
Oxidative stress in fish exposed to model xenobiotics. Oxidatively modified forms of...
16
ELSEVIER Chemico-Biological Interactions 98 (1995) 267-282 Oxidative stress in fish exposed to model xenobiotics. Oxidatively modified forms of Cu,Zn-superoxide dismutase as potential biomarkers J.R. Pedrajas, J. Peinado*, J. Lbpez-Barea Departamento de Bioquimica y Biologia Molecular. Institute de Biologia Bksica y Aplicada, Facultad de Veterinaria, Universidad de Cdrdoba. Avenida de Medina Azahara sn, 14071 Cdrdoba. Spain Received 13 February 1995; revision received 17 June 1995; accepted 22 June 1995 Abstract Fish (Sparus aurata) were intraperitoneally injected with model xenobiotics and several biomarkers of oxidative stress were analysed after 2 and 7 days exposure. The levels of soluble thiobarbituric acid reactive substances (TBARS) increased markedly in animals treated with polar xenobiotics, CuCI, or paraquat; exposure to the apolar xenobiotics, dieldrin or malathion, enhanced significantly the microsomal TBARS while decreasing the microsomal glutathione transferase activity. The specific superoxide dismutase (SOD) activity increased in Cu(Il)-injected animals but diminished in fish exposed to paraquat. After isoelectrofocus- ing separation and activity staining cell-free extracts of lish exposed to Cu(II), dieldrin or malathion displayed two new Cu,Zn-SOD isoforms of intermediate pl. An additional Mn- SOD was observed in dieldrin-injected fish, but only a faint new acidic isoform was observed in paraquat-injected animals. The new SOD bands were reproduced in vitro by incubation 01 cell-free extracts with systems generating superoxide anion or hydrogen peroxide and with a terr-butyl hydroperoxide/ADP-Fe system. Metallothionein induction was observed in Cu(II) or paraquat-exposed fish, but not in animals injected with apolar xenobiotics. So, the new SOD bands are possibly oxidized forms of this enzyme and can be considered as useful earl) biomarkers of oxidative stress due to transition metals or organic xenobiotics. Keywords: Fish; Pollutants; Oxidative damage; Thiobarbituric acid reactive substances; Superoxide dismutase * Corresponding author. 0009-279719YSO9.50 0 1995 Elsevier Science Ireland Ltd. All rights reserved SSDI 0009-2797(95)03651-2
Transcript of Oxidative stress in fish exposed to model xenobiotics. Oxidatively modified forms of...

ELSEVIER Chemico-Biological Interactions 98 (1995) 267-282
Oxidative stress in fish exposed to model xenobiotics. Oxidatively modified forms of Cu,Zn-superoxide dismutase as potential
biomarkers
J.R. Pedrajas, J. Peinado*, J. Lbpez-Barea
Departamento de Bioquimica y Biologia Molecular. Institute de Biologia Bksica y Aplicada, Facultad de
Veterinaria, Universidad de Cdrdoba. Avenida de Medina Azahara sn, 14071 Cdrdoba. Spain
Received 13 February 1995; revision received 17 June 1995; accepted 22 June 1995
Abstract
Fish (Sparus aurata) were intraperitoneally injected with model xenobiotics and several biomarkers of oxidative stress were analysed after 2 and 7 days exposure. The levels of soluble thiobarbituric acid reactive substances (TBARS) increased markedly in animals treated with polar xenobiotics, CuCI, or paraquat; exposure to the apolar xenobiotics, dieldrin or
malathion, enhanced significantly the microsomal TBARS while decreasing the microsomal glutathione transferase activity. The specific superoxide dismutase (SOD) activity increased in Cu(Il)-injected animals but diminished in fish exposed to paraquat. After isoelectrofocus- ing separation and activity staining cell-free extracts of lish exposed to Cu(II), dieldrin or
malathion displayed two new Cu,Zn-SOD isoforms of intermediate pl. An additional Mn- SOD was observed in dieldrin-injected fish, but only a faint new acidic isoform was observed in paraquat-injected animals. The new SOD bands were reproduced in vitro by incubation 01 cell-free extracts with systems generating superoxide anion or hydrogen peroxide and with a
terr-butyl hydroperoxide/ADP-Fe system. Metallothionein induction was observed in Cu(II) or paraquat-exposed fish, but not in animals injected with apolar xenobiotics. So, the new SOD bands are possibly oxidized forms of this enzyme and can be considered as useful earl) biomarkers of oxidative stress due to transition metals or organic xenobiotics.
Keywords: Fish; Pollutants; Oxidative damage; Thiobarbituric acid reactive substances; Superoxide dismutase
* Corresponding author.
0009-279719YSO9.50 0 1995 Elsevier Science Ireland Ltd. All rights reserved SSDI 0009-2797(95)03651-2

268 J. R. Pedraps et al. / Chemico-Biological lnteracrions 98 (1995) 267-282
1. Introduction
Oxidative damages to living beings affect nucleic acids, proteins, lipids and carbo- hydrates [l]. Formation of oxidized DNA or protein adducts has been proposed as biomarkers of oxidative stress [ 1,2]. Lipids, especially the polyunsaturated so abun- dant in fish, are highly prone to oxidative attack generating malondialdehyde and other aldehydes [3], which upon reaction with thiobarbituric acid yield coloured products which absorb at 535 nm, named thiobarbituric acid reactive substances (TBARS). Spectrophotometric determination of TBARS is generally considered as an indication of oxidative damage to membrane lipids [4], although this procedure is not without limitations [5]. A membrane-bound glutathione transferase with per- oxidase activity protects microsomal and plasma membranes against lipid peroxida- tion [6,7].
Superoxide dismutases (SOD) are metalloenzymes playing a key role in the de- fence against the toxic effects of reactive oxygen species by disproportionating super- oxide anion, according to the reaction [8]:
Oz5 +02: +2H+ - 02+HzOz
Eukaryotes have three SOD isoenzymes, a Cu,Zn-SOD present in cytosol, nucleus and peroxisomes, readily inhibited by CN-, a mitochondrial Mn-SOD, refractory to CN- inhibition [8] and an extracellular SOD, mainly located in the interstitial space of tissues [9]. Cu,Zn-SOD is prone to oxidative modification and inactivation due to reactive oxygen species [lO,l 11. So, HzOz reacts with the enzyme-bound Cu’ (via Fenton reaction) generating Cu 2+-OH. which could then oxidatively attack an adjacent histidine and thus inactivate the enzyme [ 121.
The biochemical changes associated with environmental pollution have been recently studied in fish (Mugif sp.) from sites of the Spanish littoral with different pollution levels. The contaminated site contained high levels of Cu and other metals, polyaromatic hydrocarbons, polychlorinated biphenyls and pesticides [ 131, whose toxicity is mediated through reactive oxygen species [ 14,151. The oxidized glutathione redox status and the increased activities of several antioxidative enzymes in polluted fish confirmed that they were subjected to oxidative stress [ 131. New Cu,Zn-SOD forms of intermediate pl appear in hepatic extracts of Mugif sp. from polluted sites, and have been purified to homogeneity. New acidic SOD bands, close- ly resembling those of polluted fish, are formed when the most active pure forms are incubated with hydrogen peroxide. These new SOD bands have been proposed as biomarkers of pollutants producing oxidative stress [ 161.
Metallothioneins are cysteine-rich low MW proteins which bind transition metals, regulate endogenous Cu and Zn, detoxicate excess of these and other pollutant met- als, act as free radical scavengers, and are induced as a part of a general stress re- sponse [ 171. Increased transcription of metallothionein genes by several metals is the basis for its use as a biomarker [ 181.
In the present work, fish (Sparus aurata) have been intraperitoneally injected with some of the more representative pollutants detected in our previous studies. The

J. R. Pedrajas et al. / Chemico-Biological Interactions 98 (I 995) 267-282 269
following chemicals were selected as model xenobiotics: copper (II), a redox-active transition metal; paraquat, a herbicide generating reactive oxygen species through redox cycling [19]; dieldrin, an organochlorine cyclodiene epoxide which, as its stereoisomer endrin, could be toxic through reactive oxygen species [20,21]; and the organophosphorous pesticide malathion. The polychlorinated biphenyl Aroclor 1254 was used as a positive control to validate the experiment. The reliability of the new acidic SOD bands as biomarkers of oxidative stress has been contrasted with the levels of soluble and microsomal TBARS and with metallothionein induction.
2. Materials and methods
2.1. Chemicals The following compounds were purchased from the indicated commercial sources:
Aroclor 1254, from Chem Service (Phase Separations Ltd., UK); malathion and di- eldrin, from Supelco (USA); type I glucose oxidase, from Boehringer (Germany); tert-butylhydroperoxide (tBuOOH), from Aldrich (England); glucose and FeC13, from Merck (Germany). Phenylmethylsulfonylfluoride (PMSF), benzamidine, ADP, azide, CuC12, cytochrome c, xanthine, xanthine oxidase, nitroblue tetrazolium, N,N,N’,N’-tetramethylethylendiamine (TEMED), bovine serum albumin and thio- barbituric acid were from Sigma (St. Louis, USA).
2.2. Animals and experimental exposure to xenobiotics Sexually immature gilthead seabrams (Sparus aurata) of 150-170 g weight were
maintained in 1800 1 tanks at 15°C with constant aeration and 3600 1 daily water renovation. Fish, previously accommodated to the tanks for 2 weeks, were daily fed with feed (Dibaq-Diproteg, Madrid) at a rate of 0.5% total weight of animals in the tank. The experience started by netting a convenient number of animals; to avoid excessive damage during handling the fish were introduced for a few minutes into a 20 1 container with 10 mg/l of quinaldine sulphate. The anaesthetised fish were in- traperitoneally injected with the model xenobiotics and passed into independent 1800 1 tanks. The tested doses (mg/kg of body weight) were: 0.2 and 1.0 for Cu(II), 50 for paraquat, 500 and 1000 for Aroclor 1254, 0.15 and 1.0 for dieldrin, and 1.5 and 7.5 for malathion. Paraquat and CuC12 were dissolved in 0.9% NaCl, while Aroclor 1254, dieldrin and malathion were dissolved in corn oil. Controls were ob- tained by injecting saline solution or oil alone. Ten animals were used for each con- trol or treatment, and 5 animals were captured after 2 and 7 days. Immediately, their spinal cords were severed and livers were excised, frozen in liquid Nz and kept at -80°C. Livers were ground in a mortar with liquid N2 and kept at -80°C until being analysed. Assays were carried out in pools made from 5 livers of each sample.
2.3. Ultracentrifuged cell-free extract and microsome preparations Ground pooled livers were washed with ice-cold 0.9% NaCl (w/v) and disrupted
in 50 mM K-phosphate buffer (pH 7.8) containing 0.1 mM EDTA and 1 mM PMSF, using an Ultra-Turrax homogeniser. The homogenate was further sonicated by three 20 s pulses in a Braun Labsonic 2000 U apparatus at 28 W output. The homogenate

270 J. R. Pedrajas et al. / Chemico-Biological Interactions 9X (1995) 267-282
was cleared by ultracentrifugation at 66 000 x g for 110 min in a Beckman L8-80 M ultracentrifuge. The resulting cell-free extract was stored at -80°C until used for analysis of SOD bands and biochemical determinations.
Microsomes were prepared as previously described [22]: ground liver washed with 0.9% NaCl (w/v) was mixed with 10 mM Tris-HCl buffer (pH 7.5,4 ml/g) containing 1 mM EDTA, 5 mM GSH, 2 mM DTT, 1 mM PMSF and 0.25 M sucrose and disrupted in a glass-teflon Potter-Elvehjem. The homogenate was diluted with 3
volumes of ice-cold 1.15% KC1 (w/v), centrifuged at 9000 x g for 10 min, and the supernatant ultracentrifuged at 105 000 x g for 60 min. The pellet was resuspended in 15 ml of 1.15% KC1 and resedimented at 105 000 x g for 30 min. The washed
microsomes were resuspended in 0.1 M phosphate buffer (pH 7.4, 0.5 ml/g of liver)
and stored at -80°C until used for TBARS and microsomal glutathione transferase
determinations.
2.4. Incubation of cell-free extracts with oxidizing species
A cell-free extract was especially prepared from 1.5 g of ground fish liver after 2 days injection with saline solution, homogenized with 6 ml of 50 mM Tris-HCl pH 7.8, containing 0.1 mM EDTA and 1 mM of PMSF and benzamidine, and centrifug-
ed at 15 000 x g for 20 min. Samples with 32 mg of total protein were incubated at 37°C with the different sources of free radicals in 1.5 ml final volume. At the in- dicated times 75 ~1 and 140 ~1 of each incubation mixture were used for analysis of SOD bands by IEF, and for TBARS determinations, respectively. Azide (1 mM) was
added to inhibit catalase released from peroxisomes. Superoxide anion was generated with a system formed by 1 mM xanthine and 0.15 U xanthine oxidase; hy-
drogen peroxide was generated using 5 mM glucose and 2.25 U glucose oxidase [lo]. When indicated, tBuOOH was added at 0.5 or 1.5 mM, in the presence of 1.5 mM
ADP (previously neutralized at pH 7.8) and 10 PM FeC& [23].
2.5. Biochemical determinations
For determination of TBARS 0.25 ml of sample containing 3 mg of protein from
cell-free extract, 4 mg of microsomal protein or 6 mg of protein from the ultracen- trifuged cell-free extract were assayed according to the procedure previously describ- ed [4].
SOD activity was measured spectrophotometrically by the xanthine-xanthine
oxidase-cytochrome c assay [24]. Mn-SOD was determined in the presence of 1 mM KCN (similar activity was observed with 2 or 4 mM KCN) and Cu,Zn-SOD as the difference between the activity determined in the absence (total activity) and the presence of cyanide. Microsomal glutathione-S-transferase was assayed with 1 mM
I-chloro-2,4_dinitrobenzene and 1 mM GSH [25]. Protein was determined by the biuret-phenol method, with BSA as standard [26]. Isoelectrofocusing was carried out in a PhastSystem equipment (Pharmacia), using gels with pH gradient 6.5-4.0 (PhastGel IEF) and the pl values determined with an IEF calibration kit (Phar-
macia). In situ staining for SOD activity was done immediately after electrophoresis using NBT, riboflavin and TEMED [27].
For separation of metallothioneins by size-exclusion chromatography, 2 g of fish

J. R. Pedrajas et al. / Chemico-Biological Interactions 98 (199s) 267-282 271
liver were mixed with 8 ml of 10 mM Tris-HCl (pH 8.0) prepared with ultrapure water (Mini-Q, Millipore) and previously deaerated. The samples were gently homogenised by hand in a glass-teflon Potter-Elvehjem and sonicated with three 20 s pulses at 28 W (Labsonic U, Braun). The homogenate was ultracentrifuged at 105 000 x g for 2 h. Then, 5.5 ml of supernatant were loaded into a G-75 Sephadex column (2.5 x 33 cm) and eluted at 10 ml/h with 10 mM Tris-HCl buffer (pH 8.0). The absorbance at 230 nm was measured in each sample, and the Zn and Cu content determined by atomic absorption spectrometry (model SP 9, Pye-Unicam).
3. Results
The present work intended to study the toxic effects of several xenobiotics on the fish Sparus aurutu under controlled exposure, including Aroclor 1254 as a positive control. The ethoxyresorufin-O-deethylase (EROD) activity of cytochrome P4501A1, a well established biomarker of xenobiotic exposure [28], increased 3.1 and 5.7-fold (results not shown) in fish injected with Aroclor after 2 or 7 days, respectively, thus confirming the success of the experiment. The toxicity of most xenobiotics studied can be related with reactive oxygen species [14,20,21,29]. Thus, exposed animals were analysed for signs of oxidative stress, such as increased thio- barbituric acid reactive substances (TBARS), changes in the activities of microsomal glutathione-S-transferase (mGST) and superoxide dismutase (SOD) and formation of new SOD forms. Induction of metallothionein-like proteins was also studied.
3.1. TBARS levels and mGST activities
Table 1 shows the soluble and microsomal TBARS detected in Spat-us aurata after exposure to CuC12 or paraquat. Soluble TBARS increased to 185% and 136% of the control values in fish injected with 0.2 mg/kg CuC12 after 2 and 7 days, respectively. Soluble TBARS increased to 509% in fish exposed to 50 mg/kg paraquat after 2 days, although they showed extensive liver damage and none survived up to the seventh day. Nevertheless, microsomal TBARS contents were not markedly altered by expo- sure to either of these two polar xenobiotics.
Table I TBARS in fish injected with polar xenobiotics
Treatment (dose in mg/kg) Soluble TBARS Microsomal TBARS
2 days 7 days 2 days 1 days
Control (saline solution) 100 100 100 100
+ CUCI, (0.2) I85 136 II4 71
+ Paraquat (50) 509 $ 129 $
The data are shown as percentage of the corresponding control, 0. I I4 or 0.210, for the soluble TBARS
after 2 or 7 days injection, and 0.136 or 0.150 for microsomal TBARS after 2 or 7 days treatment.
$Fish died 6 days after paraquat injection.

272 J. R. Pedrajas et al. / Chemico-Biological Interactions 98 (199s) 267-282
Table 2 Microsomal TBARS and glutathione transferase activity in fish injected with apolar model xenobiotics
Treatment (dose After 2 days After 7 days
in m&g) TBARSa mGSTb TBARS” mGST b
Control (corn oil) 0.10 f 0.01 0.26 f 0.03 0.29 * 0.02 0.18 + 0.01
(100) (100) (100) (100) + Dieldrin (0.15) 0.30 f 0.04*** 0.14 f 0.02** 0.21 f 0.031 0.15 f 0.01*
(300) (54) (72) (831 (0.67) 0.18 f 0.02” 0.19 f 0.02’ 0.84 f O.Ol*** 0. IO l 0.003***
(180) (73) (290) (56) + Malathion (1.5) 0.21 f 0.05; 0.16 zt 0.004** 0.14 f 0.03** 0.17 l 0.01
(210) (62) (48) (94) (7.5) 0.09 f 0.01 0.23 zt 0.01 0.46 zt 0.05** 0. I I l 0.002***
(90) (88) (158) (61) + Aroclor 1254 0. II f 0.03 0.26 zt 0.01 0.07 l 0.001*** 0.28 zt O.Ol***
(500) (110) (100) (24) (156) (1000) 0.13 l 0.01 0.21 f 0.003* 0.15 l 0.07; 0.19 f 0.01
(130) (81) (52) (106)
aAbsorbance at 535 nm.
bEnzymatic activity values, expressed as U/mg of protein, are indicated as mean * SD., for 4 indepen-
dent determinations. The relative value of each parameter is shown in parenthesis as percentage of the
corresponding controls. Asterisks indicate statistical significance of differences between treatment and
corresponding controls: *P < 0.05. **P < 0.01, ***P < 0.001.
On the contrary, significant changes were observed in the content of microsomal
TBARS (mTBARS) in fish exposed to apolar xenobiotics. Table 2 shows that signifi-
cant mTBARS increases were detected in fish injected with dieldrin after 2 days (300% and 180% low and high dose, respectively) and 7 days (290%, high dose). Similarly, malathion-injected fish also contained higher mTBARS contents than the corresponding controls after 2 days at low dose (210%) and 7 days at high dose
(158%). Although low dieldrin and malathion doses yielded after 7 days values even below those of control, the results of Table 2 indicate that fish injected with these two xenobiotics displayed, in general, higher mTBARS contents; nevertheless, a clear dose-response relationship was lacking. Fish injected with Aroclor 1254 show-
ed mTBARS levels similar to control after 2 days, but significant decreases after 7 days exposure. It should be noticed that fish injected with corn oil showed after 7 days a 2.9-fold increase in mTBARS in comparison with the control at 2 days, thus suggesting that this vehicle by itself produced some oxidative stress.
In addition, Table 2 shows the inverse relationship between the levels of TBARS and glutathione-S-transferase activity found in microsomes. Thus, the mTBARS in- creases in fish exposed to dieldrin, malathion (and also to corn oil) were paralleled with significant decreases in mGST activity, while the decrease of mTBARS
observed in fish injected with 500 mg/kg Aroclor after 7 days was linked to signifi- cant increases in mGST activity.

J.R. Pedrajas et al. / Chemico-Biological Interactions 98 (1995) 267-282 213
Table 3
Cu,Zn- and Mn-superoxide dismutase activities in fish injected with model xenobiotics
Treatment (dose Enzymatic activity (%) in mg/kg)
After 2 days
Total Cu,Zn-SOD
After I days
Mn-SOD Total Cu,Zn-SOD Mn-SOD
Control 12.9 f 0.8
+CuCI,
(0.2) 14.1 f 0.3
(1.0) 16.6 l 1.5;
+ Paraquat
(50) 9.0 f 0.3**
Control 14.4 * 1.6
+ Dieldrin
(0.15) 16.6 * 1.5
(0.67) 14.4 f 1.6
+ Malathion
(1.5) 14.0 f 1.5
(7.5) 15.6 f 0.2
+ Aroclor 1254
(500) 12.0 * 0.3
(1000) 14.1 f 0.8
10.6 f 0.7 2.3 f 0.3
11.3 f 0.2 2.8 + 0.2
13.5 f 1.5’ 3.1 l 0.7
6.7 f 0.2*** 2.3 f 0.3
11.5 f I.0 2.9 + 0.6
13.2 f 1.1 3.4 l 0.3
10.9 * 1.4 3.5 l 0.4
11.5 f I.1 2.5 f 0.4
12.7 f 0.5 2.9 f 0.3
9.3 f 0.3; 2.7 * 0.1
11.4 f 0.7 2.7 f 0.1
15.1 f 1.3 13.0 f 1.5
15.4 f 1.7 13.2 f I.1
16.5 f 0.7 13.6 + I.0
15.8 f 1.0 13.2 + I.0
15.3 l 1.5 12.4 f 1.0
15.1 f 0.9 12.3 f 0.7
15.3 f 1.1 12.4 f 1.4
14.2 + 0.9 12.1 f 0.9
14.7 f 0.8 12.7 f 1.1
15.2 f 0.9 12.9 f 0.8
2.1 f 0.3
2.2 + 0.6
2.9 f 0.6
2.6 f 0.1
2.9 f 0.5
2.8 f 0.4
2.9 f 0.5
2.1 l 0.2*
2.0 f 0.4
2.3 f 0.6
Mn-SOD values were obtained when SOD activities were measured in the presence of I mM KCN, and
Cu,Zn-SOD was calculated as the difference of SOD activity values obtained in the absence (total activity)
minus that obtained in the presence of KCN. Enzymatic activity is indicated in Uimg, as mean f S.D.
for 3 independent determinations in two different cell-free extracts prepared for each sample. Asterisks
indicate statistical significance of differences between treatment and corresponding controls: *P < 0.05, **p < 0.01, l **p < 0.001.
3.2. SOD activity and formation of new SOD isoforms
Table 3 summarizes the speciIic activity of total, Cu,Zn- and Mn-SOD in fish in- jected with model xenobiotics. In response to Cu(II) and paraquat, significant varia- tions were observed in total SOD activity due to changes of Cu,Zn-SOD. Thus, 2 days after injection with the high Cu(I1) dose the Cu,Zn-SOD activity was
significantly increased, while a very significant decrease of such activity was observed in fish injected with paraquat. No significant changes were, in general, observed in fish exposed to apolar xenobiotics, with the only exceptions of malathion (high dose, 7 days) and Aroclor (low dose, 2 days) which diminished
significantly the Mn-SOD and Cu,Zn-SOD activities, respectively. The small changes observed in SOD activities prompted us to investigate whether
the oxidative stress indicated by the changes in mTBARS levels could be reflected in the possible appearance of new SOD isoforms. Fig. 1 shows the SOD bands in
cell-free extracts of fish exposed to the model xenobiotics. Extracts of control fish displayed 7 isoforms with pZ6.5, 6.4, 6.25, 6.15, 5.4, 5.2 and 5.1 (A and C, control).
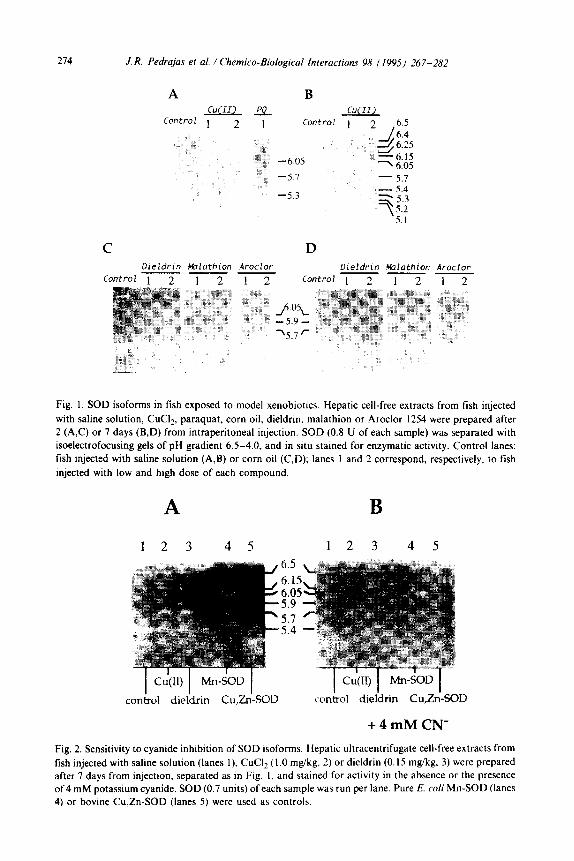
274 J. R. Pedrajas et al. / Chemico-Biological Interacrions 98 (1995) 267-282
A B CUCII) & CUCII)
Control , 2 , Control , * 6.5
&s &: J’ j
x -6.05 =; Z:E -5.7 - 5.7
-5.3 - 5.4
Fig. 1. SOD isoforms in fish exposed to model xenobiotics. Hepatic cell-free extracts from fish injected
with saline solution, CuClz, paraquat, corn oil, dieldrin. malathion or Aroclor 1254 were prepared after
2 (A,C) or 7 days (B.D) from intraperitoneal injection. SOD (0.8 U of each sample) was separated with
isoelectrofocusing gels of pH gradient 6.5-4.0, and in situ stained for enzymatic activity. Control lanes:
fish injected with saline solution (A,B) or corn oil (CD); lanes 1 and 2 correspond, respectively, to fish
injected with low and high dose of each compound.
A
123 45 123 45
6.5 6.15
E5 5.7 5.4
control dieldrin Cu,Zn-SOD
+4mMCN-
Fig. 2. Sensitivity to cyanide inhibition of SOD isoforms. Hepatic ultracentrifugate cell-free extracts from
fish injected with saline solution (lanes l), CuCI, (1 .O mg/kg. 2) or dieldrin (0. I5 mgkg, 3) were prepared after 7 days from injection, separated as in Fig. I. and stained for activity in the absence or the presence
of4 mM potassium cyanide. SOD (0.7 units) of each sample was run per lane. Pure E. co/i Mn-SOD (lanes
4) or bovine Cu,Zn-SOD (lanes 5) were used as controls.

0
A
n
k, ”
I
I r
I I
I I
1 2
3 4
time
(hou
rs)
lh
2h
4h
Fig.
3.
For
mat
ion
of T
BA
RS
and
new
SO
D
band
s up
on
incu
batio
n of
cel
l-fr
ee
extr
acts
w
ith
oxid
izin
g sp
ecie
s.
A c
ell-
free
ex
trac
t fr
om
fish
in
ject
ed
with
sa
line
solu
tion
was
in
cuba
ted
at
37°C
w
ith
0,
T o
r H
,Oz,
ge
nera
ted
by
the
xant
hine
-xan
thin
e ox
idas
e or
gl
ucos
e-gl
ucos
e ox
idas
e sy
stem
s,
resp
ectiv
ely.
or
w
ith
AD
P-Fe
pl
us
r-bu
tyl
hydr
oper
oxid
e at
tw
o co
ncen
trat
ions
. (A
) T
ime-
cour
se
of T
BA
RS
form
atio
n in
the
pr
esen
ce
of
AD
P-Fe
pl
us
I .5
mM
(@
or
0.
5 m
M
( l ) t
BuO
OH
, xa
nthi
ne
plus
xa
nthi
ne
oxid
ase
(0).
gl
ucos
e pl
us
gluc
ose
oxid
ase
(A)
or
buff
er
(0).
T
he
resu
lts
are
expr
esse
d as
mea
n va
lues
of
tw
o se
para
te
expe
rim
ents
, (B
) Is
oele
ctro
focu
sing
se
para
tion
(pH
6.
5-4.
0)
and
in s
itu
stai
ning
fo
r SO
D
activ
ity
of c
ell-
free
ex
trac
t in
cuba
ted
as i
ndic
ated
fo
r di
ffer
ent
exte
nts.

276 J. R. Pedrujas et al. / Chemico-Biological Interactions 98 11995) 267-282
After 2 and 7 days treatment, fish injected with the higher Cu(II) dose showed two new SOD forms (pZ 6.05 and 5.7) this last being quite visible on staining (A and B,
2). In contrast to the marked effect of Cu(I1) ions, only a faint new form of pl 5.3 was observed in paraquat-injected animals (A, PQ). The SOD band pattern changed also in fish injected with apolar xenobiotics. Two new SOD bands (~16.05 and 5.7) were observed in fish injected with dieldrin and malathion after 2 days with high
dose (Fig. lC, 2), with dieldrin after 7 days with low dose (D, dieldrin l), and those injected with corn oil after 7 days (D, control), while one additional new SOD form (PI 5.9) was observed after 7 days exposure with low dieldrin dose (D, dieldrin I).
Aroclor 1254 did not produce any new SOD band.
Fig. 2 shows the characterisation of the SOD isoforms of fish exposed to Cu(I1) and dieldrin, based on their different sensitivity to CN- inhibition. The complete inhibition by CN- of the two new SOD bands (p/ 6.05 and 5.7) confirmed that they corresponded to Cu,Zn-SOD, while the additional band (pl 5.9) seen in dieldrin-
exposed fish was assigned to Mn-SOD. Similarly, the bands of ~16.5 and 6.15 were only slightly inhibited by cyanide, thus corresponding to Mn-SOD.
3.3. Incubation of cell-free extracts with oxidizing systems
The putative oxidative origin of the new intermediate SOD bands was subsequent- ly studied. Cell-free extracts from control fish were incubated with continuous fluxes of 02: or HzOz, generated by the xanthine oxidase/xanthine or glucose oxi- dase/glucose systems, respectively [lo]. The effect of lipid peroxidation was also
tested by using the t-butylhydroperoxide/ADP-Fe(II1) system [23]. At different times of incubation the TBARS levels and the SOD band profile was determined (Fig. 3). In relation to the TBARS levels (Fig. 3A), as expected, the tBuOOH-ADP-Fe system
enhanced severely lipid peroxidation, with a IO-fold increase observed after 2 h in- cubation with 1.5 mM tBuOOH and a lower but continuous rise detected with 0.5 mM tBuOOH (3.4-fold after 4 h). In extracts continuously exposed to O? : or H20? the TBARS increased linearly to 2. I- and 1.8-fold, respectively, while in controls the
TBARS decreased slightly. The SOD activity was not altered upon incubation with the different oxidizing species (not shown). Fig. 3B shows the SOD isoform profile at different incubation times, folIowing IEF separation and in situ staining of activ- ity. Two new SOD bands were visible after 2 h incubation, especially with 1.5 mM
tBuOOH, in contrast with the TBARS level which was largely increased after just 1 h incubation under these conditions (Fig. 3A). After 4 h incubation with 1.5 mM tBuOOH four new intermediate bands were visible in extracts incubated with all oxi- dant systems. These new SOD bands, with pl values of 6.05. 5.9, 5.7 and 5.3, were
identical to those found in fish injected with different xenobiotics (Fig. 1).
3.4. Metallothionein-like proteins Changes in metallothionein-like proteins (MT) were also examined in fish exposed
to the above mentioned compounds. Fig. 4 shows the elution patterns of extracts from fish injected with saline solution, Cu(I1) or paraquat after chromatography in Sephadex G-75 and determination of protein, Zn and Cu. An important increase of low MW proteins with high Cu content, characteristics typical of MTs [30], was

J. R. Pedrajas et al. / Chemico-Biological Interactions 98 (1995) 267-282 277
1.5
1.0
0.5
0 0.0
5 2o 2.0
z 15 1.5 8 2 e I 1.0 51 10
2 5 0.5
0 0.0
20 2.0
E c 0 F-? 15 1.5
8 2 * I 10 1.0
P
9 5 0.5
0 0.0
E? ,a I e”
0 10 20
Fraction (6 ml)
30
Fig. 4. Metallothionein-like proteins in fish injected with polar xenobiotics. Hepatic cell-free extracts (5.5
ml) of fish after 2 days injection with saline solution, CuC12 {I.0 mgikg) or paraquat (50 m&kg) were loaded into a G-75 Sephadex column (2.5 x 33 cm) and eluted with IO mM Tris-HCI pH 8.0 at 10 ml/h.
Ultraviolet absorbance was determined at 230 nm and Zn and Cu were determined by atomic absorption
spectrometry.
observed after 2 days exposure to high Cu(II) dose. These results indicated a rise of MT-like proteins to bind the excess of this metal, although they did not fully protect the fish from oxidative stress, according to the changes observed in SOD isoforms and their increased content of soluble TBARS. The level of Zn-MT augmented also remarkably in fish injected with paraquat, in agreement with previous results in rats

278 J. R. Pedrajas et al. / Chemico-Biological fnreraciions 98 (1995) 267-282
[31]. None of the apolar xenobiotics tested, dieldrin, malathion or Aroclor, produc- ed significant changes in the MT-like proteins, although a clear increase was
observed in the levels of Cu and Zn bound to the high MW proteins, particularly in the case of dieldrin and malathion (results not shown).
4. Discussion
Thiobarbituric acid (TBA) is a well known reagent for determination of breakdown products of lipid peroxides. Malondialdehyde (MDA) is stable in micro- somes and the conjugate TBA-MDA can be used as an index of lipid peroxidation
and oxidative stress [5]. Cu,Zn-superoxide dismutase is oxidatively modified and in- activated by reactive oxygen species. New more negative and less active isoforms are generated when pure Cu,Zn-SOD is incubated with moderate H202 concentrations, in a time- and concentration-dependent process [ 10,l l] attributed to the generation
of hydroxyl radical in its active center [12]. Changes in SOD structure involve the oxidation of histidine-118 [32], one of the Cu ligands, and the subsequent release of Cu [10,33]. Cell-free extracts of fish (Mugil sp.) living in polluted sites show new Cu,Zn-SOD isoforms after isoelectrofocusing. When the more basic Cu,Zn-SOD
isoform previously purified was incubated with H202 also generated these new bands of more acidic PI, in parallel with some loss of activity. Thus, the new Cu,Zn- SOD isoforms of intermediate pl were proposed as useful biomarkers for en- vironmental pollution [ 16,341.
Soluble Cu(I1) ions catalyse the generation of the highly reactive hydroxyl radical [l]. In addition, Cu(I1) binds to metallothioneins and to apo-SOD, increasing the activity of this enzyme. Thus, Cu-deficient rats have only 50% of the SOD activity
of controls, but similar apo-SOD levels [35]. In consequence, the higher Cu,Zn-SOD activity of Cu-injected fish could be explained by increased binding of this metal to apo-SOD. Nevertheless, the higher levels of soluble TBARS found in these animals suggest that they were subjected to an oxidative stress, explaining the formation of
the new Cu,Zn-SOD isoforms by oxidative attack [ 161, in agreement with the highly oxidized glutathione redox status found in such animals [36].
The bipyridyl herbicide paraquat produces its toxic effects generating 02: and other reactive oxygen species by redox cycling [ 19,371, and SOD attenuates paraquat
toxicity [38]. Thus, this compound has been extensively used to generate oxidative stress both in vivo and in vitro [19]. The 5-fold increase of soluble TBARS in paraquat-injected fish indicate oxidative stress, while SOD activity decreased
significantly. Nevertheless, paraquat only promoted the appearance of a faint new Cu,Zn-SOD isoform, and did not significantly change the levels of mTBARS, in agreement with the polar nature and the lack of effect of paraquat on lipid peroxida- tion previously reported by other authors [39]. It should be noticed also that the in
vivo generation of hydroxyl radical by ESR spin trapping technique in rats treated with Cu(I1) plus paraquat has been described, while no such radicals were detected when Cu(II) or paraquat were excluded [40]. The marked increase in metallothionein-bound Zn observed in paraquat injected fish agrees with the induc- tion of hepatic metallothionein by paraquat in the mouse [31].

J. R. Pedrajas et al. / Chemico-Biological Interactions 98 (I 995) 26 7-282 279
When injected in Sparus aurata, dieldrin and malathion promoted similar changes. Dieldrin is an organochlorine cyclodiene epoxide similar to its stereoisomer endrin, whose toxicity is mediated by reactive oxygen species. It has been reported that lipid peroxidation, cytochrome c reduction, membrane fluidity and oxidative DNA damage are increased in peritoneal macrophages and hepatic mitochondria and mi- crosomes of endrin-exposed rats [20,21]. Dieldrin was the xenobiotic producing the most marked effects in Sparus auruta, thus suggesting that it could act through generation of reactive oxygen species. On the other hand, it has been also reported that the brain and heart of rats poisoned with malathion undergo lipid peroxidation, in a process associated with hypothermia [29], a phenomenon closely associated with oxidative damage [41]. In consequence, malathion could also have induced in fish oxidative damages, thus explaining the formation of new SOD isoforms and the in- creased microsomal lipid peroxidation. The increased mTBARS observed with low dieldrin or malathion doses after 2 days exposure (not paralleled with new SOD bands) decreased below control values after 7 days. This could be explained by bio- transformation of these two xenobiotics or by the repair of the oxidative damages induced in membrane lipids.
Based on the peroxidase activity displayed by microsomal glutathione-S- transferase it was proposed that the physiological role of such an enzyme could be to protect membranes from lipid peroxidation [6]. Evidence has been provided recently that rat liver mGST is indeed responsible for GSH-dependent protection from lipid peroxidation [7]. The inverse relationship found between mGST activity and mTBARS, with decreased mGST activity paralleling higher mTBARS levels and vice versa confirms this idea. Alternatively, mGST decreases could be explained by the membrane damages accompanying the increased mTBARS levels.
Aroclor 1254 is a mixture of PCBs acting as a general inducer of different cytochrome P450 isoenzymes, and this increase subsequently produces reactive oxy- gen species. It is known that Aroclor induces lipid peroxidation, but only if ad- ministered with the diet. Thus, stimulation of lipid peroxidation has been observed in Atlantic croaker fed with Aroclor 1254, while no lipid peroxidation has been described in fish or other animals injected with this PCB mixture [14]. This could explain the lack of oxidative stress signs obtained in our experiment.
It should be noticed that corn oil, the solvent used for apolar model xenobiotics, produced effects similar to those of dieldrin or malathion, although to a lesser ex- tent. Thus, after 7 days exposure corn oil promoted the appearance of the new Cu,Zn-SOD isoforms, increased mTBARS levels and decreased mGST activity. Corn oil contains a high proportion of polyunsaturated fatty acids, which activate the peroxisomal fatty acid oxidation system and lead to oxidative stress [42]. Al- though corn oil has been commonly used as a vehicle for xenobiotic injection in many studies, our results suggest that its use should be discouraged, in agreement with other authors reporting appreciable amounts of mTBARS in control animals injected with this solvent, among other indicators of oxidative stress [21].
The oxidative origin of the new SOD bands was confirmed by incubation of cell free extracts with different systems generating reactive oxygen species, O2 7, HzOz or tBuOOH/ADP-Fe. The Fe- or Cu-catalyzed reaction of O2 ; and HIOZ, leading

280 J.R. Pedrajas et al. / Chemico-Biological Inleractions 98 (1995) 267-282
to hydroxyl radical formation [I], occurs also at the active center of Cu,Zn-SOD [12] and oxidizes the enzyme [ 10, 1 1,161, although the new more acidic SOD forms remain
still active [ 11,161. The in vitro generation of new active SOD bands of intermediate pl by incubation with O2 T and H202 confirms their oxidative origin. Iron could have a direct role in lipid peroxidation mediated by the tBuOOH/ADP-Fe system, and the direct participation of hydroxyl radical has been recently questioned [23,43].
The oxidative conditions reproduced with crude extracts could occur also in vivo in fish injected with Cu(II), dieldrin, malathion, paraquat or corn oil. In conclusion, we propose the new Cu,Zn-SOD isoforms of intermediate pl observed in fish living in contaminated environments or exposed under controlled conditions to several
model xenobiotics should be considered as a useful early biomarker of exposure to transition metals or organic xenobiotics known to generate oxidative stress.
Acknowledgements
This work was subsidized by grants from CICYT AMB-628193, Plan Andaluz de Investigation (Group 3106) and Consejeria de Agricultura y Pesca, Junta de An- dalucia (Spain). Controlled exposure experiments were carried out at the CICEM ‘Agua de1 Pino’, Consejeria de Agricultura y Pesca, Junta de Andalucia, Aptdo. 104,
21071 Huelva (Spain). We also acknowledge Antonio Rodriguez-Ariza for measur- ing microsomal glutathione transferase activities.
References
[II 121
131
141
151
I61
[71
181 191
1101
1111
H. Sies, Biochemistry of oxidative stress, Angew. Chem. Int. Ed., 25 (1986) 1058-1071.
J. Lopez-Barea, Biomarkers in ecotoxicolgy: an overview, in: G.H. Dcgen, J.P. Seiler and P. Bentley
(Eds.), Archives of Toxicology suppl. 17. Springer. Berlin, 1994, pp. 57-79.
H. Esterbauer and H. Zollner, Methods for determination of aldehydic lipid peroxidation, Free
Rad. Biol. Med., 7 (1989) 197-203.
J.A. Buege and SD. Aust, Microsomal lipid peroxidation, m: S. Fleischer and L. Packer (Eds.),
Methods in Enzymology, Vol 52, Academic Press. New York, 1978, pp. 302-310.
H.H. Draper, E.J. Squires, H. Mahmoodi, S. Wu, S. Agarwal and M. Hadley, A comparative evalu-
ation of thiobarbituric acid methods for the determination of malondialdehyde in biological materi-
als, Free Rad. Biol. Med., 15 (1993) 353-363.
E. Mosialou and R. Morgenstern. Activity of rat liver microsomal glutathione transferase towards
products of lipid peroxidation and studies of the effect of inhibitors on glutathione-dependent pro-
tection against lipid peroxidation. Arch. Biochem. Biophys.. 275 (1989) 289-294.
E. Mosialou. G. Ekstrom, A.E.P. Adaud and R. Morgenstern, Evidence that rat liver microsomal
glutathione transferase is responsible for glutathione dependent protection against lipid peroxida-
tion. B&hem. Pharmacol., 45 (1993) 1645-1651.
I. Fridovich. Superoxide dismutases. J. Biol. Chem.. 264 (1989) 7761-7764.
S.L. Marklund. Extracellular superoxide dismutase and other superoxide dismutase isoenzymes in
tissues from nine mammalian species, Biochem. J.. 222 (1984) 649-655.
D.C. Sale, R.E. Pacitici, SW. Lin. C. Giulivi and K.J.A. Davies, Superoxide dismutase undergoes
proteolysis and fragmentation following oxidative modification and inactivation. J. Biol. Chem.. 265 (1990) 11919-l 1927.
B.P. Sharonov and I.V. Churilova. Inactivation and oxidative modification of Cu.Zn superoxtde
dismutase by stimulated neutrophils: the appearance of new catalytically active structures,
Biochem. Biophys. Res. Commun.. 189 (1992) 1129- 1135.

J. R. Pedrajas et al. / Chemico-Biological Interactions 98 (199.5) 267-282 281
[12] E.K. Hodgson and I. Fridovich, The interaction of bovine erythrocyte superoxide dismutase with
hydrogen peroxide: inactivation of the enzyme, Biochemistry, 14 (1975) 5294-5299.
[13] A. Rodriguez-Ariza, J. Peinado, C. Pt,teyo and J. Lopez-Barea, Biochemical indicators of oxidative
stress in fish from polluted littoral areas, Can. J. Fish. Aquat. Sci., 50 (1993) 1-6. [I41 H.W. Wofford and P. Thomas, Effect of xenobiotics on peroxidation of hepatic microsomal lipids
from striped mullet (Mugil cephalus) and Atlantic croaker (Micropogonias undulafus), Mar. En-
viron. Res., 24 (1988) 285-289.
[I51 G.W. Winston and R.T. di Giulio, Prooxidant and antioxidant mechanisms in aquatic organisms.
Aquat. Toxicol., 19 (1991) 137-161.
[16] J.R. Pedrajas, J. Peinado and J. Lopez-Barea, Purification of Cu,Zn-superoxide dismutase isoen-
zymes from fish liver: appearance of new isoforms as consequence of pollution, Free Rad. Res.
Commun., 19 (1993) 29-41.
[17] J.H.R. Kagi and Y. Kojima, Chemistry and biochemistry of metallothioneins, in: J.H.R. Kagi and
Y. Kojima (Eds.), Metallothionein II, Birkhauser-Verlag, Basel, 1987, pp. 25-52.
[18] P. Kille, J. Kay, M. Leaver and S. George, Induction of piscine metallothionein as a primary re-
sponse to heavy metal pollutants: applicability of new sensitive molecular probes, Aquat. Toxicol.,
22 (1992) 279-286.
[l9] J.S. Bus, J.E. Gibson, Paraquat: model for oxidant-initiated toxicity, Environ. Health Perspect., 55
(1984) 37-46. [20] M. Bagchi and S.J. Stohs, In vitro induction of reactive species by 2,3,7,8-tetrachlorodibenzo-p-
dioxin, endrin, and lindane in rat peritoneal macrophages, and hepatic mitochondria and micro-
somes, Free Rad. Biol. Med., 14 (1993) 1 I-18.
[21] M. Bagchi, E.A. Hassoun, D. Bagchi and S.J. Stohs, Production of reactive oxygen species by
peritoneal macrophages and hepatic mitochondria and microsomes from endrin-treated rats, Free
Rad. Biol. Med., 14 (1993) 149-155.
[22] L.M. Bomheim, A.N. Kotake and M.A. Correia, Differential haemin-mediated restoration of
cytochrome P450 N-demethylases after inactivation by allylisopropylacetamide, Biochem. J., 227
(1985) 277-286.
[23] G. Minotti, rerr-Butyl hydroperoxide-dependent microsomal release of iron and lipid peroxidation.
Arch. B&hem. Biophys., 273 (1989) 144-147.
[24] J.M. McCord and I. Fridovich, Superoxide dismutase: an enzymic function for erythrocuprein, J.
Biol. Chem., 244 (1969) 6049-6055.
[25] A. Rodriguez-Ariza, N. Abril, J.I. Navas, G. Dorado, J. Lopez-Barea and C. Pueyo. Metal, muta-
genicity, and biochemical studies on bivalve molluscs from Spanish coasts, Environ. Mol. Muta- gen., 19 (1992) 112-124.
[26] O.H. Lowry, M.J. Rosebrough, A.L. Farr and R.J. Randall, Protein measurement with the Folin
phenol reagents, J. Biol. Chem., 193 (1951) 265-275.
[27] C.O. Beauchamp and I. Fridovich, Superoxide dismutase improved assays and an assay applicable
to acrylamide gels, Anal. Biochem., 44 (1971) 276-287.
1281 A. Goksoyr and L. Fiirlin, The cytochrome P450 system in fish aquatic toxicology and environmen-
tal monitoring, Aquat. Toxicol., 22 (1992) 287-312.
1291 V.A. Myshkin, A.F. Vakarina, S.A. Bashkatov, G.A. Sofronov and D.A. Enikeev, Postintoxication
hypothermia and free-radical lipid oxidation in the brain and heart of rats poisoned with
organophosphorous cholinesterase inhibitors, Bull. Exp. Biol. Med., I I3 (1992) 664-665.
[30] Y. Kojima, Definitions and nomenclature of metallothioneins, in: J.F. Riordan and B.L. Vallee
(Eds.), Methods in Enzymology, Vol 205, Academic Press, San Diego, 1991, pp. 8-10.
[3l] J.W. Bauman, C. Madhu, J.M. McKim, Y. Liu and C.D. Klaasen, Induction of hepatic metallo-
thionein by paraquat, Toxicol. Appl. Pharmacol., I I7 (1992) 233-241.
1321 K. Uchida and S. Kawakishi, Identification of oxidized histidine generated at the active-site of
Cu,Zn-superoxide dismutase exposed to H,O, - Selective generation of 2-oxo-histidine at the
histidine-I 18, J. Biol. Chem., 269 (1994) 2405-2410.
1331 K. Sato, T. Akaike, M. Kohno, M. Ando, H. Maeda, Hydroxyl radical production by HzO, plus
Cu,Zn-superoxide dismutase reflects the activity of free copper released from the oxidatively
damaged protein, J. Biol. Chem., 267 (1992) 25371-25377.

282 J.R. Pedrajas et al. / Chemico-Biological Interacbons 98 (1995) 267-282
I341
[351
1361
1371
f381
[391
1401
[4ll
1421
[431
A. Rodriguez-Ariza, E. Martinez-Lara, P. Pascual, J.R. Pedrajas, N. Abril, G. Dorado, F. Toribio,
J.A. Barcena, J. Peinado, C. Pueyo, J. Lopez-Barea, Biochemical and genetic indices of marine pol-
lution in Spanish littoral, Sci. Total Environ,, Suppl. 1993 (1993) 109-I 16.
K. Chung, N. Romero, D. Tinker, C.L. Keen, K. Amemiya and R. Rucker. Role of copper in the
regulation and accumulation of superoxide dismutase and metaliothionein in rat liver, J. Nutr.. I I8
(1988) 859-864.
A. Rodriguez-Ariza, F. Toribio and J. Lopez-Barea. Rapid determination of glutathione status in
fish liver using HPLC and electrochemical detection, J. Chromatogr. Biomed., 656 (1994) 3 I I-318.
L.-L. Smith, M.-S. Rose and I. Wiatt. The pathology and biochemistry of paraquat, in: Oxygen
Free Radicals and Tissue Damage, CIBA Foundation Series, 65, CIBA Foundation, London, 1979,
pp. 321-341.
A.P. Autor, Reduction of paraquat toxicity by superoxide dismutase. Life Sci.. 14 (1974)
1309-1319.
C. Steffen and K.J. Netter, On the mechanism of paraquat action on microsomal oxygen reduction
and its relation to lipid peroxidation, Toxicol. Appl. Pharmacol., 47 (1979) 593-602.
R.P. Mason, P.M. Hanna, M.B. Kadiiska. In vivo ESR spin trapping evidence for hydroxyl radical
mediated toxicity of paraquat and copper in rats. Free Rad. Biol. Med., 15 (1993) 480.
D.P. Jones, The role of oxygen concentration in oxidative stress: hypoxic and hyperoxic models,
in: H. Sies (Ed.). Oxidative Stress, Academic Press, London, 1985, pp. I51 -193.
H. Keller, C. Dreyer, J. Medin, A. Mahfoudi, K. Ozato and W. Wahli, Fatty acids and retinoids
control lipid metabolism through activation of peroxisome proliferator-activated receptor-retinoid
X receptor heterodimers. Proc. Natl. Acad. Sci. USA, 90 (1993) 2160-2164.
D.M. Miller, T.A. Grover, N. Nayini and SD. Aust, Xanthine oxidase- and iron-dependent lipid
peroxidation, Arch. B&hem. Biophys.. 301 (1993) 1-7.




















