
Biphasic Mechanisms of Amphetamine Action at the Dopamine Terminal
Upload
independentCategory
view
1download
0
Osteoclastic Potential of Human CFU-GM: Biphasic Effect of GM-CSF
Jason M Hodge, Mark A Kirkland, Cathy J Aitken, Caryll M Waugh, Damian E Myers, Carolina M Lopez,Brendan E Adams, and Geoffrey C Nicholson
ABSTRACT: Human osteoclasts can be efficiently generated in vitro from cord blood mononuclear cells andderived CFU-GM colonies. However, CFU-M colonies are poorly osteoclastogenic. Short-term (2–48 h)treatment with GM-CSF stimulates osteoclast formation by proliferating precursors, whereas longer exposurefavors dendritic cell formation.
Introduction: Osteoclasts (OC) differentiate from cells of the myelomonocytic lineage under the influence ofmacrophage-colony stimulating factor (M-CSF) and RANKL. However, cells of this lineage can also differentiate tomacrophages and dendritic cells (DC) depending on the cytokine environment. The aims of this study were todevelop an efficient human osteoclastogenesis model and to investigate the roles of granulocyte macrophage-colonystimulating factor (GM-CSF) and M-CSF in human OC differentiation.Materials and Methods: A human osteoclastogenesis model, using as precursors colony forming unit-granulocytemacrophage (CFU-GM) colonies generated from umbilical cord mononuclear cells cultured in methylcellulose withGM-CSF, interleukin (IL)-3 and stem cell factor (SCF), has been developed. CFU-GM, colony forming unit-macrophage (CFU-M), or mixed colonies were cultured on dentine with soluble RANKL (sRANKL) and humanM-CSF with and without GM-CSF. Major endpoints were OC number, dentine resorption, and CD1a� DC clusters.Results: Osteoclast generation from CFU-GM and mixed colonies treated with M-CSF and sRANKL for 7–14 dayswas highly efficient, but CFU-M colonies were poorly osteoclastogenic under these conditions. Pretreatment ofprecursors with M-CSF for 7 or 14 days maintained the precursor pool, but OCs were smaller and resorption wasreduced. The effect of GM-CSF treatment was biphasic, depending on the timing and duration of exposure.Short-term treatment (2–48 h) at the beginning of the culture stimulated cell proliferation and enhanced OCformation up to 100%, independent of sRANKL. Longer-term GM-CSF treatment in the presence of sRANKL,however, inhibited OC generation with the formation of extensive CD1a� DC clusters, accompanied by downregu-lation of c-Fos mRNA. Delaying the addition of GM-CSF resulted in progressively less inhibition of osteoclasto-genesis.Conclusions: Human CFU-GM, but not CFU-M, progenitors have high osteoclastogenic potential. GM-CSF playsan important role in osteoclastogenesis and has a biphasic effect: Short-term treatment potentiates OC differentiationby proliferating precursors, but persistent exposure favors DC formation.J Bone Miner Res 2004;19:190–199. Published online on December 16, 2003; doi: 10.1359/JBMR.0301232
Key words: osteoclasts, granulocyte macrophage-colony stimulating factor, macrophage-colony stimulatingfactor, colony forming unit-granulocyte macrophage, differentiation
INTRODUCTION
OSTEOCLASTS (OC) are specialized cells responsible forbone resorption that differentiate from hematopoietic
precursors in the presence of RANKL and macrophage-colony stimulating factor (M-CSF). The precise lineage ofthe OC and its relationship to other hematopoietic cellsremains uncertain, although there is evidence that OC,monocyte/macrophages, and dendritic cells (DC) all derivefrom myelomonocytic precursors.(1–3) There have been con-
flicting reports that colony forming unit-macrophage (CFU-M)–derived cells have either low(4,5) or high(6) potential toform OCs. Preparations of differentiated cells of this lin-eage, including CD14� monocytes and macrophages, canform OCs but at very low efficiency,(7,8) and the possibilitythat immature cells present in these preparations are theactual OC progenitors has not been excluded.
Lineage commitment of OC is governed by a number ofgrowth factors and cytokines. M-CSF induces differentiationof macrophages but has also been demonstrated to be impor-tant in osteoclastogenesis(9–11) in conjunction with direct con-tact of OC precursors with cells of the stromal/osteoblast
The authors have no conflict of interest.
Department of Clinical and Biomedical Sciences: Barwon Health, The Geelong Hospital, University of Melbourne, Geelong, Victoria,Australia.
JOURNAL OF BONE AND MINERAL RESEARCHVolume 19, Number 2, 2004Published online on December 16, 2003; doi: 10.1359/JBMR.0301232© 2004 American Society for Bone and Mineral Research
190

lineage expressing RANKL.(12–14) Granulocyte macrophage-colony stimulating factor (GM-CSF) is also a macrophage-inducing cytokine that can stimulate OC progenitorproliferation(15–17) but inhibit differentiation,(15,17–19) and incombination with RANKL, promote DC formation.(17)
Previous studies have aided in the understanding of keystages in OC differentiation and the relative importance ofcytokines and growth factors to this process.(20,21) However,the majority of this knowledge derives from in vivo and invitro animal models, with limited data available from hu-man systems. The aim of this study was to develop anefficient and reliable in vitro human osteoclastogenesismodel to investigate the ontogeny of human OCs and theroles of colony-stimulating factors M-CSF and GM-CSF inOC differentiation.
MATERIALS AND METHODS
Eagle’s minimum essential media (MEM), penicillin/streptomycin, bromodeoxyuridine (BrdU), paraformalde-hyde, and nocodazole were purchased from Sigma. Non-essential amino acids (100�) and fetal bovine serum (FBS)were purchased from CSL Biosciences (Parkville, Austra-lia). Ficoll-Paque was purchased from Pharmacia Biotech.Methocult GF H4534 (Iscove’s MDM containing 1% meth-ylcellulose, 30% FBS, 1% bovine serum albumin [BSA],10�4 M 2-mercaptoethanol, 2 mM L-glutamine, 10 ng/mlrecombinant human (rh)GM-CSF, 10 ng/ml rhIL-3, and 50ng/ml rh stem cell factor [SCF]) was purchased from Stem-Cell Technologies. rhGM-CSF was purchased from Chemi-con. rhM-CSF was generously provided by Genetics Insti-tute. Soluble hRANKL coupled to GST fusion protein(sRANKL) was generously provided by Drs MatthewGillespie and Julian Quinn (St Vincent’s Institute of Med-ical Research, Melbourne, Australia). Monoclonal antibodyanti-BrdU-FITC was purchased from Becton Dickinson,anti-CD11b and CD14 from Immunotech and anti-CD1afrom Chemicon.
Cell preparation
Human umbilical cord blood and adult peripheral bloodwere obtained from healthy donors under a protocol ap-proved by Barwon Health Research and Ethics AdvisoryCommittee. A mononuclear cell fraction containing mono-cytes and lymphocytes from cord blood (CBMC) was iso-lated by Ficoll-Paque density gradient centrifugation.CD14� monocytes were prepared from adult peripheralblood as previously described.(22)
Colony-forming assays
CBMCs (1 � 106 cells/culture) were suspended in 1.0 mlMethocult GF H4534 in 35-mm dishes and incubated at37°C in humidified atmosphere of 5% CO2-air for 7–14days. Pooled colonies were harvested into PBS. Alterna-tively, individual colonies, visually identified as colonyforming unit-granulocyte macrophage (CFU-GM) orCFU-M, were picked using fine pipettes.
Osteoclast formation assays
Precursors (4 � 104–105 cells/well) were seeded into96-well tissue culture plates containing 4 � 4 � 0.1-mmslices of sperm whale dentine. CBMC cells were settled for1 h and the medium was aspirated to remove contaminatinglymphocytes. The cells were cultured in 200 �l MEMcontaining 10% FBS, nonessential amino acids, penicillin50 U/ml, streptomycin 50 �g/ml, 2 mM L-glutamine,M-CSF (25 ng/ml), and sRANKL (125 ng/ml). The cultureswere refreshed twice weekly by replacing additives in one-half volume of media. At 7, 14, or 21 days, the cells werefixed in 1% formalin and reacted for TRACP activity. Theformation of OCs was assessed by transmission light mi-croscopy and quantified using microcomputer image anal-ysis software (MCID-Imaging Research Inc., Ontario, Can-ada). In selected experiments, calcitonin receptor (CTR)expression was determined using 125I-salmon calcitonin(125I-sCT) autoradiography as previously described.(23)
Osteoclast resorption assays
Cells were removed from dentine slices by brief sonica-tion in chloroform:methanol 2:1. Xylene-free black ink wasapplied to the resorbed surface of each slice, and residualink was removed by wiping against absorbent paper. Re-sorption was assessed by transmission light microscopy,and the percentage area resorbed was quantified usingMCID software. This method was validated by comparisonwith scanning electron microscopy.(22)
Cyto- and immunocytochemical analyses
Pooled colonies were seeded into 96-well tissue cultureplates containing dentine slices as indicated previously andcultured in the presence of M-CSF (25 ng/ml) and sRANKL(125 ng/ml) with or without GM-CSF (10 ng/ml) for 7–14days. Cultures were fixed in 4% paraformaldehyde for 10minutes before subsequent staining. Fixed cells were testedfor �-napthyl acetate esterase (NSE) activity according tothe manufacturers instructions (Sigma). Indirect immuno-peroxidase cytochemistry (Dako LSAB2 System) was usedto determine the proportion of cells on the slices expressingthe markers CD1a, CD11b, or CD14. The formation ofCD1a� DC clusters was assessed by transmission lightmicroscopy and quantified using MCID software.
Real-time polymerase chain reaction analysis of c-FosmRNA expression
Pooled colonies were plated at 6 � 106 cells/75 cm2 flaskin the presence of M-CSF (25 ng/ml) or sRANKL (125ng/ml) with or without GM-CSF (10 ng/ml). Total RNAwas isolated at 24 and 72 h by lysing cells in RNAzolB andreverse transcribed (RT) to cDNA as previously de-scribed.(24) Real-time polymerase chain reaction (PCR;SYBR Green) was performed using an ABI PRISM 7700Sequence Detection System. Actin and c-Fos primers werefrom GENSET and were designed using Primer Expresssoftware (Applied Biosystems) sequences:
Actin P1 5�-GACAGGATGCAGAAGGAGATT-3�Actin P2 5�-TGATCCACATCTGCTGGAAGG-3�
191BIPHASIC EFFECT OF GM-CSF ON OSTEOCLASTOGENESIS

cDNA-Fos P1 5�-CGTTGTGAAGACCATGACAGG-AGG-3�
cDNA-Fos P2 5�-CATTTGGCTGCAGCCATCTTA-TTC-3�
Cell proliferation assays
Pooled colonies were plated at 7 � 105 cells/well in12-well plates containing M-CSF (25 ng/ml) with or with-out GM-CSF (10 ng/ml) and/or sRANKL (125 ng/ml) for16 h. Cells were then pulse-labeled with BrdU as previouslydescribed.(25) The proportion of cells in S-phase was deter-mined by flow cytometry using a FACSCalibur (BectonDickinson). To determine the effect of cell cycle arrest,precursors were treated with M-CSF with or without GM-CSF for 24 h in the presence or absence of nocodazole (40ng/ml).
Statistical analysis
Data are expressed as the mean � SE where applicable.Differences between groups were determined using one-way ANOVA, followed by Fisher’s multiple comparisontest, two-way ANOVA, followed by Tukey’s post hoc testor unpaired t-test. Statistical significance was set at p �0.05.
RESULTS
Osteoclastogenic potential of CD14�, CBMCs, andcells derived from CFU-GM colonies
When CD14� or CBMCs were incubated with M-CSFand sRANKL, large numbers of TRACP� multinucleatedcells and substantial resorption of dentine was seen at 21days (Table 1). Consistent with our previous experiencewith CD14�,(22) CBMCs are a superior source of humanOC precursors. Clonal expansion of CBMC in methylcel-lulose in the presence of GM-CSF, interleukin (IL)-3, andSCF to produce CFU-GM colonies enhanced the osteoclas-tic potential of the precursors and shortened the time re-quired to generate mature OC capable of resorption (Table1). Optimum OC generation was achieved at a seedingdensity of 4 � 104 cells/well in 96-well plates. A linearrelationship existed between the seeding density and OCsyield over the range 2–8 � 104 cells/well. Seeding densitiesbelow 2 � 104 cells/well did not yield significant OCs.After 2 h of settlement, 22% of the cells adhered, and at 14days, these cells were poorly osteoclastogenic compared
with the nonadherent fraction (nonadherent, 583.5 � 52.8OC/slice; adherent, 87.0 � 30.1 OC/slice; n � 3, p �0.0001). However, with continued culture for a further 7days, significant OC formation and resorption occurredfrom the adherent fraction (nonadherent, 419.0 � 50.0OC/slice, 95 � 2.9% resorption; adherent, 526.0 � 32.5OC/slice, 56.7 � 13.6% resorption).
Comparison of the osteoclastogenic potential ofCFU-GM and CFU-M colonies
Progenitor assays of CBMC cultured in Methocult GFH4534 produced predominantly CFU-GM colonies (�80%).CFU-M colonies (5–10%) and others of indeterminate originwere also present. Typical morphology of CFU-GM andCFU-M colonies, as previously described,(26,27) are shown inFig. 1A. CFU-GM colonies displayed densely compactedcores with abundant, small cells (plan area � 100 �m2) dif-fusing out from the core in radial patterns. CFU-M coloniescontained fewer, much larger cells (plan area � 300 �m2) withno dense core. Culture of selected CFU-GM or CFU-M colo-nies on dentine slices in the presence of sRANKL and M-CSFshowed that CFU-GM colonies were enriched for OC precur-sors compared with pooled colonies, whereas CFU-M colonieswere poorly osteoclastogenic (Fig. 1B). Because selection ofspecific colonies is time-consuming and the majority are CFU-GMs, pooled colonies were used in the remainder of theexperiments.
Characterization of osteoclastogenesis from pooledcolonies
When precursors were seeded at 4 � 104 cells/well andtreated with M-CSF and sRANKL, multinucleated cells(MNC) were first observed forming on plastic adjacent todentine slices at day 4 of culture. At day 7, numerousTRACP� mononuclear cells (plan area � 80 �m2) andTRACP� MNC were observed (Fig. 2A). At days 14 and21, the number of TRACP� mononuclear cells and MNCsdecreased. The progressive reduction in the number ofTRACP� MNC number was associated with a progressiveincrease in cell plan area over the 21 days, indicating thatthe formation of small TRACP� MNCs was occurring earlyin the culture period, with fusion of these cells to form largecells by day 21 (Figs. 2A and 2B). Resorption lacunae weredetected on dentine slices from day 7 (Fig. 2A). Approxi-mately 70% of the area resorbed by OCs over the 3-weekperiod occurred between the first and second weeks (Fig.
TABLE 1. OSTEOCLAST FORMATION BY HUMAN PERIPHERAL BLOOD CD14� MONOCYTES, UMBILICAL CORD BLOOD MONONUCLEAR CELLS, AND
CFU-GM–DERIVED CELLS
Source of cellsNo. of cells
seededCulture
duration (days)
TRACP�
MNC/dentineslice
Resorptionarea/dentine
slice (%)Average nuclei
number/OCEfficiency
(%)
CD14� 1 � 105 21 349 � 45 51 � 18 5.9 � 0.3 4.9CBMC 1 � 105 21 837 � 80 18 � 7 9.6 � 0.8 19.1CFU-GM 4 � 104 14 775 � 58 51 � 18 14.2 � 1.8 65.5
Results represent mean � SEM of three separate experiments. Efficiency of OC formation was calculated as the number of OC nuclei per dentine slicedivided by the total number of nuclei seeded per slice.
192 HODGE ET AL.

2B). The osteoclastogenic effect of sRANKL wasconcentration-dependent over the range of 8–125 ng/ml andwas inhibited by OPG in a concentration-dependent manner(IC50, 1 ng/ml; data not shown). Mature OCs, formed fromexposure to sRANKL and M-CSF for 14 days, were positivefor CTR as assessed by 125I-sCT autoradiography (data notshown). With M-CSF alone (25 ng/ml), without sRANKL,the cells remained mononuclear, adhered to dentine but didnot resorb it, and were TRACP� at day 14 (data not shown).The mononuclear cells could be divided into two pheno-types; smaller mononuclear cells predominated that wereCD14� and NSE� (3312 � 366 cells/slice; 8.2 � 1.2 �mdiameter; mean � SE) and macrophages (large mononu-clear cells with extensive cytoplasm) were positive forCD11b� and NSE� but CD14� (39.7 � 11.6 cells/slice;48.9 � 16.4 �m diameter; mean � SE).
Effects of pretreatment of precursors with M-CSF
Pretreatment of precursors with M-CSF for 7 or 14 daysbefore addition of sRANKL reduced the size, number ofnuclei, and resorption capacity of the generated OCs (Fig.3). Pretreatment for 7 or 14 days had similar effects on theseparameters; however, a 7-day, but not 14-day, pretreatmentresulted in a transient increase in the number of thesesmaller OCs (Fig. 3).
Effects of GM-CSF on osteoclastogenesis
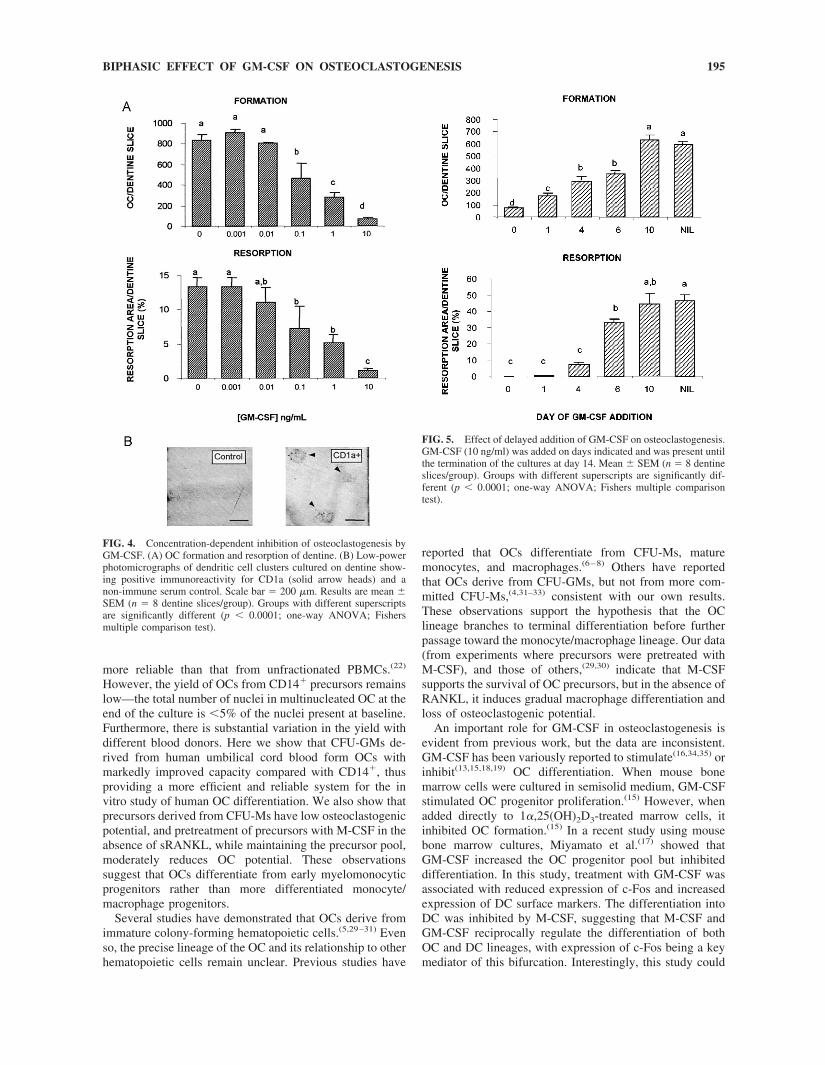
When GM-CSF was present for the entire 14-day periodin cultures treated with M-CSF and sRANKL, OC forma-tion and resorption activity was potently inhibited in aconcentration-dependent manner (Fig. 4A). The IC50 wasapproximately 0.1 ng/ml, with maximal inhibition occurringat 10 ng/ml. Clustering of mononuclear TRACP�/CD1a�
cells, indicative of DC activation,(28) was prominent ondentine slices treated with GM-CSF 10 ng/ml (59.8 � 12.0clusters/slice; 3186 � 326 �m2 cluster area; mean � SE),but such clusters were rarely seen and were much smallerwhen GM-CSF was not present (0.5 � 0.3 clusters/slice;396 � 255 �m2 cluster area; mean � SE). The morphologyand specific CD1a immunoreactivity of the clusters isshown in Fig. 4B. The inhibitory action of GM-CSF wasmaximal when added at the commencement of the cultureand progressively decreased with delayed addition (Fig. 5).No inhibition of OC number or resorption was observedwith addition at day 10, suggesting that GM-CSF does notact at the late stage of OC differentiation or on mature OCs.However, clustering of mononuclear TRACP� cells wasevident, suggesting that the cell pool contains discrete OCand DC precursor populations. When GM-CSF was addedat the commencement of the cultures and removed afterincreasing times ranging from 10 minutes to 6 days, abiphasic effect was observed (Fig. 6). Short-term treatmentstimulated both OC generation and resorption, whereastreatment for longer than 3 days inhibited as describedabove. Expression of c-Fos as assessed by real-time PCRwas not altered by the addition of GM-CSF at 24 h ofculture but was reduced 6.5-fold at 3 days of continuedexposure to GM-CSF. The stimulatory effect of short-termGM-CSF treatment was maximal at 2 days of treatment,when OC number was increased by 75% and resorption by172%. To determine whether the stimulatory effects ofGM-CSF were dependent on the presence of sRANKL inthe culture, precursors were pretreated with M-CSF alone orwith GM-CSF and M-CSF for 2, 24, or 72 h. After thispretreatment, media was replaced with sRANKL andM-CSF for 14 days. We found that short-term GM-CSFpretreatment also markedly stimulated OC generation andresorption under these conditions (Fig. 7).
To investigate the mechanism of GM-CSF–induced stim-ulation, OC precursors were incubated with M-CSF, with orwithout GM-CSF and/or sRANKL, for 16 h, and pulsed-labeled with BrdU for 30 minutes. Treatment with GM-CSFincreased cell proliferation in the nonadherent fraction by82% compared with cultures treated with M-CSF alone, andby 23% compared with those treated with M-CSF andsRANKL (Table 2). sRANKL also caused cell proliferation,
FIG. 1. Relative osteoclastogenic potential of CFU-GM and CFU-M.(A) Low power (left, bar � 500 �m) and high power (right, bar � 100�m) magnification of CFU-GM and CFU-M colonies. (B) Comparisonof OC formation and resorption from pooled, CFU-GM, and CFU-Mcolonies. Mean � SEM (n � 4 dentine slices/group). Groups withdifferent superscripts are significantly different (p � 0.001; one-wayANOVA; Fishers multiple comparison test).
193BIPHASIC EFFECT OF GM-CSF ON OSTEOCLASTOGENESIS

but maximal cell proliferation caused by GM-CSF wassRANKL independent. Treatment with GM-CSF did notalter the number of cells adherent to dentine after 1 day ofculture, but there was a small but significant (15%) increasein adherent cell number at day 2 (data not shown). Cotreat-ment of OC precursors with the reversible cell cycle arrestagent, nocodazole, had no significant effect on cell prolif-eration in control (M-CSF treated) cultures but completelyblocked the increase in proliferation produced by short-term
GM-CSF treatment (Table 2). Similarly, the stimulation ofOC formation and dentine resorption observed with short-term GM-CSF pretreatment was prevented by nocodazole(Fig. 8).
DISCUSSION
Previously we have shown that OC generation from theCD14� fraction of adult PBMCs is both more efficient and
FIG. 3. Effect of M-CSF pre-treatment on osteoclastogenesis.Precursors were pretreated withM-CSF (25 ng/ml) for 0, 1, or 2weeks and incubated with M-CSFand sRANKL for 1, 2, or 3weeks. Mean � SEM (n � 4 den-tine slices/group). Groups at eachweek of sRANKL exposure withdifferent superscripts are signifi-cantly different (p � 0.0001;two-way ANOVA; Tukey’s posthoc test).
FIG. 2. Time course of humanosteoclast differentiation frompooled colonies. (A) Low-powerphotomicrographs of OC forma-tion and resorption on dentineslices. Scale bar � 200 �m. (B)OC formation, mean plan area,and resorption after 1, 2, and 3weeks of culture. Mean � SEM(n � 4 dentine slices/group).Groups with different super-scripts are significantly different(p � 0.001; one-way ANOVA;Fishers multiple comparisontest).
194 HODGE ET AL.
more reliable than that from unfractionated PBMCs.(22)
However, the yield of OCs from CD14� precursors remainslow—the total number of nuclei in multinucleated OC at theend of the culture is �5% of the nuclei present at baseline.Furthermore, there is substantial variation in the yield withdifferent blood donors. Here we show that CFU-GMs de-rived from human umbilical cord blood form OCs withmarkedly improved capacity compared with CD14�, thusproviding a more efficient and reliable system for the invitro study of human OC differentiation. We also show thatprecursors derived from CFU-Ms have low osteoclastogenicpotential, and pretreatment of precursors with M-CSF in theabsence of sRANKL, while maintaining the precursor pool,moderately reduces OC potential. These observationssuggest that OCs differentiate from early myelomonocyticprogenitors rather than more differentiated monocyte/macrophage progenitors.
Several studies have demonstrated that OCs derive fromimmature colony-forming hematopoietic cells.(5,29–31) Evenso, the precise lineage of the OC and its relationship to otherhematopoietic cells remain unclear. Previous studies have
reported that OCs differentiate from CFU-Ms, maturemonocytes, and macrophages.(6–8) Others have reportedthat OCs derive from CFU-GMs, but not from more com-mitted CFU-Ms,(4,31–33) consistent with our own results.These observations support the hypothesis that the OClineage branches to terminal differentiation before furtherpassage toward the monocyte/macrophage lineage. Our data(from experiments where precursors were pretreated withM-CSF), and those of others,(29,30) indicate that M-CSFsupports the survival of OC precursors, but in the absence ofRANKL, it induces gradual macrophage differentiation andloss of osteoclastogenic potential.
An important role for GM-CSF in osteoclastogenesis isevident from previous work, but the data are inconsistent.GM-CSF has been variously reported to stimulate(16,34,35) orinhibit(13,15,18,19) OC differentiation. When mouse bonemarrow cells were cultured in semisolid medium, GM-CSFstimulated OC progenitor proliferation.(15) However, whenadded directly to 1�,25(OH)2D3-treated marrow cells, itinhibited OC formation.(15) In a recent study using mousebone marrow cultures, Miyamato et al.(17) showed thatGM-CSF increased the OC progenitor pool but inhibiteddifferentiation. In this study, treatment with GM-CSF wasassociated with reduced expression of c-Fos and increasedexpression of DC surface markers. The differentiation intoDC was inhibited by M-CSF, suggesting that M-CSF andGM-CSF reciprocally regulate the differentiation of bothOC and DC lineages, with expression of c-Fos being a keymediator of this bifurcation. Interestingly, this study could
FIG. 4. Concentration-dependent inhibition of osteoclastogenesis byGM-CSF. (A) OC formation and resorption of dentine. (B) Low-powerphotomicrographs of dendritic cell clusters cultured on dentine show-ing positive immunoreactivity for CD1a (solid arrow heads) and anon-immune serum control. Scale bar � 200 �m. Results are mean �SEM (n � 8 dentine slices/group). Groups with different superscriptsare significantly different (p � 0.0001; one-way ANOVA; Fishersmultiple comparison test).
FIG. 5. Effect of delayed addition of GM-CSF on osteoclastogenesis.GM-CSF (10 ng/ml) was added on days indicated and was present untilthe termination of the cultures at day 14. Mean � SEM (n � 8 dentineslices/group). Groups with different superscripts are significantly dif-ferent (p � 0.0001; one-way ANOVA; Fishers multiple comparisontest).
195BIPHASIC EFFECT OF GM-CSF ON OSTEOCLASTOGENESIS

not show a similar effect in a human model, and the authorssuggested that species variability was the likely explanation.In contrast, Quinn et al.(13) have shown an inhibitory effectof GM-CSF in a human model employing monocyte pre-cursors.
We have demonstrated that prolonged GM-CSF treatmentinhibits human osteoclastogenesis while promoting the for-mation of CD1a�/TRACP� DC clusters. The inhibitoryaction of GM-CSF on osteoclastogenesis was limited to thefirst 7 days of the 14-day culture period, indicating that itacts at an early stage of OC differentiation. Consistent withthe data of Miyamato et al.,(17) we found that expression ofc-Fos mRNA was strongly suppressed by GM-CSF. Ofinterest, DC cell proliferation occurred without any effecton OC number or resorption when GM-CSF was added tomature (10 day) OC cultures, suggesting that a discretepopulation of DC precursors remain at this late stage.
Our data clearly show that the timing and duration ofexposure to GM-CSF is vital in determining its effect onterminal differentiation outcomes of OC progenitors. Arecent study compared the osteoclastogenic capacity of pre-cursors from murine bone marrow in semisolid (adherence-independent) or liquid (adherence-dependent) culture con-ditions.(36) These authors proposed a two-stage model: thefirst consisting of multipotential precursors capable of pro-liferating in an adherence-independent manner; the secondconsisting of committed OC precursors whose subsequentdifferentiation was adherence-dependent. Under our cultureconditions, GM-CSF caused proliferation of nonadherentcells that could be blocked by nocodazole. However, GM-CSF did not influence the number of adherent cells at day 1,
suggesting that adherence and interaction with integrins(36)
are key events in determining whether the response toGM-CSF will be proliferation or DC differentiation.
Murine OC precursors exclusively express �v�5 integrin,which declines with commitment to the OC phenotype,whereas levels of �v�3 increase concomitantly.(37) Unlike�v�3, �v�5 mediates both attachment and spreading onligand of marrow macrophages, suggesting �v�3 may serveto mediate initial recognition of bone matrix by OC precur-sors.(37) Treatment of OC precursors with GM-CSF leads toalterations in �v�5 and �v�3 expression, paralleling thoseoccurring during early OC formation, further highlightingthe importance of timing of exposure to GM-CSF.
Many cells in bone marrow and bone, including mac-rophages, monocytes, fibroblasts, endothelial cells, chon-drocytes, natural killer (NK) cells, mast cells, andT-lymphocytes, secrete GM-CSF when stimulated.(38,39)
Osteoblasts also produce GM-CSF when stimulated byTNF�,(40) parathyroid hormone (PTH), and lipopolysac-charide (LPS).(41) GM-CSF production by humanT-lymphocytes has been shown to mediate IL-18 –induced inhibition(18) and annexin II–induced stimulationof osteoclastogenesis.(42) The actions of GM-CSF in the
FIG. 6. Biphasic action of GM-CSF on osteoclastogenesis. GM-CSF(10 ng/ml) was added from the commencement of culture and removedat times indicated. Mean � SEM (n � 6 dentine slices/group). Groupswith different superscripts are significantly different (p � 0.0001;one-way ANOVA; Fishers multiple comparison test).
FIG. 7. Effect of pretreatment of OC precursors with GM-CSFbefore addition of sRANKL. Precursors were cultured in the presenceof M-CSF (25 ng/ml) with or without GM-CSF (10 ng/ml) for theindicated time. After this pretreatment phase, media were replaced withsRANKL (125 ng/ml) and M-CSF (25 ng/ml) for 14 days. Mean �SEM (n � 6 dentine slices/group). *Significantly different to corre-sponding group without GM-CSF (p � 0.05; two-sample t-test).
196 HODGE ET AL.

bone marrow environment are clearly complex. GM-CSFis the pivotal mediator of the maturation and function ofDC, the most important cell type for the induction ofprimary T-cell immune responses.(43) Release of RANKLby activated T-cells would provide a pro-osteoclastogenic signal to OC precursors in addition tothe direct proliferative action of short-term exposure to
GM-CSF, demonstrated in the current study. However,persistent exposure to GM-CSF would result in prefer-ential differentiation of DC, which in turn may furtherinhibit osteoclastogenesis by their production of inhibi-tory cytokines such as IL-12.(44)
Mice with a defective M-CSF receptor caused by auto-somal recessive inactivation in the cfms gene (op/op) haveimpairment of both macrophage and OC formation and areosteopetrotic.(45–48) The severity of the osteopetrosis re-duces with age,(49–51) suggesting that an alternative molec-ular mechanism exists, possibly mediated by GM-CSF.However, administration of high-dose GM-CSF does notcorrect osteopetrosis in these mice,(52) and op/op mice thatare also GM-CSF–deficient recover with aging.(53) Subse-quently, Myint et al.(54) showed that administration of lowdoses of GM-CSF and/or IL-3 does correct osteopetrosis inyoung op/op mice and noted that GM-CSF–deficient op/opmice show a marked delay in recovery from osteoporosiscompared with single mutant op/op mice, presumably be-cause of the GM-CSF deficiency.
In conclusion, we have shown that CFU-GMs generatedfrom human cord blood, which is readily available, providean enriched pool of OC precursors that reliably, efficiently,and rapidly differentiate into mature bone-resorbing OCs.However, more committed CFU-Ms have low potential forOC differentiation. This study has identified GM-CSF as apotent biphasic regulator of osteoclast differentiation. Be-cause GM-CSF is expressed by multiple cells types in thebone marrow, including T-cells, it is likely that it plays asignificant role in modulation of osteoclastogenesis by cellsof the hematopoietic and immune systems.
ACKNOWLEDGMENTS
The authors thank Drs Matthew Gillespie and JulianQuinn (St Vincent’s Institute of Medical Research, Mel-bourne, Australia) and Genetics Institute for generouslyproviding sRANKL and recombinant human M-CSF, re-spectively. This study was supported by the National Healthand Medical Research Council of Australia.
REFERENCES
1. Akagawa KS, Takasuka N, Nozaki Y, Komuro I, Azuma M, UedaM, Naito M, Takahashi K 1996 Generation of CD1�RelB� den-dritic cells and tartrate-resistant acid phosphatase-positiveosteoclast-like multinucleated giant cells from human monocytes.Blood 88:4029–4039.
2. Ash P, Loutit JF, Townsend KMS 1980 Osteoclasts derived fromhaematopoietic stem cells. Nature 283:669–670.
3. Scheven BAA, Visser JWM, Nijweide PJ 1986 In vitro osteoclastgeneration from different bone marrow fractions, including ahighly enriched haematopoietic stem cell population. Nature 321:79–81.
4. Menaa C, Kurihara N, Roodman GD 2000 CFU-GM-derived cellsform osteoclasts at a very high efficiency. Biochem Biophys ResCommun 267:943–946.
5. Kerby JA, Hattersley G, Collins DA, Chambers TJ 1992 Deriva-tion of osteoclasts from hematopoietic colony-forming cells inculture. J Bone Miner Res 7:353–362.
6. Yamazaki H, Kunisada T, Yamane T, Hayashi SI 2001 Presence ofosteoclast precursors in colonies cloned in the presence of hema-topoietic colony-stimulating factors. Exp Hematol 29:68–76.
7. Fujikawa Y, Quinn JM, Sabokbar A, McGee JO, Athanasou NA1996 The human osteoclast precursor circulates in the monocytefraction. Endocrinology 137:4058–4060.
TABLE 2. EFFECT OF SHORT-TERM PRETREATMENT OF OCPRECURSORS WITH GM-CSF ON CELL PROLIFERATION
TreatmentProportion of cells
in S-phase (%)*
Experiment 1M-CSF 11.1 � 0.4c
M-CSF � GM-CSF 20.2 � 0.1a
M-CSF � sRANKL 16.7 � 0.7b
M-CSF � sRANKL � GM-CSF 20.5 � 0.8a
Experiment 2M-CSF 5.9 � 0.6b
M-CSF � GM-CSF 14.1 � 1.7a
M-CSF � nocodazole 2.8 � 0.6b
M-CSF � GM-CSF � nocodazole 5.5 � 0.5b
* Assessed by BrdU incorporation.Results are given as the mean � SEM (n � 3 wells/group). Groups with
different superscripts within experiments are significantly different; p �0.0001; one-way ANOVA; Fishers multiple comparison test.
FIG. 8. Effect of cell-cycle arrest on the ability of short-term pre-treatment with GM-CSF to enhance osteoclastogenesis. Precursorswere pretreated with M-CSF (25 ng/ml), with or without GM-CSF (10ng/ml), and/or nocodazole (40 ng/ml) for 48 h. They were incubatedwith M-CSF for a further 24 h and incubated with M-CSF andsRANKL for 1 week. Mean � SEM (n � 6 dentine slices/group).Groups with different superscripts are significantly different (p �0.006; one-way ANOVA; Fishers multiple comparison test).
197BIPHASIC EFFECT OF GM-CSF ON OSTEOCLASTOGENESIS

8. Udagawa N, Takahashi N, Akatsu T, Tanaka H, Sasaki T, Nishi-hara T, Koga T, Martin TJ, Suda T 1990 Origin of osteoclasts:Mature monocytes and macrophages are capable of differentiatinginto osteoclasts under a suitable microenvironment prepared bybone marrow-derived stromal cells. Proc Natl Acad Sci USA87:7260–7264.
9. Tanaka S, Takahashi N, Udagawa N, Tamura T, Akatsu T, StanleyER, Kurokawa T, Suda T 1993 Macrophage colony-stimulatingfactor is indispensable for both proliferation and differentiation ofosteoclast progenitors. J Clin Invest 91:257–263.
10. Yoshida H, Hayashi S-I, Kunisada T, Ogawa M, Nishikawa S,Okamura H, Sudo T, Shultz LD, Nishikawa S-I 1990 The murinemutation osteopetrosis is in the coding region of the macrophagecolony stimulating factor gene. Nature 345:442–444.
11. Kodama H, Nose M, Niida S, Yamasaki A 1991 Essential role ofmacrophage colony-stimulating factor in the osteoclast differenti-ation supported by stromal cells. J Exp Med 173:1291–1293.
12. Matsuzaki K, Udagawa N, Takahashi N, Yamaguchi K, Yasuda H,Shima N, Morinaga T, Toyama Y, Yabe Y, Higashio K, Suda T1998 Osteoclast differentiation factor (ODF) induces osteoclast-like cell formation in human peripheral blood mononuclear cellcultures. Biochem Biophys Res Commun 246:199–204.
13. Quinn JM, Elliott J, Gillespie MT, Martin TJ 1998 A combinationof osteoclast differentiation factor and macrophage-colony stimu-lating factor is sufficient for both human and mouse osteoclastformation in vitro. Endocrinology 139:4424–4427.
14. Takahashi N, Udagawa N, Suda T 1999 A new member of tumornecrosis factor ligand family, ODF/OPGL/TRANCE/RANKL,regulates osteoclast differentiation and function. Biochem BiophysRes Commun 256:449–455.
15. Takahashi N, Udagawa N, Akatsu T, Tanaka H, Shionome M,Suda T 1991 Role of colony-stimulating factors in osteoclastdevelopment. J Bone Miner Res 6:977–985.
16. Matayoshi A, Brown C, DiPersio JF, Haug J, Abu-Amer Y, LiapisH, Kuestner R, Pacifici R 1996 Human blood-mobilized hemato-poietic precursors differentiate into osteoclasts in the absence ofstromal cells. Proc Natl Acad Sci USA 93:10785–10790.
17. Miyamoto T, Ohneda O, Arai F, Iwamoto K, Okada S, Takagi K,Anderson DM, Suda T 2001 Bifurcation of osteoclasts and den-dritic cells from common progenitors. Blood 98:2544–2554.
18. Horwood NJ, Udagawa N, Elliott J, Grail D, Okamura H, Kuri-moto M, Dunn AR, Martin T, Gillespie MT 1998 Interleukin 18inhibits osteoclast formation via T cell production of granulocytemacrophage colony-stimulating factor. J Clin Invest 101:595–603.
19. Shinar DM, Sato M, Rodan GA 1990 The effect of hemopoieticgrowth factors on the generation of osteoclast-like cells in mousebone marrow cultures. Endocrinology 126:1728–1735.
20. Chambers TJ 2000 Regulation of the differentiation and functionof osteoclasts. J Pathol 192:4–13.
21. Boyle WJ, Simonet WS, Lacey DL 2003 Osteoclast differentiationand activation. Nature 423:337–342.
22. Nicholson GC, Malakellis M, Collier FM, Cameron PU, HollowayWR, Gough TJ, Gregorio-King C, Kirkland MA, Myers DE 2000Induction of osteoclasts from CD14-positive human peripheralblood mononuclear cells by receptor activator of nuclear factorkappaB ligand (RANKL). Clin Sci (Lond) 99:133–140.
23. Nicholson GC, Moseley JM, Sexton PM, Mendelsohn FAO, Mar-tin TJ 1986 Abundant calcitonin receptors in isolated rat oste-oclasts. Biochemical and autoradiographic characterization. J ClinInvest 78:355–360.
24. Myers DE, Collier FM, Minkin C, Wang H, Holloway WR,Malakellis M, Nicholson GC 1999 Expression of functionalRANK on mature rat and human osteoclasts. FEBS Lett 463:295–300.
25. Gratzner HG 1982 Monoclonal antibody to 5-bromo- and 5-io-dodeoxyuridine: A new reagent for detection of DNA replication.Science 218:474–475.
26. Nakahata T, Ogawa M 1982 Hemopoietic colony-forming cells inumbilical cord blood with extensive capability to generate mono-and multipotential hemopoietic progenitors. J Clin Invest 70:1324–1328.
27. Metcalf D, Johnson GR, Mandel TE 1979 Colony formation inagar by multipotential hemopoietic cells. J Cell Physiol 98:401–420.
28. Anderson DM, Maraskovsky E, Billingsley WL, Dougall WC,Tometsko ME, Roux ER, Teepe MC, DuBose RF, Cosman D,Galibert L 1997 A homologue of the TNF receptor and its ligand
enhance T-cell growth and dendritic-cell function. Nature 390:175–179.
29. Hattersley G, Kerby JA, Chambers TJ 1991 Identification of oste-oclast precursors in multilineage hemopoietic colonies. Endocri-nology 128:259–262.
30. Kurihara N, Suda T, Miura Y, Nakauchi H, Kodama H, Hiura K,Hakeda Y, Kumegawa M 1989 Generation of osteoclasts fromisolated hematopoietic progenitor cells. Blood 74:1295–1302.
31. Kurihara N, Chenu C, Miller M, Civin C, Roodman GD 1990Identification of committed mononuclear precursors for osteoclast-like cells formed in long term human marrow cultures. Endocri-nology 126:2733–2741.
32. Schneider GB, Relfson M 1988 The effects of transplantation ofgranulocyte-macrophage progenitors on bone resorption in osteo-petrotic rats. J Bone Miner Res 3:225–232.
33. Schneider GB, Relfson M 1988 A bone marrow fraction enrichedfor granulocyte-macrophage progenitors gives rise to osteoclasts invitro. Bone 9:303–308.
34. Fujikawa Y, Sabokbar A, Neale SD, Itonaga I, Torisu T, Athana-sou NA 2001 The effect of macrophage-colony stimulating factorand other humoral factors (interleukin-1, -3, -6, and -11, tumornecrosis factor-alpha, and granulocyte macrophage-colony stimu-lating factor) on human osteoclast formation from circulating cells.Bone 28:261–267.
35. Toyosaki-Maeda T, Takano H, Tomita T, Tsuruta Y, Maeda-Tanimura M, Shimaoka Y, Takahashi T, Itoh T, Suzuki R, Ochi T2001 Differentiation of monocytes into multinucleated giant bone-resorbing cells: Two-step differentiation induced by nurse-likecells and cytokines. Arthritis Res 3:306–310.
36. Miyamoto T, Arai F, Ohneda O, Takagi K, Anderson DM, Suda T2000 An adherent condition is required for formation of multinu-clear osteoclasts in the presence of macrophage colony-stimulatingfactor and receptor activator of nuclear factor kappa B ligand.Blood 96:4335–4343.
37. Inoue M, Namba N, Chappel J, Teitelbaum SL, Ross FP 1998Granulocyte macrophage-colony stimulating factor reciprocallyregulates alphav-associated integrins on murine osteoclast precur-sors. Mol Endocrinol 12:1955–1962.
38. Hamilton JA 2002 GM-CSF in inflammation and autoimmunity.Trends Immunol 23:403–408.
39. Nimer SD, Uchida H 1995 Regulation of granulocyte-macrophagecolony-stimulating factor and interleukin 3 expression. Stem Cells13:324–335.
40. Felix R, Cecchini MG, Hofstetter W, Guenther HL, Fleisch H 1991Production of granulocyte-macrophage (GM-CSF) and granulo-cyte colony-stimulating factor (G-CSF) by rat clonal osteoblasticcell population CRP 10/30 and the immortalized cell line IRC10/30-myc1 stimulated by tumor necrosis factor alpha. Endocrinology128:661–667.
41. Weir EC, Insogna KL, Horowitz MC 1989 Osteoblast-like cellssecrete granulocyte-macrophage colony-stimulating factor in re-sponse to parathyroid hormone and lipopolysaccharide. Endocri-nology 124:899–904.
42. Menaa C, Devlin RD, Reddy SV, Gazitt Y, Choi SJ, Roodman GD1999 Annexin II increases osteoclast formation by stimulating theproliferation of osteoclast precursors in human marrow cultures.J Clin Invest 103:1605–1613.
43. Tarr PE 1996 Granulocyte-macrophage colony-stimulating factorand the immune system. Med Oncol 13:133–140.
44. Horwood NJ, Elliott J, Martin TJ, Gillespie MT 2001 IL-12 aloneand in synergy with IL-18 inhibits osteoclast formation in vitro.J Immunol 166:4915–4921.
45. Marks SC Jr 1987 Osteopetrosis–multiple pathways for the inter-ception of osteoclast function. Appl Pathol 5:172–183.
46. Marks SC Jr 1982 Morphological evidence of reduced bone re-sorption in osteopetrotic (op) mice. Am J Anat 163:157–167.
47. Marks SC Jr, Lane PW 1976 Osteopetrosis, a new recessiveskeletal mutation on chromosome 12 of the mouse. J Hered 67:11–18.
48. Wiktor-Jedrzejczak WW, Ahmed A, Szczylik C, Skelly RR 1982Hematological characterization of congenital osteopetrosis inop/op mouse. Possible mechanism for abnormal macrophage dif-ferentiation. J Exp Med 156:1516–1527.
49. Begg SK, Bertoncello I 1993 The hematopoietic deficiencies inosteopetrotic (op/op) mice are not permanent, but progressivelycorrect with age. Exp Hematol 21:493–495.
198 HODGE ET AL.

50. Begg SK, Radley JM, Pollard JW, Chisholm OT, Stanley ER,Bertoncello I 1993 Delayed hematopoietic development in osteo-petrotic (op/op) mice. J Exp Med 177:237–242.
51. Nilsson SK, Bertoncello I 1994 Age-related changes in extramed-ullary hematopoiesis in the spleen of normal and perturbed osteo-petrotic (op/op) mice. Exp Hematol 22:377–383.
52. Wiktor-Jedrzejczak W, Urbanowska E, Szperl M 1994Granulocyte-macrophage colony-stimulating factor corrects mac-rophage deficiencies, but not osteopetrosis, in the colony-stimulating factor-1-deficient op/op mouse. Endocrinology 134:1932–1935.
53. Nilsson SK, Lieschke GJ, Garcia-Wijnen CC, Williams B, Tzel-epis D, Hodgson G, Grail D, Dunn AR, Bertoncello I 1995Granulocyte-macrophage colony-stimulating factor is not respon-sible for the correction of hematopoietic deficiencies in the matur-ing op/op mouse. Blood 86:66–72.
54. Myint YY, Miyakawa K, Naito M, Shultz LD, Oike Y, YamamuraK, Takahashi K 1999 Granulocyte/macrophage colony-stimulating
factor and interleukin-3 correct osteopetrosis in mice with osteo-petrosis mutation. Am J Pathol 154:553–566.
Address reprint requests to:Geoffrey Charles Nicholson, MBBS, PhD
The University of MelbourneDepartment of Clinical and Biomedical Sciences: Barwon
HealthThe Geelong Hospital
Geelong, Victoria 3220, AustraliaE-mail: [email protected]
Received in original form June 7, 2003; in revised form August 18,2003; accepted September 5, 2003.
199BIPHASIC EFFECT OF GM-CSF ON OSTEOCLASTOGENESIS
Copyright © 2022 FDOKUMEN




















