemicalsand the variables such as PNP concentration, exposure time,and microalgal species. Total protein and carotenoids wereexcluded from the analysis as they were measured only after96 h of incubation. Principal component analysis (PCA) wasdone to analyze the relationship between different biochem-icals, which were grouped based on the significant relation-ship. A correlation coefficient matrix was constructed usingPearson’s product moment correlation to study the significantcorrelation among the biochemicals chosen for the study.All statistical analyses were carried out using Minitab 16.1.1statistical software and PASW statistics 18.
RESULTS
Effect of PNP on the enzymatic antioxidant system
All three microalgae chosen for the study responded differ-entially to different PNP concentrations as well as times ofexposure. In most cases, antioxidant enzymes produced by thealgae grown in PNP were significantly higher compared withthose from algae grown without PNP. This clearly indicates thatPNP induced oxidative stress in microalgae. A nearly onefoldrise in CAT activity was observed in Chlorella sp. exposed to20mgL�1 PNP over untreated control (Fig. 1A). Interestingly,Chlorococcum sp. exposed to 10mgL�1 PNP showed higherCAT activity compared to those exposed to 20mgL�1 PNP(Fig. 1B). Levels of CAT showed a time- and dose-specificincrement in Heterochlamydomonas sp. on exposure to PNP(Fig. 1C). An inhibitory trend for GR was noticed in Chlorellasp. and Chlorococum sp. when exposed to 10 or 20mgL�1 PNP(Fig. 2A and B). Indeed, there was no significant difference inthe activity of GR when the cells of Chlorella sp. were grown inthe presence of 10 or 20mgL�1. However, Heterochlamydo-monas sp. showed a time-specific (Table 1) and dose-specific(Fig. 2C) increase in GR activity.
A time- and dose-dependent increase in POX activity wasobserved inChlorella sp.When the cells were grown even in thepresence of 20mgL�1 PNP, the activity of POX was nearlydoubled compared to cells not exposed to PNP (Fig. 3A andTable 1). Although 20mgL�1 PNP-grown Chlorococcum sp.showed a decrease in POX activity at the end of 96 h, theenzyme activity in Heterochlamydomonas sp. was not affectedat concentrations of 10 or 20mgL�1 PNP during the corre-sponding period of incubation (Fig. 3B and C). No significantdifference in POX activity was observed between untreatedcontrol and 20mgL�1 PNP-grown Heterochlamydomonas sp.
However, in Chlorella sp. there was a considerable increase inPOX levels on exposure to 5mgL�1 PNP, while the enzymeactivity was not significantly increased by exposure to 10 or20mgL�1 PNP after 24 h. The response of SOD activity toPNP treatments (Fig. 4) was a reversal of that of POX activity.OnlyChlorococum sp. andHeterochlamydomonas sp. showed atime- and dose-specific increase in SOD activity.
Effect of PNP on proline, LPO, protein, and carotenoids
Remarkably, proline content showed a time- and dose-specific increase in all the microalgal species used in the study.A significant difference in proline content was observedbetween most of the treatments (Fig. 5). Cells of Chlorellasp. exposed to 20mgL�1 PNP for 96 h showed nearly a fivefoldincrease in proline compared to those grown in the absence ofPNP. A similar trend was also noticed inHeterochlamydomonassp. Overall, the proline content was higher in Chlorella sp.,followed by Heterochlamydomonas sp. and Chlorococcum sp.(Table 2).
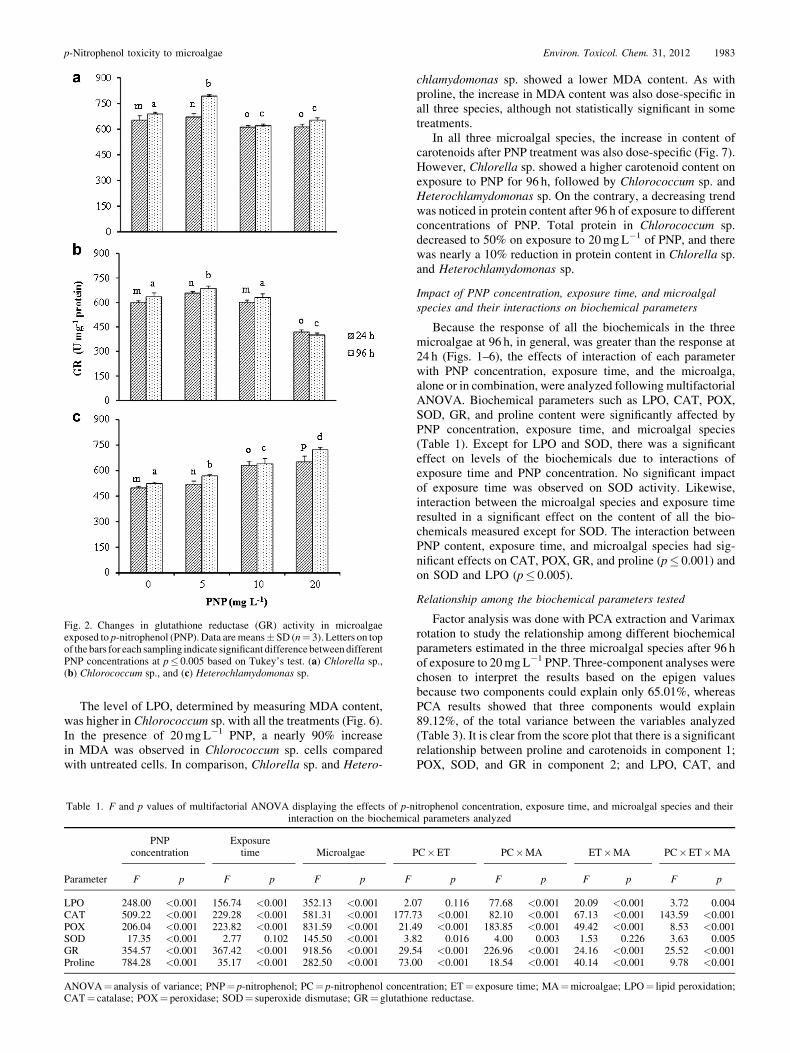
Fig. 2. Changes in glutathione reductase (GR) activity in microalgaeexposed to p-nitrophenol (PNP). Data aremeans� SD (n¼ 3). Letters on topof the bars for each sampling indicate significant difference betweendifferentPNP concentrations at p� 0.005 based on Tukey’s test. (a) Chlorella sp.,(b) Chlorococcum sp., and (c) Heterochlamydomonas sp.
p-Nitrophenol toxicity to microalgae Environ. Toxicol. Chem. 31, 2012 1983
The level of LPO, determined by measuring MDA content,was higher in Chlorococcum sp. with all the treatments (Fig. 6).In the presence of 20mgL�1 PNP, a nearly 90% increasein MDA was observed in Chlorococcum sp. cells comparedwith untreated cells. In comparison, Chlorella sp. and Hetero-
Table 1. F and p values of multifactorial ANOVA displaying the effects of p-ninteraction on the biochemica
Parameter
PNPconcentration
Exposuretime Microalgae
F p F p F p F
LPO 248.00 <0.001 156.74 <0.001 352.13 <0.001 2.0CAT 509.22 <0.001 229.28 <0.001 581.31 <0.001 177.7POX 206.04 <0.001 223.82 <0.001 831.59 <0.001 21.4SOD 17.35 <0.001 2.77 0.102 145.50 <0.001 3.8GR 354.57 <0.001 367.42 <0.001 918.56 <0.001 29.5Proline 784.28 <0.001 35.17 <0.001 282.50 <0.001 73.0
ANOVA¼ analysis of variance; PNP¼ p-nitrophenol; PC¼ p-nitrophenol concenCAT¼ catalase; POX¼ peroxidase; SOD¼ superoxide dismutase; GR¼ glutathi
chlamydomonas sp. showed a lower MDA content. As withproline, the increase in MDA content was also dose-specific inall three species, although not statistically significant in sometreatments.
In all three microalgal species, the increase in content ofcarotenoids after PNP treatment was also dose-specific (Fig. 7).However, Chlorella sp. showed a higher carotenoid content onexposure to PNP for 96 h, followed by Chlorococcum sp. andHeterochlamydomonas sp. On the contrary, a decreasing trendwas noticed in protein content after 96 h of exposure to differentconcentrations of PNP. Total protein in Chlorococcum sp.decreased to 50% on exposure to 20mgL�1 of PNP, and therewas nearly a 10% reduction in protein content in Chlorella sp.and Heterochlamydomonas sp.
Impact of PNP concentration, exposure time, and microalgalspecies and their interactions on biochemical parameters
Because the response of all the biochemicals in the threemicroalgae at 96 h, in general, was greater than the response at24 h (Figs. 1–6), the effects of interaction of each parameterwith PNP concentration, exposure time, and the microalga,alone or in combination, were analyzed following multifactorialANOVA. Biochemical parameters such as LPO, CAT, POX,SOD, GR, and proline content were significantly affected byPNP concentration, exposure time, and microalgal species(Table 1). Except for LPO and SOD, there was a significanteffect on levels of the biochemicals due to interactions ofexposure time and PNP concentration. No significant impactof exposure time was observed on SOD activity. Likewise,interaction between the microalgal species and exposure timeresulted in a significant effect on the content of all the bio-chemicals measured except for SOD. The interaction betweenPNP content, exposure time, and microalgal species had sig-nificant effects on CAT, POX, GR, and proline (p� 0.001) andon SOD and LPO (p� 0.005).
Relationship among the biochemical parameters tested
Factor analysis was done with PCA extraction and Varimaxrotation to study the relationship among different biochemicalparameters estimated in the three microalgal species after 96 hof exposure to 20mgL�1 PNP. Three-component analyses werechosen to interpret the results based on the epigen valuesbecause two components could explain only 65.01%, whereasPCA results showed that three components would explain89.12%, of the total variance between the variables analyzed(Table 3). It is clear from the score plot that there is a significantrelationship between proline and carotenoids in component 1;POX, SOD, and GR in component 2; and LPO, CAT, and
itrophenol concentration, exposure time, and microalgal species and theirl parameters analyzed
PC�ET PC�MA ET�MA PC�ET�MA
p F p F p F p
7 0.116 77.68 <0.001 20.09 <0.001 3.72 0.0043 <0.001 82.10 <0.001 67.13 <0.001 143.59 <0.0019 <0.001 183.85 <0.001 49.42 <0.001 8.53 <0.0012 0.016 4.00 0.003 1.53 0.226 3.63 0.0054 <0.001 226.96 <0.001 24.16 <0.001 25.52 <0.0010 <0.001 18.54 <0.001 40.14 <0.001 9.78 <0.001
tration; ET¼ exposure time; MA¼microalgae; LPO¼ lipid peroxidation;one reductase.

Fig. 4. Changes in superoxide dismutase (SOD) activity in microalgaeexposed to p-nitrophenol (PNP). Data aremeans� SD (n¼ 3). Letters on topof the bars for each sampling indicate significant difference betweendifferentPNP concentrations at p� 0.005 based on Tukey’s test. (a) Chlorella sp.,(b) Chlorococcum sp., and (c) Heterochlamydomonas sp.
Fig. 3. Changes in peroxidase (POX) activity in microalgae exposed top-nitrophenol (PNP). Data are means�SD (n¼ 3). Letters on top of the barsfor each sampling indicate significant difference between different PNPconcentrations at p� 0.005 based on Tukey’s test. (a) Chlorella sp.,(b) Chlorococcum sp., and (c) Heterochlamydomonas sp.
1984 Environ. Toxicol. Chem. 31, 2012 S.R. Subashchandrabose et al.
protein in component 3 (Fig. 8). Pearson’s correlation coeffi-cient matrix showed that there was a significant correlation (p�0.001) between LPO and CAT, POX and SOD, proline andSOD, and carotenoids and proline (Table 4). However, corre-lations were significantly negative between protein and prolineand between protein and carotenoids.
Removal of PNP by microalgal species
The addition of 20mgL�1 PNP to the culture mediumresulted in nearly 60, 70, and 90% reduction in growth ofChlorella sp., Heterochlamydomonas sp., and Chlorococcumsp., respectively, compared with untreated controls (data notshown). However, in cultures supplemented with 20mgL�1
PNP there was a decrease in cell number in Chlorella sp.and Heterochlamydomonas sp., while a decrease followed bya slight increase in growth of Chlorococcum sp. was observedafter 7 d of incubation (Fig. 9). At the end of 7 d, growth nearlydoubled in Chlorella sp. and there was a 50% increase inHeterochlamydomonas sp., whereas a close to 50% reductionin cell number was observed in Chlorococcum sp. The abilityof microalgae to grow in the presence of PNP was alsocorrelated with their potential to remove PNP. Removal ofPNP started only after 24 h, and the percentage of removal of
20mgL�1 PNP by Chlorella sp., Heterochlamydomonas sp.,and Chlorococcum sp. was 39, 18, and 4%, respectively, after7 d of incubation.
DISCUSSION
External stress from both biotic and abiotic origins resultsin the production of reactive oxygen species (ROS) in cells.Formation of ROS damages the most vital components of thecells like DNA, protein, and lipids. Reactive oxygen species isalso an important signaling molecule that triggers the expres-sion of various antioxidant defence systems [23]. Microalgaehave a well-developed antioxidant system, as do other eukar-yotes. In microalgae, ROS is produced not only as a result ofrespiration or photosynthesis but also as a response to variousexternal stressors. It is established that several environmentalfactors induce oxidative stress in microalgae including physicalfactors like UV light [11] and ultrasound [24,25]. Much empha-sis has been given to heavy metal–induced oxidative stress inmicroalgae [12]. Only a very few studies have been publishedon the oxidative and biochemical changes induced by organiccompounds in microalgae. Again, in these studies much focuswas given to organic pesticides, polyaromatic hydrocarbons,oil, dioxins, and polychlorinated biphenyls. A constant searchhas been ongoing for valuable biomarkers in microalgae to
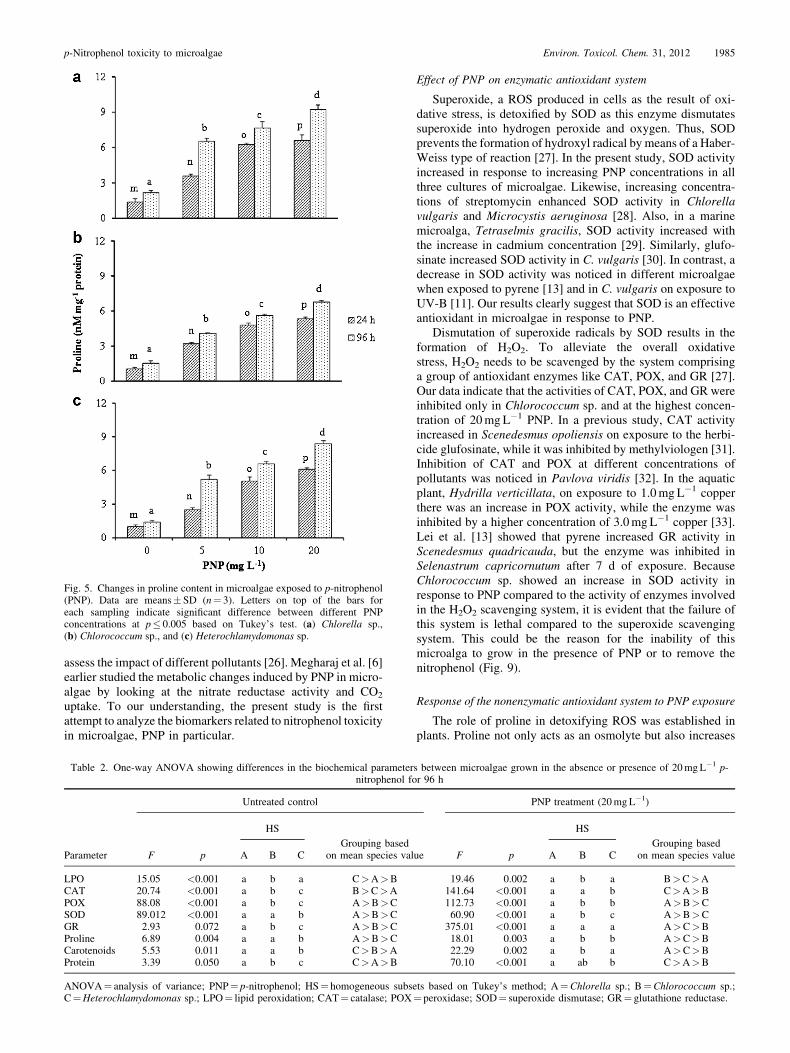
Fig. 5. Changes in proline content in microalgae exposed to p-nitrophenol(PNP). Data are means� SD (n¼ 3). Letters on top of the bars foreach sampling indicate significant difference between different PNPconcentrations at p� 0.005 based on Tukey’s test. (a) Chlorella sp.,(b) Chlorococcum sp., and (c) Heterochlamydomonas sp.
p-Nitrophenol toxicity to microalgae Environ. Toxicol. Chem. 31, 2012 1985
assess the impact of different pollutants [26]. Megharaj et al. [6]earlier studied the metabolic changes induced by PNP in micro-algae by looking at the nitrate reductase activity and CO2
uptake. To our understanding, the present study is the firstattempt to analyze the biomarkers related to nitrophenol toxicityin microalgae, PNP in particular.
Table 2. One-way ANOVA showing differences in the biochemical parameternitrophenol fo
Parameter
Untreated control
F p
HS
Grouping basedon mean species valA B C
LPO 15.05 <0.001 a b a C>A>BCAT 20.74 <0.001 a b c B>C>APOX 88.08 <0.001 a b c A>B>CSOD 89.012 <0.001 a a b A>B>CGR 2.93 0.072 a b c A>B>CProline 6.89 0.004 a a b A>B>CCarotenoids 5.53 0.011 a a b C>B>AProtein 3.39 0.050 a b c C>A>B
ANOVA¼ analysis of variance; PNP¼ p-nitrophenol; HS¼ homogeneous subsC¼Heterochlamydomonas sp.; LPO¼ lipid peroxidation; CAT¼ catalase; POX
Effect of PNP on enzymatic antioxidant system
Superoxide, a ROS produced in cells as the result of oxi-dative stress, is detoxified by SOD as this enzyme dismutatessuperoxide into hydrogen peroxide and oxygen. Thus, SODprevents the formation of hydroxyl radical by means of a Haber-Weiss type of reaction [27]. In the present study, SOD activityincreased in response to increasing PNP concentrations in allthree cultures of microalgae. Likewise, increasing concentra-tions of streptomycin enhanced SOD activity in Chlorellavulgaris and Microcystis aeruginosa [28]. Also, in a marinemicroalga, Tetraselmis gracilis, SOD activity increased withthe increase in cadmium concentration [29]. Similarly, glufo-sinate increased SOD activity in C. vulgaris [30]. In contrast, adecrease in SOD activity was noticed in different microalgaewhen exposed to pyrene [13] and in C. vulgaris on exposure toUV-B [11]. Our results clearly suggest that SOD is an effectiveantioxidant in microalgae in response to PNP.
Dismutation of superoxide radicals by SOD results in theformation of H2O2. To alleviate the overall oxidativestress, H2O2 needs to be scavenged by the system comprisinga group of antioxidant enzymes like CAT, POX, and GR [27].Our data indicate that the activities of CAT, POX, and GR wereinhibited only in Chlorococcum sp. and at the highest concen-tration of 20mgL�1 PNP. In a previous study, CAT activityincreased in Scenedesmus opoliensis on exposure to the herbi-cide glufosinate, while it was inhibited by methylviologen [31].Inhibition of CAT and POX at different concentrations ofpollutants was noticed in Pavlova viridis [32]. In the aquaticplant, Hydrilla verticillata, on exposure to 1.0mgL�1 copperthere was an increase in POX activity, while the enzyme wasinhibited by a higher concentration of 3.0mgL�1 copper [33].Lei et al. [13] showed that pyrene increased GR activity inScenedesmus quadricauda, but the enzyme was inhibited inSelenastrum capricornutum after 7 d of exposure. BecauseChlorococcum sp. showed an increase in SOD activity inresponse to PNP compared to the activity of enzymes involvedin the H2O2 scavenging system, it is evident that the failure ofthis system is lethal compared to the superoxide scavengingsystem. This could be the reason for the inability of thismicroalga to grow in the presence of PNP or to remove thenitrophenol (Fig. 9).
Response of the nonenzymatic antioxidant system to PNP exposure
The role of proline in detoxifying ROS was established inplants. Proline not only acts as an osmolyte but also increases
s between microalgae grown in the absence or presence of 20mgL�1 p-r 96 h
PNP treatment (20mgL�1)
ue F p
HS
Grouping basedon mean species valueA B C
19.46 0.002 a b a B>C>A141.64 <0.001 a a b C>A>B112.73 <0.001 a b b A>B>C60.90 <0.001 a b c A>B>C375.01 <0.001 a a a A>C>B18.01 0.003 a b b A>C>B22.29 0.002 a b a A>C>B70.10 <0.001 a ab b C>A>B
ets based on Tukey’s method; A¼Chlorella sp.; B¼Chlorococcum sp.;¼ peroxidase; SOD¼ superoxide dismutase; GR¼ glutathione reductase.

Fig. 6. Changes in malondialdehyde (MDA) content in microalgae exposedto p-nitrophenol (PNP). Data are means�SD (n¼ 3). Letters on top ofthe bars for each sampling indicate significant difference between differentPNP concentrations at p� 0.005 based on Tukey’s test. (a) Chlorella sp.,(b) Chlorococcum sp., and (c) Heterochlamydomonas sp.
Table 3. Component score coefficient matrix of the biochemicalparameters at 96 h of exposure to 20mgL�1 p-nitrophenol in all three
microalgae using principal component analysis through Varimax rotationand Kaiser normalization
Parameter
Component
1 2 3
LPO 0.110 �0.087 0.393CAT �0.090 �0.014 0.469POX �0.026 0.388 �0.079SOD 0.073 0.366 �0.071GR �0.199 0.289 0.268Proline 0.254 0.174 0.173Carotenoids 0.369 �0.120 �0.053Protein �0.336 �0.043 0.056
LPO¼ lipid peroxidation; CAT¼ catalase; POX¼ peroxidase; SOD¼superoxide dismutase; GR¼ glutathione reductase.
Fig. 7. Influence of p-nitrophenol (PNP) in altering the contents of proteinand carotenoids in microalgae exposed to PNP for 96 h. (Carotenoids areshown as solid lines and protein as dotted lines.)
1986 Environ. Toxicol. Chem. 31, 2012 S.R. Subashchandrabose et al.
the solubility of proteins by interacting with the hydrophobicresidues [34]. Plants and microalgae respond to heavy metals byaccumulating proline [35]. In earlier studies, levels of proline inmicroalgae exposed to varied pollutants was assessed, but therole of proline in protecting the microalgae from the stressinduced by environmental pollutants was not shown clearly.Carotenoids are antioxidants in microalgae and play a vital rolein protecting the photosynthetic system from oxidative damage.Carotenoids like astaxanthin play a significant antioxidant rolein green alga by rapidly detoxifying singlet oxygen, superoxideanion radical, H2O2, and peroxy radical [36]. We, therefore,looked at the response of free proline and carotenoids, thebiochemicals of the nonenzymatic antioxidant system, to PNPexposure. Dose- and time-specific increases in proline contentin all three microalgal cultures indicate that proline is an activedetoxifier of ROS in microalgae induced by PNP. Tripathi andGaur [37] demonstrated that copper at a 10-mM concentrationgreatly increased the intracellular proline content in Scenedes-mus sp.; pretreatment of the alga with proline also significantlyreduced the oxidative stress induced by copper. However, a
Fig. 8. Score plot, based on principal component analysis, showing clustersof the biochemical parameters at 96 h of exposure to 0 to 20mgL�1
p-nitrophenol (PNP) in threemicroalgae. CAT¼ catalase; GR¼ glutathionereductase; LPO¼ lipid peroxidation; POX¼ peroxidase; SOD¼ superoxidedismutase.
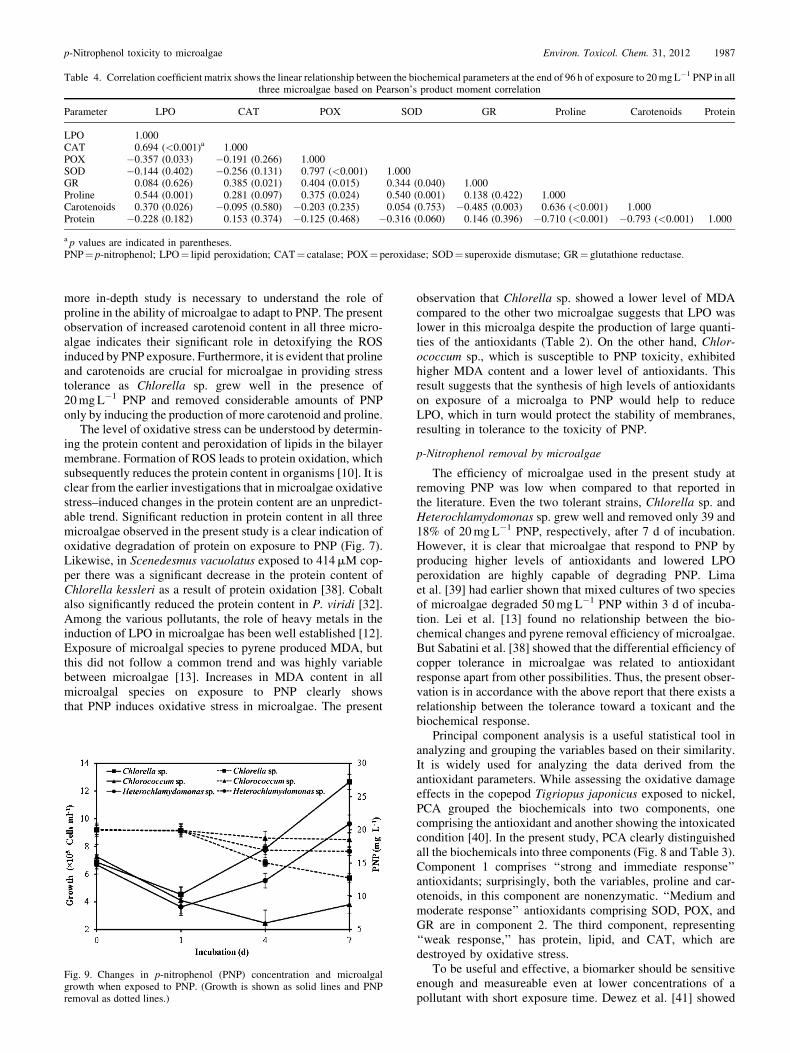
Table 4. Correlation coefficient matrix shows the linear relationship between the biochemical parameters at the end of 96 h of exposure to 20mgL�1 PNP in allthree microalgae based on Pearson’s product moment correlation
Parameter LPO CAT POX SOD GR Proline Carotenoids Protein
LPO 1.000CAT 0.694 (<0.001)a 1.000POX �0.357 (0.033) �0.191 (0.266) 1.000SOD �0.144 (0.402) �0.256 (0.131) 0.797 (<0.001) 1.000GR 0.084 (0.626) 0.385 (0.021) 0.404 (0.015) 0.344 (0.040) 1.000Proline 0.544 (0.001) 0.281 (0.097) 0.375 (0.024) 0.540 (0.001) 0.138 (0.422) 1.000Carotenoids 0.370 (0.026) �0.095 (0.580) �0.203 (0.235) 0.054 (0.753) �0.485 (0.003) 0.636 (<0.001) 1.000Protein �0.228 (0.182) 0.153 (0.374) �0.125 (0.468) �0.316 (0.060) 0.146 (0.396) �0.710 (<0.001) �0.793 (<0.001) 1.000
a p values are indicated in parentheses.PNP¼ p-nitrophenol; LPO¼ lipid peroxidation; CAT¼ catalase; POX¼ peroxidase; SOD¼ superoxide dismutase; GR¼ glutathione reductase.
p-Nitrophenol toxicity to microalgae Environ. Toxicol. Chem. 31, 2012 1987
more in-depth study is necessary to understand the role ofproline in the ability of microalgae to adapt to PNP. The presentobservation of increased carotenoid content in all three micro-algae indicates their significant role in detoxifying the ROSinduced by PNP exposure. Furthermore, it is evident that prolineand carotenoids are crucial for microalgae in providing stresstolerance as Chlorella sp. grew well in the presence of20mgL�1 PNP and removed considerable amounts of PNPonly by inducing the production of more carotenoid and proline.
The level of oxidative stress can be understood by determin-ing the protein content and peroxidation of lipids in the bilayermembrane. Formation of ROS leads to protein oxidation, whichsubsequently reduces the protein content in organisms [10]. It isclear from the earlier investigations that in microalgae oxidativestress–induced changes in the protein content are an unpredict-able trend. Significant reduction in protein content in all threemicroalgae observed in the present study is a clear indication ofoxidative degradation of protein on exposure to PNP (Fig. 7).Likewise, in Scenedesmus vacuolatus exposed to 414mM cop-per there was a significant decrease in the protein content ofChlorella kessleri as a result of protein oxidation [38]. Cobaltalso significantly reduced the protein content in P. viridi [32].Among the various pollutants, the role of heavy metals in theinduction of LPO in microalgae has been well established [12].Exposure of microalgal species to pyrene produced MDA, butthis did not follow a common trend and was highly variablebetween microalgae [13]. Increases in MDA content in allmicroalgal species on exposure to PNP clearly showsthat PNP induces oxidative stress in microalgae. The present
Fig. 9. Changes in p-nitrophenol (PNP) concentration and microalgalgrowth when exposed to PNP. (Growth is shown as solid lines and PNPremoval as dotted lines.)
observation that Chlorella sp. showed a lower level of MDAcompared to the other two microalgae suggests that LPO waslower in this microalga despite the production of large quanti-ties of the antioxidants (Table 2). On the other hand, Chlor-ococcum sp., which is susceptible to PNP toxicity, exhibitedhigher MDA content and a lower level of antioxidants. Thisresult suggests that the synthesis of high levels of antioxidantson exposure of a microalga to PNP would help to reduceLPO, which in turn would protect the stability of membranes,resulting in tolerance to the toxicity of PNP.
p-Nitrophenol removal by microalgae
The efficiency of microalgae used in the present study atremoving PNP was low when compared to that reported inthe literature. Even the two tolerant strains, Chlorella sp. andHeterochlamydomonas sp. grew well and removed only 39 and18% of 20mgL�1 PNP, respectively, after 7 d of incubation.However, it is clear that microalgae that respond to PNP byproducing higher levels of antioxidants and lowered LPOperoxidation are highly capable of degrading PNP. Limaet al. [39] had earlier shown that mixed cultures of two speciesof microalgae degraded 50mgL�1 PNP within 3 d of incuba-tion. Lei et al. [13] found no relationship between the bio-chemical changes and pyrene removal efficiency of microalgae.But Sabatini et al. [38] showed that the differential efficiency ofcopper tolerance in microalgae was related to antioxidantresponse apart from other possibilities. Thus, the present obser-vation is in accordance with the above report that there exists arelationship between the tolerance toward a toxicant and thebiochemical response.
Principal component analysis is a useful statistical tool inanalyzing and grouping the variables based on their similarity.It is widely used for analyzing the data derived from theantioxidant parameters. While assessing the oxidative damageeffects in the copepod Tigriopus japonicus exposed to nickel,PCA grouped the biochemicals into two components, onecomprising the antioxidant and another showing the intoxicatedcondition [40]. In the present study, PCA clearly distinguishedall the biochemicals into three components (Fig. 8 and Table 3).Component 1 comprises ‘‘strong and immediate response’’antioxidants; surprisingly, both the variables, proline and car-otenoids, in this component are nonenzymatic. ‘‘Medium andmoderate response’’ antioxidants comprising SOD, POX, andGR are in component 2. The third component, representing‘‘weak response,’’ has protein, lipid, and CAT, which aredestroyed by oxidative stress.
To be useful and effective, a biomarker should be sensitiveenough and measureable even at lower concentrations of apollutant with short exposure time. Dewez et al. [41] showed

1988 Environ. Toxicol. Chem. 31, 2012 S.R. Subashchandrabose et al.
that CAT is a highly sensitive enzymatic biomarker againstcopper and fludioxonil in Scenedesmus obliquus. The presentstudy clearly suggests that LPO, carotenoids, and proline couldbe effective biomarkers in microalgae to assess the extent ofPNP pollution. A more detailed study on these biochemicals inmicroalgae will be useful for developing biomarkers requiredfor screening the impact of PNP pollution. Also, furthervalidation of these biochemicals in different microalgae fromdifferent geographical locations could be useful for their effec-tive use as biomarkers.
Acknowledgement—S.R. Subashchandrabose acknowledges UniSA forproviding UPS scholarship and Cooperative Research Centre for Conta-mination Assessment and Remediation of the Environment for PhDscholarship, and K. Venkateswarlu thanks the government of Australia(Department of Education, Employment and Workplace Relations) for theEndeavour Executive Award.
REFERENCES
1. Oubina A, Ballesteros B, Galve R, Barcelo D, Marco M. 1999.Development and optimization of an indirect enzyme-linked immuno-sorbent assay for 4-nitrophenol. Application to the analysis of certifiedwater samples. Anal Chim Acta 387:255–266.
2. Kadiyala V, Spain JC. 1998. A two-component monooxygenasecatalyzes both the hydroxylation of p-nitrophenol and the oxidativerelease of nitrite from 4-nitrocatechol in Bacillus sphaericus JS905.Appl Environ Microbiol 64:2479–2484.
3. Verchot L, Borelli T. 2005. Application of para-nitrophenol (pNP)enzymeassays in degraded tropical soils.Soil BiolBiochem37:625–633.
4. Bhatti ZI, Toda H, Furukawa K. 2002. p-Nitrophenol degradation byactivated sludge attached on nonwovens. Water Res 36:1135–1142.
5. Esteban E, Rubin C, Hill R, Olson D, Pearce K. 1996. Associationbetween indoor residential contamination with methyl parathion andurinary para-nitrophenol. J Expo Anal Environ Epidemiol 6:375–387.
6. Megharaj M, Pearson H, Venkateswarlu K. 1992. Effects of phenoliccompounds on growth andmetabolic activities ofChlorella vulgaris andScenedesmus bijugatus isolated from soil. Plant Soil 140:25–34.
7. Megharaj M, Pearson HW, Venkateswarlu K. 1991. Toxicity ofp-aminophenol and p-nitrophenol to Chlorella vulgaris and two speciesof Nostoc isolated from soil. Pestic Biochem Physiol 40:266–273.
8. Megharaj M, Madhavi DR, Sreenivasulu C, Umamaheswari A,Venkateswarlu K. 1994. Biodegradation of methyl parathion by soilisolates of microalgae and cyanobacteria. Bull Environ Contam Toxicol53:292–297.
9. RamakrishnanB,MegharajM, Venkateswarlu K, Naidu R, SethunathanN. 2010. The impacts of environmental pollutants on microalgae andcyanobacteria. Crit Rev Environ Sci Technol 40:699–821.
10. Mittler R. 2002. Oxidative stress, antioxidants and stress tolerance.Trends Plant Sci 7:405–410.
11. Malanga G, Calmanovici G, Puntarulo S. 1997. Oxidative damage tochloroplasts from Chlorella vulgaris exposed to ultraviolet-B radiation.Physiol Plant 101:455–462.
12. Pinto E, Sigaud-kutner TCS, Leitao MAS, Okamoto OK, Morse D,Colepicolo P. 2003. Heavy metal–induced oxidative stress in algae.J Phycol 39:1008–1018.
13. LeiA,HuZ,WongY,TamN.2006.Antioxidant responses ofmicroalgalspecies to pyrene. J Appl Phycol 18:67–78.
14. Megharaj M, Venkateswarlu K, Rao A. 1986. Growth response of fourspecies of soil algae to monocrotophos and quinalphos. Environ Pollut42:15–22.
15. Chance B, Maehly AC. 1955. Assay of catalases and peroxidases.Methods Enzymol 2:773–775.
16. Beers RF, Sizer IW. 1952. A spectrophotometric method for measuringthe breakdown of hydrogen peroxide by catalase. J Biol Chem 195:133–140.
17. Mavis R, Stellwagen E. 1968. Purification and subunit structure ofglutathione reductase from bakers’ yeast. J Biol Chem 243:809–814.
18. McCord J, Fridovich I. 1969. Superoxide dismutase. J Biol Chem 244:6049–6055.
19. Heath R, Packer L. 1968. Photoperoxidation in isolated chloroplasts: I.Kinetics and stoichiometry of fatty acid peroxidation. Arch BiochemBiophys 125:189–198.
20. Bates LS, Waldren RP, Teare ID. 1973. Rapid determination of freeproline for water-stress studies. Plant Soil 39:205–207.
21. Bradford M. 1976. A rapid and sensitive method for the quantitation ofmicrogram quantities of protein utilizing the principle of protein-dyebinding. Anal Biochem 72:248–254.
22. WellburnA. 1994.The spectral determination of chlorophyllsa and b, aswell as total carotenoids, usingvarious solventswith spectrophotometersof different resolution. Plant Physiol 144:307–313.
23. Apel K, Hirt H. 2004. Reactive oxygen species: Metabolism, oxidativestress, and signal transduction. Plant Biol 55:373–399.
24. Malanga G, Puntarulo S. 1995. Oxidative stress and antioxidant contentin Chlorella vulgaris after exposure to ultraviolet-B radiation. PhysiolPlant 94:672–679.
25. Chen B, Huang J, Wang J, Huang L. 2008. Ultrasound effects on theantioxidative defense systems of Porphyridium cruentum. ColloidsSurf, B 61:88–92.
26. Torres MA, Barros MP, Campos SCG, Pinto E, Rajamani S, Sayre RT,Colepicolo P. 2008. Biochemical biomarkers in algae and marinepollution: A review. Ecotoxicol Environ Saf 71:1–15.
27. Arora A, Sairam R, Srivastava G. 2002. Oxidative stress andantioxidative system in plants. Curr Sci 82:1227–1238.
28. Qian H, Li J, Pan X, Sun Z, Ye C, Jin G, Fu Z. 2012. Effects ofstreptomycin on growth of algae Chlorella vulgaris and Microcystisaeruginosa. Environ Toxicol 27:229–237.
29. Okamoto OK, Asano CS, Aidar E, Colepicolo P. 1996. Effectsof cadmium on growth and superoxide dismutase activity of themarine microalga Tetraselmis gracilis (Prasinophyceae). J Phycol 32:74–79.
30. Qian H, Chen W, Sheng GD, Xu X, Liu W, Fu Z. 2008. Effects ofglufosinate on antioxidant enzymes, subcellular structure, and geneexpression in the unicellular green alga Chlorella vulgaris. AquatToxicol 88:301–307.
31. Fodorpataki L, Bartha C, Keresztes Z. 2009. Stress–physiologicalreactions of the green alga Scenedesmus opoliensis to water pollutionwith herbicides. Analele Universitatii din Oradea, Fascicula Biologie17:51–56.
32. Li M, Zhu Q, Hu C-w, Chen L, Liu Z-l, Kong Z-m. 2007. Cobalt andmanganese stress in the microalgaPavlova viridis (Prymnesiophyceae):Effects on lipid peroxidation and antioxidant enzymes. J Environ Sci19:1330–1335.
33. Byl T, Klaine S. 1991. Peroxidase activity as an indicator of sublethalstress in the aquatic plant Hydrilla verticillata (Royle). Plants ToxicAssess 2:101–106.
34. Kocsy G, Laurie R, Szalai G, Szilagyi V, Simon Sarkadi L, Galiba G,De Ronde J. 2005. Genetic manipulation of proline levels affectsantioxidants in soybean subjected to simultaneous drought and heatstresses. Physiol Plant 124:227–235.
35. Siripornadulsil S, Traina S, Verma DPS, Sayre RT. 2002. Molecularmechanisms of proline-mediated tolerance to toxic heavy metals intransgenic microalgae. Plant Cell 14:2837–2847.
36. Kobayashi M. 2000. In vivo antioxidant role of astaxanthin underoxidative stress in the green alga Haematococcus pluvialis. ApplMicrobiol Biotechnol 54:550–555.
37. Tripathi BN, Gaur JP. 2004. Relationship between copper- and zinc-induced oxidative stress and proline accumulation in Scenedesmus sp.Planta 219:397–404.
38. Sabatini SE, Juarez AB, Eppis MR, Bianchi L, Luquet CM, Rıos deMolina MdC. 2009. Oxidative stress and antioxidant defenses in twogreen microalgae exposed to copper. Ecotoxicol Environ Saf 72:1200–1206.
39. Lima SAC, Castro PML, Morais R. 2003. Biodegradation ofp-nitrophenol by microalgae. J Appl Phycol 15:137–142.
40. Wang M, Wang G. 2010. Oxidative damage effects in the copepodTigriopus japonicus Mori experimentally exposed to nickel. Ecotox-icology 19:273–284.
41. Dewez D, Geoffroy L, Vernet G, Popovic R. 2005. Determination ofphotosynthetic and enzymatic biomarkers sensitivity used to evaluatetoxic effects of copper and fludioxonil in alga Scenedesmus obliquus.Aquat Toxicol 74:150–159.
Copyright © 2022 FDOKUMEN




















