
Oxalate contributes to the resistance of Gaillardia grandiflora and Lupinus sericeus to a phytotoxin...
11
ORIGINAL ARTICLE Tiffany L. Weir Harsh Pal Bais Valerie J. Stull Ragan M. Callaway Giles C. Thelen Wendy M. Ridenour Suresh Bhamidi Frank R. Stermitz Jorge M. Vivanco Oxalate contributes to the resistance of Gaillardia grandiflora and Lupinus sericeus to a phytotoxin produced by Centaurea maculosa Received: 21 October 2005 / Accepted: 16 November 2005 Ó Springer-Verlag 2005 Abstract Centaurea maculosa Lam. is a noxious weed in western North America that produces a phytotoxin, (±)-catechin, which is thought to contribute to its invasiveness. Areas invaded by C. maculosa often result in monocultures of the weed, however; in some areas, North American natives stand their ground against C. maculosa and show varying degrees of resistance to its phytotoxin. Two of these resistant native species, Lupinus sericeus Pursh and Gaillardia grandiflora Van Houtte, were found to secrete increased amounts of oxalate in response to catechin exposure. Mechanisti- cally, we found that oxalate works exogenously by blocking generation of reactive oxygen species in sus- ceptible plants and reducing oxidative damage gener- ated in response to catechin. Furthermore, field experiments show that L. sericeus indirectly facilitates native grasses in grasslands invaded by C. maculosa, and this facilitation can be correlated with the presence of oxalate in soil. Addition of exogenous oxalate to native grasses and Arabidopsis thaliana (L.) Heynh grown in vitro alleviated the phytotoxic effects of cat- echin, supporting the field experiments and suggesting that root-secreted oxalate may also act as a chemical facilitator for plant species that do not secrete the compound. Keywords Allelopathy Catechin Centaurea Oxalate Resistance Abbreviations HPLC: High pressure liquid chromatography ROS: Reactive oxygen species Introduction Centaurea maculosa Lam., spotted knapweed, is one of the most destructive invasive weeds in western North America. The invasiveness of this plant is due, in part, to a root-secreted phytotoxin, (±)-catechin (Bais et al. 2002, 2003). In its native Eurasian habitat, C. maculosa is a relatively minor member of diverse native grassland communities, but in North America, this species can create monocultures by excluding almost all native species (Callaway et al. 1999). Furthermore, the phy- totoxic effect of C. maculosa (and that of the related C. diffusa) is much stronger against grass species from North America than closely related grass species in Europe (Callaway and Aschehoug 2000; Bais et al. 2003). This suggests the evolution of traits that bestow tolerance to the chemical effects of these Centaurea spp. However, not all North American natives are equally susceptible to the competitive effects of C. maculosa (Callaway et al. 2004). Gaillardia spp. and Lupinus sericeus Pursh are good competitors against T. L. Weir J. M. Vivanco (&) Department of Horticulture and Landscape Architecture, Graduate Program in Cell and Molecular Biology, Colorado State University, 1173 Horticulture and Landscape Architecture, Fort Collins, CO 80523, USA E-mail: [email protected] Tel.: +1-970-4917170 Fax: +1-970-4917745 V. J. Stull T. L. Weir J. M. Vivanco Center for Rhizosphere Biology, Colorado State University, Fort Collins, CO 80523, USA S. Bhamidi Department of Microbiology, Immunology, and Pathology, Colorado State University, Fort Collins, CO 80523, USA F. R. Stermitz V. J. Stull Department of Chemistry, Colorado State University, Fort Collins, CO 80523, USA H. P. Bais Department of Plant and Soil, Delaware Biotechnology Institute, University of Delaware, Newark, DE 19702, USA R. M. Callaway G. C. Thelen W. M. Ridenour The Division of Biological Sciences, The University of Montana, Missoula, MT 59812, USA Planta (2005) DOI 10.1007/s00425-005-0192-x
Transcript of Oxalate contributes to the resistance of Gaillardia grandiflora and Lupinus sericeus to a phytotoxin...

ORIGINAL ARTICLE
Tiffany L. Weir Æ Harsh Pal Bais Æ Valerie J. Stull
Ragan M. Callaway Æ Giles C. Thelen
Wendy M. Ridenour Æ Suresh Bhamidi
Frank R. Stermitz Æ Jorge M. Vivanco
Oxalate contributes to the resistance of Gaillardia grandifloraand Lupinus sericeus to a phytotoxin produced by Centaurea maculosa
Received: 21 October 2005 / Accepted: 16 November 2005� Springer-Verlag 2005
Abstract Centaurea maculosa Lam. is a noxious weed inwestern North America that produces a phytotoxin,(±)-catechin, which is thought to contribute to itsinvasiveness. Areas invaded by C. maculosa often resultin monocultures of the weed, however; in some areas,North American natives stand their ground against C.maculosa and show varying degrees of resistance to itsphytotoxin. Two of these resistant native species,Lupinus sericeus Pursh and Gaillardia grandiflora VanHoutte, were found to secrete increased amounts ofoxalate in response to catechin exposure. Mechanisti-cally, we found that oxalate works exogenously byblocking generation of reactive oxygen species in sus-ceptible plants and reducing oxidative damage gener-ated in response to catechin. Furthermore, field
experiments show that L. sericeus indirectly facilitatesnative grasses in grasslands invaded by C. maculosa,and this facilitation can be correlated with the presenceof oxalate in soil. Addition of exogenous oxalate tonative grasses and Arabidopsis thaliana (L.) Heynhgrown in vitro alleviated the phytotoxic effects of cat-echin, supporting the field experiments and suggestingthat root-secreted oxalate may also act as a chemicalfacilitator for plant species that do not secrete thecompound.
Keywords Allelopathy Æ Catechin Æ Centaurea ÆOxalate Æ Resistance
Abbreviations HPLC: High pressure liquidchromatography Æ ROS: Reactive oxygen species
Introduction
Centaurea maculosa Lam., spotted knapweed, is one ofthe most destructive invasive weeds in western NorthAmerica. The invasiveness of this plant is due, in part,to a root-secreted phytotoxin, (±)-catechin (Bais et al.2002, 2003). In its native Eurasian habitat, C. maculosais a relatively minor member of diverse native grasslandcommunities, but in North America, this species cancreate monocultures by excluding almost all nativespecies (Callaway et al. 1999). Furthermore, the phy-totoxic effect of C. maculosa (and that of the relatedC. diffusa) is much stronger against grass species fromNorth America than closely related grass species inEurope (Callaway and Aschehoug 2000; Bais et al.2003). This suggests the evolution of traits that bestowtolerance to the chemical effects of these Centaureaspp. However, not all North American natives areequally susceptible to the competitive effects ofC. maculosa (Callaway et al. 2004). Gaillardia spp. andLupinus sericeus Pursh are good competitors against
T. L. Weir Æ J. M. Vivanco (&)Department of Horticulture and Landscape Architecture,Graduate Program in Cell and Molecular Biology,Colorado State University, 1173 Horticulture and LandscapeArchitecture, Fort Collins, CO 80523, USAE-mail: [email protected].: +1-970-4917170Fax: +1-970-4917745
V. J. Stull Æ T. L. Weir Æ J. M. VivancoCenter for Rhizosphere Biology, Colorado State University,Fort Collins, CO 80523, USA
S. BhamidiDepartment of Microbiology, Immunology, and Pathology,Colorado State University, Fort Collins, CO 80523, USA
F. R. Stermitz Æ V. J. StullDepartment of Chemistry, Colorado State University,Fort Collins, CO 80523, USA
H. P. BaisDepartment of Plant and Soil, Delaware Biotechnology Institute,University of Delaware, Newark, DE 19702, USA
R. M. Callaway Æ G. C. Thelen Æ W. M. RidenourThe Division of Biological Sciences, The University of Montana,Missoula, MT 59812, USA
Planta (2005)DOI 10.1007/s00425-005-0192-x

C. maculosa, and are less affected by the phytotoxic ef-fects of catechin (Weir et al. 2003; Thelen et al. 2005;Perry et al. 2005), suggesting that some North Americannatives may possess general defenses to catechin toxicity.A characteristic effect of catechin toxicity is the pro-duction of a wave of reactive oxygen species (ROS),followed by an increase in cytoplasmic Ca2+ and acidi-fication of the cytoplasm, leading to cell death whencatechin is added exogenously to the roots of susceptibleplants (Bais et al. 2003). However, this deadly cascade isblocked, and cell death prevented, when the antioxidantascorbate is added simultaneously (Bais et al. 2003). Herewe provide evidence that some catechin-resistant speciesnaturally mimic this mechanism by launching a moreeffective antioxidant response, thereby counteracting theeffects of the phytotoxin.
The root-mediated secretion of organic acids is acommon response to a number of environmental andbiological conditions. Secretion of citrate and malateincreases the mobilization of sparingly soluble phos-phate (P) in the rhizosphere (Gardner et al. 1983;Keerthisinghe et al. 1998; Neumann et al. 1999). En-hanced secretion of citrate, malate, and oxalate has beenimplicated in conferring aluminum (Al) tolerance to avariety of plants through exclusion of Al from root tips(Ryan et al. 1995; Ma et al. 1997; Zheng et al. 1998), andmore recently in conferring Pb resistance to rice seed-lings (Yang et al. 2000). Therefore we analyzed the rootexudates of catechin-resistant plants L. sericeus andGaillardia grandiflora Van Houtte, as well as Arabidopsisthaliana, Festuca idahoensis Elmer, and Pseudoroegnariaspicata Pursh, species shown to be susceptible toC. maculosa’s root exudates, to ascertain if root-secretedorganic acids conferred resistance to catechin.
Materials and methods
Plant culture and growth conditions
Seeds of wild-type A. thaliana (L.) Heynh. ecotypeColumbia (Col-0) were obtained from Lehle Seeds(Round Rock, TX). Seeds of G. grandifloraVan Houttewere obtained from Botanical Interests (Broomfield,CO), and L. sericeus Pursh, F. idahoensis Elmer, andP. spicata Pursh seeds were purchased from Wind RiverSeed (Manderson, WY). C. maculosa Lam. seeds werecollected by Dr. Ragan Callaway (University of Mon-tana) from wild populations in Missoula, Montana.Seeds were washed in running tap water and surfacesterilized using sodium hypochlorite (0.3% v/v) for30 min, followed by 3–4 washes in sterile distilled water.Surface sterilized seeds were germinated on solid Mu-rashige and Skoog (Murashige and Skoog 1962) basalmedium in petri dishes. The temperature within thegrowth chamber was 25±2�C and light intensity was4.41 J m�2 s�1 , with a 16 h light, 8 h dark photoperiod.Unless specified, liquid bioassays were performed using10-day-old seedlings transferred to sterile 12-well plates
with 2 ml of liquid MS basal media. Plants were thenplaced at 25±2�C on an orbital platform shaker set at90 rpm and exposed to 16 h daily of 4.41 J m�2 s�1 lightintensity for further bioassays.
Root exudate assays
Gaillardia root exudate assay
Twenty-one-day-old plants of G. grandiflora were grownin 250-ml flasks containing 50 ml of liquid MS mediumand treated with 200 lg ml�1 of (±)-catechin (Sigma,St. Louis, MO, USA; Shivambu International, Hima-chel Pradesh, India) dissolved in 10% MeOH, a cate-chin concentration previously shown to have littlephytotoxic effect on Gaillardia spp. (Weir et al. 2003).After 1 week, medium containing catechin and G.grandiflora root exudates was removed, filter sterilizedwith a 0.22 lm filter, and 2 ml were added to 12-wellplates. Five-day-old seedlings of A. thaliana were grownin the collected medium, MS medium treated with theappropriate amount of High pressure liquid chroma-tography (HPLC) grade MeOH (Fisher Scientific, FairLawn, NJ, USA), or fresh MS medium amended with200 lg ml�1 of catechin dissolved in 10% MeOH; acatechin concentration sufficient to result in phytotox-icity to A. thaliana (Bais et al. 2003). After 7 days,plants were examined for mortality by staining the rootsfor 15 min with 0.1% (w/v) lactophenol tryphan blue(Sigma, St. Louis, MO, USA), which marks dead cellswith a blue color. Stained roots were examined at 20·and 40· magnifications under an Olympus BX60microscope equipped with Cool Snap imaging software(MediaCybernetics, San Diego, CA).
Bromocresol purple assay
Variations in pH around roots of G. grandiflora, andA. thaliana were visualized in MS basal medium con-taining 90 mg l�1 of bromocresol purple (Sigma, St.Louis, MO, USA; pK=6.4) and adjusted to pH 6.3.Prepared medium was cooled to 38�C and poured into15-mm petri dishes, root systems were pressed onto asolidified medium and placed in a growth chamber andinspected and photographed over 48 h to determine thesecretion of acidic compounds. To determine if catechininduced the secretion of acidic compounds, this proce-dure was repeated on plants with a filter paper disksaturated with 200 lg ml�1 of catechin placed at the tipof the primary root.
Chemical extraction and HPLC analysis
Root exudates of G. grandiflora, L. sericeus, A. thaliana,F. idahoensis, Pseudoroegnaia spicata (non-elicited andelicited with 200 lg ml�1 catechin) were collected by

growing ten plants of each species in 200 ml of MSmedium. Collected media were extracted as described inYang et al. (2000) by passing through an octadecylsilanecartridge column (Sep-Pak C18, Waters, Milford, MA,USA) and eluting with 50% (v/v) MeOH. Samples wereconcentrated by drying and resuspended in sterile dis-tilled water. Organic acids were quantified by HPLCanalysis as described by Coppola and Starr (1986). Or-ganic acids were separated using two reverse phase C18
columns in tandem (Supelco, State College, PA, USA)with UV detection at 210 nm (Dionex Summit System,Sunnyvale, CA, HPLC operating conditions were asfollows: flow rate 0.8 ml min�1; 214 nm referencewavelength; ambient temperature; sensitivity 0.1 AUFS;mobile phase was 0.2 M KH2PO4, pH 2.4; gradient wasisocratic. Soil samples were extracted for organic acidsusing 50% (v/v) MeOH, concentrated and resuspendedin sterile distilled water and analyzed by HPLC as de-scribed above. Compound identity was confirmed usingnegative ion electron spray mass spectrometry (FisonsVG Quattro, Suffolk, UK). Mass and fragmentationpattern were compared with an oxalate standard. Cate-chin samples were extracted from soil and lyophilizedmedia using 100% MeOH and quantified using HPLCas described by Bais et al. (2002). All organic solventswere of HPLC grade (Fisher Scientific, Fair Lawn, NJ,USA).
Organic acid assays
Addition of oxalate to susceptible plant species
A. thaliana plants were cultured as previously described.Plants were then treated with 10, 20, 50, 100, or 200 lMoxalate (Sigma, St. Louis, MO, USA), these same con-centrations of oxalate and 100 lg ml�1 of catechin,100 lg ml�1 of catechin alone, or 100 lg ml�1 of 10%MeOH (control). After 7 days, the plants were visuallyexamined to determine mortality and weighed to obtaintotal biomass (g). This assay was repeated using A. tha-liana, P. spicata, and F. idahoensis treated with 100 lMof oxalate and 100 lg ml�1 of catechin, 100 lg ml�1 ofcatechin alone, and 100 lg ml�1 of 10% MeOH.
Prevention of oxalate secretion from G. grandiflora
In order to further clarify the role of oxalate in pro-tecting plants from catechin, a resistant plant, G. gran-diflora was treated with 400 lg ml�1 of catechin alongwith 1 mM sodium orthovanadate (Na3VO4), an anionchannel blocker that inhibits the natural release ofoxalate from the plant roots (Yan et al. 2002). Controlplants were treated with 1 mM of Na3VO4, 400 lg ml�1
of catechin, or 100 lg ml�1 of 10% MeOH. Plants wereexamined for mortality and total biomass (g) wasobtained.
Antioxidant assays
Visualization of oxygen radicals
Seedlings of A. thaliana were treated as described aboveand stained with 0.1% (w/v) nitroblue tetrazolium dye(Sigma, St. Louis, MO, USA) for 1 h. Roots weremounted on slides and examined at 40· magnificationunder an Olympus BX60 microscope equipped withCool Snap imaging software (MediaCybernetics, SanDiego, CA, USA). Leaves were destained for 30 minwith 100% ethanol and photographed by using a Sonydigital camera.
Lipid peroxidation analysis
To determine if A. thaliana plants suffered membranedamage caused by the generation of free radicals in re-sponse to catechin treatment, we quantified the presenceof conjugated dienes, a product of lipid peroxidation, byadapting the method described by Recknagel andGlende (1984). Briefly, approximately 200 mg of plantmaterial subjected to treatment with 100 lg ml�1 ofracemic catechin, 100 lM of oxalate, or both com-pounds was homogenized in 1 ml of H2O and 4 ml ofchloroform:methanol (2:1 v/v) was added. Samples wereplaced on ice for 30 min and centrifuged for 5 min at260g. The organic layer was transferred to a clean tubeand the chloroform was evaporated off in a 50�C waterbath. Lipids were dissolved in 2 ml of cyclohexane andthe optical density at 230 nm was recorded.
Field experiments
We grew the native grasses P. spicata and F. idahoensisfrom local seed for 5 weeks in the greenhouse and thentransplanted these seedlings into the field on April 25,1996. Fifteen individuals of each species were planted10 cm from a mature L. sericeus plant growing in denseC. maculosa cover and 15 more were planted 10 cm froma mature L. sericeus plant with no C. maculosa within2 m. As controls we planted 15 individuals of eachspecies without L. sericeus in dense C. maculosa coverand 15 more without L. sericeus but with no C. maculosawithin 2 m. The aboveground tissues of the grasses wereharvested on June 17, 1996, dried at 60�C and weighed.
Statistical analysis
All statistical analyses of in vitro experiments wereperformed using XLStat Pro 7.5.2 (Addinsoft, Paris,France). Analysis of variance (ANOVA) was performedfor all experiments. Significant differences betweentreatment means were determined using Tukey’s HSD ata 95% confidence interval, and significant differencesbetween treatments and controls were determined using

Dunnet’s two-tailed test set at a 95% confidence inter-val. All experiments were performed in duplicate withthe exception of root exudate analyses, which wereperformed in triplicate. The abundances of P. spicataand F. idahoensis with and without L. sericeus in patchesof C. maculosa were tested with independent sample ttests. The biomasses of P. spicata and F. idahoensis (withand without L. sericeus, and in or out of C. maculosapatches) or not were tested with a two-way ANOVAwith +L. sericeus and �L. sericeus and +C. maculosaand �C. maculosa as fixed factors.
Results
Root exudate assays
Several Gaillardia sp. have demonstrated resistance toC. maculosa’s phytotoxic effects in the field (Callawayet al. 2004) and to direct challenge by its phytotoxin,(±)-catechin (Weir et al. 2003; Perry et al. 2005). Todetermine if a root-secreted compound contributed to G.grandiflora’s ability to resist catechin, we collected Mu-rashige and Skoog (1962) growth medium (MS) that hadbeen conditioned by G. grandiflora plants for 1 week.This conditioned medium containing G. grandiflora rootexudates and supplemented with 200 lg ml�1 of (±)-catechin was used to grow 5-day-old A. thaliana plants.Control plants were grown in MS liquid medium andwere treated with 200 lg ml�1 of (±)-catechin ormethanol (10% v/v). Seven days post-treatment, roots ofcontrol plants treated with 200 lg ml�1 of (±)-catechinhad succumbed to the toxin but those grown in themedium containing G. grandiflora root exudates sur-vived. Root mortality was confirmed using the vitalstain, lactophenol tryphan blue (0.1% w/v) (Fig. 1a–d).These results indicated that a component of the rootexudates from G. grandiflora conferred resistance toA. thaliana.
Bais et al. (2003) reported data indicating that theorganic acid, ascorbate, was capable of blocking theseries of events that lead to catechin-induced cell deathwhen applied exogenously to A. thaliana roots. Fur-thermore, increased production of lateral roots, a phe-notype also seen in resistant plant species in response tocatechin (data not shown), was linked to increasedsecretion of oxalate in rice seedlings exposed to lead (Pb)toxicity (Yang et al. 2000). These data suggested that aroot-secreted organic acid might contribute to the cate-chin-resistance of some plant species. To examine thishypothesis, we placed G. grandiflora and A. thalianaseedlings on a solid pH indicator medium (90 mg l�1
bromcresol purple) to monitor secretion of acidic com-pounds in the root exudates. As expected, resistantG. grandiflora plants constitutively secreted acidic com-pounds from their roots, particularly in the areas aroundlateral roots, with maximum acidification occurringwithin 48 h after the plants were placed on the medium.When a filter disk saturated with (±)-catechin was
placed at the tip of the primary root, increased secretionwas induced, with a visible acidification of the mediumoccurring as early as 1-h post-treatment (Fig. 1e, f).Conversely, catechin-sensitive A. thaliana plants showedvery little medium acidification by 48 h and addition ofthe phytotoxin did not elicit an apparent change in rootsecretion (Fig. 1g, h). These data, particularly the in-crease in secretion of acids by the resistant plant specieselicited by catechin, gave us further indication that anorganic acid may be involved in catechin resistance.
Identification of oxalate in root exudates of resistantplants using HPLC and MS
To determine if organic acids were present at higherlevels in catechin-resistant plant species, we used solidphase extraction (Yang et al. 2000) and a HPLC pro-tocol (Coppola and Starr 1986) designed to specificallyseparate and detect different organic acids. Briefly, wecollected root exudates from the catechin-resistant plantspecies, G. grandiflora and L. sericius, and from thecatechin-sensitive plant species, A. thaliana, as well astwo native grass species Pseudorognaria spicata Pursh(bluebunch wheatgrass) and F. idahoensis Elmer (Idahofescue), which are typically displaced by C. maculosainvasion. We also collected root exudates from each ofthese plants after the growth medium was treated with200 lg ml�1 (±)-catechin to quantify the increase insecretion due to elicitation. Based on retention times ofknown standards and verification by mass spectrometrywe determined that substantial amounts of oxalate werepresent in the root exudates of both G. grandiflora andL. sericeus and that the amount of oxalate present inthese root exudates significantly increased when theplants were treated with (±)-catechin during culture(PGAGF, LUSE<0.001) (Table 1). Both P. spicata andA. thaliana secreted relatively small amounts of oxalate(Table 1) and the amount present in elicited root exu-dates did not differ significantly from the unelicitedsamples (PARTH=0.147; PPSSP=0.910). In contrast, thecatechin-elicited root exudates of F. idahoensis containedsignificantly greater amounts of oxalate per sample thanF. idahoensis unelicited exudates (PFEID=0.012) andexudates of other susceptible species (P<0.001), but thiswas only approximately equal to basal levels seen in theresistant plant species (Table 1). This was particularlyinteresting as there have been conflicting reports on thesusceptibility of this plant species to catechin toxicity(Ridenour and Callaway 2003; Weir et al. 2003; Blairet al. 2005) and these results suggest that F. idahoensismay secrete enough oxalate to provide some level ofprotection against catechin toxicity depending on thespecific laboratory and/or environmental conditions.There was a small amount of ascorbic acid(0.014±0.004 mg sample�1) present in elicited root ex-udates of A. thaliana, but otherwise, oxalic acid was theonly organic acid detected in any root exudates samples.It is possible that oxalate production in plants is favored

over that of other organic acids because it providesmaximum charge balance per carbon atom (Libert andFranceschi 1980). Further, ascorbate can spontaneouslydegrade to oxalate in solutions with a pH>7 (Palgi et al.2005), suggesting that ascorbate might be present inhigher amounts in the root exudates, but that it degradesto oxalate during the extraction procedure.
Oxalate provides protection against (±)-catechinphytotoxicity
We tested the specific ability of pure oxalate to facilitateresistance to catechin in susceptible species. Concen-trations ranging from 10 to 200 lM of commerciallypurchased oxalate (Sigma-Aldrich,St. Louis, MO) wereadded alone or simultaneously with 100 lg ml�1 of (±)-catechin to in vitro grown A. thaliana plants. Oxalatetreatments of 50–100 lM protected A. thaliana fromcatechin rhizotoxicity without any significant ill effectson biomass when compared to untreated controls(P=0.899) (Fig. 2a). Addition of 200 lM of oxalatealone caused severe chlorosis and ultimately resulted incomplete mortality of A. thaliana plants, probably dueto the low pH of the growth medium. Plants treatedwith 10–50 lM oxalate and treated with catechin re-mained viable but were significantly smaller than con-trols (P=0.001); however, when plants were treatedwith 100 lM of oxalate and catechin, there were nosignificant reductions in biomass (P=0.295) and noapparent damage to the root systems (Fig. 2a). Growthmedia supplemented with oxalate was also able to pro-tect native grasses, P. spicata and F. idahoensis fromcatechin phytotoxicity (Fig. 2b). Seedlings of these two
Fig 1 a Five-day-old seedlings of A. thaliana were grown in mediacontaining root exudates of G. grandiflora and 200 lg ml�1 ofcatechin (top), MS medium (center), or MS medium amended with200 lg ml�1 of catechin (bottom). After 5 days, the roots ofb untreated controls and c seedlings treated with catechin butgrown in Gaillardia root exdates remained viable, but d roots ofArabidopsis treated with catechin alone were dead, as evidencedby staining with the vitality stain lactophenol tryphan blue.e G. grandiflora grown on a pH indicator medium containing
bromocresol purple caused dramatic acidification of the mediawithin 48 h, particularly from the areas around the lateral roots.f However, when root tips were exposed to a filter disk saturatedwith (±)-catechin, the time required for visible acidification of themedia was reduced to 1 h. g A. thaliana grown on a pH indicatormedium containing bromocresol purple showed little acidificationof the media after 48 h. h Exposure of root tips to a filter disksaturated with catechin did not induce secretion of acidiccompounds
Table 1 Root exudates of several catechin resistant and susceptiblespecies were analyzed for the presence of oxalate
Sample Oxalate (mg)/sample
Unelicited Elicited [200 lg ml�1
(±)-catechin]
G. grandiflora 0.416±0.032 1.796±0.024a
L. serecius 0.125±0.001 5.070±0.249a
A. thaliana 0.008±0.004 0.019±0.005b
P. spicata 0.005±0.005 0.008±0.008b
F. idahoensis 0.007±0.003 0.144±0.015a
Resistant plant species had higher basal levels, which increasedsignificantly when these species were exposed to catechinaRepresents means that are significantly different between elicitedand unelicited samplesbRepresents means that are not significantly different betweenelicited and unelicited samples

catechin-susceptible native grasses, along with A. thali-ana, were treated with 100 lg ml�1 of (±)-catechin and100 lM of oxalate or with 100 lg ml�1 of (±)-catechinalone. After 7 days, plants treated with only100 lg ml�1 of (±)-catechin showed significantly lessgrowth as indicated by fresh weight (g) than the controlplants or those grown in oxalate supplemented growthmedium (PPSSP=0.001; PFEID=0.002; PARTH=0.001)suggesting that the oxalate reduces the phytotoxic effectof (±)-catechin on these species (Fig. 2b). To furthertest the potential for oxalate to protect plant speciesfrom catechin-mediated root phytotoxicity, we treatedthe resistant species G. grandiflora with 1 mM sodiumorthovanadate (Na3VO4) (Sigma-Aldrich, St. Louis,MO), an anion channel blocker that inhibits the secre-tion of organic acids (Yan et al. 2002). G. grandifloraplants were then treated with 400 lg ml�1 of (±)-cate-
chin, a level insufficient to cause mortality in this speciesin vitro. As expected plants treated with both 1 mMNa3VO4 and 400 lg ml�1 of (±)-catechin showed re-duced growth relative to plants subjected to othertreatments (P<0.001), and were beginning to succumbto the phytotoxin by displaying signs of root phyto-toxicity (Fig. 2c). These results indicate that root-med-iated secretion of oxalate is at least partially responsiblefor catechin resistance in G. grandiflora and L. sericeus.
Oxalate inhibits catechin phytotoxicity by actingas an antioxidant
The evidence that root-secreted oxalate protects plantsagainst oxidative damage caused by catechin is strong.After staining with the reducing dye nitroblue tetrazo-
Fig 2 a Total biomass, measured as fresh weight (g), wasdetermined for A. thaliana plants treated with several concentra-tions of oxalate alone or in combination with 200 lg ml�1 catechin.Error bars represent 1 SE. Different letters denote means that aresignificantly different, as determined using Tukey’s HSD. bSeedlings of two catechin-susceptible native grasses, P. spicataand F. idahoensis, and A. thaliana were treated with 100 lg ml�1 of(±)-catechin and 100 lM of oxalate or with 100 lg ml�1 of (±)-catechin alone. Error bars show (±)1 SE. Means that weresignificantly different from control means are denoted with an
asterisk. c 1 mM of an anion channel blocker, Na3VO4, was addedto the resistant plant G. grandiflora to inhibit the secretion ofoxalate. Plants were then treated with 400 lg ml�1 of (±)-catechin,a level typically insufficient to cause mortality in this species invitro. Plants treated with both 1 mM Na3VO4 and 400 lg ml�1 of(±)-catechin were significantly smaller than control plants treatedwith methanol, 1 mM Na3VO4, or 400 lg ml�1 of (±)-catechinalone. Error bars represent (±)1 SE. Means that were significantlydifferent from the control, as determined by Dunnet’s two-tailedtest at 95% confidence are denoted with a asterisk

lium (0.1% w/v), A. thaliana plants treated with100 lg ml�1 of (±)-catechin displayed blue colorationin the roots and leaves, indicating the presence of O2
radicals; whereas untreated control plants and thosetreated with oxalate showed no staining (Fig. 3a–c).When 10 lM oxalate and 100 lg ml�1 of (±)-catechinwere both added to A. thaliana there was no visible O2
generation in the leaves, as was seen when catechin wasadded alone, but individual root cells showed accumu-lation of O2 radicals (Fig. 3d). As in the untreatedcontrols, no intracellular O2 radicals were visible inplants treated with 100 lM of oxalate and 100 lg ml�1
of (±)-catechin (Fig. 3e).The exact cellular targets of most allelochemicals
have not yet been elucidated (Weir et al. 2004). How-ever, oxidative damage leading to lipid peroxidation,which subsequently decreases the integrity of cell mem-branes and ultimately results in cytotoxicity, has beenimplicated as a direct or indirect effect of exposure to anumber of allelotoxins (Yu et al. 2003; Cruz-Ortegaet al. 2002). To determine if the free radicals generated in
response to catechin treatment resulted in lipid peroxi-dation, we quantified conjugated dienes, a product oflipid peroxidation, from A. thaliana plants in response to100 lg ml�1 of (±)-catechin, 100 lM of oxalate, orboth chemicals added simultaneously (Table 2). Therewas no difference in the amount of conjugated dienespresent in plant material that had been treated withoxalate or oxalate and catechin, however; plants treatedwith catechin alone showed a 1.5-fold increase in thepresence of conjugated dienes 24 h after exposure to thephytotoxin, indicating that the cytotoxic effects of cat-echin are at least partially due to oxidative stress, cor-raborating previously published reports (Bais et al.2003).
Oxalate may influence plant interactions in naturalecosystems
The relationship between oxalate-secreting plantsand catechin resistance appears to affect ecological
Fig 3 Using the reducing dye nitroblue tetrazolium, we stained A.thaliana plants that had been treated with different concentrationsof oxalate, with and without the addition of 100 lg/ml of catechin,to visualize the generation of O2 radicals. a The MeOH treatedcontrol and c plants treated with 100-lM oxalate showed noaccumulation of O2. b Plants treated only with catechin accumu-
lated O2 in the roots and the shoots. d In the presence of 10-lMoxalate and catechin, O2 was visible in individual root cells, but wasnot apparent in the shoots. e 100 lM of oxalate completelyprevented O2 accumulation in catechin treated plants. Scale barsrepresent 10 lm

interactions in the field. As noted above, G. grandifloraand L. sericeus are unusually good competitors againstC. maculosa (Ridenour and Callaway 2001; Callawayet al. 2004), and field measurements show that nativegrasses are much more abundant near L. sericeus inareas heavily invaded by C. maculosa (Fig. 4a). Of 100(0.25 m2) plots, seven contained L. sericeus and of thesesix (86%) contained P. spicata and five (71%) containedF. idahoensis. Of the 93 plots without L. sericeus, 14(15%) contained P. spicata and 15 (16%) containedF. idahoensis. This suggests that these root exudatechemical counter-measures of catechin-resistant plantsmay facilitate other species. We tested this hypothesis ina field experiment and found that in C. maculosa in-vaded patches, native P. spicata (FCEMA=18.97,df=1,59, P<0.001; FLUSE · CEMA=24.50, df=1,59,P<0.001) and F. idahoensis (FCEMA=27.43, df=1,59,P<0.001; FLUSE · CEMA=11.32, df=1,59, P=0.001)transplanted within 10 cm of a L. sericeus plant grewsignificantly larger than those planted more than 1 m
from L. sericeus (Fig. 4b, upper). However, this indirectfacilitative relationship only occurred in dense C. mac-ulosa patches; when P. spicata (F=3.56, df=1,59,P=0.064) and F. idahoensis (F=0.24, df=1,59,P=0.877) were transplanted next to L. sericeus in theabsence of C. maculosa the effect of L. sericeus wascompetitive (Fig. 4b, lower). In other words, L. sericeusonly had facilitative effects in the presence of C. macul-osa. These results suggest that the beneficial effects ofL. sericeus on native grass survival in C. maculosa standsis not entirely due to increased availability of nitrogenprovided by the legume.
The difficulty of relating laboratory observations toplant behavior under field conditions has been a per-sistant constraint in studying allelopathic interactions.In an attempt to relate our laboratory results with fieldconditions, we collected soil samples from the rhizo-spheres of L. sericeus growing alone, L. sericeus growingclosely together with C. maculosa, and C. maculosagrowing alone, and analyzed the soil for catechin andoxalate. We did not look for oxalate from soil ofC. maculosa-only stands because no oxalate was de-tected in in vitro grown exudates of this plant (data notshown). Further, we did not examine soil samples ofL. sericeus growing alone for catechin because of therelative rareity of this compound in plants. There areonly a few reports of catechin-producing plants in theliterature, and it is not present in the exudates ofC. maculosa’s close relatives C. diffusa and Acroptilin(Centaurea) repens (Stermitz et al. 2003; Vivanco et al.2004). Although we obtained poor oxalate recoveryfrom our soil samples (<10% recovery from spikedsamples), we were able to detect significantly greaterquantities of oxalate in soil samples collected near
Table 2 Arabidopsis thaliana plants were treated with (±)-cate-chin, oxalate, or both compounds and extracted for conjugateddienes, an indicator of lipid peroxidation
Lipid peroxidation
Treatment Conjugateddienes (OD230)
200 lg ml�1 catechin 0.56±0.02a
100 lM oxalic acid 0.34±0.02b
200 lg ml�1 catechinand 100 lM oxalic acid
0.35±0.01b
Different letters represent significantly different meansC
ove
r (%
)
0
5
10
15
20Lupinus in plotNo Lupinus in plot
Pseudoroegneria Festuca
*
*
0
1
2
3
4
0
1
2
3
4With LupinusWithout Lupinus
Pseudoroegneria Festuca
Not in Centaurea stand
In Centaurea stand
Bio
mas
s (g
)
a bFig 4 a Percent cover ofPseudoroegneria spicata andF. idahoensis in intermountaingrassland heavily invaded byC. maculosa, in plots either withor without L. sericeus. Errorbars show 1 SE and asteriskdesignate a significantdifference of P<0.001 in a ttest. b Aboveground biomass ofP. spicata and F. idahoensistransplanted either next to orfar from L. sericeus, and eitherin intermountain grasslandheavily invaded by C. maculosa(�100% cover (not relativized)or in grassland with �2% coverof C. maculosa.) Error barsshow (±)1 SE
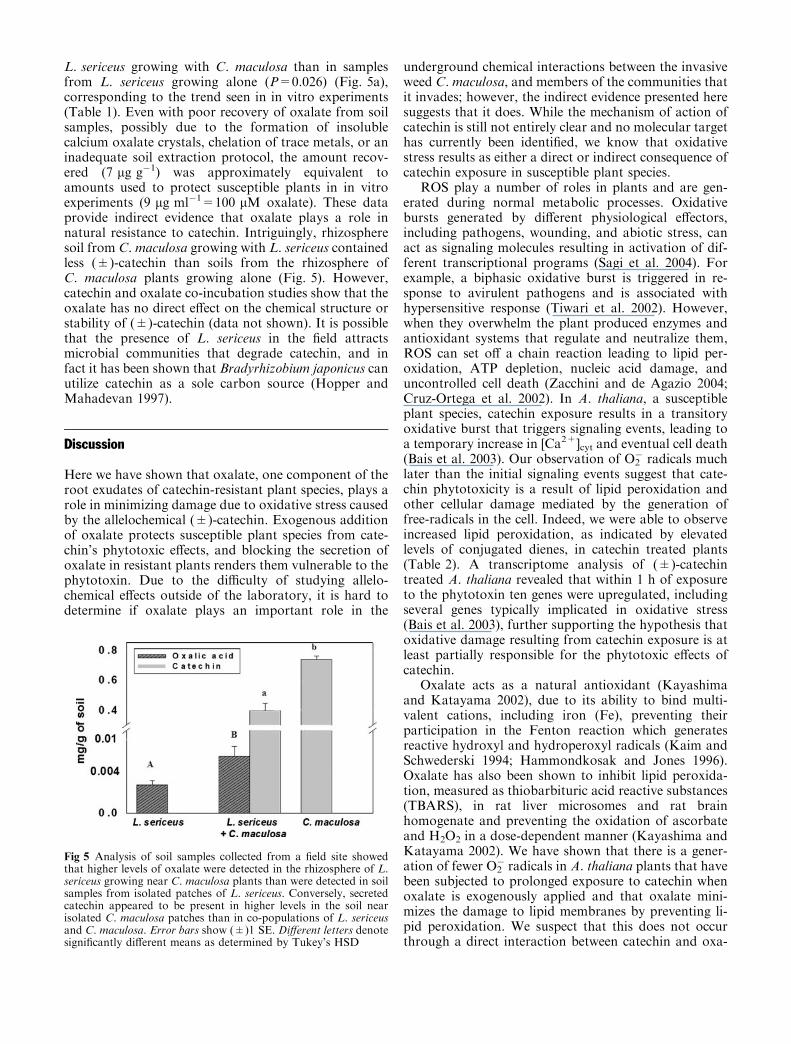
L. sericeus growing with C. maculosa than in samplesfrom L. sericeus growing alone (P=0.026) (Fig. 5a),corresponding to the trend seen in in vitro experiments(Table 1). Even with poor recovery of oxalate from soilsamples, possibly due to the formation of insolublecalcium oxalate crystals, chelation of trace metals, or aninadequate soil extraction protocol, the amount recov-ered (7 lg g�1) was approximately equivalent toamounts used to protect susceptible plants in in vitroexperiments (9 lg ml�1=100 lM oxalate). These dataprovide indirect evidence that oxalate plays a role innatural resistance to catechin. Intriguingly, rhizospheresoil from C. maculosa growing with L. sericeus containedless (±)-catechin than soils from the rhizosphere ofC. maculosa plants growing alone (Fig. 5). However,catechin and oxalate co-incubation studies show that theoxalate has no direct effect on the chemical structure orstability of (±)-catechin (data not shown). It is possiblethat the presence of L. sericeus in the field attractsmicrobial communities that degrade catechin, and infact it has been shown that Bradyrhizobium japonicus canutilize catechin as a sole carbon source (Hopper andMahadevan 1997).
Discussion
Here we have shown that oxalate, one component of theroot exudates of catechin-resistant plant species, plays arole in minimizing damage due to oxidative stress causedby the allelochemical (±)-catechin. Exogenous additionof oxalate protects susceptible plant species from cate-chin’s phytotoxic effects, and blocking the secretion ofoxalate in resistant plants renders them vulnerable to thephytotoxin. Due to the difficulty of studying allelo-chemical effects outside of the laboratory, it is hard todetermine if oxalate plays an important role in the
underground chemical interactions between the invasiveweed C. maculosa, and members of the communities thatit invades; however, the indirect evidence presented heresuggests that it does. While the mechanism of action ofcatechin is still not entirely clear and no molecular targethas currently been identified, we know that oxidativestress results as either a direct or indirect consequence ofcatechin exposure in susceptible plant species.
ROS play a number of roles in plants and are gen-erated during normal metabolic processes. Oxidativebursts generated by different physiological effectors,including pathogens, wounding, and abiotic stress, canact as signaling molecules resulting in activation of dif-ferent transcriptional programs (Sagi et al. 2004). Forexample, a biphasic oxidative burst is triggered in re-sponse to avirulent pathogens and is associated withhypersensitive response (Tiwari et al. 2002). However,when they overwhelm the plant produced enzymes andantioxidant systems that regulate and neutralize them,ROS can set off a chain reaction leading to lipid per-oxidation, ATP depletion, nucleic acid damage, anduncontrolled cell death (Zacchini and de Agazio 2004;Cruz-Ortega et al. 2002). In A. thaliana, a susceptibleplant species, catechin exposure results in a transitoryoxidative burst that triggers signaling events, leading toa temporary increase in [Ca2+]cyt and eventual cell death(Bais et al. 2003). Our observation of O2
� radicals muchlater than the initial signaling events suggest that cate-chin phytotoxicity is a result of lipid peroxidation andother cellular damage mediated by the generation offree-radicals in the cell. Indeed, we were able to observeincreased lipid peroxidation, as indicated by elevatedlevels of conjugated dienes, in catechin treated plants(Table 2). A transcriptome analysis of (±)-catechintreated A. thaliana revealed that within 1 h of exposureto the phytotoxin ten genes were upregulated, includingseveral genes typically implicated in oxidative stress(Bais et al. 2003), further supporting the hypothesis thatoxidative damage resulting from catechin exposure is atleast partially responsible for the phytotoxic effects ofcatechin.
Oxalate acts as a natural antioxidant (Kayashimaand Katayama 2002), due to its ability to bind multi-valent cations, including iron (Fe), preventing theirparticipation in the Fenton reaction which generatesreactive hydroxyl and hydroperoxyl radicals (Kaim andSchwederski 1994; Hammondkosak and Jones 1996).Oxalate has also been shown to inhibit lipid peroxida-tion, measured as thiobarbituric acid reactive substances(TBARS), in rat liver microsomes and rat brainhomogenate and preventing the oxidation of ascorbateand H2O2 in a dose-dependent manner (Kayashima andKatayama 2002). We have shown that there is a gener-ation of fewer O2
� radicals in A. thaliana plants that havebeen subjected to prolonged exposure to catechin whenoxalate is exogenously applied and that oxalate mini-mizes the damage to lipid membranes by preventing li-pid peroxidation. We suspect that this does not occurthrough a direct interaction between catechin and oxa-
Fig 5 Analysis of soil samples collected from a field site showedthat higher levels of oxalate were detected in the rhizosphere of L.sericeus growing near C. maculosa plants than were detected in soilsamples from isolated patches of L. sericeus. Conversely, secretedcatechin appeared to be present in higher levels in the soil nearisolated C. maculosa patches than in co-populations of L. sericeusand C. maculosa. Error bars show (±)1 SE. Different letters denotesignificantly different means as determined by Tukey’s HSD

late, but that oxalate simply aids in scavenging freeradicals generated by or in response to catechin expo-sure. Oxalate is the metabolic degradation product ofanother antioxidant, ascorbate (Yang and Loewus 1975;Green and Fry 2005), and it is possible that the increasedsecretion of oxalate in resistant plants is due to an in-crease in ascorbate, produced as part of the plant cellantioxidant system (Foyer et al. 1994), to counteract theoxidative stress caused by catechin. However, the resultsof Bais et al. (2003) showing that ROS signaling isblocked by exogenous addition of ascorbate, and ourresults showing that growth medium supplemented withoxalate protects susceptible plant species suggest that itis not merely the internal efficiency of antioxidant sys-tems that affects the resistance of a plant species tocatechin phytotoxicity.
In summation, we have provided insight into thephytotoxic mechanism of (±)-catechin secreted by theinvasive weed C. maculosa, and suggested a potentialmeans by which some plant species in native communi-ties may defend against this invader. Our data indicatethat chemically mediated attack and counter measurebetween C. maculosa and C. maculosa-resistant nativeplants appears to play a role in competitive interactionsand may mediate indirect facilitation of plants that aremuch weaker competitors with C. maculosa. This sug-gests that plant–plant interactions may be far morecomplicated and species-specific than resource competi-tion alone. Furthermore, defensive chemical responsesshed light on how native communities may meet thechallenges of allelopathic invaders.
Acknowledgements This work was supported by grants fromUSDA-WRIPM (grant no. 2003-05060, J.M.V.), National ScienceFoundation- (grant no. NSF-IBN 0335203, J.M.V.), U.S. Depart-ment of Defense SERDP (CS1388 to J.M.V. and R.M.C.) andInvasive Weeds Initiative of the State of Colorado (J.M.V. andF.R.S.). JMV is an NSF-CAREER faculty fellow (grant no. MCB093014).
References
Bais HP, Walker TS, Stermitz FR, Hufbauer R, Vivanco JM (2002)Enantiomeric-dependant phytotoxic and antimicrobial activityof (+/�)-catechin: a rhizosecreted racemic mixture from spot-ted knapweed. Plant Physiol 128:1173–1179
Bais HP, Vepachedu R, Gilroy S, Callaway RM, Vivanco JM(2003) Allelopathy and exotic plant invasion: from moleculesand genes to species interactions. Science 301:1377–1380
Blair AC, Hanson BD, Brunk GR, Marrs RA, Westra P, NissenSJ, Hufbauer RA (2005) New techniques and findings in thestudy of a candidate allelochemical implicated in invasionsuccess. Ecol Lett 8:1039–1047
Callaway RM, Aschehoug ET (2000) Invasive plants versus theirnew and old neighbours: a mechanism for exotic invasion.Science 290:521–523
Callaway RM, Deluca TH, Belliveau WM (1999) Biological controlherbivores may increase competitive ability of the noxious weedCentaurea maculosa. Ecology 80:1196–1201
Callaway RM, Thelen GC, Barth S, Ramsey PW,Gannon JE (2004)Soil fungi alter interactions between the invader Centaureamaculosa and North American natives. Ecology 85:1062–1071
Coppola ED, Starr MS (1986) Liquid chromatographic determina-tion of major organic acids in apple juice and cranberry juicecocktail: collaborative study. J Assoc OffAnal Chem 69:594–598
Cruz-Ortega R, Ayala-Cordero G, Anaya AL (2002) Allelochem-ical stress produced by the aqueous leachate of Callicarpa ac-uminata: effects on roots of bean, maize, and tomato. PhysiolPlant 116:20–27
Foyer CH, Descourvieres P, Kunert KJ (1994) Protection againstoxygen radicals: important defence mechanism studied intransgenic plants. Plant Cell Environ 17:507–523
Gardner WK, Barber DA, Parbery DG (1983) The acquisition ofphosphorus by Lupinus alba L.: II. The probable mechanism bywhich phosphorus movement in the soil/root interface is en-hanced. Plant Soil 70:107–124
Green MA, Fry SC (2005) Vitamin C degradation in plant cells viaenzymatic hydrolysis of 4-O-oxalyl-L-threonate. Nature 433:83–87
Hammondkosak KE, Jones JDG (1996) Resistance gene-depen-dent plant defense responses. Plant Cell 8:1773–1791
Hopper W, Mahadevan A (1997) Degradation of catechin byBradyrhizobium japonicum. Biodegradation 8:159–165
Kaim W, Schwederski B (1994) Bioinorganic chemistry: inorganicelements in the chemistry of life. Wiley, New York
Kayashima T, Katayama T (2002) Oxalate is available as a naturalantioxidant in some systems. Biochem Biophys Acta 1537:1–3
Keerthisinghe G, Hocking PJ, Ryan PR, Delhaize E (1998) Effectof phosphorus supply on the formation and function of proteidroots of white lupine (Lupinus albus L.). Plant Cell Environ21:467–478
Libert B, Franceschi VR (1980) Oxalate in crop plants. J AgricFood Chem 35:926–938
Ma JF, Zheng SJ, Matsumoto H (1997) Specific secretion of citricacid induced by Al stress in Cassia tora L. Plant Cell Physiol38:1019–1025
Murashige T, Skoog F (1962) A revised medium for rapid growthand bioassay with tissue culture. Physiol Plant 15:473
Neumann G, Massonneau A, Martinoia E, Romheld V (1999)Physiological adaptations to phosphorus deficiency duringproteoid root development in white lupine. Planta 208:373–328
Palgi N, Vatnik I, Pinshow B (2005) Oxalate, calcium and ashintake and excretion balances in fat sand rats (Psammomysobesus) feeding on two different diets. Compar Biochem PhysiolDOI 10.1016/j.cbpb.2005.03.011
Perry LG, Thelen GC, Ridenour WM, Weir TL, Callaway RMet al (2005) Dual role for an allelochemical: (+/�)-catechinfrom Centaurea maculosa root exudates regulates conspecificseedling establishment. J Ecol 93:1126–1135
Recknagel RO, Glende EA (1984) Spectrophotometric detection oflipid conjugated dienes. Method Enzymol 105:331–337
Ridenour WM, Callaway RM (2001) The relative importance ofallelopathy in interference: the effects of an invasive weed on anative bunchgrass. Oecologia 124:444–450
Ridenour WM, Callaway RM (2003) Root herbivores, pathogenicfungi, and competition between Centaurea maculosa and Fest-uca idahoensis. Plant Ecol 169:161–170
Ryan PR, Delhaize E, Randall PJ (1995) Malate efflux from rootapices and tolerance to aluminum are highly correlated inwheat. Aust J Plant Physiol 22:531–563
Sagi M, Davydov O, Orazova S, Yesbergenova Z, Ophir R,Stratmann JW, Fluhrb R (2004) Plant respiratory burst oxidasehomologs impinge on wound responsiveness and developmentin lycopersicon esculentum. Plant Cell 16:616–628
Stermitz FR, Bais HP, Foderaro TA, Vivanco JM (2003) 7,8-Benzoflavone: a phytotoxin from root exudates of invasiveRussian knapweed. Phytochemistry 64:493–497
Thelen GC, Vivanco JM, Newingham B, Good W, Bais HP et al(2005) Insect herbivory stimulates allelopathic exudation by aninvasive plant and the suppression of natives. Ecol Lett 8:209–217
Tiwari BS, Belenghi B, Levine A (2002) Oxidative stress increasedrespiration and generation of reactive oxygen species, resulting inATP depletion, opening of mitochondrial permeability transition,and programmed cell death. Plant Physiol 128:1271–1281

Vivanco JM, Bais HP, Stermitz FR, Thelen GC, Callaway RM(2004) Biogeographical variation in community response toroot allelochemistry: novel weapons and exotic invasion. EcolLett 7:285–292
Weir TL, Bais HP, Vivanco JM (2003) Intraspecific and interspe-cific interactions mediated by a phytotoxin, (�)-catechin, se-creted by the roots of Centaurea maculosa (spotted knapweed).J Chem Ecol 29:2397–2412
Weir TL, Park S-W, Vivanco JM (2004) Biochemical and physio-logical mechanisms mediated by allelochemicals. Curr OpinPlant Biol 7:472–479
Yan F, Zhu YY, Muller C, Zorb C, Schubert S (2002) Adaptationof H+-pumping and plasma membrane H+ ATPase activity inproteoid roots of white lupin under phosphate deficiency. PlantPhysiol 129:50–63
Yang JC, Loewus FA (1975) Metabolic conversion of L-ascorbic-acid to oxalic-acid in oxalate-accumulating plants. Plant Phys-iol 56:283–285
Yang YY, Jung JY, Song WY, Suh HS, Lee Y (2000) Identificationof rice varieties with high tolerance or sensitivity to lead andcharacterization of the mechanism of tolerance. Plant Physiol124:1019–1026
Yu JQ, Ye SF, Zhang MF, Hu WH (2003) Effects of root exudatesand aqueous root extracts of cucumber (Cucumis sativus) andallelochemicals, on photosynthesis and antioxidant enzymes incucumber. Biochem Syst Ecol 31:129–139
Zacchini M, Agazio M (2004) Spread of oxidative damageand antioxidative responses through cell layers of tobaccocallus after UV-C treatment. Plant Physiol Biochem 42:445–450
Zheng SJ, Ma MF, Matsumoto H (1998) High aluminum resis-tance in buckwheat: I. Al-induced specific secretion of oxalatefrom root tips. Plant Physiol 117:745–751




















