
Effect of water column phosphorus reduction on competitive outcome and traits of Ludwigia...
10
Aquatic Invasions (2014) Volume 9, Issue 2: 157–166 doi: http://dx.doi.org/10.3391/ai.2014.9.2.04 © 2014 The Author(s). Journal compilation © 2014 REABIC Open Access 157 Research Article Effect of water column phosphorus reduction on competitive outcome and traits of Ludwigia grandiflora and L. peploides, invasive species in Europe Joëlle Gérard 1 *, Natacha Brion 2 and Ludwig Triest 1 1 Plant Biology and Nature Management (APNA), Vrije Universiteit Brussel, Pleinlaan 2, B-1050 Brussels, Belgium 2 Analytical and Environmental Geochemistry (AMGC), Vrije Universiteit Brussel, Pleinlaan 2, B-1050 Brussels, Belgium E-mail: [email protected] (JG), [email protected] (NB), [email protected] (LT) *Corresponding author Received: 14 December 2013 / Accepted: 24 April 2014 / Published online: 14 May 2014 Handling editor: Vadim Panov Abstract The invasion of alien macrophytes in aquatic ecosystems has caused serious ecological and economic impacts. Their introduction often leads to competition with natives and already established invasive species. Competition among invasive species is assumed to be greater than among native and invasive species, especially for plants with similar growth form and position in the water column. Many freshwater bodies are eutrophicated due to high phosphorus inputs, which have led to a competitive advantage for invasive macrophytes. A decrease in water column phosphorus load might lead to a change in competitive performance and reveal apparent variation in those traits promoting invasiveness. We investigated the effect of a water column phosphorus reduction on the growth, competitive outcome, traits and nutrient uptake of two related invasive Ludwigia species, L. grandiflora and L. peploides. We performed indoor competition experiments in eutrophic and mesotrophic conditions following a complete additive design. L. grandiflora always was the better competitor in both trophic conditions and had higher trait values compared to L. peploides. Both species relative growth rates (RGR) and trait values were affected by a P reduction, especially the number of branches and stem length. Although both species prefer higher water column phosphorus loadings, they can also colonize habitats of lower mesotrophic phosphorus concentrations. L. peploides was able to store more P in its tissue compared to L. grandiflora, thereby possibly outcompeting L. grandiflora in even lower P concentrations. Our results show a water column P reduction to lower branch numbers, stem length and biomass, which could reduce mat formation and allow for more effective control of both species. Key words: macrophyte, alien, eutrophication, growth, freshwater, competition, RGR Introduction The invasion of alien plant species has become a worldwide problem and is considered a severe threat to biodiversity (Mack et al. 2000). During the last few decades, a high number of macrophyte species, especially ornamental plants, have been introduced due to increases in travel and trade (Brunel 2009; Hulme 2009). The presence of invasive alien species often leads to competition with native species, usually resulting in a partial or complete loss of the latter. Competition among invasive species will be higher, particularly between related species with similar growth form residing in the same position in the water column (Gopal and Goel 1993; Cahill et al. 2008). Invasive plants are characterized by higher growth rates (Fogarty and Facelli 1999; Burns 2004; Funk and Vitousek 2007; Grotkopp and Rejmanek 2007) and other traits which enhance their invasiveness and show plasticity with resource availability (Hastwell et al. 2008; Fan et al. 2013). Many studies have investigated traits associated with invasiveness, however a universally applicable set of predictive traits is still elusive, especially for aquatic plants (Daehler 2003; Mony et al. 2006; Richardson and Pysek 2006). Trait studies focusing on competition between related invasive species with the same growth form are more suitable to reveal which species will be a better competitor and which traits promote invasiveness (Pysek and Richardson 2007; Feng and Fu 2008). Impacts caused by invasive plants underline the importance of understanding the factors that influence a habitat’s invasibility (Chadwell and Engelhardt 2008). Eutrophication of freshwater ecosystems as a result of human activities has facilitated such invasions and enhanced their adverse effects (Dukes and Mooney 1999; Daehler 2003), and is currently viewed as the second
Transcript of Effect of water column phosphorus reduction on competitive outcome and traits of Ludwigia...

Aquatic Invasions (2014) Volume 9, Issue 2: 157–166 doi: http://dx.doi.org/10.3391/ai.2014.9.2.04
© 2014 The Author(s). Journal compilation © 2014 REABIC
Open Access
157
Research Article
Effect of water column phosphorus reduction on competitive outcome and traits of Ludwigia grandiflora and L. peploides, invasive species in Europe
Joëlle Gérard1*, Natacha Brion2 and Ludwig Triest1
1Plant Biology and Nature Management (APNA), Vrije Universiteit Brussel, Pleinlaan 2, B-1050 Brussels, Belgium 2Analytical and Environmental Geochemistry (AMGC), Vrije Universiteit Brussel, Pleinlaan 2, B-1050 Brussels, Belgium
E-mail: [email protected] (JG), [email protected] (NB), [email protected] (LT)
*Corresponding author
Received: 14 December 2013 / Accepted: 24 April 2014 / Published online: 14 May 2014
Handling editor: Vadim Panov
Abstract
The invasion of alien macrophytes in aquatic ecosystems has caused serious ecological and economic impacts. Their introduction often leads to competition with natives and already established invasive species. Competition among invasive species is assumed to be greater than among native and invasive species, especially for plants with similar growth form and position in the water column. Many freshwater bodies are eutrophicated due to high phosphorus inputs, which have led to a competitive advantage for invasive macrophytes. A decrease in water column phosphorus load might lead to a change in competitive performance and reveal apparent variation in those traits promoting invasiveness. We investigated the effect of a water column phosphorus reduction on the growth, competitive outcome, traits and nutrient uptake of two related invasive Ludwigia species, L. grandiflora and L. peploides. We performed indoor competition experiments in eutrophic and mesotrophic conditions following a complete additive design. L. grandiflora always was the better competitor in both trophic conditions and had higher trait values compared to L. peploides. Both species relative growth rates (RGR) and trait values were affected by a P reduction, especially the number of branches and stem length. Although both species prefer higher water column phosphorus loadings, they can also colonize habitats of lower mesotrophic phosphorus concentrations. L. peploides was able to store more P in its tissue compared to L. grandiflora, thereby possibly outcompeting L. grandiflora in even lower P concentrations. Our results show a water column P reduction to lower branch numbers, stem length and biomass, which could reduce mat formation and allow for more effective control of both species.
Key words: macrophyte, alien, eutrophication, growth, freshwater, competition, RGR
Introduction
The invasion of alien plant species has become a worldwide problem and is considered a severe threat to biodiversity (Mack et al. 2000). During the last few decades, a high number of macrophyte species, especially ornamental plants, have been introduced due to increases in travel and trade (Brunel 2009; Hulme 2009). The presence of invasive alien species often leads to competition with native species, usually resulting in a partial or complete loss of the latter. Competition among invasive species will be higher, particularly between related species with similar growth form residing in the same position in the water column (Gopal and Goel 1993; Cahill et al. 2008). Invasive plants are characterized by higher growth rates (Fogarty and Facelli 1999; Burns 2004; Funk and Vitousek 2007; Grotkopp and Rejmanek 2007) and other traits which enhance their invasiveness
and show plasticity with resource availability (Hastwell et al. 2008; Fan et al. 2013). Many studies have investigated traits associated with invasiveness, however a universally applicable set of predictive traits is still elusive, especially for aquatic plants (Daehler 2003; Mony et al. 2006; Richardson and Pysek 2006). Trait studies focusing on competition between related invasive species with the same growth form are more suitable to reveal which species will be a better competitor and which traits promote invasiveness (Pysek and Richardson 2007; Feng and Fu 2008).
Impacts caused by invasive plants underline the importance of understanding the factors that influence a habitat’s invasibility (Chadwell and Engelhardt 2008). Eutrophication of freshwater ecosystems as a result of human activities has facilitated such invasions and enhanced their adverse effects (Dukes and Mooney 1999; Daehler 2003), and is currently viewed as the second

J. Gérard et al.
158
major threat to biodiversity (Van et al. 1999; Daehler 2003). High water column nutrient loadings favor invasives by leading to increased growth and a change in competitive balance between plant species (Smith et al. 1999). Several studies have shown invasives to be able to take up more nutrients at high nutrient levels, while natives are likely to have adapted to low nutrient conditions and show low growth (Fogarty and Facelli 1999; Burns 2004; Funk and Vitousek 2007; Grotkopp and Rejmanek 2007; Hastwell et al. 2008; Fan et al. 2013). In freshwater ecosystems, P is usually the main limiting element, and eutrophi-cation is thus enhanced by P inputs (Schindler 1977; Sharpley et al. 2003). Several studies have however shown N limitation to occur as frequent as P limitation (Elser et al. 2007; Lewis and Wurtsbaugh 2008). Nevertheless, even if N is primarily limiting, controlling P levels will generally be the most successful and low-cost method for reducing algal populations (Lewis and Wurtsbaugh 2008). Hence, a water column phosphorus reduction might affect macrophyte species’ P uptake, growth rate and change the competitiveness of the invasive species, since species might vary in their ability to cope with low P (Koerselman and Meuleman 1996). A reduction in nutrients may also affect the amount of internally stored nutrient concentrations (Chapin 1991). Invasive species are often able to better exploit high nutrients or compete by pre-empting the supply of limited nutrients (Craine et al. 2005). Plants with high storage ability should occur over a wide ecological range and are consequently highly competitive (Garbey et al. 2004; Thiébaut 2005). We therefore hypothesize that the better competitor will have the highest tissue P concentration.
Ludwigia grandiflora ((Michaux) Greuter and Burdet, 1987) and Ludwigia peploides ((Kunth) P.H. Raven, 1963) are two related invasive macrophyte species originating from North and South America, and are now both present in several European countries including Belgium (Bauchau et al. 1984; Dandelot 2004; Hussner 2012). The distribution of the two species has been intensely studied in France, where L. peploides has extended more throughout the Mediterranean area, while L. grandiflora has spread more to the Eastern and Northern parts of France, however the species are often found to co-occur (Ancrenaz and Dutartre 2002; Dandelot et al. 2005b). In Belgium and other European countries, the distribution of L. peploides is still limited (EPPO
2011), but since the two species are able to exist in a wide range of habitats (Haury et al. 2010; Hussner 2010), they could ultimately co-occur in invaded areas. The spread of the species could also be underestimated since L. peploides and L. grandiflora are two morphologically very similar species that are difficult to distinguish and often confused when not flowering (Dandelot et al. 2008).
Both Ludwigia species are able to spread quickly due to their rapid growth and propagation by fragments (Dandelot et al. 2005b; Verloove 2006). They can laterally extend their shoots on top of the water column, thereby producing dense mats, which are known to reduce native biodiversity, degrade water quality by reducing oxygen and silt up ponds (Dutartre et al. 1997; Dandelot et al. 2005b; Dandelot et al. 2008; Stiers 2011). Both species produce two types of roots: branched downwards growing roots and spongy unbranched upwards growing roots for aeration (Ellmore 1981). The species are very vigorous, since they are able to grow in several sediment types and can inhabit wet terrestrial habitats as well as aquatic habitats (Matrat et al. 2006; Hussner 2010).
In this paper, we investigated the effect of a water column phosphorus reduction on growth, strategies and competition between two invasive related species Ludwigia grandiflora and Ludwigia peploides with similar growth form and position in the water column. Both species were grown in monocultures and mixtures in water column P concentrations concurring with eutrophic and mesotrophic conditions. Our goals are to determine (1) the effects of a water column phosphorus reduction on growth; (2) the better competitor in eutrophic and mesotrophic conditions and hence the effect of a phosphorus reduction on competitive outcome; (3) whether the better competitor exhibits different traits related to nutrient acquisition and invasiveness; (4) whether traits show trait plasticity with decreasing P; and (5) whether the competitive strength of the species is related to the amount of phosphorus acquired.
Materials and methods
Plant material
We studied two invasive species: Ludwigia gran-diflora (Michaux) Greuter and Burdet and Ludwigia peploides (Kunth) P.H. Raven. L. grandiflora was collected from De Spicht – Goorbroek in Belgium. L. peploides was obtained from the Plant

Effect of phosphorus reduction on competition and traits
159
Protection Service of the Netherlands (collected from former polder Alblas, the Netherlands).
Experimental design
All plants were cut into 10±1 cm long apices and placed into 100 L containers filled with rain water, to allow for root growth. After 3 weeks, all plants (average height 15.0 cm) had grown roots and were planted in monocultures and mixtures in 1L pots (height 10 cm) following a complete additive design (Spitters 1983). L. grandiflora : L. peploides planting densities were 0:2, 2:0, 2:2, 2:4, 4:2, 4:4, 4:0, 0:4, each combination replicated 5 times. Although we do not have any data on field densities, the same initial densities were used in Ludwigia competition experiments performed by Njambuya and Triest (2010) and Thouvenot et al. (2013). Pots were filled with sand and KNO3 mixture (1g KNO3/l sand) and covered with 3 cm of clay to reduce N flow to the water column. Pots were then placed into 12 l buckets (height 24,5 cm) filled with 10 l solution, such that the plants were submerged only a few centimeters below the water surface and were able to emerge during the first week. We used a 3x diluted Nitrogen lacking Hoagland and Arnon (1950) solution. P concentrations were adjusted to 30 (mesotrophic) and 100 µg/l (eutrophic condition) with KH2PO4. All buckets were placed randomly in a greenhouse under 12:12 hr day: night cycle with Photosynthetically Active Radiation (PAR) ranging from 105 to 197 µmol m-2 s-1. All solutions were replenished and thoroughly mixed once a week. At the end of the experiment, various biological traits were measured including stem length, number of leaves, number of shoots, and length and width of the leaves (leaf at 3 cm from the plant apex). The average pH (average eutrophic 7.50, mesotrophic 7.44), water temperature (23.8°C) and air temperature (29.4°C) were recorded. After 8 weeks, all plants were harvested and separated into roots and shoots. All biomass was dried for 72 hours at 70°C and weighed. We calculated the root/shoot ratio as root weight (g)/shoot weight (g). Particulate organic nitrogen (PON) and carbon (POC) concentrations in plant tissue were analyzed simultaneously with a Flash EA 1112 (Thermo) elemental analyzer (Nieuwenhuize et al. 1994). The analysis combines conversion of PON and POC into N2 and CO2, separation on a chromatographic solid phase column and detection by thermal conductivity (Pella 1990). Concentrations of PON and POC were obtained by standardization with
acetanilide. Total phosphorus in plants was analyzed using a modified digestion protocol (Johengen 1996) after which digestions were analyzed with a segmented flow auto-analyzer QuAAtro (SEAL Analytical).
Statistical analysis
The relative growth rate (RGR) of each species was calculated according to the formula: RGRln Y/y /t , where y is the species’ initial aboveground biomass or length and Y the species’ aboveground biomass or length at the end of the experimental period t. We determined the relative competitive strength as formulated by olde Venterink and Güsewell (2010): in each treatment, we calculated the ratio of a species’ biomass in a mixed pot by the biomass in a monoculture pot. Monoculture and mixture pots were randomly coupled. The superior competitor was the species which produced significantly more biomass in mixture than in monoculture (competitive response > 1) and/or caused the other species to produce significantly less biomass in mixture than in monoculture (CR<1) (olde Venterink and Güsewell 2010). By calculating the L. grandiflora / L. peploides biomass ratio, we examined the relative dominance of the two species in competition (olde Venterink and Güsewell 2010). We analyzed the differences in biomass, weight RGR and length RGR between separate initial planting densities using non-parametric Mann-Whitney U-tests for monocultures and Kruskall-Wallis tests for mixtures due to the non-normal distribution of the data. For all the parameters, there was no significant difference between the initial planting densities in monocultures or in mixtures and therefore the different densities of monocultures and mixtures were pooled. The differences in RGR between species and between nutrient levels were analyzed with parametric t-tests. Differences in other traits, biomass, competitive response, biomass ratio and tissue nutrient concentrations did not pass the assumptions of normality or equal variances and were therefore analyzed with non-parametric Mann-Whitney U-tests.
Results
Species biomass and competitive strength
We found L. grandiflora to produce more biomass than L. peploides at the end of each trophic experiment, both in monocultures and in mixtures (Eutrophic: monoculture p<0.001, mixed culture p<0.0001; Mesotrophic: monoculture p<0.05, mixed

J. Gérard et al.
160
Figure 1. Effect of phosphorus reduction on L. grandiflora and L. peploides total biomass production in monocultures (grey boxes) and mixtures (white boxes). □ represents the median, the box the interquartile range and the vertical lines the minimum and the maximum non-outlier values. Outliers are represented by ○. Significant differences are indicated as ** p < 0.01, * p < 0.05.
culture p<0.0001). For both nutrient levels, the biomass of L. grandiflora remained similar in monocultures and in mixtures. The biomass of L. peploides however was reduced in mixed cultures compared to monocultures in both trophic conditions, indicating that L. grandiflora was the superior competitor in the tested conditions (Figure 1). The relative competitive strength of both species and biomass ratio of L. grandiflora / L. peploides in mixtures was not affected by a phosphorus reduction, confirming the superiority of L. grandiflora under the conditions of this study.
Strategy
Relative growth rate
In both trophic conditions, Ludwigia grandiflora generally had a higher biomass relative growth rate (RGR) than L. peploides, in monocultures and in mixtures (Figure 2A). The length RGR was higher for L. grandiflora in monocultures and mixtures in eutrophic condition, but in mesotrophic condition both species performed similarly (Figure 2B). When compared between monocultures and mixtures in eutrophic condition, both species’ weight RGR was higher in monocultures than in mixtures. In mesotrophic condition however, the two species performed similarly in monocultures and mixtures. A comparison of the length RGR indicated that both species performed similar in mixtures and monocultures for both trophic conditions. Growing in nutrient reduced conditions had a negative effect on both species length and weight RGR, except for L. grandiflora weight RGR in mixtures.
Plant traits
Six plant traits were measured during the experiment (Table 1). When compared between species, we found that in both trophic conditions, values of traits of L. grandiflora were almost always higher than the ones of L. peploides. L. grandiflora produced longer stems (Eutrophic U= 790.5, p<0.0001; Mesotrophic: U= 732.0, p<0.001) and more leaves per stem (Eutrophic U= 443.5, p<0.0001; Mesotrophic U= 486.5, p<0.0001). The average number of branches produced was higher for L. grandiflora in eutrophic conditions, however in mesotrophic conditions both species produced similar amounts (Eutrophic U=1119.5, p<0.01; Mesotrophic U=1339.5, p>0.05). Leaf length was the highest for L. grandiflora, in both nutrient conditions (Eutrophic U= 200.0, p<0.0001; Mesotrophic U= 188.0, p< 0.0001). The same results were found for leaf width (Eutrophic: U= 481.0, p<0.0001; Mesotrophic U= 655.0, p<0.001). The root/shoot ratio was higher for L. grandiflora in both nutrient conditions (Eutrophic U=385.5, p<0.0001; Mesotrophic U=1500.0, p<0.0001).
Growing the plants in nutrient reduced conditions (mesotrophic) had a significant negative impact, compared to eutrophic conditions, on almost all plant traits (Table 1). Both species’ stem length and number of branches were reduced, however the number of leaves per stem increased. Leaves in eutrophic condition were longer for both species and wider for L. grandiflora. The root/shoot ratio was also higher for both species in eutrophic compared to mesotrophic condition.
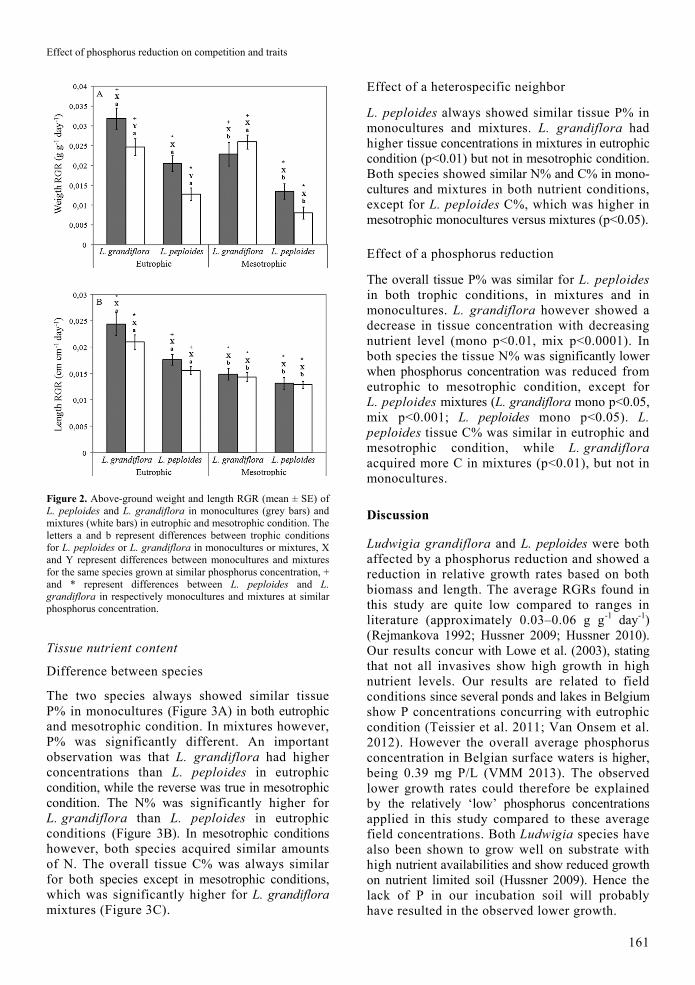
Effect of phosphorus reduction on competition and traits
161
Figure 2. Above-ground weight and length RGR (mean ± SE) of L. peploides and L. grandiflora in monocultures (grey bars) and mixtures (white bars) in eutrophic and mesotrophic condition. The letters a and b represent differences between trophic conditions for L. peploides or L. grandiflora in monocultures or mixtures, X and Y represent differences between monocultures and mixtures for the same species grown at similar phosphorus concentration, + and * represent differences between L. peploides and L. grandiflora in respectively monocultures and mixtures at similar phosphorus concentration.
Tissue nutrient content
Difference between species
The two species always showed similar tissue P% in monocultures (Figure 3A) in both eutrophic and mesotrophic condition. In mixtures however, P% was significantly different. An important observation was that L. grandiflora had higher concentrations than L. peploides in eutrophic condition, while the reverse was true in mesotrophic condition. The N% was significantly higher for L. grandiflora than L. peploides in eutrophic conditions (Figure 3B). In mesotrophic conditions however, both species acquired similar amounts of N. The overall tissue C% was always similar for both species except in mesotrophic conditions, which was significantly higher for L. grandiflora mixtures (Figure 3C).
Effect of a heterospecific neighbor
L. peploides always showed similar tissue P% in monocultures and mixtures. L. grandiflora had higher tissue concentrations in mixtures in eutrophic condition (p<0.01) but not in mesotrophic condition. Both species showed similar N% and C% in mono-cultures and mixtures in both nutrient conditions, except for L. peploides C%, which was higher in mesotrophic monocultures versus mixtures (p<0.05).
Effect of a phosphorus reduction
The overall tissue P% was similar for L. peploides in both trophic conditions, in mixtures and in monocultures. L. grandiflora however showed a decrease in tissue concentration with decreasing nutrient level (mono p<0.01, mix p<0.0001). In both species the tissue N% was significantly lower when phosphorus concentration was reduced from eutrophic to mesotrophic condition, except for L. peploides mixtures (L. grandiflora mono p<0.05, mix p<0.001; L. peploides mono p<0.05). L. peploides tissue C% was similar in eutrophic and mesotrophic condition, while L. grandiflora acquired more C in mixtures (p<0.01), but not in monocultures.
Discussion
Ludwigia grandiflora and L. peploides were both affected by a phosphorus reduction and showed a reduction in relative growth rates based on both biomass and length. The average RGRs found in this study are quite low compared to ranges in literature (approximately 0.03–0.06 g g-1 day-1) (Rejmankova 1992; Hussner 2009; Hussner 2010). Our results concur with Lowe et al. (2003), stating that not all invasives show high growth in high nutrient levels. Our results are related to field conditions since several ponds and lakes in Belgium show P concentrations concurring with eutrophic condition (Teissier et al. 2011; Van Onsem et al. 2012). However the overall average phosphorus concentration in Belgian surface waters is higher, being 0.39 mg P/L (VMM 2013). The observed lower growth rates could therefore be explained by the relatively ‘low’ phosphorus concentrations applied in this study compared to these average field concentrations. Both Ludwigia species have also been shown to grow well on substrate with high nutrient availabilities and show reduced growth on nutrient limited soil (Hussner 2009). Hence the lack of P in our incubation soil will probably have resulted in the observed lower growth.

J. Gérard et al.
162
Figure 3. Tissue P, N and C concentration (mg/g dry weight) of L. peploides (white boxes) and L. grandiflora (black boxes) in monocultures and mixtures in eutrophic and mesotrophic condition. □ represents the median, the box the interquartile range and the vertical lines the minimum and the maximum non-outlier values. Outliers are represented by ○, extremes by x. Significant differences between species are indicated as *** p < 0.001, ** p < 0.01, * p < 0.05, ns non significant.
The search for traits associated with invasiveness is crucial since a common set of traits is elusive, particularly for aquatic plants (Daehler 2003; Mony et al. 2006; Richardson and Pysek 2006). Although this approach is mainly used for explaining differences between natives and invasives, we believe it is also valid for explaining different degrees of alien species’ invasiveness. Ludwigia grandiflora and Ludwigia peploides are two very similar species which are hard to distinguish in the absence of flowers (Dandelot et al. 2005b). Even though being almost identical species, we showed that once placed in competition, both species performed differently. Under the conditions of this study, L. grandiflora was the overall better competitor, in high phosphorus concentration as well as in lower concentration. Several studies have investigated traits in native and invasive species and found invaders to have higher growth rates, leaf area, leaf mass ratio, nutrient uptake rates and other traits (Barrat-Segretain et al. 2002; Feng et al. 2007; van Kleunen et al. 2010; Matzek 2012). Our results support that Ludwigia species possess several of these traits which might explain their excessive spread in such a large range of countries (Aboucaya 1999; Cronk and Fuller 1995). When compared between species, L. grandiflora exhibited higher values of traits compared to L. peploides. L. grandiflora stems were longer, and produced more branches and leaves. Leaves were also larger, thereby having a larger photosynthetic surface. To predict future invasions and to control invasive species that are already present, it is important to study trait responses under changing conditions (Drenovsky et al. 2012) such as nutrient loading to surface water, since it is of great relevance to water quality standards (Wersal and Madsen 2011). We found that reduced phosphorus inputs in the water column affected not only species RGR but also other traits. Especially the smaller number of branches and stem length was highly reduced. This was also observed by Mony et al. (2006), who found Ranunculus peltatus traits to depend on P-availability.
When growing in high P, L. grandiflora had higher tissue P in mixture than in monoculture, even though biomass remained similar, indicating that L. grandiflora had an enhanced phosphorus accumulation capacity in the presence of L. peploides. Additionally, L. grandiflora had higher P% than L. peploides in eutrophic conditions, probably by having superior competitive ability for nutrient uptake (Aerts et al. 1999). L. peploides

Effect of phosphorus reduction on competition and traits
163
Table 1. Effect of a water column phosphorus reduction on L. grandiflora and L. peploides traits. Species are indicated by G= L. grandiflora and P= L. peploides .Values given are average values. Significant differences between nutrient levels are shown as **** p < 0.0001, *** p < 0.001, ** p < 0.01, * p < 0.05, ns non significant.
Trait Species Eutrophic condition Mesotrophic condition p U
Final length (cm) G 39.87 28.23 **** 790.5 P 29.48 24.24 **** 732.0 Number of branches G 6.12 2.68 *** 1030.5 P 3.19 2.13 ** 1114.5 Number of leaves G 19.29 21.50 * 1135.5 P 14.07 16.09 * 899.0 Leaf length (cm) G 2.92 2.28 **** 746.0 P 1.52 1.24 ** 735.0 Leaf width (cm) G 1.97 1.46 **** 799.5 P 1.28 1.07 ns 685.5 Biomass (g) G 0.32 0.25 ns 3340.5 P 0.12 0.08 ** 2575.0 Root:shoot ratio G 0.25 0.14 ** 2660.5
P 0.08 0.07 * 2480.5
on the other hand always had similar P accumulation in monocultures and mixtures, whatever the P levels, even though biomass was always reduced due to the presence of L. grandiflora. In mesotrophic conditions, L. peploides even had higher internal P compared to L. grandiflora in mixtures.
Plants with the ability of high nutrient storage are highly competitive (Garbey et al. 2004; Thiébaut 2005). Internal nutrient concentrations can be used to compare aggressiveness and competitiveness in nutrient uptake at different nutrient concen-trations, and are even recommended over nutrient uptake studies for evaluating nutrient competitive-ness (Gerloff 1975). P thresholds have to be accounted for (Demars and Edwards 2007), since they indicate interesting variations in the nutritional requirements of organisms, even within species belonging to the same taxonomic group (Gerloff 1975). For aquatic species, estimated critical P nutrient concentrations in tissues having 95% of the maximum yield (= final standing biomass) and 95% of the maximum growth are 0.10 (±0.03 SD) % DW and 0.16 (±0.05 SD) % DW, respectively (Gerloff 1975; Demars and Edwards 2007). Our results showed both species to always contain P levels above the maximum biomass threshold. However, in mesotrophic condition 50% of the L. grandiflora internal P concentration was lower than the maximum growth threshold, indicating that P could have been growth-limiting. However this value needs to be interpreted with care since P thresholds are species-specific (Gerloff 1975). Demars and Edwards (2007) also found external nutrient availability for aquatic
vascular plants to influence tissue nutrient concentrations. Additionally, a low P availability has often been found to reduce tissue P concentrations of other aquatic plants (Xu et al. 2006; Rejmankova and Snyder 2008; Zhang and Liu 2011).
L. grandiflora was probably able to take up more nutrients than L. peploides due to the production of more and longer roots. Both species produced branched downwards growing roots and unbranched spongy upwards growing roots. An important observation in our study was that in both trophic conditions, L. peploides showed red leaves and reddish downward growing roots not embedded in the soil. The coloration of the roots is often found in both Ludwigia species (Ellmore 1981; Hussner 2010; van Valkenburg pers. comm.). This could be caused by an accumulation of anthocyanin, a pigment with a wide range of functions associated to UV absorption and nutrient stress (Stewart et al. 2001; Kliebenstein 2004). Anthocyanin is known to accumulate in plants when P is limiting (Jiang et al. 2007). Anthocyanin can also accumulate to protect the plant from high-light damage by absorbing light (Chalker-Scott 1999). Since roots were only colored above-ground and individuals in field conditions growing out of the water have also shown pink roots (van Valkenburg pers. comm.), this explanation seems more appropriate. Additionally, Ellmore (1981) found the nodes of L. peploides plants to show red pigmentation on the dorsal sides illuminated by the sun, while the shaded ventral sides remained green.

J. Gérard et al.
164
Both species also produced spongy white upwards growing roots, known as pneumatophores, which improve gas exchange during anoxic conditions, occurring mostly in summer (Ellmore 1981; Dandelot et al. 2005a). These roots were frequently observed for L. grandiflora, but not often for L. peploides. Both species have also been found to release allelopathic chemicals in the environment (Dandelot et al. 2008), which may have played a role in the competitive outcome. Other studies also mention complex interactions with bacteria that produce toxic compounds (Ahmed et al. 2005; Dandelot et al. 2005a; Dandelot et al. 2008). However this toxicity due to the proliferation of bacteria, has been found to be higher in L. grandiflora compared to L. peploides stands (Dandelot et al. 2005a).
Conclusion
Since eutrophication by increased phosphorus inputs has a facilitating effect on invasives, a P reduction in the water column is needed to reduce invasive species’ performance. This study has demonstrated that a water column phosphorus reduction clearly affects invasive Ludwigia species growth, competition and traits. The effect on traits is very important in controlling both species, since a reduction in stem length and branch number could reduce the matting effect of both species and even lead to a reduction in plant parts able to fragment and spread. Targets for total P in water in Belgian water quality legislation is around 0.03–0.14 mg P/L depending on the watercourse types. Our results show that a water column phosphorus reduction to 0.03 mg P/L has a large effect on Ludwigia species and therefore support the lower nutrient threshold.
In most eutrophicated lakes and ponds sediments are heavily nutrient loaded which may influence the results of a water column P input reduction. Sediment removal may be applied to counter these problems but should be applied together with load reduction (Yenilmez and Aksoy 2013). The relevance of our study applies to lakes and ponds where the water column has been loaded with phosphorus but the sediment is low in phosphorus (for instance after sediment removal), or where plants have no contact with the nutrient-rich sediment. Further research with nutrient-loaded sediments should be performed to clarify the influence of sediment on a water column phosphorus reduction.
Acknowledgements
We would like to thank D. Maes, J. Appah and J. van Valkenburg for supplying L. peploides. We are grateful to the anonymous reviewers for their comments on the manuscript. This PhD research is funded by a scholarship provided by the government agency for Innovation by Science and Technology and by the Vrije Universiteit Brussel (BAS42).
References
Aboucaya A (1999) Premier bilan dune enqueˆ te nationale 363 destiné à identifier les xénophytes invasifs sur le territoire métropolitain français (Corse comprise). Bulletin de la Société Botanique du Centre-Ouest 19: 463–482
Aerts R (1999) Interspecific competition in natural plant communities: mechanisms, trade-offs and plant–soil feedbacks. Journal of Experimental Botany 50: 29–37, http://dx.doi.org/10.1093/jxb/50.330.29
Ahmed F, Selim MST, Shilpi JA (2005) Antibacterial activity of Ludwigia adscendens. Fitoterapia 76: 473–475, http://dx.doi.org/10.1016/j.fitote.2005.04.007
Ancrenaz K, Dutartre A (2002) Cartographie des Jussies (Ludwigia spp) en France métropolitaine. Cemagref, Unité de Recherche Qualité des Eaux, étude n 73, Département Gestion des Milieux Aquatiques, Ed. CEMAGREF, 18 pp
Barrat-Segretain MH, Elger A, Sagnes P, Puijalon S (2002) Comparison of three life-history traits of invasive Elodea canadensis Michx. and Elodea nuttallii (Planch.) H. St. John. Aquatic Botany 74: 299–313, http://dx.doi.org/10.1016/S0304-3770(02)00106-7
Bauchau V, Lejeune A, Bouharmont J (1984) Maintien et expansion de Ludwigia uruguayensis (Camb.) Hara en Brabant. Dumortiera 28: 8–9
Brunel S (2009) Pathway analysis: aquatic plants imported in 10 EPPO countries. EPPO Bulletin 39: 201–213, http://dx.doi.org/10.1111/j.1365-2338.2009.02291.x
Burns JH (2004) A comparison of invasive and non-invasive dayflowers (Commelinaceae) across experimental nutrient and water gradients. Diversity and Distributions 10: 387–397, http://dx.doi.org/10.1111/j.1366-9516.2004.00105.x
Cahill DM, Rookes JE, Wilson BA, Gibson L, Mcdougall KL (2008) Phytophthora cinnamomi and Australia's biodiversity: impacts, predictions and progress towards control. Australian Journal of Botany 56: 279–310, http://dx.doi.org/10.1071/BT 07159
Chadwell TB, Engelhardt K (2008) Effects of pre-existing submersed vegetation and propagule pressure on the invasion success of Hydrilla verticillata. Journal of Applied Ecology 45: 515–523, http://dx.doi.org/10.1111/j.1365-2664.2007.01384.x
Chalker-Scott L (1999) Environmental significance of anthocyanins in plant stress responses. Photochemistry and Photobiology 70: 1–9, http://dx.doi.org/10.1111/j.1751-1097.1999. tb01944.x
Chapin FS (1991) Integrated Responses of Plants to Stress. BioScience 41: 29–36, http://dx.doi.org/10.2307/1311538
Craine JM, Fargione J, Sugita S (2005) Supply pre-emption, not concentration reduction, is the mechanism of competition for nutrients. New Phytologist 166: 933–940, http://dx.doi.org/10.11 11/j.1469-8137.2005.01386.x
Cronk QCB, Fuller JL (1995) Plant invaders: the threat to natural ecosystems. Chapman and Hall, London, UK, 241 pp
Daehler CC (2003) Performance comparisons of co-occurring native and alien invasive plants: implications for conservation and restoration. Annual Review of Ecology, Evolution, and Systematics 34: 183–211, http://dx.doi.org/10.1146/annurev. ecolsys.34.011802.132403

Effect of phosphorus reduction on competition and traits
165
Dandelot S (2004) Les Ludwigia spp. invasives du Sud de la France: Historique, Biosystématique, Biologie et Ecologie. PhD Thesis, University Aix-Marseille-III, France, 207 pp
Dandelot S, Matheron R, le Petit J, Verlaque R, Cazaubon A (2005a) Variations temporelles des paramètres physico-chimiques et microbiologiques de trois écosystèmes aquatiques (Sud-Est de la France) envahis par des Ludwigia. Comptes Rendus Biologies 328: 991–999, http://dx.doi.org/10. 1016/j.crvi.2005.09.007
Dandelot S, Verlaque R, Dutarte A, Cazaubon A (2005b) Ecological, dynamic and taxonomic problems due to Ludwigia (Onagraceae) in France. Hydrobiologia 551: 131–136, http://dx.doi.org/10.1007/s10750-005-4455-0
Dandelot S, Robles C, Pech N, Cazaubon A, Verlaque R (2008) Allelopathic potential of two invasive alien Ludwigia spp. Aquatic Botany 88: 311–316, http://dx.doi.org/10.1016/j.aquabot. 2007.12.004
Demars BOL, Edwards AC (2007) Tissue nutrient concentrations in freshwater aquatic macrophytes: high inter-taxon differences and low phenotypic response to nutrient supply. Freshwater Biology 52: 2073–2086, http://dx.doi.org/10.1111/j. 1365-2427.2007.01817.x
Drenovsky RE, Grewell BJ, D’Antonio CM, Funk JL, James JJ, Molinari N, Parker IM, Richards CL (2012) A functional trait perspective on plant invasion. Annals of Botany 110: 141–153, http://dx.doi.org/10.1093/aob/mcs100
Dukes JS, Mooney HA (1999) Does global change increase the success of biological invaders? Trends in Ecology & Evolution 14: 135–139, http://dx.doi.org/10.1016/S0169-5347(98) 01554-7
Dutartre A, Haury J, Planty-Tabacchi AM (1997) introductions de macrophytes aquatiques et riverains dans les hydrosystèmes Français métropolitains: essai de bilan. Bulletin Francais De La Peche Et De La Pisciculture 344/345: 407–426, http://dx.doi.org/10.1051/kmae:1997038
Ellmore GS (1981) Root dimorphism in Ludwigia peploides (Onagraceae): structure and gas content of mature roots. American Journal of Botany 68: 557–568, http://dx.doi.org/ 10.2307/2443032
Elser JJ, Bracken ME, Cleland EE, Gruner DS, Harpole WS, Hillebrand H, Ngai JT, Seabloom EW, Shurin JB, Smith JE (2007) Global analysis of nitrogen and phosphorus limitation of primary producers in fresh-water, marine and terrestrial ecosystems. Ecology Letters 10: 1135–1142, http://dx.doi.org/ 10.1111/j.1461-0248.2007.01113.x
EPPO (European and Mediterranean Plant Protection Organi-zation) (2011) EPPO Data sheet on invasive alien plants Ludwigia grandiflora and L. peploides Onagraceae – Water primroses. EPPO Bulletin 41: 414–418, http://dx.doi.org/ 10.1111/j.1365-2338.2011.02511.x
Fan SF, Liu CH, Yu D, Xie D (2013) Differences in leaf nitrogen content, photosynthesis, and resource-use efficiency between Eichhornia crassipes and a native plant Monochoria vaginalis in response to altered sediment nutrient levels. Hydrobiologia 711: 129–137, http://dx.doi.org/10.1007/s10750-013-1471-3
Feng Y, Wang J, Sang W (2007) Biomass allocation, morphology and photosynthesis of invasive and noninvasive exotic species grown at four irradiance levels. Acta Oecologica 31: 40–47, http://dx.doi.org/10.1016/j.actao.2006.03.009
Feng YL, Fu GL (2008) Nitrogen allocation, partitioning and use efficiency in three invasive plant species in comparison with their native congeners. Biological Invasions 10: 891–902, http://dx.doi.org/10.1007/s10530-008-9240-3
Fogarti G, Facelli JM (1999) Growth and competition of Cytisus scoparius, an invasive shrub and Australian native shrubs. Plant Ecology 144: 27–35, http://dx.doi.org/10.1023/A:10098 08116068
Funk JL, Vitousek PM (2007) Resource-use efficiency and plant invasion in low resource systems. Nature 446: 1079–1081l, http://dx.doi.org/10.1038/nature05719
Garbey C, Murphy KJ, Thiébaut G, Muller S (2004) Variation in P-content in aquatic plant tissues offers an efficient tool for determining plant growth strategies along a resource gradient. Freshwater Biology 49: 346–356, http://dx.doi.org/10.1111/j. 1365-2427.2004.01188.x
Gerloff GC (1975) Nutritional ecology of nuisance aquatic plants. U.S. Environmental Protection Agency, Corvallis, Oregon, USA, 78 pp
Gopal B, Goel U (1993) Competition and allelopathy in aquatic plant communities. The Botanical Review 59: 155–186, http://dx.doi.org/10.1007/BF02856599
Grotkopp E, Rejmanek M (2007) High seedling relative growth rate and specific leaf area are traits of invasive species: Phylogenetically independent contrasts of woody angiosperms. American Journal of Botany 94: 526–532, http://dx.doi.org/10.3732/ajb.94.4.526
Hastwell GT, Daniel AJ, Vivian-Smith G (2008) Predicting invasiveness in exotic species: do subtropical native and invasive exotic aquatic plants differ in their growth responses to macronutrients? Diversity and Distributions 14: 243–251, http://dx.doi.org/10.1111/j.1472-4642.2007.00367.x
Haury J, Hudin S, Matrat R, Anras L et al. (2010) Manuel de gestion des plantes exotiques envahissant les milieux aquatiques et les berges du bassin Loire-Bretagne, Fédération des conservatoires d'espaces naturels, 136 pp
Hoagland R, Arnon I (1950) The water-culture method for growing plants without soil. Circular 347: 1–32
Hulme PE (2009) Trade, transport and trouble: managing invasive species pathways in an era of globalization. Journal of Applied Ecology 46: 10–18, http://dx.doi.org/10.1111/j.1365-2664.2008.01600.x
Hussner A (2009) Growth and photosynthesis of four invasive aquatic plant species in Europe. Weed Research 49: 506–515, http://dx.doi.org/10.1111/j.1365-3180.2009.00721.x
Hussner A (2010) Growth response and root system development of the invasive Ludwigia grandiflora and Ludwigia peploides to nutrient availability and water level. Fundamental and Applied Limnology 177: 189–196, http://dx.doi.org/10.1127/ 1863-9135/2010/0177-0189
Hussner A (2012) Alien aquatic plant species in European countries. Weed Research 52: 297–306, http://dx.doi.org/ 10.1111/j.1365-3180.2012.00926.x
Jiang C, Gao X, Liao L, Harberd NP, Fu X (2007) Phosphate starvation root architecture and anthocyanin accumulation responses are modulated by the gibberellin-DELLA signaling pathway in Arabidopsis. Plant Physiology 145: 1460–1470, http://dx.doi.org/10.1104/pp.107.103788
Johengen T (1996) Standard operating procedures for determining total phosphorus, available phosphorus, and biogenic silica concentrations of lake Michigan sediments and sediment trap material. In: Lake Michigan Mass Balance Metodhs Compedium, 1997, Volume 3 Metals, Conventionals, Radio-chemistry and Biomonitoring Sample Analysis Techniques, GLERL-SED NUTRIENT-96, pp 3–307–3–312
Kliebenstein DJ (2004) Secondary metabolites and plant/environment interactions: a view through Arabidopsis thaliana tinged glasses. Plant, Cell and Environment 27: 675–684, http://dx.doi.org/10.1111/j.1365-3040.2004.01180.x
Koerselman W, Meuleman AFM (1996) The Vegetation N:P Ratio: a New Tool to Detect the Nature of Nutrient Limitation. Journal of Applied Ecology 33: 1441–1450, http://dx.doi.org/10.2307/2404783
Lewis WMJ, Wurtsbaugh WA (2008) Control of lacustrine phytoplankton by nutrients: erosion of the phosphorus paradigm. International Review of Hydrobiology 93: 446–465, http://dx.doi.org/10.1002/iroh.200811065

J. Gérard et al.
166
Lowe PN, Lauenroth WK, Burke IC (2003) Effects of nitrogen availability on competition between Bromus tectorum and Bouteloua gracilis. Plant Ecology 167: 247–254, http://dx.doi.org/10.1023/A:1023934515420
Mack RN, Simberloff D, Lonsdale WM, Evans H, Clout M, Bazzaz FA (2000) Biotic invasions: causes, epidemiology, global consequences, and control. Ecological Applications 10: 689–710, http://dx.doi.org/10.1890/1051-0761(2000)010[0689: BICEGC]2.0.CO;2
Matrat R, Anras L, Vienne L, Hervochon F, Pineau C, Bastian S, Dutartre A, Haury J, Lambert E, Gilet H, Lacroix P, Maman L (2006) Gestion des plantes exotiques envahissantes en cours d’eau et zones humides – Guide technique. Comité des Pays de la Loire, Nantes, France, 68 pp
Matzek V (2012) Trait values, not trait plasticity, best explain invasive species’ performance in a changing environment. PLoS ONE 7: e48821, http://dx.doi.org/10.1371/journal.pone. 0048821
Mony C, Thiebaut G, Muller S (2006) Changes in morphological and physiological traits of the freshwater plant Ranunculus peltatus with the phosphorus bioavailability. Plant Ecology 191: 109–118, http://dx.doi.org/10.1007/s11258-006-9219-z
Nieuwenhuize J, Maas Y, Middelburg JJ (1994) Rapid analysis of organic carbon and nitrogen in particulate materials. Marine Chemistry 45: 217–224, http://dx.doi.org/10.1016/0304-4203(94) 90005-1
Njambuya J, Triest L (2010) Comparative performance of invasive alien Eichhorniacrassipes and native Ludwigia stolonifera under non-limiting nutrient conditions in Lake Naivasha, Kenya. Hydrobiologia 656: 221–231, http://dx.doi.org/10.1007/s10750-010-0430-5
Olde Venterink H, Güsewell S (2010) Competitive interactions between two meadow grasses under nitrogen and phosphorus limitation. Functional Ecology 24: 877–886, http://dx.doi.org/ 10.1111/j.1365-2435.2010.01692.x
Pella E (1990) Elemental organic analysis: Part 2. State of the art. American Laboratory 22: 28–32
Pysek P, Richardson DM (2007) Traits associated with invasi-veness in alien plants: Where do we stand? In: Nentwig W (ed), Biological Invasions, Berlin/Heidelberg, Germany, Vol. 193, pp 97–125, http://dx.doi.org/10.1007/978-3-540-36920-2_7
Rejmánková E (1992) Ecology of creeping macrophytes with special reference to Ludwigia peploides (H. B. K.) Raven. Aquatic Botany 43: 283–299, http://dx.doi.org/10.1016/0304-3770(92)90073-R
Rejmánková E, Snyder JM (2008) Emergent macrophytes in phosphorus limited marshes: do phosphorus usage strategies change after nutrient addition? Plant Soil 313: 141–153, http://dx.doi.org/10.1007/s11104-008-9687-0
Richardson DM, Pysek P (2006) Plant invasions: merging the concepts of species invasiveness and community invasibility. Progress in Physical Geography 30: 409–431, http://dx.doi.org/ 10.1191/0309133306pp490pr
Schindler DW (1977) Evolution of phosphorus limitation in lakes. Science 195: 260–262, http://dx.doi.org/10.1126/science.195. 4275.260
Sharpley AN, Daniel T, Sims T, Lemunyon J, Stevens R, Parry R (2003) Agricultural phosphorus and eutrophication, U.S. Department of Agriculture, Agricultural Research Service 149, 44 pp
Smith VH, Tilman GD, Nekola JC (1999) Eutrophication: impacts of excess nutrient inputs on freshwater, marine, and terrestrial ecosystems. Environmental Pollution 100: 179–196, http://dx.doi.org/10.1016/S0269-7491(99)00091-3
Spitters CJT (1983) An alternative approach to the analyses of mixed cropping experiments. 1. Estimation of competition effects. Netherlands Journal of Agricultural Science 31: 1–11
Stewart AJ, Chapman W, Jenkins GI, Graham I, Martin T, Crozier A (2001) The effect of nitrogen and phosphorus deficiency on Xavonol accumulation in plant tissues. Plant, Cell and Environment 24: 1189–1197, http://dx.doi.org/10.10 46/j.1365-3040.2001.00768.x
Stiers I, Crohain N, Josens G, Triest L (2011) Impact of three aquatic invasive species on native plants and macroinvertebrates in temperate ponds. Biological Invasions 13: 2715–2726, http://dx.doi.org/10.1007/s10530-011-9942-9
Thiébaut G (2005) Does competition for phosphate supply explain the invasion pattern of Elodea species? Water Research 39: 3385–3393, http://dx.doi.org/10.1016/j.watres.2005. 05.036
Thouvenot L, Puech C, Martineza L, Haury J, Thiébaut G (2013) Strategies of the invasive macrophyte Ludwigia grandiflora in its introduced range: Competition, facilitation or coexistence with native and exotic species? Aquatic Botany 107: 8–16, http://dx.doi.org/10.1016/j.aquabot.2013.01.003
Teissier S, Peretyatko A, De Backer S, Triest L (2011) Strength of phytoplankton–nutrient relationship: evidence from 13 biomanipulated ponds. Hydrobiologia 689: 147–159, http://dx.doi.org/10.1007/s10750-011-0726-0
Van TK, Wheeler GS, Center TD (1999) Competition between Hydrilla verticillata and Vallisneria americana as influenced by soil fertility. Aquatic Botany 62: 225–233, http://dx.doi.org/ 10.1016/S0304-3770(98)00100-4
van Kleunen M, Weber E, Fischer M (2010) A meta-analysis of trait differences between invasive and non-invasive plant species. Ecology Letters 13: 235–245, http://dx.doi.org/10.1111/ j.1461-0248.2009.01418.x
van Onsem S, Triest L, Crohain N, Josens G (2012) Beoordeling van de ecologische kwaliteit van waterlichamen in het Brussels Hoofdstedelijk Gewest in uitvoering van de Europese Kaderrichtlijn Water - Macrofyten, fytobenthos, fytoplankton & macro-invertebraten. Brussels Instituut voor Milieubeheer (BIM) Report, 113 pp
Verloove F (2006) Catalogue of neophytes in Belgium (1800-2005). Scripta Botanica Belgica 39: 3–89
VMM (2013) Vlaamse Milieumaatschappij (VMM). http://www. vmm.be/water/kwaliteit-oppervlaktewater/toestand-oppervlaktewater/ fysisch-chemische-toestand/nutrienten (Accessed 7 Sept. 2013)
Wersal RM, Madsen JD (2011) Influences of water column nutrient loading on growth characteristics of the invasive aquatic macrophyte Myriophyllum aquaticum (Vell.) Verdc. Hydrobiologia 665: 93–105, http://dx.doi.org/10.1007/s10750-011-0607-6
Xu ZG, Yan BX, He Y, Zhai JL, Song CC (2006) Effect of nitrogen and phosphorus on tissue nutrition and biomass of freshwater wetland plant in Sanjiang Plain, Northeast China. Chinese Geographical Science 16: 270–275, http://dx.doi.org/ 10.1007/s11769-006-0270-4
Yenilmez F, Aksoy A (2013) Comparison of phosphorus reduction alternatives in control of nutrient concentrations in Lake Uluabat (Bursa, Turkey): Partial versus full sediment dredging. Limnology 43: 1–9, http://dx.doi.org/10.1016/j.limno. 2012.05.003
Zhang X, Liu Z (2011) Interspecific competition effects on phosphorus accumulation by Hydrilla verticillata and Vallisneria natans. Journal of Environmental Science 23: 1274–1278




















