
Organization and Elemental Analysis of P-, S-, and Fe-rich Inclusions in a Population of Freshwater...
20
Geomicrobiology Journal, 19:387 – 406, 2002 Copyright C ° 2002 Taylor & Francis 0149-0451/02 $12.00 + .00 DOI: 10.1080/0149045029009850 4 Organization and Elemental Analysis of P-, S-, and Fe-rich Inclusions in a Population of Freshwater Magnetococci B. LEA COX RADU POPA Center for Life Detection Jet Propulsion Laboratory and California Institute of Technology Pasadena, California, USA DENNIS A. BAZYLINSKI Department of Microbiology Iowa State University Ames, Iowa, USA BRIAN LANOIL SUSANNE DOUGLAS ANDREA BELZ DIANE L. ENGLER KENNETH H. NEALSON Center for Life Detection Jet Propulsion Laboratory and California Institute of Technology Pasadena, California, USA We characterized a population of bilophotrichously agellated freshwater magnetotac- tic cocci (MC), referred to as ARB-1, morphologically, chemically, and phylogenetically. Cells examined using light microscopy, uorescence microscopy, environmental scan- ning electron microscopy (ESEM), and transmission electron microscopy (TEM) con- tained three types of intracellular inclusions placed in a speci c arrangement within the cell. Elemental compositions of the inclusions were determined using energy dispersive X-ray spectroscopy (EDS) from both ESEM and TEM. The spherical to ovoid cells con- tained two large phosphorus-rich inclusions that occupied most of the cell volume and Received 15 March 2001; accepted 29 May 2001. We thank J. Kirschvink (California Institute of Technology) for suggesting the eld location, for valuable discussions, and for laboratory space. H. Vali (McGill University) provided many TEM images including an HRTEM image of one of the magnetite crystals showing lattice fringes, as well as helpful discussions and sug- gestions. P. Koen, J. Edens, and C. Garland (California Institute of Technology) provided technical assistance at the TEM facilities. K. Venkateswaran and W. Schubert (Jet Propulsion Laboratory) provided help with the live- dead stained images. P. Atkins and the staff at the Los Angeles Arboretum allowed us to access the lake for our numerous collections. NASA provided funding under the Jet Propulsion Laboratory Astrobiology grant 10417. D. A. Bazylinski was supported by NASA Johnson Space Center grant NAG 9-1115 and U.S. National Science Foundation grant CHE-9714101. Address correspondenc e to B. Lea Cox, University of Southern California, Earth Sciences, Science Hall 117, 3651 Trousdale Parkway, Los Angeles, CA 90089-0740 , USA. E-mail: [email protected] 387
-
Upload
independent -
Category
Documents
-
view
0 -
download
0
Transcript of Organization and Elemental Analysis of P-, S-, and Fe-rich Inclusions in a Population of Freshwater...

Geomicrobiology Journal 19387ndash406 2002Copyright Cdeg 2002 Taylor amp Francis0149-0451 02 $1200 + 00DOI 1010800149045029009850 4
Organization and Elemental Analysis of P- S-and Fe-rich Inclusions in a Population
of Freshwater Magnetococci
B LEA COXRADU POPA
Center for Life DetectionJet Propulsion Laboratory and California Institute of TechnologyPasadena California USA
DENNIS A BAZYLINSKI
Department of MicrobiologyIowa State UniversityAmes Iowa USA
BRIAN LANOILSUSANNE DOUGLASANDREA BELZDIANE L ENGLERKENNETH H NEALSON
Center for Life DetectionJet Propulsion Laboratory and California Institute of TechnologyPasadena California USA
We characterized a population of bilophotrichously agellated freshwater magnetotac-tic cocci (MC) referred to as ARB-1 morphologically chemically and phylogeneticallyCells examined using light microscopy uorescence microscopy environmental scan-ning electron microscopy (ESEM) and transmission electron microscopy (TEM) con-tained three types of intracellular inclusions placed in a speci c arrangement within thecell Elemental compositions of the inclusions were determined using energy dispersiveX-ray spectroscopy (EDS) from both ESEM and TEM The spherical to ovoid cells con-tained two large phosphorus-rich inclusions that occupied most of the cell volume and
Received 15 March 2001 accepted 29 May 2001We thank J Kirschvink (California Institute of Technology) for suggesting the eld location for valuable
discussions and for laboratory space H Vali (McGill University) provided many TEM images including anHRTEM image of one of the magnetite crystals showing lattice fringes as well as helpful discussions and sug-gestions P Koen J Edens and C Garland (California Institute of Technology) provided technical assistance atthe TEM facilities K Venkateswaran and W Schubert (Jet Propulsion Laboratory) provided help with the live-dead stained images P Atkins and the staff at the Los Angeles Arboretum allowed us to access the lake for ournumerous collections NASA provided funding under the Jet Propulsion Laboratory Astrobiology grant 10417D A Bazylinski was supported by NASA Johnson Space Center grant NAG 9-1115 and US National ScienceFoundation grant CHE-9714101
Address correspondenc e to B Lea Cox University of Southern California Earth Sciences Science Hall117 3651 Trousdale Parkway Los Angeles CA 90089-0740 USA E-mail blcoxuscedu
387
388 B L Cox et al
appeared to be enclosed in a membrane or coating Several smaller sulfur-rich inclusionswere located at the end of the cell opposite the agellar bundles The magnetosomesarranged either as a cluster a chain or a combination of both were located proximal tothe two agellar bundles Magnetite was identi ed as the mineral phase of the magneto-somes using selected area electron diffraction (SAED) and by measuring lattice fringespacings of the crystals The magnetite crystals were hexagonal prisms that averaged82 nm in length and thus t into the single-magnetic-domain size range Phylogeneticanalysis of the 16S rRNA gene sequences suggests that it is a mixed population of MCthat form a monophyletic clade distinct from but similar to other uncultured MC
Keywords magnetococci magnetite ESEM TEM sulfur inclusions phosphorusinclusions
Introduction
The magnetotactic bacteria rst discovered by Richard Blakemore in 1974 are prokaryoticmicroorganisms whose direction of motility is in uenced by magnetic elds (Blakemore1975) They are ubiquitous and abundant in aquatic environments including salt marshesfreshwater swamps (Blakemore et al 1979) and sewage oxidation ponds (Moench andKonetzka 1978) These bacteria are diverse morphologically physiologically and phylo-genetically but share a few key characteristics 1) the presence of intracellular membrane-bound magnetic crystals called magnetosomes (Balkwill et al 1980) that are usually butnot always arranged in chains within the cell 2) motility by means of agella and 3) mi-croaerophilic andor anaerobic physiology The function of the magnetosomes appears tobe in the cellrsquos locating and maintaining an optimal position in vertical chemical gradientscommon in aquatic habitats (Frankel et al 1997) The mechanism of formation of the mag-netosomes has not been completely elucidated The biogeochemical roles these bacteriaplay in natural environments are also not understood
Most researchers accept the idea that the primary function of magnetosomes is naviga-tion (Bazylinski and Frankel 2000) although some cells appear to contain more magneto-somes than would be required for navigation (Vali and Kirschvink 1990) Other suggestedfunctions for magnetosomes are iron storage prevention of iron toxicity the use of iron as anenergy source and destruction of toxic oxygen radicals produced during oxygen respiration(Blakemore 1982) It is unlikely that magnetite-containing magnetosomes could be used foriron storage or as an energy source because the iron in magnetosomes is not used by cellseven when they are starved for iron and several species limit their own growth by incor-porating iron into magnetosomes rather than into essential iron-containing proteins (D ABazylinski unpublished results) Many magnetotactic bacteria such as the ones discussedhere also contain inclusions of phosphorus (Blakemore 1982 Lins and Farina 1999) andorsulfur (Moench 1988 Iida and Akai 1996 Bazylinski and Frankel 2000) although moststudies have focussed on magnetosomes In this paper we describe and discuss all threekinds of intracellular structures present in a group of magnetotactic cocci (MC) abundantin a freshwater lake
A major obstacle to studying magnetic bacteria is that they are generally very dif cultto cultivate in pure culture and thus very few pure cultures exist The cultured magne-totactic bacteria include the freshwater spirilla Magnetospirillum magnetotacticum MS-1(Blakemore et al 1979) M magnetotacticum strain AMB1 (Matsunaga et al 1991) andM gryphiswaldense (Schleifer et al 1991 Schuler et al 1999) the marine vibrios MV-1and MV-2 (Bazylinski et al 1988 Meldrum et al 1993b) the curved sulfate-reducing rodstrain RS-1 (Sakaguchi et al 1993) the magnetotactic coccoid strains MC-1 (Meldrumet al 1993a) MC-2 (Devouard et al 1998) and MC-3 (Dennis Bazylinski unpublished)
Organization of Inclusions in Magnetococci 389
The MC were the rst magnetotactic bacteria to be described (Blakemore 1975) and ap-pear to be the most abundant magnetotactic bacteria in freshwater environments with a highorganic content (Schuler 1999)
We report here on a group of uncultured freshwater MC we refer to as ARB-1 whichappear to exhibit some novel behaviors and characteristics These include the reversal ofswimming direction under different conditions possible diurnal effects on the swimmingdirection and a tendency to die sooner if they are in a magnetically shielded environmentAs yet it has not been possible to grow ARB-1 in pure culture However microscopic andchemical studies show that ARB-1 cells have a remarkable organization of interior andexternal structures that might indicate physiological and biogeochemical features of thispopulation
Description of Field Location and Conditions
Cells of ARB-1 were discovered in Baldwin Lake a small lake located within the LosAngeles Arboretum (Arcadia CA USA) Baldwin Lake has been described as a sag pond(Buwalda 1940) and is located on the upthrust side of the Raymond Fault a reverse thrustfault with left-lateral movement (Crook et al 1987) The lake is underlain by Quaternarygravel and alluvium eroded and transported from granodiorite quartz diorite and gneissicsource rocks containing abundant iron-rich hornblende and biotite The shore was arti ciallyplanted with Mediterranean and tropical plants and various chemicals and fertilizers usedin garden maintenance are periodically introduced to the lake ARB-1 cells appeared tobe particularly abundant in locations where the sediments were rich in large fragments ofmetallic iron and magnetic particles
The temperature during the collection period (July 1999ndashOctober 2000) varied from10plusmnC to 28plusmnC The pH of water collected from the lake was 72 to 76 (cells remained viablein bottles in the laboratory at pHs up to 83) The Eh at the surface of the sediment rangedfrom D iexcl315 mV to iexcl183 mV and oxygen from 012 to 017 mgL (38 to 53 sup1M)
Methods
Extraction of the Bacteria
Samples used for cell extraction were taken from 2-liter bottles lled with approximately50 surface sediment and 50 overlying lake water Bottles were taken back to the labora-tory shaken vigorously and magnetic stir bars placed at the watersediment interface alongthe Earthrsquos magnetic eld with the south pole of one magnet on the north side of the bottleand the north pole of the other magnet on the south side of the jar Typically cells accumu-lated as a pinkish pellet at the south-seeking side The pellet was removed with a Pasteurpipette and released into a scintillation vial containing a small amount of lter-sterilizedlake water Magnets were placed on the scintillation vial as described previously and theprocess repeated to further purify the cells
In some experiments we used a modi cation of the racetrack method (Wolfe et al1987 Schuler et al 1999) to separate the magnetic bacteria from nonmagnetic cells Inthis method a sterile cotton plug acting as a barrier for nonmagnetotactic bacteria wasinserted into the wide end of a sealed sterile Pasteur pipette The magnetotactic bacteriaswam to the sealed tip toward the south end of a bar magnet and then the tip containing thecells was broken off and the cells removed The MC typically swim much faster than othermagnetotactic and nonmagnetotactic cells and thus are the rst cells to appear at the tip ofthe pipette
390 B L Cox et al
Optical and Fluorescence Light Microscopy
Optical light microscopy was performed with fresh cells (within 24 h of collection) using aNikon Eclipse E600 microscope with phase contrast Images were obtained with a JavelinMOS solid state camera Flagella were visualized via the Gray staining method using amethod described by Murray and Robinow (1994) Loef errsquos methylene blue stain was usedto test for polyphosphate inclusions and safranin was used to visualize inclusion membranes(Murray and Robinow 1994) Viability was assessed using the Baclite bacterial viabilitykit (Molecular Probes L-7012 stain) This method employs two nucleic acid dyes bothof which are excited with blue (490 nm) light The green dye (cytonine) labels all cellsdead and live while the red dye (propidium iodate) is excluded from living cells but canpenetrate the damaged membranes of dead cells Thus living cells appear green while deadcells appear red
Transmission Electron Microscopy (TEM)
We used a Philips EM 201C microscope operating at 80 kV to examine stained thin sectionsand whole mounts of cell suspensions Cells for thin sections were xed in glutaraldehyderinsed in malate buffer post xed in osmium tetroxide stained with uranyl acetate dehy-drated with ethanol and embedded in EPON resin (Beveridge et al 1994) For negativestains whole cells were deposited on Cu grids coated with Formvar and stained with 2uranyl acetate
A Philips 430 TEM with a single crystal lanthanum hexaboride lament operatingat 200 kV was used for microscopy energy dispersive X-ray spectroscopy (EDS) andselected area electron diffraction (SAED) on fresh (collected within 24 h) unstained cellsuspensions that were deposited on carbon-coated copper grids The area sampled by theEDAX 9900 spectrum analyzer is approximately 200 nm The electron beam was defocusedat the condenser lens to maintain counting rates below 1 kHz and live time ef ciency greaterthan 95 The SAED patterns were recorded on Kodak electron image lm and digitizedfor analysis
Environmental Scanning Electron Microscopy (ESEM)
A Philips XL30 ESEM with a eld emission gun was used to observe hydrated un xed cellsImages were acquired at 20 kV in ldquowet moderdquo (3plusmnC 53 Torr chamber and water vaporpressure) at a relative humidity in the sample chamber of 93 The EDS was performedon fully hydrated uncoated and un xed specimens using an accelerating voltage of 20 kVand counting for 100 s (live time) The spot size sampled by the EDS detector was 20 nmbut because of beam spread signals from elements within a radius of 100 nm may also becounted In some cases cells were xed in glutaraldehyde prior to observation (images notshown) This had a tendency to shrink the cells to 60 of their original size For all otherexperiments fresh cells (within 24 h of collection) were used
Molecular Diversity
Bacterial small subunit (SSU) rRNA genes were PCR-ampli ed using the primers B11Fand B1512r (Giovannoni 1991) Five aliquots of cells (ca 108 each) were placed in separatetubes containing 18 sup1l of PCR cocktail (1 pound Taq DNA polymerase buffer with MgCl2 1 Uof Taq polymerase [Promega Madison WI USA] 200 sup1M dNTPs 750 nM each primer)Cells were lysed by incubation for 6 min at 94plusmnC and then subjected to PCR thermalcycling PCR conditions were 30 sec 94plusmnC denaturation 1 min 50plusmnC annealing and
Organization of Inclusions in Magnetococci 391
2 min 72plusmnC elongation for 35 cycles in an MJ Research thermal cycler (Waltham MAUSA) After a nal 10 min incubation at 72plusmnC the products were combined and puri edby gel extraction as recommended by the manufacturer (Qiagen Chatsworth CA) Am-pli cation products were cloned into the plasmid vector pCR21 as recommended by themanufacturer (Invitrogen Carlsbad CA)
SSU rDNA inserts were PCR-ampli ed under the same conditions as above Theproduct was double-digested with HhaI (New England Biolabs Beverly MA) and RsaI(Promega) restriction endonucleases in 1 pound Promega digestion buffer at 37plusmnC for 3 h Thedigested DNA was separated by electrophoresis through low melting point agarose (20wtvol IBI Shelton Scienti c Shelton CT USA) The banding patterns were grouped ac-cording to similarity and representative members of groups displaying patterns similar tothose of MC we had previously sequenced (data not shown) were partially sequenced Sixpartial sequences that were tentatively identi ed as MC and as different groups were sub-sequently fully bidirectionally sequenced with either an ABI 3700 (Applied BiosystemsFoster City CA) or a Licor 4200 (Licor Lincoln NE USA) automated DNA sequencerand dye terminator chemistry
Sequences were initially aligned to their nearest neighbor using the program ARB pro-vided by Wolfgang Ludwig and Oliver Strunk (Technische Universitat Munchen MunichGermany program available at httpwwwtumunchendeARB) The sequences were fur-ther manually aligned to sequences obtained from the GenBank database using the GeneticData Environment (GDE) v 20 sequence analysis software package provided by SteveSmith (Millipore Corporation Bedford MA) and described by Rappe et al (1997) Phy-logenetic inference and evolutionary distance calculations were performed as described inRappe et al (1997) Regions of ambiguous alignment were excluded from analyses
Results and Discussion
General Description of Cells
Cells of ARB-1 were spherical to oblate spheroidal Gram-negative cells ranging from1 to 4 sup1m (Figure 1) and averaging 25 sup1m In the ESEM the average size of cells was16 sup1m if xed with glutaraldehyde (aspect ratio 085) and 27 sup1m if not xed (aspectratio 085) The wide range of cells sizes (1ndash4 sup1m) might re ect the phylogenetic diversity(described below) or indicate that there are different phases in the cell cycle re ected incell size or that cells simply exhibit budding Cells contained two large phosphorus-richsymmetrical intracellular bodies that took up a large fraction of the cell volume Otherobserved inclusions were sulfur globules and magnetosomes Phosphorus-rich inclusionsin ESEM images (Figure 1d) appeared to be wider apart at one end forming a crevicewhere many small sulfur-containing inclusions were located On the opposite side of mostcells where the phosphorus inclusions appeared to be closer together magnetosomes werearranged as a cluster of disorganized crystals as chains or both (Figure 1d)
Several cells in the TEM (Figure 2) and in the ESEM appeared to be in the processof dividing In some cases indentations preceding cell division appeared to occur betweenthe two phosphorus inclusions in others these constrictions seemed to separate the sulfurinclusions from the rest of the cell (Figure 2) We also observed protrusions or extensionson some of the dividing cells (Figure 2c d) The large differences in cell size may well bedue to unequal division (budding) observed in some cells during division (Figure 2) Thecylindrical structure of one of these (Figure 2d) is similar to the basal body of a agellabundle Other possible explanations for these protrusions are that they are membrane vesicles(Beveridge 1999) or some artifact of electron microscopy or staining
392 B L Cox et al
FIGURE 1 Images of ARB-1 cells showing inclusions containing phosphorus (P) sul-fur (S) and magnetite (M) and variations in cell sizes a) Phase contrast light microscopyimage of live cells b) TEM image of unstained cells xed in glutaraldehyde (image taken byH Vali) c) TEM image of unstained and un xed cell showing S-rich globules P-rich in-clusions and magnetosomes (M) in chains d) ESEM image of hydrated un xed cells on amembrane lter H indicates ldquohingedrdquo P inclusions
The swimming speed of ARB-1 cells in wet mounts or in capillary racetracks wasextremely rapid up to a millimeter or 1000 body lengths per second for fresh cells whendirected by magnetic stir bars The phosphorus-rich inclusions appeared to wobble from sideto side or may even have opened and closed slightly as if hinged on one side (Figure 1d)as the cells swam
Organization of Inclusions in Magnetococci 393
FIGURE 2 Images of ARB-1 cells showing cell division a) TEM image of cells stainedwith uranyl acetate showing division across rather than between P inclusions b) ESEMimage showing division of cells between P inclusions c) d) TEM images of cells stainedwith uranyl acetate showing unequal division of cells with protrusions
Flagella and Fimbriae or Pili
Like other MC (Blakemore 1975 1982 Moench 1988 Frankel et al 1997) cells of ARB-1were bilophotrichously agellated having two agellar bundles on one side of the cell(Figure 3) Each bundle contained about 10 unsheathed agella that were 13 nm in diameterCells also appeared to have other smaller protrusions that resemble pili (Figure 3b) Thetendency for cells of ARB-1 to stick to glass slides and cover slips and to each other supportsthe presence of mbriae or pili Pili have been observed on other uncultured (Blakemore1982) and cultured (Frankel et al 1997) MC
Magnetosomes
Magnetosome crystals in ARB-1 cells occurred in chains and clusters and ranged in lengthfrom 20 to 133 nm and in width from 10 to 84 nm with an average (sect standard deviation)length and width of 82 sect 23 nm and 49 sect 15 nm respectively (n D 341) The average lengthof clustered magnetosomes (n D 260) was 83 sect 24 nearly the same as the average lengthof magnetosomes from chains (n D 81 mean D 80 sect 19) The range of lengths of magneto-somes in chains (40ndash120 nm) was narrower than that of clustered magnetosomes The size
394 B L Cox et al
FIGURE 3 Images of ARB-1 cells showing agella andor pili a) Light microscopy imageof cells stained for agella Arrows indicate possible cell division plane b) Thin section of acell stained with uranyl acetate showing spaces where P inclusions had been S inclusionsmagnetosomes (M) and pili c) TEM image of whole cells with clustered magnetosomecrystals (M) negatively stained with uranyl acetate Notice haloes around crystals whichmight represent a membrane or coating
distributions (length and width) of all 341 magnetite crystals from cells of ARB-1 are plot-ted in Figure 4 Values plotted for ARB-1 magnetite crystals showed that like in virtually allother magnetotactic bacteria the magnetite particles in ARB-1 are single-magnetic-domaincrystals (Butler and Banerjee 1975 Bazylinski and Frankel 2000) These results were verysimilar to those obtained for the uncultured coccus Bilophococcus magnetotacticus (Toweand Moench 1981) which also possessed clumps of magnetosomes at one end of the cellrather than chains The average length of ARB-1 magnetosomes was also similar to thatof the cultured MC-1 However the shape of MC-1 magnetosomes was less elongated(Meldrum et al 1993a) It appears that the size distributions and aspect ratios for this typeof MC are distinct from those of other magnetotactic bacteria (eg MV-1 and MC-1) aswell as from those in the Mars meteorite ALH84001 (Thomas-Keprta et al 2000)
Morphologically the magnetosome crystals in ARB-1 were rectangular in projectionwith an hexagonal cross-section There were irregularities in the edges and interiors of the
FIG
UR
E4
Dis
trib
utio
nof
leng
ths
and
wid
ths
of34
1m
agne
tite
crys
tals
from
clus
ters
and
chai
nsin
AR
B-1
cells
a) F
requ
ency
asa
func
tion
ofle
ngth
b)
Freq
uenc
yas
afu
nctio
nof
wid
thc
) Wid
thas
afu
nctio
nof
leng
th
395
396 B L Cox et al
FIGURE 5 TEM image of a thin section of a cell of ARB-1 stained with uranyl acetateshowing magnetosomes before a) and after b) beam damage leaving crystal ldquodoubletsrdquo b)Notice disappearance of electron dense material in b)
crystals (Figure 3) The edges were often rounded and the particles occasionally con-stricted slightly in the middle The crystals sometimes had a mottled appearance and mayhave had fringes around the edges Once during TEM analysis of thin sections (80 kV)magnetosome crystals sustained signi cant beam damage after several minutes of viewingat a magni cation of 100000 (Figure 5) The crystals appeared to have broken in halfpossibly along twin planes in the (111) direction or along ldquowaistedrdquo regions where thecrystals showed some constriction and there appeared to be a loss of material Beam dam-age has been previously observed during TEM of greigite (Fe3S4) magnetosome particlesbut not in magnetite crystals (D Bazylinski H Vali and J Kirschvink personal commu-nications January 2001) except when viewed as whole mounts at 400 kV (Taylor et al2001) Taylor et al (2001) speculated that water vapor from the cells underwent radiolysisin the electron-beam producing a plasma that rapidly melted the magnetosomes
EDS analyses with both TEM and ESEM of the magnetosome particles (Figure 6c 6f)showed the particles were iron-rich with some extraneous signals including sulfur (Figure 6)A strong sulfur signal would be expected from EDS analyses of the magnetic mineral greigitealso known to be produced by some magnetotactic bacteria (Mann et al 1990) In ARB-1however these extraneous signals are likely due to the sulfur- and phosphorus-containinginclusions also present in the cell SAED patterns obtained from the crystals (Figure 7)and lattice fringes from high resolution TEM with spacings of 048 nm elongated in (111)direction (Figure 7) were consistent with the mineral magnetite
Magnetosomes within cells of ARB-1 were found in three different arrangements1) as a polar cluster forming a sort of ldquocaprdquo usually at the agellar end of the cell 2) in partialor complete chains at the same end of the cell or 3) a combination of both arrangementsThe polar clusters appear to be disorganized in many images (eg Figures 1 2 and 3)However the attraction of these cells to a magnet indicates they have permanent magneticdipole moments implying that they are not disorganized We are currently pursuing thisquestion of whether or not the clusters are disorganized
In cells having both chains and clusters it is dif cult to determine whether chainswere forming from the clustered crystals or whether the polar clusters of particles werefrom a chain that was disrupted during dehydration of the cell during preparation for TEMexamination However polar clusters of crystals were observed in ESEM images of many
Organization of Inclusions in Magnetococci 397
FIGURE 6 EDS spectra from TEM a) b) c) and ESEM d) e) f) showing elementalcomposition of P-rich a) d) and S-rich b) e) inclusions and magnetosomes c) f) Elementalcompositions of clustered magnetosomes were identical to those of magnetosomes in chainsAll y-axes are linear except for a) and b) which are log scales Noise in f ) is due to scattering
hydrated cells (Figure 1d) suggesting that they were not caused by dehydration In manyof the cells viewed by TEM it seemed that the chains tended to occur in smaller morespherical cells while clumps of magnetosomes occurred more frequently in elongatedcells In order to see if there was any relationship between cell size and shape and the threetypes of magnetosome arrangements the size and aspect ratio for cells with these differentmagnetosome arrangements were measured (Figure 8) The cell size distribution appearedto be narrower for cells with chains than for the cells with disorganized crystals Cells withchains were predominantly smaller than 25 sup1m (Figure 8a) The aspect ratios of the cellswith chains only was smaller than that of cells with disorganized magnetosomes and wasshifted toward spherical aspect ratios (Figure 8b)
One explanation for these observations is that there are different stages of magnetosomeformation in a single species of MC The disorganized magnetosomes might form near thecell membrane between the agellar bundles and then extend into chains Alternativelymagnetosomes might rst form as chains in smaller cells At a later stage as cells increasein size the membranes linking the chains dissolve or become stressed and the magneto-somes somehow migrate to the agellar end of the cell Because magnetosomes appear tobe anchored to the inner surface of the cell membrane in virtually all species examined(Bazylinski and Frankel 2000) and are never free- oating in the cytoplasm migration ofmagnetosomes through the cytoplasm seems unlikely Instead we are presently designinghybridization experiments to address the hypothesis that the relationships between cell sizeshape and magnetosome arrangement correlate with the genotypical variations in ARB-1described later Spring et al (1994) analyzed communities of MC found in freshwater lakesin Germany using uorescent oligonucleotide probes They found different genotypes ofMC (similarity below 95) but these genotypes were not phenotypically expressed asdifferent magnetosome organizations but as preferences for different concentrations of O2
398 B L Cox et al
FIGURE 7 Evidence supporting magnetite as the magnetosome mineral in ARB-1 cellsa) Image of a single crystal of magnetite within a cell in an unstained glutaraldehyde- xedwhole cell mount taken with High Resolution TEM by H Vali b) Part of the magnetitecrystal shown in a) magni ed to show the distance between lattice fringes c) Selectedarea electron diffraction (SAED) of a group of magnetosomes in a cell from an un xedunstained whole cell mount showing the indices of some of the diffraction spots
Therefore we might expect no correlation between magnetite organization and phyloge-netic diversity of ARB-1 cells but perhaps a correlation between oxygen preference andphylogeny
A third possibility is that chains and clumps of magnetosomes form as a result ofdifferent environmental conditions Since our sampling methods were not selective as tothe cellsrsquo location in the vertical chemical (eg oxygen) and redox gradients that are sure to
Organization of Inclusions in Magnetococci 399
FIGURE 8 Relationships between organization of magnetosomes and cell size a) and cellaspect ratio b) Measurements were taken from both TEM and ESEM images Solid patternis for cells with clustered magnetosomes crossed pattern is for cells with magnetosomesin chains only Patterned bars originate at the top of solid bars
exist at the site the environmental conditions of our samples represent a relatively large rangeof oxygen and perhaps other chemical concentrations as well as redox conditions Magneticbacteria in culture are known to vary the size and number of magnetosomes in responseto the media composition (Meldrum et al 1993a) and particularly oxygen concentration(Blakemore et al 1985) Another possibility is that the clustered magnetosomes are formedby a different biological mechanism and serve a different function than magnetosomes inchains
Phosphorus (P) Inclusions
The two very large inclusion bodies seen in each cell always occurred in pairs in cellsand seemed to extend to the cell membrane and to be in contact with each other Thus themaximum dimension of the P-rich inclusions was the same as the minimum dimension ofthe cell and their combined width was approximately half of the long dimension of the cellIn ESEM images the two P-containing inclusions appeared to be attached in the middle insome cells and separated in others (Figure 1d) The P-containing inclusions appeared to beself-contained and perhaps bounded by some sort of coating or membrane in TEM images(Figure 2) and in light microscopy images of cells stained with safranin (not shown)
These inclusions were shown by both ESEM and TEM EDS to contain phosphorus with generally small amounts of potassium calcium magnesium sulfur and iron the lattertwo elements likely being due to the close proximity of the P-rich inclusions to sulfurglobules and magnetosomes (Figures 1 and 6) A very large calcium peak was observedwhen these large inclusions were analyzed in the TEM (Figure 6a) The samples shown inFigures 6a and 6d were collected at different times of the year which may explain some ofthe differences observed in these spectra Other EDS data from the ESEM (not shown) hadhigher calcium peaks than those shown in Figure 6d indicating that relative proportions ofP and Ca in P-rich inclusions vary even within the same sample
The inclusions were electron dense in TEM and had a mottled appearance due toablation by the electron beam Similar inclusions have been described for uncultured MCfrom the Itaupo Lagoon near Rio de Janeiro Brazil (Lins and Farina 1999) and from MillPond New Hampshire (Blakemore 1982) Like the structures in ARB-1 polyP granules aretranslucent in visible light (Spring et al 1994) dark (electron dense) in the TEM and oftenhave a mottled appearance (Harold 1966) The P-rich inclusions in ARB-1 cells stained
400 B L Cox et al
with methylene blue (Murray and Robinson 1994) which is suggestive of the presence ofpolyP but not con rmatory
PolyP is composed of linear chains of tetrahedral phosphate residues linked throughcommon oxygen atoms by phosphoanhydrid e bonds At physiological pH each unit carriesa monovalent negative charge giving polyP the property of having a large cation exchangecapacity The binding energy of polyP allows it to sequester Ca2C and Mg2C (Reusch 2000)consistent with the Ca2C and Mg2C signals observed in the EDS spectra of the ARB-1inclusions (Figure 6a 6d) The binding of these metals to the polyP granules contributesto their high electron density PolyP is often associated with polyhydroxybutyrat e (PHB)PHBs solvate (dissolve) cations and associate withpolyP to form Ca2C selective ion channelsacross plasma membranes (Reusch 2000)
The presence of polyP bodies is common in bacteria often resulting as a bacterialresponse to stress and they may play a role in cell motility quorum sensing and bio lmformation (Kornberg and Fraley 2000) Sulfate starvation is known to lead to rapid andmassive accumulation of polyP in some bacteria (Stanier et al 1986) When cells regainaccess to sulfate 32P uptake studies showed that PO3iexcl
4 present in the polyP bodies wasincorporated into nucleic acids PolyP is synthesized from ATP by the enzyme polyP kinasePolyP granules have been reported to be associated with nucleic acids (Harold 1966) butit is not clear whether the association occurs in vivo or after extraction This associationwith nucleic acids would be consistent with staining patterns seen when ARB-1 cells wereexamined with a uorescent livedead stain (nucleic acid stain) displaying a double-lobedstructure in all of the cells (Figure 9) This dumbbell shape appears to extend inside andacross the two phosphorus inclusions
The physiological function of the P inclusions in ARB-1 is not yet known but fromother cell systems several possibilities can be considered These inclusions might serveas a phosphorus reserve for the biosynthesis of nucleic acids and phospholipids underconditions of phosphorus starvation (Harold 1966) Still another possibility is buoyancyregulation Cells can vary their density by as much as 7 by incorporating substances intoinclusions (Guerrero et al 1985) A more speculative possibility is that ARB-1 cells canderive energy from polyP Acinetobacter johnsonii 210a is known to couple the excretion ofa metal phosphate chelate and HC generating a proton motive force (VanVeen et al 1994)A johnsonii takes up P from the aquatic environment by using two Pi transport systemsand stores it as intracellular granules of polyP
Sulfur (S) Inclusions
Cells of ARB-1 contained 5 to 10 spherical S-rich inclusions (Figures 1d) These rangedin size from approximately 100 to 700 nm although most of the S-rich inclusions in asingle cell were close to the same size and were located at one end of the cell oppositethe side where the magnetosomes were located and agellar bundles attached Like theP-rich inclusions the S-rich inclusions appeared to be enclosed in a coating or membrane(Figure 1d) EDS analysis from the ESEM showed these structures contained mostly S withsome carbon and oxygen (Figure 6e) EDS analysis from the TEM also showed S but evenhigher counts for Ca and P (Figure 6b) Beam spreading for the two methods is within aradius of 200 nm The X-rays produced by the 200-keV TEM beam sample the entire verticalextent of the cell unlike the 20 keV SEM beam which loses most of its energy Thereforethe EDS spectra collected at the TEM likely have sampled elements from the cytoplasmand cell membrane components both above and below the inclusion as well as from theP-rich inclusions nearby Other explanations are that hydrated sulfur inclusions changedtheir composition upon dehydration or that these two samples collected at different times
Organization of Inclusions in Magnetococci 401
FIGURE 9 Fluorescence micrograph of an ARB-1 cell suspension treated with Bacliteviability dyes showing that the nucleic acid staining pattern corresponds with the locationof the P-rich inclusions Baclite stains nucleic acids with cytonine (all cells) and propidiumiodate (dead or damaged cells) Bright double-lobed patterns are from ldquoliverdquo cells
of the day and the year may re ect a seasonal diurnal or cell cycle variability in the relativeproportions of the elements
Sulfur inclusions have been found in other uncultured magnetotactic bacteria includ-ing Magnetobacterium bavaricum (Spring et al 1993) Bilophococcus magnetotacticum(Moench 1988) and others (Iida and Akai 1996) and cultured species including the vibrioidstrains MV-1 and MV-2 the coccus strain MC-1 and the spirillum strain MV-4 (Bazylinskiand Frankel 2000) However only in Bilophococcus magnetotacticum was there a similarpolarization of sulfur-containing inclusions from the clustered magnetosomes within thecell (Moench 1988)
It seems likely that ARB-1 oxidizes H2S or other reduced sulfur compounds as anenergy source depositing ldquoelementalrdquo sulfur in intracellular inclusions like many othersul de-oxidizing bacteria including the anaerobic phototrophic purple sulfur bacteria suchas Chromatium vinosum and Thiocapsa roseopersicina the nonsulfur bacteria Beggiatoaspp and Thiothrix spp (Shively 1974 Friedrich 1998) and the magnetotactic bacteriamentioned before In these organisms the presence of sulfur globules is transitory andtakes place when the growth medium contains sul de andor thiosulfate When the sul deis completely utilized the sulfur is converted to sulfate (Stanier et al 1986)
Elemental S in sulfur globules is typically in a liquid state and analysis of in vivoglobules from some members of the purple and green sulfur bacteria by X-ray absorptionnear edge spectroscopy (Prange et al 1999) indicated the sulfur was in a chain structure
402 B L Cox et al
FIGURE 10 Phylogenetic position of six SSU rRNA gene clones related to previouslystudied MC This phylogenetic tree was rooted with Escherichia coli and constructed bythe neighbor joining method with the Kimura 2-parameter model for nucleotide change Amask including all nonambiguously aligned positions for 1427 nucleotides spanning nearlythe full length of the SSU rRNA gene was included Bootstrap values (100 replications)generated by the neighbor joining method are shown above relevant nodes and bootstrapvalues generated by maximum-parsimony analysis are shown below relevant nodes Onlybootstrap values above 50 are shown GenBank accession numbers are MC-CS92 X81182MC-pa119 Y13214 MC-CS81 X81184 MC-TB12 X81183 MC-MP17 X80996 MC-PA11 Y13212 MC12 L06456 partial sequence extended with data from MC-1 genomeRhizobium leguminosarum U31074 Agrobacterium tumefaciens D13294 Rhodobactersphaeroides X53854 Paracoccus denitri cans X59159 Zymomonas mobilis AF281034Sphingomonas paucimobilis D84528 Caulobacter crescentus M83799 Magnetospiril-lum magnetotacticum strain AMB1 D17514 S88429 Magnetospirillum magnetotacticumM10110 Rhodospirillum fulvum M59065 Magnetospirillum gryphiswaldense Y10109Magnetic Vibrio MV-1 LO6455 Oceanospirillum pusillum AB000482 Rhodospirillumrubrum D30778 Roseococcus thiosulfatophilus X72908 Escherichia coli J01695
Organization of Inclusions in Magnetococci 403
and was possibly complexed with water and organic residues If the S globules in ARB-1cells are analogous to those in the purple sulfur bacteria Chromatium vinosum and Thio-capsa roseopersicina they may contain a signi cant protein content Sulfur globules fromthe anaerobic phototrophs C vinosum and T roseopersicina were found to contain upto 52 protein that formed an envelope enclosing the S globules (Brune 1995) Theseproteins were isolated and analyzed and found to be glycine and tyrosine-rich structuralproteins which were homologous for the two species suggesting they are highly con-served (Brune 1995) Phylogenetically C vinosum and T roseopersicina are members ofthe deg -Proteobacteria whereas MC are members of the reg-Proteobacteria To our knowledgemagnetotactic bacteria may be the only members of the reg-Proteobacteria that make internalsulfur globules
Phylogenetic Af liation
We wished to detemine whether ARB-1 was a single species or a mixed population of MCToward that end 45 SSU rRNA genes were cloned from partially puri ed ARB-1 cells30 of which displayed similar restriction fragment length patterns (RFLP) to previouslycharacterized MC (data not shown) The nding that 15 patterns were not related to pre-viously characterized MC was not surprising given that the initial sample was not a pureculture but rather a population magnetically enriched from a sample originating from arelatively large vertical column containing both sediments and overlying water Fourteenof the MC-related clones representing different RFLP pattern groups were chosen for par-tial sequencing 10 of these sequences showed high similarity to previously characterizedMC Six of these sequences were phylogenetically distinct and were fully bidirectionallysequenced
ARB-1 MC SSU rRNA genes formed a monophyletic cluster related to but dis-tinct from other MC (Figure 10) Within the cluster the similarity between SSU rRNAgene sequences ranged from 97 (MC-H1arb and MC-g6arb) to 92 (MC-h1arb andMC-g2arb) The nearest neighbor to the ARB-1 cluster MC-CS92 was ca 89 similarand the nearest cultured relative MC-1 was ca 88 similar to the ARB-1 cluster As arule of thumb for SSU rRNA genes gt97 similarity is considered the same species andgt93ndash95 similarity is considered the same genus (Stackebrandt and Goebel 1994) Thesedata strongly suggest that ARB-1 represents a population of different diverse MC Therewas signi cant variation within the ARB-1 cluster and therefore we do not consider ARB-1to consist of bacteria of a single species or genus Correlations if any between phylogeneticand phenotypic variations in the ARB-1 cells have yet to be determined
Summary
We have partially characterized a population of magnetotactic cocci (MC) referred to asARB-1 which was particularly abundant in iron-rich sediments of a small eutrophic lake andmorphologically and structurally similar to an uncultured species of MC called Bilophococ-cus magnetotacticus The latter organism was found in large numbers in a sewage treatmentpond Cells of ARB-1 had a relatively wide size range of 1 to 4 sup1m They were motileGram-negative and bilophotrichously agellated Cells of this uncultured magnetotacticcoccus contained three types of intracellular inclusions that appeared to be in a consistentarrangement within the cell The magnetosomes contained crystals of magnetite and wereeither in chains andor polar clumps at the end of the cell where the agellar bundles werelocated Large P-rich bodies occupied the majority of the cell volume and appeared to sepa-rate the S-containing inclusions from the magnetosomes The presence of these large P-rich
404 B L Cox et al
inclusions might be an indication of cell stress or may be important in cell buoyancy or inenergy generation The presence of S-rich inclusions located at the end of the cell oppositethe magnetosomes strongly suggests that ARB-1 cells are sul de-oxidizing bacteria andthat the S might be useful as a stored electron donor when cells travel from sul de-rich tosul de-depleted zones in the water column The wide range of the cell sizes the differentmagnetosome arrangements and the large number of different retrieved 16S rRNA genesequences suggest that ARB-1 represents a population of at least several different MC 16SrRNA sequences obtained from cells of ARB-1 appeared to form a monophyletic clusterrelated to but distinct from other uncultured and cultured MC Future work using uores-cent oligonucleotide probes microscopy and physiological measurements should clarifyrelationships between phylogeny cell morphology and organization of inclusions
References
Balkwill DL Maratea D Blakemore RP 1980 Ultrastructure of a magnetotactic spirillum J Bacteriol141(3)1399ndash1408
Bazylinski DA Frankel RB 2000 Biologically controlled mineralization of magnetic iron mineralsby magnetotactic bacteria In Lovley DR editor Environmental microbe-metal interactionsWashington DC ASM Press p 109ndash143
Bazylinski DA Frankel RB Jannasch HW 1988 Anaerobic magnetite production by a marinemagnetotactic bacterium Nature 334518ndash519
Beveridge T 1999 Structures of gram-negative cell walls and their derived membrane vesiclesJ Bacteriol 181(6)4725ndash4733
Beveridge TJ Popkin TJ Cole RM 1994 Electron microscopy In Gerhardt P editor Methods forgeneral and molecular bacteriology Washington DC American Society for Microbiology p 25
Blakemore R 1975 Magnetotactic bacteria Science 190377ndash379Blakemore RP 1982 Magnetotactic bacteria Ann Rev Microbiol 36217ndash238Blakemore RP Maratea D Wolfe RS 1979 Isolation and pure culture of a freshwater magnetic
spirillum in chemically de ned medium J Bacteriol 140(2)720ndash729Blakemore RP Short KA Bazylinski DA Rosenblatt C Frankel RB 1985 Microaerobic conditions
are required for magnetite formation within Aquaspirillum magnetotacticum Geomicrobiol J4(1)53ndash71
Brune DC 1995 Isolation and characterization of sulfur globule proteins from Chromatium vinosumand Thiocapsa roseopersicina Arch Microbiol 163(1)391ndash399
Butler RF Banerjee SK 1975 Theoretical single-domain grain size range in magnetite and titano-magnetite J Geophys Res 804049ndash4058
Buwalda JP 1940 Geology of the Raymond Basin report to Pasadena Water DepartmentCrook R Allen CR Kamb B Payne CM Procter RJ 1987 Recent reverse faulting in the Transverse
Ranges California Pasadena CA US Geol Surv Prof Paper 133927ndash63Devouard B Posfai M Hua X Bazylinski DA Frankel RB Buseck PR 1998 Magnetite from
magnetotactic bacteria size distribution and twinning Am Mineral 831387ndash1398Frankel RB Bazylinski DA Johnson MS Taylor BL 1997 Magneto-Aerotaxis in marine coccoid
bacteria Biophys J 73994ndash1000Friedrich CG 1998 Physiology and genetics of sulfur-oxidizing bacteria Adv Microbial Physiol
39235ndash289Giovannoni SJ 1991 The polymerase chain reaction In Stackebrandt E Goodfellow M editors
Sequencing and hybridization techniques in bacterial systematics New York John Wiley andSons p 177ndash201
Guerrero R Pedros-Alio C Schmidt TM Mas J 1985 A survey of buoyant density of microorganismsin pure cultures and natural samples Microbiologia Sept 1(1ndash2)53ndash65
Harold FM 1966 Inorganic polyphosphates in biology structure metabolism and function BacteriolRev 30(4)772ndash794
Organization of Inclusions in Magnetococci 405
Iida A Akai J 1996 Crystalline sulfur inclusions in magnetotactic bacteria Sci Rep Nigata Univ SerE 1135ndash42
Kornberg A Fraley CD 2000 Inorganic polyphosphate a molecular fossil come to life ASM News66(5)275ndash280
Lins U Farina M 1999 Phosphorus-rich granules in uncultured magnetotactic bacteria FEMSMicrobiol Lett 172(2)23ndash28
Mann S Sparks NHC Frankel RB Bazylinski DA Jannasch HW 1990 Biomineralization of ferri-magnetic greigite (Fe3O4 and iron pyrite (FeS2 ) in a magnetotactic bacterium Nature 343258ndash
261Matsunaga T Sakaguchi T Tadokoro F 1991 Magnetite formation by a magnetic bacterium capable
of growing aerobically Appl Microbiol Biotechnol 35651ndash655Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993a Electron microscopy
study of magnetosomes in a cultured coccoid magnetotactic bacterium Proc Roy Soc Lond B251231ndash236
Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993b Electron microscopystudy of magnetosomes in two cultured vibrioid magnetotactic bacteria Proc Roy Soc Lond B251237ndash242
Moench TT 1988 Bilophococcus magnetotacticus gen nov sp nov a motile magnetic coccus Antvan Leeuwenhoek 54483ndash496
Moench TT Konetzka WA 1978 A novel method for the isolation and study of a magnetotacticbacterium Arch Microbiol 119203ndash212
Murray RGE Robinow CF 1994 Optical Microscopy In Gerhardt P editor Methods for generaland molecular bacteriology Washington DC American Society for Microbiology p 35ndash36
Prange A Arzberger I Engemann C Modrow H Schumann O Truper HG Steudel R Dahl C andHormes J 1999 In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteriaby X-ray absorption near edge spectroscopy Biochim Biophys Acta 1428446ndash454
Rappe MS Kemp PF Giovannoni SJ 1997 Phylogenetic diversity of marine coastal picoplankton16S rRNA genes cloned from the continental shelf off Cape Hatteras North Carolina LimnolOceanogr 42811ndash826
Reusch RN 2000 Transmembrane ion transport by polyphosphatepoly-(R)-3-hydroxybutyratecomplexes Biochemistry (Moscow) 65(3)335ndash352
Sakaguchi T Burgess JG Matsunaga T 1993 Magnetite formation by a sulphate-reducing bacteriumNature 36547ndash49
Schleifer KHSchuler D Spring S Weizenegger M Amann R 1991 The genus Magnetospirillum gennov description of Magnetospirillum gryphiswaldense sp nov and transfer of Aquaspirillummagnetotacticum to Magnetospirillum magnetotacticum comb nov System Appl Microbiol14379ndash385
Schuler D 1999 Formation of magnetosomes in magnetotactic bacteria J Mol Microbiol Biotechnol1(1)79ndash86
Schuler D Spring S Bazylinski DA 1999 Improved technique for the isolation of magnetotactic spir-illa from a freshwater sediment and their phylogenetic characterization System Appl Microbiol22466ndash471
Shively JM 1974 Inclusion bodies of prokaryotes Ann Rev Microbiol 28167ndash187Spring S Amann R Ludwig W Schleifer KH van Gemerden H Petersen N 1993 Dominating role
of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment ApplEnviron Microbiol 59(8)2397ndash2403
Spring S Amann R Ludwig W Schleifer KH Schuler D Poralla K Petersen N 1994 Phylogeneticanalysis of uncultured magnetotactic baceria from the Alpha-subclass of Proteobacteria SystemAppl Microbiol 17501ndash508
Stackebrandt E Goebel BM 1994 Taxonomic note a place for DNA-DNA reassociation and 16SrRNA sequence analysis in the present species de nition in bacteriology Int J System Bacteriol44846ndash849
Stanier RY Ingraham JL Wheelis ML Painter PR 1986 The microbial world 5th edition EnglewoodCliffs NJ Prentice Hall p 179
406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35

388 B L Cox et al
appeared to be enclosed in a membrane or coating Several smaller sulfur-rich inclusionswere located at the end of the cell opposite the agellar bundles The magnetosomesarranged either as a cluster a chain or a combination of both were located proximal tothe two agellar bundles Magnetite was identi ed as the mineral phase of the magneto-somes using selected area electron diffraction (SAED) and by measuring lattice fringespacings of the crystals The magnetite crystals were hexagonal prisms that averaged82 nm in length and thus t into the single-magnetic-domain size range Phylogeneticanalysis of the 16S rRNA gene sequences suggests that it is a mixed population of MCthat form a monophyletic clade distinct from but similar to other uncultured MC
Keywords magnetococci magnetite ESEM TEM sulfur inclusions phosphorusinclusions
Introduction
The magnetotactic bacteria rst discovered by Richard Blakemore in 1974 are prokaryoticmicroorganisms whose direction of motility is in uenced by magnetic elds (Blakemore1975) They are ubiquitous and abundant in aquatic environments including salt marshesfreshwater swamps (Blakemore et al 1979) and sewage oxidation ponds (Moench andKonetzka 1978) These bacteria are diverse morphologically physiologically and phylo-genetically but share a few key characteristics 1) the presence of intracellular membrane-bound magnetic crystals called magnetosomes (Balkwill et al 1980) that are usually butnot always arranged in chains within the cell 2) motility by means of agella and 3) mi-croaerophilic andor anaerobic physiology The function of the magnetosomes appears tobe in the cellrsquos locating and maintaining an optimal position in vertical chemical gradientscommon in aquatic habitats (Frankel et al 1997) The mechanism of formation of the mag-netosomes has not been completely elucidated The biogeochemical roles these bacteriaplay in natural environments are also not understood
Most researchers accept the idea that the primary function of magnetosomes is naviga-tion (Bazylinski and Frankel 2000) although some cells appear to contain more magneto-somes than would be required for navigation (Vali and Kirschvink 1990) Other suggestedfunctions for magnetosomes are iron storage prevention of iron toxicity the use of iron as anenergy source and destruction of toxic oxygen radicals produced during oxygen respiration(Blakemore 1982) It is unlikely that magnetite-containing magnetosomes could be used foriron storage or as an energy source because the iron in magnetosomes is not used by cellseven when they are starved for iron and several species limit their own growth by incor-porating iron into magnetosomes rather than into essential iron-containing proteins (D ABazylinski unpublished results) Many magnetotactic bacteria such as the ones discussedhere also contain inclusions of phosphorus (Blakemore 1982 Lins and Farina 1999) andorsulfur (Moench 1988 Iida and Akai 1996 Bazylinski and Frankel 2000) although moststudies have focussed on magnetosomes In this paper we describe and discuss all threekinds of intracellular structures present in a group of magnetotactic cocci (MC) abundantin a freshwater lake
A major obstacle to studying magnetic bacteria is that they are generally very dif cultto cultivate in pure culture and thus very few pure cultures exist The cultured magne-totactic bacteria include the freshwater spirilla Magnetospirillum magnetotacticum MS-1(Blakemore et al 1979) M magnetotacticum strain AMB1 (Matsunaga et al 1991) andM gryphiswaldense (Schleifer et al 1991 Schuler et al 1999) the marine vibrios MV-1and MV-2 (Bazylinski et al 1988 Meldrum et al 1993b) the curved sulfate-reducing rodstrain RS-1 (Sakaguchi et al 1993) the magnetotactic coccoid strains MC-1 (Meldrumet al 1993a) MC-2 (Devouard et al 1998) and MC-3 (Dennis Bazylinski unpublished)
Organization of Inclusions in Magnetococci 389
The MC were the rst magnetotactic bacteria to be described (Blakemore 1975) and ap-pear to be the most abundant magnetotactic bacteria in freshwater environments with a highorganic content (Schuler 1999)
We report here on a group of uncultured freshwater MC we refer to as ARB-1 whichappear to exhibit some novel behaviors and characteristics These include the reversal ofswimming direction under different conditions possible diurnal effects on the swimmingdirection and a tendency to die sooner if they are in a magnetically shielded environmentAs yet it has not been possible to grow ARB-1 in pure culture However microscopic andchemical studies show that ARB-1 cells have a remarkable organization of interior andexternal structures that might indicate physiological and biogeochemical features of thispopulation
Description of Field Location and Conditions
Cells of ARB-1 were discovered in Baldwin Lake a small lake located within the LosAngeles Arboretum (Arcadia CA USA) Baldwin Lake has been described as a sag pond(Buwalda 1940) and is located on the upthrust side of the Raymond Fault a reverse thrustfault with left-lateral movement (Crook et al 1987) The lake is underlain by Quaternarygravel and alluvium eroded and transported from granodiorite quartz diorite and gneissicsource rocks containing abundant iron-rich hornblende and biotite The shore was arti ciallyplanted with Mediterranean and tropical plants and various chemicals and fertilizers usedin garden maintenance are periodically introduced to the lake ARB-1 cells appeared tobe particularly abundant in locations where the sediments were rich in large fragments ofmetallic iron and magnetic particles
The temperature during the collection period (July 1999ndashOctober 2000) varied from10plusmnC to 28plusmnC The pH of water collected from the lake was 72 to 76 (cells remained viablein bottles in the laboratory at pHs up to 83) The Eh at the surface of the sediment rangedfrom D iexcl315 mV to iexcl183 mV and oxygen from 012 to 017 mgL (38 to 53 sup1M)
Methods
Extraction of the Bacteria
Samples used for cell extraction were taken from 2-liter bottles lled with approximately50 surface sediment and 50 overlying lake water Bottles were taken back to the labora-tory shaken vigorously and magnetic stir bars placed at the watersediment interface alongthe Earthrsquos magnetic eld with the south pole of one magnet on the north side of the bottleand the north pole of the other magnet on the south side of the jar Typically cells accumu-lated as a pinkish pellet at the south-seeking side The pellet was removed with a Pasteurpipette and released into a scintillation vial containing a small amount of lter-sterilizedlake water Magnets were placed on the scintillation vial as described previously and theprocess repeated to further purify the cells
In some experiments we used a modi cation of the racetrack method (Wolfe et al1987 Schuler et al 1999) to separate the magnetic bacteria from nonmagnetic cells Inthis method a sterile cotton plug acting as a barrier for nonmagnetotactic bacteria wasinserted into the wide end of a sealed sterile Pasteur pipette The magnetotactic bacteriaswam to the sealed tip toward the south end of a bar magnet and then the tip containing thecells was broken off and the cells removed The MC typically swim much faster than othermagnetotactic and nonmagnetotactic cells and thus are the rst cells to appear at the tip ofthe pipette
390 B L Cox et al
Optical and Fluorescence Light Microscopy
Optical light microscopy was performed with fresh cells (within 24 h of collection) using aNikon Eclipse E600 microscope with phase contrast Images were obtained with a JavelinMOS solid state camera Flagella were visualized via the Gray staining method using amethod described by Murray and Robinow (1994) Loef errsquos methylene blue stain was usedto test for polyphosphate inclusions and safranin was used to visualize inclusion membranes(Murray and Robinow 1994) Viability was assessed using the Baclite bacterial viabilitykit (Molecular Probes L-7012 stain) This method employs two nucleic acid dyes bothof which are excited with blue (490 nm) light The green dye (cytonine) labels all cellsdead and live while the red dye (propidium iodate) is excluded from living cells but canpenetrate the damaged membranes of dead cells Thus living cells appear green while deadcells appear red
Transmission Electron Microscopy (TEM)
We used a Philips EM 201C microscope operating at 80 kV to examine stained thin sectionsand whole mounts of cell suspensions Cells for thin sections were xed in glutaraldehyderinsed in malate buffer post xed in osmium tetroxide stained with uranyl acetate dehy-drated with ethanol and embedded in EPON resin (Beveridge et al 1994) For negativestains whole cells were deposited on Cu grids coated with Formvar and stained with 2uranyl acetate
A Philips 430 TEM with a single crystal lanthanum hexaboride lament operatingat 200 kV was used for microscopy energy dispersive X-ray spectroscopy (EDS) andselected area electron diffraction (SAED) on fresh (collected within 24 h) unstained cellsuspensions that were deposited on carbon-coated copper grids The area sampled by theEDAX 9900 spectrum analyzer is approximately 200 nm The electron beam was defocusedat the condenser lens to maintain counting rates below 1 kHz and live time ef ciency greaterthan 95 The SAED patterns were recorded on Kodak electron image lm and digitizedfor analysis
Environmental Scanning Electron Microscopy (ESEM)
A Philips XL30 ESEM with a eld emission gun was used to observe hydrated un xed cellsImages were acquired at 20 kV in ldquowet moderdquo (3plusmnC 53 Torr chamber and water vaporpressure) at a relative humidity in the sample chamber of 93 The EDS was performedon fully hydrated uncoated and un xed specimens using an accelerating voltage of 20 kVand counting for 100 s (live time) The spot size sampled by the EDS detector was 20 nmbut because of beam spread signals from elements within a radius of 100 nm may also becounted In some cases cells were xed in glutaraldehyde prior to observation (images notshown) This had a tendency to shrink the cells to 60 of their original size For all otherexperiments fresh cells (within 24 h of collection) were used
Molecular Diversity
Bacterial small subunit (SSU) rRNA genes were PCR-ampli ed using the primers B11Fand B1512r (Giovannoni 1991) Five aliquots of cells (ca 108 each) were placed in separatetubes containing 18 sup1l of PCR cocktail (1 pound Taq DNA polymerase buffer with MgCl2 1 Uof Taq polymerase [Promega Madison WI USA] 200 sup1M dNTPs 750 nM each primer)Cells were lysed by incubation for 6 min at 94plusmnC and then subjected to PCR thermalcycling PCR conditions were 30 sec 94plusmnC denaturation 1 min 50plusmnC annealing and
Organization of Inclusions in Magnetococci 391
2 min 72plusmnC elongation for 35 cycles in an MJ Research thermal cycler (Waltham MAUSA) After a nal 10 min incubation at 72plusmnC the products were combined and puri edby gel extraction as recommended by the manufacturer (Qiagen Chatsworth CA) Am-pli cation products were cloned into the plasmid vector pCR21 as recommended by themanufacturer (Invitrogen Carlsbad CA)
SSU rDNA inserts were PCR-ampli ed under the same conditions as above Theproduct was double-digested with HhaI (New England Biolabs Beverly MA) and RsaI(Promega) restriction endonucleases in 1 pound Promega digestion buffer at 37plusmnC for 3 h Thedigested DNA was separated by electrophoresis through low melting point agarose (20wtvol IBI Shelton Scienti c Shelton CT USA) The banding patterns were grouped ac-cording to similarity and representative members of groups displaying patterns similar tothose of MC we had previously sequenced (data not shown) were partially sequenced Sixpartial sequences that were tentatively identi ed as MC and as different groups were sub-sequently fully bidirectionally sequenced with either an ABI 3700 (Applied BiosystemsFoster City CA) or a Licor 4200 (Licor Lincoln NE USA) automated DNA sequencerand dye terminator chemistry
Sequences were initially aligned to their nearest neighbor using the program ARB pro-vided by Wolfgang Ludwig and Oliver Strunk (Technische Universitat Munchen MunichGermany program available at httpwwwtumunchendeARB) The sequences were fur-ther manually aligned to sequences obtained from the GenBank database using the GeneticData Environment (GDE) v 20 sequence analysis software package provided by SteveSmith (Millipore Corporation Bedford MA) and described by Rappe et al (1997) Phy-logenetic inference and evolutionary distance calculations were performed as described inRappe et al (1997) Regions of ambiguous alignment were excluded from analyses
Results and Discussion
General Description of Cells
Cells of ARB-1 were spherical to oblate spheroidal Gram-negative cells ranging from1 to 4 sup1m (Figure 1) and averaging 25 sup1m In the ESEM the average size of cells was16 sup1m if xed with glutaraldehyde (aspect ratio 085) and 27 sup1m if not xed (aspectratio 085) The wide range of cells sizes (1ndash4 sup1m) might re ect the phylogenetic diversity(described below) or indicate that there are different phases in the cell cycle re ected incell size or that cells simply exhibit budding Cells contained two large phosphorus-richsymmetrical intracellular bodies that took up a large fraction of the cell volume Otherobserved inclusions were sulfur globules and magnetosomes Phosphorus-rich inclusionsin ESEM images (Figure 1d) appeared to be wider apart at one end forming a crevicewhere many small sulfur-containing inclusions were located On the opposite side of mostcells where the phosphorus inclusions appeared to be closer together magnetosomes werearranged as a cluster of disorganized crystals as chains or both (Figure 1d)
Several cells in the TEM (Figure 2) and in the ESEM appeared to be in the processof dividing In some cases indentations preceding cell division appeared to occur betweenthe two phosphorus inclusions in others these constrictions seemed to separate the sulfurinclusions from the rest of the cell (Figure 2) We also observed protrusions or extensionson some of the dividing cells (Figure 2c d) The large differences in cell size may well bedue to unequal division (budding) observed in some cells during division (Figure 2) Thecylindrical structure of one of these (Figure 2d) is similar to the basal body of a agellabundle Other possible explanations for these protrusions are that they are membrane vesicles(Beveridge 1999) or some artifact of electron microscopy or staining
392 B L Cox et al
FIGURE 1 Images of ARB-1 cells showing inclusions containing phosphorus (P) sul-fur (S) and magnetite (M) and variations in cell sizes a) Phase contrast light microscopyimage of live cells b) TEM image of unstained cells xed in glutaraldehyde (image taken byH Vali) c) TEM image of unstained and un xed cell showing S-rich globules P-rich in-clusions and magnetosomes (M) in chains d) ESEM image of hydrated un xed cells on amembrane lter H indicates ldquohingedrdquo P inclusions
The swimming speed of ARB-1 cells in wet mounts or in capillary racetracks wasextremely rapid up to a millimeter or 1000 body lengths per second for fresh cells whendirected by magnetic stir bars The phosphorus-rich inclusions appeared to wobble from sideto side or may even have opened and closed slightly as if hinged on one side (Figure 1d)as the cells swam
Organization of Inclusions in Magnetococci 393
FIGURE 2 Images of ARB-1 cells showing cell division a) TEM image of cells stainedwith uranyl acetate showing division across rather than between P inclusions b) ESEMimage showing division of cells between P inclusions c) d) TEM images of cells stainedwith uranyl acetate showing unequal division of cells with protrusions
Flagella and Fimbriae or Pili
Like other MC (Blakemore 1975 1982 Moench 1988 Frankel et al 1997) cells of ARB-1were bilophotrichously agellated having two agellar bundles on one side of the cell(Figure 3) Each bundle contained about 10 unsheathed agella that were 13 nm in diameterCells also appeared to have other smaller protrusions that resemble pili (Figure 3b) Thetendency for cells of ARB-1 to stick to glass slides and cover slips and to each other supportsthe presence of mbriae or pili Pili have been observed on other uncultured (Blakemore1982) and cultured (Frankel et al 1997) MC
Magnetosomes
Magnetosome crystals in ARB-1 cells occurred in chains and clusters and ranged in lengthfrom 20 to 133 nm and in width from 10 to 84 nm with an average (sect standard deviation)length and width of 82 sect 23 nm and 49 sect 15 nm respectively (n D 341) The average lengthof clustered magnetosomes (n D 260) was 83 sect 24 nearly the same as the average lengthof magnetosomes from chains (n D 81 mean D 80 sect 19) The range of lengths of magneto-somes in chains (40ndash120 nm) was narrower than that of clustered magnetosomes The size
394 B L Cox et al
FIGURE 3 Images of ARB-1 cells showing agella andor pili a) Light microscopy imageof cells stained for agella Arrows indicate possible cell division plane b) Thin section of acell stained with uranyl acetate showing spaces where P inclusions had been S inclusionsmagnetosomes (M) and pili c) TEM image of whole cells with clustered magnetosomecrystals (M) negatively stained with uranyl acetate Notice haloes around crystals whichmight represent a membrane or coating
distributions (length and width) of all 341 magnetite crystals from cells of ARB-1 are plot-ted in Figure 4 Values plotted for ARB-1 magnetite crystals showed that like in virtually allother magnetotactic bacteria the magnetite particles in ARB-1 are single-magnetic-domaincrystals (Butler and Banerjee 1975 Bazylinski and Frankel 2000) These results were verysimilar to those obtained for the uncultured coccus Bilophococcus magnetotacticus (Toweand Moench 1981) which also possessed clumps of magnetosomes at one end of the cellrather than chains The average length of ARB-1 magnetosomes was also similar to thatof the cultured MC-1 However the shape of MC-1 magnetosomes was less elongated(Meldrum et al 1993a) It appears that the size distributions and aspect ratios for this typeof MC are distinct from those of other magnetotactic bacteria (eg MV-1 and MC-1) aswell as from those in the Mars meteorite ALH84001 (Thomas-Keprta et al 2000)
Morphologically the magnetosome crystals in ARB-1 were rectangular in projectionwith an hexagonal cross-section There were irregularities in the edges and interiors of the
FIG
UR
E4
Dis
trib
utio
nof
leng
ths
and
wid
ths
of34
1m
agne
tite
crys
tals
from
clus
ters
and
chai
nsin
AR
B-1
cells
a) F
requ
ency
asa
func
tion
ofle
ngth
b)
Freq
uenc
yas
afu
nctio
nof
wid
thc
) Wid
thas
afu
nctio
nof
leng
th
395
396 B L Cox et al
FIGURE 5 TEM image of a thin section of a cell of ARB-1 stained with uranyl acetateshowing magnetosomes before a) and after b) beam damage leaving crystal ldquodoubletsrdquo b)Notice disappearance of electron dense material in b)
crystals (Figure 3) The edges were often rounded and the particles occasionally con-stricted slightly in the middle The crystals sometimes had a mottled appearance and mayhave had fringes around the edges Once during TEM analysis of thin sections (80 kV)magnetosome crystals sustained signi cant beam damage after several minutes of viewingat a magni cation of 100000 (Figure 5) The crystals appeared to have broken in halfpossibly along twin planes in the (111) direction or along ldquowaistedrdquo regions where thecrystals showed some constriction and there appeared to be a loss of material Beam dam-age has been previously observed during TEM of greigite (Fe3S4) magnetosome particlesbut not in magnetite crystals (D Bazylinski H Vali and J Kirschvink personal commu-nications January 2001) except when viewed as whole mounts at 400 kV (Taylor et al2001) Taylor et al (2001) speculated that water vapor from the cells underwent radiolysisin the electron-beam producing a plasma that rapidly melted the magnetosomes
EDS analyses with both TEM and ESEM of the magnetosome particles (Figure 6c 6f)showed the particles were iron-rich with some extraneous signals including sulfur (Figure 6)A strong sulfur signal would be expected from EDS analyses of the magnetic mineral greigitealso known to be produced by some magnetotactic bacteria (Mann et al 1990) In ARB-1however these extraneous signals are likely due to the sulfur- and phosphorus-containinginclusions also present in the cell SAED patterns obtained from the crystals (Figure 7)and lattice fringes from high resolution TEM with spacings of 048 nm elongated in (111)direction (Figure 7) were consistent with the mineral magnetite
Magnetosomes within cells of ARB-1 were found in three different arrangements1) as a polar cluster forming a sort of ldquocaprdquo usually at the agellar end of the cell 2) in partialor complete chains at the same end of the cell or 3) a combination of both arrangementsThe polar clusters appear to be disorganized in many images (eg Figures 1 2 and 3)However the attraction of these cells to a magnet indicates they have permanent magneticdipole moments implying that they are not disorganized We are currently pursuing thisquestion of whether or not the clusters are disorganized
In cells having both chains and clusters it is dif cult to determine whether chainswere forming from the clustered crystals or whether the polar clusters of particles werefrom a chain that was disrupted during dehydration of the cell during preparation for TEMexamination However polar clusters of crystals were observed in ESEM images of many
Organization of Inclusions in Magnetococci 397
FIGURE 6 EDS spectra from TEM a) b) c) and ESEM d) e) f) showing elementalcomposition of P-rich a) d) and S-rich b) e) inclusions and magnetosomes c) f) Elementalcompositions of clustered magnetosomes were identical to those of magnetosomes in chainsAll y-axes are linear except for a) and b) which are log scales Noise in f ) is due to scattering
hydrated cells (Figure 1d) suggesting that they were not caused by dehydration In manyof the cells viewed by TEM it seemed that the chains tended to occur in smaller morespherical cells while clumps of magnetosomes occurred more frequently in elongatedcells In order to see if there was any relationship between cell size and shape and the threetypes of magnetosome arrangements the size and aspect ratio for cells with these differentmagnetosome arrangements were measured (Figure 8) The cell size distribution appearedto be narrower for cells with chains than for the cells with disorganized crystals Cells withchains were predominantly smaller than 25 sup1m (Figure 8a) The aspect ratios of the cellswith chains only was smaller than that of cells with disorganized magnetosomes and wasshifted toward spherical aspect ratios (Figure 8b)
One explanation for these observations is that there are different stages of magnetosomeformation in a single species of MC The disorganized magnetosomes might form near thecell membrane between the agellar bundles and then extend into chains Alternativelymagnetosomes might rst form as chains in smaller cells At a later stage as cells increasein size the membranes linking the chains dissolve or become stressed and the magneto-somes somehow migrate to the agellar end of the cell Because magnetosomes appear tobe anchored to the inner surface of the cell membrane in virtually all species examined(Bazylinski and Frankel 2000) and are never free- oating in the cytoplasm migration ofmagnetosomes through the cytoplasm seems unlikely Instead we are presently designinghybridization experiments to address the hypothesis that the relationships between cell sizeshape and magnetosome arrangement correlate with the genotypical variations in ARB-1described later Spring et al (1994) analyzed communities of MC found in freshwater lakesin Germany using uorescent oligonucleotide probes They found different genotypes ofMC (similarity below 95) but these genotypes were not phenotypically expressed asdifferent magnetosome organizations but as preferences for different concentrations of O2
398 B L Cox et al
FIGURE 7 Evidence supporting magnetite as the magnetosome mineral in ARB-1 cellsa) Image of a single crystal of magnetite within a cell in an unstained glutaraldehyde- xedwhole cell mount taken with High Resolution TEM by H Vali b) Part of the magnetitecrystal shown in a) magni ed to show the distance between lattice fringes c) Selectedarea electron diffraction (SAED) of a group of magnetosomes in a cell from an un xedunstained whole cell mount showing the indices of some of the diffraction spots
Therefore we might expect no correlation between magnetite organization and phyloge-netic diversity of ARB-1 cells but perhaps a correlation between oxygen preference andphylogeny
A third possibility is that chains and clumps of magnetosomes form as a result ofdifferent environmental conditions Since our sampling methods were not selective as tothe cellsrsquo location in the vertical chemical (eg oxygen) and redox gradients that are sure to
Organization of Inclusions in Magnetococci 399
FIGURE 8 Relationships between organization of magnetosomes and cell size a) and cellaspect ratio b) Measurements were taken from both TEM and ESEM images Solid patternis for cells with clustered magnetosomes crossed pattern is for cells with magnetosomesin chains only Patterned bars originate at the top of solid bars
exist at the site the environmental conditions of our samples represent a relatively large rangeof oxygen and perhaps other chemical concentrations as well as redox conditions Magneticbacteria in culture are known to vary the size and number of magnetosomes in responseto the media composition (Meldrum et al 1993a) and particularly oxygen concentration(Blakemore et al 1985) Another possibility is that the clustered magnetosomes are formedby a different biological mechanism and serve a different function than magnetosomes inchains
Phosphorus (P) Inclusions
The two very large inclusion bodies seen in each cell always occurred in pairs in cellsand seemed to extend to the cell membrane and to be in contact with each other Thus themaximum dimension of the P-rich inclusions was the same as the minimum dimension ofthe cell and their combined width was approximately half of the long dimension of the cellIn ESEM images the two P-containing inclusions appeared to be attached in the middle insome cells and separated in others (Figure 1d) The P-containing inclusions appeared to beself-contained and perhaps bounded by some sort of coating or membrane in TEM images(Figure 2) and in light microscopy images of cells stained with safranin (not shown)
These inclusions were shown by both ESEM and TEM EDS to contain phosphorus with generally small amounts of potassium calcium magnesium sulfur and iron the lattertwo elements likely being due to the close proximity of the P-rich inclusions to sulfurglobules and magnetosomes (Figures 1 and 6) A very large calcium peak was observedwhen these large inclusions were analyzed in the TEM (Figure 6a) The samples shown inFigures 6a and 6d were collected at different times of the year which may explain some ofthe differences observed in these spectra Other EDS data from the ESEM (not shown) hadhigher calcium peaks than those shown in Figure 6d indicating that relative proportions ofP and Ca in P-rich inclusions vary even within the same sample
The inclusions were electron dense in TEM and had a mottled appearance due toablation by the electron beam Similar inclusions have been described for uncultured MCfrom the Itaupo Lagoon near Rio de Janeiro Brazil (Lins and Farina 1999) and from MillPond New Hampshire (Blakemore 1982) Like the structures in ARB-1 polyP granules aretranslucent in visible light (Spring et al 1994) dark (electron dense) in the TEM and oftenhave a mottled appearance (Harold 1966) The P-rich inclusions in ARB-1 cells stained
400 B L Cox et al
with methylene blue (Murray and Robinson 1994) which is suggestive of the presence ofpolyP but not con rmatory
PolyP is composed of linear chains of tetrahedral phosphate residues linked throughcommon oxygen atoms by phosphoanhydrid e bonds At physiological pH each unit carriesa monovalent negative charge giving polyP the property of having a large cation exchangecapacity The binding energy of polyP allows it to sequester Ca2C and Mg2C (Reusch 2000)consistent with the Ca2C and Mg2C signals observed in the EDS spectra of the ARB-1inclusions (Figure 6a 6d) The binding of these metals to the polyP granules contributesto their high electron density PolyP is often associated with polyhydroxybutyrat e (PHB)PHBs solvate (dissolve) cations and associate withpolyP to form Ca2C selective ion channelsacross plasma membranes (Reusch 2000)
The presence of polyP bodies is common in bacteria often resulting as a bacterialresponse to stress and they may play a role in cell motility quorum sensing and bio lmformation (Kornberg and Fraley 2000) Sulfate starvation is known to lead to rapid andmassive accumulation of polyP in some bacteria (Stanier et al 1986) When cells regainaccess to sulfate 32P uptake studies showed that PO3iexcl
4 present in the polyP bodies wasincorporated into nucleic acids PolyP is synthesized from ATP by the enzyme polyP kinasePolyP granules have been reported to be associated with nucleic acids (Harold 1966) butit is not clear whether the association occurs in vivo or after extraction This associationwith nucleic acids would be consistent with staining patterns seen when ARB-1 cells wereexamined with a uorescent livedead stain (nucleic acid stain) displaying a double-lobedstructure in all of the cells (Figure 9) This dumbbell shape appears to extend inside andacross the two phosphorus inclusions
The physiological function of the P inclusions in ARB-1 is not yet known but fromother cell systems several possibilities can be considered These inclusions might serveas a phosphorus reserve for the biosynthesis of nucleic acids and phospholipids underconditions of phosphorus starvation (Harold 1966) Still another possibility is buoyancyregulation Cells can vary their density by as much as 7 by incorporating substances intoinclusions (Guerrero et al 1985) A more speculative possibility is that ARB-1 cells canderive energy from polyP Acinetobacter johnsonii 210a is known to couple the excretion ofa metal phosphate chelate and HC generating a proton motive force (VanVeen et al 1994)A johnsonii takes up P from the aquatic environment by using two Pi transport systemsand stores it as intracellular granules of polyP
Sulfur (S) Inclusions
Cells of ARB-1 contained 5 to 10 spherical S-rich inclusions (Figures 1d) These rangedin size from approximately 100 to 700 nm although most of the S-rich inclusions in asingle cell were close to the same size and were located at one end of the cell oppositethe side where the magnetosomes were located and agellar bundles attached Like theP-rich inclusions the S-rich inclusions appeared to be enclosed in a coating or membrane(Figure 1d) EDS analysis from the ESEM showed these structures contained mostly S withsome carbon and oxygen (Figure 6e) EDS analysis from the TEM also showed S but evenhigher counts for Ca and P (Figure 6b) Beam spreading for the two methods is within aradius of 200 nm The X-rays produced by the 200-keV TEM beam sample the entire verticalextent of the cell unlike the 20 keV SEM beam which loses most of its energy Thereforethe EDS spectra collected at the TEM likely have sampled elements from the cytoplasmand cell membrane components both above and below the inclusion as well as from theP-rich inclusions nearby Other explanations are that hydrated sulfur inclusions changedtheir composition upon dehydration or that these two samples collected at different times
Organization of Inclusions in Magnetococci 401
FIGURE 9 Fluorescence micrograph of an ARB-1 cell suspension treated with Bacliteviability dyes showing that the nucleic acid staining pattern corresponds with the locationof the P-rich inclusions Baclite stains nucleic acids with cytonine (all cells) and propidiumiodate (dead or damaged cells) Bright double-lobed patterns are from ldquoliverdquo cells
of the day and the year may re ect a seasonal diurnal or cell cycle variability in the relativeproportions of the elements
Sulfur inclusions have been found in other uncultured magnetotactic bacteria includ-ing Magnetobacterium bavaricum (Spring et al 1993) Bilophococcus magnetotacticum(Moench 1988) and others (Iida and Akai 1996) and cultured species including the vibrioidstrains MV-1 and MV-2 the coccus strain MC-1 and the spirillum strain MV-4 (Bazylinskiand Frankel 2000) However only in Bilophococcus magnetotacticum was there a similarpolarization of sulfur-containing inclusions from the clustered magnetosomes within thecell (Moench 1988)
It seems likely that ARB-1 oxidizes H2S or other reduced sulfur compounds as anenergy source depositing ldquoelementalrdquo sulfur in intracellular inclusions like many othersul de-oxidizing bacteria including the anaerobic phototrophic purple sulfur bacteria suchas Chromatium vinosum and Thiocapsa roseopersicina the nonsulfur bacteria Beggiatoaspp and Thiothrix spp (Shively 1974 Friedrich 1998) and the magnetotactic bacteriamentioned before In these organisms the presence of sulfur globules is transitory andtakes place when the growth medium contains sul de andor thiosulfate When the sul deis completely utilized the sulfur is converted to sulfate (Stanier et al 1986)
Elemental S in sulfur globules is typically in a liquid state and analysis of in vivoglobules from some members of the purple and green sulfur bacteria by X-ray absorptionnear edge spectroscopy (Prange et al 1999) indicated the sulfur was in a chain structure
402 B L Cox et al
FIGURE 10 Phylogenetic position of six SSU rRNA gene clones related to previouslystudied MC This phylogenetic tree was rooted with Escherichia coli and constructed bythe neighbor joining method with the Kimura 2-parameter model for nucleotide change Amask including all nonambiguously aligned positions for 1427 nucleotides spanning nearlythe full length of the SSU rRNA gene was included Bootstrap values (100 replications)generated by the neighbor joining method are shown above relevant nodes and bootstrapvalues generated by maximum-parsimony analysis are shown below relevant nodes Onlybootstrap values above 50 are shown GenBank accession numbers are MC-CS92 X81182MC-pa119 Y13214 MC-CS81 X81184 MC-TB12 X81183 MC-MP17 X80996 MC-PA11 Y13212 MC12 L06456 partial sequence extended with data from MC-1 genomeRhizobium leguminosarum U31074 Agrobacterium tumefaciens D13294 Rhodobactersphaeroides X53854 Paracoccus denitri cans X59159 Zymomonas mobilis AF281034Sphingomonas paucimobilis D84528 Caulobacter crescentus M83799 Magnetospiril-lum magnetotacticum strain AMB1 D17514 S88429 Magnetospirillum magnetotacticumM10110 Rhodospirillum fulvum M59065 Magnetospirillum gryphiswaldense Y10109Magnetic Vibrio MV-1 LO6455 Oceanospirillum pusillum AB000482 Rhodospirillumrubrum D30778 Roseococcus thiosulfatophilus X72908 Escherichia coli J01695
Organization of Inclusions in Magnetococci 403
and was possibly complexed with water and organic residues If the S globules in ARB-1cells are analogous to those in the purple sulfur bacteria Chromatium vinosum and Thio-capsa roseopersicina they may contain a signi cant protein content Sulfur globules fromthe anaerobic phototrophs C vinosum and T roseopersicina were found to contain upto 52 protein that formed an envelope enclosing the S globules (Brune 1995) Theseproteins were isolated and analyzed and found to be glycine and tyrosine-rich structuralproteins which were homologous for the two species suggesting they are highly con-served (Brune 1995) Phylogenetically C vinosum and T roseopersicina are members ofthe deg -Proteobacteria whereas MC are members of the reg-Proteobacteria To our knowledgemagnetotactic bacteria may be the only members of the reg-Proteobacteria that make internalsulfur globules
Phylogenetic Af liation
We wished to detemine whether ARB-1 was a single species or a mixed population of MCToward that end 45 SSU rRNA genes were cloned from partially puri ed ARB-1 cells30 of which displayed similar restriction fragment length patterns (RFLP) to previouslycharacterized MC (data not shown) The nding that 15 patterns were not related to pre-viously characterized MC was not surprising given that the initial sample was not a pureculture but rather a population magnetically enriched from a sample originating from arelatively large vertical column containing both sediments and overlying water Fourteenof the MC-related clones representing different RFLP pattern groups were chosen for par-tial sequencing 10 of these sequences showed high similarity to previously characterizedMC Six of these sequences were phylogenetically distinct and were fully bidirectionallysequenced
ARB-1 MC SSU rRNA genes formed a monophyletic cluster related to but dis-tinct from other MC (Figure 10) Within the cluster the similarity between SSU rRNAgene sequences ranged from 97 (MC-H1arb and MC-g6arb) to 92 (MC-h1arb andMC-g2arb) The nearest neighbor to the ARB-1 cluster MC-CS92 was ca 89 similarand the nearest cultured relative MC-1 was ca 88 similar to the ARB-1 cluster As arule of thumb for SSU rRNA genes gt97 similarity is considered the same species andgt93ndash95 similarity is considered the same genus (Stackebrandt and Goebel 1994) Thesedata strongly suggest that ARB-1 represents a population of different diverse MC Therewas signi cant variation within the ARB-1 cluster and therefore we do not consider ARB-1to consist of bacteria of a single species or genus Correlations if any between phylogeneticand phenotypic variations in the ARB-1 cells have yet to be determined
Summary
We have partially characterized a population of magnetotactic cocci (MC) referred to asARB-1 which was particularly abundant in iron-rich sediments of a small eutrophic lake andmorphologically and structurally similar to an uncultured species of MC called Bilophococ-cus magnetotacticus The latter organism was found in large numbers in a sewage treatmentpond Cells of ARB-1 had a relatively wide size range of 1 to 4 sup1m They were motileGram-negative and bilophotrichously agellated Cells of this uncultured magnetotacticcoccus contained three types of intracellular inclusions that appeared to be in a consistentarrangement within the cell The magnetosomes contained crystals of magnetite and wereeither in chains andor polar clumps at the end of the cell where the agellar bundles werelocated Large P-rich bodies occupied the majority of the cell volume and appeared to sepa-rate the S-containing inclusions from the magnetosomes The presence of these large P-rich
404 B L Cox et al
inclusions might be an indication of cell stress or may be important in cell buoyancy or inenergy generation The presence of S-rich inclusions located at the end of the cell oppositethe magnetosomes strongly suggests that ARB-1 cells are sul de-oxidizing bacteria andthat the S might be useful as a stored electron donor when cells travel from sul de-rich tosul de-depleted zones in the water column The wide range of the cell sizes the differentmagnetosome arrangements and the large number of different retrieved 16S rRNA genesequences suggest that ARB-1 represents a population of at least several different MC 16SrRNA sequences obtained from cells of ARB-1 appeared to form a monophyletic clusterrelated to but distinct from other uncultured and cultured MC Future work using uores-cent oligonucleotide probes microscopy and physiological measurements should clarifyrelationships between phylogeny cell morphology and organization of inclusions
References
Balkwill DL Maratea D Blakemore RP 1980 Ultrastructure of a magnetotactic spirillum J Bacteriol141(3)1399ndash1408
Bazylinski DA Frankel RB 2000 Biologically controlled mineralization of magnetic iron mineralsby magnetotactic bacteria In Lovley DR editor Environmental microbe-metal interactionsWashington DC ASM Press p 109ndash143
Bazylinski DA Frankel RB Jannasch HW 1988 Anaerobic magnetite production by a marinemagnetotactic bacterium Nature 334518ndash519
Beveridge T 1999 Structures of gram-negative cell walls and their derived membrane vesiclesJ Bacteriol 181(6)4725ndash4733
Beveridge TJ Popkin TJ Cole RM 1994 Electron microscopy In Gerhardt P editor Methods forgeneral and molecular bacteriology Washington DC American Society for Microbiology p 25
Blakemore R 1975 Magnetotactic bacteria Science 190377ndash379Blakemore RP 1982 Magnetotactic bacteria Ann Rev Microbiol 36217ndash238Blakemore RP Maratea D Wolfe RS 1979 Isolation and pure culture of a freshwater magnetic
spirillum in chemically de ned medium J Bacteriol 140(2)720ndash729Blakemore RP Short KA Bazylinski DA Rosenblatt C Frankel RB 1985 Microaerobic conditions
are required for magnetite formation within Aquaspirillum magnetotacticum Geomicrobiol J4(1)53ndash71
Brune DC 1995 Isolation and characterization of sulfur globule proteins from Chromatium vinosumand Thiocapsa roseopersicina Arch Microbiol 163(1)391ndash399
Butler RF Banerjee SK 1975 Theoretical single-domain grain size range in magnetite and titano-magnetite J Geophys Res 804049ndash4058
Buwalda JP 1940 Geology of the Raymond Basin report to Pasadena Water DepartmentCrook R Allen CR Kamb B Payne CM Procter RJ 1987 Recent reverse faulting in the Transverse
Ranges California Pasadena CA US Geol Surv Prof Paper 133927ndash63Devouard B Posfai M Hua X Bazylinski DA Frankel RB Buseck PR 1998 Magnetite from
magnetotactic bacteria size distribution and twinning Am Mineral 831387ndash1398Frankel RB Bazylinski DA Johnson MS Taylor BL 1997 Magneto-Aerotaxis in marine coccoid
bacteria Biophys J 73994ndash1000Friedrich CG 1998 Physiology and genetics of sulfur-oxidizing bacteria Adv Microbial Physiol
39235ndash289Giovannoni SJ 1991 The polymerase chain reaction In Stackebrandt E Goodfellow M editors
Sequencing and hybridization techniques in bacterial systematics New York John Wiley andSons p 177ndash201
Guerrero R Pedros-Alio C Schmidt TM Mas J 1985 A survey of buoyant density of microorganismsin pure cultures and natural samples Microbiologia Sept 1(1ndash2)53ndash65
Harold FM 1966 Inorganic polyphosphates in biology structure metabolism and function BacteriolRev 30(4)772ndash794
Organization of Inclusions in Magnetococci 405
Iida A Akai J 1996 Crystalline sulfur inclusions in magnetotactic bacteria Sci Rep Nigata Univ SerE 1135ndash42
Kornberg A Fraley CD 2000 Inorganic polyphosphate a molecular fossil come to life ASM News66(5)275ndash280
Lins U Farina M 1999 Phosphorus-rich granules in uncultured magnetotactic bacteria FEMSMicrobiol Lett 172(2)23ndash28
Mann S Sparks NHC Frankel RB Bazylinski DA Jannasch HW 1990 Biomineralization of ferri-magnetic greigite (Fe3O4 and iron pyrite (FeS2 ) in a magnetotactic bacterium Nature 343258ndash
261Matsunaga T Sakaguchi T Tadokoro F 1991 Magnetite formation by a magnetic bacterium capable
of growing aerobically Appl Microbiol Biotechnol 35651ndash655Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993a Electron microscopy
study of magnetosomes in a cultured coccoid magnetotactic bacterium Proc Roy Soc Lond B251231ndash236
Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993b Electron microscopystudy of magnetosomes in two cultured vibrioid magnetotactic bacteria Proc Roy Soc Lond B251237ndash242
Moench TT 1988 Bilophococcus magnetotacticus gen nov sp nov a motile magnetic coccus Antvan Leeuwenhoek 54483ndash496
Moench TT Konetzka WA 1978 A novel method for the isolation and study of a magnetotacticbacterium Arch Microbiol 119203ndash212
Murray RGE Robinow CF 1994 Optical Microscopy In Gerhardt P editor Methods for generaland molecular bacteriology Washington DC American Society for Microbiology p 35ndash36
Prange A Arzberger I Engemann C Modrow H Schumann O Truper HG Steudel R Dahl C andHormes J 1999 In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteriaby X-ray absorption near edge spectroscopy Biochim Biophys Acta 1428446ndash454
Rappe MS Kemp PF Giovannoni SJ 1997 Phylogenetic diversity of marine coastal picoplankton16S rRNA genes cloned from the continental shelf off Cape Hatteras North Carolina LimnolOceanogr 42811ndash826
Reusch RN 2000 Transmembrane ion transport by polyphosphatepoly-(R)-3-hydroxybutyratecomplexes Biochemistry (Moscow) 65(3)335ndash352
Sakaguchi T Burgess JG Matsunaga T 1993 Magnetite formation by a sulphate-reducing bacteriumNature 36547ndash49
Schleifer KHSchuler D Spring S Weizenegger M Amann R 1991 The genus Magnetospirillum gennov description of Magnetospirillum gryphiswaldense sp nov and transfer of Aquaspirillummagnetotacticum to Magnetospirillum magnetotacticum comb nov System Appl Microbiol14379ndash385
Schuler D 1999 Formation of magnetosomes in magnetotactic bacteria J Mol Microbiol Biotechnol1(1)79ndash86
Schuler D Spring S Bazylinski DA 1999 Improved technique for the isolation of magnetotactic spir-illa from a freshwater sediment and their phylogenetic characterization System Appl Microbiol22466ndash471
Shively JM 1974 Inclusion bodies of prokaryotes Ann Rev Microbiol 28167ndash187Spring S Amann R Ludwig W Schleifer KH van Gemerden H Petersen N 1993 Dominating role
of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment ApplEnviron Microbiol 59(8)2397ndash2403
Spring S Amann R Ludwig W Schleifer KH Schuler D Poralla K Petersen N 1994 Phylogeneticanalysis of uncultured magnetotactic baceria from the Alpha-subclass of Proteobacteria SystemAppl Microbiol 17501ndash508
Stackebrandt E Goebel BM 1994 Taxonomic note a place for DNA-DNA reassociation and 16SrRNA sequence analysis in the present species de nition in bacteriology Int J System Bacteriol44846ndash849
Stanier RY Ingraham JL Wheelis ML Painter PR 1986 The microbial world 5th edition EnglewoodCliffs NJ Prentice Hall p 179
406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35

Organization of Inclusions in Magnetococci 389
The MC were the rst magnetotactic bacteria to be described (Blakemore 1975) and ap-pear to be the most abundant magnetotactic bacteria in freshwater environments with a highorganic content (Schuler 1999)
We report here on a group of uncultured freshwater MC we refer to as ARB-1 whichappear to exhibit some novel behaviors and characteristics These include the reversal ofswimming direction under different conditions possible diurnal effects on the swimmingdirection and a tendency to die sooner if they are in a magnetically shielded environmentAs yet it has not been possible to grow ARB-1 in pure culture However microscopic andchemical studies show that ARB-1 cells have a remarkable organization of interior andexternal structures that might indicate physiological and biogeochemical features of thispopulation
Description of Field Location and Conditions
Cells of ARB-1 were discovered in Baldwin Lake a small lake located within the LosAngeles Arboretum (Arcadia CA USA) Baldwin Lake has been described as a sag pond(Buwalda 1940) and is located on the upthrust side of the Raymond Fault a reverse thrustfault with left-lateral movement (Crook et al 1987) The lake is underlain by Quaternarygravel and alluvium eroded and transported from granodiorite quartz diorite and gneissicsource rocks containing abundant iron-rich hornblende and biotite The shore was arti ciallyplanted with Mediterranean and tropical plants and various chemicals and fertilizers usedin garden maintenance are periodically introduced to the lake ARB-1 cells appeared tobe particularly abundant in locations where the sediments were rich in large fragments ofmetallic iron and magnetic particles
The temperature during the collection period (July 1999ndashOctober 2000) varied from10plusmnC to 28plusmnC The pH of water collected from the lake was 72 to 76 (cells remained viablein bottles in the laboratory at pHs up to 83) The Eh at the surface of the sediment rangedfrom D iexcl315 mV to iexcl183 mV and oxygen from 012 to 017 mgL (38 to 53 sup1M)
Methods
Extraction of the Bacteria
Samples used for cell extraction were taken from 2-liter bottles lled with approximately50 surface sediment and 50 overlying lake water Bottles were taken back to the labora-tory shaken vigorously and magnetic stir bars placed at the watersediment interface alongthe Earthrsquos magnetic eld with the south pole of one magnet on the north side of the bottleand the north pole of the other magnet on the south side of the jar Typically cells accumu-lated as a pinkish pellet at the south-seeking side The pellet was removed with a Pasteurpipette and released into a scintillation vial containing a small amount of lter-sterilizedlake water Magnets were placed on the scintillation vial as described previously and theprocess repeated to further purify the cells
In some experiments we used a modi cation of the racetrack method (Wolfe et al1987 Schuler et al 1999) to separate the magnetic bacteria from nonmagnetic cells Inthis method a sterile cotton plug acting as a barrier for nonmagnetotactic bacteria wasinserted into the wide end of a sealed sterile Pasteur pipette The magnetotactic bacteriaswam to the sealed tip toward the south end of a bar magnet and then the tip containing thecells was broken off and the cells removed The MC typically swim much faster than othermagnetotactic and nonmagnetotactic cells and thus are the rst cells to appear at the tip ofthe pipette
390 B L Cox et al
Optical and Fluorescence Light Microscopy
Optical light microscopy was performed with fresh cells (within 24 h of collection) using aNikon Eclipse E600 microscope with phase contrast Images were obtained with a JavelinMOS solid state camera Flagella were visualized via the Gray staining method using amethod described by Murray and Robinow (1994) Loef errsquos methylene blue stain was usedto test for polyphosphate inclusions and safranin was used to visualize inclusion membranes(Murray and Robinow 1994) Viability was assessed using the Baclite bacterial viabilitykit (Molecular Probes L-7012 stain) This method employs two nucleic acid dyes bothof which are excited with blue (490 nm) light The green dye (cytonine) labels all cellsdead and live while the red dye (propidium iodate) is excluded from living cells but canpenetrate the damaged membranes of dead cells Thus living cells appear green while deadcells appear red
Transmission Electron Microscopy (TEM)
We used a Philips EM 201C microscope operating at 80 kV to examine stained thin sectionsand whole mounts of cell suspensions Cells for thin sections were xed in glutaraldehyderinsed in malate buffer post xed in osmium tetroxide stained with uranyl acetate dehy-drated with ethanol and embedded in EPON resin (Beveridge et al 1994) For negativestains whole cells were deposited on Cu grids coated with Formvar and stained with 2uranyl acetate
A Philips 430 TEM with a single crystal lanthanum hexaboride lament operatingat 200 kV was used for microscopy energy dispersive X-ray spectroscopy (EDS) andselected area electron diffraction (SAED) on fresh (collected within 24 h) unstained cellsuspensions that were deposited on carbon-coated copper grids The area sampled by theEDAX 9900 spectrum analyzer is approximately 200 nm The electron beam was defocusedat the condenser lens to maintain counting rates below 1 kHz and live time ef ciency greaterthan 95 The SAED patterns were recorded on Kodak electron image lm and digitizedfor analysis
Environmental Scanning Electron Microscopy (ESEM)
A Philips XL30 ESEM with a eld emission gun was used to observe hydrated un xed cellsImages were acquired at 20 kV in ldquowet moderdquo (3plusmnC 53 Torr chamber and water vaporpressure) at a relative humidity in the sample chamber of 93 The EDS was performedon fully hydrated uncoated and un xed specimens using an accelerating voltage of 20 kVand counting for 100 s (live time) The spot size sampled by the EDS detector was 20 nmbut because of beam spread signals from elements within a radius of 100 nm may also becounted In some cases cells were xed in glutaraldehyde prior to observation (images notshown) This had a tendency to shrink the cells to 60 of their original size For all otherexperiments fresh cells (within 24 h of collection) were used
Molecular Diversity
Bacterial small subunit (SSU) rRNA genes were PCR-ampli ed using the primers B11Fand B1512r (Giovannoni 1991) Five aliquots of cells (ca 108 each) were placed in separatetubes containing 18 sup1l of PCR cocktail (1 pound Taq DNA polymerase buffer with MgCl2 1 Uof Taq polymerase [Promega Madison WI USA] 200 sup1M dNTPs 750 nM each primer)Cells were lysed by incubation for 6 min at 94plusmnC and then subjected to PCR thermalcycling PCR conditions were 30 sec 94plusmnC denaturation 1 min 50plusmnC annealing and
Organization of Inclusions in Magnetococci 391
2 min 72plusmnC elongation for 35 cycles in an MJ Research thermal cycler (Waltham MAUSA) After a nal 10 min incubation at 72plusmnC the products were combined and puri edby gel extraction as recommended by the manufacturer (Qiagen Chatsworth CA) Am-pli cation products were cloned into the plasmid vector pCR21 as recommended by themanufacturer (Invitrogen Carlsbad CA)
SSU rDNA inserts were PCR-ampli ed under the same conditions as above Theproduct was double-digested with HhaI (New England Biolabs Beverly MA) and RsaI(Promega) restriction endonucleases in 1 pound Promega digestion buffer at 37plusmnC for 3 h Thedigested DNA was separated by electrophoresis through low melting point agarose (20wtvol IBI Shelton Scienti c Shelton CT USA) The banding patterns were grouped ac-cording to similarity and representative members of groups displaying patterns similar tothose of MC we had previously sequenced (data not shown) were partially sequenced Sixpartial sequences that were tentatively identi ed as MC and as different groups were sub-sequently fully bidirectionally sequenced with either an ABI 3700 (Applied BiosystemsFoster City CA) or a Licor 4200 (Licor Lincoln NE USA) automated DNA sequencerand dye terminator chemistry
Sequences were initially aligned to their nearest neighbor using the program ARB pro-vided by Wolfgang Ludwig and Oliver Strunk (Technische Universitat Munchen MunichGermany program available at httpwwwtumunchendeARB) The sequences were fur-ther manually aligned to sequences obtained from the GenBank database using the GeneticData Environment (GDE) v 20 sequence analysis software package provided by SteveSmith (Millipore Corporation Bedford MA) and described by Rappe et al (1997) Phy-logenetic inference and evolutionary distance calculations were performed as described inRappe et al (1997) Regions of ambiguous alignment were excluded from analyses
Results and Discussion
General Description of Cells
Cells of ARB-1 were spherical to oblate spheroidal Gram-negative cells ranging from1 to 4 sup1m (Figure 1) and averaging 25 sup1m In the ESEM the average size of cells was16 sup1m if xed with glutaraldehyde (aspect ratio 085) and 27 sup1m if not xed (aspectratio 085) The wide range of cells sizes (1ndash4 sup1m) might re ect the phylogenetic diversity(described below) or indicate that there are different phases in the cell cycle re ected incell size or that cells simply exhibit budding Cells contained two large phosphorus-richsymmetrical intracellular bodies that took up a large fraction of the cell volume Otherobserved inclusions were sulfur globules and magnetosomes Phosphorus-rich inclusionsin ESEM images (Figure 1d) appeared to be wider apart at one end forming a crevicewhere many small sulfur-containing inclusions were located On the opposite side of mostcells where the phosphorus inclusions appeared to be closer together magnetosomes werearranged as a cluster of disorganized crystals as chains or both (Figure 1d)
Several cells in the TEM (Figure 2) and in the ESEM appeared to be in the processof dividing In some cases indentations preceding cell division appeared to occur betweenthe two phosphorus inclusions in others these constrictions seemed to separate the sulfurinclusions from the rest of the cell (Figure 2) We also observed protrusions or extensionson some of the dividing cells (Figure 2c d) The large differences in cell size may well bedue to unequal division (budding) observed in some cells during division (Figure 2) Thecylindrical structure of one of these (Figure 2d) is similar to the basal body of a agellabundle Other possible explanations for these protrusions are that they are membrane vesicles(Beveridge 1999) or some artifact of electron microscopy or staining
392 B L Cox et al
FIGURE 1 Images of ARB-1 cells showing inclusions containing phosphorus (P) sul-fur (S) and magnetite (M) and variations in cell sizes a) Phase contrast light microscopyimage of live cells b) TEM image of unstained cells xed in glutaraldehyde (image taken byH Vali) c) TEM image of unstained and un xed cell showing S-rich globules P-rich in-clusions and magnetosomes (M) in chains d) ESEM image of hydrated un xed cells on amembrane lter H indicates ldquohingedrdquo P inclusions
The swimming speed of ARB-1 cells in wet mounts or in capillary racetracks wasextremely rapid up to a millimeter or 1000 body lengths per second for fresh cells whendirected by magnetic stir bars The phosphorus-rich inclusions appeared to wobble from sideto side or may even have opened and closed slightly as if hinged on one side (Figure 1d)as the cells swam
Organization of Inclusions in Magnetococci 393
FIGURE 2 Images of ARB-1 cells showing cell division a) TEM image of cells stainedwith uranyl acetate showing division across rather than between P inclusions b) ESEMimage showing division of cells between P inclusions c) d) TEM images of cells stainedwith uranyl acetate showing unequal division of cells with protrusions
Flagella and Fimbriae or Pili
Like other MC (Blakemore 1975 1982 Moench 1988 Frankel et al 1997) cells of ARB-1were bilophotrichously agellated having two agellar bundles on one side of the cell(Figure 3) Each bundle contained about 10 unsheathed agella that were 13 nm in diameterCells also appeared to have other smaller protrusions that resemble pili (Figure 3b) Thetendency for cells of ARB-1 to stick to glass slides and cover slips and to each other supportsthe presence of mbriae or pili Pili have been observed on other uncultured (Blakemore1982) and cultured (Frankel et al 1997) MC
Magnetosomes
Magnetosome crystals in ARB-1 cells occurred in chains and clusters and ranged in lengthfrom 20 to 133 nm and in width from 10 to 84 nm with an average (sect standard deviation)length and width of 82 sect 23 nm and 49 sect 15 nm respectively (n D 341) The average lengthof clustered magnetosomes (n D 260) was 83 sect 24 nearly the same as the average lengthof magnetosomes from chains (n D 81 mean D 80 sect 19) The range of lengths of magneto-somes in chains (40ndash120 nm) was narrower than that of clustered magnetosomes The size
394 B L Cox et al
FIGURE 3 Images of ARB-1 cells showing agella andor pili a) Light microscopy imageof cells stained for agella Arrows indicate possible cell division plane b) Thin section of acell stained with uranyl acetate showing spaces where P inclusions had been S inclusionsmagnetosomes (M) and pili c) TEM image of whole cells with clustered magnetosomecrystals (M) negatively stained with uranyl acetate Notice haloes around crystals whichmight represent a membrane or coating
distributions (length and width) of all 341 magnetite crystals from cells of ARB-1 are plot-ted in Figure 4 Values plotted for ARB-1 magnetite crystals showed that like in virtually allother magnetotactic bacteria the magnetite particles in ARB-1 are single-magnetic-domaincrystals (Butler and Banerjee 1975 Bazylinski and Frankel 2000) These results were verysimilar to those obtained for the uncultured coccus Bilophococcus magnetotacticus (Toweand Moench 1981) which also possessed clumps of magnetosomes at one end of the cellrather than chains The average length of ARB-1 magnetosomes was also similar to thatof the cultured MC-1 However the shape of MC-1 magnetosomes was less elongated(Meldrum et al 1993a) It appears that the size distributions and aspect ratios for this typeof MC are distinct from those of other magnetotactic bacteria (eg MV-1 and MC-1) aswell as from those in the Mars meteorite ALH84001 (Thomas-Keprta et al 2000)
Morphologically the magnetosome crystals in ARB-1 were rectangular in projectionwith an hexagonal cross-section There were irregularities in the edges and interiors of the
FIG
UR
E4
Dis
trib
utio
nof
leng
ths
and
wid
ths
of34
1m
agne
tite
crys
tals
from
clus
ters
and
chai
nsin
AR
B-1
cells
a) F
requ
ency
asa
func
tion
ofle
ngth
b)
Freq
uenc
yas
afu
nctio
nof
wid
thc
) Wid
thas
afu
nctio
nof
leng
th
395
396 B L Cox et al
FIGURE 5 TEM image of a thin section of a cell of ARB-1 stained with uranyl acetateshowing magnetosomes before a) and after b) beam damage leaving crystal ldquodoubletsrdquo b)Notice disappearance of electron dense material in b)
crystals (Figure 3) The edges were often rounded and the particles occasionally con-stricted slightly in the middle The crystals sometimes had a mottled appearance and mayhave had fringes around the edges Once during TEM analysis of thin sections (80 kV)magnetosome crystals sustained signi cant beam damage after several minutes of viewingat a magni cation of 100000 (Figure 5) The crystals appeared to have broken in halfpossibly along twin planes in the (111) direction or along ldquowaistedrdquo regions where thecrystals showed some constriction and there appeared to be a loss of material Beam dam-age has been previously observed during TEM of greigite (Fe3S4) magnetosome particlesbut not in magnetite crystals (D Bazylinski H Vali and J Kirschvink personal commu-nications January 2001) except when viewed as whole mounts at 400 kV (Taylor et al2001) Taylor et al (2001) speculated that water vapor from the cells underwent radiolysisin the electron-beam producing a plasma that rapidly melted the magnetosomes
EDS analyses with both TEM and ESEM of the magnetosome particles (Figure 6c 6f)showed the particles were iron-rich with some extraneous signals including sulfur (Figure 6)A strong sulfur signal would be expected from EDS analyses of the magnetic mineral greigitealso known to be produced by some magnetotactic bacteria (Mann et al 1990) In ARB-1however these extraneous signals are likely due to the sulfur- and phosphorus-containinginclusions also present in the cell SAED patterns obtained from the crystals (Figure 7)and lattice fringes from high resolution TEM with spacings of 048 nm elongated in (111)direction (Figure 7) were consistent with the mineral magnetite
Magnetosomes within cells of ARB-1 were found in three different arrangements1) as a polar cluster forming a sort of ldquocaprdquo usually at the agellar end of the cell 2) in partialor complete chains at the same end of the cell or 3) a combination of both arrangementsThe polar clusters appear to be disorganized in many images (eg Figures 1 2 and 3)However the attraction of these cells to a magnet indicates they have permanent magneticdipole moments implying that they are not disorganized We are currently pursuing thisquestion of whether or not the clusters are disorganized
In cells having both chains and clusters it is dif cult to determine whether chainswere forming from the clustered crystals or whether the polar clusters of particles werefrom a chain that was disrupted during dehydration of the cell during preparation for TEMexamination However polar clusters of crystals were observed in ESEM images of many
Organization of Inclusions in Magnetococci 397
FIGURE 6 EDS spectra from TEM a) b) c) and ESEM d) e) f) showing elementalcomposition of P-rich a) d) and S-rich b) e) inclusions and magnetosomes c) f) Elementalcompositions of clustered magnetosomes were identical to those of magnetosomes in chainsAll y-axes are linear except for a) and b) which are log scales Noise in f ) is due to scattering
hydrated cells (Figure 1d) suggesting that they were not caused by dehydration In manyof the cells viewed by TEM it seemed that the chains tended to occur in smaller morespherical cells while clumps of magnetosomes occurred more frequently in elongatedcells In order to see if there was any relationship between cell size and shape and the threetypes of magnetosome arrangements the size and aspect ratio for cells with these differentmagnetosome arrangements were measured (Figure 8) The cell size distribution appearedto be narrower for cells with chains than for the cells with disorganized crystals Cells withchains were predominantly smaller than 25 sup1m (Figure 8a) The aspect ratios of the cellswith chains only was smaller than that of cells with disorganized magnetosomes and wasshifted toward spherical aspect ratios (Figure 8b)
One explanation for these observations is that there are different stages of magnetosomeformation in a single species of MC The disorganized magnetosomes might form near thecell membrane between the agellar bundles and then extend into chains Alternativelymagnetosomes might rst form as chains in smaller cells At a later stage as cells increasein size the membranes linking the chains dissolve or become stressed and the magneto-somes somehow migrate to the agellar end of the cell Because magnetosomes appear tobe anchored to the inner surface of the cell membrane in virtually all species examined(Bazylinski and Frankel 2000) and are never free- oating in the cytoplasm migration ofmagnetosomes through the cytoplasm seems unlikely Instead we are presently designinghybridization experiments to address the hypothesis that the relationships between cell sizeshape and magnetosome arrangement correlate with the genotypical variations in ARB-1described later Spring et al (1994) analyzed communities of MC found in freshwater lakesin Germany using uorescent oligonucleotide probes They found different genotypes ofMC (similarity below 95) but these genotypes were not phenotypically expressed asdifferent magnetosome organizations but as preferences for different concentrations of O2
398 B L Cox et al
FIGURE 7 Evidence supporting magnetite as the magnetosome mineral in ARB-1 cellsa) Image of a single crystal of magnetite within a cell in an unstained glutaraldehyde- xedwhole cell mount taken with High Resolution TEM by H Vali b) Part of the magnetitecrystal shown in a) magni ed to show the distance between lattice fringes c) Selectedarea electron diffraction (SAED) of a group of magnetosomes in a cell from an un xedunstained whole cell mount showing the indices of some of the diffraction spots
Therefore we might expect no correlation between magnetite organization and phyloge-netic diversity of ARB-1 cells but perhaps a correlation between oxygen preference andphylogeny
A third possibility is that chains and clumps of magnetosomes form as a result ofdifferent environmental conditions Since our sampling methods were not selective as tothe cellsrsquo location in the vertical chemical (eg oxygen) and redox gradients that are sure to
Organization of Inclusions in Magnetococci 399
FIGURE 8 Relationships between organization of magnetosomes and cell size a) and cellaspect ratio b) Measurements were taken from both TEM and ESEM images Solid patternis for cells with clustered magnetosomes crossed pattern is for cells with magnetosomesin chains only Patterned bars originate at the top of solid bars
exist at the site the environmental conditions of our samples represent a relatively large rangeof oxygen and perhaps other chemical concentrations as well as redox conditions Magneticbacteria in culture are known to vary the size and number of magnetosomes in responseto the media composition (Meldrum et al 1993a) and particularly oxygen concentration(Blakemore et al 1985) Another possibility is that the clustered magnetosomes are formedby a different biological mechanism and serve a different function than magnetosomes inchains
Phosphorus (P) Inclusions
The two very large inclusion bodies seen in each cell always occurred in pairs in cellsand seemed to extend to the cell membrane and to be in contact with each other Thus themaximum dimension of the P-rich inclusions was the same as the minimum dimension ofthe cell and their combined width was approximately half of the long dimension of the cellIn ESEM images the two P-containing inclusions appeared to be attached in the middle insome cells and separated in others (Figure 1d) The P-containing inclusions appeared to beself-contained and perhaps bounded by some sort of coating or membrane in TEM images(Figure 2) and in light microscopy images of cells stained with safranin (not shown)
These inclusions were shown by both ESEM and TEM EDS to contain phosphorus with generally small amounts of potassium calcium magnesium sulfur and iron the lattertwo elements likely being due to the close proximity of the P-rich inclusions to sulfurglobules and magnetosomes (Figures 1 and 6) A very large calcium peak was observedwhen these large inclusions were analyzed in the TEM (Figure 6a) The samples shown inFigures 6a and 6d were collected at different times of the year which may explain some ofthe differences observed in these spectra Other EDS data from the ESEM (not shown) hadhigher calcium peaks than those shown in Figure 6d indicating that relative proportions ofP and Ca in P-rich inclusions vary even within the same sample
The inclusions were electron dense in TEM and had a mottled appearance due toablation by the electron beam Similar inclusions have been described for uncultured MCfrom the Itaupo Lagoon near Rio de Janeiro Brazil (Lins and Farina 1999) and from MillPond New Hampshire (Blakemore 1982) Like the structures in ARB-1 polyP granules aretranslucent in visible light (Spring et al 1994) dark (electron dense) in the TEM and oftenhave a mottled appearance (Harold 1966) The P-rich inclusions in ARB-1 cells stained
400 B L Cox et al
with methylene blue (Murray and Robinson 1994) which is suggestive of the presence ofpolyP but not con rmatory
PolyP is composed of linear chains of tetrahedral phosphate residues linked throughcommon oxygen atoms by phosphoanhydrid e bonds At physiological pH each unit carriesa monovalent negative charge giving polyP the property of having a large cation exchangecapacity The binding energy of polyP allows it to sequester Ca2C and Mg2C (Reusch 2000)consistent with the Ca2C and Mg2C signals observed in the EDS spectra of the ARB-1inclusions (Figure 6a 6d) The binding of these metals to the polyP granules contributesto their high electron density PolyP is often associated with polyhydroxybutyrat e (PHB)PHBs solvate (dissolve) cations and associate withpolyP to form Ca2C selective ion channelsacross plasma membranes (Reusch 2000)
The presence of polyP bodies is common in bacteria often resulting as a bacterialresponse to stress and they may play a role in cell motility quorum sensing and bio lmformation (Kornberg and Fraley 2000) Sulfate starvation is known to lead to rapid andmassive accumulation of polyP in some bacteria (Stanier et al 1986) When cells regainaccess to sulfate 32P uptake studies showed that PO3iexcl
4 present in the polyP bodies wasincorporated into nucleic acids PolyP is synthesized from ATP by the enzyme polyP kinasePolyP granules have been reported to be associated with nucleic acids (Harold 1966) butit is not clear whether the association occurs in vivo or after extraction This associationwith nucleic acids would be consistent with staining patterns seen when ARB-1 cells wereexamined with a uorescent livedead stain (nucleic acid stain) displaying a double-lobedstructure in all of the cells (Figure 9) This dumbbell shape appears to extend inside andacross the two phosphorus inclusions
The physiological function of the P inclusions in ARB-1 is not yet known but fromother cell systems several possibilities can be considered These inclusions might serveas a phosphorus reserve for the biosynthesis of nucleic acids and phospholipids underconditions of phosphorus starvation (Harold 1966) Still another possibility is buoyancyregulation Cells can vary their density by as much as 7 by incorporating substances intoinclusions (Guerrero et al 1985) A more speculative possibility is that ARB-1 cells canderive energy from polyP Acinetobacter johnsonii 210a is known to couple the excretion ofa metal phosphate chelate and HC generating a proton motive force (VanVeen et al 1994)A johnsonii takes up P from the aquatic environment by using two Pi transport systemsand stores it as intracellular granules of polyP
Sulfur (S) Inclusions
Cells of ARB-1 contained 5 to 10 spherical S-rich inclusions (Figures 1d) These rangedin size from approximately 100 to 700 nm although most of the S-rich inclusions in asingle cell were close to the same size and were located at one end of the cell oppositethe side where the magnetosomes were located and agellar bundles attached Like theP-rich inclusions the S-rich inclusions appeared to be enclosed in a coating or membrane(Figure 1d) EDS analysis from the ESEM showed these structures contained mostly S withsome carbon and oxygen (Figure 6e) EDS analysis from the TEM also showed S but evenhigher counts for Ca and P (Figure 6b) Beam spreading for the two methods is within aradius of 200 nm The X-rays produced by the 200-keV TEM beam sample the entire verticalextent of the cell unlike the 20 keV SEM beam which loses most of its energy Thereforethe EDS spectra collected at the TEM likely have sampled elements from the cytoplasmand cell membrane components both above and below the inclusion as well as from theP-rich inclusions nearby Other explanations are that hydrated sulfur inclusions changedtheir composition upon dehydration or that these two samples collected at different times
Organization of Inclusions in Magnetococci 401
FIGURE 9 Fluorescence micrograph of an ARB-1 cell suspension treated with Bacliteviability dyes showing that the nucleic acid staining pattern corresponds with the locationof the P-rich inclusions Baclite stains nucleic acids with cytonine (all cells) and propidiumiodate (dead or damaged cells) Bright double-lobed patterns are from ldquoliverdquo cells
of the day and the year may re ect a seasonal diurnal or cell cycle variability in the relativeproportions of the elements
Sulfur inclusions have been found in other uncultured magnetotactic bacteria includ-ing Magnetobacterium bavaricum (Spring et al 1993) Bilophococcus magnetotacticum(Moench 1988) and others (Iida and Akai 1996) and cultured species including the vibrioidstrains MV-1 and MV-2 the coccus strain MC-1 and the spirillum strain MV-4 (Bazylinskiand Frankel 2000) However only in Bilophococcus magnetotacticum was there a similarpolarization of sulfur-containing inclusions from the clustered magnetosomes within thecell (Moench 1988)
It seems likely that ARB-1 oxidizes H2S or other reduced sulfur compounds as anenergy source depositing ldquoelementalrdquo sulfur in intracellular inclusions like many othersul de-oxidizing bacteria including the anaerobic phototrophic purple sulfur bacteria suchas Chromatium vinosum and Thiocapsa roseopersicina the nonsulfur bacteria Beggiatoaspp and Thiothrix spp (Shively 1974 Friedrich 1998) and the magnetotactic bacteriamentioned before In these organisms the presence of sulfur globules is transitory andtakes place when the growth medium contains sul de andor thiosulfate When the sul deis completely utilized the sulfur is converted to sulfate (Stanier et al 1986)
Elemental S in sulfur globules is typically in a liquid state and analysis of in vivoglobules from some members of the purple and green sulfur bacteria by X-ray absorptionnear edge spectroscopy (Prange et al 1999) indicated the sulfur was in a chain structure
402 B L Cox et al
FIGURE 10 Phylogenetic position of six SSU rRNA gene clones related to previouslystudied MC This phylogenetic tree was rooted with Escherichia coli and constructed bythe neighbor joining method with the Kimura 2-parameter model for nucleotide change Amask including all nonambiguously aligned positions for 1427 nucleotides spanning nearlythe full length of the SSU rRNA gene was included Bootstrap values (100 replications)generated by the neighbor joining method are shown above relevant nodes and bootstrapvalues generated by maximum-parsimony analysis are shown below relevant nodes Onlybootstrap values above 50 are shown GenBank accession numbers are MC-CS92 X81182MC-pa119 Y13214 MC-CS81 X81184 MC-TB12 X81183 MC-MP17 X80996 MC-PA11 Y13212 MC12 L06456 partial sequence extended with data from MC-1 genomeRhizobium leguminosarum U31074 Agrobacterium tumefaciens D13294 Rhodobactersphaeroides X53854 Paracoccus denitri cans X59159 Zymomonas mobilis AF281034Sphingomonas paucimobilis D84528 Caulobacter crescentus M83799 Magnetospiril-lum magnetotacticum strain AMB1 D17514 S88429 Magnetospirillum magnetotacticumM10110 Rhodospirillum fulvum M59065 Magnetospirillum gryphiswaldense Y10109Magnetic Vibrio MV-1 LO6455 Oceanospirillum pusillum AB000482 Rhodospirillumrubrum D30778 Roseococcus thiosulfatophilus X72908 Escherichia coli J01695
Organization of Inclusions in Magnetococci 403
and was possibly complexed with water and organic residues If the S globules in ARB-1cells are analogous to those in the purple sulfur bacteria Chromatium vinosum and Thio-capsa roseopersicina they may contain a signi cant protein content Sulfur globules fromthe anaerobic phototrophs C vinosum and T roseopersicina were found to contain upto 52 protein that formed an envelope enclosing the S globules (Brune 1995) Theseproteins were isolated and analyzed and found to be glycine and tyrosine-rich structuralproteins which were homologous for the two species suggesting they are highly con-served (Brune 1995) Phylogenetically C vinosum and T roseopersicina are members ofthe deg -Proteobacteria whereas MC are members of the reg-Proteobacteria To our knowledgemagnetotactic bacteria may be the only members of the reg-Proteobacteria that make internalsulfur globules
Phylogenetic Af liation
We wished to detemine whether ARB-1 was a single species or a mixed population of MCToward that end 45 SSU rRNA genes were cloned from partially puri ed ARB-1 cells30 of which displayed similar restriction fragment length patterns (RFLP) to previouslycharacterized MC (data not shown) The nding that 15 patterns were not related to pre-viously characterized MC was not surprising given that the initial sample was not a pureculture but rather a population magnetically enriched from a sample originating from arelatively large vertical column containing both sediments and overlying water Fourteenof the MC-related clones representing different RFLP pattern groups were chosen for par-tial sequencing 10 of these sequences showed high similarity to previously characterizedMC Six of these sequences were phylogenetically distinct and were fully bidirectionallysequenced
ARB-1 MC SSU rRNA genes formed a monophyletic cluster related to but dis-tinct from other MC (Figure 10) Within the cluster the similarity between SSU rRNAgene sequences ranged from 97 (MC-H1arb and MC-g6arb) to 92 (MC-h1arb andMC-g2arb) The nearest neighbor to the ARB-1 cluster MC-CS92 was ca 89 similarand the nearest cultured relative MC-1 was ca 88 similar to the ARB-1 cluster As arule of thumb for SSU rRNA genes gt97 similarity is considered the same species andgt93ndash95 similarity is considered the same genus (Stackebrandt and Goebel 1994) Thesedata strongly suggest that ARB-1 represents a population of different diverse MC Therewas signi cant variation within the ARB-1 cluster and therefore we do not consider ARB-1to consist of bacteria of a single species or genus Correlations if any between phylogeneticand phenotypic variations in the ARB-1 cells have yet to be determined
Summary
We have partially characterized a population of magnetotactic cocci (MC) referred to asARB-1 which was particularly abundant in iron-rich sediments of a small eutrophic lake andmorphologically and structurally similar to an uncultured species of MC called Bilophococ-cus magnetotacticus The latter organism was found in large numbers in a sewage treatmentpond Cells of ARB-1 had a relatively wide size range of 1 to 4 sup1m They were motileGram-negative and bilophotrichously agellated Cells of this uncultured magnetotacticcoccus contained three types of intracellular inclusions that appeared to be in a consistentarrangement within the cell The magnetosomes contained crystals of magnetite and wereeither in chains andor polar clumps at the end of the cell where the agellar bundles werelocated Large P-rich bodies occupied the majority of the cell volume and appeared to sepa-rate the S-containing inclusions from the magnetosomes The presence of these large P-rich
404 B L Cox et al
inclusions might be an indication of cell stress or may be important in cell buoyancy or inenergy generation The presence of S-rich inclusions located at the end of the cell oppositethe magnetosomes strongly suggests that ARB-1 cells are sul de-oxidizing bacteria andthat the S might be useful as a stored electron donor when cells travel from sul de-rich tosul de-depleted zones in the water column The wide range of the cell sizes the differentmagnetosome arrangements and the large number of different retrieved 16S rRNA genesequences suggest that ARB-1 represents a population of at least several different MC 16SrRNA sequences obtained from cells of ARB-1 appeared to form a monophyletic clusterrelated to but distinct from other uncultured and cultured MC Future work using uores-cent oligonucleotide probes microscopy and physiological measurements should clarifyrelationships between phylogeny cell morphology and organization of inclusions
References
Balkwill DL Maratea D Blakemore RP 1980 Ultrastructure of a magnetotactic spirillum J Bacteriol141(3)1399ndash1408
Bazylinski DA Frankel RB 2000 Biologically controlled mineralization of magnetic iron mineralsby magnetotactic bacteria In Lovley DR editor Environmental microbe-metal interactionsWashington DC ASM Press p 109ndash143
Bazylinski DA Frankel RB Jannasch HW 1988 Anaerobic magnetite production by a marinemagnetotactic bacterium Nature 334518ndash519
Beveridge T 1999 Structures of gram-negative cell walls and their derived membrane vesiclesJ Bacteriol 181(6)4725ndash4733
Beveridge TJ Popkin TJ Cole RM 1994 Electron microscopy In Gerhardt P editor Methods forgeneral and molecular bacteriology Washington DC American Society for Microbiology p 25
Blakemore R 1975 Magnetotactic bacteria Science 190377ndash379Blakemore RP 1982 Magnetotactic bacteria Ann Rev Microbiol 36217ndash238Blakemore RP Maratea D Wolfe RS 1979 Isolation and pure culture of a freshwater magnetic
spirillum in chemically de ned medium J Bacteriol 140(2)720ndash729Blakemore RP Short KA Bazylinski DA Rosenblatt C Frankel RB 1985 Microaerobic conditions
are required for magnetite formation within Aquaspirillum magnetotacticum Geomicrobiol J4(1)53ndash71
Brune DC 1995 Isolation and characterization of sulfur globule proteins from Chromatium vinosumand Thiocapsa roseopersicina Arch Microbiol 163(1)391ndash399
Butler RF Banerjee SK 1975 Theoretical single-domain grain size range in magnetite and titano-magnetite J Geophys Res 804049ndash4058
Buwalda JP 1940 Geology of the Raymond Basin report to Pasadena Water DepartmentCrook R Allen CR Kamb B Payne CM Procter RJ 1987 Recent reverse faulting in the Transverse
Ranges California Pasadena CA US Geol Surv Prof Paper 133927ndash63Devouard B Posfai M Hua X Bazylinski DA Frankel RB Buseck PR 1998 Magnetite from
magnetotactic bacteria size distribution and twinning Am Mineral 831387ndash1398Frankel RB Bazylinski DA Johnson MS Taylor BL 1997 Magneto-Aerotaxis in marine coccoid
bacteria Biophys J 73994ndash1000Friedrich CG 1998 Physiology and genetics of sulfur-oxidizing bacteria Adv Microbial Physiol
39235ndash289Giovannoni SJ 1991 The polymerase chain reaction In Stackebrandt E Goodfellow M editors
Sequencing and hybridization techniques in bacterial systematics New York John Wiley andSons p 177ndash201
Guerrero R Pedros-Alio C Schmidt TM Mas J 1985 A survey of buoyant density of microorganismsin pure cultures and natural samples Microbiologia Sept 1(1ndash2)53ndash65
Harold FM 1966 Inorganic polyphosphates in biology structure metabolism and function BacteriolRev 30(4)772ndash794
Organization of Inclusions in Magnetococci 405
Iida A Akai J 1996 Crystalline sulfur inclusions in magnetotactic bacteria Sci Rep Nigata Univ SerE 1135ndash42
Kornberg A Fraley CD 2000 Inorganic polyphosphate a molecular fossil come to life ASM News66(5)275ndash280
Lins U Farina M 1999 Phosphorus-rich granules in uncultured magnetotactic bacteria FEMSMicrobiol Lett 172(2)23ndash28
Mann S Sparks NHC Frankel RB Bazylinski DA Jannasch HW 1990 Biomineralization of ferri-magnetic greigite (Fe3O4 and iron pyrite (FeS2 ) in a magnetotactic bacterium Nature 343258ndash
261Matsunaga T Sakaguchi T Tadokoro F 1991 Magnetite formation by a magnetic bacterium capable
of growing aerobically Appl Microbiol Biotechnol 35651ndash655Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993a Electron microscopy
study of magnetosomes in a cultured coccoid magnetotactic bacterium Proc Roy Soc Lond B251231ndash236
Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993b Electron microscopystudy of magnetosomes in two cultured vibrioid magnetotactic bacteria Proc Roy Soc Lond B251237ndash242
Moench TT 1988 Bilophococcus magnetotacticus gen nov sp nov a motile magnetic coccus Antvan Leeuwenhoek 54483ndash496
Moench TT Konetzka WA 1978 A novel method for the isolation and study of a magnetotacticbacterium Arch Microbiol 119203ndash212
Murray RGE Robinow CF 1994 Optical Microscopy In Gerhardt P editor Methods for generaland molecular bacteriology Washington DC American Society for Microbiology p 35ndash36
Prange A Arzberger I Engemann C Modrow H Schumann O Truper HG Steudel R Dahl C andHormes J 1999 In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteriaby X-ray absorption near edge spectroscopy Biochim Biophys Acta 1428446ndash454
Rappe MS Kemp PF Giovannoni SJ 1997 Phylogenetic diversity of marine coastal picoplankton16S rRNA genes cloned from the continental shelf off Cape Hatteras North Carolina LimnolOceanogr 42811ndash826
Reusch RN 2000 Transmembrane ion transport by polyphosphatepoly-(R)-3-hydroxybutyratecomplexes Biochemistry (Moscow) 65(3)335ndash352
Sakaguchi T Burgess JG Matsunaga T 1993 Magnetite formation by a sulphate-reducing bacteriumNature 36547ndash49
Schleifer KHSchuler D Spring S Weizenegger M Amann R 1991 The genus Magnetospirillum gennov description of Magnetospirillum gryphiswaldense sp nov and transfer of Aquaspirillummagnetotacticum to Magnetospirillum magnetotacticum comb nov System Appl Microbiol14379ndash385
Schuler D 1999 Formation of magnetosomes in magnetotactic bacteria J Mol Microbiol Biotechnol1(1)79ndash86
Schuler D Spring S Bazylinski DA 1999 Improved technique for the isolation of magnetotactic spir-illa from a freshwater sediment and their phylogenetic characterization System Appl Microbiol22466ndash471
Shively JM 1974 Inclusion bodies of prokaryotes Ann Rev Microbiol 28167ndash187Spring S Amann R Ludwig W Schleifer KH van Gemerden H Petersen N 1993 Dominating role
of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment ApplEnviron Microbiol 59(8)2397ndash2403
Spring S Amann R Ludwig W Schleifer KH Schuler D Poralla K Petersen N 1994 Phylogeneticanalysis of uncultured magnetotactic baceria from the Alpha-subclass of Proteobacteria SystemAppl Microbiol 17501ndash508
Stackebrandt E Goebel BM 1994 Taxonomic note a place for DNA-DNA reassociation and 16SrRNA sequence analysis in the present species de nition in bacteriology Int J System Bacteriol44846ndash849
Stanier RY Ingraham JL Wheelis ML Painter PR 1986 The microbial world 5th edition EnglewoodCliffs NJ Prentice Hall p 179
406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35

390 B L Cox et al
Optical and Fluorescence Light Microscopy
Optical light microscopy was performed with fresh cells (within 24 h of collection) using aNikon Eclipse E600 microscope with phase contrast Images were obtained with a JavelinMOS solid state camera Flagella were visualized via the Gray staining method using amethod described by Murray and Robinow (1994) Loef errsquos methylene blue stain was usedto test for polyphosphate inclusions and safranin was used to visualize inclusion membranes(Murray and Robinow 1994) Viability was assessed using the Baclite bacterial viabilitykit (Molecular Probes L-7012 stain) This method employs two nucleic acid dyes bothof which are excited with blue (490 nm) light The green dye (cytonine) labels all cellsdead and live while the red dye (propidium iodate) is excluded from living cells but canpenetrate the damaged membranes of dead cells Thus living cells appear green while deadcells appear red
Transmission Electron Microscopy (TEM)
We used a Philips EM 201C microscope operating at 80 kV to examine stained thin sectionsand whole mounts of cell suspensions Cells for thin sections were xed in glutaraldehyderinsed in malate buffer post xed in osmium tetroxide stained with uranyl acetate dehy-drated with ethanol and embedded in EPON resin (Beveridge et al 1994) For negativestains whole cells were deposited on Cu grids coated with Formvar and stained with 2uranyl acetate
A Philips 430 TEM with a single crystal lanthanum hexaboride lament operatingat 200 kV was used for microscopy energy dispersive X-ray spectroscopy (EDS) andselected area electron diffraction (SAED) on fresh (collected within 24 h) unstained cellsuspensions that were deposited on carbon-coated copper grids The area sampled by theEDAX 9900 spectrum analyzer is approximately 200 nm The electron beam was defocusedat the condenser lens to maintain counting rates below 1 kHz and live time ef ciency greaterthan 95 The SAED patterns were recorded on Kodak electron image lm and digitizedfor analysis
Environmental Scanning Electron Microscopy (ESEM)
A Philips XL30 ESEM with a eld emission gun was used to observe hydrated un xed cellsImages were acquired at 20 kV in ldquowet moderdquo (3plusmnC 53 Torr chamber and water vaporpressure) at a relative humidity in the sample chamber of 93 The EDS was performedon fully hydrated uncoated and un xed specimens using an accelerating voltage of 20 kVand counting for 100 s (live time) The spot size sampled by the EDS detector was 20 nmbut because of beam spread signals from elements within a radius of 100 nm may also becounted In some cases cells were xed in glutaraldehyde prior to observation (images notshown) This had a tendency to shrink the cells to 60 of their original size For all otherexperiments fresh cells (within 24 h of collection) were used
Molecular Diversity
Bacterial small subunit (SSU) rRNA genes were PCR-ampli ed using the primers B11Fand B1512r (Giovannoni 1991) Five aliquots of cells (ca 108 each) were placed in separatetubes containing 18 sup1l of PCR cocktail (1 pound Taq DNA polymerase buffer with MgCl2 1 Uof Taq polymerase [Promega Madison WI USA] 200 sup1M dNTPs 750 nM each primer)Cells were lysed by incubation for 6 min at 94plusmnC and then subjected to PCR thermalcycling PCR conditions were 30 sec 94plusmnC denaturation 1 min 50plusmnC annealing and
Organization of Inclusions in Magnetococci 391
2 min 72plusmnC elongation for 35 cycles in an MJ Research thermal cycler (Waltham MAUSA) After a nal 10 min incubation at 72plusmnC the products were combined and puri edby gel extraction as recommended by the manufacturer (Qiagen Chatsworth CA) Am-pli cation products were cloned into the plasmid vector pCR21 as recommended by themanufacturer (Invitrogen Carlsbad CA)
SSU rDNA inserts were PCR-ampli ed under the same conditions as above Theproduct was double-digested with HhaI (New England Biolabs Beverly MA) and RsaI(Promega) restriction endonucleases in 1 pound Promega digestion buffer at 37plusmnC for 3 h Thedigested DNA was separated by electrophoresis through low melting point agarose (20wtvol IBI Shelton Scienti c Shelton CT USA) The banding patterns were grouped ac-cording to similarity and representative members of groups displaying patterns similar tothose of MC we had previously sequenced (data not shown) were partially sequenced Sixpartial sequences that were tentatively identi ed as MC and as different groups were sub-sequently fully bidirectionally sequenced with either an ABI 3700 (Applied BiosystemsFoster City CA) or a Licor 4200 (Licor Lincoln NE USA) automated DNA sequencerand dye terminator chemistry
Sequences were initially aligned to their nearest neighbor using the program ARB pro-vided by Wolfgang Ludwig and Oliver Strunk (Technische Universitat Munchen MunichGermany program available at httpwwwtumunchendeARB) The sequences were fur-ther manually aligned to sequences obtained from the GenBank database using the GeneticData Environment (GDE) v 20 sequence analysis software package provided by SteveSmith (Millipore Corporation Bedford MA) and described by Rappe et al (1997) Phy-logenetic inference and evolutionary distance calculations were performed as described inRappe et al (1997) Regions of ambiguous alignment were excluded from analyses
Results and Discussion
General Description of Cells
Cells of ARB-1 were spherical to oblate spheroidal Gram-negative cells ranging from1 to 4 sup1m (Figure 1) and averaging 25 sup1m In the ESEM the average size of cells was16 sup1m if xed with glutaraldehyde (aspect ratio 085) and 27 sup1m if not xed (aspectratio 085) The wide range of cells sizes (1ndash4 sup1m) might re ect the phylogenetic diversity(described below) or indicate that there are different phases in the cell cycle re ected incell size or that cells simply exhibit budding Cells contained two large phosphorus-richsymmetrical intracellular bodies that took up a large fraction of the cell volume Otherobserved inclusions were sulfur globules and magnetosomes Phosphorus-rich inclusionsin ESEM images (Figure 1d) appeared to be wider apart at one end forming a crevicewhere many small sulfur-containing inclusions were located On the opposite side of mostcells where the phosphorus inclusions appeared to be closer together magnetosomes werearranged as a cluster of disorganized crystals as chains or both (Figure 1d)
Several cells in the TEM (Figure 2) and in the ESEM appeared to be in the processof dividing In some cases indentations preceding cell division appeared to occur betweenthe two phosphorus inclusions in others these constrictions seemed to separate the sulfurinclusions from the rest of the cell (Figure 2) We also observed protrusions or extensionson some of the dividing cells (Figure 2c d) The large differences in cell size may well bedue to unequal division (budding) observed in some cells during division (Figure 2) Thecylindrical structure of one of these (Figure 2d) is similar to the basal body of a agellabundle Other possible explanations for these protrusions are that they are membrane vesicles(Beveridge 1999) or some artifact of electron microscopy or staining
392 B L Cox et al
FIGURE 1 Images of ARB-1 cells showing inclusions containing phosphorus (P) sul-fur (S) and magnetite (M) and variations in cell sizes a) Phase contrast light microscopyimage of live cells b) TEM image of unstained cells xed in glutaraldehyde (image taken byH Vali) c) TEM image of unstained and un xed cell showing S-rich globules P-rich in-clusions and magnetosomes (M) in chains d) ESEM image of hydrated un xed cells on amembrane lter H indicates ldquohingedrdquo P inclusions
The swimming speed of ARB-1 cells in wet mounts or in capillary racetracks wasextremely rapid up to a millimeter or 1000 body lengths per second for fresh cells whendirected by magnetic stir bars The phosphorus-rich inclusions appeared to wobble from sideto side or may even have opened and closed slightly as if hinged on one side (Figure 1d)as the cells swam
Organization of Inclusions in Magnetococci 393
FIGURE 2 Images of ARB-1 cells showing cell division a) TEM image of cells stainedwith uranyl acetate showing division across rather than between P inclusions b) ESEMimage showing division of cells between P inclusions c) d) TEM images of cells stainedwith uranyl acetate showing unequal division of cells with protrusions
Flagella and Fimbriae or Pili
Like other MC (Blakemore 1975 1982 Moench 1988 Frankel et al 1997) cells of ARB-1were bilophotrichously agellated having two agellar bundles on one side of the cell(Figure 3) Each bundle contained about 10 unsheathed agella that were 13 nm in diameterCells also appeared to have other smaller protrusions that resemble pili (Figure 3b) Thetendency for cells of ARB-1 to stick to glass slides and cover slips and to each other supportsthe presence of mbriae or pili Pili have been observed on other uncultured (Blakemore1982) and cultured (Frankel et al 1997) MC
Magnetosomes
Magnetosome crystals in ARB-1 cells occurred in chains and clusters and ranged in lengthfrom 20 to 133 nm and in width from 10 to 84 nm with an average (sect standard deviation)length and width of 82 sect 23 nm and 49 sect 15 nm respectively (n D 341) The average lengthof clustered magnetosomes (n D 260) was 83 sect 24 nearly the same as the average lengthof magnetosomes from chains (n D 81 mean D 80 sect 19) The range of lengths of magneto-somes in chains (40ndash120 nm) was narrower than that of clustered magnetosomes The size
394 B L Cox et al
FIGURE 3 Images of ARB-1 cells showing agella andor pili a) Light microscopy imageof cells stained for agella Arrows indicate possible cell division plane b) Thin section of acell stained with uranyl acetate showing spaces where P inclusions had been S inclusionsmagnetosomes (M) and pili c) TEM image of whole cells with clustered magnetosomecrystals (M) negatively stained with uranyl acetate Notice haloes around crystals whichmight represent a membrane or coating
distributions (length and width) of all 341 magnetite crystals from cells of ARB-1 are plot-ted in Figure 4 Values plotted for ARB-1 magnetite crystals showed that like in virtually allother magnetotactic bacteria the magnetite particles in ARB-1 are single-magnetic-domaincrystals (Butler and Banerjee 1975 Bazylinski and Frankel 2000) These results were verysimilar to those obtained for the uncultured coccus Bilophococcus magnetotacticus (Toweand Moench 1981) which also possessed clumps of magnetosomes at one end of the cellrather than chains The average length of ARB-1 magnetosomes was also similar to thatof the cultured MC-1 However the shape of MC-1 magnetosomes was less elongated(Meldrum et al 1993a) It appears that the size distributions and aspect ratios for this typeof MC are distinct from those of other magnetotactic bacteria (eg MV-1 and MC-1) aswell as from those in the Mars meteorite ALH84001 (Thomas-Keprta et al 2000)
Morphologically the magnetosome crystals in ARB-1 were rectangular in projectionwith an hexagonal cross-section There were irregularities in the edges and interiors of the
FIG
UR
E4
Dis
trib
utio
nof
leng
ths
and
wid
ths
of34
1m
agne
tite
crys
tals
from
clus
ters
and
chai
nsin
AR
B-1
cells
a) F
requ
ency
asa
func
tion
ofle
ngth
b)
Freq
uenc
yas
afu
nctio
nof
wid
thc
) Wid
thas
afu
nctio
nof
leng
th
395
396 B L Cox et al
FIGURE 5 TEM image of a thin section of a cell of ARB-1 stained with uranyl acetateshowing magnetosomes before a) and after b) beam damage leaving crystal ldquodoubletsrdquo b)Notice disappearance of electron dense material in b)
crystals (Figure 3) The edges were often rounded and the particles occasionally con-stricted slightly in the middle The crystals sometimes had a mottled appearance and mayhave had fringes around the edges Once during TEM analysis of thin sections (80 kV)magnetosome crystals sustained signi cant beam damage after several minutes of viewingat a magni cation of 100000 (Figure 5) The crystals appeared to have broken in halfpossibly along twin planes in the (111) direction or along ldquowaistedrdquo regions where thecrystals showed some constriction and there appeared to be a loss of material Beam dam-age has been previously observed during TEM of greigite (Fe3S4) magnetosome particlesbut not in magnetite crystals (D Bazylinski H Vali and J Kirschvink personal commu-nications January 2001) except when viewed as whole mounts at 400 kV (Taylor et al2001) Taylor et al (2001) speculated that water vapor from the cells underwent radiolysisin the electron-beam producing a plasma that rapidly melted the magnetosomes
EDS analyses with both TEM and ESEM of the magnetosome particles (Figure 6c 6f)showed the particles were iron-rich with some extraneous signals including sulfur (Figure 6)A strong sulfur signal would be expected from EDS analyses of the magnetic mineral greigitealso known to be produced by some magnetotactic bacteria (Mann et al 1990) In ARB-1however these extraneous signals are likely due to the sulfur- and phosphorus-containinginclusions also present in the cell SAED patterns obtained from the crystals (Figure 7)and lattice fringes from high resolution TEM with spacings of 048 nm elongated in (111)direction (Figure 7) were consistent with the mineral magnetite
Magnetosomes within cells of ARB-1 were found in three different arrangements1) as a polar cluster forming a sort of ldquocaprdquo usually at the agellar end of the cell 2) in partialor complete chains at the same end of the cell or 3) a combination of both arrangementsThe polar clusters appear to be disorganized in many images (eg Figures 1 2 and 3)However the attraction of these cells to a magnet indicates they have permanent magneticdipole moments implying that they are not disorganized We are currently pursuing thisquestion of whether or not the clusters are disorganized
In cells having both chains and clusters it is dif cult to determine whether chainswere forming from the clustered crystals or whether the polar clusters of particles werefrom a chain that was disrupted during dehydration of the cell during preparation for TEMexamination However polar clusters of crystals were observed in ESEM images of many
Organization of Inclusions in Magnetococci 397
FIGURE 6 EDS spectra from TEM a) b) c) and ESEM d) e) f) showing elementalcomposition of P-rich a) d) and S-rich b) e) inclusions and magnetosomes c) f) Elementalcompositions of clustered magnetosomes were identical to those of magnetosomes in chainsAll y-axes are linear except for a) and b) which are log scales Noise in f ) is due to scattering
hydrated cells (Figure 1d) suggesting that they were not caused by dehydration In manyof the cells viewed by TEM it seemed that the chains tended to occur in smaller morespherical cells while clumps of magnetosomes occurred more frequently in elongatedcells In order to see if there was any relationship between cell size and shape and the threetypes of magnetosome arrangements the size and aspect ratio for cells with these differentmagnetosome arrangements were measured (Figure 8) The cell size distribution appearedto be narrower for cells with chains than for the cells with disorganized crystals Cells withchains were predominantly smaller than 25 sup1m (Figure 8a) The aspect ratios of the cellswith chains only was smaller than that of cells with disorganized magnetosomes and wasshifted toward spherical aspect ratios (Figure 8b)
One explanation for these observations is that there are different stages of magnetosomeformation in a single species of MC The disorganized magnetosomes might form near thecell membrane between the agellar bundles and then extend into chains Alternativelymagnetosomes might rst form as chains in smaller cells At a later stage as cells increasein size the membranes linking the chains dissolve or become stressed and the magneto-somes somehow migrate to the agellar end of the cell Because magnetosomes appear tobe anchored to the inner surface of the cell membrane in virtually all species examined(Bazylinski and Frankel 2000) and are never free- oating in the cytoplasm migration ofmagnetosomes through the cytoplasm seems unlikely Instead we are presently designinghybridization experiments to address the hypothesis that the relationships between cell sizeshape and magnetosome arrangement correlate with the genotypical variations in ARB-1described later Spring et al (1994) analyzed communities of MC found in freshwater lakesin Germany using uorescent oligonucleotide probes They found different genotypes ofMC (similarity below 95) but these genotypes were not phenotypically expressed asdifferent magnetosome organizations but as preferences for different concentrations of O2
398 B L Cox et al
FIGURE 7 Evidence supporting magnetite as the magnetosome mineral in ARB-1 cellsa) Image of a single crystal of magnetite within a cell in an unstained glutaraldehyde- xedwhole cell mount taken with High Resolution TEM by H Vali b) Part of the magnetitecrystal shown in a) magni ed to show the distance between lattice fringes c) Selectedarea electron diffraction (SAED) of a group of magnetosomes in a cell from an un xedunstained whole cell mount showing the indices of some of the diffraction spots
Therefore we might expect no correlation between magnetite organization and phyloge-netic diversity of ARB-1 cells but perhaps a correlation between oxygen preference andphylogeny
A third possibility is that chains and clumps of magnetosomes form as a result ofdifferent environmental conditions Since our sampling methods were not selective as tothe cellsrsquo location in the vertical chemical (eg oxygen) and redox gradients that are sure to
Organization of Inclusions in Magnetococci 399
FIGURE 8 Relationships between organization of magnetosomes and cell size a) and cellaspect ratio b) Measurements were taken from both TEM and ESEM images Solid patternis for cells with clustered magnetosomes crossed pattern is for cells with magnetosomesin chains only Patterned bars originate at the top of solid bars
exist at the site the environmental conditions of our samples represent a relatively large rangeof oxygen and perhaps other chemical concentrations as well as redox conditions Magneticbacteria in culture are known to vary the size and number of magnetosomes in responseto the media composition (Meldrum et al 1993a) and particularly oxygen concentration(Blakemore et al 1985) Another possibility is that the clustered magnetosomes are formedby a different biological mechanism and serve a different function than magnetosomes inchains
Phosphorus (P) Inclusions
The two very large inclusion bodies seen in each cell always occurred in pairs in cellsand seemed to extend to the cell membrane and to be in contact with each other Thus themaximum dimension of the P-rich inclusions was the same as the minimum dimension ofthe cell and their combined width was approximately half of the long dimension of the cellIn ESEM images the two P-containing inclusions appeared to be attached in the middle insome cells and separated in others (Figure 1d) The P-containing inclusions appeared to beself-contained and perhaps bounded by some sort of coating or membrane in TEM images(Figure 2) and in light microscopy images of cells stained with safranin (not shown)
These inclusions were shown by both ESEM and TEM EDS to contain phosphorus with generally small amounts of potassium calcium magnesium sulfur and iron the lattertwo elements likely being due to the close proximity of the P-rich inclusions to sulfurglobules and magnetosomes (Figures 1 and 6) A very large calcium peak was observedwhen these large inclusions were analyzed in the TEM (Figure 6a) The samples shown inFigures 6a and 6d were collected at different times of the year which may explain some ofthe differences observed in these spectra Other EDS data from the ESEM (not shown) hadhigher calcium peaks than those shown in Figure 6d indicating that relative proportions ofP and Ca in P-rich inclusions vary even within the same sample
The inclusions were electron dense in TEM and had a mottled appearance due toablation by the electron beam Similar inclusions have been described for uncultured MCfrom the Itaupo Lagoon near Rio de Janeiro Brazil (Lins and Farina 1999) and from MillPond New Hampshire (Blakemore 1982) Like the structures in ARB-1 polyP granules aretranslucent in visible light (Spring et al 1994) dark (electron dense) in the TEM and oftenhave a mottled appearance (Harold 1966) The P-rich inclusions in ARB-1 cells stained
400 B L Cox et al
with methylene blue (Murray and Robinson 1994) which is suggestive of the presence ofpolyP but not con rmatory
PolyP is composed of linear chains of tetrahedral phosphate residues linked throughcommon oxygen atoms by phosphoanhydrid e bonds At physiological pH each unit carriesa monovalent negative charge giving polyP the property of having a large cation exchangecapacity The binding energy of polyP allows it to sequester Ca2C and Mg2C (Reusch 2000)consistent with the Ca2C and Mg2C signals observed in the EDS spectra of the ARB-1inclusions (Figure 6a 6d) The binding of these metals to the polyP granules contributesto their high electron density PolyP is often associated with polyhydroxybutyrat e (PHB)PHBs solvate (dissolve) cations and associate withpolyP to form Ca2C selective ion channelsacross plasma membranes (Reusch 2000)
The presence of polyP bodies is common in bacteria often resulting as a bacterialresponse to stress and they may play a role in cell motility quorum sensing and bio lmformation (Kornberg and Fraley 2000) Sulfate starvation is known to lead to rapid andmassive accumulation of polyP in some bacteria (Stanier et al 1986) When cells regainaccess to sulfate 32P uptake studies showed that PO3iexcl
4 present in the polyP bodies wasincorporated into nucleic acids PolyP is synthesized from ATP by the enzyme polyP kinasePolyP granules have been reported to be associated with nucleic acids (Harold 1966) butit is not clear whether the association occurs in vivo or after extraction This associationwith nucleic acids would be consistent with staining patterns seen when ARB-1 cells wereexamined with a uorescent livedead stain (nucleic acid stain) displaying a double-lobedstructure in all of the cells (Figure 9) This dumbbell shape appears to extend inside andacross the two phosphorus inclusions
The physiological function of the P inclusions in ARB-1 is not yet known but fromother cell systems several possibilities can be considered These inclusions might serveas a phosphorus reserve for the biosynthesis of nucleic acids and phospholipids underconditions of phosphorus starvation (Harold 1966) Still another possibility is buoyancyregulation Cells can vary their density by as much as 7 by incorporating substances intoinclusions (Guerrero et al 1985) A more speculative possibility is that ARB-1 cells canderive energy from polyP Acinetobacter johnsonii 210a is known to couple the excretion ofa metal phosphate chelate and HC generating a proton motive force (VanVeen et al 1994)A johnsonii takes up P from the aquatic environment by using two Pi transport systemsand stores it as intracellular granules of polyP
Sulfur (S) Inclusions
Cells of ARB-1 contained 5 to 10 spherical S-rich inclusions (Figures 1d) These rangedin size from approximately 100 to 700 nm although most of the S-rich inclusions in asingle cell were close to the same size and were located at one end of the cell oppositethe side where the magnetosomes were located and agellar bundles attached Like theP-rich inclusions the S-rich inclusions appeared to be enclosed in a coating or membrane(Figure 1d) EDS analysis from the ESEM showed these structures contained mostly S withsome carbon and oxygen (Figure 6e) EDS analysis from the TEM also showed S but evenhigher counts for Ca and P (Figure 6b) Beam spreading for the two methods is within aradius of 200 nm The X-rays produced by the 200-keV TEM beam sample the entire verticalextent of the cell unlike the 20 keV SEM beam which loses most of its energy Thereforethe EDS spectra collected at the TEM likely have sampled elements from the cytoplasmand cell membrane components both above and below the inclusion as well as from theP-rich inclusions nearby Other explanations are that hydrated sulfur inclusions changedtheir composition upon dehydration or that these two samples collected at different times
Organization of Inclusions in Magnetococci 401
FIGURE 9 Fluorescence micrograph of an ARB-1 cell suspension treated with Bacliteviability dyes showing that the nucleic acid staining pattern corresponds with the locationof the P-rich inclusions Baclite stains nucleic acids with cytonine (all cells) and propidiumiodate (dead or damaged cells) Bright double-lobed patterns are from ldquoliverdquo cells
of the day and the year may re ect a seasonal diurnal or cell cycle variability in the relativeproportions of the elements
Sulfur inclusions have been found in other uncultured magnetotactic bacteria includ-ing Magnetobacterium bavaricum (Spring et al 1993) Bilophococcus magnetotacticum(Moench 1988) and others (Iida and Akai 1996) and cultured species including the vibrioidstrains MV-1 and MV-2 the coccus strain MC-1 and the spirillum strain MV-4 (Bazylinskiand Frankel 2000) However only in Bilophococcus magnetotacticum was there a similarpolarization of sulfur-containing inclusions from the clustered magnetosomes within thecell (Moench 1988)
It seems likely that ARB-1 oxidizes H2S or other reduced sulfur compounds as anenergy source depositing ldquoelementalrdquo sulfur in intracellular inclusions like many othersul de-oxidizing bacteria including the anaerobic phototrophic purple sulfur bacteria suchas Chromatium vinosum and Thiocapsa roseopersicina the nonsulfur bacteria Beggiatoaspp and Thiothrix spp (Shively 1974 Friedrich 1998) and the magnetotactic bacteriamentioned before In these organisms the presence of sulfur globules is transitory andtakes place when the growth medium contains sul de andor thiosulfate When the sul deis completely utilized the sulfur is converted to sulfate (Stanier et al 1986)
Elemental S in sulfur globules is typically in a liquid state and analysis of in vivoglobules from some members of the purple and green sulfur bacteria by X-ray absorptionnear edge spectroscopy (Prange et al 1999) indicated the sulfur was in a chain structure
402 B L Cox et al
FIGURE 10 Phylogenetic position of six SSU rRNA gene clones related to previouslystudied MC This phylogenetic tree was rooted with Escherichia coli and constructed bythe neighbor joining method with the Kimura 2-parameter model for nucleotide change Amask including all nonambiguously aligned positions for 1427 nucleotides spanning nearlythe full length of the SSU rRNA gene was included Bootstrap values (100 replications)generated by the neighbor joining method are shown above relevant nodes and bootstrapvalues generated by maximum-parsimony analysis are shown below relevant nodes Onlybootstrap values above 50 are shown GenBank accession numbers are MC-CS92 X81182MC-pa119 Y13214 MC-CS81 X81184 MC-TB12 X81183 MC-MP17 X80996 MC-PA11 Y13212 MC12 L06456 partial sequence extended with data from MC-1 genomeRhizobium leguminosarum U31074 Agrobacterium tumefaciens D13294 Rhodobactersphaeroides X53854 Paracoccus denitri cans X59159 Zymomonas mobilis AF281034Sphingomonas paucimobilis D84528 Caulobacter crescentus M83799 Magnetospiril-lum magnetotacticum strain AMB1 D17514 S88429 Magnetospirillum magnetotacticumM10110 Rhodospirillum fulvum M59065 Magnetospirillum gryphiswaldense Y10109Magnetic Vibrio MV-1 LO6455 Oceanospirillum pusillum AB000482 Rhodospirillumrubrum D30778 Roseococcus thiosulfatophilus X72908 Escherichia coli J01695
Organization of Inclusions in Magnetococci 403
and was possibly complexed with water and organic residues If the S globules in ARB-1cells are analogous to those in the purple sulfur bacteria Chromatium vinosum and Thio-capsa roseopersicina they may contain a signi cant protein content Sulfur globules fromthe anaerobic phototrophs C vinosum and T roseopersicina were found to contain upto 52 protein that formed an envelope enclosing the S globules (Brune 1995) Theseproteins were isolated and analyzed and found to be glycine and tyrosine-rich structuralproteins which were homologous for the two species suggesting they are highly con-served (Brune 1995) Phylogenetically C vinosum and T roseopersicina are members ofthe deg -Proteobacteria whereas MC are members of the reg-Proteobacteria To our knowledgemagnetotactic bacteria may be the only members of the reg-Proteobacteria that make internalsulfur globules
Phylogenetic Af liation
We wished to detemine whether ARB-1 was a single species or a mixed population of MCToward that end 45 SSU rRNA genes were cloned from partially puri ed ARB-1 cells30 of which displayed similar restriction fragment length patterns (RFLP) to previouslycharacterized MC (data not shown) The nding that 15 patterns were not related to pre-viously characterized MC was not surprising given that the initial sample was not a pureculture but rather a population magnetically enriched from a sample originating from arelatively large vertical column containing both sediments and overlying water Fourteenof the MC-related clones representing different RFLP pattern groups were chosen for par-tial sequencing 10 of these sequences showed high similarity to previously characterizedMC Six of these sequences were phylogenetically distinct and were fully bidirectionallysequenced
ARB-1 MC SSU rRNA genes formed a monophyletic cluster related to but dis-tinct from other MC (Figure 10) Within the cluster the similarity between SSU rRNAgene sequences ranged from 97 (MC-H1arb and MC-g6arb) to 92 (MC-h1arb andMC-g2arb) The nearest neighbor to the ARB-1 cluster MC-CS92 was ca 89 similarand the nearest cultured relative MC-1 was ca 88 similar to the ARB-1 cluster As arule of thumb for SSU rRNA genes gt97 similarity is considered the same species andgt93ndash95 similarity is considered the same genus (Stackebrandt and Goebel 1994) Thesedata strongly suggest that ARB-1 represents a population of different diverse MC Therewas signi cant variation within the ARB-1 cluster and therefore we do not consider ARB-1to consist of bacteria of a single species or genus Correlations if any between phylogeneticand phenotypic variations in the ARB-1 cells have yet to be determined
Summary
We have partially characterized a population of magnetotactic cocci (MC) referred to asARB-1 which was particularly abundant in iron-rich sediments of a small eutrophic lake andmorphologically and structurally similar to an uncultured species of MC called Bilophococ-cus magnetotacticus The latter organism was found in large numbers in a sewage treatmentpond Cells of ARB-1 had a relatively wide size range of 1 to 4 sup1m They were motileGram-negative and bilophotrichously agellated Cells of this uncultured magnetotacticcoccus contained three types of intracellular inclusions that appeared to be in a consistentarrangement within the cell The magnetosomes contained crystals of magnetite and wereeither in chains andor polar clumps at the end of the cell where the agellar bundles werelocated Large P-rich bodies occupied the majority of the cell volume and appeared to sepa-rate the S-containing inclusions from the magnetosomes The presence of these large P-rich
404 B L Cox et al
inclusions might be an indication of cell stress or may be important in cell buoyancy or inenergy generation The presence of S-rich inclusions located at the end of the cell oppositethe magnetosomes strongly suggests that ARB-1 cells are sul de-oxidizing bacteria andthat the S might be useful as a stored electron donor when cells travel from sul de-rich tosul de-depleted zones in the water column The wide range of the cell sizes the differentmagnetosome arrangements and the large number of different retrieved 16S rRNA genesequences suggest that ARB-1 represents a population of at least several different MC 16SrRNA sequences obtained from cells of ARB-1 appeared to form a monophyletic clusterrelated to but distinct from other uncultured and cultured MC Future work using uores-cent oligonucleotide probes microscopy and physiological measurements should clarifyrelationships between phylogeny cell morphology and organization of inclusions
References
Balkwill DL Maratea D Blakemore RP 1980 Ultrastructure of a magnetotactic spirillum J Bacteriol141(3)1399ndash1408
Bazylinski DA Frankel RB 2000 Biologically controlled mineralization of magnetic iron mineralsby magnetotactic bacteria In Lovley DR editor Environmental microbe-metal interactionsWashington DC ASM Press p 109ndash143
Bazylinski DA Frankel RB Jannasch HW 1988 Anaerobic magnetite production by a marinemagnetotactic bacterium Nature 334518ndash519
Beveridge T 1999 Structures of gram-negative cell walls and their derived membrane vesiclesJ Bacteriol 181(6)4725ndash4733
Beveridge TJ Popkin TJ Cole RM 1994 Electron microscopy In Gerhardt P editor Methods forgeneral and molecular bacteriology Washington DC American Society for Microbiology p 25
Blakemore R 1975 Magnetotactic bacteria Science 190377ndash379Blakemore RP 1982 Magnetotactic bacteria Ann Rev Microbiol 36217ndash238Blakemore RP Maratea D Wolfe RS 1979 Isolation and pure culture of a freshwater magnetic
spirillum in chemically de ned medium J Bacteriol 140(2)720ndash729Blakemore RP Short KA Bazylinski DA Rosenblatt C Frankel RB 1985 Microaerobic conditions
are required for magnetite formation within Aquaspirillum magnetotacticum Geomicrobiol J4(1)53ndash71
Brune DC 1995 Isolation and characterization of sulfur globule proteins from Chromatium vinosumand Thiocapsa roseopersicina Arch Microbiol 163(1)391ndash399
Butler RF Banerjee SK 1975 Theoretical single-domain grain size range in magnetite and titano-magnetite J Geophys Res 804049ndash4058
Buwalda JP 1940 Geology of the Raymond Basin report to Pasadena Water DepartmentCrook R Allen CR Kamb B Payne CM Procter RJ 1987 Recent reverse faulting in the Transverse
Ranges California Pasadena CA US Geol Surv Prof Paper 133927ndash63Devouard B Posfai M Hua X Bazylinski DA Frankel RB Buseck PR 1998 Magnetite from
magnetotactic bacteria size distribution and twinning Am Mineral 831387ndash1398Frankel RB Bazylinski DA Johnson MS Taylor BL 1997 Magneto-Aerotaxis in marine coccoid
bacteria Biophys J 73994ndash1000Friedrich CG 1998 Physiology and genetics of sulfur-oxidizing bacteria Adv Microbial Physiol
39235ndash289Giovannoni SJ 1991 The polymerase chain reaction In Stackebrandt E Goodfellow M editors
Sequencing and hybridization techniques in bacterial systematics New York John Wiley andSons p 177ndash201
Guerrero R Pedros-Alio C Schmidt TM Mas J 1985 A survey of buoyant density of microorganismsin pure cultures and natural samples Microbiologia Sept 1(1ndash2)53ndash65
Harold FM 1966 Inorganic polyphosphates in biology structure metabolism and function BacteriolRev 30(4)772ndash794
Organization of Inclusions in Magnetococci 405
Iida A Akai J 1996 Crystalline sulfur inclusions in magnetotactic bacteria Sci Rep Nigata Univ SerE 1135ndash42
Kornberg A Fraley CD 2000 Inorganic polyphosphate a molecular fossil come to life ASM News66(5)275ndash280
Lins U Farina M 1999 Phosphorus-rich granules in uncultured magnetotactic bacteria FEMSMicrobiol Lett 172(2)23ndash28
Mann S Sparks NHC Frankel RB Bazylinski DA Jannasch HW 1990 Biomineralization of ferri-magnetic greigite (Fe3O4 and iron pyrite (FeS2 ) in a magnetotactic bacterium Nature 343258ndash
261Matsunaga T Sakaguchi T Tadokoro F 1991 Magnetite formation by a magnetic bacterium capable
of growing aerobically Appl Microbiol Biotechnol 35651ndash655Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993a Electron microscopy
study of magnetosomes in a cultured coccoid magnetotactic bacterium Proc Roy Soc Lond B251231ndash236
Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993b Electron microscopystudy of magnetosomes in two cultured vibrioid magnetotactic bacteria Proc Roy Soc Lond B251237ndash242
Moench TT 1988 Bilophococcus magnetotacticus gen nov sp nov a motile magnetic coccus Antvan Leeuwenhoek 54483ndash496
Moench TT Konetzka WA 1978 A novel method for the isolation and study of a magnetotacticbacterium Arch Microbiol 119203ndash212
Murray RGE Robinow CF 1994 Optical Microscopy In Gerhardt P editor Methods for generaland molecular bacteriology Washington DC American Society for Microbiology p 35ndash36
Prange A Arzberger I Engemann C Modrow H Schumann O Truper HG Steudel R Dahl C andHormes J 1999 In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteriaby X-ray absorption near edge spectroscopy Biochim Biophys Acta 1428446ndash454
Rappe MS Kemp PF Giovannoni SJ 1997 Phylogenetic diversity of marine coastal picoplankton16S rRNA genes cloned from the continental shelf off Cape Hatteras North Carolina LimnolOceanogr 42811ndash826
Reusch RN 2000 Transmembrane ion transport by polyphosphatepoly-(R)-3-hydroxybutyratecomplexes Biochemistry (Moscow) 65(3)335ndash352
Sakaguchi T Burgess JG Matsunaga T 1993 Magnetite formation by a sulphate-reducing bacteriumNature 36547ndash49
Schleifer KHSchuler D Spring S Weizenegger M Amann R 1991 The genus Magnetospirillum gennov description of Magnetospirillum gryphiswaldense sp nov and transfer of Aquaspirillummagnetotacticum to Magnetospirillum magnetotacticum comb nov System Appl Microbiol14379ndash385
Schuler D 1999 Formation of magnetosomes in magnetotactic bacteria J Mol Microbiol Biotechnol1(1)79ndash86
Schuler D Spring S Bazylinski DA 1999 Improved technique for the isolation of magnetotactic spir-illa from a freshwater sediment and their phylogenetic characterization System Appl Microbiol22466ndash471
Shively JM 1974 Inclusion bodies of prokaryotes Ann Rev Microbiol 28167ndash187Spring S Amann R Ludwig W Schleifer KH van Gemerden H Petersen N 1993 Dominating role
of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment ApplEnviron Microbiol 59(8)2397ndash2403
Spring S Amann R Ludwig W Schleifer KH Schuler D Poralla K Petersen N 1994 Phylogeneticanalysis of uncultured magnetotactic baceria from the Alpha-subclass of Proteobacteria SystemAppl Microbiol 17501ndash508
Stackebrandt E Goebel BM 1994 Taxonomic note a place for DNA-DNA reassociation and 16SrRNA sequence analysis in the present species de nition in bacteriology Int J System Bacteriol44846ndash849
Stanier RY Ingraham JL Wheelis ML Painter PR 1986 The microbial world 5th edition EnglewoodCliffs NJ Prentice Hall p 179
406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35

Organization of Inclusions in Magnetococci 391
2 min 72plusmnC elongation for 35 cycles in an MJ Research thermal cycler (Waltham MAUSA) After a nal 10 min incubation at 72plusmnC the products were combined and puri edby gel extraction as recommended by the manufacturer (Qiagen Chatsworth CA) Am-pli cation products were cloned into the plasmid vector pCR21 as recommended by themanufacturer (Invitrogen Carlsbad CA)
SSU rDNA inserts were PCR-ampli ed under the same conditions as above Theproduct was double-digested with HhaI (New England Biolabs Beverly MA) and RsaI(Promega) restriction endonucleases in 1 pound Promega digestion buffer at 37plusmnC for 3 h Thedigested DNA was separated by electrophoresis through low melting point agarose (20wtvol IBI Shelton Scienti c Shelton CT USA) The banding patterns were grouped ac-cording to similarity and representative members of groups displaying patterns similar tothose of MC we had previously sequenced (data not shown) were partially sequenced Sixpartial sequences that were tentatively identi ed as MC and as different groups were sub-sequently fully bidirectionally sequenced with either an ABI 3700 (Applied BiosystemsFoster City CA) or a Licor 4200 (Licor Lincoln NE USA) automated DNA sequencerand dye terminator chemistry
Sequences were initially aligned to their nearest neighbor using the program ARB pro-vided by Wolfgang Ludwig and Oliver Strunk (Technische Universitat Munchen MunichGermany program available at httpwwwtumunchendeARB) The sequences were fur-ther manually aligned to sequences obtained from the GenBank database using the GeneticData Environment (GDE) v 20 sequence analysis software package provided by SteveSmith (Millipore Corporation Bedford MA) and described by Rappe et al (1997) Phy-logenetic inference and evolutionary distance calculations were performed as described inRappe et al (1997) Regions of ambiguous alignment were excluded from analyses
Results and Discussion
General Description of Cells
Cells of ARB-1 were spherical to oblate spheroidal Gram-negative cells ranging from1 to 4 sup1m (Figure 1) and averaging 25 sup1m In the ESEM the average size of cells was16 sup1m if xed with glutaraldehyde (aspect ratio 085) and 27 sup1m if not xed (aspectratio 085) The wide range of cells sizes (1ndash4 sup1m) might re ect the phylogenetic diversity(described below) or indicate that there are different phases in the cell cycle re ected incell size or that cells simply exhibit budding Cells contained two large phosphorus-richsymmetrical intracellular bodies that took up a large fraction of the cell volume Otherobserved inclusions were sulfur globules and magnetosomes Phosphorus-rich inclusionsin ESEM images (Figure 1d) appeared to be wider apart at one end forming a crevicewhere many small sulfur-containing inclusions were located On the opposite side of mostcells where the phosphorus inclusions appeared to be closer together magnetosomes werearranged as a cluster of disorganized crystals as chains or both (Figure 1d)
Several cells in the TEM (Figure 2) and in the ESEM appeared to be in the processof dividing In some cases indentations preceding cell division appeared to occur betweenthe two phosphorus inclusions in others these constrictions seemed to separate the sulfurinclusions from the rest of the cell (Figure 2) We also observed protrusions or extensionson some of the dividing cells (Figure 2c d) The large differences in cell size may well bedue to unequal division (budding) observed in some cells during division (Figure 2) Thecylindrical structure of one of these (Figure 2d) is similar to the basal body of a agellabundle Other possible explanations for these protrusions are that they are membrane vesicles(Beveridge 1999) or some artifact of electron microscopy or staining
392 B L Cox et al
FIGURE 1 Images of ARB-1 cells showing inclusions containing phosphorus (P) sul-fur (S) and magnetite (M) and variations in cell sizes a) Phase contrast light microscopyimage of live cells b) TEM image of unstained cells xed in glutaraldehyde (image taken byH Vali) c) TEM image of unstained and un xed cell showing S-rich globules P-rich in-clusions and magnetosomes (M) in chains d) ESEM image of hydrated un xed cells on amembrane lter H indicates ldquohingedrdquo P inclusions
The swimming speed of ARB-1 cells in wet mounts or in capillary racetracks wasextremely rapid up to a millimeter or 1000 body lengths per second for fresh cells whendirected by magnetic stir bars The phosphorus-rich inclusions appeared to wobble from sideto side or may even have opened and closed slightly as if hinged on one side (Figure 1d)as the cells swam
Organization of Inclusions in Magnetococci 393
FIGURE 2 Images of ARB-1 cells showing cell division a) TEM image of cells stainedwith uranyl acetate showing division across rather than between P inclusions b) ESEMimage showing division of cells between P inclusions c) d) TEM images of cells stainedwith uranyl acetate showing unequal division of cells with protrusions
Flagella and Fimbriae or Pili
Like other MC (Blakemore 1975 1982 Moench 1988 Frankel et al 1997) cells of ARB-1were bilophotrichously agellated having two agellar bundles on one side of the cell(Figure 3) Each bundle contained about 10 unsheathed agella that were 13 nm in diameterCells also appeared to have other smaller protrusions that resemble pili (Figure 3b) Thetendency for cells of ARB-1 to stick to glass slides and cover slips and to each other supportsthe presence of mbriae or pili Pili have been observed on other uncultured (Blakemore1982) and cultured (Frankel et al 1997) MC
Magnetosomes
Magnetosome crystals in ARB-1 cells occurred in chains and clusters and ranged in lengthfrom 20 to 133 nm and in width from 10 to 84 nm with an average (sect standard deviation)length and width of 82 sect 23 nm and 49 sect 15 nm respectively (n D 341) The average lengthof clustered magnetosomes (n D 260) was 83 sect 24 nearly the same as the average lengthof magnetosomes from chains (n D 81 mean D 80 sect 19) The range of lengths of magneto-somes in chains (40ndash120 nm) was narrower than that of clustered magnetosomes The size
394 B L Cox et al
FIGURE 3 Images of ARB-1 cells showing agella andor pili a) Light microscopy imageof cells stained for agella Arrows indicate possible cell division plane b) Thin section of acell stained with uranyl acetate showing spaces where P inclusions had been S inclusionsmagnetosomes (M) and pili c) TEM image of whole cells with clustered magnetosomecrystals (M) negatively stained with uranyl acetate Notice haloes around crystals whichmight represent a membrane or coating
distributions (length and width) of all 341 magnetite crystals from cells of ARB-1 are plot-ted in Figure 4 Values plotted for ARB-1 magnetite crystals showed that like in virtually allother magnetotactic bacteria the magnetite particles in ARB-1 are single-magnetic-domaincrystals (Butler and Banerjee 1975 Bazylinski and Frankel 2000) These results were verysimilar to those obtained for the uncultured coccus Bilophococcus magnetotacticus (Toweand Moench 1981) which also possessed clumps of magnetosomes at one end of the cellrather than chains The average length of ARB-1 magnetosomes was also similar to thatof the cultured MC-1 However the shape of MC-1 magnetosomes was less elongated(Meldrum et al 1993a) It appears that the size distributions and aspect ratios for this typeof MC are distinct from those of other magnetotactic bacteria (eg MV-1 and MC-1) aswell as from those in the Mars meteorite ALH84001 (Thomas-Keprta et al 2000)
Morphologically the magnetosome crystals in ARB-1 were rectangular in projectionwith an hexagonal cross-section There were irregularities in the edges and interiors of the
FIG
UR
E4
Dis
trib
utio
nof
leng
ths
and
wid
ths
of34
1m
agne
tite
crys
tals
from
clus
ters
and
chai
nsin
AR
B-1
cells
a) F
requ
ency
asa
func
tion
ofle
ngth
b)
Freq
uenc
yas
afu
nctio
nof
wid
thc
) Wid
thas
afu
nctio
nof
leng
th
395
396 B L Cox et al
FIGURE 5 TEM image of a thin section of a cell of ARB-1 stained with uranyl acetateshowing magnetosomes before a) and after b) beam damage leaving crystal ldquodoubletsrdquo b)Notice disappearance of electron dense material in b)
crystals (Figure 3) The edges were often rounded and the particles occasionally con-stricted slightly in the middle The crystals sometimes had a mottled appearance and mayhave had fringes around the edges Once during TEM analysis of thin sections (80 kV)magnetosome crystals sustained signi cant beam damage after several minutes of viewingat a magni cation of 100000 (Figure 5) The crystals appeared to have broken in halfpossibly along twin planes in the (111) direction or along ldquowaistedrdquo regions where thecrystals showed some constriction and there appeared to be a loss of material Beam dam-age has been previously observed during TEM of greigite (Fe3S4) magnetosome particlesbut not in magnetite crystals (D Bazylinski H Vali and J Kirschvink personal commu-nications January 2001) except when viewed as whole mounts at 400 kV (Taylor et al2001) Taylor et al (2001) speculated that water vapor from the cells underwent radiolysisin the electron-beam producing a plasma that rapidly melted the magnetosomes
EDS analyses with both TEM and ESEM of the magnetosome particles (Figure 6c 6f)showed the particles were iron-rich with some extraneous signals including sulfur (Figure 6)A strong sulfur signal would be expected from EDS analyses of the magnetic mineral greigitealso known to be produced by some magnetotactic bacteria (Mann et al 1990) In ARB-1however these extraneous signals are likely due to the sulfur- and phosphorus-containinginclusions also present in the cell SAED patterns obtained from the crystals (Figure 7)and lattice fringes from high resolution TEM with spacings of 048 nm elongated in (111)direction (Figure 7) were consistent with the mineral magnetite
Magnetosomes within cells of ARB-1 were found in three different arrangements1) as a polar cluster forming a sort of ldquocaprdquo usually at the agellar end of the cell 2) in partialor complete chains at the same end of the cell or 3) a combination of both arrangementsThe polar clusters appear to be disorganized in many images (eg Figures 1 2 and 3)However the attraction of these cells to a magnet indicates they have permanent magneticdipole moments implying that they are not disorganized We are currently pursuing thisquestion of whether or not the clusters are disorganized
In cells having both chains and clusters it is dif cult to determine whether chainswere forming from the clustered crystals or whether the polar clusters of particles werefrom a chain that was disrupted during dehydration of the cell during preparation for TEMexamination However polar clusters of crystals were observed in ESEM images of many
Organization of Inclusions in Magnetococci 397
FIGURE 6 EDS spectra from TEM a) b) c) and ESEM d) e) f) showing elementalcomposition of P-rich a) d) and S-rich b) e) inclusions and magnetosomes c) f) Elementalcompositions of clustered magnetosomes were identical to those of magnetosomes in chainsAll y-axes are linear except for a) and b) which are log scales Noise in f ) is due to scattering
hydrated cells (Figure 1d) suggesting that they were not caused by dehydration In manyof the cells viewed by TEM it seemed that the chains tended to occur in smaller morespherical cells while clumps of magnetosomes occurred more frequently in elongatedcells In order to see if there was any relationship between cell size and shape and the threetypes of magnetosome arrangements the size and aspect ratio for cells with these differentmagnetosome arrangements were measured (Figure 8) The cell size distribution appearedto be narrower for cells with chains than for the cells with disorganized crystals Cells withchains were predominantly smaller than 25 sup1m (Figure 8a) The aspect ratios of the cellswith chains only was smaller than that of cells with disorganized magnetosomes and wasshifted toward spherical aspect ratios (Figure 8b)
One explanation for these observations is that there are different stages of magnetosomeformation in a single species of MC The disorganized magnetosomes might form near thecell membrane between the agellar bundles and then extend into chains Alternativelymagnetosomes might rst form as chains in smaller cells At a later stage as cells increasein size the membranes linking the chains dissolve or become stressed and the magneto-somes somehow migrate to the agellar end of the cell Because magnetosomes appear tobe anchored to the inner surface of the cell membrane in virtually all species examined(Bazylinski and Frankel 2000) and are never free- oating in the cytoplasm migration ofmagnetosomes through the cytoplasm seems unlikely Instead we are presently designinghybridization experiments to address the hypothesis that the relationships between cell sizeshape and magnetosome arrangement correlate with the genotypical variations in ARB-1described later Spring et al (1994) analyzed communities of MC found in freshwater lakesin Germany using uorescent oligonucleotide probes They found different genotypes ofMC (similarity below 95) but these genotypes were not phenotypically expressed asdifferent magnetosome organizations but as preferences for different concentrations of O2
398 B L Cox et al
FIGURE 7 Evidence supporting magnetite as the magnetosome mineral in ARB-1 cellsa) Image of a single crystal of magnetite within a cell in an unstained glutaraldehyde- xedwhole cell mount taken with High Resolution TEM by H Vali b) Part of the magnetitecrystal shown in a) magni ed to show the distance between lattice fringes c) Selectedarea electron diffraction (SAED) of a group of magnetosomes in a cell from an un xedunstained whole cell mount showing the indices of some of the diffraction spots
Therefore we might expect no correlation between magnetite organization and phyloge-netic diversity of ARB-1 cells but perhaps a correlation between oxygen preference andphylogeny
A third possibility is that chains and clumps of magnetosomes form as a result ofdifferent environmental conditions Since our sampling methods were not selective as tothe cellsrsquo location in the vertical chemical (eg oxygen) and redox gradients that are sure to
Organization of Inclusions in Magnetococci 399
FIGURE 8 Relationships between organization of magnetosomes and cell size a) and cellaspect ratio b) Measurements were taken from both TEM and ESEM images Solid patternis for cells with clustered magnetosomes crossed pattern is for cells with magnetosomesin chains only Patterned bars originate at the top of solid bars
exist at the site the environmental conditions of our samples represent a relatively large rangeof oxygen and perhaps other chemical concentrations as well as redox conditions Magneticbacteria in culture are known to vary the size and number of magnetosomes in responseto the media composition (Meldrum et al 1993a) and particularly oxygen concentration(Blakemore et al 1985) Another possibility is that the clustered magnetosomes are formedby a different biological mechanism and serve a different function than magnetosomes inchains
Phosphorus (P) Inclusions
The two very large inclusion bodies seen in each cell always occurred in pairs in cellsand seemed to extend to the cell membrane and to be in contact with each other Thus themaximum dimension of the P-rich inclusions was the same as the minimum dimension ofthe cell and their combined width was approximately half of the long dimension of the cellIn ESEM images the two P-containing inclusions appeared to be attached in the middle insome cells and separated in others (Figure 1d) The P-containing inclusions appeared to beself-contained and perhaps bounded by some sort of coating or membrane in TEM images(Figure 2) and in light microscopy images of cells stained with safranin (not shown)
These inclusions were shown by both ESEM and TEM EDS to contain phosphorus with generally small amounts of potassium calcium magnesium sulfur and iron the lattertwo elements likely being due to the close proximity of the P-rich inclusions to sulfurglobules and magnetosomes (Figures 1 and 6) A very large calcium peak was observedwhen these large inclusions were analyzed in the TEM (Figure 6a) The samples shown inFigures 6a and 6d were collected at different times of the year which may explain some ofthe differences observed in these spectra Other EDS data from the ESEM (not shown) hadhigher calcium peaks than those shown in Figure 6d indicating that relative proportions ofP and Ca in P-rich inclusions vary even within the same sample
The inclusions were electron dense in TEM and had a mottled appearance due toablation by the electron beam Similar inclusions have been described for uncultured MCfrom the Itaupo Lagoon near Rio de Janeiro Brazil (Lins and Farina 1999) and from MillPond New Hampshire (Blakemore 1982) Like the structures in ARB-1 polyP granules aretranslucent in visible light (Spring et al 1994) dark (electron dense) in the TEM and oftenhave a mottled appearance (Harold 1966) The P-rich inclusions in ARB-1 cells stained
400 B L Cox et al
with methylene blue (Murray and Robinson 1994) which is suggestive of the presence ofpolyP but not con rmatory
PolyP is composed of linear chains of tetrahedral phosphate residues linked throughcommon oxygen atoms by phosphoanhydrid e bonds At physiological pH each unit carriesa monovalent negative charge giving polyP the property of having a large cation exchangecapacity The binding energy of polyP allows it to sequester Ca2C and Mg2C (Reusch 2000)consistent with the Ca2C and Mg2C signals observed in the EDS spectra of the ARB-1inclusions (Figure 6a 6d) The binding of these metals to the polyP granules contributesto their high electron density PolyP is often associated with polyhydroxybutyrat e (PHB)PHBs solvate (dissolve) cations and associate withpolyP to form Ca2C selective ion channelsacross plasma membranes (Reusch 2000)
The presence of polyP bodies is common in bacteria often resulting as a bacterialresponse to stress and they may play a role in cell motility quorum sensing and bio lmformation (Kornberg and Fraley 2000) Sulfate starvation is known to lead to rapid andmassive accumulation of polyP in some bacteria (Stanier et al 1986) When cells regainaccess to sulfate 32P uptake studies showed that PO3iexcl
4 present in the polyP bodies wasincorporated into nucleic acids PolyP is synthesized from ATP by the enzyme polyP kinasePolyP granules have been reported to be associated with nucleic acids (Harold 1966) butit is not clear whether the association occurs in vivo or after extraction This associationwith nucleic acids would be consistent with staining patterns seen when ARB-1 cells wereexamined with a uorescent livedead stain (nucleic acid stain) displaying a double-lobedstructure in all of the cells (Figure 9) This dumbbell shape appears to extend inside andacross the two phosphorus inclusions
The physiological function of the P inclusions in ARB-1 is not yet known but fromother cell systems several possibilities can be considered These inclusions might serveas a phosphorus reserve for the biosynthesis of nucleic acids and phospholipids underconditions of phosphorus starvation (Harold 1966) Still another possibility is buoyancyregulation Cells can vary their density by as much as 7 by incorporating substances intoinclusions (Guerrero et al 1985) A more speculative possibility is that ARB-1 cells canderive energy from polyP Acinetobacter johnsonii 210a is known to couple the excretion ofa metal phosphate chelate and HC generating a proton motive force (VanVeen et al 1994)A johnsonii takes up P from the aquatic environment by using two Pi transport systemsand stores it as intracellular granules of polyP
Sulfur (S) Inclusions
Cells of ARB-1 contained 5 to 10 spherical S-rich inclusions (Figures 1d) These rangedin size from approximately 100 to 700 nm although most of the S-rich inclusions in asingle cell were close to the same size and were located at one end of the cell oppositethe side where the magnetosomes were located and agellar bundles attached Like theP-rich inclusions the S-rich inclusions appeared to be enclosed in a coating or membrane(Figure 1d) EDS analysis from the ESEM showed these structures contained mostly S withsome carbon and oxygen (Figure 6e) EDS analysis from the TEM also showed S but evenhigher counts for Ca and P (Figure 6b) Beam spreading for the two methods is within aradius of 200 nm The X-rays produced by the 200-keV TEM beam sample the entire verticalextent of the cell unlike the 20 keV SEM beam which loses most of its energy Thereforethe EDS spectra collected at the TEM likely have sampled elements from the cytoplasmand cell membrane components both above and below the inclusion as well as from theP-rich inclusions nearby Other explanations are that hydrated sulfur inclusions changedtheir composition upon dehydration or that these two samples collected at different times
Organization of Inclusions in Magnetococci 401
FIGURE 9 Fluorescence micrograph of an ARB-1 cell suspension treated with Bacliteviability dyes showing that the nucleic acid staining pattern corresponds with the locationof the P-rich inclusions Baclite stains nucleic acids with cytonine (all cells) and propidiumiodate (dead or damaged cells) Bright double-lobed patterns are from ldquoliverdquo cells
of the day and the year may re ect a seasonal diurnal or cell cycle variability in the relativeproportions of the elements
Sulfur inclusions have been found in other uncultured magnetotactic bacteria includ-ing Magnetobacterium bavaricum (Spring et al 1993) Bilophococcus magnetotacticum(Moench 1988) and others (Iida and Akai 1996) and cultured species including the vibrioidstrains MV-1 and MV-2 the coccus strain MC-1 and the spirillum strain MV-4 (Bazylinskiand Frankel 2000) However only in Bilophococcus magnetotacticum was there a similarpolarization of sulfur-containing inclusions from the clustered magnetosomes within thecell (Moench 1988)
It seems likely that ARB-1 oxidizes H2S or other reduced sulfur compounds as anenergy source depositing ldquoelementalrdquo sulfur in intracellular inclusions like many othersul de-oxidizing bacteria including the anaerobic phototrophic purple sulfur bacteria suchas Chromatium vinosum and Thiocapsa roseopersicina the nonsulfur bacteria Beggiatoaspp and Thiothrix spp (Shively 1974 Friedrich 1998) and the magnetotactic bacteriamentioned before In these organisms the presence of sulfur globules is transitory andtakes place when the growth medium contains sul de andor thiosulfate When the sul deis completely utilized the sulfur is converted to sulfate (Stanier et al 1986)
Elemental S in sulfur globules is typically in a liquid state and analysis of in vivoglobules from some members of the purple and green sulfur bacteria by X-ray absorptionnear edge spectroscopy (Prange et al 1999) indicated the sulfur was in a chain structure
402 B L Cox et al
FIGURE 10 Phylogenetic position of six SSU rRNA gene clones related to previouslystudied MC This phylogenetic tree was rooted with Escherichia coli and constructed bythe neighbor joining method with the Kimura 2-parameter model for nucleotide change Amask including all nonambiguously aligned positions for 1427 nucleotides spanning nearlythe full length of the SSU rRNA gene was included Bootstrap values (100 replications)generated by the neighbor joining method are shown above relevant nodes and bootstrapvalues generated by maximum-parsimony analysis are shown below relevant nodes Onlybootstrap values above 50 are shown GenBank accession numbers are MC-CS92 X81182MC-pa119 Y13214 MC-CS81 X81184 MC-TB12 X81183 MC-MP17 X80996 MC-PA11 Y13212 MC12 L06456 partial sequence extended with data from MC-1 genomeRhizobium leguminosarum U31074 Agrobacterium tumefaciens D13294 Rhodobactersphaeroides X53854 Paracoccus denitri cans X59159 Zymomonas mobilis AF281034Sphingomonas paucimobilis D84528 Caulobacter crescentus M83799 Magnetospiril-lum magnetotacticum strain AMB1 D17514 S88429 Magnetospirillum magnetotacticumM10110 Rhodospirillum fulvum M59065 Magnetospirillum gryphiswaldense Y10109Magnetic Vibrio MV-1 LO6455 Oceanospirillum pusillum AB000482 Rhodospirillumrubrum D30778 Roseococcus thiosulfatophilus X72908 Escherichia coli J01695
Organization of Inclusions in Magnetococci 403
and was possibly complexed with water and organic residues If the S globules in ARB-1cells are analogous to those in the purple sulfur bacteria Chromatium vinosum and Thio-capsa roseopersicina they may contain a signi cant protein content Sulfur globules fromthe anaerobic phototrophs C vinosum and T roseopersicina were found to contain upto 52 protein that formed an envelope enclosing the S globules (Brune 1995) Theseproteins were isolated and analyzed and found to be glycine and tyrosine-rich structuralproteins which were homologous for the two species suggesting they are highly con-served (Brune 1995) Phylogenetically C vinosum and T roseopersicina are members ofthe deg -Proteobacteria whereas MC are members of the reg-Proteobacteria To our knowledgemagnetotactic bacteria may be the only members of the reg-Proteobacteria that make internalsulfur globules
Phylogenetic Af liation
We wished to detemine whether ARB-1 was a single species or a mixed population of MCToward that end 45 SSU rRNA genes were cloned from partially puri ed ARB-1 cells30 of which displayed similar restriction fragment length patterns (RFLP) to previouslycharacterized MC (data not shown) The nding that 15 patterns were not related to pre-viously characterized MC was not surprising given that the initial sample was not a pureculture but rather a population magnetically enriched from a sample originating from arelatively large vertical column containing both sediments and overlying water Fourteenof the MC-related clones representing different RFLP pattern groups were chosen for par-tial sequencing 10 of these sequences showed high similarity to previously characterizedMC Six of these sequences were phylogenetically distinct and were fully bidirectionallysequenced
ARB-1 MC SSU rRNA genes formed a monophyletic cluster related to but dis-tinct from other MC (Figure 10) Within the cluster the similarity between SSU rRNAgene sequences ranged from 97 (MC-H1arb and MC-g6arb) to 92 (MC-h1arb andMC-g2arb) The nearest neighbor to the ARB-1 cluster MC-CS92 was ca 89 similarand the nearest cultured relative MC-1 was ca 88 similar to the ARB-1 cluster As arule of thumb for SSU rRNA genes gt97 similarity is considered the same species andgt93ndash95 similarity is considered the same genus (Stackebrandt and Goebel 1994) Thesedata strongly suggest that ARB-1 represents a population of different diverse MC Therewas signi cant variation within the ARB-1 cluster and therefore we do not consider ARB-1to consist of bacteria of a single species or genus Correlations if any between phylogeneticand phenotypic variations in the ARB-1 cells have yet to be determined
Summary
We have partially characterized a population of magnetotactic cocci (MC) referred to asARB-1 which was particularly abundant in iron-rich sediments of a small eutrophic lake andmorphologically and structurally similar to an uncultured species of MC called Bilophococ-cus magnetotacticus The latter organism was found in large numbers in a sewage treatmentpond Cells of ARB-1 had a relatively wide size range of 1 to 4 sup1m They were motileGram-negative and bilophotrichously agellated Cells of this uncultured magnetotacticcoccus contained three types of intracellular inclusions that appeared to be in a consistentarrangement within the cell The magnetosomes contained crystals of magnetite and wereeither in chains andor polar clumps at the end of the cell where the agellar bundles werelocated Large P-rich bodies occupied the majority of the cell volume and appeared to sepa-rate the S-containing inclusions from the magnetosomes The presence of these large P-rich
404 B L Cox et al
inclusions might be an indication of cell stress or may be important in cell buoyancy or inenergy generation The presence of S-rich inclusions located at the end of the cell oppositethe magnetosomes strongly suggests that ARB-1 cells are sul de-oxidizing bacteria andthat the S might be useful as a stored electron donor when cells travel from sul de-rich tosul de-depleted zones in the water column The wide range of the cell sizes the differentmagnetosome arrangements and the large number of different retrieved 16S rRNA genesequences suggest that ARB-1 represents a population of at least several different MC 16SrRNA sequences obtained from cells of ARB-1 appeared to form a monophyletic clusterrelated to but distinct from other uncultured and cultured MC Future work using uores-cent oligonucleotide probes microscopy and physiological measurements should clarifyrelationships between phylogeny cell morphology and organization of inclusions
References
Balkwill DL Maratea D Blakemore RP 1980 Ultrastructure of a magnetotactic spirillum J Bacteriol141(3)1399ndash1408
Bazylinski DA Frankel RB 2000 Biologically controlled mineralization of magnetic iron mineralsby magnetotactic bacteria In Lovley DR editor Environmental microbe-metal interactionsWashington DC ASM Press p 109ndash143
Bazylinski DA Frankel RB Jannasch HW 1988 Anaerobic magnetite production by a marinemagnetotactic bacterium Nature 334518ndash519
Beveridge T 1999 Structures of gram-negative cell walls and their derived membrane vesiclesJ Bacteriol 181(6)4725ndash4733
Beveridge TJ Popkin TJ Cole RM 1994 Electron microscopy In Gerhardt P editor Methods forgeneral and molecular bacteriology Washington DC American Society for Microbiology p 25
Blakemore R 1975 Magnetotactic bacteria Science 190377ndash379Blakemore RP 1982 Magnetotactic bacteria Ann Rev Microbiol 36217ndash238Blakemore RP Maratea D Wolfe RS 1979 Isolation and pure culture of a freshwater magnetic
spirillum in chemically de ned medium J Bacteriol 140(2)720ndash729Blakemore RP Short KA Bazylinski DA Rosenblatt C Frankel RB 1985 Microaerobic conditions
are required for magnetite formation within Aquaspirillum magnetotacticum Geomicrobiol J4(1)53ndash71
Brune DC 1995 Isolation and characterization of sulfur globule proteins from Chromatium vinosumand Thiocapsa roseopersicina Arch Microbiol 163(1)391ndash399
Butler RF Banerjee SK 1975 Theoretical single-domain grain size range in magnetite and titano-magnetite J Geophys Res 804049ndash4058
Buwalda JP 1940 Geology of the Raymond Basin report to Pasadena Water DepartmentCrook R Allen CR Kamb B Payne CM Procter RJ 1987 Recent reverse faulting in the Transverse
Ranges California Pasadena CA US Geol Surv Prof Paper 133927ndash63Devouard B Posfai M Hua X Bazylinski DA Frankel RB Buseck PR 1998 Magnetite from
magnetotactic bacteria size distribution and twinning Am Mineral 831387ndash1398Frankel RB Bazylinski DA Johnson MS Taylor BL 1997 Magneto-Aerotaxis in marine coccoid
bacteria Biophys J 73994ndash1000Friedrich CG 1998 Physiology and genetics of sulfur-oxidizing bacteria Adv Microbial Physiol
39235ndash289Giovannoni SJ 1991 The polymerase chain reaction In Stackebrandt E Goodfellow M editors
Sequencing and hybridization techniques in bacterial systematics New York John Wiley andSons p 177ndash201
Guerrero R Pedros-Alio C Schmidt TM Mas J 1985 A survey of buoyant density of microorganismsin pure cultures and natural samples Microbiologia Sept 1(1ndash2)53ndash65
Harold FM 1966 Inorganic polyphosphates in biology structure metabolism and function BacteriolRev 30(4)772ndash794
Organization of Inclusions in Magnetococci 405
Iida A Akai J 1996 Crystalline sulfur inclusions in magnetotactic bacteria Sci Rep Nigata Univ SerE 1135ndash42
Kornberg A Fraley CD 2000 Inorganic polyphosphate a molecular fossil come to life ASM News66(5)275ndash280
Lins U Farina M 1999 Phosphorus-rich granules in uncultured magnetotactic bacteria FEMSMicrobiol Lett 172(2)23ndash28
Mann S Sparks NHC Frankel RB Bazylinski DA Jannasch HW 1990 Biomineralization of ferri-magnetic greigite (Fe3O4 and iron pyrite (FeS2 ) in a magnetotactic bacterium Nature 343258ndash
261Matsunaga T Sakaguchi T Tadokoro F 1991 Magnetite formation by a magnetic bacterium capable
of growing aerobically Appl Microbiol Biotechnol 35651ndash655Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993a Electron microscopy
study of magnetosomes in a cultured coccoid magnetotactic bacterium Proc Roy Soc Lond B251231ndash236
Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993b Electron microscopystudy of magnetosomes in two cultured vibrioid magnetotactic bacteria Proc Roy Soc Lond B251237ndash242
Moench TT 1988 Bilophococcus magnetotacticus gen nov sp nov a motile magnetic coccus Antvan Leeuwenhoek 54483ndash496
Moench TT Konetzka WA 1978 A novel method for the isolation and study of a magnetotacticbacterium Arch Microbiol 119203ndash212
Murray RGE Robinow CF 1994 Optical Microscopy In Gerhardt P editor Methods for generaland molecular bacteriology Washington DC American Society for Microbiology p 35ndash36
Prange A Arzberger I Engemann C Modrow H Schumann O Truper HG Steudel R Dahl C andHormes J 1999 In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteriaby X-ray absorption near edge spectroscopy Biochim Biophys Acta 1428446ndash454
Rappe MS Kemp PF Giovannoni SJ 1997 Phylogenetic diversity of marine coastal picoplankton16S rRNA genes cloned from the continental shelf off Cape Hatteras North Carolina LimnolOceanogr 42811ndash826
Reusch RN 2000 Transmembrane ion transport by polyphosphatepoly-(R)-3-hydroxybutyratecomplexes Biochemistry (Moscow) 65(3)335ndash352
Sakaguchi T Burgess JG Matsunaga T 1993 Magnetite formation by a sulphate-reducing bacteriumNature 36547ndash49
Schleifer KHSchuler D Spring S Weizenegger M Amann R 1991 The genus Magnetospirillum gennov description of Magnetospirillum gryphiswaldense sp nov and transfer of Aquaspirillummagnetotacticum to Magnetospirillum magnetotacticum comb nov System Appl Microbiol14379ndash385
Schuler D 1999 Formation of magnetosomes in magnetotactic bacteria J Mol Microbiol Biotechnol1(1)79ndash86
Schuler D Spring S Bazylinski DA 1999 Improved technique for the isolation of magnetotactic spir-illa from a freshwater sediment and their phylogenetic characterization System Appl Microbiol22466ndash471
Shively JM 1974 Inclusion bodies of prokaryotes Ann Rev Microbiol 28167ndash187Spring S Amann R Ludwig W Schleifer KH van Gemerden H Petersen N 1993 Dominating role
of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment ApplEnviron Microbiol 59(8)2397ndash2403
Spring S Amann R Ludwig W Schleifer KH Schuler D Poralla K Petersen N 1994 Phylogeneticanalysis of uncultured magnetotactic baceria from the Alpha-subclass of Proteobacteria SystemAppl Microbiol 17501ndash508
Stackebrandt E Goebel BM 1994 Taxonomic note a place for DNA-DNA reassociation and 16SrRNA sequence analysis in the present species de nition in bacteriology Int J System Bacteriol44846ndash849
Stanier RY Ingraham JL Wheelis ML Painter PR 1986 The microbial world 5th edition EnglewoodCliffs NJ Prentice Hall p 179
406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35
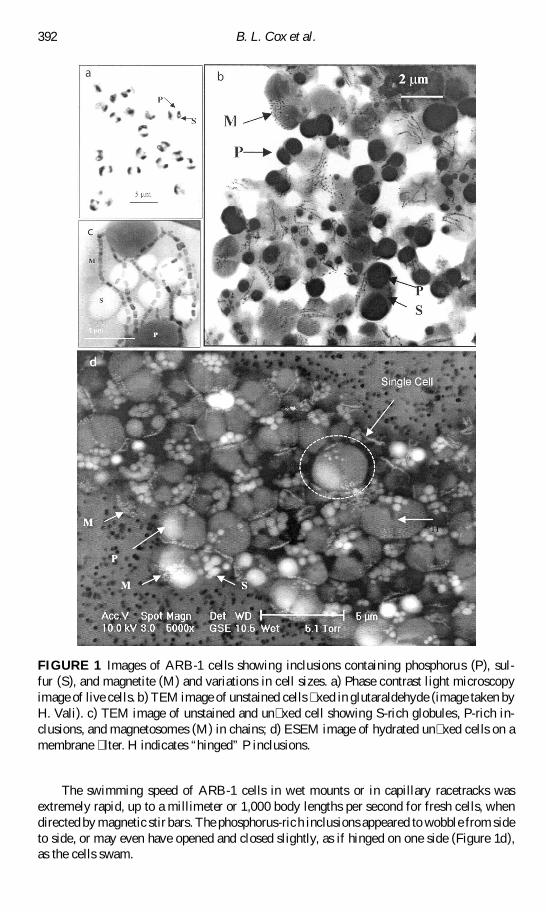
392 B L Cox et al
FIGURE 1 Images of ARB-1 cells showing inclusions containing phosphorus (P) sul-fur (S) and magnetite (M) and variations in cell sizes a) Phase contrast light microscopyimage of live cells b) TEM image of unstained cells xed in glutaraldehyde (image taken byH Vali) c) TEM image of unstained and un xed cell showing S-rich globules P-rich in-clusions and magnetosomes (M) in chains d) ESEM image of hydrated un xed cells on amembrane lter H indicates ldquohingedrdquo P inclusions
The swimming speed of ARB-1 cells in wet mounts or in capillary racetracks wasextremely rapid up to a millimeter or 1000 body lengths per second for fresh cells whendirected by magnetic stir bars The phosphorus-rich inclusions appeared to wobble from sideto side or may even have opened and closed slightly as if hinged on one side (Figure 1d)as the cells swam
Organization of Inclusions in Magnetococci 393
FIGURE 2 Images of ARB-1 cells showing cell division a) TEM image of cells stainedwith uranyl acetate showing division across rather than between P inclusions b) ESEMimage showing division of cells between P inclusions c) d) TEM images of cells stainedwith uranyl acetate showing unequal division of cells with protrusions
Flagella and Fimbriae or Pili
Like other MC (Blakemore 1975 1982 Moench 1988 Frankel et al 1997) cells of ARB-1were bilophotrichously agellated having two agellar bundles on one side of the cell(Figure 3) Each bundle contained about 10 unsheathed agella that were 13 nm in diameterCells also appeared to have other smaller protrusions that resemble pili (Figure 3b) Thetendency for cells of ARB-1 to stick to glass slides and cover slips and to each other supportsthe presence of mbriae or pili Pili have been observed on other uncultured (Blakemore1982) and cultured (Frankel et al 1997) MC
Magnetosomes
Magnetosome crystals in ARB-1 cells occurred in chains and clusters and ranged in lengthfrom 20 to 133 nm and in width from 10 to 84 nm with an average (sect standard deviation)length and width of 82 sect 23 nm and 49 sect 15 nm respectively (n D 341) The average lengthof clustered magnetosomes (n D 260) was 83 sect 24 nearly the same as the average lengthof magnetosomes from chains (n D 81 mean D 80 sect 19) The range of lengths of magneto-somes in chains (40ndash120 nm) was narrower than that of clustered magnetosomes The size
394 B L Cox et al
FIGURE 3 Images of ARB-1 cells showing agella andor pili a) Light microscopy imageof cells stained for agella Arrows indicate possible cell division plane b) Thin section of acell stained with uranyl acetate showing spaces where P inclusions had been S inclusionsmagnetosomes (M) and pili c) TEM image of whole cells with clustered magnetosomecrystals (M) negatively stained with uranyl acetate Notice haloes around crystals whichmight represent a membrane or coating
distributions (length and width) of all 341 magnetite crystals from cells of ARB-1 are plot-ted in Figure 4 Values plotted for ARB-1 magnetite crystals showed that like in virtually allother magnetotactic bacteria the magnetite particles in ARB-1 are single-magnetic-domaincrystals (Butler and Banerjee 1975 Bazylinski and Frankel 2000) These results were verysimilar to those obtained for the uncultured coccus Bilophococcus magnetotacticus (Toweand Moench 1981) which also possessed clumps of magnetosomes at one end of the cellrather than chains The average length of ARB-1 magnetosomes was also similar to thatof the cultured MC-1 However the shape of MC-1 magnetosomes was less elongated(Meldrum et al 1993a) It appears that the size distributions and aspect ratios for this typeof MC are distinct from those of other magnetotactic bacteria (eg MV-1 and MC-1) aswell as from those in the Mars meteorite ALH84001 (Thomas-Keprta et al 2000)
Morphologically the magnetosome crystals in ARB-1 were rectangular in projectionwith an hexagonal cross-section There were irregularities in the edges and interiors of the
FIG
UR
E4
Dis
trib
utio
nof
leng
ths
and
wid
ths
of34
1m
agne
tite
crys
tals
from
clus
ters
and
chai
nsin
AR
B-1
cells
a) F
requ
ency
asa
func
tion
ofle
ngth
b)
Freq
uenc
yas
afu
nctio
nof
wid
thc
) Wid
thas
afu
nctio
nof
leng
th
395
396 B L Cox et al
FIGURE 5 TEM image of a thin section of a cell of ARB-1 stained with uranyl acetateshowing magnetosomes before a) and after b) beam damage leaving crystal ldquodoubletsrdquo b)Notice disappearance of electron dense material in b)
crystals (Figure 3) The edges were often rounded and the particles occasionally con-stricted slightly in the middle The crystals sometimes had a mottled appearance and mayhave had fringes around the edges Once during TEM analysis of thin sections (80 kV)magnetosome crystals sustained signi cant beam damage after several minutes of viewingat a magni cation of 100000 (Figure 5) The crystals appeared to have broken in halfpossibly along twin planes in the (111) direction or along ldquowaistedrdquo regions where thecrystals showed some constriction and there appeared to be a loss of material Beam dam-age has been previously observed during TEM of greigite (Fe3S4) magnetosome particlesbut not in magnetite crystals (D Bazylinski H Vali and J Kirschvink personal commu-nications January 2001) except when viewed as whole mounts at 400 kV (Taylor et al2001) Taylor et al (2001) speculated that water vapor from the cells underwent radiolysisin the electron-beam producing a plasma that rapidly melted the magnetosomes
EDS analyses with both TEM and ESEM of the magnetosome particles (Figure 6c 6f)showed the particles were iron-rich with some extraneous signals including sulfur (Figure 6)A strong sulfur signal would be expected from EDS analyses of the magnetic mineral greigitealso known to be produced by some magnetotactic bacteria (Mann et al 1990) In ARB-1however these extraneous signals are likely due to the sulfur- and phosphorus-containinginclusions also present in the cell SAED patterns obtained from the crystals (Figure 7)and lattice fringes from high resolution TEM with spacings of 048 nm elongated in (111)direction (Figure 7) were consistent with the mineral magnetite
Magnetosomes within cells of ARB-1 were found in three different arrangements1) as a polar cluster forming a sort of ldquocaprdquo usually at the agellar end of the cell 2) in partialor complete chains at the same end of the cell or 3) a combination of both arrangementsThe polar clusters appear to be disorganized in many images (eg Figures 1 2 and 3)However the attraction of these cells to a magnet indicates they have permanent magneticdipole moments implying that they are not disorganized We are currently pursuing thisquestion of whether or not the clusters are disorganized
In cells having both chains and clusters it is dif cult to determine whether chainswere forming from the clustered crystals or whether the polar clusters of particles werefrom a chain that was disrupted during dehydration of the cell during preparation for TEMexamination However polar clusters of crystals were observed in ESEM images of many
Organization of Inclusions in Magnetococci 397
FIGURE 6 EDS spectra from TEM a) b) c) and ESEM d) e) f) showing elementalcomposition of P-rich a) d) and S-rich b) e) inclusions and magnetosomes c) f) Elementalcompositions of clustered magnetosomes were identical to those of magnetosomes in chainsAll y-axes are linear except for a) and b) which are log scales Noise in f ) is due to scattering
hydrated cells (Figure 1d) suggesting that they were not caused by dehydration In manyof the cells viewed by TEM it seemed that the chains tended to occur in smaller morespherical cells while clumps of magnetosomes occurred more frequently in elongatedcells In order to see if there was any relationship between cell size and shape and the threetypes of magnetosome arrangements the size and aspect ratio for cells with these differentmagnetosome arrangements were measured (Figure 8) The cell size distribution appearedto be narrower for cells with chains than for the cells with disorganized crystals Cells withchains were predominantly smaller than 25 sup1m (Figure 8a) The aspect ratios of the cellswith chains only was smaller than that of cells with disorganized magnetosomes and wasshifted toward spherical aspect ratios (Figure 8b)
One explanation for these observations is that there are different stages of magnetosomeformation in a single species of MC The disorganized magnetosomes might form near thecell membrane between the agellar bundles and then extend into chains Alternativelymagnetosomes might rst form as chains in smaller cells At a later stage as cells increasein size the membranes linking the chains dissolve or become stressed and the magneto-somes somehow migrate to the agellar end of the cell Because magnetosomes appear tobe anchored to the inner surface of the cell membrane in virtually all species examined(Bazylinski and Frankel 2000) and are never free- oating in the cytoplasm migration ofmagnetosomes through the cytoplasm seems unlikely Instead we are presently designinghybridization experiments to address the hypothesis that the relationships between cell sizeshape and magnetosome arrangement correlate with the genotypical variations in ARB-1described later Spring et al (1994) analyzed communities of MC found in freshwater lakesin Germany using uorescent oligonucleotide probes They found different genotypes ofMC (similarity below 95) but these genotypes were not phenotypically expressed asdifferent magnetosome organizations but as preferences for different concentrations of O2
398 B L Cox et al
FIGURE 7 Evidence supporting magnetite as the magnetosome mineral in ARB-1 cellsa) Image of a single crystal of magnetite within a cell in an unstained glutaraldehyde- xedwhole cell mount taken with High Resolution TEM by H Vali b) Part of the magnetitecrystal shown in a) magni ed to show the distance between lattice fringes c) Selectedarea electron diffraction (SAED) of a group of magnetosomes in a cell from an un xedunstained whole cell mount showing the indices of some of the diffraction spots
Therefore we might expect no correlation between magnetite organization and phyloge-netic diversity of ARB-1 cells but perhaps a correlation between oxygen preference andphylogeny
A third possibility is that chains and clumps of magnetosomes form as a result ofdifferent environmental conditions Since our sampling methods were not selective as tothe cellsrsquo location in the vertical chemical (eg oxygen) and redox gradients that are sure to
Organization of Inclusions in Magnetococci 399
FIGURE 8 Relationships between organization of magnetosomes and cell size a) and cellaspect ratio b) Measurements were taken from both TEM and ESEM images Solid patternis for cells with clustered magnetosomes crossed pattern is for cells with magnetosomesin chains only Patterned bars originate at the top of solid bars
exist at the site the environmental conditions of our samples represent a relatively large rangeof oxygen and perhaps other chemical concentrations as well as redox conditions Magneticbacteria in culture are known to vary the size and number of magnetosomes in responseto the media composition (Meldrum et al 1993a) and particularly oxygen concentration(Blakemore et al 1985) Another possibility is that the clustered magnetosomes are formedby a different biological mechanism and serve a different function than magnetosomes inchains
Phosphorus (P) Inclusions
The two very large inclusion bodies seen in each cell always occurred in pairs in cellsand seemed to extend to the cell membrane and to be in contact with each other Thus themaximum dimension of the P-rich inclusions was the same as the minimum dimension ofthe cell and their combined width was approximately half of the long dimension of the cellIn ESEM images the two P-containing inclusions appeared to be attached in the middle insome cells and separated in others (Figure 1d) The P-containing inclusions appeared to beself-contained and perhaps bounded by some sort of coating or membrane in TEM images(Figure 2) and in light microscopy images of cells stained with safranin (not shown)
These inclusions were shown by both ESEM and TEM EDS to contain phosphorus with generally small amounts of potassium calcium magnesium sulfur and iron the lattertwo elements likely being due to the close proximity of the P-rich inclusions to sulfurglobules and magnetosomes (Figures 1 and 6) A very large calcium peak was observedwhen these large inclusions were analyzed in the TEM (Figure 6a) The samples shown inFigures 6a and 6d were collected at different times of the year which may explain some ofthe differences observed in these spectra Other EDS data from the ESEM (not shown) hadhigher calcium peaks than those shown in Figure 6d indicating that relative proportions ofP and Ca in P-rich inclusions vary even within the same sample
The inclusions were electron dense in TEM and had a mottled appearance due toablation by the electron beam Similar inclusions have been described for uncultured MCfrom the Itaupo Lagoon near Rio de Janeiro Brazil (Lins and Farina 1999) and from MillPond New Hampshire (Blakemore 1982) Like the structures in ARB-1 polyP granules aretranslucent in visible light (Spring et al 1994) dark (electron dense) in the TEM and oftenhave a mottled appearance (Harold 1966) The P-rich inclusions in ARB-1 cells stained
400 B L Cox et al
with methylene blue (Murray and Robinson 1994) which is suggestive of the presence ofpolyP but not con rmatory
PolyP is composed of linear chains of tetrahedral phosphate residues linked throughcommon oxygen atoms by phosphoanhydrid e bonds At physiological pH each unit carriesa monovalent negative charge giving polyP the property of having a large cation exchangecapacity The binding energy of polyP allows it to sequester Ca2C and Mg2C (Reusch 2000)consistent with the Ca2C and Mg2C signals observed in the EDS spectra of the ARB-1inclusions (Figure 6a 6d) The binding of these metals to the polyP granules contributesto their high electron density PolyP is often associated with polyhydroxybutyrat e (PHB)PHBs solvate (dissolve) cations and associate withpolyP to form Ca2C selective ion channelsacross plasma membranes (Reusch 2000)
The presence of polyP bodies is common in bacteria often resulting as a bacterialresponse to stress and they may play a role in cell motility quorum sensing and bio lmformation (Kornberg and Fraley 2000) Sulfate starvation is known to lead to rapid andmassive accumulation of polyP in some bacteria (Stanier et al 1986) When cells regainaccess to sulfate 32P uptake studies showed that PO3iexcl
4 present in the polyP bodies wasincorporated into nucleic acids PolyP is synthesized from ATP by the enzyme polyP kinasePolyP granules have been reported to be associated with nucleic acids (Harold 1966) butit is not clear whether the association occurs in vivo or after extraction This associationwith nucleic acids would be consistent with staining patterns seen when ARB-1 cells wereexamined with a uorescent livedead stain (nucleic acid stain) displaying a double-lobedstructure in all of the cells (Figure 9) This dumbbell shape appears to extend inside andacross the two phosphorus inclusions
The physiological function of the P inclusions in ARB-1 is not yet known but fromother cell systems several possibilities can be considered These inclusions might serveas a phosphorus reserve for the biosynthesis of nucleic acids and phospholipids underconditions of phosphorus starvation (Harold 1966) Still another possibility is buoyancyregulation Cells can vary their density by as much as 7 by incorporating substances intoinclusions (Guerrero et al 1985) A more speculative possibility is that ARB-1 cells canderive energy from polyP Acinetobacter johnsonii 210a is known to couple the excretion ofa metal phosphate chelate and HC generating a proton motive force (VanVeen et al 1994)A johnsonii takes up P from the aquatic environment by using two Pi transport systemsand stores it as intracellular granules of polyP
Sulfur (S) Inclusions
Cells of ARB-1 contained 5 to 10 spherical S-rich inclusions (Figures 1d) These rangedin size from approximately 100 to 700 nm although most of the S-rich inclusions in asingle cell were close to the same size and were located at one end of the cell oppositethe side where the magnetosomes were located and agellar bundles attached Like theP-rich inclusions the S-rich inclusions appeared to be enclosed in a coating or membrane(Figure 1d) EDS analysis from the ESEM showed these structures contained mostly S withsome carbon and oxygen (Figure 6e) EDS analysis from the TEM also showed S but evenhigher counts for Ca and P (Figure 6b) Beam spreading for the two methods is within aradius of 200 nm The X-rays produced by the 200-keV TEM beam sample the entire verticalextent of the cell unlike the 20 keV SEM beam which loses most of its energy Thereforethe EDS spectra collected at the TEM likely have sampled elements from the cytoplasmand cell membrane components both above and below the inclusion as well as from theP-rich inclusions nearby Other explanations are that hydrated sulfur inclusions changedtheir composition upon dehydration or that these two samples collected at different times
Organization of Inclusions in Magnetococci 401
FIGURE 9 Fluorescence micrograph of an ARB-1 cell suspension treated with Bacliteviability dyes showing that the nucleic acid staining pattern corresponds with the locationof the P-rich inclusions Baclite stains nucleic acids with cytonine (all cells) and propidiumiodate (dead or damaged cells) Bright double-lobed patterns are from ldquoliverdquo cells
of the day and the year may re ect a seasonal diurnal or cell cycle variability in the relativeproportions of the elements
Sulfur inclusions have been found in other uncultured magnetotactic bacteria includ-ing Magnetobacterium bavaricum (Spring et al 1993) Bilophococcus magnetotacticum(Moench 1988) and others (Iida and Akai 1996) and cultured species including the vibrioidstrains MV-1 and MV-2 the coccus strain MC-1 and the spirillum strain MV-4 (Bazylinskiand Frankel 2000) However only in Bilophococcus magnetotacticum was there a similarpolarization of sulfur-containing inclusions from the clustered magnetosomes within thecell (Moench 1988)
It seems likely that ARB-1 oxidizes H2S or other reduced sulfur compounds as anenergy source depositing ldquoelementalrdquo sulfur in intracellular inclusions like many othersul de-oxidizing bacteria including the anaerobic phototrophic purple sulfur bacteria suchas Chromatium vinosum and Thiocapsa roseopersicina the nonsulfur bacteria Beggiatoaspp and Thiothrix spp (Shively 1974 Friedrich 1998) and the magnetotactic bacteriamentioned before In these organisms the presence of sulfur globules is transitory andtakes place when the growth medium contains sul de andor thiosulfate When the sul deis completely utilized the sulfur is converted to sulfate (Stanier et al 1986)
Elemental S in sulfur globules is typically in a liquid state and analysis of in vivoglobules from some members of the purple and green sulfur bacteria by X-ray absorptionnear edge spectroscopy (Prange et al 1999) indicated the sulfur was in a chain structure
402 B L Cox et al
FIGURE 10 Phylogenetic position of six SSU rRNA gene clones related to previouslystudied MC This phylogenetic tree was rooted with Escherichia coli and constructed bythe neighbor joining method with the Kimura 2-parameter model for nucleotide change Amask including all nonambiguously aligned positions for 1427 nucleotides spanning nearlythe full length of the SSU rRNA gene was included Bootstrap values (100 replications)generated by the neighbor joining method are shown above relevant nodes and bootstrapvalues generated by maximum-parsimony analysis are shown below relevant nodes Onlybootstrap values above 50 are shown GenBank accession numbers are MC-CS92 X81182MC-pa119 Y13214 MC-CS81 X81184 MC-TB12 X81183 MC-MP17 X80996 MC-PA11 Y13212 MC12 L06456 partial sequence extended with data from MC-1 genomeRhizobium leguminosarum U31074 Agrobacterium tumefaciens D13294 Rhodobactersphaeroides X53854 Paracoccus denitri cans X59159 Zymomonas mobilis AF281034Sphingomonas paucimobilis D84528 Caulobacter crescentus M83799 Magnetospiril-lum magnetotacticum strain AMB1 D17514 S88429 Magnetospirillum magnetotacticumM10110 Rhodospirillum fulvum M59065 Magnetospirillum gryphiswaldense Y10109Magnetic Vibrio MV-1 LO6455 Oceanospirillum pusillum AB000482 Rhodospirillumrubrum D30778 Roseococcus thiosulfatophilus X72908 Escherichia coli J01695
Organization of Inclusions in Magnetococci 403
and was possibly complexed with water and organic residues If the S globules in ARB-1cells are analogous to those in the purple sulfur bacteria Chromatium vinosum and Thio-capsa roseopersicina they may contain a signi cant protein content Sulfur globules fromthe anaerobic phototrophs C vinosum and T roseopersicina were found to contain upto 52 protein that formed an envelope enclosing the S globules (Brune 1995) Theseproteins were isolated and analyzed and found to be glycine and tyrosine-rich structuralproteins which were homologous for the two species suggesting they are highly con-served (Brune 1995) Phylogenetically C vinosum and T roseopersicina are members ofthe deg -Proteobacteria whereas MC are members of the reg-Proteobacteria To our knowledgemagnetotactic bacteria may be the only members of the reg-Proteobacteria that make internalsulfur globules
Phylogenetic Af liation
We wished to detemine whether ARB-1 was a single species or a mixed population of MCToward that end 45 SSU rRNA genes were cloned from partially puri ed ARB-1 cells30 of which displayed similar restriction fragment length patterns (RFLP) to previouslycharacterized MC (data not shown) The nding that 15 patterns were not related to pre-viously characterized MC was not surprising given that the initial sample was not a pureculture but rather a population magnetically enriched from a sample originating from arelatively large vertical column containing both sediments and overlying water Fourteenof the MC-related clones representing different RFLP pattern groups were chosen for par-tial sequencing 10 of these sequences showed high similarity to previously characterizedMC Six of these sequences were phylogenetically distinct and were fully bidirectionallysequenced
ARB-1 MC SSU rRNA genes formed a monophyletic cluster related to but dis-tinct from other MC (Figure 10) Within the cluster the similarity between SSU rRNAgene sequences ranged from 97 (MC-H1arb and MC-g6arb) to 92 (MC-h1arb andMC-g2arb) The nearest neighbor to the ARB-1 cluster MC-CS92 was ca 89 similarand the nearest cultured relative MC-1 was ca 88 similar to the ARB-1 cluster As arule of thumb for SSU rRNA genes gt97 similarity is considered the same species andgt93ndash95 similarity is considered the same genus (Stackebrandt and Goebel 1994) Thesedata strongly suggest that ARB-1 represents a population of different diverse MC Therewas signi cant variation within the ARB-1 cluster and therefore we do not consider ARB-1to consist of bacteria of a single species or genus Correlations if any between phylogeneticand phenotypic variations in the ARB-1 cells have yet to be determined
Summary
We have partially characterized a population of magnetotactic cocci (MC) referred to asARB-1 which was particularly abundant in iron-rich sediments of a small eutrophic lake andmorphologically and structurally similar to an uncultured species of MC called Bilophococ-cus magnetotacticus The latter organism was found in large numbers in a sewage treatmentpond Cells of ARB-1 had a relatively wide size range of 1 to 4 sup1m They were motileGram-negative and bilophotrichously agellated Cells of this uncultured magnetotacticcoccus contained three types of intracellular inclusions that appeared to be in a consistentarrangement within the cell The magnetosomes contained crystals of magnetite and wereeither in chains andor polar clumps at the end of the cell where the agellar bundles werelocated Large P-rich bodies occupied the majority of the cell volume and appeared to sepa-rate the S-containing inclusions from the magnetosomes The presence of these large P-rich
404 B L Cox et al
inclusions might be an indication of cell stress or may be important in cell buoyancy or inenergy generation The presence of S-rich inclusions located at the end of the cell oppositethe magnetosomes strongly suggests that ARB-1 cells are sul de-oxidizing bacteria andthat the S might be useful as a stored electron donor when cells travel from sul de-rich tosul de-depleted zones in the water column The wide range of the cell sizes the differentmagnetosome arrangements and the large number of different retrieved 16S rRNA genesequences suggest that ARB-1 represents a population of at least several different MC 16SrRNA sequences obtained from cells of ARB-1 appeared to form a monophyletic clusterrelated to but distinct from other uncultured and cultured MC Future work using uores-cent oligonucleotide probes microscopy and physiological measurements should clarifyrelationships between phylogeny cell morphology and organization of inclusions
References
Balkwill DL Maratea D Blakemore RP 1980 Ultrastructure of a magnetotactic spirillum J Bacteriol141(3)1399ndash1408
Bazylinski DA Frankel RB 2000 Biologically controlled mineralization of magnetic iron mineralsby magnetotactic bacteria In Lovley DR editor Environmental microbe-metal interactionsWashington DC ASM Press p 109ndash143
Bazylinski DA Frankel RB Jannasch HW 1988 Anaerobic magnetite production by a marinemagnetotactic bacterium Nature 334518ndash519
Beveridge T 1999 Structures of gram-negative cell walls and their derived membrane vesiclesJ Bacteriol 181(6)4725ndash4733
Beveridge TJ Popkin TJ Cole RM 1994 Electron microscopy In Gerhardt P editor Methods forgeneral and molecular bacteriology Washington DC American Society for Microbiology p 25
Blakemore R 1975 Magnetotactic bacteria Science 190377ndash379Blakemore RP 1982 Magnetotactic bacteria Ann Rev Microbiol 36217ndash238Blakemore RP Maratea D Wolfe RS 1979 Isolation and pure culture of a freshwater magnetic
spirillum in chemically de ned medium J Bacteriol 140(2)720ndash729Blakemore RP Short KA Bazylinski DA Rosenblatt C Frankel RB 1985 Microaerobic conditions
are required for magnetite formation within Aquaspirillum magnetotacticum Geomicrobiol J4(1)53ndash71
Brune DC 1995 Isolation and characterization of sulfur globule proteins from Chromatium vinosumand Thiocapsa roseopersicina Arch Microbiol 163(1)391ndash399
Butler RF Banerjee SK 1975 Theoretical single-domain grain size range in magnetite and titano-magnetite J Geophys Res 804049ndash4058
Buwalda JP 1940 Geology of the Raymond Basin report to Pasadena Water DepartmentCrook R Allen CR Kamb B Payne CM Procter RJ 1987 Recent reverse faulting in the Transverse
Ranges California Pasadena CA US Geol Surv Prof Paper 133927ndash63Devouard B Posfai M Hua X Bazylinski DA Frankel RB Buseck PR 1998 Magnetite from
magnetotactic bacteria size distribution and twinning Am Mineral 831387ndash1398Frankel RB Bazylinski DA Johnson MS Taylor BL 1997 Magneto-Aerotaxis in marine coccoid
bacteria Biophys J 73994ndash1000Friedrich CG 1998 Physiology and genetics of sulfur-oxidizing bacteria Adv Microbial Physiol
39235ndash289Giovannoni SJ 1991 The polymerase chain reaction In Stackebrandt E Goodfellow M editors
Sequencing and hybridization techniques in bacterial systematics New York John Wiley andSons p 177ndash201
Guerrero R Pedros-Alio C Schmidt TM Mas J 1985 A survey of buoyant density of microorganismsin pure cultures and natural samples Microbiologia Sept 1(1ndash2)53ndash65
Harold FM 1966 Inorganic polyphosphates in biology structure metabolism and function BacteriolRev 30(4)772ndash794
Organization of Inclusions in Magnetococci 405
Iida A Akai J 1996 Crystalline sulfur inclusions in magnetotactic bacteria Sci Rep Nigata Univ SerE 1135ndash42
Kornberg A Fraley CD 2000 Inorganic polyphosphate a molecular fossil come to life ASM News66(5)275ndash280
Lins U Farina M 1999 Phosphorus-rich granules in uncultured magnetotactic bacteria FEMSMicrobiol Lett 172(2)23ndash28
Mann S Sparks NHC Frankel RB Bazylinski DA Jannasch HW 1990 Biomineralization of ferri-magnetic greigite (Fe3O4 and iron pyrite (FeS2 ) in a magnetotactic bacterium Nature 343258ndash
261Matsunaga T Sakaguchi T Tadokoro F 1991 Magnetite formation by a magnetic bacterium capable
of growing aerobically Appl Microbiol Biotechnol 35651ndash655Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993a Electron microscopy
study of magnetosomes in a cultured coccoid magnetotactic bacterium Proc Roy Soc Lond B251231ndash236
Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993b Electron microscopystudy of magnetosomes in two cultured vibrioid magnetotactic bacteria Proc Roy Soc Lond B251237ndash242
Moench TT 1988 Bilophococcus magnetotacticus gen nov sp nov a motile magnetic coccus Antvan Leeuwenhoek 54483ndash496
Moench TT Konetzka WA 1978 A novel method for the isolation and study of a magnetotacticbacterium Arch Microbiol 119203ndash212
Murray RGE Robinow CF 1994 Optical Microscopy In Gerhardt P editor Methods for generaland molecular bacteriology Washington DC American Society for Microbiology p 35ndash36
Prange A Arzberger I Engemann C Modrow H Schumann O Truper HG Steudel R Dahl C andHormes J 1999 In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteriaby X-ray absorption near edge spectroscopy Biochim Biophys Acta 1428446ndash454
Rappe MS Kemp PF Giovannoni SJ 1997 Phylogenetic diversity of marine coastal picoplankton16S rRNA genes cloned from the continental shelf off Cape Hatteras North Carolina LimnolOceanogr 42811ndash826
Reusch RN 2000 Transmembrane ion transport by polyphosphatepoly-(R)-3-hydroxybutyratecomplexes Biochemistry (Moscow) 65(3)335ndash352
Sakaguchi T Burgess JG Matsunaga T 1993 Magnetite formation by a sulphate-reducing bacteriumNature 36547ndash49
Schleifer KHSchuler D Spring S Weizenegger M Amann R 1991 The genus Magnetospirillum gennov description of Magnetospirillum gryphiswaldense sp nov and transfer of Aquaspirillummagnetotacticum to Magnetospirillum magnetotacticum comb nov System Appl Microbiol14379ndash385
Schuler D 1999 Formation of magnetosomes in magnetotactic bacteria J Mol Microbiol Biotechnol1(1)79ndash86
Schuler D Spring S Bazylinski DA 1999 Improved technique for the isolation of magnetotactic spir-illa from a freshwater sediment and their phylogenetic characterization System Appl Microbiol22466ndash471
Shively JM 1974 Inclusion bodies of prokaryotes Ann Rev Microbiol 28167ndash187Spring S Amann R Ludwig W Schleifer KH van Gemerden H Petersen N 1993 Dominating role
of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment ApplEnviron Microbiol 59(8)2397ndash2403
Spring S Amann R Ludwig W Schleifer KH Schuler D Poralla K Petersen N 1994 Phylogeneticanalysis of uncultured magnetotactic baceria from the Alpha-subclass of Proteobacteria SystemAppl Microbiol 17501ndash508
Stackebrandt E Goebel BM 1994 Taxonomic note a place for DNA-DNA reassociation and 16SrRNA sequence analysis in the present species de nition in bacteriology Int J System Bacteriol44846ndash849
Stanier RY Ingraham JL Wheelis ML Painter PR 1986 The microbial world 5th edition EnglewoodCliffs NJ Prentice Hall p 179
406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35
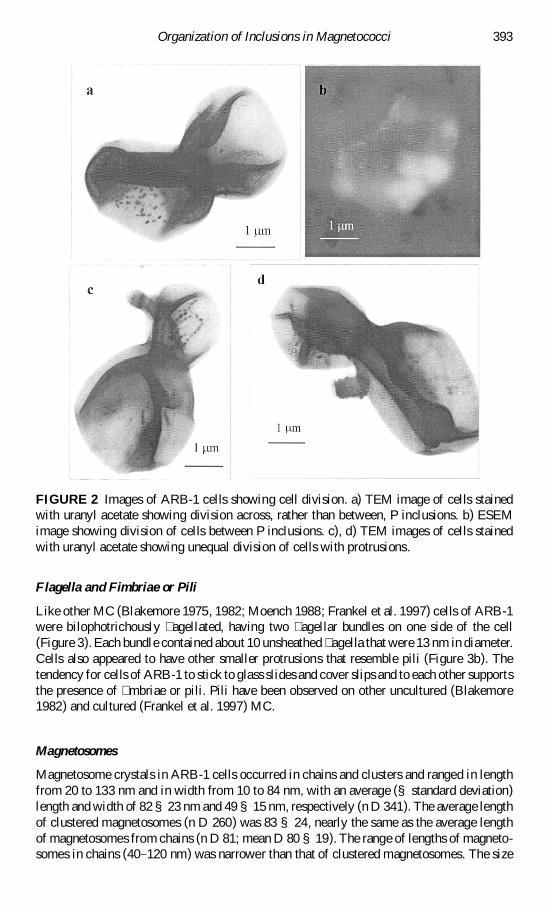
Organization of Inclusions in Magnetococci 393
FIGURE 2 Images of ARB-1 cells showing cell division a) TEM image of cells stainedwith uranyl acetate showing division across rather than between P inclusions b) ESEMimage showing division of cells between P inclusions c) d) TEM images of cells stainedwith uranyl acetate showing unequal division of cells with protrusions
Flagella and Fimbriae or Pili
Like other MC (Blakemore 1975 1982 Moench 1988 Frankel et al 1997) cells of ARB-1were bilophotrichously agellated having two agellar bundles on one side of the cell(Figure 3) Each bundle contained about 10 unsheathed agella that were 13 nm in diameterCells also appeared to have other smaller protrusions that resemble pili (Figure 3b) Thetendency for cells of ARB-1 to stick to glass slides and cover slips and to each other supportsthe presence of mbriae or pili Pili have been observed on other uncultured (Blakemore1982) and cultured (Frankel et al 1997) MC
Magnetosomes
Magnetosome crystals in ARB-1 cells occurred in chains and clusters and ranged in lengthfrom 20 to 133 nm and in width from 10 to 84 nm with an average (sect standard deviation)length and width of 82 sect 23 nm and 49 sect 15 nm respectively (n D 341) The average lengthof clustered magnetosomes (n D 260) was 83 sect 24 nearly the same as the average lengthof magnetosomes from chains (n D 81 mean D 80 sect 19) The range of lengths of magneto-somes in chains (40ndash120 nm) was narrower than that of clustered magnetosomes The size
394 B L Cox et al
FIGURE 3 Images of ARB-1 cells showing agella andor pili a) Light microscopy imageof cells stained for agella Arrows indicate possible cell division plane b) Thin section of acell stained with uranyl acetate showing spaces where P inclusions had been S inclusionsmagnetosomes (M) and pili c) TEM image of whole cells with clustered magnetosomecrystals (M) negatively stained with uranyl acetate Notice haloes around crystals whichmight represent a membrane or coating
distributions (length and width) of all 341 magnetite crystals from cells of ARB-1 are plot-ted in Figure 4 Values plotted for ARB-1 magnetite crystals showed that like in virtually allother magnetotactic bacteria the magnetite particles in ARB-1 are single-magnetic-domaincrystals (Butler and Banerjee 1975 Bazylinski and Frankel 2000) These results were verysimilar to those obtained for the uncultured coccus Bilophococcus magnetotacticus (Toweand Moench 1981) which also possessed clumps of magnetosomes at one end of the cellrather than chains The average length of ARB-1 magnetosomes was also similar to thatof the cultured MC-1 However the shape of MC-1 magnetosomes was less elongated(Meldrum et al 1993a) It appears that the size distributions and aspect ratios for this typeof MC are distinct from those of other magnetotactic bacteria (eg MV-1 and MC-1) aswell as from those in the Mars meteorite ALH84001 (Thomas-Keprta et al 2000)
Morphologically the magnetosome crystals in ARB-1 were rectangular in projectionwith an hexagonal cross-section There were irregularities in the edges and interiors of the
FIG
UR
E4
Dis
trib
utio
nof
leng
ths
and
wid
ths
of34
1m
agne
tite
crys
tals
from
clus
ters
and
chai
nsin
AR
B-1
cells
a) F
requ
ency
asa
func
tion
ofle
ngth
b)
Freq
uenc
yas
afu
nctio
nof
wid
thc
) Wid
thas
afu
nctio
nof
leng
th
395
396 B L Cox et al
FIGURE 5 TEM image of a thin section of a cell of ARB-1 stained with uranyl acetateshowing magnetosomes before a) and after b) beam damage leaving crystal ldquodoubletsrdquo b)Notice disappearance of electron dense material in b)
crystals (Figure 3) The edges were often rounded and the particles occasionally con-stricted slightly in the middle The crystals sometimes had a mottled appearance and mayhave had fringes around the edges Once during TEM analysis of thin sections (80 kV)magnetosome crystals sustained signi cant beam damage after several minutes of viewingat a magni cation of 100000 (Figure 5) The crystals appeared to have broken in halfpossibly along twin planes in the (111) direction or along ldquowaistedrdquo regions where thecrystals showed some constriction and there appeared to be a loss of material Beam dam-age has been previously observed during TEM of greigite (Fe3S4) magnetosome particlesbut not in magnetite crystals (D Bazylinski H Vali and J Kirschvink personal commu-nications January 2001) except when viewed as whole mounts at 400 kV (Taylor et al2001) Taylor et al (2001) speculated that water vapor from the cells underwent radiolysisin the electron-beam producing a plasma that rapidly melted the magnetosomes
EDS analyses with both TEM and ESEM of the magnetosome particles (Figure 6c 6f)showed the particles were iron-rich with some extraneous signals including sulfur (Figure 6)A strong sulfur signal would be expected from EDS analyses of the magnetic mineral greigitealso known to be produced by some magnetotactic bacteria (Mann et al 1990) In ARB-1however these extraneous signals are likely due to the sulfur- and phosphorus-containinginclusions also present in the cell SAED patterns obtained from the crystals (Figure 7)and lattice fringes from high resolution TEM with spacings of 048 nm elongated in (111)direction (Figure 7) were consistent with the mineral magnetite
Magnetosomes within cells of ARB-1 were found in three different arrangements1) as a polar cluster forming a sort of ldquocaprdquo usually at the agellar end of the cell 2) in partialor complete chains at the same end of the cell or 3) a combination of both arrangementsThe polar clusters appear to be disorganized in many images (eg Figures 1 2 and 3)However the attraction of these cells to a magnet indicates they have permanent magneticdipole moments implying that they are not disorganized We are currently pursuing thisquestion of whether or not the clusters are disorganized
In cells having both chains and clusters it is dif cult to determine whether chainswere forming from the clustered crystals or whether the polar clusters of particles werefrom a chain that was disrupted during dehydration of the cell during preparation for TEMexamination However polar clusters of crystals were observed in ESEM images of many
Organization of Inclusions in Magnetococci 397
FIGURE 6 EDS spectra from TEM a) b) c) and ESEM d) e) f) showing elementalcomposition of P-rich a) d) and S-rich b) e) inclusions and magnetosomes c) f) Elementalcompositions of clustered magnetosomes were identical to those of magnetosomes in chainsAll y-axes are linear except for a) and b) which are log scales Noise in f ) is due to scattering
hydrated cells (Figure 1d) suggesting that they were not caused by dehydration In manyof the cells viewed by TEM it seemed that the chains tended to occur in smaller morespherical cells while clumps of magnetosomes occurred more frequently in elongatedcells In order to see if there was any relationship between cell size and shape and the threetypes of magnetosome arrangements the size and aspect ratio for cells with these differentmagnetosome arrangements were measured (Figure 8) The cell size distribution appearedto be narrower for cells with chains than for the cells with disorganized crystals Cells withchains were predominantly smaller than 25 sup1m (Figure 8a) The aspect ratios of the cellswith chains only was smaller than that of cells with disorganized magnetosomes and wasshifted toward spherical aspect ratios (Figure 8b)
One explanation for these observations is that there are different stages of magnetosomeformation in a single species of MC The disorganized magnetosomes might form near thecell membrane between the agellar bundles and then extend into chains Alternativelymagnetosomes might rst form as chains in smaller cells At a later stage as cells increasein size the membranes linking the chains dissolve or become stressed and the magneto-somes somehow migrate to the agellar end of the cell Because magnetosomes appear tobe anchored to the inner surface of the cell membrane in virtually all species examined(Bazylinski and Frankel 2000) and are never free- oating in the cytoplasm migration ofmagnetosomes through the cytoplasm seems unlikely Instead we are presently designinghybridization experiments to address the hypothesis that the relationships between cell sizeshape and magnetosome arrangement correlate with the genotypical variations in ARB-1described later Spring et al (1994) analyzed communities of MC found in freshwater lakesin Germany using uorescent oligonucleotide probes They found different genotypes ofMC (similarity below 95) but these genotypes were not phenotypically expressed asdifferent magnetosome organizations but as preferences for different concentrations of O2
398 B L Cox et al
FIGURE 7 Evidence supporting magnetite as the magnetosome mineral in ARB-1 cellsa) Image of a single crystal of magnetite within a cell in an unstained glutaraldehyde- xedwhole cell mount taken with High Resolution TEM by H Vali b) Part of the magnetitecrystal shown in a) magni ed to show the distance between lattice fringes c) Selectedarea electron diffraction (SAED) of a group of magnetosomes in a cell from an un xedunstained whole cell mount showing the indices of some of the diffraction spots
Therefore we might expect no correlation between magnetite organization and phyloge-netic diversity of ARB-1 cells but perhaps a correlation between oxygen preference andphylogeny
A third possibility is that chains and clumps of magnetosomes form as a result ofdifferent environmental conditions Since our sampling methods were not selective as tothe cellsrsquo location in the vertical chemical (eg oxygen) and redox gradients that are sure to
Organization of Inclusions in Magnetococci 399
FIGURE 8 Relationships between organization of magnetosomes and cell size a) and cellaspect ratio b) Measurements were taken from both TEM and ESEM images Solid patternis for cells with clustered magnetosomes crossed pattern is for cells with magnetosomesin chains only Patterned bars originate at the top of solid bars
exist at the site the environmental conditions of our samples represent a relatively large rangeof oxygen and perhaps other chemical concentrations as well as redox conditions Magneticbacteria in culture are known to vary the size and number of magnetosomes in responseto the media composition (Meldrum et al 1993a) and particularly oxygen concentration(Blakemore et al 1985) Another possibility is that the clustered magnetosomes are formedby a different biological mechanism and serve a different function than magnetosomes inchains
Phosphorus (P) Inclusions
The two very large inclusion bodies seen in each cell always occurred in pairs in cellsand seemed to extend to the cell membrane and to be in contact with each other Thus themaximum dimension of the P-rich inclusions was the same as the minimum dimension ofthe cell and their combined width was approximately half of the long dimension of the cellIn ESEM images the two P-containing inclusions appeared to be attached in the middle insome cells and separated in others (Figure 1d) The P-containing inclusions appeared to beself-contained and perhaps bounded by some sort of coating or membrane in TEM images(Figure 2) and in light microscopy images of cells stained with safranin (not shown)
These inclusions were shown by both ESEM and TEM EDS to contain phosphorus with generally small amounts of potassium calcium magnesium sulfur and iron the lattertwo elements likely being due to the close proximity of the P-rich inclusions to sulfurglobules and magnetosomes (Figures 1 and 6) A very large calcium peak was observedwhen these large inclusions were analyzed in the TEM (Figure 6a) The samples shown inFigures 6a and 6d were collected at different times of the year which may explain some ofthe differences observed in these spectra Other EDS data from the ESEM (not shown) hadhigher calcium peaks than those shown in Figure 6d indicating that relative proportions ofP and Ca in P-rich inclusions vary even within the same sample
The inclusions were electron dense in TEM and had a mottled appearance due toablation by the electron beam Similar inclusions have been described for uncultured MCfrom the Itaupo Lagoon near Rio de Janeiro Brazil (Lins and Farina 1999) and from MillPond New Hampshire (Blakemore 1982) Like the structures in ARB-1 polyP granules aretranslucent in visible light (Spring et al 1994) dark (electron dense) in the TEM and oftenhave a mottled appearance (Harold 1966) The P-rich inclusions in ARB-1 cells stained
400 B L Cox et al
with methylene blue (Murray and Robinson 1994) which is suggestive of the presence ofpolyP but not con rmatory
PolyP is composed of linear chains of tetrahedral phosphate residues linked throughcommon oxygen atoms by phosphoanhydrid e bonds At physiological pH each unit carriesa monovalent negative charge giving polyP the property of having a large cation exchangecapacity The binding energy of polyP allows it to sequester Ca2C and Mg2C (Reusch 2000)consistent with the Ca2C and Mg2C signals observed in the EDS spectra of the ARB-1inclusions (Figure 6a 6d) The binding of these metals to the polyP granules contributesto their high electron density PolyP is often associated with polyhydroxybutyrat e (PHB)PHBs solvate (dissolve) cations and associate withpolyP to form Ca2C selective ion channelsacross plasma membranes (Reusch 2000)
The presence of polyP bodies is common in bacteria often resulting as a bacterialresponse to stress and they may play a role in cell motility quorum sensing and bio lmformation (Kornberg and Fraley 2000) Sulfate starvation is known to lead to rapid andmassive accumulation of polyP in some bacteria (Stanier et al 1986) When cells regainaccess to sulfate 32P uptake studies showed that PO3iexcl
4 present in the polyP bodies wasincorporated into nucleic acids PolyP is synthesized from ATP by the enzyme polyP kinasePolyP granules have been reported to be associated with nucleic acids (Harold 1966) butit is not clear whether the association occurs in vivo or after extraction This associationwith nucleic acids would be consistent with staining patterns seen when ARB-1 cells wereexamined with a uorescent livedead stain (nucleic acid stain) displaying a double-lobedstructure in all of the cells (Figure 9) This dumbbell shape appears to extend inside andacross the two phosphorus inclusions
The physiological function of the P inclusions in ARB-1 is not yet known but fromother cell systems several possibilities can be considered These inclusions might serveas a phosphorus reserve for the biosynthesis of nucleic acids and phospholipids underconditions of phosphorus starvation (Harold 1966) Still another possibility is buoyancyregulation Cells can vary their density by as much as 7 by incorporating substances intoinclusions (Guerrero et al 1985) A more speculative possibility is that ARB-1 cells canderive energy from polyP Acinetobacter johnsonii 210a is known to couple the excretion ofa metal phosphate chelate and HC generating a proton motive force (VanVeen et al 1994)A johnsonii takes up P from the aquatic environment by using two Pi transport systemsand stores it as intracellular granules of polyP
Sulfur (S) Inclusions
Cells of ARB-1 contained 5 to 10 spherical S-rich inclusions (Figures 1d) These rangedin size from approximately 100 to 700 nm although most of the S-rich inclusions in asingle cell were close to the same size and were located at one end of the cell oppositethe side where the magnetosomes were located and agellar bundles attached Like theP-rich inclusions the S-rich inclusions appeared to be enclosed in a coating or membrane(Figure 1d) EDS analysis from the ESEM showed these structures contained mostly S withsome carbon and oxygen (Figure 6e) EDS analysis from the TEM also showed S but evenhigher counts for Ca and P (Figure 6b) Beam spreading for the two methods is within aradius of 200 nm The X-rays produced by the 200-keV TEM beam sample the entire verticalextent of the cell unlike the 20 keV SEM beam which loses most of its energy Thereforethe EDS spectra collected at the TEM likely have sampled elements from the cytoplasmand cell membrane components both above and below the inclusion as well as from theP-rich inclusions nearby Other explanations are that hydrated sulfur inclusions changedtheir composition upon dehydration or that these two samples collected at different times
Organization of Inclusions in Magnetococci 401
FIGURE 9 Fluorescence micrograph of an ARB-1 cell suspension treated with Bacliteviability dyes showing that the nucleic acid staining pattern corresponds with the locationof the P-rich inclusions Baclite stains nucleic acids with cytonine (all cells) and propidiumiodate (dead or damaged cells) Bright double-lobed patterns are from ldquoliverdquo cells
of the day and the year may re ect a seasonal diurnal or cell cycle variability in the relativeproportions of the elements
Sulfur inclusions have been found in other uncultured magnetotactic bacteria includ-ing Magnetobacterium bavaricum (Spring et al 1993) Bilophococcus magnetotacticum(Moench 1988) and others (Iida and Akai 1996) and cultured species including the vibrioidstrains MV-1 and MV-2 the coccus strain MC-1 and the spirillum strain MV-4 (Bazylinskiand Frankel 2000) However only in Bilophococcus magnetotacticum was there a similarpolarization of sulfur-containing inclusions from the clustered magnetosomes within thecell (Moench 1988)
It seems likely that ARB-1 oxidizes H2S or other reduced sulfur compounds as anenergy source depositing ldquoelementalrdquo sulfur in intracellular inclusions like many othersul de-oxidizing bacteria including the anaerobic phototrophic purple sulfur bacteria suchas Chromatium vinosum and Thiocapsa roseopersicina the nonsulfur bacteria Beggiatoaspp and Thiothrix spp (Shively 1974 Friedrich 1998) and the magnetotactic bacteriamentioned before In these organisms the presence of sulfur globules is transitory andtakes place when the growth medium contains sul de andor thiosulfate When the sul deis completely utilized the sulfur is converted to sulfate (Stanier et al 1986)
Elemental S in sulfur globules is typically in a liquid state and analysis of in vivoglobules from some members of the purple and green sulfur bacteria by X-ray absorptionnear edge spectroscopy (Prange et al 1999) indicated the sulfur was in a chain structure
402 B L Cox et al
FIGURE 10 Phylogenetic position of six SSU rRNA gene clones related to previouslystudied MC This phylogenetic tree was rooted with Escherichia coli and constructed bythe neighbor joining method with the Kimura 2-parameter model for nucleotide change Amask including all nonambiguously aligned positions for 1427 nucleotides spanning nearlythe full length of the SSU rRNA gene was included Bootstrap values (100 replications)generated by the neighbor joining method are shown above relevant nodes and bootstrapvalues generated by maximum-parsimony analysis are shown below relevant nodes Onlybootstrap values above 50 are shown GenBank accession numbers are MC-CS92 X81182MC-pa119 Y13214 MC-CS81 X81184 MC-TB12 X81183 MC-MP17 X80996 MC-PA11 Y13212 MC12 L06456 partial sequence extended with data from MC-1 genomeRhizobium leguminosarum U31074 Agrobacterium tumefaciens D13294 Rhodobactersphaeroides X53854 Paracoccus denitri cans X59159 Zymomonas mobilis AF281034Sphingomonas paucimobilis D84528 Caulobacter crescentus M83799 Magnetospiril-lum magnetotacticum strain AMB1 D17514 S88429 Magnetospirillum magnetotacticumM10110 Rhodospirillum fulvum M59065 Magnetospirillum gryphiswaldense Y10109Magnetic Vibrio MV-1 LO6455 Oceanospirillum pusillum AB000482 Rhodospirillumrubrum D30778 Roseococcus thiosulfatophilus X72908 Escherichia coli J01695
Organization of Inclusions in Magnetococci 403
and was possibly complexed with water and organic residues If the S globules in ARB-1cells are analogous to those in the purple sulfur bacteria Chromatium vinosum and Thio-capsa roseopersicina they may contain a signi cant protein content Sulfur globules fromthe anaerobic phototrophs C vinosum and T roseopersicina were found to contain upto 52 protein that formed an envelope enclosing the S globules (Brune 1995) Theseproteins were isolated and analyzed and found to be glycine and tyrosine-rich structuralproteins which were homologous for the two species suggesting they are highly con-served (Brune 1995) Phylogenetically C vinosum and T roseopersicina are members ofthe deg -Proteobacteria whereas MC are members of the reg-Proteobacteria To our knowledgemagnetotactic bacteria may be the only members of the reg-Proteobacteria that make internalsulfur globules
Phylogenetic Af liation
We wished to detemine whether ARB-1 was a single species or a mixed population of MCToward that end 45 SSU rRNA genes were cloned from partially puri ed ARB-1 cells30 of which displayed similar restriction fragment length patterns (RFLP) to previouslycharacterized MC (data not shown) The nding that 15 patterns were not related to pre-viously characterized MC was not surprising given that the initial sample was not a pureculture but rather a population magnetically enriched from a sample originating from arelatively large vertical column containing both sediments and overlying water Fourteenof the MC-related clones representing different RFLP pattern groups were chosen for par-tial sequencing 10 of these sequences showed high similarity to previously characterizedMC Six of these sequences were phylogenetically distinct and were fully bidirectionallysequenced
ARB-1 MC SSU rRNA genes formed a monophyletic cluster related to but dis-tinct from other MC (Figure 10) Within the cluster the similarity between SSU rRNAgene sequences ranged from 97 (MC-H1arb and MC-g6arb) to 92 (MC-h1arb andMC-g2arb) The nearest neighbor to the ARB-1 cluster MC-CS92 was ca 89 similarand the nearest cultured relative MC-1 was ca 88 similar to the ARB-1 cluster As arule of thumb for SSU rRNA genes gt97 similarity is considered the same species andgt93ndash95 similarity is considered the same genus (Stackebrandt and Goebel 1994) Thesedata strongly suggest that ARB-1 represents a population of different diverse MC Therewas signi cant variation within the ARB-1 cluster and therefore we do not consider ARB-1to consist of bacteria of a single species or genus Correlations if any between phylogeneticand phenotypic variations in the ARB-1 cells have yet to be determined
Summary
We have partially characterized a population of magnetotactic cocci (MC) referred to asARB-1 which was particularly abundant in iron-rich sediments of a small eutrophic lake andmorphologically and structurally similar to an uncultured species of MC called Bilophococ-cus magnetotacticus The latter organism was found in large numbers in a sewage treatmentpond Cells of ARB-1 had a relatively wide size range of 1 to 4 sup1m They were motileGram-negative and bilophotrichously agellated Cells of this uncultured magnetotacticcoccus contained three types of intracellular inclusions that appeared to be in a consistentarrangement within the cell The magnetosomes contained crystals of magnetite and wereeither in chains andor polar clumps at the end of the cell where the agellar bundles werelocated Large P-rich bodies occupied the majority of the cell volume and appeared to sepa-rate the S-containing inclusions from the magnetosomes The presence of these large P-rich
404 B L Cox et al
inclusions might be an indication of cell stress or may be important in cell buoyancy or inenergy generation The presence of S-rich inclusions located at the end of the cell oppositethe magnetosomes strongly suggests that ARB-1 cells are sul de-oxidizing bacteria andthat the S might be useful as a stored electron donor when cells travel from sul de-rich tosul de-depleted zones in the water column The wide range of the cell sizes the differentmagnetosome arrangements and the large number of different retrieved 16S rRNA genesequences suggest that ARB-1 represents a population of at least several different MC 16SrRNA sequences obtained from cells of ARB-1 appeared to form a monophyletic clusterrelated to but distinct from other uncultured and cultured MC Future work using uores-cent oligonucleotide probes microscopy and physiological measurements should clarifyrelationships between phylogeny cell morphology and organization of inclusions
References
Balkwill DL Maratea D Blakemore RP 1980 Ultrastructure of a magnetotactic spirillum J Bacteriol141(3)1399ndash1408
Bazylinski DA Frankel RB 2000 Biologically controlled mineralization of magnetic iron mineralsby magnetotactic bacteria In Lovley DR editor Environmental microbe-metal interactionsWashington DC ASM Press p 109ndash143
Bazylinski DA Frankel RB Jannasch HW 1988 Anaerobic magnetite production by a marinemagnetotactic bacterium Nature 334518ndash519
Beveridge T 1999 Structures of gram-negative cell walls and their derived membrane vesiclesJ Bacteriol 181(6)4725ndash4733
Beveridge TJ Popkin TJ Cole RM 1994 Electron microscopy In Gerhardt P editor Methods forgeneral and molecular bacteriology Washington DC American Society for Microbiology p 25
Blakemore R 1975 Magnetotactic bacteria Science 190377ndash379Blakemore RP 1982 Magnetotactic bacteria Ann Rev Microbiol 36217ndash238Blakemore RP Maratea D Wolfe RS 1979 Isolation and pure culture of a freshwater magnetic
spirillum in chemically de ned medium J Bacteriol 140(2)720ndash729Blakemore RP Short KA Bazylinski DA Rosenblatt C Frankel RB 1985 Microaerobic conditions
are required for magnetite formation within Aquaspirillum magnetotacticum Geomicrobiol J4(1)53ndash71
Brune DC 1995 Isolation and characterization of sulfur globule proteins from Chromatium vinosumand Thiocapsa roseopersicina Arch Microbiol 163(1)391ndash399
Butler RF Banerjee SK 1975 Theoretical single-domain grain size range in magnetite and titano-magnetite J Geophys Res 804049ndash4058
Buwalda JP 1940 Geology of the Raymond Basin report to Pasadena Water DepartmentCrook R Allen CR Kamb B Payne CM Procter RJ 1987 Recent reverse faulting in the Transverse
Ranges California Pasadena CA US Geol Surv Prof Paper 133927ndash63Devouard B Posfai M Hua X Bazylinski DA Frankel RB Buseck PR 1998 Magnetite from
magnetotactic bacteria size distribution and twinning Am Mineral 831387ndash1398Frankel RB Bazylinski DA Johnson MS Taylor BL 1997 Magneto-Aerotaxis in marine coccoid
bacteria Biophys J 73994ndash1000Friedrich CG 1998 Physiology and genetics of sulfur-oxidizing bacteria Adv Microbial Physiol
39235ndash289Giovannoni SJ 1991 The polymerase chain reaction In Stackebrandt E Goodfellow M editors
Sequencing and hybridization techniques in bacterial systematics New York John Wiley andSons p 177ndash201
Guerrero R Pedros-Alio C Schmidt TM Mas J 1985 A survey of buoyant density of microorganismsin pure cultures and natural samples Microbiologia Sept 1(1ndash2)53ndash65
Harold FM 1966 Inorganic polyphosphates in biology structure metabolism and function BacteriolRev 30(4)772ndash794
Organization of Inclusions in Magnetococci 405
Iida A Akai J 1996 Crystalline sulfur inclusions in magnetotactic bacteria Sci Rep Nigata Univ SerE 1135ndash42
Kornberg A Fraley CD 2000 Inorganic polyphosphate a molecular fossil come to life ASM News66(5)275ndash280
Lins U Farina M 1999 Phosphorus-rich granules in uncultured magnetotactic bacteria FEMSMicrobiol Lett 172(2)23ndash28
Mann S Sparks NHC Frankel RB Bazylinski DA Jannasch HW 1990 Biomineralization of ferri-magnetic greigite (Fe3O4 and iron pyrite (FeS2 ) in a magnetotactic bacterium Nature 343258ndash
261Matsunaga T Sakaguchi T Tadokoro F 1991 Magnetite formation by a magnetic bacterium capable
of growing aerobically Appl Microbiol Biotechnol 35651ndash655Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993a Electron microscopy
study of magnetosomes in a cultured coccoid magnetotactic bacterium Proc Roy Soc Lond B251231ndash236
Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993b Electron microscopystudy of magnetosomes in two cultured vibrioid magnetotactic bacteria Proc Roy Soc Lond B251237ndash242
Moench TT 1988 Bilophococcus magnetotacticus gen nov sp nov a motile magnetic coccus Antvan Leeuwenhoek 54483ndash496
Moench TT Konetzka WA 1978 A novel method for the isolation and study of a magnetotacticbacterium Arch Microbiol 119203ndash212
Murray RGE Robinow CF 1994 Optical Microscopy In Gerhardt P editor Methods for generaland molecular bacteriology Washington DC American Society for Microbiology p 35ndash36
Prange A Arzberger I Engemann C Modrow H Schumann O Truper HG Steudel R Dahl C andHormes J 1999 In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteriaby X-ray absorption near edge spectroscopy Biochim Biophys Acta 1428446ndash454
Rappe MS Kemp PF Giovannoni SJ 1997 Phylogenetic diversity of marine coastal picoplankton16S rRNA genes cloned from the continental shelf off Cape Hatteras North Carolina LimnolOceanogr 42811ndash826
Reusch RN 2000 Transmembrane ion transport by polyphosphatepoly-(R)-3-hydroxybutyratecomplexes Biochemistry (Moscow) 65(3)335ndash352
Sakaguchi T Burgess JG Matsunaga T 1993 Magnetite formation by a sulphate-reducing bacteriumNature 36547ndash49
Schleifer KHSchuler D Spring S Weizenegger M Amann R 1991 The genus Magnetospirillum gennov description of Magnetospirillum gryphiswaldense sp nov and transfer of Aquaspirillummagnetotacticum to Magnetospirillum magnetotacticum comb nov System Appl Microbiol14379ndash385
Schuler D 1999 Formation of magnetosomes in magnetotactic bacteria J Mol Microbiol Biotechnol1(1)79ndash86
Schuler D Spring S Bazylinski DA 1999 Improved technique for the isolation of magnetotactic spir-illa from a freshwater sediment and their phylogenetic characterization System Appl Microbiol22466ndash471
Shively JM 1974 Inclusion bodies of prokaryotes Ann Rev Microbiol 28167ndash187Spring S Amann R Ludwig W Schleifer KH van Gemerden H Petersen N 1993 Dominating role
of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment ApplEnviron Microbiol 59(8)2397ndash2403
Spring S Amann R Ludwig W Schleifer KH Schuler D Poralla K Petersen N 1994 Phylogeneticanalysis of uncultured magnetotactic baceria from the Alpha-subclass of Proteobacteria SystemAppl Microbiol 17501ndash508
Stackebrandt E Goebel BM 1994 Taxonomic note a place for DNA-DNA reassociation and 16SrRNA sequence analysis in the present species de nition in bacteriology Int J System Bacteriol44846ndash849
Stanier RY Ingraham JL Wheelis ML Painter PR 1986 The microbial world 5th edition EnglewoodCliffs NJ Prentice Hall p 179
406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35
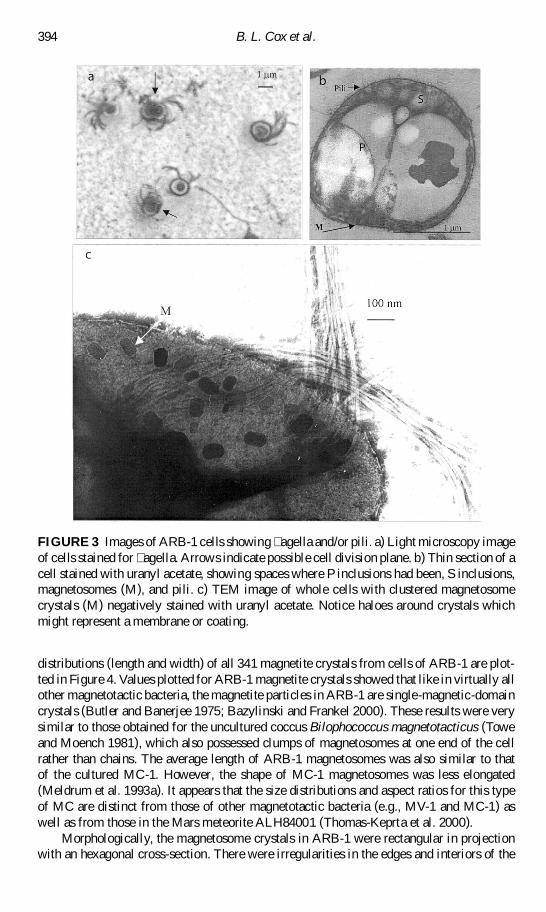
394 B L Cox et al
FIGURE 3 Images of ARB-1 cells showing agella andor pili a) Light microscopy imageof cells stained for agella Arrows indicate possible cell division plane b) Thin section of acell stained with uranyl acetate showing spaces where P inclusions had been S inclusionsmagnetosomes (M) and pili c) TEM image of whole cells with clustered magnetosomecrystals (M) negatively stained with uranyl acetate Notice haloes around crystals whichmight represent a membrane or coating
distributions (length and width) of all 341 magnetite crystals from cells of ARB-1 are plot-ted in Figure 4 Values plotted for ARB-1 magnetite crystals showed that like in virtually allother magnetotactic bacteria the magnetite particles in ARB-1 are single-magnetic-domaincrystals (Butler and Banerjee 1975 Bazylinski and Frankel 2000) These results were verysimilar to those obtained for the uncultured coccus Bilophococcus magnetotacticus (Toweand Moench 1981) which also possessed clumps of magnetosomes at one end of the cellrather than chains The average length of ARB-1 magnetosomes was also similar to thatof the cultured MC-1 However the shape of MC-1 magnetosomes was less elongated(Meldrum et al 1993a) It appears that the size distributions and aspect ratios for this typeof MC are distinct from those of other magnetotactic bacteria (eg MV-1 and MC-1) aswell as from those in the Mars meteorite ALH84001 (Thomas-Keprta et al 2000)
Morphologically the magnetosome crystals in ARB-1 were rectangular in projectionwith an hexagonal cross-section There were irregularities in the edges and interiors of the
FIG
UR
E4
Dis
trib
utio
nof
leng
ths
and
wid
ths
of34
1m
agne
tite
crys
tals
from
clus
ters
and
chai
nsin
AR
B-1
cells
a) F
requ
ency
asa
func
tion
ofle
ngth
b)
Freq
uenc
yas
afu
nctio
nof
wid
thc
) Wid
thas
afu
nctio
nof
leng
th
395
396 B L Cox et al
FIGURE 5 TEM image of a thin section of a cell of ARB-1 stained with uranyl acetateshowing magnetosomes before a) and after b) beam damage leaving crystal ldquodoubletsrdquo b)Notice disappearance of electron dense material in b)
crystals (Figure 3) The edges were often rounded and the particles occasionally con-stricted slightly in the middle The crystals sometimes had a mottled appearance and mayhave had fringes around the edges Once during TEM analysis of thin sections (80 kV)magnetosome crystals sustained signi cant beam damage after several minutes of viewingat a magni cation of 100000 (Figure 5) The crystals appeared to have broken in halfpossibly along twin planes in the (111) direction or along ldquowaistedrdquo regions where thecrystals showed some constriction and there appeared to be a loss of material Beam dam-age has been previously observed during TEM of greigite (Fe3S4) magnetosome particlesbut not in magnetite crystals (D Bazylinski H Vali and J Kirschvink personal commu-nications January 2001) except when viewed as whole mounts at 400 kV (Taylor et al2001) Taylor et al (2001) speculated that water vapor from the cells underwent radiolysisin the electron-beam producing a plasma that rapidly melted the magnetosomes
EDS analyses with both TEM and ESEM of the magnetosome particles (Figure 6c 6f)showed the particles were iron-rich with some extraneous signals including sulfur (Figure 6)A strong sulfur signal would be expected from EDS analyses of the magnetic mineral greigitealso known to be produced by some magnetotactic bacteria (Mann et al 1990) In ARB-1however these extraneous signals are likely due to the sulfur- and phosphorus-containinginclusions also present in the cell SAED patterns obtained from the crystals (Figure 7)and lattice fringes from high resolution TEM with spacings of 048 nm elongated in (111)direction (Figure 7) were consistent with the mineral magnetite
Magnetosomes within cells of ARB-1 were found in three different arrangements1) as a polar cluster forming a sort of ldquocaprdquo usually at the agellar end of the cell 2) in partialor complete chains at the same end of the cell or 3) a combination of both arrangementsThe polar clusters appear to be disorganized in many images (eg Figures 1 2 and 3)However the attraction of these cells to a magnet indicates they have permanent magneticdipole moments implying that they are not disorganized We are currently pursuing thisquestion of whether or not the clusters are disorganized
In cells having both chains and clusters it is dif cult to determine whether chainswere forming from the clustered crystals or whether the polar clusters of particles werefrom a chain that was disrupted during dehydration of the cell during preparation for TEMexamination However polar clusters of crystals were observed in ESEM images of many
Organization of Inclusions in Magnetococci 397
FIGURE 6 EDS spectra from TEM a) b) c) and ESEM d) e) f) showing elementalcomposition of P-rich a) d) and S-rich b) e) inclusions and magnetosomes c) f) Elementalcompositions of clustered magnetosomes were identical to those of magnetosomes in chainsAll y-axes are linear except for a) and b) which are log scales Noise in f ) is due to scattering
hydrated cells (Figure 1d) suggesting that they were not caused by dehydration In manyof the cells viewed by TEM it seemed that the chains tended to occur in smaller morespherical cells while clumps of magnetosomes occurred more frequently in elongatedcells In order to see if there was any relationship between cell size and shape and the threetypes of magnetosome arrangements the size and aspect ratio for cells with these differentmagnetosome arrangements were measured (Figure 8) The cell size distribution appearedto be narrower for cells with chains than for the cells with disorganized crystals Cells withchains were predominantly smaller than 25 sup1m (Figure 8a) The aspect ratios of the cellswith chains only was smaller than that of cells with disorganized magnetosomes and wasshifted toward spherical aspect ratios (Figure 8b)
One explanation for these observations is that there are different stages of magnetosomeformation in a single species of MC The disorganized magnetosomes might form near thecell membrane between the agellar bundles and then extend into chains Alternativelymagnetosomes might rst form as chains in smaller cells At a later stage as cells increasein size the membranes linking the chains dissolve or become stressed and the magneto-somes somehow migrate to the agellar end of the cell Because magnetosomes appear tobe anchored to the inner surface of the cell membrane in virtually all species examined(Bazylinski and Frankel 2000) and are never free- oating in the cytoplasm migration ofmagnetosomes through the cytoplasm seems unlikely Instead we are presently designinghybridization experiments to address the hypothesis that the relationships between cell sizeshape and magnetosome arrangement correlate with the genotypical variations in ARB-1described later Spring et al (1994) analyzed communities of MC found in freshwater lakesin Germany using uorescent oligonucleotide probes They found different genotypes ofMC (similarity below 95) but these genotypes were not phenotypically expressed asdifferent magnetosome organizations but as preferences for different concentrations of O2
398 B L Cox et al
FIGURE 7 Evidence supporting magnetite as the magnetosome mineral in ARB-1 cellsa) Image of a single crystal of magnetite within a cell in an unstained glutaraldehyde- xedwhole cell mount taken with High Resolution TEM by H Vali b) Part of the magnetitecrystal shown in a) magni ed to show the distance between lattice fringes c) Selectedarea electron diffraction (SAED) of a group of magnetosomes in a cell from an un xedunstained whole cell mount showing the indices of some of the diffraction spots
Therefore we might expect no correlation between magnetite organization and phyloge-netic diversity of ARB-1 cells but perhaps a correlation between oxygen preference andphylogeny
A third possibility is that chains and clumps of magnetosomes form as a result ofdifferent environmental conditions Since our sampling methods were not selective as tothe cellsrsquo location in the vertical chemical (eg oxygen) and redox gradients that are sure to
Organization of Inclusions in Magnetococci 399
FIGURE 8 Relationships between organization of magnetosomes and cell size a) and cellaspect ratio b) Measurements were taken from both TEM and ESEM images Solid patternis for cells with clustered magnetosomes crossed pattern is for cells with magnetosomesin chains only Patterned bars originate at the top of solid bars
exist at the site the environmental conditions of our samples represent a relatively large rangeof oxygen and perhaps other chemical concentrations as well as redox conditions Magneticbacteria in culture are known to vary the size and number of magnetosomes in responseto the media composition (Meldrum et al 1993a) and particularly oxygen concentration(Blakemore et al 1985) Another possibility is that the clustered magnetosomes are formedby a different biological mechanism and serve a different function than magnetosomes inchains
Phosphorus (P) Inclusions
The two very large inclusion bodies seen in each cell always occurred in pairs in cellsand seemed to extend to the cell membrane and to be in contact with each other Thus themaximum dimension of the P-rich inclusions was the same as the minimum dimension ofthe cell and their combined width was approximately half of the long dimension of the cellIn ESEM images the two P-containing inclusions appeared to be attached in the middle insome cells and separated in others (Figure 1d) The P-containing inclusions appeared to beself-contained and perhaps bounded by some sort of coating or membrane in TEM images(Figure 2) and in light microscopy images of cells stained with safranin (not shown)
These inclusions were shown by both ESEM and TEM EDS to contain phosphorus with generally small amounts of potassium calcium magnesium sulfur and iron the lattertwo elements likely being due to the close proximity of the P-rich inclusions to sulfurglobules and magnetosomes (Figures 1 and 6) A very large calcium peak was observedwhen these large inclusions were analyzed in the TEM (Figure 6a) The samples shown inFigures 6a and 6d were collected at different times of the year which may explain some ofthe differences observed in these spectra Other EDS data from the ESEM (not shown) hadhigher calcium peaks than those shown in Figure 6d indicating that relative proportions ofP and Ca in P-rich inclusions vary even within the same sample
The inclusions were electron dense in TEM and had a mottled appearance due toablation by the electron beam Similar inclusions have been described for uncultured MCfrom the Itaupo Lagoon near Rio de Janeiro Brazil (Lins and Farina 1999) and from MillPond New Hampshire (Blakemore 1982) Like the structures in ARB-1 polyP granules aretranslucent in visible light (Spring et al 1994) dark (electron dense) in the TEM and oftenhave a mottled appearance (Harold 1966) The P-rich inclusions in ARB-1 cells stained
400 B L Cox et al
with methylene blue (Murray and Robinson 1994) which is suggestive of the presence ofpolyP but not con rmatory
PolyP is composed of linear chains of tetrahedral phosphate residues linked throughcommon oxygen atoms by phosphoanhydrid e bonds At physiological pH each unit carriesa monovalent negative charge giving polyP the property of having a large cation exchangecapacity The binding energy of polyP allows it to sequester Ca2C and Mg2C (Reusch 2000)consistent with the Ca2C and Mg2C signals observed in the EDS spectra of the ARB-1inclusions (Figure 6a 6d) The binding of these metals to the polyP granules contributesto their high electron density PolyP is often associated with polyhydroxybutyrat e (PHB)PHBs solvate (dissolve) cations and associate withpolyP to form Ca2C selective ion channelsacross plasma membranes (Reusch 2000)
The presence of polyP bodies is common in bacteria often resulting as a bacterialresponse to stress and they may play a role in cell motility quorum sensing and bio lmformation (Kornberg and Fraley 2000) Sulfate starvation is known to lead to rapid andmassive accumulation of polyP in some bacteria (Stanier et al 1986) When cells regainaccess to sulfate 32P uptake studies showed that PO3iexcl
4 present in the polyP bodies wasincorporated into nucleic acids PolyP is synthesized from ATP by the enzyme polyP kinasePolyP granules have been reported to be associated with nucleic acids (Harold 1966) butit is not clear whether the association occurs in vivo or after extraction This associationwith nucleic acids would be consistent with staining patterns seen when ARB-1 cells wereexamined with a uorescent livedead stain (nucleic acid stain) displaying a double-lobedstructure in all of the cells (Figure 9) This dumbbell shape appears to extend inside andacross the two phosphorus inclusions
The physiological function of the P inclusions in ARB-1 is not yet known but fromother cell systems several possibilities can be considered These inclusions might serveas a phosphorus reserve for the biosynthesis of nucleic acids and phospholipids underconditions of phosphorus starvation (Harold 1966) Still another possibility is buoyancyregulation Cells can vary their density by as much as 7 by incorporating substances intoinclusions (Guerrero et al 1985) A more speculative possibility is that ARB-1 cells canderive energy from polyP Acinetobacter johnsonii 210a is known to couple the excretion ofa metal phosphate chelate and HC generating a proton motive force (VanVeen et al 1994)A johnsonii takes up P from the aquatic environment by using two Pi transport systemsand stores it as intracellular granules of polyP
Sulfur (S) Inclusions
Cells of ARB-1 contained 5 to 10 spherical S-rich inclusions (Figures 1d) These rangedin size from approximately 100 to 700 nm although most of the S-rich inclusions in asingle cell were close to the same size and were located at one end of the cell oppositethe side where the magnetosomes were located and agellar bundles attached Like theP-rich inclusions the S-rich inclusions appeared to be enclosed in a coating or membrane(Figure 1d) EDS analysis from the ESEM showed these structures contained mostly S withsome carbon and oxygen (Figure 6e) EDS analysis from the TEM also showed S but evenhigher counts for Ca and P (Figure 6b) Beam spreading for the two methods is within aradius of 200 nm The X-rays produced by the 200-keV TEM beam sample the entire verticalextent of the cell unlike the 20 keV SEM beam which loses most of its energy Thereforethe EDS spectra collected at the TEM likely have sampled elements from the cytoplasmand cell membrane components both above and below the inclusion as well as from theP-rich inclusions nearby Other explanations are that hydrated sulfur inclusions changedtheir composition upon dehydration or that these two samples collected at different times
Organization of Inclusions in Magnetococci 401
FIGURE 9 Fluorescence micrograph of an ARB-1 cell suspension treated with Bacliteviability dyes showing that the nucleic acid staining pattern corresponds with the locationof the P-rich inclusions Baclite stains nucleic acids with cytonine (all cells) and propidiumiodate (dead or damaged cells) Bright double-lobed patterns are from ldquoliverdquo cells
of the day and the year may re ect a seasonal diurnal or cell cycle variability in the relativeproportions of the elements
Sulfur inclusions have been found in other uncultured magnetotactic bacteria includ-ing Magnetobacterium bavaricum (Spring et al 1993) Bilophococcus magnetotacticum(Moench 1988) and others (Iida and Akai 1996) and cultured species including the vibrioidstrains MV-1 and MV-2 the coccus strain MC-1 and the spirillum strain MV-4 (Bazylinskiand Frankel 2000) However only in Bilophococcus magnetotacticum was there a similarpolarization of sulfur-containing inclusions from the clustered magnetosomes within thecell (Moench 1988)
It seems likely that ARB-1 oxidizes H2S or other reduced sulfur compounds as anenergy source depositing ldquoelementalrdquo sulfur in intracellular inclusions like many othersul de-oxidizing bacteria including the anaerobic phototrophic purple sulfur bacteria suchas Chromatium vinosum and Thiocapsa roseopersicina the nonsulfur bacteria Beggiatoaspp and Thiothrix spp (Shively 1974 Friedrich 1998) and the magnetotactic bacteriamentioned before In these organisms the presence of sulfur globules is transitory andtakes place when the growth medium contains sul de andor thiosulfate When the sul deis completely utilized the sulfur is converted to sulfate (Stanier et al 1986)
Elemental S in sulfur globules is typically in a liquid state and analysis of in vivoglobules from some members of the purple and green sulfur bacteria by X-ray absorptionnear edge spectroscopy (Prange et al 1999) indicated the sulfur was in a chain structure
402 B L Cox et al
FIGURE 10 Phylogenetic position of six SSU rRNA gene clones related to previouslystudied MC This phylogenetic tree was rooted with Escherichia coli and constructed bythe neighbor joining method with the Kimura 2-parameter model for nucleotide change Amask including all nonambiguously aligned positions for 1427 nucleotides spanning nearlythe full length of the SSU rRNA gene was included Bootstrap values (100 replications)generated by the neighbor joining method are shown above relevant nodes and bootstrapvalues generated by maximum-parsimony analysis are shown below relevant nodes Onlybootstrap values above 50 are shown GenBank accession numbers are MC-CS92 X81182MC-pa119 Y13214 MC-CS81 X81184 MC-TB12 X81183 MC-MP17 X80996 MC-PA11 Y13212 MC12 L06456 partial sequence extended with data from MC-1 genomeRhizobium leguminosarum U31074 Agrobacterium tumefaciens D13294 Rhodobactersphaeroides X53854 Paracoccus denitri cans X59159 Zymomonas mobilis AF281034Sphingomonas paucimobilis D84528 Caulobacter crescentus M83799 Magnetospiril-lum magnetotacticum strain AMB1 D17514 S88429 Magnetospirillum magnetotacticumM10110 Rhodospirillum fulvum M59065 Magnetospirillum gryphiswaldense Y10109Magnetic Vibrio MV-1 LO6455 Oceanospirillum pusillum AB000482 Rhodospirillumrubrum D30778 Roseococcus thiosulfatophilus X72908 Escherichia coli J01695
Organization of Inclusions in Magnetococci 403
and was possibly complexed with water and organic residues If the S globules in ARB-1cells are analogous to those in the purple sulfur bacteria Chromatium vinosum and Thio-capsa roseopersicina they may contain a signi cant protein content Sulfur globules fromthe anaerobic phototrophs C vinosum and T roseopersicina were found to contain upto 52 protein that formed an envelope enclosing the S globules (Brune 1995) Theseproteins were isolated and analyzed and found to be glycine and tyrosine-rich structuralproteins which were homologous for the two species suggesting they are highly con-served (Brune 1995) Phylogenetically C vinosum and T roseopersicina are members ofthe deg -Proteobacteria whereas MC are members of the reg-Proteobacteria To our knowledgemagnetotactic bacteria may be the only members of the reg-Proteobacteria that make internalsulfur globules
Phylogenetic Af liation
We wished to detemine whether ARB-1 was a single species or a mixed population of MCToward that end 45 SSU rRNA genes were cloned from partially puri ed ARB-1 cells30 of which displayed similar restriction fragment length patterns (RFLP) to previouslycharacterized MC (data not shown) The nding that 15 patterns were not related to pre-viously characterized MC was not surprising given that the initial sample was not a pureculture but rather a population magnetically enriched from a sample originating from arelatively large vertical column containing both sediments and overlying water Fourteenof the MC-related clones representing different RFLP pattern groups were chosen for par-tial sequencing 10 of these sequences showed high similarity to previously characterizedMC Six of these sequences were phylogenetically distinct and were fully bidirectionallysequenced
ARB-1 MC SSU rRNA genes formed a monophyletic cluster related to but dis-tinct from other MC (Figure 10) Within the cluster the similarity between SSU rRNAgene sequences ranged from 97 (MC-H1arb and MC-g6arb) to 92 (MC-h1arb andMC-g2arb) The nearest neighbor to the ARB-1 cluster MC-CS92 was ca 89 similarand the nearest cultured relative MC-1 was ca 88 similar to the ARB-1 cluster As arule of thumb for SSU rRNA genes gt97 similarity is considered the same species andgt93ndash95 similarity is considered the same genus (Stackebrandt and Goebel 1994) Thesedata strongly suggest that ARB-1 represents a population of different diverse MC Therewas signi cant variation within the ARB-1 cluster and therefore we do not consider ARB-1to consist of bacteria of a single species or genus Correlations if any between phylogeneticand phenotypic variations in the ARB-1 cells have yet to be determined
Summary
We have partially characterized a population of magnetotactic cocci (MC) referred to asARB-1 which was particularly abundant in iron-rich sediments of a small eutrophic lake andmorphologically and structurally similar to an uncultured species of MC called Bilophococ-cus magnetotacticus The latter organism was found in large numbers in a sewage treatmentpond Cells of ARB-1 had a relatively wide size range of 1 to 4 sup1m They were motileGram-negative and bilophotrichously agellated Cells of this uncultured magnetotacticcoccus contained three types of intracellular inclusions that appeared to be in a consistentarrangement within the cell The magnetosomes contained crystals of magnetite and wereeither in chains andor polar clumps at the end of the cell where the agellar bundles werelocated Large P-rich bodies occupied the majority of the cell volume and appeared to sepa-rate the S-containing inclusions from the magnetosomes The presence of these large P-rich
404 B L Cox et al
inclusions might be an indication of cell stress or may be important in cell buoyancy or inenergy generation The presence of S-rich inclusions located at the end of the cell oppositethe magnetosomes strongly suggests that ARB-1 cells are sul de-oxidizing bacteria andthat the S might be useful as a stored electron donor when cells travel from sul de-rich tosul de-depleted zones in the water column The wide range of the cell sizes the differentmagnetosome arrangements and the large number of different retrieved 16S rRNA genesequences suggest that ARB-1 represents a population of at least several different MC 16SrRNA sequences obtained from cells of ARB-1 appeared to form a monophyletic clusterrelated to but distinct from other uncultured and cultured MC Future work using uores-cent oligonucleotide probes microscopy and physiological measurements should clarifyrelationships between phylogeny cell morphology and organization of inclusions
References
Balkwill DL Maratea D Blakemore RP 1980 Ultrastructure of a magnetotactic spirillum J Bacteriol141(3)1399ndash1408
Bazylinski DA Frankel RB 2000 Biologically controlled mineralization of magnetic iron mineralsby magnetotactic bacteria In Lovley DR editor Environmental microbe-metal interactionsWashington DC ASM Press p 109ndash143
Bazylinski DA Frankel RB Jannasch HW 1988 Anaerobic magnetite production by a marinemagnetotactic bacterium Nature 334518ndash519
Beveridge T 1999 Structures of gram-negative cell walls and their derived membrane vesiclesJ Bacteriol 181(6)4725ndash4733
Beveridge TJ Popkin TJ Cole RM 1994 Electron microscopy In Gerhardt P editor Methods forgeneral and molecular bacteriology Washington DC American Society for Microbiology p 25
Blakemore R 1975 Magnetotactic bacteria Science 190377ndash379Blakemore RP 1982 Magnetotactic bacteria Ann Rev Microbiol 36217ndash238Blakemore RP Maratea D Wolfe RS 1979 Isolation and pure culture of a freshwater magnetic
spirillum in chemically de ned medium J Bacteriol 140(2)720ndash729Blakemore RP Short KA Bazylinski DA Rosenblatt C Frankel RB 1985 Microaerobic conditions
are required for magnetite formation within Aquaspirillum magnetotacticum Geomicrobiol J4(1)53ndash71
Brune DC 1995 Isolation and characterization of sulfur globule proteins from Chromatium vinosumand Thiocapsa roseopersicina Arch Microbiol 163(1)391ndash399
Butler RF Banerjee SK 1975 Theoretical single-domain grain size range in magnetite and titano-magnetite J Geophys Res 804049ndash4058
Buwalda JP 1940 Geology of the Raymond Basin report to Pasadena Water DepartmentCrook R Allen CR Kamb B Payne CM Procter RJ 1987 Recent reverse faulting in the Transverse
Ranges California Pasadena CA US Geol Surv Prof Paper 133927ndash63Devouard B Posfai M Hua X Bazylinski DA Frankel RB Buseck PR 1998 Magnetite from
magnetotactic bacteria size distribution and twinning Am Mineral 831387ndash1398Frankel RB Bazylinski DA Johnson MS Taylor BL 1997 Magneto-Aerotaxis in marine coccoid
bacteria Biophys J 73994ndash1000Friedrich CG 1998 Physiology and genetics of sulfur-oxidizing bacteria Adv Microbial Physiol
39235ndash289Giovannoni SJ 1991 The polymerase chain reaction In Stackebrandt E Goodfellow M editors
Sequencing and hybridization techniques in bacterial systematics New York John Wiley andSons p 177ndash201
Guerrero R Pedros-Alio C Schmidt TM Mas J 1985 A survey of buoyant density of microorganismsin pure cultures and natural samples Microbiologia Sept 1(1ndash2)53ndash65
Harold FM 1966 Inorganic polyphosphates in biology structure metabolism and function BacteriolRev 30(4)772ndash794
Organization of Inclusions in Magnetococci 405
Iida A Akai J 1996 Crystalline sulfur inclusions in magnetotactic bacteria Sci Rep Nigata Univ SerE 1135ndash42
Kornberg A Fraley CD 2000 Inorganic polyphosphate a molecular fossil come to life ASM News66(5)275ndash280
Lins U Farina M 1999 Phosphorus-rich granules in uncultured magnetotactic bacteria FEMSMicrobiol Lett 172(2)23ndash28
Mann S Sparks NHC Frankel RB Bazylinski DA Jannasch HW 1990 Biomineralization of ferri-magnetic greigite (Fe3O4 and iron pyrite (FeS2 ) in a magnetotactic bacterium Nature 343258ndash
261Matsunaga T Sakaguchi T Tadokoro F 1991 Magnetite formation by a magnetic bacterium capable
of growing aerobically Appl Microbiol Biotechnol 35651ndash655Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993a Electron microscopy
study of magnetosomes in a cultured coccoid magnetotactic bacterium Proc Roy Soc Lond B251231ndash236
Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993b Electron microscopystudy of magnetosomes in two cultured vibrioid magnetotactic bacteria Proc Roy Soc Lond B251237ndash242
Moench TT 1988 Bilophococcus magnetotacticus gen nov sp nov a motile magnetic coccus Antvan Leeuwenhoek 54483ndash496
Moench TT Konetzka WA 1978 A novel method for the isolation and study of a magnetotacticbacterium Arch Microbiol 119203ndash212
Murray RGE Robinow CF 1994 Optical Microscopy In Gerhardt P editor Methods for generaland molecular bacteriology Washington DC American Society for Microbiology p 35ndash36
Prange A Arzberger I Engemann C Modrow H Schumann O Truper HG Steudel R Dahl C andHormes J 1999 In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteriaby X-ray absorption near edge spectroscopy Biochim Biophys Acta 1428446ndash454
Rappe MS Kemp PF Giovannoni SJ 1997 Phylogenetic diversity of marine coastal picoplankton16S rRNA genes cloned from the continental shelf off Cape Hatteras North Carolina LimnolOceanogr 42811ndash826
Reusch RN 2000 Transmembrane ion transport by polyphosphatepoly-(R)-3-hydroxybutyratecomplexes Biochemistry (Moscow) 65(3)335ndash352
Sakaguchi T Burgess JG Matsunaga T 1993 Magnetite formation by a sulphate-reducing bacteriumNature 36547ndash49
Schleifer KHSchuler D Spring S Weizenegger M Amann R 1991 The genus Magnetospirillum gennov description of Magnetospirillum gryphiswaldense sp nov and transfer of Aquaspirillummagnetotacticum to Magnetospirillum magnetotacticum comb nov System Appl Microbiol14379ndash385
Schuler D 1999 Formation of magnetosomes in magnetotactic bacteria J Mol Microbiol Biotechnol1(1)79ndash86
Schuler D Spring S Bazylinski DA 1999 Improved technique for the isolation of magnetotactic spir-illa from a freshwater sediment and their phylogenetic characterization System Appl Microbiol22466ndash471
Shively JM 1974 Inclusion bodies of prokaryotes Ann Rev Microbiol 28167ndash187Spring S Amann R Ludwig W Schleifer KH van Gemerden H Petersen N 1993 Dominating role
of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment ApplEnviron Microbiol 59(8)2397ndash2403
Spring S Amann R Ludwig W Schleifer KH Schuler D Poralla K Petersen N 1994 Phylogeneticanalysis of uncultured magnetotactic baceria from the Alpha-subclass of Proteobacteria SystemAppl Microbiol 17501ndash508
Stackebrandt E Goebel BM 1994 Taxonomic note a place for DNA-DNA reassociation and 16SrRNA sequence analysis in the present species de nition in bacteriology Int J System Bacteriol44846ndash849
Stanier RY Ingraham JL Wheelis ML Painter PR 1986 The microbial world 5th edition EnglewoodCliffs NJ Prentice Hall p 179
406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35
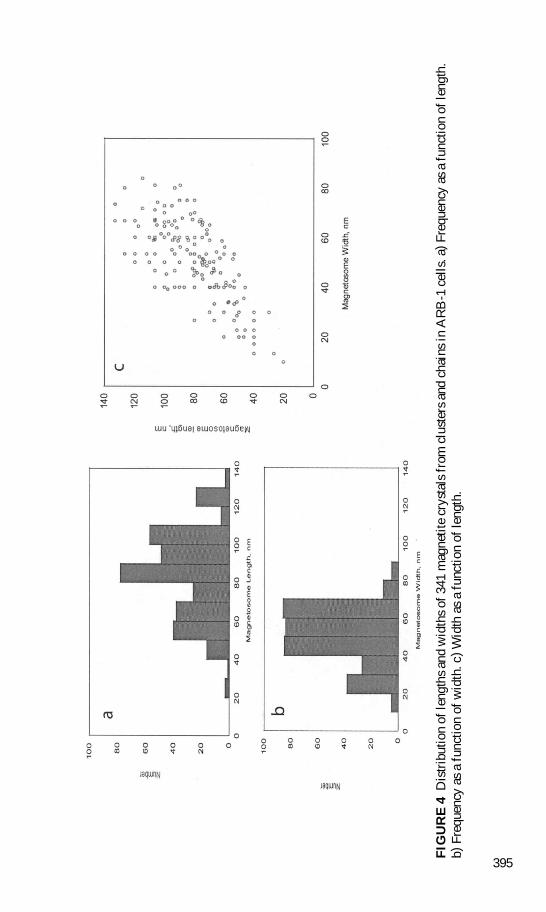
FIG
UR
E4
Dis
trib
utio
nof
leng
ths
and
wid
ths
of34
1m
agne
tite
crys
tals
from
clus
ters
and
chai
nsin
AR
B-1
cells
a) F
requ
ency
asa
func
tion
ofle
ngth
b)
Freq
uenc
yas
afu
nctio
nof
wid
thc
) Wid
thas
afu
nctio
nof
leng
th
395
396 B L Cox et al
FIGURE 5 TEM image of a thin section of a cell of ARB-1 stained with uranyl acetateshowing magnetosomes before a) and after b) beam damage leaving crystal ldquodoubletsrdquo b)Notice disappearance of electron dense material in b)
crystals (Figure 3) The edges were often rounded and the particles occasionally con-stricted slightly in the middle The crystals sometimes had a mottled appearance and mayhave had fringes around the edges Once during TEM analysis of thin sections (80 kV)magnetosome crystals sustained signi cant beam damage after several minutes of viewingat a magni cation of 100000 (Figure 5) The crystals appeared to have broken in halfpossibly along twin planes in the (111) direction or along ldquowaistedrdquo regions where thecrystals showed some constriction and there appeared to be a loss of material Beam dam-age has been previously observed during TEM of greigite (Fe3S4) magnetosome particlesbut not in magnetite crystals (D Bazylinski H Vali and J Kirschvink personal commu-nications January 2001) except when viewed as whole mounts at 400 kV (Taylor et al2001) Taylor et al (2001) speculated that water vapor from the cells underwent radiolysisin the electron-beam producing a plasma that rapidly melted the magnetosomes
EDS analyses with both TEM and ESEM of the magnetosome particles (Figure 6c 6f)showed the particles were iron-rich with some extraneous signals including sulfur (Figure 6)A strong sulfur signal would be expected from EDS analyses of the magnetic mineral greigitealso known to be produced by some magnetotactic bacteria (Mann et al 1990) In ARB-1however these extraneous signals are likely due to the sulfur- and phosphorus-containinginclusions also present in the cell SAED patterns obtained from the crystals (Figure 7)and lattice fringes from high resolution TEM with spacings of 048 nm elongated in (111)direction (Figure 7) were consistent with the mineral magnetite
Magnetosomes within cells of ARB-1 were found in three different arrangements1) as a polar cluster forming a sort of ldquocaprdquo usually at the agellar end of the cell 2) in partialor complete chains at the same end of the cell or 3) a combination of both arrangementsThe polar clusters appear to be disorganized in many images (eg Figures 1 2 and 3)However the attraction of these cells to a magnet indicates they have permanent magneticdipole moments implying that they are not disorganized We are currently pursuing thisquestion of whether or not the clusters are disorganized
In cells having both chains and clusters it is dif cult to determine whether chainswere forming from the clustered crystals or whether the polar clusters of particles werefrom a chain that was disrupted during dehydration of the cell during preparation for TEMexamination However polar clusters of crystals were observed in ESEM images of many
Organization of Inclusions in Magnetococci 397
FIGURE 6 EDS spectra from TEM a) b) c) and ESEM d) e) f) showing elementalcomposition of P-rich a) d) and S-rich b) e) inclusions and magnetosomes c) f) Elementalcompositions of clustered magnetosomes were identical to those of magnetosomes in chainsAll y-axes are linear except for a) and b) which are log scales Noise in f ) is due to scattering
hydrated cells (Figure 1d) suggesting that they were not caused by dehydration In manyof the cells viewed by TEM it seemed that the chains tended to occur in smaller morespherical cells while clumps of magnetosomes occurred more frequently in elongatedcells In order to see if there was any relationship between cell size and shape and the threetypes of magnetosome arrangements the size and aspect ratio for cells with these differentmagnetosome arrangements were measured (Figure 8) The cell size distribution appearedto be narrower for cells with chains than for the cells with disorganized crystals Cells withchains were predominantly smaller than 25 sup1m (Figure 8a) The aspect ratios of the cellswith chains only was smaller than that of cells with disorganized magnetosomes and wasshifted toward spherical aspect ratios (Figure 8b)
One explanation for these observations is that there are different stages of magnetosomeformation in a single species of MC The disorganized magnetosomes might form near thecell membrane between the agellar bundles and then extend into chains Alternativelymagnetosomes might rst form as chains in smaller cells At a later stage as cells increasein size the membranes linking the chains dissolve or become stressed and the magneto-somes somehow migrate to the agellar end of the cell Because magnetosomes appear tobe anchored to the inner surface of the cell membrane in virtually all species examined(Bazylinski and Frankel 2000) and are never free- oating in the cytoplasm migration ofmagnetosomes through the cytoplasm seems unlikely Instead we are presently designinghybridization experiments to address the hypothesis that the relationships between cell sizeshape and magnetosome arrangement correlate with the genotypical variations in ARB-1described later Spring et al (1994) analyzed communities of MC found in freshwater lakesin Germany using uorescent oligonucleotide probes They found different genotypes ofMC (similarity below 95) but these genotypes were not phenotypically expressed asdifferent magnetosome organizations but as preferences for different concentrations of O2
398 B L Cox et al
FIGURE 7 Evidence supporting magnetite as the magnetosome mineral in ARB-1 cellsa) Image of a single crystal of magnetite within a cell in an unstained glutaraldehyde- xedwhole cell mount taken with High Resolution TEM by H Vali b) Part of the magnetitecrystal shown in a) magni ed to show the distance between lattice fringes c) Selectedarea electron diffraction (SAED) of a group of magnetosomes in a cell from an un xedunstained whole cell mount showing the indices of some of the diffraction spots
Therefore we might expect no correlation between magnetite organization and phyloge-netic diversity of ARB-1 cells but perhaps a correlation between oxygen preference andphylogeny
A third possibility is that chains and clumps of magnetosomes form as a result ofdifferent environmental conditions Since our sampling methods were not selective as tothe cellsrsquo location in the vertical chemical (eg oxygen) and redox gradients that are sure to
Organization of Inclusions in Magnetococci 399
FIGURE 8 Relationships between organization of magnetosomes and cell size a) and cellaspect ratio b) Measurements were taken from both TEM and ESEM images Solid patternis for cells with clustered magnetosomes crossed pattern is for cells with magnetosomesin chains only Patterned bars originate at the top of solid bars
exist at the site the environmental conditions of our samples represent a relatively large rangeof oxygen and perhaps other chemical concentrations as well as redox conditions Magneticbacteria in culture are known to vary the size and number of magnetosomes in responseto the media composition (Meldrum et al 1993a) and particularly oxygen concentration(Blakemore et al 1985) Another possibility is that the clustered magnetosomes are formedby a different biological mechanism and serve a different function than magnetosomes inchains
Phosphorus (P) Inclusions
The two very large inclusion bodies seen in each cell always occurred in pairs in cellsand seemed to extend to the cell membrane and to be in contact with each other Thus themaximum dimension of the P-rich inclusions was the same as the minimum dimension ofthe cell and their combined width was approximately half of the long dimension of the cellIn ESEM images the two P-containing inclusions appeared to be attached in the middle insome cells and separated in others (Figure 1d) The P-containing inclusions appeared to beself-contained and perhaps bounded by some sort of coating or membrane in TEM images(Figure 2) and in light microscopy images of cells stained with safranin (not shown)
These inclusions were shown by both ESEM and TEM EDS to contain phosphorus with generally small amounts of potassium calcium magnesium sulfur and iron the lattertwo elements likely being due to the close proximity of the P-rich inclusions to sulfurglobules and magnetosomes (Figures 1 and 6) A very large calcium peak was observedwhen these large inclusions were analyzed in the TEM (Figure 6a) The samples shown inFigures 6a and 6d were collected at different times of the year which may explain some ofthe differences observed in these spectra Other EDS data from the ESEM (not shown) hadhigher calcium peaks than those shown in Figure 6d indicating that relative proportions ofP and Ca in P-rich inclusions vary even within the same sample
The inclusions were electron dense in TEM and had a mottled appearance due toablation by the electron beam Similar inclusions have been described for uncultured MCfrom the Itaupo Lagoon near Rio de Janeiro Brazil (Lins and Farina 1999) and from MillPond New Hampshire (Blakemore 1982) Like the structures in ARB-1 polyP granules aretranslucent in visible light (Spring et al 1994) dark (electron dense) in the TEM and oftenhave a mottled appearance (Harold 1966) The P-rich inclusions in ARB-1 cells stained
400 B L Cox et al
with methylene blue (Murray and Robinson 1994) which is suggestive of the presence ofpolyP but not con rmatory
PolyP is composed of linear chains of tetrahedral phosphate residues linked throughcommon oxygen atoms by phosphoanhydrid e bonds At physiological pH each unit carriesa monovalent negative charge giving polyP the property of having a large cation exchangecapacity The binding energy of polyP allows it to sequester Ca2C and Mg2C (Reusch 2000)consistent with the Ca2C and Mg2C signals observed in the EDS spectra of the ARB-1inclusions (Figure 6a 6d) The binding of these metals to the polyP granules contributesto their high electron density PolyP is often associated with polyhydroxybutyrat e (PHB)PHBs solvate (dissolve) cations and associate withpolyP to form Ca2C selective ion channelsacross plasma membranes (Reusch 2000)
The presence of polyP bodies is common in bacteria often resulting as a bacterialresponse to stress and they may play a role in cell motility quorum sensing and bio lmformation (Kornberg and Fraley 2000) Sulfate starvation is known to lead to rapid andmassive accumulation of polyP in some bacteria (Stanier et al 1986) When cells regainaccess to sulfate 32P uptake studies showed that PO3iexcl
4 present in the polyP bodies wasincorporated into nucleic acids PolyP is synthesized from ATP by the enzyme polyP kinasePolyP granules have been reported to be associated with nucleic acids (Harold 1966) butit is not clear whether the association occurs in vivo or after extraction This associationwith nucleic acids would be consistent with staining patterns seen when ARB-1 cells wereexamined with a uorescent livedead stain (nucleic acid stain) displaying a double-lobedstructure in all of the cells (Figure 9) This dumbbell shape appears to extend inside andacross the two phosphorus inclusions
The physiological function of the P inclusions in ARB-1 is not yet known but fromother cell systems several possibilities can be considered These inclusions might serveas a phosphorus reserve for the biosynthesis of nucleic acids and phospholipids underconditions of phosphorus starvation (Harold 1966) Still another possibility is buoyancyregulation Cells can vary their density by as much as 7 by incorporating substances intoinclusions (Guerrero et al 1985) A more speculative possibility is that ARB-1 cells canderive energy from polyP Acinetobacter johnsonii 210a is known to couple the excretion ofa metal phosphate chelate and HC generating a proton motive force (VanVeen et al 1994)A johnsonii takes up P from the aquatic environment by using two Pi transport systemsand stores it as intracellular granules of polyP
Sulfur (S) Inclusions
Cells of ARB-1 contained 5 to 10 spherical S-rich inclusions (Figures 1d) These rangedin size from approximately 100 to 700 nm although most of the S-rich inclusions in asingle cell were close to the same size and were located at one end of the cell oppositethe side where the magnetosomes were located and agellar bundles attached Like theP-rich inclusions the S-rich inclusions appeared to be enclosed in a coating or membrane(Figure 1d) EDS analysis from the ESEM showed these structures contained mostly S withsome carbon and oxygen (Figure 6e) EDS analysis from the TEM also showed S but evenhigher counts for Ca and P (Figure 6b) Beam spreading for the two methods is within aradius of 200 nm The X-rays produced by the 200-keV TEM beam sample the entire verticalextent of the cell unlike the 20 keV SEM beam which loses most of its energy Thereforethe EDS spectra collected at the TEM likely have sampled elements from the cytoplasmand cell membrane components both above and below the inclusion as well as from theP-rich inclusions nearby Other explanations are that hydrated sulfur inclusions changedtheir composition upon dehydration or that these two samples collected at different times
Organization of Inclusions in Magnetococci 401
FIGURE 9 Fluorescence micrograph of an ARB-1 cell suspension treated with Bacliteviability dyes showing that the nucleic acid staining pattern corresponds with the locationof the P-rich inclusions Baclite stains nucleic acids with cytonine (all cells) and propidiumiodate (dead or damaged cells) Bright double-lobed patterns are from ldquoliverdquo cells
of the day and the year may re ect a seasonal diurnal or cell cycle variability in the relativeproportions of the elements
Sulfur inclusions have been found in other uncultured magnetotactic bacteria includ-ing Magnetobacterium bavaricum (Spring et al 1993) Bilophococcus magnetotacticum(Moench 1988) and others (Iida and Akai 1996) and cultured species including the vibrioidstrains MV-1 and MV-2 the coccus strain MC-1 and the spirillum strain MV-4 (Bazylinskiand Frankel 2000) However only in Bilophococcus magnetotacticum was there a similarpolarization of sulfur-containing inclusions from the clustered magnetosomes within thecell (Moench 1988)
It seems likely that ARB-1 oxidizes H2S or other reduced sulfur compounds as anenergy source depositing ldquoelementalrdquo sulfur in intracellular inclusions like many othersul de-oxidizing bacteria including the anaerobic phototrophic purple sulfur bacteria suchas Chromatium vinosum and Thiocapsa roseopersicina the nonsulfur bacteria Beggiatoaspp and Thiothrix spp (Shively 1974 Friedrich 1998) and the magnetotactic bacteriamentioned before In these organisms the presence of sulfur globules is transitory andtakes place when the growth medium contains sul de andor thiosulfate When the sul deis completely utilized the sulfur is converted to sulfate (Stanier et al 1986)
Elemental S in sulfur globules is typically in a liquid state and analysis of in vivoglobules from some members of the purple and green sulfur bacteria by X-ray absorptionnear edge spectroscopy (Prange et al 1999) indicated the sulfur was in a chain structure
402 B L Cox et al
FIGURE 10 Phylogenetic position of six SSU rRNA gene clones related to previouslystudied MC This phylogenetic tree was rooted with Escherichia coli and constructed bythe neighbor joining method with the Kimura 2-parameter model for nucleotide change Amask including all nonambiguously aligned positions for 1427 nucleotides spanning nearlythe full length of the SSU rRNA gene was included Bootstrap values (100 replications)generated by the neighbor joining method are shown above relevant nodes and bootstrapvalues generated by maximum-parsimony analysis are shown below relevant nodes Onlybootstrap values above 50 are shown GenBank accession numbers are MC-CS92 X81182MC-pa119 Y13214 MC-CS81 X81184 MC-TB12 X81183 MC-MP17 X80996 MC-PA11 Y13212 MC12 L06456 partial sequence extended with data from MC-1 genomeRhizobium leguminosarum U31074 Agrobacterium tumefaciens D13294 Rhodobactersphaeroides X53854 Paracoccus denitri cans X59159 Zymomonas mobilis AF281034Sphingomonas paucimobilis D84528 Caulobacter crescentus M83799 Magnetospiril-lum magnetotacticum strain AMB1 D17514 S88429 Magnetospirillum magnetotacticumM10110 Rhodospirillum fulvum M59065 Magnetospirillum gryphiswaldense Y10109Magnetic Vibrio MV-1 LO6455 Oceanospirillum pusillum AB000482 Rhodospirillumrubrum D30778 Roseococcus thiosulfatophilus X72908 Escherichia coli J01695
Organization of Inclusions in Magnetococci 403
and was possibly complexed with water and organic residues If the S globules in ARB-1cells are analogous to those in the purple sulfur bacteria Chromatium vinosum and Thio-capsa roseopersicina they may contain a signi cant protein content Sulfur globules fromthe anaerobic phototrophs C vinosum and T roseopersicina were found to contain upto 52 protein that formed an envelope enclosing the S globules (Brune 1995) Theseproteins were isolated and analyzed and found to be glycine and tyrosine-rich structuralproteins which were homologous for the two species suggesting they are highly con-served (Brune 1995) Phylogenetically C vinosum and T roseopersicina are members ofthe deg -Proteobacteria whereas MC are members of the reg-Proteobacteria To our knowledgemagnetotactic bacteria may be the only members of the reg-Proteobacteria that make internalsulfur globules
Phylogenetic Af liation
We wished to detemine whether ARB-1 was a single species or a mixed population of MCToward that end 45 SSU rRNA genes were cloned from partially puri ed ARB-1 cells30 of which displayed similar restriction fragment length patterns (RFLP) to previouslycharacterized MC (data not shown) The nding that 15 patterns were not related to pre-viously characterized MC was not surprising given that the initial sample was not a pureculture but rather a population magnetically enriched from a sample originating from arelatively large vertical column containing both sediments and overlying water Fourteenof the MC-related clones representing different RFLP pattern groups were chosen for par-tial sequencing 10 of these sequences showed high similarity to previously characterizedMC Six of these sequences were phylogenetically distinct and were fully bidirectionallysequenced
ARB-1 MC SSU rRNA genes formed a monophyletic cluster related to but dis-tinct from other MC (Figure 10) Within the cluster the similarity between SSU rRNAgene sequences ranged from 97 (MC-H1arb and MC-g6arb) to 92 (MC-h1arb andMC-g2arb) The nearest neighbor to the ARB-1 cluster MC-CS92 was ca 89 similarand the nearest cultured relative MC-1 was ca 88 similar to the ARB-1 cluster As arule of thumb for SSU rRNA genes gt97 similarity is considered the same species andgt93ndash95 similarity is considered the same genus (Stackebrandt and Goebel 1994) Thesedata strongly suggest that ARB-1 represents a population of different diverse MC Therewas signi cant variation within the ARB-1 cluster and therefore we do not consider ARB-1to consist of bacteria of a single species or genus Correlations if any between phylogeneticand phenotypic variations in the ARB-1 cells have yet to be determined
Summary
We have partially characterized a population of magnetotactic cocci (MC) referred to asARB-1 which was particularly abundant in iron-rich sediments of a small eutrophic lake andmorphologically and structurally similar to an uncultured species of MC called Bilophococ-cus magnetotacticus The latter organism was found in large numbers in a sewage treatmentpond Cells of ARB-1 had a relatively wide size range of 1 to 4 sup1m They were motileGram-negative and bilophotrichously agellated Cells of this uncultured magnetotacticcoccus contained three types of intracellular inclusions that appeared to be in a consistentarrangement within the cell The magnetosomes contained crystals of magnetite and wereeither in chains andor polar clumps at the end of the cell where the agellar bundles werelocated Large P-rich bodies occupied the majority of the cell volume and appeared to sepa-rate the S-containing inclusions from the magnetosomes The presence of these large P-rich
404 B L Cox et al
inclusions might be an indication of cell stress or may be important in cell buoyancy or inenergy generation The presence of S-rich inclusions located at the end of the cell oppositethe magnetosomes strongly suggests that ARB-1 cells are sul de-oxidizing bacteria andthat the S might be useful as a stored electron donor when cells travel from sul de-rich tosul de-depleted zones in the water column The wide range of the cell sizes the differentmagnetosome arrangements and the large number of different retrieved 16S rRNA genesequences suggest that ARB-1 represents a population of at least several different MC 16SrRNA sequences obtained from cells of ARB-1 appeared to form a monophyletic clusterrelated to but distinct from other uncultured and cultured MC Future work using uores-cent oligonucleotide probes microscopy and physiological measurements should clarifyrelationships between phylogeny cell morphology and organization of inclusions
References
Balkwill DL Maratea D Blakemore RP 1980 Ultrastructure of a magnetotactic spirillum J Bacteriol141(3)1399ndash1408
Bazylinski DA Frankel RB 2000 Biologically controlled mineralization of magnetic iron mineralsby magnetotactic bacteria In Lovley DR editor Environmental microbe-metal interactionsWashington DC ASM Press p 109ndash143
Bazylinski DA Frankel RB Jannasch HW 1988 Anaerobic magnetite production by a marinemagnetotactic bacterium Nature 334518ndash519
Beveridge T 1999 Structures of gram-negative cell walls and their derived membrane vesiclesJ Bacteriol 181(6)4725ndash4733
Beveridge TJ Popkin TJ Cole RM 1994 Electron microscopy In Gerhardt P editor Methods forgeneral and molecular bacteriology Washington DC American Society for Microbiology p 25
Blakemore R 1975 Magnetotactic bacteria Science 190377ndash379Blakemore RP 1982 Magnetotactic bacteria Ann Rev Microbiol 36217ndash238Blakemore RP Maratea D Wolfe RS 1979 Isolation and pure culture of a freshwater magnetic
spirillum in chemically de ned medium J Bacteriol 140(2)720ndash729Blakemore RP Short KA Bazylinski DA Rosenblatt C Frankel RB 1985 Microaerobic conditions
are required for magnetite formation within Aquaspirillum magnetotacticum Geomicrobiol J4(1)53ndash71
Brune DC 1995 Isolation and characterization of sulfur globule proteins from Chromatium vinosumand Thiocapsa roseopersicina Arch Microbiol 163(1)391ndash399
Butler RF Banerjee SK 1975 Theoretical single-domain grain size range in magnetite and titano-magnetite J Geophys Res 804049ndash4058
Buwalda JP 1940 Geology of the Raymond Basin report to Pasadena Water DepartmentCrook R Allen CR Kamb B Payne CM Procter RJ 1987 Recent reverse faulting in the Transverse
Ranges California Pasadena CA US Geol Surv Prof Paper 133927ndash63Devouard B Posfai M Hua X Bazylinski DA Frankel RB Buseck PR 1998 Magnetite from
magnetotactic bacteria size distribution and twinning Am Mineral 831387ndash1398Frankel RB Bazylinski DA Johnson MS Taylor BL 1997 Magneto-Aerotaxis in marine coccoid
bacteria Biophys J 73994ndash1000Friedrich CG 1998 Physiology and genetics of sulfur-oxidizing bacteria Adv Microbial Physiol
39235ndash289Giovannoni SJ 1991 The polymerase chain reaction In Stackebrandt E Goodfellow M editors
Sequencing and hybridization techniques in bacterial systematics New York John Wiley andSons p 177ndash201
Guerrero R Pedros-Alio C Schmidt TM Mas J 1985 A survey of buoyant density of microorganismsin pure cultures and natural samples Microbiologia Sept 1(1ndash2)53ndash65
Harold FM 1966 Inorganic polyphosphates in biology structure metabolism and function BacteriolRev 30(4)772ndash794
Organization of Inclusions in Magnetococci 405
Iida A Akai J 1996 Crystalline sulfur inclusions in magnetotactic bacteria Sci Rep Nigata Univ SerE 1135ndash42
Kornberg A Fraley CD 2000 Inorganic polyphosphate a molecular fossil come to life ASM News66(5)275ndash280
Lins U Farina M 1999 Phosphorus-rich granules in uncultured magnetotactic bacteria FEMSMicrobiol Lett 172(2)23ndash28
Mann S Sparks NHC Frankel RB Bazylinski DA Jannasch HW 1990 Biomineralization of ferri-magnetic greigite (Fe3O4 and iron pyrite (FeS2 ) in a magnetotactic bacterium Nature 343258ndash
261Matsunaga T Sakaguchi T Tadokoro F 1991 Magnetite formation by a magnetic bacterium capable
of growing aerobically Appl Microbiol Biotechnol 35651ndash655Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993a Electron microscopy
study of magnetosomes in a cultured coccoid magnetotactic bacterium Proc Roy Soc Lond B251231ndash236
Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993b Electron microscopystudy of magnetosomes in two cultured vibrioid magnetotactic bacteria Proc Roy Soc Lond B251237ndash242
Moench TT 1988 Bilophococcus magnetotacticus gen nov sp nov a motile magnetic coccus Antvan Leeuwenhoek 54483ndash496
Moench TT Konetzka WA 1978 A novel method for the isolation and study of a magnetotacticbacterium Arch Microbiol 119203ndash212
Murray RGE Robinow CF 1994 Optical Microscopy In Gerhardt P editor Methods for generaland molecular bacteriology Washington DC American Society for Microbiology p 35ndash36
Prange A Arzberger I Engemann C Modrow H Schumann O Truper HG Steudel R Dahl C andHormes J 1999 In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteriaby X-ray absorption near edge spectroscopy Biochim Biophys Acta 1428446ndash454
Rappe MS Kemp PF Giovannoni SJ 1997 Phylogenetic diversity of marine coastal picoplankton16S rRNA genes cloned from the continental shelf off Cape Hatteras North Carolina LimnolOceanogr 42811ndash826
Reusch RN 2000 Transmembrane ion transport by polyphosphatepoly-(R)-3-hydroxybutyratecomplexes Biochemistry (Moscow) 65(3)335ndash352
Sakaguchi T Burgess JG Matsunaga T 1993 Magnetite formation by a sulphate-reducing bacteriumNature 36547ndash49
Schleifer KHSchuler D Spring S Weizenegger M Amann R 1991 The genus Magnetospirillum gennov description of Magnetospirillum gryphiswaldense sp nov and transfer of Aquaspirillummagnetotacticum to Magnetospirillum magnetotacticum comb nov System Appl Microbiol14379ndash385
Schuler D 1999 Formation of magnetosomes in magnetotactic bacteria J Mol Microbiol Biotechnol1(1)79ndash86
Schuler D Spring S Bazylinski DA 1999 Improved technique for the isolation of magnetotactic spir-illa from a freshwater sediment and their phylogenetic characterization System Appl Microbiol22466ndash471
Shively JM 1974 Inclusion bodies of prokaryotes Ann Rev Microbiol 28167ndash187Spring S Amann R Ludwig W Schleifer KH van Gemerden H Petersen N 1993 Dominating role
of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment ApplEnviron Microbiol 59(8)2397ndash2403
Spring S Amann R Ludwig W Schleifer KH Schuler D Poralla K Petersen N 1994 Phylogeneticanalysis of uncultured magnetotactic baceria from the Alpha-subclass of Proteobacteria SystemAppl Microbiol 17501ndash508
Stackebrandt E Goebel BM 1994 Taxonomic note a place for DNA-DNA reassociation and 16SrRNA sequence analysis in the present species de nition in bacteriology Int J System Bacteriol44846ndash849
Stanier RY Ingraham JL Wheelis ML Painter PR 1986 The microbial world 5th edition EnglewoodCliffs NJ Prentice Hall p 179
406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35

396 B L Cox et al
FIGURE 5 TEM image of a thin section of a cell of ARB-1 stained with uranyl acetateshowing magnetosomes before a) and after b) beam damage leaving crystal ldquodoubletsrdquo b)Notice disappearance of electron dense material in b)
crystals (Figure 3) The edges were often rounded and the particles occasionally con-stricted slightly in the middle The crystals sometimes had a mottled appearance and mayhave had fringes around the edges Once during TEM analysis of thin sections (80 kV)magnetosome crystals sustained signi cant beam damage after several minutes of viewingat a magni cation of 100000 (Figure 5) The crystals appeared to have broken in halfpossibly along twin planes in the (111) direction or along ldquowaistedrdquo regions where thecrystals showed some constriction and there appeared to be a loss of material Beam dam-age has been previously observed during TEM of greigite (Fe3S4) magnetosome particlesbut not in magnetite crystals (D Bazylinski H Vali and J Kirschvink personal commu-nications January 2001) except when viewed as whole mounts at 400 kV (Taylor et al2001) Taylor et al (2001) speculated that water vapor from the cells underwent radiolysisin the electron-beam producing a plasma that rapidly melted the magnetosomes
EDS analyses with both TEM and ESEM of the magnetosome particles (Figure 6c 6f)showed the particles were iron-rich with some extraneous signals including sulfur (Figure 6)A strong sulfur signal would be expected from EDS analyses of the magnetic mineral greigitealso known to be produced by some magnetotactic bacteria (Mann et al 1990) In ARB-1however these extraneous signals are likely due to the sulfur- and phosphorus-containinginclusions also present in the cell SAED patterns obtained from the crystals (Figure 7)and lattice fringes from high resolution TEM with spacings of 048 nm elongated in (111)direction (Figure 7) were consistent with the mineral magnetite
Magnetosomes within cells of ARB-1 were found in three different arrangements1) as a polar cluster forming a sort of ldquocaprdquo usually at the agellar end of the cell 2) in partialor complete chains at the same end of the cell or 3) a combination of both arrangementsThe polar clusters appear to be disorganized in many images (eg Figures 1 2 and 3)However the attraction of these cells to a magnet indicates they have permanent magneticdipole moments implying that they are not disorganized We are currently pursuing thisquestion of whether or not the clusters are disorganized
In cells having both chains and clusters it is dif cult to determine whether chainswere forming from the clustered crystals or whether the polar clusters of particles werefrom a chain that was disrupted during dehydration of the cell during preparation for TEMexamination However polar clusters of crystals were observed in ESEM images of many
Organization of Inclusions in Magnetococci 397
FIGURE 6 EDS spectra from TEM a) b) c) and ESEM d) e) f) showing elementalcomposition of P-rich a) d) and S-rich b) e) inclusions and magnetosomes c) f) Elementalcompositions of clustered magnetosomes were identical to those of magnetosomes in chainsAll y-axes are linear except for a) and b) which are log scales Noise in f ) is due to scattering
hydrated cells (Figure 1d) suggesting that they were not caused by dehydration In manyof the cells viewed by TEM it seemed that the chains tended to occur in smaller morespherical cells while clumps of magnetosomes occurred more frequently in elongatedcells In order to see if there was any relationship between cell size and shape and the threetypes of magnetosome arrangements the size and aspect ratio for cells with these differentmagnetosome arrangements were measured (Figure 8) The cell size distribution appearedto be narrower for cells with chains than for the cells with disorganized crystals Cells withchains were predominantly smaller than 25 sup1m (Figure 8a) The aspect ratios of the cellswith chains only was smaller than that of cells with disorganized magnetosomes and wasshifted toward spherical aspect ratios (Figure 8b)
One explanation for these observations is that there are different stages of magnetosomeformation in a single species of MC The disorganized magnetosomes might form near thecell membrane between the agellar bundles and then extend into chains Alternativelymagnetosomes might rst form as chains in smaller cells At a later stage as cells increasein size the membranes linking the chains dissolve or become stressed and the magneto-somes somehow migrate to the agellar end of the cell Because magnetosomes appear tobe anchored to the inner surface of the cell membrane in virtually all species examined(Bazylinski and Frankel 2000) and are never free- oating in the cytoplasm migration ofmagnetosomes through the cytoplasm seems unlikely Instead we are presently designinghybridization experiments to address the hypothesis that the relationships between cell sizeshape and magnetosome arrangement correlate with the genotypical variations in ARB-1described later Spring et al (1994) analyzed communities of MC found in freshwater lakesin Germany using uorescent oligonucleotide probes They found different genotypes ofMC (similarity below 95) but these genotypes were not phenotypically expressed asdifferent magnetosome organizations but as preferences for different concentrations of O2
398 B L Cox et al
FIGURE 7 Evidence supporting magnetite as the magnetosome mineral in ARB-1 cellsa) Image of a single crystal of magnetite within a cell in an unstained glutaraldehyde- xedwhole cell mount taken with High Resolution TEM by H Vali b) Part of the magnetitecrystal shown in a) magni ed to show the distance between lattice fringes c) Selectedarea electron diffraction (SAED) of a group of magnetosomes in a cell from an un xedunstained whole cell mount showing the indices of some of the diffraction spots
Therefore we might expect no correlation between magnetite organization and phyloge-netic diversity of ARB-1 cells but perhaps a correlation between oxygen preference andphylogeny
A third possibility is that chains and clumps of magnetosomes form as a result ofdifferent environmental conditions Since our sampling methods were not selective as tothe cellsrsquo location in the vertical chemical (eg oxygen) and redox gradients that are sure to
Organization of Inclusions in Magnetococci 399
FIGURE 8 Relationships between organization of magnetosomes and cell size a) and cellaspect ratio b) Measurements were taken from both TEM and ESEM images Solid patternis for cells with clustered magnetosomes crossed pattern is for cells with magnetosomesin chains only Patterned bars originate at the top of solid bars
exist at the site the environmental conditions of our samples represent a relatively large rangeof oxygen and perhaps other chemical concentrations as well as redox conditions Magneticbacteria in culture are known to vary the size and number of magnetosomes in responseto the media composition (Meldrum et al 1993a) and particularly oxygen concentration(Blakemore et al 1985) Another possibility is that the clustered magnetosomes are formedby a different biological mechanism and serve a different function than magnetosomes inchains
Phosphorus (P) Inclusions
The two very large inclusion bodies seen in each cell always occurred in pairs in cellsand seemed to extend to the cell membrane and to be in contact with each other Thus themaximum dimension of the P-rich inclusions was the same as the minimum dimension ofthe cell and their combined width was approximately half of the long dimension of the cellIn ESEM images the two P-containing inclusions appeared to be attached in the middle insome cells and separated in others (Figure 1d) The P-containing inclusions appeared to beself-contained and perhaps bounded by some sort of coating or membrane in TEM images(Figure 2) and in light microscopy images of cells stained with safranin (not shown)
These inclusions were shown by both ESEM and TEM EDS to contain phosphorus with generally small amounts of potassium calcium magnesium sulfur and iron the lattertwo elements likely being due to the close proximity of the P-rich inclusions to sulfurglobules and magnetosomes (Figures 1 and 6) A very large calcium peak was observedwhen these large inclusions were analyzed in the TEM (Figure 6a) The samples shown inFigures 6a and 6d were collected at different times of the year which may explain some ofthe differences observed in these spectra Other EDS data from the ESEM (not shown) hadhigher calcium peaks than those shown in Figure 6d indicating that relative proportions ofP and Ca in P-rich inclusions vary even within the same sample
The inclusions were electron dense in TEM and had a mottled appearance due toablation by the electron beam Similar inclusions have been described for uncultured MCfrom the Itaupo Lagoon near Rio de Janeiro Brazil (Lins and Farina 1999) and from MillPond New Hampshire (Blakemore 1982) Like the structures in ARB-1 polyP granules aretranslucent in visible light (Spring et al 1994) dark (electron dense) in the TEM and oftenhave a mottled appearance (Harold 1966) The P-rich inclusions in ARB-1 cells stained
400 B L Cox et al
with methylene blue (Murray and Robinson 1994) which is suggestive of the presence ofpolyP but not con rmatory
PolyP is composed of linear chains of tetrahedral phosphate residues linked throughcommon oxygen atoms by phosphoanhydrid e bonds At physiological pH each unit carriesa monovalent negative charge giving polyP the property of having a large cation exchangecapacity The binding energy of polyP allows it to sequester Ca2C and Mg2C (Reusch 2000)consistent with the Ca2C and Mg2C signals observed in the EDS spectra of the ARB-1inclusions (Figure 6a 6d) The binding of these metals to the polyP granules contributesto their high electron density PolyP is often associated with polyhydroxybutyrat e (PHB)PHBs solvate (dissolve) cations and associate withpolyP to form Ca2C selective ion channelsacross plasma membranes (Reusch 2000)
The presence of polyP bodies is common in bacteria often resulting as a bacterialresponse to stress and they may play a role in cell motility quorum sensing and bio lmformation (Kornberg and Fraley 2000) Sulfate starvation is known to lead to rapid andmassive accumulation of polyP in some bacteria (Stanier et al 1986) When cells regainaccess to sulfate 32P uptake studies showed that PO3iexcl
4 present in the polyP bodies wasincorporated into nucleic acids PolyP is synthesized from ATP by the enzyme polyP kinasePolyP granules have been reported to be associated with nucleic acids (Harold 1966) butit is not clear whether the association occurs in vivo or after extraction This associationwith nucleic acids would be consistent with staining patterns seen when ARB-1 cells wereexamined with a uorescent livedead stain (nucleic acid stain) displaying a double-lobedstructure in all of the cells (Figure 9) This dumbbell shape appears to extend inside andacross the two phosphorus inclusions
The physiological function of the P inclusions in ARB-1 is not yet known but fromother cell systems several possibilities can be considered These inclusions might serveas a phosphorus reserve for the biosynthesis of nucleic acids and phospholipids underconditions of phosphorus starvation (Harold 1966) Still another possibility is buoyancyregulation Cells can vary their density by as much as 7 by incorporating substances intoinclusions (Guerrero et al 1985) A more speculative possibility is that ARB-1 cells canderive energy from polyP Acinetobacter johnsonii 210a is known to couple the excretion ofa metal phosphate chelate and HC generating a proton motive force (VanVeen et al 1994)A johnsonii takes up P from the aquatic environment by using two Pi transport systemsand stores it as intracellular granules of polyP
Sulfur (S) Inclusions
Cells of ARB-1 contained 5 to 10 spherical S-rich inclusions (Figures 1d) These rangedin size from approximately 100 to 700 nm although most of the S-rich inclusions in asingle cell were close to the same size and were located at one end of the cell oppositethe side where the magnetosomes were located and agellar bundles attached Like theP-rich inclusions the S-rich inclusions appeared to be enclosed in a coating or membrane(Figure 1d) EDS analysis from the ESEM showed these structures contained mostly S withsome carbon and oxygen (Figure 6e) EDS analysis from the TEM also showed S but evenhigher counts for Ca and P (Figure 6b) Beam spreading for the two methods is within aradius of 200 nm The X-rays produced by the 200-keV TEM beam sample the entire verticalextent of the cell unlike the 20 keV SEM beam which loses most of its energy Thereforethe EDS spectra collected at the TEM likely have sampled elements from the cytoplasmand cell membrane components both above and below the inclusion as well as from theP-rich inclusions nearby Other explanations are that hydrated sulfur inclusions changedtheir composition upon dehydration or that these two samples collected at different times
Organization of Inclusions in Magnetococci 401
FIGURE 9 Fluorescence micrograph of an ARB-1 cell suspension treated with Bacliteviability dyes showing that the nucleic acid staining pattern corresponds with the locationof the P-rich inclusions Baclite stains nucleic acids with cytonine (all cells) and propidiumiodate (dead or damaged cells) Bright double-lobed patterns are from ldquoliverdquo cells
of the day and the year may re ect a seasonal diurnal or cell cycle variability in the relativeproportions of the elements
Sulfur inclusions have been found in other uncultured magnetotactic bacteria includ-ing Magnetobacterium bavaricum (Spring et al 1993) Bilophococcus magnetotacticum(Moench 1988) and others (Iida and Akai 1996) and cultured species including the vibrioidstrains MV-1 and MV-2 the coccus strain MC-1 and the spirillum strain MV-4 (Bazylinskiand Frankel 2000) However only in Bilophococcus magnetotacticum was there a similarpolarization of sulfur-containing inclusions from the clustered magnetosomes within thecell (Moench 1988)
It seems likely that ARB-1 oxidizes H2S or other reduced sulfur compounds as anenergy source depositing ldquoelementalrdquo sulfur in intracellular inclusions like many othersul de-oxidizing bacteria including the anaerobic phototrophic purple sulfur bacteria suchas Chromatium vinosum and Thiocapsa roseopersicina the nonsulfur bacteria Beggiatoaspp and Thiothrix spp (Shively 1974 Friedrich 1998) and the magnetotactic bacteriamentioned before In these organisms the presence of sulfur globules is transitory andtakes place when the growth medium contains sul de andor thiosulfate When the sul deis completely utilized the sulfur is converted to sulfate (Stanier et al 1986)
Elemental S in sulfur globules is typically in a liquid state and analysis of in vivoglobules from some members of the purple and green sulfur bacteria by X-ray absorptionnear edge spectroscopy (Prange et al 1999) indicated the sulfur was in a chain structure
402 B L Cox et al
FIGURE 10 Phylogenetic position of six SSU rRNA gene clones related to previouslystudied MC This phylogenetic tree was rooted with Escherichia coli and constructed bythe neighbor joining method with the Kimura 2-parameter model for nucleotide change Amask including all nonambiguously aligned positions for 1427 nucleotides spanning nearlythe full length of the SSU rRNA gene was included Bootstrap values (100 replications)generated by the neighbor joining method are shown above relevant nodes and bootstrapvalues generated by maximum-parsimony analysis are shown below relevant nodes Onlybootstrap values above 50 are shown GenBank accession numbers are MC-CS92 X81182MC-pa119 Y13214 MC-CS81 X81184 MC-TB12 X81183 MC-MP17 X80996 MC-PA11 Y13212 MC12 L06456 partial sequence extended with data from MC-1 genomeRhizobium leguminosarum U31074 Agrobacterium tumefaciens D13294 Rhodobactersphaeroides X53854 Paracoccus denitri cans X59159 Zymomonas mobilis AF281034Sphingomonas paucimobilis D84528 Caulobacter crescentus M83799 Magnetospiril-lum magnetotacticum strain AMB1 D17514 S88429 Magnetospirillum magnetotacticumM10110 Rhodospirillum fulvum M59065 Magnetospirillum gryphiswaldense Y10109Magnetic Vibrio MV-1 LO6455 Oceanospirillum pusillum AB000482 Rhodospirillumrubrum D30778 Roseococcus thiosulfatophilus X72908 Escherichia coli J01695
Organization of Inclusions in Magnetococci 403
and was possibly complexed with water and organic residues If the S globules in ARB-1cells are analogous to those in the purple sulfur bacteria Chromatium vinosum and Thio-capsa roseopersicina they may contain a signi cant protein content Sulfur globules fromthe anaerobic phototrophs C vinosum and T roseopersicina were found to contain upto 52 protein that formed an envelope enclosing the S globules (Brune 1995) Theseproteins were isolated and analyzed and found to be glycine and tyrosine-rich structuralproteins which were homologous for the two species suggesting they are highly con-served (Brune 1995) Phylogenetically C vinosum and T roseopersicina are members ofthe deg -Proteobacteria whereas MC are members of the reg-Proteobacteria To our knowledgemagnetotactic bacteria may be the only members of the reg-Proteobacteria that make internalsulfur globules
Phylogenetic Af liation
We wished to detemine whether ARB-1 was a single species or a mixed population of MCToward that end 45 SSU rRNA genes were cloned from partially puri ed ARB-1 cells30 of which displayed similar restriction fragment length patterns (RFLP) to previouslycharacterized MC (data not shown) The nding that 15 patterns were not related to pre-viously characterized MC was not surprising given that the initial sample was not a pureculture but rather a population magnetically enriched from a sample originating from arelatively large vertical column containing both sediments and overlying water Fourteenof the MC-related clones representing different RFLP pattern groups were chosen for par-tial sequencing 10 of these sequences showed high similarity to previously characterizedMC Six of these sequences were phylogenetically distinct and were fully bidirectionallysequenced
ARB-1 MC SSU rRNA genes formed a monophyletic cluster related to but dis-tinct from other MC (Figure 10) Within the cluster the similarity between SSU rRNAgene sequences ranged from 97 (MC-H1arb and MC-g6arb) to 92 (MC-h1arb andMC-g2arb) The nearest neighbor to the ARB-1 cluster MC-CS92 was ca 89 similarand the nearest cultured relative MC-1 was ca 88 similar to the ARB-1 cluster As arule of thumb for SSU rRNA genes gt97 similarity is considered the same species andgt93ndash95 similarity is considered the same genus (Stackebrandt and Goebel 1994) Thesedata strongly suggest that ARB-1 represents a population of different diverse MC Therewas signi cant variation within the ARB-1 cluster and therefore we do not consider ARB-1to consist of bacteria of a single species or genus Correlations if any between phylogeneticand phenotypic variations in the ARB-1 cells have yet to be determined
Summary
We have partially characterized a population of magnetotactic cocci (MC) referred to asARB-1 which was particularly abundant in iron-rich sediments of a small eutrophic lake andmorphologically and structurally similar to an uncultured species of MC called Bilophococ-cus magnetotacticus The latter organism was found in large numbers in a sewage treatmentpond Cells of ARB-1 had a relatively wide size range of 1 to 4 sup1m They were motileGram-negative and bilophotrichously agellated Cells of this uncultured magnetotacticcoccus contained three types of intracellular inclusions that appeared to be in a consistentarrangement within the cell The magnetosomes contained crystals of magnetite and wereeither in chains andor polar clumps at the end of the cell where the agellar bundles werelocated Large P-rich bodies occupied the majority of the cell volume and appeared to sepa-rate the S-containing inclusions from the magnetosomes The presence of these large P-rich
404 B L Cox et al
inclusions might be an indication of cell stress or may be important in cell buoyancy or inenergy generation The presence of S-rich inclusions located at the end of the cell oppositethe magnetosomes strongly suggests that ARB-1 cells are sul de-oxidizing bacteria andthat the S might be useful as a stored electron donor when cells travel from sul de-rich tosul de-depleted zones in the water column The wide range of the cell sizes the differentmagnetosome arrangements and the large number of different retrieved 16S rRNA genesequences suggest that ARB-1 represents a population of at least several different MC 16SrRNA sequences obtained from cells of ARB-1 appeared to form a monophyletic clusterrelated to but distinct from other uncultured and cultured MC Future work using uores-cent oligonucleotide probes microscopy and physiological measurements should clarifyrelationships between phylogeny cell morphology and organization of inclusions
References
Balkwill DL Maratea D Blakemore RP 1980 Ultrastructure of a magnetotactic spirillum J Bacteriol141(3)1399ndash1408
Bazylinski DA Frankel RB 2000 Biologically controlled mineralization of magnetic iron mineralsby magnetotactic bacteria In Lovley DR editor Environmental microbe-metal interactionsWashington DC ASM Press p 109ndash143
Bazylinski DA Frankel RB Jannasch HW 1988 Anaerobic magnetite production by a marinemagnetotactic bacterium Nature 334518ndash519
Beveridge T 1999 Structures of gram-negative cell walls and their derived membrane vesiclesJ Bacteriol 181(6)4725ndash4733
Beveridge TJ Popkin TJ Cole RM 1994 Electron microscopy In Gerhardt P editor Methods forgeneral and molecular bacteriology Washington DC American Society for Microbiology p 25
Blakemore R 1975 Magnetotactic bacteria Science 190377ndash379Blakemore RP 1982 Magnetotactic bacteria Ann Rev Microbiol 36217ndash238Blakemore RP Maratea D Wolfe RS 1979 Isolation and pure culture of a freshwater magnetic
spirillum in chemically de ned medium J Bacteriol 140(2)720ndash729Blakemore RP Short KA Bazylinski DA Rosenblatt C Frankel RB 1985 Microaerobic conditions
are required for magnetite formation within Aquaspirillum magnetotacticum Geomicrobiol J4(1)53ndash71
Brune DC 1995 Isolation and characterization of sulfur globule proteins from Chromatium vinosumand Thiocapsa roseopersicina Arch Microbiol 163(1)391ndash399
Butler RF Banerjee SK 1975 Theoretical single-domain grain size range in magnetite and titano-magnetite J Geophys Res 804049ndash4058
Buwalda JP 1940 Geology of the Raymond Basin report to Pasadena Water DepartmentCrook R Allen CR Kamb B Payne CM Procter RJ 1987 Recent reverse faulting in the Transverse
Ranges California Pasadena CA US Geol Surv Prof Paper 133927ndash63Devouard B Posfai M Hua X Bazylinski DA Frankel RB Buseck PR 1998 Magnetite from
magnetotactic bacteria size distribution and twinning Am Mineral 831387ndash1398Frankel RB Bazylinski DA Johnson MS Taylor BL 1997 Magneto-Aerotaxis in marine coccoid
bacteria Biophys J 73994ndash1000Friedrich CG 1998 Physiology and genetics of sulfur-oxidizing bacteria Adv Microbial Physiol
39235ndash289Giovannoni SJ 1991 The polymerase chain reaction In Stackebrandt E Goodfellow M editors
Sequencing and hybridization techniques in bacterial systematics New York John Wiley andSons p 177ndash201
Guerrero R Pedros-Alio C Schmidt TM Mas J 1985 A survey of buoyant density of microorganismsin pure cultures and natural samples Microbiologia Sept 1(1ndash2)53ndash65
Harold FM 1966 Inorganic polyphosphates in biology structure metabolism and function BacteriolRev 30(4)772ndash794
Organization of Inclusions in Magnetococci 405
Iida A Akai J 1996 Crystalline sulfur inclusions in magnetotactic bacteria Sci Rep Nigata Univ SerE 1135ndash42
Kornberg A Fraley CD 2000 Inorganic polyphosphate a molecular fossil come to life ASM News66(5)275ndash280
Lins U Farina M 1999 Phosphorus-rich granules in uncultured magnetotactic bacteria FEMSMicrobiol Lett 172(2)23ndash28
Mann S Sparks NHC Frankel RB Bazylinski DA Jannasch HW 1990 Biomineralization of ferri-magnetic greigite (Fe3O4 and iron pyrite (FeS2 ) in a magnetotactic bacterium Nature 343258ndash
261Matsunaga T Sakaguchi T Tadokoro F 1991 Magnetite formation by a magnetic bacterium capable
of growing aerobically Appl Microbiol Biotechnol 35651ndash655Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993a Electron microscopy
study of magnetosomes in a cultured coccoid magnetotactic bacterium Proc Roy Soc Lond B251231ndash236
Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993b Electron microscopystudy of magnetosomes in two cultured vibrioid magnetotactic bacteria Proc Roy Soc Lond B251237ndash242
Moench TT 1988 Bilophococcus magnetotacticus gen nov sp nov a motile magnetic coccus Antvan Leeuwenhoek 54483ndash496
Moench TT Konetzka WA 1978 A novel method for the isolation and study of a magnetotacticbacterium Arch Microbiol 119203ndash212
Murray RGE Robinow CF 1994 Optical Microscopy In Gerhardt P editor Methods for generaland molecular bacteriology Washington DC American Society for Microbiology p 35ndash36
Prange A Arzberger I Engemann C Modrow H Schumann O Truper HG Steudel R Dahl C andHormes J 1999 In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteriaby X-ray absorption near edge spectroscopy Biochim Biophys Acta 1428446ndash454
Rappe MS Kemp PF Giovannoni SJ 1997 Phylogenetic diversity of marine coastal picoplankton16S rRNA genes cloned from the continental shelf off Cape Hatteras North Carolina LimnolOceanogr 42811ndash826
Reusch RN 2000 Transmembrane ion transport by polyphosphatepoly-(R)-3-hydroxybutyratecomplexes Biochemistry (Moscow) 65(3)335ndash352
Sakaguchi T Burgess JG Matsunaga T 1993 Magnetite formation by a sulphate-reducing bacteriumNature 36547ndash49
Schleifer KHSchuler D Spring S Weizenegger M Amann R 1991 The genus Magnetospirillum gennov description of Magnetospirillum gryphiswaldense sp nov and transfer of Aquaspirillummagnetotacticum to Magnetospirillum magnetotacticum comb nov System Appl Microbiol14379ndash385
Schuler D 1999 Formation of magnetosomes in magnetotactic bacteria J Mol Microbiol Biotechnol1(1)79ndash86
Schuler D Spring S Bazylinski DA 1999 Improved technique for the isolation of magnetotactic spir-illa from a freshwater sediment and their phylogenetic characterization System Appl Microbiol22466ndash471
Shively JM 1974 Inclusion bodies of prokaryotes Ann Rev Microbiol 28167ndash187Spring S Amann R Ludwig W Schleifer KH van Gemerden H Petersen N 1993 Dominating role
of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment ApplEnviron Microbiol 59(8)2397ndash2403
Spring S Amann R Ludwig W Schleifer KH Schuler D Poralla K Petersen N 1994 Phylogeneticanalysis of uncultured magnetotactic baceria from the Alpha-subclass of Proteobacteria SystemAppl Microbiol 17501ndash508
Stackebrandt E Goebel BM 1994 Taxonomic note a place for DNA-DNA reassociation and 16SrRNA sequence analysis in the present species de nition in bacteriology Int J System Bacteriol44846ndash849
Stanier RY Ingraham JL Wheelis ML Painter PR 1986 The microbial world 5th edition EnglewoodCliffs NJ Prentice Hall p 179
406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35
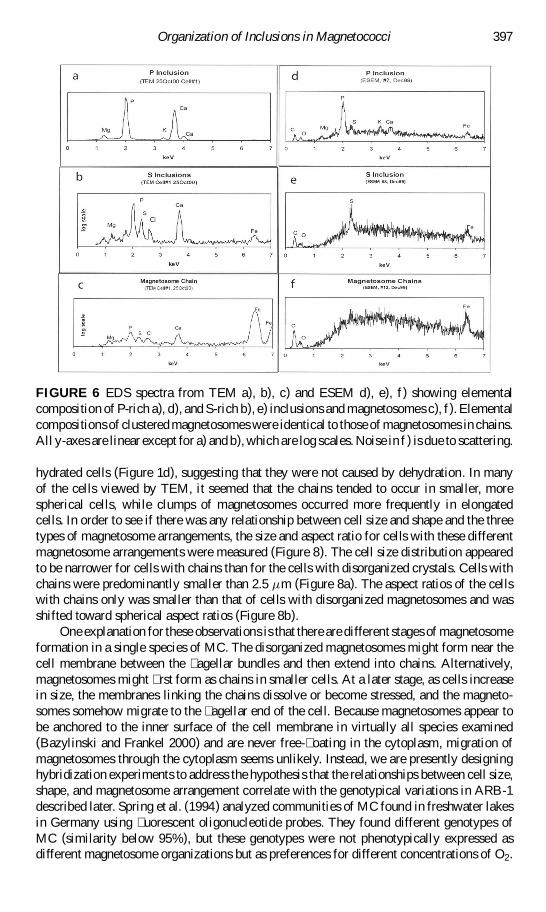
Organization of Inclusions in Magnetococci 397
FIGURE 6 EDS spectra from TEM a) b) c) and ESEM d) e) f) showing elementalcomposition of P-rich a) d) and S-rich b) e) inclusions and magnetosomes c) f) Elementalcompositions of clustered magnetosomes were identical to those of magnetosomes in chainsAll y-axes are linear except for a) and b) which are log scales Noise in f ) is due to scattering
hydrated cells (Figure 1d) suggesting that they were not caused by dehydration In manyof the cells viewed by TEM it seemed that the chains tended to occur in smaller morespherical cells while clumps of magnetosomes occurred more frequently in elongatedcells In order to see if there was any relationship between cell size and shape and the threetypes of magnetosome arrangements the size and aspect ratio for cells with these differentmagnetosome arrangements were measured (Figure 8) The cell size distribution appearedto be narrower for cells with chains than for the cells with disorganized crystals Cells withchains were predominantly smaller than 25 sup1m (Figure 8a) The aspect ratios of the cellswith chains only was smaller than that of cells with disorganized magnetosomes and wasshifted toward spherical aspect ratios (Figure 8b)
One explanation for these observations is that there are different stages of magnetosomeformation in a single species of MC The disorganized magnetosomes might form near thecell membrane between the agellar bundles and then extend into chains Alternativelymagnetosomes might rst form as chains in smaller cells At a later stage as cells increasein size the membranes linking the chains dissolve or become stressed and the magneto-somes somehow migrate to the agellar end of the cell Because magnetosomes appear tobe anchored to the inner surface of the cell membrane in virtually all species examined(Bazylinski and Frankel 2000) and are never free- oating in the cytoplasm migration ofmagnetosomes through the cytoplasm seems unlikely Instead we are presently designinghybridization experiments to address the hypothesis that the relationships between cell sizeshape and magnetosome arrangement correlate with the genotypical variations in ARB-1described later Spring et al (1994) analyzed communities of MC found in freshwater lakesin Germany using uorescent oligonucleotide probes They found different genotypes ofMC (similarity below 95) but these genotypes were not phenotypically expressed asdifferent magnetosome organizations but as preferences for different concentrations of O2
398 B L Cox et al
FIGURE 7 Evidence supporting magnetite as the magnetosome mineral in ARB-1 cellsa) Image of a single crystal of magnetite within a cell in an unstained glutaraldehyde- xedwhole cell mount taken with High Resolution TEM by H Vali b) Part of the magnetitecrystal shown in a) magni ed to show the distance between lattice fringes c) Selectedarea electron diffraction (SAED) of a group of magnetosomes in a cell from an un xedunstained whole cell mount showing the indices of some of the diffraction spots
Therefore we might expect no correlation between magnetite organization and phyloge-netic diversity of ARB-1 cells but perhaps a correlation between oxygen preference andphylogeny
A third possibility is that chains and clumps of magnetosomes form as a result ofdifferent environmental conditions Since our sampling methods were not selective as tothe cellsrsquo location in the vertical chemical (eg oxygen) and redox gradients that are sure to
Organization of Inclusions in Magnetococci 399
FIGURE 8 Relationships between organization of magnetosomes and cell size a) and cellaspect ratio b) Measurements were taken from both TEM and ESEM images Solid patternis for cells with clustered magnetosomes crossed pattern is for cells with magnetosomesin chains only Patterned bars originate at the top of solid bars
exist at the site the environmental conditions of our samples represent a relatively large rangeof oxygen and perhaps other chemical concentrations as well as redox conditions Magneticbacteria in culture are known to vary the size and number of magnetosomes in responseto the media composition (Meldrum et al 1993a) and particularly oxygen concentration(Blakemore et al 1985) Another possibility is that the clustered magnetosomes are formedby a different biological mechanism and serve a different function than magnetosomes inchains
Phosphorus (P) Inclusions
The two very large inclusion bodies seen in each cell always occurred in pairs in cellsand seemed to extend to the cell membrane and to be in contact with each other Thus themaximum dimension of the P-rich inclusions was the same as the minimum dimension ofthe cell and their combined width was approximately half of the long dimension of the cellIn ESEM images the two P-containing inclusions appeared to be attached in the middle insome cells and separated in others (Figure 1d) The P-containing inclusions appeared to beself-contained and perhaps bounded by some sort of coating or membrane in TEM images(Figure 2) and in light microscopy images of cells stained with safranin (not shown)
These inclusions were shown by both ESEM and TEM EDS to contain phosphorus with generally small amounts of potassium calcium magnesium sulfur and iron the lattertwo elements likely being due to the close proximity of the P-rich inclusions to sulfurglobules and magnetosomes (Figures 1 and 6) A very large calcium peak was observedwhen these large inclusions were analyzed in the TEM (Figure 6a) The samples shown inFigures 6a and 6d were collected at different times of the year which may explain some ofthe differences observed in these spectra Other EDS data from the ESEM (not shown) hadhigher calcium peaks than those shown in Figure 6d indicating that relative proportions ofP and Ca in P-rich inclusions vary even within the same sample
The inclusions were electron dense in TEM and had a mottled appearance due toablation by the electron beam Similar inclusions have been described for uncultured MCfrom the Itaupo Lagoon near Rio de Janeiro Brazil (Lins and Farina 1999) and from MillPond New Hampshire (Blakemore 1982) Like the structures in ARB-1 polyP granules aretranslucent in visible light (Spring et al 1994) dark (electron dense) in the TEM and oftenhave a mottled appearance (Harold 1966) The P-rich inclusions in ARB-1 cells stained
400 B L Cox et al
with methylene blue (Murray and Robinson 1994) which is suggestive of the presence ofpolyP but not con rmatory
PolyP is composed of linear chains of tetrahedral phosphate residues linked throughcommon oxygen atoms by phosphoanhydrid e bonds At physiological pH each unit carriesa monovalent negative charge giving polyP the property of having a large cation exchangecapacity The binding energy of polyP allows it to sequester Ca2C and Mg2C (Reusch 2000)consistent with the Ca2C and Mg2C signals observed in the EDS spectra of the ARB-1inclusions (Figure 6a 6d) The binding of these metals to the polyP granules contributesto their high electron density PolyP is often associated with polyhydroxybutyrat e (PHB)PHBs solvate (dissolve) cations and associate withpolyP to form Ca2C selective ion channelsacross plasma membranes (Reusch 2000)
The presence of polyP bodies is common in bacteria often resulting as a bacterialresponse to stress and they may play a role in cell motility quorum sensing and bio lmformation (Kornberg and Fraley 2000) Sulfate starvation is known to lead to rapid andmassive accumulation of polyP in some bacteria (Stanier et al 1986) When cells regainaccess to sulfate 32P uptake studies showed that PO3iexcl
4 present in the polyP bodies wasincorporated into nucleic acids PolyP is synthesized from ATP by the enzyme polyP kinasePolyP granules have been reported to be associated with nucleic acids (Harold 1966) butit is not clear whether the association occurs in vivo or after extraction This associationwith nucleic acids would be consistent with staining patterns seen when ARB-1 cells wereexamined with a uorescent livedead stain (nucleic acid stain) displaying a double-lobedstructure in all of the cells (Figure 9) This dumbbell shape appears to extend inside andacross the two phosphorus inclusions
The physiological function of the P inclusions in ARB-1 is not yet known but fromother cell systems several possibilities can be considered These inclusions might serveas a phosphorus reserve for the biosynthesis of nucleic acids and phospholipids underconditions of phosphorus starvation (Harold 1966) Still another possibility is buoyancyregulation Cells can vary their density by as much as 7 by incorporating substances intoinclusions (Guerrero et al 1985) A more speculative possibility is that ARB-1 cells canderive energy from polyP Acinetobacter johnsonii 210a is known to couple the excretion ofa metal phosphate chelate and HC generating a proton motive force (VanVeen et al 1994)A johnsonii takes up P from the aquatic environment by using two Pi transport systemsand stores it as intracellular granules of polyP
Sulfur (S) Inclusions
Cells of ARB-1 contained 5 to 10 spherical S-rich inclusions (Figures 1d) These rangedin size from approximately 100 to 700 nm although most of the S-rich inclusions in asingle cell were close to the same size and were located at one end of the cell oppositethe side where the magnetosomes were located and agellar bundles attached Like theP-rich inclusions the S-rich inclusions appeared to be enclosed in a coating or membrane(Figure 1d) EDS analysis from the ESEM showed these structures contained mostly S withsome carbon and oxygen (Figure 6e) EDS analysis from the TEM also showed S but evenhigher counts for Ca and P (Figure 6b) Beam spreading for the two methods is within aradius of 200 nm The X-rays produced by the 200-keV TEM beam sample the entire verticalextent of the cell unlike the 20 keV SEM beam which loses most of its energy Thereforethe EDS spectra collected at the TEM likely have sampled elements from the cytoplasmand cell membrane components both above and below the inclusion as well as from theP-rich inclusions nearby Other explanations are that hydrated sulfur inclusions changedtheir composition upon dehydration or that these two samples collected at different times
Organization of Inclusions in Magnetococci 401
FIGURE 9 Fluorescence micrograph of an ARB-1 cell suspension treated with Bacliteviability dyes showing that the nucleic acid staining pattern corresponds with the locationof the P-rich inclusions Baclite stains nucleic acids with cytonine (all cells) and propidiumiodate (dead or damaged cells) Bright double-lobed patterns are from ldquoliverdquo cells
of the day and the year may re ect a seasonal diurnal or cell cycle variability in the relativeproportions of the elements
Sulfur inclusions have been found in other uncultured magnetotactic bacteria includ-ing Magnetobacterium bavaricum (Spring et al 1993) Bilophococcus magnetotacticum(Moench 1988) and others (Iida and Akai 1996) and cultured species including the vibrioidstrains MV-1 and MV-2 the coccus strain MC-1 and the spirillum strain MV-4 (Bazylinskiand Frankel 2000) However only in Bilophococcus magnetotacticum was there a similarpolarization of sulfur-containing inclusions from the clustered magnetosomes within thecell (Moench 1988)
It seems likely that ARB-1 oxidizes H2S or other reduced sulfur compounds as anenergy source depositing ldquoelementalrdquo sulfur in intracellular inclusions like many othersul de-oxidizing bacteria including the anaerobic phototrophic purple sulfur bacteria suchas Chromatium vinosum and Thiocapsa roseopersicina the nonsulfur bacteria Beggiatoaspp and Thiothrix spp (Shively 1974 Friedrich 1998) and the magnetotactic bacteriamentioned before In these organisms the presence of sulfur globules is transitory andtakes place when the growth medium contains sul de andor thiosulfate When the sul deis completely utilized the sulfur is converted to sulfate (Stanier et al 1986)
Elemental S in sulfur globules is typically in a liquid state and analysis of in vivoglobules from some members of the purple and green sulfur bacteria by X-ray absorptionnear edge spectroscopy (Prange et al 1999) indicated the sulfur was in a chain structure
402 B L Cox et al
FIGURE 10 Phylogenetic position of six SSU rRNA gene clones related to previouslystudied MC This phylogenetic tree was rooted with Escherichia coli and constructed bythe neighbor joining method with the Kimura 2-parameter model for nucleotide change Amask including all nonambiguously aligned positions for 1427 nucleotides spanning nearlythe full length of the SSU rRNA gene was included Bootstrap values (100 replications)generated by the neighbor joining method are shown above relevant nodes and bootstrapvalues generated by maximum-parsimony analysis are shown below relevant nodes Onlybootstrap values above 50 are shown GenBank accession numbers are MC-CS92 X81182MC-pa119 Y13214 MC-CS81 X81184 MC-TB12 X81183 MC-MP17 X80996 MC-PA11 Y13212 MC12 L06456 partial sequence extended with data from MC-1 genomeRhizobium leguminosarum U31074 Agrobacterium tumefaciens D13294 Rhodobactersphaeroides X53854 Paracoccus denitri cans X59159 Zymomonas mobilis AF281034Sphingomonas paucimobilis D84528 Caulobacter crescentus M83799 Magnetospiril-lum magnetotacticum strain AMB1 D17514 S88429 Magnetospirillum magnetotacticumM10110 Rhodospirillum fulvum M59065 Magnetospirillum gryphiswaldense Y10109Magnetic Vibrio MV-1 LO6455 Oceanospirillum pusillum AB000482 Rhodospirillumrubrum D30778 Roseococcus thiosulfatophilus X72908 Escherichia coli J01695
Organization of Inclusions in Magnetococci 403
and was possibly complexed with water and organic residues If the S globules in ARB-1cells are analogous to those in the purple sulfur bacteria Chromatium vinosum and Thio-capsa roseopersicina they may contain a signi cant protein content Sulfur globules fromthe anaerobic phototrophs C vinosum and T roseopersicina were found to contain upto 52 protein that formed an envelope enclosing the S globules (Brune 1995) Theseproteins were isolated and analyzed and found to be glycine and tyrosine-rich structuralproteins which were homologous for the two species suggesting they are highly con-served (Brune 1995) Phylogenetically C vinosum and T roseopersicina are members ofthe deg -Proteobacteria whereas MC are members of the reg-Proteobacteria To our knowledgemagnetotactic bacteria may be the only members of the reg-Proteobacteria that make internalsulfur globules
Phylogenetic Af liation
We wished to detemine whether ARB-1 was a single species or a mixed population of MCToward that end 45 SSU rRNA genes were cloned from partially puri ed ARB-1 cells30 of which displayed similar restriction fragment length patterns (RFLP) to previouslycharacterized MC (data not shown) The nding that 15 patterns were not related to pre-viously characterized MC was not surprising given that the initial sample was not a pureculture but rather a population magnetically enriched from a sample originating from arelatively large vertical column containing both sediments and overlying water Fourteenof the MC-related clones representing different RFLP pattern groups were chosen for par-tial sequencing 10 of these sequences showed high similarity to previously characterizedMC Six of these sequences were phylogenetically distinct and were fully bidirectionallysequenced
ARB-1 MC SSU rRNA genes formed a monophyletic cluster related to but dis-tinct from other MC (Figure 10) Within the cluster the similarity between SSU rRNAgene sequences ranged from 97 (MC-H1arb and MC-g6arb) to 92 (MC-h1arb andMC-g2arb) The nearest neighbor to the ARB-1 cluster MC-CS92 was ca 89 similarand the nearest cultured relative MC-1 was ca 88 similar to the ARB-1 cluster As arule of thumb for SSU rRNA genes gt97 similarity is considered the same species andgt93ndash95 similarity is considered the same genus (Stackebrandt and Goebel 1994) Thesedata strongly suggest that ARB-1 represents a population of different diverse MC Therewas signi cant variation within the ARB-1 cluster and therefore we do not consider ARB-1to consist of bacteria of a single species or genus Correlations if any between phylogeneticand phenotypic variations in the ARB-1 cells have yet to be determined
Summary
We have partially characterized a population of magnetotactic cocci (MC) referred to asARB-1 which was particularly abundant in iron-rich sediments of a small eutrophic lake andmorphologically and structurally similar to an uncultured species of MC called Bilophococ-cus magnetotacticus The latter organism was found in large numbers in a sewage treatmentpond Cells of ARB-1 had a relatively wide size range of 1 to 4 sup1m They were motileGram-negative and bilophotrichously agellated Cells of this uncultured magnetotacticcoccus contained three types of intracellular inclusions that appeared to be in a consistentarrangement within the cell The magnetosomes contained crystals of magnetite and wereeither in chains andor polar clumps at the end of the cell where the agellar bundles werelocated Large P-rich bodies occupied the majority of the cell volume and appeared to sepa-rate the S-containing inclusions from the magnetosomes The presence of these large P-rich
404 B L Cox et al
inclusions might be an indication of cell stress or may be important in cell buoyancy or inenergy generation The presence of S-rich inclusions located at the end of the cell oppositethe magnetosomes strongly suggests that ARB-1 cells are sul de-oxidizing bacteria andthat the S might be useful as a stored electron donor when cells travel from sul de-rich tosul de-depleted zones in the water column The wide range of the cell sizes the differentmagnetosome arrangements and the large number of different retrieved 16S rRNA genesequences suggest that ARB-1 represents a population of at least several different MC 16SrRNA sequences obtained from cells of ARB-1 appeared to form a monophyletic clusterrelated to but distinct from other uncultured and cultured MC Future work using uores-cent oligonucleotide probes microscopy and physiological measurements should clarifyrelationships between phylogeny cell morphology and organization of inclusions
References
Balkwill DL Maratea D Blakemore RP 1980 Ultrastructure of a magnetotactic spirillum J Bacteriol141(3)1399ndash1408
Bazylinski DA Frankel RB 2000 Biologically controlled mineralization of magnetic iron mineralsby magnetotactic bacteria In Lovley DR editor Environmental microbe-metal interactionsWashington DC ASM Press p 109ndash143
Bazylinski DA Frankel RB Jannasch HW 1988 Anaerobic magnetite production by a marinemagnetotactic bacterium Nature 334518ndash519
Beveridge T 1999 Structures of gram-negative cell walls and their derived membrane vesiclesJ Bacteriol 181(6)4725ndash4733
Beveridge TJ Popkin TJ Cole RM 1994 Electron microscopy In Gerhardt P editor Methods forgeneral and molecular bacteriology Washington DC American Society for Microbiology p 25
Blakemore R 1975 Magnetotactic bacteria Science 190377ndash379Blakemore RP 1982 Magnetotactic bacteria Ann Rev Microbiol 36217ndash238Blakemore RP Maratea D Wolfe RS 1979 Isolation and pure culture of a freshwater magnetic
spirillum in chemically de ned medium J Bacteriol 140(2)720ndash729Blakemore RP Short KA Bazylinski DA Rosenblatt C Frankel RB 1985 Microaerobic conditions
are required for magnetite formation within Aquaspirillum magnetotacticum Geomicrobiol J4(1)53ndash71
Brune DC 1995 Isolation and characterization of sulfur globule proteins from Chromatium vinosumand Thiocapsa roseopersicina Arch Microbiol 163(1)391ndash399
Butler RF Banerjee SK 1975 Theoretical single-domain grain size range in magnetite and titano-magnetite J Geophys Res 804049ndash4058
Buwalda JP 1940 Geology of the Raymond Basin report to Pasadena Water DepartmentCrook R Allen CR Kamb B Payne CM Procter RJ 1987 Recent reverse faulting in the Transverse
Ranges California Pasadena CA US Geol Surv Prof Paper 133927ndash63Devouard B Posfai M Hua X Bazylinski DA Frankel RB Buseck PR 1998 Magnetite from
magnetotactic bacteria size distribution and twinning Am Mineral 831387ndash1398Frankel RB Bazylinski DA Johnson MS Taylor BL 1997 Magneto-Aerotaxis in marine coccoid
bacteria Biophys J 73994ndash1000Friedrich CG 1998 Physiology and genetics of sulfur-oxidizing bacteria Adv Microbial Physiol
39235ndash289Giovannoni SJ 1991 The polymerase chain reaction In Stackebrandt E Goodfellow M editors
Sequencing and hybridization techniques in bacterial systematics New York John Wiley andSons p 177ndash201
Guerrero R Pedros-Alio C Schmidt TM Mas J 1985 A survey of buoyant density of microorganismsin pure cultures and natural samples Microbiologia Sept 1(1ndash2)53ndash65
Harold FM 1966 Inorganic polyphosphates in biology structure metabolism and function BacteriolRev 30(4)772ndash794
Organization of Inclusions in Magnetococci 405
Iida A Akai J 1996 Crystalline sulfur inclusions in magnetotactic bacteria Sci Rep Nigata Univ SerE 1135ndash42
Kornberg A Fraley CD 2000 Inorganic polyphosphate a molecular fossil come to life ASM News66(5)275ndash280
Lins U Farina M 1999 Phosphorus-rich granules in uncultured magnetotactic bacteria FEMSMicrobiol Lett 172(2)23ndash28
Mann S Sparks NHC Frankel RB Bazylinski DA Jannasch HW 1990 Biomineralization of ferri-magnetic greigite (Fe3O4 and iron pyrite (FeS2 ) in a magnetotactic bacterium Nature 343258ndash
261Matsunaga T Sakaguchi T Tadokoro F 1991 Magnetite formation by a magnetic bacterium capable
of growing aerobically Appl Microbiol Biotechnol 35651ndash655Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993a Electron microscopy
study of magnetosomes in a cultured coccoid magnetotactic bacterium Proc Roy Soc Lond B251231ndash236
Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993b Electron microscopystudy of magnetosomes in two cultured vibrioid magnetotactic bacteria Proc Roy Soc Lond B251237ndash242
Moench TT 1988 Bilophococcus magnetotacticus gen nov sp nov a motile magnetic coccus Antvan Leeuwenhoek 54483ndash496
Moench TT Konetzka WA 1978 A novel method for the isolation and study of a magnetotacticbacterium Arch Microbiol 119203ndash212
Murray RGE Robinow CF 1994 Optical Microscopy In Gerhardt P editor Methods for generaland molecular bacteriology Washington DC American Society for Microbiology p 35ndash36
Prange A Arzberger I Engemann C Modrow H Schumann O Truper HG Steudel R Dahl C andHormes J 1999 In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteriaby X-ray absorption near edge spectroscopy Biochim Biophys Acta 1428446ndash454
Rappe MS Kemp PF Giovannoni SJ 1997 Phylogenetic diversity of marine coastal picoplankton16S rRNA genes cloned from the continental shelf off Cape Hatteras North Carolina LimnolOceanogr 42811ndash826
Reusch RN 2000 Transmembrane ion transport by polyphosphatepoly-(R)-3-hydroxybutyratecomplexes Biochemistry (Moscow) 65(3)335ndash352
Sakaguchi T Burgess JG Matsunaga T 1993 Magnetite formation by a sulphate-reducing bacteriumNature 36547ndash49
Schleifer KHSchuler D Spring S Weizenegger M Amann R 1991 The genus Magnetospirillum gennov description of Magnetospirillum gryphiswaldense sp nov and transfer of Aquaspirillummagnetotacticum to Magnetospirillum magnetotacticum comb nov System Appl Microbiol14379ndash385
Schuler D 1999 Formation of magnetosomes in magnetotactic bacteria J Mol Microbiol Biotechnol1(1)79ndash86
Schuler D Spring S Bazylinski DA 1999 Improved technique for the isolation of magnetotactic spir-illa from a freshwater sediment and their phylogenetic characterization System Appl Microbiol22466ndash471
Shively JM 1974 Inclusion bodies of prokaryotes Ann Rev Microbiol 28167ndash187Spring S Amann R Ludwig W Schleifer KH van Gemerden H Petersen N 1993 Dominating role
of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment ApplEnviron Microbiol 59(8)2397ndash2403
Spring S Amann R Ludwig W Schleifer KH Schuler D Poralla K Petersen N 1994 Phylogeneticanalysis of uncultured magnetotactic baceria from the Alpha-subclass of Proteobacteria SystemAppl Microbiol 17501ndash508
Stackebrandt E Goebel BM 1994 Taxonomic note a place for DNA-DNA reassociation and 16SrRNA sequence analysis in the present species de nition in bacteriology Int J System Bacteriol44846ndash849
Stanier RY Ingraham JL Wheelis ML Painter PR 1986 The microbial world 5th edition EnglewoodCliffs NJ Prentice Hall p 179
406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35
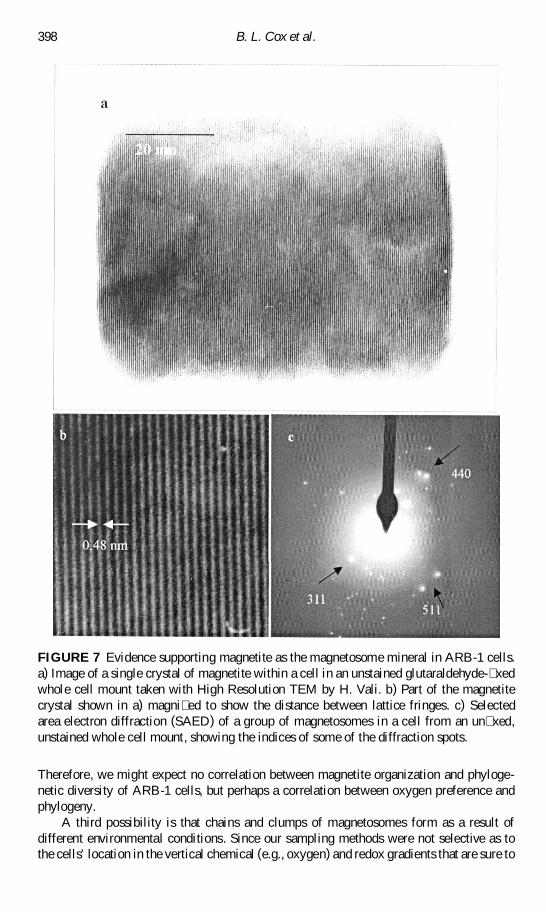
398 B L Cox et al
FIGURE 7 Evidence supporting magnetite as the magnetosome mineral in ARB-1 cellsa) Image of a single crystal of magnetite within a cell in an unstained glutaraldehyde- xedwhole cell mount taken with High Resolution TEM by H Vali b) Part of the magnetitecrystal shown in a) magni ed to show the distance between lattice fringes c) Selectedarea electron diffraction (SAED) of a group of magnetosomes in a cell from an un xedunstained whole cell mount showing the indices of some of the diffraction spots
Therefore we might expect no correlation between magnetite organization and phyloge-netic diversity of ARB-1 cells but perhaps a correlation between oxygen preference andphylogeny
A third possibility is that chains and clumps of magnetosomes form as a result ofdifferent environmental conditions Since our sampling methods were not selective as tothe cellsrsquo location in the vertical chemical (eg oxygen) and redox gradients that are sure to
Organization of Inclusions in Magnetococci 399
FIGURE 8 Relationships between organization of magnetosomes and cell size a) and cellaspect ratio b) Measurements were taken from both TEM and ESEM images Solid patternis for cells with clustered magnetosomes crossed pattern is for cells with magnetosomesin chains only Patterned bars originate at the top of solid bars
exist at the site the environmental conditions of our samples represent a relatively large rangeof oxygen and perhaps other chemical concentrations as well as redox conditions Magneticbacteria in culture are known to vary the size and number of magnetosomes in responseto the media composition (Meldrum et al 1993a) and particularly oxygen concentration(Blakemore et al 1985) Another possibility is that the clustered magnetosomes are formedby a different biological mechanism and serve a different function than magnetosomes inchains
Phosphorus (P) Inclusions
The two very large inclusion bodies seen in each cell always occurred in pairs in cellsand seemed to extend to the cell membrane and to be in contact with each other Thus themaximum dimension of the P-rich inclusions was the same as the minimum dimension ofthe cell and their combined width was approximately half of the long dimension of the cellIn ESEM images the two P-containing inclusions appeared to be attached in the middle insome cells and separated in others (Figure 1d) The P-containing inclusions appeared to beself-contained and perhaps bounded by some sort of coating or membrane in TEM images(Figure 2) and in light microscopy images of cells stained with safranin (not shown)
These inclusions were shown by both ESEM and TEM EDS to contain phosphorus with generally small amounts of potassium calcium magnesium sulfur and iron the lattertwo elements likely being due to the close proximity of the P-rich inclusions to sulfurglobules and magnetosomes (Figures 1 and 6) A very large calcium peak was observedwhen these large inclusions were analyzed in the TEM (Figure 6a) The samples shown inFigures 6a and 6d were collected at different times of the year which may explain some ofthe differences observed in these spectra Other EDS data from the ESEM (not shown) hadhigher calcium peaks than those shown in Figure 6d indicating that relative proportions ofP and Ca in P-rich inclusions vary even within the same sample
The inclusions were electron dense in TEM and had a mottled appearance due toablation by the electron beam Similar inclusions have been described for uncultured MCfrom the Itaupo Lagoon near Rio de Janeiro Brazil (Lins and Farina 1999) and from MillPond New Hampshire (Blakemore 1982) Like the structures in ARB-1 polyP granules aretranslucent in visible light (Spring et al 1994) dark (electron dense) in the TEM and oftenhave a mottled appearance (Harold 1966) The P-rich inclusions in ARB-1 cells stained
400 B L Cox et al
with methylene blue (Murray and Robinson 1994) which is suggestive of the presence ofpolyP but not con rmatory
PolyP is composed of linear chains of tetrahedral phosphate residues linked throughcommon oxygen atoms by phosphoanhydrid e bonds At physiological pH each unit carriesa monovalent negative charge giving polyP the property of having a large cation exchangecapacity The binding energy of polyP allows it to sequester Ca2C and Mg2C (Reusch 2000)consistent with the Ca2C and Mg2C signals observed in the EDS spectra of the ARB-1inclusions (Figure 6a 6d) The binding of these metals to the polyP granules contributesto their high electron density PolyP is often associated with polyhydroxybutyrat e (PHB)PHBs solvate (dissolve) cations and associate withpolyP to form Ca2C selective ion channelsacross plasma membranes (Reusch 2000)
The presence of polyP bodies is common in bacteria often resulting as a bacterialresponse to stress and they may play a role in cell motility quorum sensing and bio lmformation (Kornberg and Fraley 2000) Sulfate starvation is known to lead to rapid andmassive accumulation of polyP in some bacteria (Stanier et al 1986) When cells regainaccess to sulfate 32P uptake studies showed that PO3iexcl
4 present in the polyP bodies wasincorporated into nucleic acids PolyP is synthesized from ATP by the enzyme polyP kinasePolyP granules have been reported to be associated with nucleic acids (Harold 1966) butit is not clear whether the association occurs in vivo or after extraction This associationwith nucleic acids would be consistent with staining patterns seen when ARB-1 cells wereexamined with a uorescent livedead stain (nucleic acid stain) displaying a double-lobedstructure in all of the cells (Figure 9) This dumbbell shape appears to extend inside andacross the two phosphorus inclusions
The physiological function of the P inclusions in ARB-1 is not yet known but fromother cell systems several possibilities can be considered These inclusions might serveas a phosphorus reserve for the biosynthesis of nucleic acids and phospholipids underconditions of phosphorus starvation (Harold 1966) Still another possibility is buoyancyregulation Cells can vary their density by as much as 7 by incorporating substances intoinclusions (Guerrero et al 1985) A more speculative possibility is that ARB-1 cells canderive energy from polyP Acinetobacter johnsonii 210a is known to couple the excretion ofa metal phosphate chelate and HC generating a proton motive force (VanVeen et al 1994)A johnsonii takes up P from the aquatic environment by using two Pi transport systemsand stores it as intracellular granules of polyP
Sulfur (S) Inclusions
Cells of ARB-1 contained 5 to 10 spherical S-rich inclusions (Figures 1d) These rangedin size from approximately 100 to 700 nm although most of the S-rich inclusions in asingle cell were close to the same size and were located at one end of the cell oppositethe side where the magnetosomes were located and agellar bundles attached Like theP-rich inclusions the S-rich inclusions appeared to be enclosed in a coating or membrane(Figure 1d) EDS analysis from the ESEM showed these structures contained mostly S withsome carbon and oxygen (Figure 6e) EDS analysis from the TEM also showed S but evenhigher counts for Ca and P (Figure 6b) Beam spreading for the two methods is within aradius of 200 nm The X-rays produced by the 200-keV TEM beam sample the entire verticalextent of the cell unlike the 20 keV SEM beam which loses most of its energy Thereforethe EDS spectra collected at the TEM likely have sampled elements from the cytoplasmand cell membrane components both above and below the inclusion as well as from theP-rich inclusions nearby Other explanations are that hydrated sulfur inclusions changedtheir composition upon dehydration or that these two samples collected at different times
Organization of Inclusions in Magnetococci 401
FIGURE 9 Fluorescence micrograph of an ARB-1 cell suspension treated with Bacliteviability dyes showing that the nucleic acid staining pattern corresponds with the locationof the P-rich inclusions Baclite stains nucleic acids with cytonine (all cells) and propidiumiodate (dead or damaged cells) Bright double-lobed patterns are from ldquoliverdquo cells
of the day and the year may re ect a seasonal diurnal or cell cycle variability in the relativeproportions of the elements
Sulfur inclusions have been found in other uncultured magnetotactic bacteria includ-ing Magnetobacterium bavaricum (Spring et al 1993) Bilophococcus magnetotacticum(Moench 1988) and others (Iida and Akai 1996) and cultured species including the vibrioidstrains MV-1 and MV-2 the coccus strain MC-1 and the spirillum strain MV-4 (Bazylinskiand Frankel 2000) However only in Bilophococcus magnetotacticum was there a similarpolarization of sulfur-containing inclusions from the clustered magnetosomes within thecell (Moench 1988)
It seems likely that ARB-1 oxidizes H2S or other reduced sulfur compounds as anenergy source depositing ldquoelementalrdquo sulfur in intracellular inclusions like many othersul de-oxidizing bacteria including the anaerobic phototrophic purple sulfur bacteria suchas Chromatium vinosum and Thiocapsa roseopersicina the nonsulfur bacteria Beggiatoaspp and Thiothrix spp (Shively 1974 Friedrich 1998) and the magnetotactic bacteriamentioned before In these organisms the presence of sulfur globules is transitory andtakes place when the growth medium contains sul de andor thiosulfate When the sul deis completely utilized the sulfur is converted to sulfate (Stanier et al 1986)
Elemental S in sulfur globules is typically in a liquid state and analysis of in vivoglobules from some members of the purple and green sulfur bacteria by X-ray absorptionnear edge spectroscopy (Prange et al 1999) indicated the sulfur was in a chain structure
402 B L Cox et al
FIGURE 10 Phylogenetic position of six SSU rRNA gene clones related to previouslystudied MC This phylogenetic tree was rooted with Escherichia coli and constructed bythe neighbor joining method with the Kimura 2-parameter model for nucleotide change Amask including all nonambiguously aligned positions for 1427 nucleotides spanning nearlythe full length of the SSU rRNA gene was included Bootstrap values (100 replications)generated by the neighbor joining method are shown above relevant nodes and bootstrapvalues generated by maximum-parsimony analysis are shown below relevant nodes Onlybootstrap values above 50 are shown GenBank accession numbers are MC-CS92 X81182MC-pa119 Y13214 MC-CS81 X81184 MC-TB12 X81183 MC-MP17 X80996 MC-PA11 Y13212 MC12 L06456 partial sequence extended with data from MC-1 genomeRhizobium leguminosarum U31074 Agrobacterium tumefaciens D13294 Rhodobactersphaeroides X53854 Paracoccus denitri cans X59159 Zymomonas mobilis AF281034Sphingomonas paucimobilis D84528 Caulobacter crescentus M83799 Magnetospiril-lum magnetotacticum strain AMB1 D17514 S88429 Magnetospirillum magnetotacticumM10110 Rhodospirillum fulvum M59065 Magnetospirillum gryphiswaldense Y10109Magnetic Vibrio MV-1 LO6455 Oceanospirillum pusillum AB000482 Rhodospirillumrubrum D30778 Roseococcus thiosulfatophilus X72908 Escherichia coli J01695
Organization of Inclusions in Magnetococci 403
and was possibly complexed with water and organic residues If the S globules in ARB-1cells are analogous to those in the purple sulfur bacteria Chromatium vinosum and Thio-capsa roseopersicina they may contain a signi cant protein content Sulfur globules fromthe anaerobic phototrophs C vinosum and T roseopersicina were found to contain upto 52 protein that formed an envelope enclosing the S globules (Brune 1995) Theseproteins were isolated and analyzed and found to be glycine and tyrosine-rich structuralproteins which were homologous for the two species suggesting they are highly con-served (Brune 1995) Phylogenetically C vinosum and T roseopersicina are members ofthe deg -Proteobacteria whereas MC are members of the reg-Proteobacteria To our knowledgemagnetotactic bacteria may be the only members of the reg-Proteobacteria that make internalsulfur globules
Phylogenetic Af liation
We wished to detemine whether ARB-1 was a single species or a mixed population of MCToward that end 45 SSU rRNA genes were cloned from partially puri ed ARB-1 cells30 of which displayed similar restriction fragment length patterns (RFLP) to previouslycharacterized MC (data not shown) The nding that 15 patterns were not related to pre-viously characterized MC was not surprising given that the initial sample was not a pureculture but rather a population magnetically enriched from a sample originating from arelatively large vertical column containing both sediments and overlying water Fourteenof the MC-related clones representing different RFLP pattern groups were chosen for par-tial sequencing 10 of these sequences showed high similarity to previously characterizedMC Six of these sequences were phylogenetically distinct and were fully bidirectionallysequenced
ARB-1 MC SSU rRNA genes formed a monophyletic cluster related to but dis-tinct from other MC (Figure 10) Within the cluster the similarity between SSU rRNAgene sequences ranged from 97 (MC-H1arb and MC-g6arb) to 92 (MC-h1arb andMC-g2arb) The nearest neighbor to the ARB-1 cluster MC-CS92 was ca 89 similarand the nearest cultured relative MC-1 was ca 88 similar to the ARB-1 cluster As arule of thumb for SSU rRNA genes gt97 similarity is considered the same species andgt93ndash95 similarity is considered the same genus (Stackebrandt and Goebel 1994) Thesedata strongly suggest that ARB-1 represents a population of different diverse MC Therewas signi cant variation within the ARB-1 cluster and therefore we do not consider ARB-1to consist of bacteria of a single species or genus Correlations if any between phylogeneticand phenotypic variations in the ARB-1 cells have yet to be determined
Summary
We have partially characterized a population of magnetotactic cocci (MC) referred to asARB-1 which was particularly abundant in iron-rich sediments of a small eutrophic lake andmorphologically and structurally similar to an uncultured species of MC called Bilophococ-cus magnetotacticus The latter organism was found in large numbers in a sewage treatmentpond Cells of ARB-1 had a relatively wide size range of 1 to 4 sup1m They were motileGram-negative and bilophotrichously agellated Cells of this uncultured magnetotacticcoccus contained three types of intracellular inclusions that appeared to be in a consistentarrangement within the cell The magnetosomes contained crystals of magnetite and wereeither in chains andor polar clumps at the end of the cell where the agellar bundles werelocated Large P-rich bodies occupied the majority of the cell volume and appeared to sepa-rate the S-containing inclusions from the magnetosomes The presence of these large P-rich
404 B L Cox et al
inclusions might be an indication of cell stress or may be important in cell buoyancy or inenergy generation The presence of S-rich inclusions located at the end of the cell oppositethe magnetosomes strongly suggests that ARB-1 cells are sul de-oxidizing bacteria andthat the S might be useful as a stored electron donor when cells travel from sul de-rich tosul de-depleted zones in the water column The wide range of the cell sizes the differentmagnetosome arrangements and the large number of different retrieved 16S rRNA genesequences suggest that ARB-1 represents a population of at least several different MC 16SrRNA sequences obtained from cells of ARB-1 appeared to form a monophyletic clusterrelated to but distinct from other uncultured and cultured MC Future work using uores-cent oligonucleotide probes microscopy and physiological measurements should clarifyrelationships between phylogeny cell morphology and organization of inclusions
References
Balkwill DL Maratea D Blakemore RP 1980 Ultrastructure of a magnetotactic spirillum J Bacteriol141(3)1399ndash1408
Bazylinski DA Frankel RB 2000 Biologically controlled mineralization of magnetic iron mineralsby magnetotactic bacteria In Lovley DR editor Environmental microbe-metal interactionsWashington DC ASM Press p 109ndash143
Bazylinski DA Frankel RB Jannasch HW 1988 Anaerobic magnetite production by a marinemagnetotactic bacterium Nature 334518ndash519
Beveridge T 1999 Structures of gram-negative cell walls and their derived membrane vesiclesJ Bacteriol 181(6)4725ndash4733
Beveridge TJ Popkin TJ Cole RM 1994 Electron microscopy In Gerhardt P editor Methods forgeneral and molecular bacteriology Washington DC American Society for Microbiology p 25
Blakemore R 1975 Magnetotactic bacteria Science 190377ndash379Blakemore RP 1982 Magnetotactic bacteria Ann Rev Microbiol 36217ndash238Blakemore RP Maratea D Wolfe RS 1979 Isolation and pure culture of a freshwater magnetic
spirillum in chemically de ned medium J Bacteriol 140(2)720ndash729Blakemore RP Short KA Bazylinski DA Rosenblatt C Frankel RB 1985 Microaerobic conditions
are required for magnetite formation within Aquaspirillum magnetotacticum Geomicrobiol J4(1)53ndash71
Brune DC 1995 Isolation and characterization of sulfur globule proteins from Chromatium vinosumand Thiocapsa roseopersicina Arch Microbiol 163(1)391ndash399
Butler RF Banerjee SK 1975 Theoretical single-domain grain size range in magnetite and titano-magnetite J Geophys Res 804049ndash4058
Buwalda JP 1940 Geology of the Raymond Basin report to Pasadena Water DepartmentCrook R Allen CR Kamb B Payne CM Procter RJ 1987 Recent reverse faulting in the Transverse
Ranges California Pasadena CA US Geol Surv Prof Paper 133927ndash63Devouard B Posfai M Hua X Bazylinski DA Frankel RB Buseck PR 1998 Magnetite from
magnetotactic bacteria size distribution and twinning Am Mineral 831387ndash1398Frankel RB Bazylinski DA Johnson MS Taylor BL 1997 Magneto-Aerotaxis in marine coccoid
bacteria Biophys J 73994ndash1000Friedrich CG 1998 Physiology and genetics of sulfur-oxidizing bacteria Adv Microbial Physiol
39235ndash289Giovannoni SJ 1991 The polymerase chain reaction In Stackebrandt E Goodfellow M editors
Sequencing and hybridization techniques in bacterial systematics New York John Wiley andSons p 177ndash201
Guerrero R Pedros-Alio C Schmidt TM Mas J 1985 A survey of buoyant density of microorganismsin pure cultures and natural samples Microbiologia Sept 1(1ndash2)53ndash65
Harold FM 1966 Inorganic polyphosphates in biology structure metabolism and function BacteriolRev 30(4)772ndash794
Organization of Inclusions in Magnetococci 405
Iida A Akai J 1996 Crystalline sulfur inclusions in magnetotactic bacteria Sci Rep Nigata Univ SerE 1135ndash42
Kornberg A Fraley CD 2000 Inorganic polyphosphate a molecular fossil come to life ASM News66(5)275ndash280
Lins U Farina M 1999 Phosphorus-rich granules in uncultured magnetotactic bacteria FEMSMicrobiol Lett 172(2)23ndash28
Mann S Sparks NHC Frankel RB Bazylinski DA Jannasch HW 1990 Biomineralization of ferri-magnetic greigite (Fe3O4 and iron pyrite (FeS2 ) in a magnetotactic bacterium Nature 343258ndash
261Matsunaga T Sakaguchi T Tadokoro F 1991 Magnetite formation by a magnetic bacterium capable
of growing aerobically Appl Microbiol Biotechnol 35651ndash655Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993a Electron microscopy
study of magnetosomes in a cultured coccoid magnetotactic bacterium Proc Roy Soc Lond B251231ndash236
Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993b Electron microscopystudy of magnetosomes in two cultured vibrioid magnetotactic bacteria Proc Roy Soc Lond B251237ndash242
Moench TT 1988 Bilophococcus magnetotacticus gen nov sp nov a motile magnetic coccus Antvan Leeuwenhoek 54483ndash496
Moench TT Konetzka WA 1978 A novel method for the isolation and study of a magnetotacticbacterium Arch Microbiol 119203ndash212
Murray RGE Robinow CF 1994 Optical Microscopy In Gerhardt P editor Methods for generaland molecular bacteriology Washington DC American Society for Microbiology p 35ndash36
Prange A Arzberger I Engemann C Modrow H Schumann O Truper HG Steudel R Dahl C andHormes J 1999 In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteriaby X-ray absorption near edge spectroscopy Biochim Biophys Acta 1428446ndash454
Rappe MS Kemp PF Giovannoni SJ 1997 Phylogenetic diversity of marine coastal picoplankton16S rRNA genes cloned from the continental shelf off Cape Hatteras North Carolina LimnolOceanogr 42811ndash826
Reusch RN 2000 Transmembrane ion transport by polyphosphatepoly-(R)-3-hydroxybutyratecomplexes Biochemistry (Moscow) 65(3)335ndash352
Sakaguchi T Burgess JG Matsunaga T 1993 Magnetite formation by a sulphate-reducing bacteriumNature 36547ndash49
Schleifer KHSchuler D Spring S Weizenegger M Amann R 1991 The genus Magnetospirillum gennov description of Magnetospirillum gryphiswaldense sp nov and transfer of Aquaspirillummagnetotacticum to Magnetospirillum magnetotacticum comb nov System Appl Microbiol14379ndash385
Schuler D 1999 Formation of magnetosomes in magnetotactic bacteria J Mol Microbiol Biotechnol1(1)79ndash86
Schuler D Spring S Bazylinski DA 1999 Improved technique for the isolation of magnetotactic spir-illa from a freshwater sediment and their phylogenetic characterization System Appl Microbiol22466ndash471
Shively JM 1974 Inclusion bodies of prokaryotes Ann Rev Microbiol 28167ndash187Spring S Amann R Ludwig W Schleifer KH van Gemerden H Petersen N 1993 Dominating role
of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment ApplEnviron Microbiol 59(8)2397ndash2403
Spring S Amann R Ludwig W Schleifer KH Schuler D Poralla K Petersen N 1994 Phylogeneticanalysis of uncultured magnetotactic baceria from the Alpha-subclass of Proteobacteria SystemAppl Microbiol 17501ndash508
Stackebrandt E Goebel BM 1994 Taxonomic note a place for DNA-DNA reassociation and 16SrRNA sequence analysis in the present species de nition in bacteriology Int J System Bacteriol44846ndash849
Stanier RY Ingraham JL Wheelis ML Painter PR 1986 The microbial world 5th edition EnglewoodCliffs NJ Prentice Hall p 179
406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35

Organization of Inclusions in Magnetococci 399
FIGURE 8 Relationships between organization of magnetosomes and cell size a) and cellaspect ratio b) Measurements were taken from both TEM and ESEM images Solid patternis for cells with clustered magnetosomes crossed pattern is for cells with magnetosomesin chains only Patterned bars originate at the top of solid bars
exist at the site the environmental conditions of our samples represent a relatively large rangeof oxygen and perhaps other chemical concentrations as well as redox conditions Magneticbacteria in culture are known to vary the size and number of magnetosomes in responseto the media composition (Meldrum et al 1993a) and particularly oxygen concentration(Blakemore et al 1985) Another possibility is that the clustered magnetosomes are formedby a different biological mechanism and serve a different function than magnetosomes inchains
Phosphorus (P) Inclusions
The two very large inclusion bodies seen in each cell always occurred in pairs in cellsand seemed to extend to the cell membrane and to be in contact with each other Thus themaximum dimension of the P-rich inclusions was the same as the minimum dimension ofthe cell and their combined width was approximately half of the long dimension of the cellIn ESEM images the two P-containing inclusions appeared to be attached in the middle insome cells and separated in others (Figure 1d) The P-containing inclusions appeared to beself-contained and perhaps bounded by some sort of coating or membrane in TEM images(Figure 2) and in light microscopy images of cells stained with safranin (not shown)
These inclusions were shown by both ESEM and TEM EDS to contain phosphorus with generally small amounts of potassium calcium magnesium sulfur and iron the lattertwo elements likely being due to the close proximity of the P-rich inclusions to sulfurglobules and magnetosomes (Figures 1 and 6) A very large calcium peak was observedwhen these large inclusions were analyzed in the TEM (Figure 6a) The samples shown inFigures 6a and 6d were collected at different times of the year which may explain some ofthe differences observed in these spectra Other EDS data from the ESEM (not shown) hadhigher calcium peaks than those shown in Figure 6d indicating that relative proportions ofP and Ca in P-rich inclusions vary even within the same sample
The inclusions were electron dense in TEM and had a mottled appearance due toablation by the electron beam Similar inclusions have been described for uncultured MCfrom the Itaupo Lagoon near Rio de Janeiro Brazil (Lins and Farina 1999) and from MillPond New Hampshire (Blakemore 1982) Like the structures in ARB-1 polyP granules aretranslucent in visible light (Spring et al 1994) dark (electron dense) in the TEM and oftenhave a mottled appearance (Harold 1966) The P-rich inclusions in ARB-1 cells stained
400 B L Cox et al
with methylene blue (Murray and Robinson 1994) which is suggestive of the presence ofpolyP but not con rmatory
PolyP is composed of linear chains of tetrahedral phosphate residues linked throughcommon oxygen atoms by phosphoanhydrid e bonds At physiological pH each unit carriesa monovalent negative charge giving polyP the property of having a large cation exchangecapacity The binding energy of polyP allows it to sequester Ca2C and Mg2C (Reusch 2000)consistent with the Ca2C and Mg2C signals observed in the EDS spectra of the ARB-1inclusions (Figure 6a 6d) The binding of these metals to the polyP granules contributesto their high electron density PolyP is often associated with polyhydroxybutyrat e (PHB)PHBs solvate (dissolve) cations and associate withpolyP to form Ca2C selective ion channelsacross plasma membranes (Reusch 2000)
The presence of polyP bodies is common in bacteria often resulting as a bacterialresponse to stress and they may play a role in cell motility quorum sensing and bio lmformation (Kornberg and Fraley 2000) Sulfate starvation is known to lead to rapid andmassive accumulation of polyP in some bacteria (Stanier et al 1986) When cells regainaccess to sulfate 32P uptake studies showed that PO3iexcl
4 present in the polyP bodies wasincorporated into nucleic acids PolyP is synthesized from ATP by the enzyme polyP kinasePolyP granules have been reported to be associated with nucleic acids (Harold 1966) butit is not clear whether the association occurs in vivo or after extraction This associationwith nucleic acids would be consistent with staining patterns seen when ARB-1 cells wereexamined with a uorescent livedead stain (nucleic acid stain) displaying a double-lobedstructure in all of the cells (Figure 9) This dumbbell shape appears to extend inside andacross the two phosphorus inclusions
The physiological function of the P inclusions in ARB-1 is not yet known but fromother cell systems several possibilities can be considered These inclusions might serveas a phosphorus reserve for the biosynthesis of nucleic acids and phospholipids underconditions of phosphorus starvation (Harold 1966) Still another possibility is buoyancyregulation Cells can vary their density by as much as 7 by incorporating substances intoinclusions (Guerrero et al 1985) A more speculative possibility is that ARB-1 cells canderive energy from polyP Acinetobacter johnsonii 210a is known to couple the excretion ofa metal phosphate chelate and HC generating a proton motive force (VanVeen et al 1994)A johnsonii takes up P from the aquatic environment by using two Pi transport systemsand stores it as intracellular granules of polyP
Sulfur (S) Inclusions
Cells of ARB-1 contained 5 to 10 spherical S-rich inclusions (Figures 1d) These rangedin size from approximately 100 to 700 nm although most of the S-rich inclusions in asingle cell were close to the same size and were located at one end of the cell oppositethe side where the magnetosomes were located and agellar bundles attached Like theP-rich inclusions the S-rich inclusions appeared to be enclosed in a coating or membrane(Figure 1d) EDS analysis from the ESEM showed these structures contained mostly S withsome carbon and oxygen (Figure 6e) EDS analysis from the TEM also showed S but evenhigher counts for Ca and P (Figure 6b) Beam spreading for the two methods is within aradius of 200 nm The X-rays produced by the 200-keV TEM beam sample the entire verticalextent of the cell unlike the 20 keV SEM beam which loses most of its energy Thereforethe EDS spectra collected at the TEM likely have sampled elements from the cytoplasmand cell membrane components both above and below the inclusion as well as from theP-rich inclusions nearby Other explanations are that hydrated sulfur inclusions changedtheir composition upon dehydration or that these two samples collected at different times
Organization of Inclusions in Magnetococci 401
FIGURE 9 Fluorescence micrograph of an ARB-1 cell suspension treated with Bacliteviability dyes showing that the nucleic acid staining pattern corresponds with the locationof the P-rich inclusions Baclite stains nucleic acids with cytonine (all cells) and propidiumiodate (dead or damaged cells) Bright double-lobed patterns are from ldquoliverdquo cells
of the day and the year may re ect a seasonal diurnal or cell cycle variability in the relativeproportions of the elements
Sulfur inclusions have been found in other uncultured magnetotactic bacteria includ-ing Magnetobacterium bavaricum (Spring et al 1993) Bilophococcus magnetotacticum(Moench 1988) and others (Iida and Akai 1996) and cultured species including the vibrioidstrains MV-1 and MV-2 the coccus strain MC-1 and the spirillum strain MV-4 (Bazylinskiand Frankel 2000) However only in Bilophococcus magnetotacticum was there a similarpolarization of sulfur-containing inclusions from the clustered magnetosomes within thecell (Moench 1988)
It seems likely that ARB-1 oxidizes H2S or other reduced sulfur compounds as anenergy source depositing ldquoelementalrdquo sulfur in intracellular inclusions like many othersul de-oxidizing bacteria including the anaerobic phototrophic purple sulfur bacteria suchas Chromatium vinosum and Thiocapsa roseopersicina the nonsulfur bacteria Beggiatoaspp and Thiothrix spp (Shively 1974 Friedrich 1998) and the magnetotactic bacteriamentioned before In these organisms the presence of sulfur globules is transitory andtakes place when the growth medium contains sul de andor thiosulfate When the sul deis completely utilized the sulfur is converted to sulfate (Stanier et al 1986)
Elemental S in sulfur globules is typically in a liquid state and analysis of in vivoglobules from some members of the purple and green sulfur bacteria by X-ray absorptionnear edge spectroscopy (Prange et al 1999) indicated the sulfur was in a chain structure
402 B L Cox et al
FIGURE 10 Phylogenetic position of six SSU rRNA gene clones related to previouslystudied MC This phylogenetic tree was rooted with Escherichia coli and constructed bythe neighbor joining method with the Kimura 2-parameter model for nucleotide change Amask including all nonambiguously aligned positions for 1427 nucleotides spanning nearlythe full length of the SSU rRNA gene was included Bootstrap values (100 replications)generated by the neighbor joining method are shown above relevant nodes and bootstrapvalues generated by maximum-parsimony analysis are shown below relevant nodes Onlybootstrap values above 50 are shown GenBank accession numbers are MC-CS92 X81182MC-pa119 Y13214 MC-CS81 X81184 MC-TB12 X81183 MC-MP17 X80996 MC-PA11 Y13212 MC12 L06456 partial sequence extended with data from MC-1 genomeRhizobium leguminosarum U31074 Agrobacterium tumefaciens D13294 Rhodobactersphaeroides X53854 Paracoccus denitri cans X59159 Zymomonas mobilis AF281034Sphingomonas paucimobilis D84528 Caulobacter crescentus M83799 Magnetospiril-lum magnetotacticum strain AMB1 D17514 S88429 Magnetospirillum magnetotacticumM10110 Rhodospirillum fulvum M59065 Magnetospirillum gryphiswaldense Y10109Magnetic Vibrio MV-1 LO6455 Oceanospirillum pusillum AB000482 Rhodospirillumrubrum D30778 Roseococcus thiosulfatophilus X72908 Escherichia coli J01695
Organization of Inclusions in Magnetococci 403
and was possibly complexed with water and organic residues If the S globules in ARB-1cells are analogous to those in the purple sulfur bacteria Chromatium vinosum and Thio-capsa roseopersicina they may contain a signi cant protein content Sulfur globules fromthe anaerobic phototrophs C vinosum and T roseopersicina were found to contain upto 52 protein that formed an envelope enclosing the S globules (Brune 1995) Theseproteins were isolated and analyzed and found to be glycine and tyrosine-rich structuralproteins which were homologous for the two species suggesting they are highly con-served (Brune 1995) Phylogenetically C vinosum and T roseopersicina are members ofthe deg -Proteobacteria whereas MC are members of the reg-Proteobacteria To our knowledgemagnetotactic bacteria may be the only members of the reg-Proteobacteria that make internalsulfur globules
Phylogenetic Af liation
We wished to detemine whether ARB-1 was a single species or a mixed population of MCToward that end 45 SSU rRNA genes were cloned from partially puri ed ARB-1 cells30 of which displayed similar restriction fragment length patterns (RFLP) to previouslycharacterized MC (data not shown) The nding that 15 patterns were not related to pre-viously characterized MC was not surprising given that the initial sample was not a pureculture but rather a population magnetically enriched from a sample originating from arelatively large vertical column containing both sediments and overlying water Fourteenof the MC-related clones representing different RFLP pattern groups were chosen for par-tial sequencing 10 of these sequences showed high similarity to previously characterizedMC Six of these sequences were phylogenetically distinct and were fully bidirectionallysequenced
ARB-1 MC SSU rRNA genes formed a monophyletic cluster related to but dis-tinct from other MC (Figure 10) Within the cluster the similarity between SSU rRNAgene sequences ranged from 97 (MC-H1arb and MC-g6arb) to 92 (MC-h1arb andMC-g2arb) The nearest neighbor to the ARB-1 cluster MC-CS92 was ca 89 similarand the nearest cultured relative MC-1 was ca 88 similar to the ARB-1 cluster As arule of thumb for SSU rRNA genes gt97 similarity is considered the same species andgt93ndash95 similarity is considered the same genus (Stackebrandt and Goebel 1994) Thesedata strongly suggest that ARB-1 represents a population of different diverse MC Therewas signi cant variation within the ARB-1 cluster and therefore we do not consider ARB-1to consist of bacteria of a single species or genus Correlations if any between phylogeneticand phenotypic variations in the ARB-1 cells have yet to be determined
Summary
We have partially characterized a population of magnetotactic cocci (MC) referred to asARB-1 which was particularly abundant in iron-rich sediments of a small eutrophic lake andmorphologically and structurally similar to an uncultured species of MC called Bilophococ-cus magnetotacticus The latter organism was found in large numbers in a sewage treatmentpond Cells of ARB-1 had a relatively wide size range of 1 to 4 sup1m They were motileGram-negative and bilophotrichously agellated Cells of this uncultured magnetotacticcoccus contained three types of intracellular inclusions that appeared to be in a consistentarrangement within the cell The magnetosomes contained crystals of magnetite and wereeither in chains andor polar clumps at the end of the cell where the agellar bundles werelocated Large P-rich bodies occupied the majority of the cell volume and appeared to sepa-rate the S-containing inclusions from the magnetosomes The presence of these large P-rich
404 B L Cox et al
inclusions might be an indication of cell stress or may be important in cell buoyancy or inenergy generation The presence of S-rich inclusions located at the end of the cell oppositethe magnetosomes strongly suggests that ARB-1 cells are sul de-oxidizing bacteria andthat the S might be useful as a stored electron donor when cells travel from sul de-rich tosul de-depleted zones in the water column The wide range of the cell sizes the differentmagnetosome arrangements and the large number of different retrieved 16S rRNA genesequences suggest that ARB-1 represents a population of at least several different MC 16SrRNA sequences obtained from cells of ARB-1 appeared to form a monophyletic clusterrelated to but distinct from other uncultured and cultured MC Future work using uores-cent oligonucleotide probes microscopy and physiological measurements should clarifyrelationships between phylogeny cell morphology and organization of inclusions
References
Balkwill DL Maratea D Blakemore RP 1980 Ultrastructure of a magnetotactic spirillum J Bacteriol141(3)1399ndash1408
Bazylinski DA Frankel RB 2000 Biologically controlled mineralization of magnetic iron mineralsby magnetotactic bacteria In Lovley DR editor Environmental microbe-metal interactionsWashington DC ASM Press p 109ndash143
Bazylinski DA Frankel RB Jannasch HW 1988 Anaerobic magnetite production by a marinemagnetotactic bacterium Nature 334518ndash519
Beveridge T 1999 Structures of gram-negative cell walls and their derived membrane vesiclesJ Bacteriol 181(6)4725ndash4733
Beveridge TJ Popkin TJ Cole RM 1994 Electron microscopy In Gerhardt P editor Methods forgeneral and molecular bacteriology Washington DC American Society for Microbiology p 25
Blakemore R 1975 Magnetotactic bacteria Science 190377ndash379Blakemore RP 1982 Magnetotactic bacteria Ann Rev Microbiol 36217ndash238Blakemore RP Maratea D Wolfe RS 1979 Isolation and pure culture of a freshwater magnetic
spirillum in chemically de ned medium J Bacteriol 140(2)720ndash729Blakemore RP Short KA Bazylinski DA Rosenblatt C Frankel RB 1985 Microaerobic conditions
are required for magnetite formation within Aquaspirillum magnetotacticum Geomicrobiol J4(1)53ndash71
Brune DC 1995 Isolation and characterization of sulfur globule proteins from Chromatium vinosumand Thiocapsa roseopersicina Arch Microbiol 163(1)391ndash399
Butler RF Banerjee SK 1975 Theoretical single-domain grain size range in magnetite and titano-magnetite J Geophys Res 804049ndash4058
Buwalda JP 1940 Geology of the Raymond Basin report to Pasadena Water DepartmentCrook R Allen CR Kamb B Payne CM Procter RJ 1987 Recent reverse faulting in the Transverse
Ranges California Pasadena CA US Geol Surv Prof Paper 133927ndash63Devouard B Posfai M Hua X Bazylinski DA Frankel RB Buseck PR 1998 Magnetite from
magnetotactic bacteria size distribution and twinning Am Mineral 831387ndash1398Frankel RB Bazylinski DA Johnson MS Taylor BL 1997 Magneto-Aerotaxis in marine coccoid
bacteria Biophys J 73994ndash1000Friedrich CG 1998 Physiology and genetics of sulfur-oxidizing bacteria Adv Microbial Physiol
39235ndash289Giovannoni SJ 1991 The polymerase chain reaction In Stackebrandt E Goodfellow M editors
Sequencing and hybridization techniques in bacterial systematics New York John Wiley andSons p 177ndash201
Guerrero R Pedros-Alio C Schmidt TM Mas J 1985 A survey of buoyant density of microorganismsin pure cultures and natural samples Microbiologia Sept 1(1ndash2)53ndash65
Harold FM 1966 Inorganic polyphosphates in biology structure metabolism and function BacteriolRev 30(4)772ndash794
Organization of Inclusions in Magnetococci 405
Iida A Akai J 1996 Crystalline sulfur inclusions in magnetotactic bacteria Sci Rep Nigata Univ SerE 1135ndash42
Kornberg A Fraley CD 2000 Inorganic polyphosphate a molecular fossil come to life ASM News66(5)275ndash280
Lins U Farina M 1999 Phosphorus-rich granules in uncultured magnetotactic bacteria FEMSMicrobiol Lett 172(2)23ndash28
Mann S Sparks NHC Frankel RB Bazylinski DA Jannasch HW 1990 Biomineralization of ferri-magnetic greigite (Fe3O4 and iron pyrite (FeS2 ) in a magnetotactic bacterium Nature 343258ndash
261Matsunaga T Sakaguchi T Tadokoro F 1991 Magnetite formation by a magnetic bacterium capable
of growing aerobically Appl Microbiol Biotechnol 35651ndash655Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993a Electron microscopy
study of magnetosomes in a cultured coccoid magnetotactic bacterium Proc Roy Soc Lond B251231ndash236
Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993b Electron microscopystudy of magnetosomes in two cultured vibrioid magnetotactic bacteria Proc Roy Soc Lond B251237ndash242
Moench TT 1988 Bilophococcus magnetotacticus gen nov sp nov a motile magnetic coccus Antvan Leeuwenhoek 54483ndash496
Moench TT Konetzka WA 1978 A novel method for the isolation and study of a magnetotacticbacterium Arch Microbiol 119203ndash212
Murray RGE Robinow CF 1994 Optical Microscopy In Gerhardt P editor Methods for generaland molecular bacteriology Washington DC American Society for Microbiology p 35ndash36
Prange A Arzberger I Engemann C Modrow H Schumann O Truper HG Steudel R Dahl C andHormes J 1999 In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteriaby X-ray absorption near edge spectroscopy Biochim Biophys Acta 1428446ndash454
Rappe MS Kemp PF Giovannoni SJ 1997 Phylogenetic diversity of marine coastal picoplankton16S rRNA genes cloned from the continental shelf off Cape Hatteras North Carolina LimnolOceanogr 42811ndash826
Reusch RN 2000 Transmembrane ion transport by polyphosphatepoly-(R)-3-hydroxybutyratecomplexes Biochemistry (Moscow) 65(3)335ndash352
Sakaguchi T Burgess JG Matsunaga T 1993 Magnetite formation by a sulphate-reducing bacteriumNature 36547ndash49
Schleifer KHSchuler D Spring S Weizenegger M Amann R 1991 The genus Magnetospirillum gennov description of Magnetospirillum gryphiswaldense sp nov and transfer of Aquaspirillummagnetotacticum to Magnetospirillum magnetotacticum comb nov System Appl Microbiol14379ndash385
Schuler D 1999 Formation of magnetosomes in magnetotactic bacteria J Mol Microbiol Biotechnol1(1)79ndash86
Schuler D Spring S Bazylinski DA 1999 Improved technique for the isolation of magnetotactic spir-illa from a freshwater sediment and their phylogenetic characterization System Appl Microbiol22466ndash471
Shively JM 1974 Inclusion bodies of prokaryotes Ann Rev Microbiol 28167ndash187Spring S Amann R Ludwig W Schleifer KH van Gemerden H Petersen N 1993 Dominating role
of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment ApplEnviron Microbiol 59(8)2397ndash2403
Spring S Amann R Ludwig W Schleifer KH Schuler D Poralla K Petersen N 1994 Phylogeneticanalysis of uncultured magnetotactic baceria from the Alpha-subclass of Proteobacteria SystemAppl Microbiol 17501ndash508
Stackebrandt E Goebel BM 1994 Taxonomic note a place for DNA-DNA reassociation and 16SrRNA sequence analysis in the present species de nition in bacteriology Int J System Bacteriol44846ndash849
Stanier RY Ingraham JL Wheelis ML Painter PR 1986 The microbial world 5th edition EnglewoodCliffs NJ Prentice Hall p 179
406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35

400 B L Cox et al
with methylene blue (Murray and Robinson 1994) which is suggestive of the presence ofpolyP but not con rmatory
PolyP is composed of linear chains of tetrahedral phosphate residues linked throughcommon oxygen atoms by phosphoanhydrid e bonds At physiological pH each unit carriesa monovalent negative charge giving polyP the property of having a large cation exchangecapacity The binding energy of polyP allows it to sequester Ca2C and Mg2C (Reusch 2000)consistent with the Ca2C and Mg2C signals observed in the EDS spectra of the ARB-1inclusions (Figure 6a 6d) The binding of these metals to the polyP granules contributesto their high electron density PolyP is often associated with polyhydroxybutyrat e (PHB)PHBs solvate (dissolve) cations and associate withpolyP to form Ca2C selective ion channelsacross plasma membranes (Reusch 2000)
The presence of polyP bodies is common in bacteria often resulting as a bacterialresponse to stress and they may play a role in cell motility quorum sensing and bio lmformation (Kornberg and Fraley 2000) Sulfate starvation is known to lead to rapid andmassive accumulation of polyP in some bacteria (Stanier et al 1986) When cells regainaccess to sulfate 32P uptake studies showed that PO3iexcl
4 present in the polyP bodies wasincorporated into nucleic acids PolyP is synthesized from ATP by the enzyme polyP kinasePolyP granules have been reported to be associated with nucleic acids (Harold 1966) butit is not clear whether the association occurs in vivo or after extraction This associationwith nucleic acids would be consistent with staining patterns seen when ARB-1 cells wereexamined with a uorescent livedead stain (nucleic acid stain) displaying a double-lobedstructure in all of the cells (Figure 9) This dumbbell shape appears to extend inside andacross the two phosphorus inclusions
The physiological function of the P inclusions in ARB-1 is not yet known but fromother cell systems several possibilities can be considered These inclusions might serveas a phosphorus reserve for the biosynthesis of nucleic acids and phospholipids underconditions of phosphorus starvation (Harold 1966) Still another possibility is buoyancyregulation Cells can vary their density by as much as 7 by incorporating substances intoinclusions (Guerrero et al 1985) A more speculative possibility is that ARB-1 cells canderive energy from polyP Acinetobacter johnsonii 210a is known to couple the excretion ofa metal phosphate chelate and HC generating a proton motive force (VanVeen et al 1994)A johnsonii takes up P from the aquatic environment by using two Pi transport systemsand stores it as intracellular granules of polyP
Sulfur (S) Inclusions
Cells of ARB-1 contained 5 to 10 spherical S-rich inclusions (Figures 1d) These rangedin size from approximately 100 to 700 nm although most of the S-rich inclusions in asingle cell were close to the same size and were located at one end of the cell oppositethe side where the magnetosomes were located and agellar bundles attached Like theP-rich inclusions the S-rich inclusions appeared to be enclosed in a coating or membrane(Figure 1d) EDS analysis from the ESEM showed these structures contained mostly S withsome carbon and oxygen (Figure 6e) EDS analysis from the TEM also showed S but evenhigher counts for Ca and P (Figure 6b) Beam spreading for the two methods is within aradius of 200 nm The X-rays produced by the 200-keV TEM beam sample the entire verticalextent of the cell unlike the 20 keV SEM beam which loses most of its energy Thereforethe EDS spectra collected at the TEM likely have sampled elements from the cytoplasmand cell membrane components both above and below the inclusion as well as from theP-rich inclusions nearby Other explanations are that hydrated sulfur inclusions changedtheir composition upon dehydration or that these two samples collected at different times
Organization of Inclusions in Magnetococci 401
FIGURE 9 Fluorescence micrograph of an ARB-1 cell suspension treated with Bacliteviability dyes showing that the nucleic acid staining pattern corresponds with the locationof the P-rich inclusions Baclite stains nucleic acids with cytonine (all cells) and propidiumiodate (dead or damaged cells) Bright double-lobed patterns are from ldquoliverdquo cells
of the day and the year may re ect a seasonal diurnal or cell cycle variability in the relativeproportions of the elements
Sulfur inclusions have been found in other uncultured magnetotactic bacteria includ-ing Magnetobacterium bavaricum (Spring et al 1993) Bilophococcus magnetotacticum(Moench 1988) and others (Iida and Akai 1996) and cultured species including the vibrioidstrains MV-1 and MV-2 the coccus strain MC-1 and the spirillum strain MV-4 (Bazylinskiand Frankel 2000) However only in Bilophococcus magnetotacticum was there a similarpolarization of sulfur-containing inclusions from the clustered magnetosomes within thecell (Moench 1988)
It seems likely that ARB-1 oxidizes H2S or other reduced sulfur compounds as anenergy source depositing ldquoelementalrdquo sulfur in intracellular inclusions like many othersul de-oxidizing bacteria including the anaerobic phototrophic purple sulfur bacteria suchas Chromatium vinosum and Thiocapsa roseopersicina the nonsulfur bacteria Beggiatoaspp and Thiothrix spp (Shively 1974 Friedrich 1998) and the magnetotactic bacteriamentioned before In these organisms the presence of sulfur globules is transitory andtakes place when the growth medium contains sul de andor thiosulfate When the sul deis completely utilized the sulfur is converted to sulfate (Stanier et al 1986)
Elemental S in sulfur globules is typically in a liquid state and analysis of in vivoglobules from some members of the purple and green sulfur bacteria by X-ray absorptionnear edge spectroscopy (Prange et al 1999) indicated the sulfur was in a chain structure
402 B L Cox et al
FIGURE 10 Phylogenetic position of six SSU rRNA gene clones related to previouslystudied MC This phylogenetic tree was rooted with Escherichia coli and constructed bythe neighbor joining method with the Kimura 2-parameter model for nucleotide change Amask including all nonambiguously aligned positions for 1427 nucleotides spanning nearlythe full length of the SSU rRNA gene was included Bootstrap values (100 replications)generated by the neighbor joining method are shown above relevant nodes and bootstrapvalues generated by maximum-parsimony analysis are shown below relevant nodes Onlybootstrap values above 50 are shown GenBank accession numbers are MC-CS92 X81182MC-pa119 Y13214 MC-CS81 X81184 MC-TB12 X81183 MC-MP17 X80996 MC-PA11 Y13212 MC12 L06456 partial sequence extended with data from MC-1 genomeRhizobium leguminosarum U31074 Agrobacterium tumefaciens D13294 Rhodobactersphaeroides X53854 Paracoccus denitri cans X59159 Zymomonas mobilis AF281034Sphingomonas paucimobilis D84528 Caulobacter crescentus M83799 Magnetospiril-lum magnetotacticum strain AMB1 D17514 S88429 Magnetospirillum magnetotacticumM10110 Rhodospirillum fulvum M59065 Magnetospirillum gryphiswaldense Y10109Magnetic Vibrio MV-1 LO6455 Oceanospirillum pusillum AB000482 Rhodospirillumrubrum D30778 Roseococcus thiosulfatophilus X72908 Escherichia coli J01695
Organization of Inclusions in Magnetococci 403
and was possibly complexed with water and organic residues If the S globules in ARB-1cells are analogous to those in the purple sulfur bacteria Chromatium vinosum and Thio-capsa roseopersicina they may contain a signi cant protein content Sulfur globules fromthe anaerobic phototrophs C vinosum and T roseopersicina were found to contain upto 52 protein that formed an envelope enclosing the S globules (Brune 1995) Theseproteins were isolated and analyzed and found to be glycine and tyrosine-rich structuralproteins which were homologous for the two species suggesting they are highly con-served (Brune 1995) Phylogenetically C vinosum and T roseopersicina are members ofthe deg -Proteobacteria whereas MC are members of the reg-Proteobacteria To our knowledgemagnetotactic bacteria may be the only members of the reg-Proteobacteria that make internalsulfur globules
Phylogenetic Af liation
We wished to detemine whether ARB-1 was a single species or a mixed population of MCToward that end 45 SSU rRNA genes were cloned from partially puri ed ARB-1 cells30 of which displayed similar restriction fragment length patterns (RFLP) to previouslycharacterized MC (data not shown) The nding that 15 patterns were not related to pre-viously characterized MC was not surprising given that the initial sample was not a pureculture but rather a population magnetically enriched from a sample originating from arelatively large vertical column containing both sediments and overlying water Fourteenof the MC-related clones representing different RFLP pattern groups were chosen for par-tial sequencing 10 of these sequences showed high similarity to previously characterizedMC Six of these sequences were phylogenetically distinct and were fully bidirectionallysequenced
ARB-1 MC SSU rRNA genes formed a monophyletic cluster related to but dis-tinct from other MC (Figure 10) Within the cluster the similarity between SSU rRNAgene sequences ranged from 97 (MC-H1arb and MC-g6arb) to 92 (MC-h1arb andMC-g2arb) The nearest neighbor to the ARB-1 cluster MC-CS92 was ca 89 similarand the nearest cultured relative MC-1 was ca 88 similar to the ARB-1 cluster As arule of thumb for SSU rRNA genes gt97 similarity is considered the same species andgt93ndash95 similarity is considered the same genus (Stackebrandt and Goebel 1994) Thesedata strongly suggest that ARB-1 represents a population of different diverse MC Therewas signi cant variation within the ARB-1 cluster and therefore we do not consider ARB-1to consist of bacteria of a single species or genus Correlations if any between phylogeneticand phenotypic variations in the ARB-1 cells have yet to be determined
Summary
We have partially characterized a population of magnetotactic cocci (MC) referred to asARB-1 which was particularly abundant in iron-rich sediments of a small eutrophic lake andmorphologically and structurally similar to an uncultured species of MC called Bilophococ-cus magnetotacticus The latter organism was found in large numbers in a sewage treatmentpond Cells of ARB-1 had a relatively wide size range of 1 to 4 sup1m They were motileGram-negative and bilophotrichously agellated Cells of this uncultured magnetotacticcoccus contained three types of intracellular inclusions that appeared to be in a consistentarrangement within the cell The magnetosomes contained crystals of magnetite and wereeither in chains andor polar clumps at the end of the cell where the agellar bundles werelocated Large P-rich bodies occupied the majority of the cell volume and appeared to sepa-rate the S-containing inclusions from the magnetosomes The presence of these large P-rich
404 B L Cox et al
inclusions might be an indication of cell stress or may be important in cell buoyancy or inenergy generation The presence of S-rich inclusions located at the end of the cell oppositethe magnetosomes strongly suggests that ARB-1 cells are sul de-oxidizing bacteria andthat the S might be useful as a stored electron donor when cells travel from sul de-rich tosul de-depleted zones in the water column The wide range of the cell sizes the differentmagnetosome arrangements and the large number of different retrieved 16S rRNA genesequences suggest that ARB-1 represents a population of at least several different MC 16SrRNA sequences obtained from cells of ARB-1 appeared to form a monophyletic clusterrelated to but distinct from other uncultured and cultured MC Future work using uores-cent oligonucleotide probes microscopy and physiological measurements should clarifyrelationships between phylogeny cell morphology and organization of inclusions
References
Balkwill DL Maratea D Blakemore RP 1980 Ultrastructure of a magnetotactic spirillum J Bacteriol141(3)1399ndash1408
Bazylinski DA Frankel RB 2000 Biologically controlled mineralization of magnetic iron mineralsby magnetotactic bacteria In Lovley DR editor Environmental microbe-metal interactionsWashington DC ASM Press p 109ndash143
Bazylinski DA Frankel RB Jannasch HW 1988 Anaerobic magnetite production by a marinemagnetotactic bacterium Nature 334518ndash519
Beveridge T 1999 Structures of gram-negative cell walls and their derived membrane vesiclesJ Bacteriol 181(6)4725ndash4733
Beveridge TJ Popkin TJ Cole RM 1994 Electron microscopy In Gerhardt P editor Methods forgeneral and molecular bacteriology Washington DC American Society for Microbiology p 25
Blakemore R 1975 Magnetotactic bacteria Science 190377ndash379Blakemore RP 1982 Magnetotactic bacteria Ann Rev Microbiol 36217ndash238Blakemore RP Maratea D Wolfe RS 1979 Isolation and pure culture of a freshwater magnetic
spirillum in chemically de ned medium J Bacteriol 140(2)720ndash729Blakemore RP Short KA Bazylinski DA Rosenblatt C Frankel RB 1985 Microaerobic conditions
are required for magnetite formation within Aquaspirillum magnetotacticum Geomicrobiol J4(1)53ndash71
Brune DC 1995 Isolation and characterization of sulfur globule proteins from Chromatium vinosumand Thiocapsa roseopersicina Arch Microbiol 163(1)391ndash399
Butler RF Banerjee SK 1975 Theoretical single-domain grain size range in magnetite and titano-magnetite J Geophys Res 804049ndash4058
Buwalda JP 1940 Geology of the Raymond Basin report to Pasadena Water DepartmentCrook R Allen CR Kamb B Payne CM Procter RJ 1987 Recent reverse faulting in the Transverse
Ranges California Pasadena CA US Geol Surv Prof Paper 133927ndash63Devouard B Posfai M Hua X Bazylinski DA Frankel RB Buseck PR 1998 Magnetite from
magnetotactic bacteria size distribution and twinning Am Mineral 831387ndash1398Frankel RB Bazylinski DA Johnson MS Taylor BL 1997 Magneto-Aerotaxis in marine coccoid
bacteria Biophys J 73994ndash1000Friedrich CG 1998 Physiology and genetics of sulfur-oxidizing bacteria Adv Microbial Physiol
39235ndash289Giovannoni SJ 1991 The polymerase chain reaction In Stackebrandt E Goodfellow M editors
Sequencing and hybridization techniques in bacterial systematics New York John Wiley andSons p 177ndash201
Guerrero R Pedros-Alio C Schmidt TM Mas J 1985 A survey of buoyant density of microorganismsin pure cultures and natural samples Microbiologia Sept 1(1ndash2)53ndash65
Harold FM 1966 Inorganic polyphosphates in biology structure metabolism and function BacteriolRev 30(4)772ndash794
Organization of Inclusions in Magnetococci 405
Iida A Akai J 1996 Crystalline sulfur inclusions in magnetotactic bacteria Sci Rep Nigata Univ SerE 1135ndash42
Kornberg A Fraley CD 2000 Inorganic polyphosphate a molecular fossil come to life ASM News66(5)275ndash280
Lins U Farina M 1999 Phosphorus-rich granules in uncultured magnetotactic bacteria FEMSMicrobiol Lett 172(2)23ndash28
Mann S Sparks NHC Frankel RB Bazylinski DA Jannasch HW 1990 Biomineralization of ferri-magnetic greigite (Fe3O4 and iron pyrite (FeS2 ) in a magnetotactic bacterium Nature 343258ndash
261Matsunaga T Sakaguchi T Tadokoro F 1991 Magnetite formation by a magnetic bacterium capable
of growing aerobically Appl Microbiol Biotechnol 35651ndash655Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993a Electron microscopy
study of magnetosomes in a cultured coccoid magnetotactic bacterium Proc Roy Soc Lond B251231ndash236
Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993b Electron microscopystudy of magnetosomes in two cultured vibrioid magnetotactic bacteria Proc Roy Soc Lond B251237ndash242
Moench TT 1988 Bilophococcus magnetotacticus gen nov sp nov a motile magnetic coccus Antvan Leeuwenhoek 54483ndash496
Moench TT Konetzka WA 1978 A novel method for the isolation and study of a magnetotacticbacterium Arch Microbiol 119203ndash212
Murray RGE Robinow CF 1994 Optical Microscopy In Gerhardt P editor Methods for generaland molecular bacteriology Washington DC American Society for Microbiology p 35ndash36
Prange A Arzberger I Engemann C Modrow H Schumann O Truper HG Steudel R Dahl C andHormes J 1999 In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteriaby X-ray absorption near edge spectroscopy Biochim Biophys Acta 1428446ndash454
Rappe MS Kemp PF Giovannoni SJ 1997 Phylogenetic diversity of marine coastal picoplankton16S rRNA genes cloned from the continental shelf off Cape Hatteras North Carolina LimnolOceanogr 42811ndash826
Reusch RN 2000 Transmembrane ion transport by polyphosphatepoly-(R)-3-hydroxybutyratecomplexes Biochemistry (Moscow) 65(3)335ndash352
Sakaguchi T Burgess JG Matsunaga T 1993 Magnetite formation by a sulphate-reducing bacteriumNature 36547ndash49
Schleifer KHSchuler D Spring S Weizenegger M Amann R 1991 The genus Magnetospirillum gennov description of Magnetospirillum gryphiswaldense sp nov and transfer of Aquaspirillummagnetotacticum to Magnetospirillum magnetotacticum comb nov System Appl Microbiol14379ndash385
Schuler D 1999 Formation of magnetosomes in magnetotactic bacteria J Mol Microbiol Biotechnol1(1)79ndash86
Schuler D Spring S Bazylinski DA 1999 Improved technique for the isolation of magnetotactic spir-illa from a freshwater sediment and their phylogenetic characterization System Appl Microbiol22466ndash471
Shively JM 1974 Inclusion bodies of prokaryotes Ann Rev Microbiol 28167ndash187Spring S Amann R Ludwig W Schleifer KH van Gemerden H Petersen N 1993 Dominating role
of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment ApplEnviron Microbiol 59(8)2397ndash2403
Spring S Amann R Ludwig W Schleifer KH Schuler D Poralla K Petersen N 1994 Phylogeneticanalysis of uncultured magnetotactic baceria from the Alpha-subclass of Proteobacteria SystemAppl Microbiol 17501ndash508
Stackebrandt E Goebel BM 1994 Taxonomic note a place for DNA-DNA reassociation and 16SrRNA sequence analysis in the present species de nition in bacteriology Int J System Bacteriol44846ndash849
Stanier RY Ingraham JL Wheelis ML Painter PR 1986 The microbial world 5th edition EnglewoodCliffs NJ Prentice Hall p 179
406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35

Organization of Inclusions in Magnetococci 401
FIGURE 9 Fluorescence micrograph of an ARB-1 cell suspension treated with Bacliteviability dyes showing that the nucleic acid staining pattern corresponds with the locationof the P-rich inclusions Baclite stains nucleic acids with cytonine (all cells) and propidiumiodate (dead or damaged cells) Bright double-lobed patterns are from ldquoliverdquo cells
of the day and the year may re ect a seasonal diurnal or cell cycle variability in the relativeproportions of the elements
Sulfur inclusions have been found in other uncultured magnetotactic bacteria includ-ing Magnetobacterium bavaricum (Spring et al 1993) Bilophococcus magnetotacticum(Moench 1988) and others (Iida and Akai 1996) and cultured species including the vibrioidstrains MV-1 and MV-2 the coccus strain MC-1 and the spirillum strain MV-4 (Bazylinskiand Frankel 2000) However only in Bilophococcus magnetotacticum was there a similarpolarization of sulfur-containing inclusions from the clustered magnetosomes within thecell (Moench 1988)
It seems likely that ARB-1 oxidizes H2S or other reduced sulfur compounds as anenergy source depositing ldquoelementalrdquo sulfur in intracellular inclusions like many othersul de-oxidizing bacteria including the anaerobic phototrophic purple sulfur bacteria suchas Chromatium vinosum and Thiocapsa roseopersicina the nonsulfur bacteria Beggiatoaspp and Thiothrix spp (Shively 1974 Friedrich 1998) and the magnetotactic bacteriamentioned before In these organisms the presence of sulfur globules is transitory andtakes place when the growth medium contains sul de andor thiosulfate When the sul deis completely utilized the sulfur is converted to sulfate (Stanier et al 1986)
Elemental S in sulfur globules is typically in a liquid state and analysis of in vivoglobules from some members of the purple and green sulfur bacteria by X-ray absorptionnear edge spectroscopy (Prange et al 1999) indicated the sulfur was in a chain structure
402 B L Cox et al
FIGURE 10 Phylogenetic position of six SSU rRNA gene clones related to previouslystudied MC This phylogenetic tree was rooted with Escherichia coli and constructed bythe neighbor joining method with the Kimura 2-parameter model for nucleotide change Amask including all nonambiguously aligned positions for 1427 nucleotides spanning nearlythe full length of the SSU rRNA gene was included Bootstrap values (100 replications)generated by the neighbor joining method are shown above relevant nodes and bootstrapvalues generated by maximum-parsimony analysis are shown below relevant nodes Onlybootstrap values above 50 are shown GenBank accession numbers are MC-CS92 X81182MC-pa119 Y13214 MC-CS81 X81184 MC-TB12 X81183 MC-MP17 X80996 MC-PA11 Y13212 MC12 L06456 partial sequence extended with data from MC-1 genomeRhizobium leguminosarum U31074 Agrobacterium tumefaciens D13294 Rhodobactersphaeroides X53854 Paracoccus denitri cans X59159 Zymomonas mobilis AF281034Sphingomonas paucimobilis D84528 Caulobacter crescentus M83799 Magnetospiril-lum magnetotacticum strain AMB1 D17514 S88429 Magnetospirillum magnetotacticumM10110 Rhodospirillum fulvum M59065 Magnetospirillum gryphiswaldense Y10109Magnetic Vibrio MV-1 LO6455 Oceanospirillum pusillum AB000482 Rhodospirillumrubrum D30778 Roseococcus thiosulfatophilus X72908 Escherichia coli J01695
Organization of Inclusions in Magnetococci 403
and was possibly complexed with water and organic residues If the S globules in ARB-1cells are analogous to those in the purple sulfur bacteria Chromatium vinosum and Thio-capsa roseopersicina they may contain a signi cant protein content Sulfur globules fromthe anaerobic phototrophs C vinosum and T roseopersicina were found to contain upto 52 protein that formed an envelope enclosing the S globules (Brune 1995) Theseproteins were isolated and analyzed and found to be glycine and tyrosine-rich structuralproteins which were homologous for the two species suggesting they are highly con-served (Brune 1995) Phylogenetically C vinosum and T roseopersicina are members ofthe deg -Proteobacteria whereas MC are members of the reg-Proteobacteria To our knowledgemagnetotactic bacteria may be the only members of the reg-Proteobacteria that make internalsulfur globules
Phylogenetic Af liation
We wished to detemine whether ARB-1 was a single species or a mixed population of MCToward that end 45 SSU rRNA genes were cloned from partially puri ed ARB-1 cells30 of which displayed similar restriction fragment length patterns (RFLP) to previouslycharacterized MC (data not shown) The nding that 15 patterns were not related to pre-viously characterized MC was not surprising given that the initial sample was not a pureculture but rather a population magnetically enriched from a sample originating from arelatively large vertical column containing both sediments and overlying water Fourteenof the MC-related clones representing different RFLP pattern groups were chosen for par-tial sequencing 10 of these sequences showed high similarity to previously characterizedMC Six of these sequences were phylogenetically distinct and were fully bidirectionallysequenced
ARB-1 MC SSU rRNA genes formed a monophyletic cluster related to but dis-tinct from other MC (Figure 10) Within the cluster the similarity between SSU rRNAgene sequences ranged from 97 (MC-H1arb and MC-g6arb) to 92 (MC-h1arb andMC-g2arb) The nearest neighbor to the ARB-1 cluster MC-CS92 was ca 89 similarand the nearest cultured relative MC-1 was ca 88 similar to the ARB-1 cluster As arule of thumb for SSU rRNA genes gt97 similarity is considered the same species andgt93ndash95 similarity is considered the same genus (Stackebrandt and Goebel 1994) Thesedata strongly suggest that ARB-1 represents a population of different diverse MC Therewas signi cant variation within the ARB-1 cluster and therefore we do not consider ARB-1to consist of bacteria of a single species or genus Correlations if any between phylogeneticand phenotypic variations in the ARB-1 cells have yet to be determined
Summary
We have partially characterized a population of magnetotactic cocci (MC) referred to asARB-1 which was particularly abundant in iron-rich sediments of a small eutrophic lake andmorphologically and structurally similar to an uncultured species of MC called Bilophococ-cus magnetotacticus The latter organism was found in large numbers in a sewage treatmentpond Cells of ARB-1 had a relatively wide size range of 1 to 4 sup1m They were motileGram-negative and bilophotrichously agellated Cells of this uncultured magnetotacticcoccus contained three types of intracellular inclusions that appeared to be in a consistentarrangement within the cell The magnetosomes contained crystals of magnetite and wereeither in chains andor polar clumps at the end of the cell where the agellar bundles werelocated Large P-rich bodies occupied the majority of the cell volume and appeared to sepa-rate the S-containing inclusions from the magnetosomes The presence of these large P-rich
404 B L Cox et al
inclusions might be an indication of cell stress or may be important in cell buoyancy or inenergy generation The presence of S-rich inclusions located at the end of the cell oppositethe magnetosomes strongly suggests that ARB-1 cells are sul de-oxidizing bacteria andthat the S might be useful as a stored electron donor when cells travel from sul de-rich tosul de-depleted zones in the water column The wide range of the cell sizes the differentmagnetosome arrangements and the large number of different retrieved 16S rRNA genesequences suggest that ARB-1 represents a population of at least several different MC 16SrRNA sequences obtained from cells of ARB-1 appeared to form a monophyletic clusterrelated to but distinct from other uncultured and cultured MC Future work using uores-cent oligonucleotide probes microscopy and physiological measurements should clarifyrelationships between phylogeny cell morphology and organization of inclusions
References
Balkwill DL Maratea D Blakemore RP 1980 Ultrastructure of a magnetotactic spirillum J Bacteriol141(3)1399ndash1408
Bazylinski DA Frankel RB 2000 Biologically controlled mineralization of magnetic iron mineralsby magnetotactic bacteria In Lovley DR editor Environmental microbe-metal interactionsWashington DC ASM Press p 109ndash143
Bazylinski DA Frankel RB Jannasch HW 1988 Anaerobic magnetite production by a marinemagnetotactic bacterium Nature 334518ndash519
Beveridge T 1999 Structures of gram-negative cell walls and their derived membrane vesiclesJ Bacteriol 181(6)4725ndash4733
Beveridge TJ Popkin TJ Cole RM 1994 Electron microscopy In Gerhardt P editor Methods forgeneral and molecular bacteriology Washington DC American Society for Microbiology p 25
Blakemore R 1975 Magnetotactic bacteria Science 190377ndash379Blakemore RP 1982 Magnetotactic bacteria Ann Rev Microbiol 36217ndash238Blakemore RP Maratea D Wolfe RS 1979 Isolation and pure culture of a freshwater magnetic
spirillum in chemically de ned medium J Bacteriol 140(2)720ndash729Blakemore RP Short KA Bazylinski DA Rosenblatt C Frankel RB 1985 Microaerobic conditions
are required for magnetite formation within Aquaspirillum magnetotacticum Geomicrobiol J4(1)53ndash71
Brune DC 1995 Isolation and characterization of sulfur globule proteins from Chromatium vinosumand Thiocapsa roseopersicina Arch Microbiol 163(1)391ndash399
Butler RF Banerjee SK 1975 Theoretical single-domain grain size range in magnetite and titano-magnetite J Geophys Res 804049ndash4058
Buwalda JP 1940 Geology of the Raymond Basin report to Pasadena Water DepartmentCrook R Allen CR Kamb B Payne CM Procter RJ 1987 Recent reverse faulting in the Transverse
Ranges California Pasadena CA US Geol Surv Prof Paper 133927ndash63Devouard B Posfai M Hua X Bazylinski DA Frankel RB Buseck PR 1998 Magnetite from
magnetotactic bacteria size distribution and twinning Am Mineral 831387ndash1398Frankel RB Bazylinski DA Johnson MS Taylor BL 1997 Magneto-Aerotaxis in marine coccoid
bacteria Biophys J 73994ndash1000Friedrich CG 1998 Physiology and genetics of sulfur-oxidizing bacteria Adv Microbial Physiol
39235ndash289Giovannoni SJ 1991 The polymerase chain reaction In Stackebrandt E Goodfellow M editors
Sequencing and hybridization techniques in bacterial systematics New York John Wiley andSons p 177ndash201
Guerrero R Pedros-Alio C Schmidt TM Mas J 1985 A survey of buoyant density of microorganismsin pure cultures and natural samples Microbiologia Sept 1(1ndash2)53ndash65
Harold FM 1966 Inorganic polyphosphates in biology structure metabolism and function BacteriolRev 30(4)772ndash794
Organization of Inclusions in Magnetococci 405
Iida A Akai J 1996 Crystalline sulfur inclusions in magnetotactic bacteria Sci Rep Nigata Univ SerE 1135ndash42
Kornberg A Fraley CD 2000 Inorganic polyphosphate a molecular fossil come to life ASM News66(5)275ndash280
Lins U Farina M 1999 Phosphorus-rich granules in uncultured magnetotactic bacteria FEMSMicrobiol Lett 172(2)23ndash28
Mann S Sparks NHC Frankel RB Bazylinski DA Jannasch HW 1990 Biomineralization of ferri-magnetic greigite (Fe3O4 and iron pyrite (FeS2 ) in a magnetotactic bacterium Nature 343258ndash
261Matsunaga T Sakaguchi T Tadokoro F 1991 Magnetite formation by a magnetic bacterium capable
of growing aerobically Appl Microbiol Biotechnol 35651ndash655Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993a Electron microscopy
study of magnetosomes in a cultured coccoid magnetotactic bacterium Proc Roy Soc Lond B251231ndash236
Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993b Electron microscopystudy of magnetosomes in two cultured vibrioid magnetotactic bacteria Proc Roy Soc Lond B251237ndash242
Moench TT 1988 Bilophococcus magnetotacticus gen nov sp nov a motile magnetic coccus Antvan Leeuwenhoek 54483ndash496
Moench TT Konetzka WA 1978 A novel method for the isolation and study of a magnetotacticbacterium Arch Microbiol 119203ndash212
Murray RGE Robinow CF 1994 Optical Microscopy In Gerhardt P editor Methods for generaland molecular bacteriology Washington DC American Society for Microbiology p 35ndash36
Prange A Arzberger I Engemann C Modrow H Schumann O Truper HG Steudel R Dahl C andHormes J 1999 In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteriaby X-ray absorption near edge spectroscopy Biochim Biophys Acta 1428446ndash454
Rappe MS Kemp PF Giovannoni SJ 1997 Phylogenetic diversity of marine coastal picoplankton16S rRNA genes cloned from the continental shelf off Cape Hatteras North Carolina LimnolOceanogr 42811ndash826
Reusch RN 2000 Transmembrane ion transport by polyphosphatepoly-(R)-3-hydroxybutyratecomplexes Biochemistry (Moscow) 65(3)335ndash352
Sakaguchi T Burgess JG Matsunaga T 1993 Magnetite formation by a sulphate-reducing bacteriumNature 36547ndash49
Schleifer KHSchuler D Spring S Weizenegger M Amann R 1991 The genus Magnetospirillum gennov description of Magnetospirillum gryphiswaldense sp nov and transfer of Aquaspirillummagnetotacticum to Magnetospirillum magnetotacticum comb nov System Appl Microbiol14379ndash385
Schuler D 1999 Formation of magnetosomes in magnetotactic bacteria J Mol Microbiol Biotechnol1(1)79ndash86
Schuler D Spring S Bazylinski DA 1999 Improved technique for the isolation of magnetotactic spir-illa from a freshwater sediment and their phylogenetic characterization System Appl Microbiol22466ndash471
Shively JM 1974 Inclusion bodies of prokaryotes Ann Rev Microbiol 28167ndash187Spring S Amann R Ludwig W Schleifer KH van Gemerden H Petersen N 1993 Dominating role
of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment ApplEnviron Microbiol 59(8)2397ndash2403
Spring S Amann R Ludwig W Schleifer KH Schuler D Poralla K Petersen N 1994 Phylogeneticanalysis of uncultured magnetotactic baceria from the Alpha-subclass of Proteobacteria SystemAppl Microbiol 17501ndash508
Stackebrandt E Goebel BM 1994 Taxonomic note a place for DNA-DNA reassociation and 16SrRNA sequence analysis in the present species de nition in bacteriology Int J System Bacteriol44846ndash849
Stanier RY Ingraham JL Wheelis ML Painter PR 1986 The microbial world 5th edition EnglewoodCliffs NJ Prentice Hall p 179
406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35
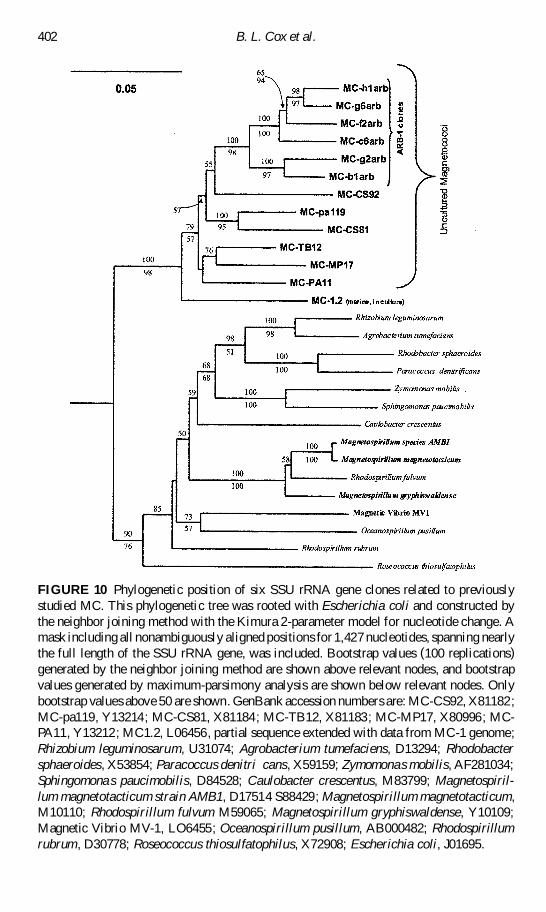
402 B L Cox et al
FIGURE 10 Phylogenetic position of six SSU rRNA gene clones related to previouslystudied MC This phylogenetic tree was rooted with Escherichia coli and constructed bythe neighbor joining method with the Kimura 2-parameter model for nucleotide change Amask including all nonambiguously aligned positions for 1427 nucleotides spanning nearlythe full length of the SSU rRNA gene was included Bootstrap values (100 replications)generated by the neighbor joining method are shown above relevant nodes and bootstrapvalues generated by maximum-parsimony analysis are shown below relevant nodes Onlybootstrap values above 50 are shown GenBank accession numbers are MC-CS92 X81182MC-pa119 Y13214 MC-CS81 X81184 MC-TB12 X81183 MC-MP17 X80996 MC-PA11 Y13212 MC12 L06456 partial sequence extended with data from MC-1 genomeRhizobium leguminosarum U31074 Agrobacterium tumefaciens D13294 Rhodobactersphaeroides X53854 Paracoccus denitri cans X59159 Zymomonas mobilis AF281034Sphingomonas paucimobilis D84528 Caulobacter crescentus M83799 Magnetospiril-lum magnetotacticum strain AMB1 D17514 S88429 Magnetospirillum magnetotacticumM10110 Rhodospirillum fulvum M59065 Magnetospirillum gryphiswaldense Y10109Magnetic Vibrio MV-1 LO6455 Oceanospirillum pusillum AB000482 Rhodospirillumrubrum D30778 Roseococcus thiosulfatophilus X72908 Escherichia coli J01695
Organization of Inclusions in Magnetococci 403
and was possibly complexed with water and organic residues If the S globules in ARB-1cells are analogous to those in the purple sulfur bacteria Chromatium vinosum and Thio-capsa roseopersicina they may contain a signi cant protein content Sulfur globules fromthe anaerobic phototrophs C vinosum and T roseopersicina were found to contain upto 52 protein that formed an envelope enclosing the S globules (Brune 1995) Theseproteins were isolated and analyzed and found to be glycine and tyrosine-rich structuralproteins which were homologous for the two species suggesting they are highly con-served (Brune 1995) Phylogenetically C vinosum and T roseopersicina are members ofthe deg -Proteobacteria whereas MC are members of the reg-Proteobacteria To our knowledgemagnetotactic bacteria may be the only members of the reg-Proteobacteria that make internalsulfur globules
Phylogenetic Af liation
We wished to detemine whether ARB-1 was a single species or a mixed population of MCToward that end 45 SSU rRNA genes were cloned from partially puri ed ARB-1 cells30 of which displayed similar restriction fragment length patterns (RFLP) to previouslycharacterized MC (data not shown) The nding that 15 patterns were not related to pre-viously characterized MC was not surprising given that the initial sample was not a pureculture but rather a population magnetically enriched from a sample originating from arelatively large vertical column containing both sediments and overlying water Fourteenof the MC-related clones representing different RFLP pattern groups were chosen for par-tial sequencing 10 of these sequences showed high similarity to previously characterizedMC Six of these sequences were phylogenetically distinct and were fully bidirectionallysequenced
ARB-1 MC SSU rRNA genes formed a monophyletic cluster related to but dis-tinct from other MC (Figure 10) Within the cluster the similarity between SSU rRNAgene sequences ranged from 97 (MC-H1arb and MC-g6arb) to 92 (MC-h1arb andMC-g2arb) The nearest neighbor to the ARB-1 cluster MC-CS92 was ca 89 similarand the nearest cultured relative MC-1 was ca 88 similar to the ARB-1 cluster As arule of thumb for SSU rRNA genes gt97 similarity is considered the same species andgt93ndash95 similarity is considered the same genus (Stackebrandt and Goebel 1994) Thesedata strongly suggest that ARB-1 represents a population of different diverse MC Therewas signi cant variation within the ARB-1 cluster and therefore we do not consider ARB-1to consist of bacteria of a single species or genus Correlations if any between phylogeneticand phenotypic variations in the ARB-1 cells have yet to be determined
Summary
We have partially characterized a population of magnetotactic cocci (MC) referred to asARB-1 which was particularly abundant in iron-rich sediments of a small eutrophic lake andmorphologically and structurally similar to an uncultured species of MC called Bilophococ-cus magnetotacticus The latter organism was found in large numbers in a sewage treatmentpond Cells of ARB-1 had a relatively wide size range of 1 to 4 sup1m They were motileGram-negative and bilophotrichously agellated Cells of this uncultured magnetotacticcoccus contained three types of intracellular inclusions that appeared to be in a consistentarrangement within the cell The magnetosomes contained crystals of magnetite and wereeither in chains andor polar clumps at the end of the cell where the agellar bundles werelocated Large P-rich bodies occupied the majority of the cell volume and appeared to sepa-rate the S-containing inclusions from the magnetosomes The presence of these large P-rich
404 B L Cox et al
inclusions might be an indication of cell stress or may be important in cell buoyancy or inenergy generation The presence of S-rich inclusions located at the end of the cell oppositethe magnetosomes strongly suggests that ARB-1 cells are sul de-oxidizing bacteria andthat the S might be useful as a stored electron donor when cells travel from sul de-rich tosul de-depleted zones in the water column The wide range of the cell sizes the differentmagnetosome arrangements and the large number of different retrieved 16S rRNA genesequences suggest that ARB-1 represents a population of at least several different MC 16SrRNA sequences obtained from cells of ARB-1 appeared to form a monophyletic clusterrelated to but distinct from other uncultured and cultured MC Future work using uores-cent oligonucleotide probes microscopy and physiological measurements should clarifyrelationships between phylogeny cell morphology and organization of inclusions
References
Balkwill DL Maratea D Blakemore RP 1980 Ultrastructure of a magnetotactic spirillum J Bacteriol141(3)1399ndash1408
Bazylinski DA Frankel RB 2000 Biologically controlled mineralization of magnetic iron mineralsby magnetotactic bacteria In Lovley DR editor Environmental microbe-metal interactionsWashington DC ASM Press p 109ndash143
Bazylinski DA Frankel RB Jannasch HW 1988 Anaerobic magnetite production by a marinemagnetotactic bacterium Nature 334518ndash519
Beveridge T 1999 Structures of gram-negative cell walls and their derived membrane vesiclesJ Bacteriol 181(6)4725ndash4733
Beveridge TJ Popkin TJ Cole RM 1994 Electron microscopy In Gerhardt P editor Methods forgeneral and molecular bacteriology Washington DC American Society for Microbiology p 25
Blakemore R 1975 Magnetotactic bacteria Science 190377ndash379Blakemore RP 1982 Magnetotactic bacteria Ann Rev Microbiol 36217ndash238Blakemore RP Maratea D Wolfe RS 1979 Isolation and pure culture of a freshwater magnetic
spirillum in chemically de ned medium J Bacteriol 140(2)720ndash729Blakemore RP Short KA Bazylinski DA Rosenblatt C Frankel RB 1985 Microaerobic conditions
are required for magnetite formation within Aquaspirillum magnetotacticum Geomicrobiol J4(1)53ndash71
Brune DC 1995 Isolation and characterization of sulfur globule proteins from Chromatium vinosumand Thiocapsa roseopersicina Arch Microbiol 163(1)391ndash399
Butler RF Banerjee SK 1975 Theoretical single-domain grain size range in magnetite and titano-magnetite J Geophys Res 804049ndash4058
Buwalda JP 1940 Geology of the Raymond Basin report to Pasadena Water DepartmentCrook R Allen CR Kamb B Payne CM Procter RJ 1987 Recent reverse faulting in the Transverse
Ranges California Pasadena CA US Geol Surv Prof Paper 133927ndash63Devouard B Posfai M Hua X Bazylinski DA Frankel RB Buseck PR 1998 Magnetite from
magnetotactic bacteria size distribution and twinning Am Mineral 831387ndash1398Frankel RB Bazylinski DA Johnson MS Taylor BL 1997 Magneto-Aerotaxis in marine coccoid
bacteria Biophys J 73994ndash1000Friedrich CG 1998 Physiology and genetics of sulfur-oxidizing bacteria Adv Microbial Physiol
39235ndash289Giovannoni SJ 1991 The polymerase chain reaction In Stackebrandt E Goodfellow M editors
Sequencing and hybridization techniques in bacterial systematics New York John Wiley andSons p 177ndash201
Guerrero R Pedros-Alio C Schmidt TM Mas J 1985 A survey of buoyant density of microorganismsin pure cultures and natural samples Microbiologia Sept 1(1ndash2)53ndash65
Harold FM 1966 Inorganic polyphosphates in biology structure metabolism and function BacteriolRev 30(4)772ndash794
Organization of Inclusions in Magnetococci 405
Iida A Akai J 1996 Crystalline sulfur inclusions in magnetotactic bacteria Sci Rep Nigata Univ SerE 1135ndash42
Kornberg A Fraley CD 2000 Inorganic polyphosphate a molecular fossil come to life ASM News66(5)275ndash280
Lins U Farina M 1999 Phosphorus-rich granules in uncultured magnetotactic bacteria FEMSMicrobiol Lett 172(2)23ndash28
Mann S Sparks NHC Frankel RB Bazylinski DA Jannasch HW 1990 Biomineralization of ferri-magnetic greigite (Fe3O4 and iron pyrite (FeS2 ) in a magnetotactic bacterium Nature 343258ndash
261Matsunaga T Sakaguchi T Tadokoro F 1991 Magnetite formation by a magnetic bacterium capable
of growing aerobically Appl Microbiol Biotechnol 35651ndash655Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993a Electron microscopy
study of magnetosomes in a cultured coccoid magnetotactic bacterium Proc Roy Soc Lond B251231ndash236
Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993b Electron microscopystudy of magnetosomes in two cultured vibrioid magnetotactic bacteria Proc Roy Soc Lond B251237ndash242
Moench TT 1988 Bilophococcus magnetotacticus gen nov sp nov a motile magnetic coccus Antvan Leeuwenhoek 54483ndash496
Moench TT Konetzka WA 1978 A novel method for the isolation and study of a magnetotacticbacterium Arch Microbiol 119203ndash212
Murray RGE Robinow CF 1994 Optical Microscopy In Gerhardt P editor Methods for generaland molecular bacteriology Washington DC American Society for Microbiology p 35ndash36
Prange A Arzberger I Engemann C Modrow H Schumann O Truper HG Steudel R Dahl C andHormes J 1999 In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteriaby X-ray absorption near edge spectroscopy Biochim Biophys Acta 1428446ndash454
Rappe MS Kemp PF Giovannoni SJ 1997 Phylogenetic diversity of marine coastal picoplankton16S rRNA genes cloned from the continental shelf off Cape Hatteras North Carolina LimnolOceanogr 42811ndash826
Reusch RN 2000 Transmembrane ion transport by polyphosphatepoly-(R)-3-hydroxybutyratecomplexes Biochemistry (Moscow) 65(3)335ndash352
Sakaguchi T Burgess JG Matsunaga T 1993 Magnetite formation by a sulphate-reducing bacteriumNature 36547ndash49
Schleifer KHSchuler D Spring S Weizenegger M Amann R 1991 The genus Magnetospirillum gennov description of Magnetospirillum gryphiswaldense sp nov and transfer of Aquaspirillummagnetotacticum to Magnetospirillum magnetotacticum comb nov System Appl Microbiol14379ndash385
Schuler D 1999 Formation of magnetosomes in magnetotactic bacteria J Mol Microbiol Biotechnol1(1)79ndash86
Schuler D Spring S Bazylinski DA 1999 Improved technique for the isolation of magnetotactic spir-illa from a freshwater sediment and their phylogenetic characterization System Appl Microbiol22466ndash471
Shively JM 1974 Inclusion bodies of prokaryotes Ann Rev Microbiol 28167ndash187Spring S Amann R Ludwig W Schleifer KH van Gemerden H Petersen N 1993 Dominating role
of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment ApplEnviron Microbiol 59(8)2397ndash2403
Spring S Amann R Ludwig W Schleifer KH Schuler D Poralla K Petersen N 1994 Phylogeneticanalysis of uncultured magnetotactic baceria from the Alpha-subclass of Proteobacteria SystemAppl Microbiol 17501ndash508
Stackebrandt E Goebel BM 1994 Taxonomic note a place for DNA-DNA reassociation and 16SrRNA sequence analysis in the present species de nition in bacteriology Int J System Bacteriol44846ndash849
Stanier RY Ingraham JL Wheelis ML Painter PR 1986 The microbial world 5th edition EnglewoodCliffs NJ Prentice Hall p 179
406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35

Organization of Inclusions in Magnetococci 403
and was possibly complexed with water and organic residues If the S globules in ARB-1cells are analogous to those in the purple sulfur bacteria Chromatium vinosum and Thio-capsa roseopersicina they may contain a signi cant protein content Sulfur globules fromthe anaerobic phototrophs C vinosum and T roseopersicina were found to contain upto 52 protein that formed an envelope enclosing the S globules (Brune 1995) Theseproteins were isolated and analyzed and found to be glycine and tyrosine-rich structuralproteins which were homologous for the two species suggesting they are highly con-served (Brune 1995) Phylogenetically C vinosum and T roseopersicina are members ofthe deg -Proteobacteria whereas MC are members of the reg-Proteobacteria To our knowledgemagnetotactic bacteria may be the only members of the reg-Proteobacteria that make internalsulfur globules
Phylogenetic Af liation
We wished to detemine whether ARB-1 was a single species or a mixed population of MCToward that end 45 SSU rRNA genes were cloned from partially puri ed ARB-1 cells30 of which displayed similar restriction fragment length patterns (RFLP) to previouslycharacterized MC (data not shown) The nding that 15 patterns were not related to pre-viously characterized MC was not surprising given that the initial sample was not a pureculture but rather a population magnetically enriched from a sample originating from arelatively large vertical column containing both sediments and overlying water Fourteenof the MC-related clones representing different RFLP pattern groups were chosen for par-tial sequencing 10 of these sequences showed high similarity to previously characterizedMC Six of these sequences were phylogenetically distinct and were fully bidirectionallysequenced
ARB-1 MC SSU rRNA genes formed a monophyletic cluster related to but dis-tinct from other MC (Figure 10) Within the cluster the similarity between SSU rRNAgene sequences ranged from 97 (MC-H1arb and MC-g6arb) to 92 (MC-h1arb andMC-g2arb) The nearest neighbor to the ARB-1 cluster MC-CS92 was ca 89 similarand the nearest cultured relative MC-1 was ca 88 similar to the ARB-1 cluster As arule of thumb for SSU rRNA genes gt97 similarity is considered the same species andgt93ndash95 similarity is considered the same genus (Stackebrandt and Goebel 1994) Thesedata strongly suggest that ARB-1 represents a population of different diverse MC Therewas signi cant variation within the ARB-1 cluster and therefore we do not consider ARB-1to consist of bacteria of a single species or genus Correlations if any between phylogeneticand phenotypic variations in the ARB-1 cells have yet to be determined
Summary
We have partially characterized a population of magnetotactic cocci (MC) referred to asARB-1 which was particularly abundant in iron-rich sediments of a small eutrophic lake andmorphologically and structurally similar to an uncultured species of MC called Bilophococ-cus magnetotacticus The latter organism was found in large numbers in a sewage treatmentpond Cells of ARB-1 had a relatively wide size range of 1 to 4 sup1m They were motileGram-negative and bilophotrichously agellated Cells of this uncultured magnetotacticcoccus contained three types of intracellular inclusions that appeared to be in a consistentarrangement within the cell The magnetosomes contained crystals of magnetite and wereeither in chains andor polar clumps at the end of the cell where the agellar bundles werelocated Large P-rich bodies occupied the majority of the cell volume and appeared to sepa-rate the S-containing inclusions from the magnetosomes The presence of these large P-rich
404 B L Cox et al
inclusions might be an indication of cell stress or may be important in cell buoyancy or inenergy generation The presence of S-rich inclusions located at the end of the cell oppositethe magnetosomes strongly suggests that ARB-1 cells are sul de-oxidizing bacteria andthat the S might be useful as a stored electron donor when cells travel from sul de-rich tosul de-depleted zones in the water column The wide range of the cell sizes the differentmagnetosome arrangements and the large number of different retrieved 16S rRNA genesequences suggest that ARB-1 represents a population of at least several different MC 16SrRNA sequences obtained from cells of ARB-1 appeared to form a monophyletic clusterrelated to but distinct from other uncultured and cultured MC Future work using uores-cent oligonucleotide probes microscopy and physiological measurements should clarifyrelationships between phylogeny cell morphology and organization of inclusions
References
Balkwill DL Maratea D Blakemore RP 1980 Ultrastructure of a magnetotactic spirillum J Bacteriol141(3)1399ndash1408
Bazylinski DA Frankel RB 2000 Biologically controlled mineralization of magnetic iron mineralsby magnetotactic bacteria In Lovley DR editor Environmental microbe-metal interactionsWashington DC ASM Press p 109ndash143
Bazylinski DA Frankel RB Jannasch HW 1988 Anaerobic magnetite production by a marinemagnetotactic bacterium Nature 334518ndash519
Beveridge T 1999 Structures of gram-negative cell walls and their derived membrane vesiclesJ Bacteriol 181(6)4725ndash4733
Beveridge TJ Popkin TJ Cole RM 1994 Electron microscopy In Gerhardt P editor Methods forgeneral and molecular bacteriology Washington DC American Society for Microbiology p 25
Blakemore R 1975 Magnetotactic bacteria Science 190377ndash379Blakemore RP 1982 Magnetotactic bacteria Ann Rev Microbiol 36217ndash238Blakemore RP Maratea D Wolfe RS 1979 Isolation and pure culture of a freshwater magnetic
spirillum in chemically de ned medium J Bacteriol 140(2)720ndash729Blakemore RP Short KA Bazylinski DA Rosenblatt C Frankel RB 1985 Microaerobic conditions
are required for magnetite formation within Aquaspirillum magnetotacticum Geomicrobiol J4(1)53ndash71
Brune DC 1995 Isolation and characterization of sulfur globule proteins from Chromatium vinosumand Thiocapsa roseopersicina Arch Microbiol 163(1)391ndash399
Butler RF Banerjee SK 1975 Theoretical single-domain grain size range in magnetite and titano-magnetite J Geophys Res 804049ndash4058
Buwalda JP 1940 Geology of the Raymond Basin report to Pasadena Water DepartmentCrook R Allen CR Kamb B Payne CM Procter RJ 1987 Recent reverse faulting in the Transverse
Ranges California Pasadena CA US Geol Surv Prof Paper 133927ndash63Devouard B Posfai M Hua X Bazylinski DA Frankel RB Buseck PR 1998 Magnetite from
magnetotactic bacteria size distribution and twinning Am Mineral 831387ndash1398Frankel RB Bazylinski DA Johnson MS Taylor BL 1997 Magneto-Aerotaxis in marine coccoid
bacteria Biophys J 73994ndash1000Friedrich CG 1998 Physiology and genetics of sulfur-oxidizing bacteria Adv Microbial Physiol
39235ndash289Giovannoni SJ 1991 The polymerase chain reaction In Stackebrandt E Goodfellow M editors
Sequencing and hybridization techniques in bacterial systematics New York John Wiley andSons p 177ndash201
Guerrero R Pedros-Alio C Schmidt TM Mas J 1985 A survey of buoyant density of microorganismsin pure cultures and natural samples Microbiologia Sept 1(1ndash2)53ndash65
Harold FM 1966 Inorganic polyphosphates in biology structure metabolism and function BacteriolRev 30(4)772ndash794
Organization of Inclusions in Magnetococci 405
Iida A Akai J 1996 Crystalline sulfur inclusions in magnetotactic bacteria Sci Rep Nigata Univ SerE 1135ndash42
Kornberg A Fraley CD 2000 Inorganic polyphosphate a molecular fossil come to life ASM News66(5)275ndash280
Lins U Farina M 1999 Phosphorus-rich granules in uncultured magnetotactic bacteria FEMSMicrobiol Lett 172(2)23ndash28
Mann S Sparks NHC Frankel RB Bazylinski DA Jannasch HW 1990 Biomineralization of ferri-magnetic greigite (Fe3O4 and iron pyrite (FeS2 ) in a magnetotactic bacterium Nature 343258ndash
261Matsunaga T Sakaguchi T Tadokoro F 1991 Magnetite formation by a magnetic bacterium capable
of growing aerobically Appl Microbiol Biotechnol 35651ndash655Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993a Electron microscopy
study of magnetosomes in a cultured coccoid magnetotactic bacterium Proc Roy Soc Lond B251231ndash236
Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993b Electron microscopystudy of magnetosomes in two cultured vibrioid magnetotactic bacteria Proc Roy Soc Lond B251237ndash242
Moench TT 1988 Bilophococcus magnetotacticus gen nov sp nov a motile magnetic coccus Antvan Leeuwenhoek 54483ndash496
Moench TT Konetzka WA 1978 A novel method for the isolation and study of a magnetotacticbacterium Arch Microbiol 119203ndash212
Murray RGE Robinow CF 1994 Optical Microscopy In Gerhardt P editor Methods for generaland molecular bacteriology Washington DC American Society for Microbiology p 35ndash36
Prange A Arzberger I Engemann C Modrow H Schumann O Truper HG Steudel R Dahl C andHormes J 1999 In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteriaby X-ray absorption near edge spectroscopy Biochim Biophys Acta 1428446ndash454
Rappe MS Kemp PF Giovannoni SJ 1997 Phylogenetic diversity of marine coastal picoplankton16S rRNA genes cloned from the continental shelf off Cape Hatteras North Carolina LimnolOceanogr 42811ndash826
Reusch RN 2000 Transmembrane ion transport by polyphosphatepoly-(R)-3-hydroxybutyratecomplexes Biochemistry (Moscow) 65(3)335ndash352
Sakaguchi T Burgess JG Matsunaga T 1993 Magnetite formation by a sulphate-reducing bacteriumNature 36547ndash49
Schleifer KHSchuler D Spring S Weizenegger M Amann R 1991 The genus Magnetospirillum gennov description of Magnetospirillum gryphiswaldense sp nov and transfer of Aquaspirillummagnetotacticum to Magnetospirillum magnetotacticum comb nov System Appl Microbiol14379ndash385
Schuler D 1999 Formation of magnetosomes in magnetotactic bacteria J Mol Microbiol Biotechnol1(1)79ndash86
Schuler D Spring S Bazylinski DA 1999 Improved technique for the isolation of magnetotactic spir-illa from a freshwater sediment and their phylogenetic characterization System Appl Microbiol22466ndash471
Shively JM 1974 Inclusion bodies of prokaryotes Ann Rev Microbiol 28167ndash187Spring S Amann R Ludwig W Schleifer KH van Gemerden H Petersen N 1993 Dominating role
of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment ApplEnviron Microbiol 59(8)2397ndash2403
Spring S Amann R Ludwig W Schleifer KH Schuler D Poralla K Petersen N 1994 Phylogeneticanalysis of uncultured magnetotactic baceria from the Alpha-subclass of Proteobacteria SystemAppl Microbiol 17501ndash508
Stackebrandt E Goebel BM 1994 Taxonomic note a place for DNA-DNA reassociation and 16SrRNA sequence analysis in the present species de nition in bacteriology Int J System Bacteriol44846ndash849
Stanier RY Ingraham JL Wheelis ML Painter PR 1986 The microbial world 5th edition EnglewoodCliffs NJ Prentice Hall p 179
406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35

404 B L Cox et al
inclusions might be an indication of cell stress or may be important in cell buoyancy or inenergy generation The presence of S-rich inclusions located at the end of the cell oppositethe magnetosomes strongly suggests that ARB-1 cells are sul de-oxidizing bacteria andthat the S might be useful as a stored electron donor when cells travel from sul de-rich tosul de-depleted zones in the water column The wide range of the cell sizes the differentmagnetosome arrangements and the large number of different retrieved 16S rRNA genesequences suggest that ARB-1 represents a population of at least several different MC 16SrRNA sequences obtained from cells of ARB-1 appeared to form a monophyletic clusterrelated to but distinct from other uncultured and cultured MC Future work using uores-cent oligonucleotide probes microscopy and physiological measurements should clarifyrelationships between phylogeny cell morphology and organization of inclusions
References
Balkwill DL Maratea D Blakemore RP 1980 Ultrastructure of a magnetotactic spirillum J Bacteriol141(3)1399ndash1408
Bazylinski DA Frankel RB 2000 Biologically controlled mineralization of magnetic iron mineralsby magnetotactic bacteria In Lovley DR editor Environmental microbe-metal interactionsWashington DC ASM Press p 109ndash143
Bazylinski DA Frankel RB Jannasch HW 1988 Anaerobic magnetite production by a marinemagnetotactic bacterium Nature 334518ndash519
Beveridge T 1999 Structures of gram-negative cell walls and their derived membrane vesiclesJ Bacteriol 181(6)4725ndash4733
Beveridge TJ Popkin TJ Cole RM 1994 Electron microscopy In Gerhardt P editor Methods forgeneral and molecular bacteriology Washington DC American Society for Microbiology p 25
Blakemore R 1975 Magnetotactic bacteria Science 190377ndash379Blakemore RP 1982 Magnetotactic bacteria Ann Rev Microbiol 36217ndash238Blakemore RP Maratea D Wolfe RS 1979 Isolation and pure culture of a freshwater magnetic
spirillum in chemically de ned medium J Bacteriol 140(2)720ndash729Blakemore RP Short KA Bazylinski DA Rosenblatt C Frankel RB 1985 Microaerobic conditions
are required for magnetite formation within Aquaspirillum magnetotacticum Geomicrobiol J4(1)53ndash71
Brune DC 1995 Isolation and characterization of sulfur globule proteins from Chromatium vinosumand Thiocapsa roseopersicina Arch Microbiol 163(1)391ndash399
Butler RF Banerjee SK 1975 Theoretical single-domain grain size range in magnetite and titano-magnetite J Geophys Res 804049ndash4058
Buwalda JP 1940 Geology of the Raymond Basin report to Pasadena Water DepartmentCrook R Allen CR Kamb B Payne CM Procter RJ 1987 Recent reverse faulting in the Transverse
Ranges California Pasadena CA US Geol Surv Prof Paper 133927ndash63Devouard B Posfai M Hua X Bazylinski DA Frankel RB Buseck PR 1998 Magnetite from
magnetotactic bacteria size distribution and twinning Am Mineral 831387ndash1398Frankel RB Bazylinski DA Johnson MS Taylor BL 1997 Magneto-Aerotaxis in marine coccoid
bacteria Biophys J 73994ndash1000Friedrich CG 1998 Physiology and genetics of sulfur-oxidizing bacteria Adv Microbial Physiol
39235ndash289Giovannoni SJ 1991 The polymerase chain reaction In Stackebrandt E Goodfellow M editors
Sequencing and hybridization techniques in bacterial systematics New York John Wiley andSons p 177ndash201
Guerrero R Pedros-Alio C Schmidt TM Mas J 1985 A survey of buoyant density of microorganismsin pure cultures and natural samples Microbiologia Sept 1(1ndash2)53ndash65
Harold FM 1966 Inorganic polyphosphates in biology structure metabolism and function BacteriolRev 30(4)772ndash794
Organization of Inclusions in Magnetococci 405
Iida A Akai J 1996 Crystalline sulfur inclusions in magnetotactic bacteria Sci Rep Nigata Univ SerE 1135ndash42
Kornberg A Fraley CD 2000 Inorganic polyphosphate a molecular fossil come to life ASM News66(5)275ndash280
Lins U Farina M 1999 Phosphorus-rich granules in uncultured magnetotactic bacteria FEMSMicrobiol Lett 172(2)23ndash28
Mann S Sparks NHC Frankel RB Bazylinski DA Jannasch HW 1990 Biomineralization of ferri-magnetic greigite (Fe3O4 and iron pyrite (FeS2 ) in a magnetotactic bacterium Nature 343258ndash
261Matsunaga T Sakaguchi T Tadokoro F 1991 Magnetite formation by a magnetic bacterium capable
of growing aerobically Appl Microbiol Biotechnol 35651ndash655Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993a Electron microscopy
study of magnetosomes in a cultured coccoid magnetotactic bacterium Proc Roy Soc Lond B251231ndash236
Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993b Electron microscopystudy of magnetosomes in two cultured vibrioid magnetotactic bacteria Proc Roy Soc Lond B251237ndash242
Moench TT 1988 Bilophococcus magnetotacticus gen nov sp nov a motile magnetic coccus Antvan Leeuwenhoek 54483ndash496
Moench TT Konetzka WA 1978 A novel method for the isolation and study of a magnetotacticbacterium Arch Microbiol 119203ndash212
Murray RGE Robinow CF 1994 Optical Microscopy In Gerhardt P editor Methods for generaland molecular bacteriology Washington DC American Society for Microbiology p 35ndash36
Prange A Arzberger I Engemann C Modrow H Schumann O Truper HG Steudel R Dahl C andHormes J 1999 In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteriaby X-ray absorption near edge spectroscopy Biochim Biophys Acta 1428446ndash454
Rappe MS Kemp PF Giovannoni SJ 1997 Phylogenetic diversity of marine coastal picoplankton16S rRNA genes cloned from the continental shelf off Cape Hatteras North Carolina LimnolOceanogr 42811ndash826
Reusch RN 2000 Transmembrane ion transport by polyphosphatepoly-(R)-3-hydroxybutyratecomplexes Biochemistry (Moscow) 65(3)335ndash352
Sakaguchi T Burgess JG Matsunaga T 1993 Magnetite formation by a sulphate-reducing bacteriumNature 36547ndash49
Schleifer KHSchuler D Spring S Weizenegger M Amann R 1991 The genus Magnetospirillum gennov description of Magnetospirillum gryphiswaldense sp nov and transfer of Aquaspirillummagnetotacticum to Magnetospirillum magnetotacticum comb nov System Appl Microbiol14379ndash385
Schuler D 1999 Formation of magnetosomes in magnetotactic bacteria J Mol Microbiol Biotechnol1(1)79ndash86
Schuler D Spring S Bazylinski DA 1999 Improved technique for the isolation of magnetotactic spir-illa from a freshwater sediment and their phylogenetic characterization System Appl Microbiol22466ndash471
Shively JM 1974 Inclusion bodies of prokaryotes Ann Rev Microbiol 28167ndash187Spring S Amann R Ludwig W Schleifer KH van Gemerden H Petersen N 1993 Dominating role
of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment ApplEnviron Microbiol 59(8)2397ndash2403
Spring S Amann R Ludwig W Schleifer KH Schuler D Poralla K Petersen N 1994 Phylogeneticanalysis of uncultured magnetotactic baceria from the Alpha-subclass of Proteobacteria SystemAppl Microbiol 17501ndash508
Stackebrandt E Goebel BM 1994 Taxonomic note a place for DNA-DNA reassociation and 16SrRNA sequence analysis in the present species de nition in bacteriology Int J System Bacteriol44846ndash849
Stanier RY Ingraham JL Wheelis ML Painter PR 1986 The microbial world 5th edition EnglewoodCliffs NJ Prentice Hall p 179
406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35

Organization of Inclusions in Magnetococci 405
Iida A Akai J 1996 Crystalline sulfur inclusions in magnetotactic bacteria Sci Rep Nigata Univ SerE 1135ndash42
Kornberg A Fraley CD 2000 Inorganic polyphosphate a molecular fossil come to life ASM News66(5)275ndash280
Lins U Farina M 1999 Phosphorus-rich granules in uncultured magnetotactic bacteria FEMSMicrobiol Lett 172(2)23ndash28
Mann S Sparks NHC Frankel RB Bazylinski DA Jannasch HW 1990 Biomineralization of ferri-magnetic greigite (Fe3O4 and iron pyrite (FeS2 ) in a magnetotactic bacterium Nature 343258ndash
261Matsunaga T Sakaguchi T Tadokoro F 1991 Magnetite formation by a magnetic bacterium capable
of growing aerobically Appl Microbiol Biotechnol 35651ndash655Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993a Electron microscopy
study of magnetosomes in a cultured coccoid magnetotactic bacterium Proc Roy Soc Lond B251231ndash236
Meldrum FC Mann S Heywood BR Frankel RB Bazylinski DA 1993b Electron microscopystudy of magnetosomes in two cultured vibrioid magnetotactic bacteria Proc Roy Soc Lond B251237ndash242
Moench TT 1988 Bilophococcus magnetotacticus gen nov sp nov a motile magnetic coccus Antvan Leeuwenhoek 54483ndash496
Moench TT Konetzka WA 1978 A novel method for the isolation and study of a magnetotacticbacterium Arch Microbiol 119203ndash212
Murray RGE Robinow CF 1994 Optical Microscopy In Gerhardt P editor Methods for generaland molecular bacteriology Washington DC American Society for Microbiology p 35ndash36
Prange A Arzberger I Engemann C Modrow H Schumann O Truper HG Steudel R Dahl C andHormes J 1999 In situ analysis of sulfur in the sulfur globules of phototrophic sulfur bacteriaby X-ray absorption near edge spectroscopy Biochim Biophys Acta 1428446ndash454
Rappe MS Kemp PF Giovannoni SJ 1997 Phylogenetic diversity of marine coastal picoplankton16S rRNA genes cloned from the continental shelf off Cape Hatteras North Carolina LimnolOceanogr 42811ndash826
Reusch RN 2000 Transmembrane ion transport by polyphosphatepoly-(R)-3-hydroxybutyratecomplexes Biochemistry (Moscow) 65(3)335ndash352
Sakaguchi T Burgess JG Matsunaga T 1993 Magnetite formation by a sulphate-reducing bacteriumNature 36547ndash49
Schleifer KHSchuler D Spring S Weizenegger M Amann R 1991 The genus Magnetospirillum gennov description of Magnetospirillum gryphiswaldense sp nov and transfer of Aquaspirillummagnetotacticum to Magnetospirillum magnetotacticum comb nov System Appl Microbiol14379ndash385
Schuler D 1999 Formation of magnetosomes in magnetotactic bacteria J Mol Microbiol Biotechnol1(1)79ndash86
Schuler D Spring S Bazylinski DA 1999 Improved technique for the isolation of magnetotactic spir-illa from a freshwater sediment and their phylogenetic characterization System Appl Microbiol22466ndash471
Shively JM 1974 Inclusion bodies of prokaryotes Ann Rev Microbiol 28167ndash187Spring S Amann R Ludwig W Schleifer KH van Gemerden H Petersen N 1993 Dominating role
of an unusual magnetotactic bacterium in the microaerobic zone of a freshwater sediment ApplEnviron Microbiol 59(8)2397ndash2403
Spring S Amann R Ludwig W Schleifer KH Schuler D Poralla K Petersen N 1994 Phylogeneticanalysis of uncultured magnetotactic baceria from the Alpha-subclass of Proteobacteria SystemAppl Microbiol 17501ndash508
Stackebrandt E Goebel BM 1994 Taxonomic note a place for DNA-DNA reassociation and 16SrRNA sequence analysis in the present species de nition in bacteriology Int J System Bacteriol44846ndash849
Stanier RY Ingraham JL Wheelis ML Painter PR 1986 The microbial world 5th edition EnglewoodCliffs NJ Prentice Hall p 179
406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35

406 B L Cox et al
Taylor AP Barry JC Webb RI 2001 Structural and morphological anomalies in magnetosomespossible biogenic origin for magnetite in ALH84001 J Microsc Oxford 201(1 Jan 2001)84ndash
106Thomas-Keprta KL Bazylinski DA Kirschvink JL Clemett SJ McKay DS Wentworth SJ Vali H
Gibson EKJ Romanek CS 2000 Elongated prismatic magnetic crystals in ALH84001 carbonateglobules potential Martian magnetofossils Geochim Cosmochim Acta 64(23)4049ndash4081
Towe KM Moench TT 1981 Electron optical characterization of bacterial magnetite Earth PlanetSci Lett 52213ndash220
Vali H Kirschvink JL 1990 Observations of magnetosome organization surface structure and ironbiomineralization of undescribed magnetic bacteria evolutionary speculations In Frankel RBBlakemore RP editors Iron biominerals New York Plenum Press p 97ndash113
Van Veen HW Abee T Kortstee GJJ Pereira H Konings WN Zehnder AJB 1994 Generation ofa proton motive force by the excretion of metal-phosphate in the polyphosphate-accumulatingAcinetobacter-Johnsonii strain 210A J Biol Chem 269(47)29509ndash29514
Wolfe RS Thauer RK Pfennig N 1987 A ldquocapillary racetrackrdquo method for isolation of magnetotacticbacteria FEMS Microbiol Ecol 4531ndash35




















