
Organic marker compounds for surface soil and fugitive dust from open lot dairies and cattle...
23
Atmospheric Environment 40 (2006) 27–49 Organic marker compounds for surface soil and fugitive dust from open lot dairies and cattle feedlots Wolfgang F. Rogge a, , Patricia M. Medeiros b , Bernd R.T. Simoneit b a Department of Civil and Environmental Engineering, Florida International University, Miami, FL 33199, USA b Environmental and Petroleum Geochemistry Group, College of Oceanic and Atmospheric Sciences, Oregon State University, Corvallis, OR 97331, USA Received 20 April 2005; received in revised form 10 July 2005; accepted 10 July 2005 Abstract Fugitive dust emissions from cattle feedlots and open lot dairies are substantial. In order to determine the contribution of intensive cattle operations on ambient PM levels, more knowledge besides the elemental composition is necessary in order to distinguish between airborne PM from nearby agricultural fields, barren lands, or dirt roads. Here, as part of the San Joaquin Valley Fugitive Dust Characterization Study, surface soil samples collected from feedlots and open lot dairy farms are investigated for potential source specific molecular marker compounds. More than 100 organic compounds were quantified including: n-alkanes, n-alkanoic acids, n-alkenoic acids, n-alkanols, n-alkanals, n-alkan-2-ones, steroids, triterpenoids, isoprenoids, and tocopherols (vitamin E) and metabolites. Biohydrogenation of plant lipids and sterols in the rumen results in distinctive alteration products. Animal and plant derived steroids are most abundant. Here, it is shown that 5b-stigmastanol and epi-5b-stigmastanol, two biohydrogenation products of sitosterol and stigmasterol, are the most distinctive molecular marker compounds. While stearic (C 18 ) and palmitic (C 16 ) acids are as individual compounds not source specific, biohydrogenation of the more abundant C 18 unsaturated fatty acids, causes the ratio of C 18 /C 16 fatty acids to shift from below 0.5 for vegetation to an average of 3.070.7. Consequently, the C 18 /C 16 fatty acid ratio is unique and can be used as well in source apportionment studies. r 2005 Elsevier Ltd. All rights reserved. Keywords: Concentrated animal feeding operations; Tocopheryl acetate metabolites; 5b-stigmastanol; epi-5b-Stigmastanol; Fatty acids; epi-Coprosteroids; Coprostanol; epi-Coprostanol; Phytosterols; Phytostanols 1. Introduction In the San Joaquin Valley of California, 40–60% of the ambient PM10 concentrations have been suggested to be the result of agricultural activities (Chow et al., 1992, 1993; Clausnitzer and Singer, 2000). Major contributions of these mainly fugitive sources are related to wind blown dust from agricultural fields post harvesting and agricultural activities such as tilling, harvesting, and traffic over unpaved roads. Likewise, cattle husbandry in open lot dairies and beef cattle feedlots are major sources of fugitive dust, including the inhalable fractions (PM10 and PM2.5). In the United States, animal farming has changed dramatically over the last century. Since World War ARTICLE IN PRESS www.elsevier.com/locate/atmosenv 1352-2310/$ - see front matter r 2005 Elsevier Ltd. All rights reserved. doi:10.1016/j.atmosenv.2005.07.076 Corresponding author. Tel.: +1 305 348 6755; fax: +1 305 348 2802. E-mail address: roggew@fiu.edu (W.F. Rogge).
Transcript of Organic marker compounds for surface soil and fugitive dust from open lot dairies and cattle...

ARTICLE IN PRESS
1352-2310/$ - se
doi:10.1016/j.at
�Correspondfax: +1305 348
E-mail addr
Atmospheric Environment 40 (2006) 27–49
www.elsevier.com/locate/atmosenv
Organic marker compounds for surface soil and fugitive dustfrom open lot dairies and cattle feedlots
Wolfgang F. Roggea,�, Patricia M. Medeirosb, Bernd R.T. Simoneitb
aDepartment of Civil and Environmental Engineering, Florida International University, Miami, FL 33199, USAbEnvironmental and Petroleum Geochemistry Group, College of Oceanic and Atmospheric Sciences, Oregon State University,
Corvallis, OR 97331, USA
Received 20 April 2005; received in revised form 10 July 2005; accepted 10 July 2005
Abstract
Fugitive dust emissions from cattle feedlots and open lot dairies are substantial. In order to determine the contribution
of intensive cattle operations on ambient PM levels, more knowledge besides the elemental composition is necessary in
order to distinguish between airborne PM from nearby agricultural fields, barren lands, or dirt roads. Here, as part of the
San Joaquin Valley Fugitive Dust Characterization Study, surface soil samples collected from feedlots and open lot dairy
farms are investigated for potential source specific molecular marker compounds. More than 100 organic compounds were
quantified including: n-alkanes, n-alkanoic acids, n-alkenoic acids, n-alkanols, n-alkanals, n-alkan-2-ones, steroids,
triterpenoids, isoprenoids, and tocopherols (vitamin E) and metabolites. Biohydrogenation of plant lipids and sterols in the
rumen results in distinctive alteration products. Animal and plant derived steroids are most abundant. Here, it is shown
that 5b-stigmastanol and epi-5b-stigmastanol, two biohydrogenation products of sitosterol and stigmasterol, are the most
distinctive molecular marker compounds. While stearic (C18) and palmitic (C16) acids are as individual compounds not
source specific, biohydrogenation of the more abundant C18 unsaturated fatty acids, causes the ratio of C18/C16 fatty acids
to shift from below 0.5 for vegetation to an average of 3.070.7. Consequently, the C18/C16 fatty acid ratio is unique and
can be used as well in source apportionment studies.
r 2005 Elsevier Ltd. All rights reserved.
Keywords: Concentrated animal feeding operations; Tocopheryl acetate metabolites; 5b-stigmastanol; epi-5b-Stigmastanol; Fatty acids;
epi-Coprosteroids; Coprostanol; epi-Coprostanol; Phytosterols; Phytostanols
1. Introduction
In the San Joaquin Valley of California, 40–60%of the ambient PM10 concentrations have beensuggested to be the result of agricultural activities(Chow et al., 1992, 1993; Clausnitzer and Singer,
e front matter r 2005 Elsevier Ltd. All rights reserved
mosenv.2005.07.076
ing author. Tel.: +1305 348 6755;
2802.
ess: [email protected] (W.F. Rogge).
2000). Major contributions of these mainly fugitivesources are related to wind blown dust fromagricultural fields post harvesting and agriculturalactivities such as tilling, harvesting, and traffic overunpaved roads. Likewise, cattle husbandry in openlot dairies and beef cattle feedlots are major sourcesof fugitive dust, including the inhalable fractions(PM10 and PM2.5).
In the United States, animal farming has changeddramatically over the last century. Since World War
.

ARTICLE IN PRESSW.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–4928
II, cattle raising is shifting away from farms thatwere limited by their own fodder production tolarge industrialized feeding operations that mainlybuy their cattle fodder (Gregory, 2000; Hinrichs andWelsh, 2003; Subak, 1999). In 2003, about 96million cattle and calves were raised in the UnitedStates in pastures, dairy farms, and feedlots(USDA, 2004). Today, more than 20 million cattleare raised annually for fattening in feedlots thatoften hold several tens of thousands of cattle at atime. Typically, cattle stay about 80–150 days forfattening in feedlots. Within the San Joaquin Valley,the cattle population consists of nearly 2 milliondairy cattle, about 400,000 range cattle, and close to300,000 beef cattle raised in feedlots.
Open lot concentrated cattle operations in theUSA generate about 18 million metric tons ofcollectable manure/year (USDA, 2000). The solidand liquid waste generated from cattle farm opera-tions have a pronounced impact on nearby surfaceand ground water quality, as well as soil contam-ination and air pollution. Major contaminants arenitrate, phosphate, veterinary agents, natural ster-oidal estrogen hormones, pesticides, pathogenicmicroorganisms and resulting endotoxins, petro-leum related exhaust from farm vehicles, organicdust from feedlot surface soil and dried manure,dust from fodder receiving, loading and processingoperations, ammonia (NH3) that leads to secondaryaerosol formation with subsequent acidic aerosoldeposition, reactive organic gases (ROG) thateventually lead to ozone and secondary organicaerosol formation, green house gases such as carbondioxide (CO2), methane (CH4), nitrous oxide (N2O),odorous compounds such as H2S, amides, volatileorganic acids, mercaptans and others (e.g., Gintinget al., 2003; Goss et al., 1998; Hanselman et al.,2003; Hartung, 1998; Hintz and Linke, 1998;Jjemba, 2002; Lammel et al., 2004; Larney et al.,2003; McGinn et al., 2002, 2003; NRC, 2003;Phetteplace et al., 2001; Saintfort et al., 1995; Scottet al., 2003; Shaw et al., 1998; Subak, 1999; Takaiet al., 1998; Todd et al., 2004; USDA, 2000).
While the impact of cattle feedlots and dairyfarms on surface and ground water quality hasreceived appreciable attention in the past, little isknown about the impact of fugitive dust on localand regional air quality. In epidemiological andclinical studies, organic dusts from animal husban-dries have been related to allergic rhinitis, occupa-tional farmer asthma, chronic obstructivepulmonary disease, other respiratory diseases, and
cancer (e.g., Bessot et al., 1996; Cathomas et al.,2002; Kauppinen et al., 1992; Molocznik, 2002;Molocznik and Zagorski, 2001; Radon et al., 2001;Rautiainen et al., 1997). Similarly, cattle haveexhibited respiratory distress that is believed to berelated to the organic dust, fungi and bacteriaassociated with manure dust particles, and otheragents (McVean et al., 1986; Purdy et al., 2004;Sorenson et al., 1992; Woolums et al., 2001).Likewise, it has been suggested that intensivelivestock operations impair the quality of life ofnearby residents mainly from airborne dust andodor nuisances (McGinn et al., 2002, 2003; Purdyet al., 2004; Sweeten and Miner, 1993; Wing andWolf, 2000).
As part of the San Joaquin Valley Fugitive DustCharacterization Study, 47 surface soil sampleswere collected by Ashbaugh and coworkers (2003)from agricultural fields growing major crops,feedlots, dairy farms, construction sites, as well asdust from paved and unpaved roads. These soil dustsamples were characterized for moisture content,particle-size distribution, dry silt content, nitrogenand carbon content, and PM10 as well as PM2.5index (Ashbaugh et al., 2003; Carvacho et al., 2001).Chow and coworkers (2003) used a resuspensionapparatus to generate PM10 samples that wereanalyzed for 40 elements, EC, OC, and seven watersoluble ions. According to the above investigations,the surface soils from the feedlots and open lotdairies were classified as mainly organic. Com-monly, soil dusts from agricultural fields are oftennot distinguishable from each other or from desertderived airborne dust particles based solely on theirmajor elemental composition. Thus, adding specificorganic marker compounds to the bulk chemical,ionic, and elemental data of the fugitive dust sourceprofiles allows discrimination of fugitive dust sourcecontributions to a given site of interest (Rogge,2000).
The detailed organic chemical composition ofsurface soils from cattle husbandries has previouslynot been determined, and therefore, it is the scope ofthis study. If unique organic compounds can bedetermined in the soil dust from open lot concen-trated cattle operations (OLCCO), then the nextstep is to investigate if these source specificmolecular markers are measurable in the airbornefugitive dust downwind from cattle operations.Should this also be the case, then the fraction ofambient aerosol derived from OLCCO can bedetermined.

ARTICLE IN PRESSW.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–49 29
2. Experimental methods
2.1. Surface soil samples
The surface soil samples were collected according toEPA guidelines for surface and bulk materials byAshbaugh and coworkers (2003) as part of the SanJoaquin Valley Fugitive Dust Characterization Studyin 1997. The samples were taken next to theconcentrated cattle feeding operations, just outsidethe corral fence. Although portions of these sampleswere size-segregated via resuspension into PM10 andPM2.5 (Ashbaugh et al., 2003; Chow et al., 2003),only the surface soil samples were available to us foranalysis. The soil samples were shipped frozen over-night to our lab at FIU, where they were stored uponarrival in a freezer at less than �21 1C. The sampledescriptions and sampling locations are given inTable 1, for more details see Ashbaugh et al. (2003).
2.2. Extraction
For each sample, small rocks, if present, were firstremoved with a solvent rinsed stainless steel tweezers,and then the remaining soil was ground to powder. Asubsample of 5 g was selected for solvent extraction.A suite of internal standards, consisting of decanoic-d19 acid, tetradecanoic-d27 acid, n-eicosane-d42, n-tetracosane-d50, and n-triacontane-d62, was addedto each sample aliquot just before solvent extraction.Every 5 g sample was extracted three times, each with30mL dichloromethane using mild ultrasonicationfor 10min. The extracts were combined, filtered andconcentrated by rotary evaporator to about 3mL,then further to about 1mL using a stream of highpurity nitrogen. Each extract was split and onealiquot derivatized with diazomethane (preparedfrom 1-methyl-3-nitro-1-nitrosoguanidine in ether)to convert organic acids to their methyl esters. Fromthe same soil samples, independently, a second set
Table 1
Soil sample locations and descriptionsa
Sample ID Source Latitude L
FDCTD1 Dairy 3610701300 N 11
FDCTD2 Dairy 3613005400 N 11
FDCTF1 Feedlot 3513000200 N 11
FDCTF2 Feedlot 3611501000 N 12
OC: Organic carbon.aAshbaugh et al. (2003).bChow et al. (2003).
was extracted to examine exclusively the polarorganic compounds fraction. These were ultrasoni-cated three times for 10min with dichloromethane:-methanol (1:1; v/v). Aliquots of the total extractswere converted to trimethylsilyl derivatives byreaction with N,O-bis-(trimethylsilyl)trifluoroaceta-mide (BSTFA) containing 1% trimethylchlorosilaneand pyridine for 3 h at 70 1C.
2.3. Instrumental analysis
All sample extracts were analyzed by gas chro-matography–mass spectrometry (GC–MS). Themethylated extracts of the soils were analyzed usingan Agilent 6890 gas chromatograph interfaced withan Agilent 5973 MSD and equipped with a Grobinjector that was operated in the pulsed splitlessmode at 25 psi until 1.0min. The injector tempera-ture was set to 300 1C. A DB5-MS capillary column(30m� 0.25mm i.d. and film thickness of 0.25 mm, J& W Scientific, Rancho Cordova, California) wasused. The column temperature program consisted ofinjection at 60 1C and hold for 2min, temperatureincrease of 6 1Cmin�1 to 300 1C, followed by anisothermal hold at 300 1C for 15min. The MSD wasoperated in the electron impact (EI) mode with anionization energy of 70 eV. The scan range was setfrom 50 to 550 daltons. The underivatized and thesilylated total extracts were also analyzed with asecond Agilent GC–MS system (GC 6890,MSD5973), but using a DB1-MS capillary column.The MS and GC operating conditions were thesame as above.
2.4. Compound identification and quantification
Data were acquired and processed with the HP-Chemstation software. Individual compounds wereidentified by comparison of mass spectra withliterature and library data, comparison of mass
ongitude County OC (%) of PM10b
913101300 W Tulare 13.9
914300400 W Fresno 24.3
910603000 W Kern 12.7
011505300 W Fresno 22.9

ARTICLE IN PRESSW.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–4930
spectra and GC retention times with authenticstandards and/or interpretation of mass spectro-metric fragmentation patterns. Relative ion countswere converted to compound mass using the areacounts of the internal recovery standards (fivedeuterated standards added to the samples beforeextraction). Relative response factors (RRFs) wereobtained by injection of external standards thatcontained the compound of interest, and 1-phenyl-dodecane as coinjection standard. Measurementuncertainty was determined using replicate solventextractions and GC/MS analyses and the standarddeviation (STD) of each of the RRFs obtained forauthentic standards.
For compounds for which no authentic standardswere available, the RRFs and measurement un-certainties were assumed to be identical to those ofavailable standard compounds with similar chemi-cal structures, polarities, and molecular weights.
3. Results and discussion
3.1. Environmental setting
Cattle confined in open lot dairies and beef cattlefeedlots can be a substantial source for fugitive dust.Fugitive dust emissions from OLCCO depend atleast on four major factors: (1) The size of thefeedlot or dairy farm, including number of cattle,and cattle density. Space allocated per head of cattledepends mainly on the annual total rainfall, withlower space requirements for arid regions. For openlot dairies the space allocated per head rangesfrom 50 to 120m2 and for feedlots from 7 to 40m2.(2) The daily temperature cycle and rainfallpotential indirectly control how long manuredroppings need to dry; this is mainly a function ofthe geographic location and climate. (3) Lot design,manure management, ground moisture control, andcleaning schedule. Typically the lots are paved on anincline to insure that the liquid portion of theexcrements can run off to a nearby wastewatertreatment facility. (4) The activity pattern of thecattle. For example, in a warm climate likeCalifornia, cattle emerge typically in the late after-noon from their lethargy and move around, causingmore dust from dried manure and soil to be kickedup in the early evening hours. With the tendency ofthe atmospheric temperature stratification to be-come more stable after sundown, dense dust cloudscan persist throughout the night until the earlymorning.
In the 1970s, Algeo et al. (1972) and Elam et al.(1971) determined the total suspended particulatematter (TSP) concentrations downwind of 25California feedlots during summer in the absenceof dust control measures. Daily peak concentrationswere typically observed after sundown between 7and 10 pm with TSP levels ranging from about 2000to 35,500 mgm�3 with an average peak concentra-tion of 14,200 mgm�3. Depending on fugitive dustcontrol practices, these very high concentrations canbe much lower, typically 10–30% of that with nodust control measures.
The particle size distribution of airborne dustfrom OLCCO has been determined by severalresearchers for different locations. The ratio ofPM10 to TSP ranges from 40% to 48.2% and theratio of PM2.5 to TSP ranges from 5% to 6%(ARB, 2003; OMNI, 1989; Sweeten et al., 1998).For the soil samples considered here, Chow andcoworkers (2003) obtained PM10 samples byresuspension of the original surface soil samplesand determined the OC content to range from12.7% to 24.3%, see also Table 1. Depending on thecorrection factor to estimate organic matter contentfrom these OC data, it can be assumed that about20–40% of the PM10 emitted from concentratedcattle operations consists of organic matter.
US-EPA emission factors for TSP and PM10 forfugitive dust from feedlot operations are availablefor dry season only; see AP-42, USEPA (1986).According to AP-42, 17.2 tons of PM10 is releasedto the atmosphere per 1000 head throughput. In afeedlot with 100,000 cattle at a time, where cattlestay 4–5 months for fattening, roughly 200,000cattle enter and leave the feedlot per year. Thisamounts to almost 3500 tons of PM10 emitted peryear for a feedlot designed to hold 100,000 head ofcattle at any time and in the absence of any fugitivedust control measures. Although this is an upperestimate for cattle PM10 contributions, it does notinclude any particulate emissions from farm vehiclesused for fodder, manure, and animal transport,feedstuff processing, and other tasks. Consequently,at such particulate matter emission rates, nearbytowns are likely to be impacted by organic dustfrom concentrated cattle operations.
3.2. Cattle diet and metabolism
The organic matter content of fugitive dustemissions from concentrated cattle operations aremainly related to the original diet and chemical

ARTICLE IN PRESSW.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–49 31
conversions in the digestive tract of cattle. Cattle areruminants and therefore have, unlike many otheranimals, a stomach with four compartments utiliz-ing specialized microorganisms for digestion of evenlow quality fodder. Therefore, feces from cattle is inmany ways chemically different from that of otheranimals raised in large farms such as pigs orchickens. Hence, a brief review on feedstuff andfeeding practice will help to aid in selecting uniqueorganic marker compounds for OLCCO.
Open lot dairy farms and feedlots are occupiedtypically by large numbers of cattle. In contrast topasture and open range cattle that feed on nativeplants by availability and preference, cattle inconcentrated cattle operations are subjected tofeeding strategies with the goal to enhance yieldand performance. For feedlots, the objective is tomaximize weight gain, while for dairy cows, tomaximize milk production. The volumetric capacityof the cattle’s gut for fodder places an upper limiton how much feedstuff can be consumed per day.The higher the content of forage and other fibrousfeed in the diet, the sooner the cattle needs to stopeating. By carefully selecting feed processing, therate and extent of digestion can be enhanced. Inmodern mass cattle feeding operations, the rumi-nant diet can consist of a great variety of feedstuffs,ranging from fresh, hay, or silage of a wide varietyof forages, processed grains, feed supplements suchas minerals, vitamins, fats (oilseeds and tallowblends), pharmaceuticals and others. In California,some of the major ruminant feeds include forages,particularly alfalfa, fresh or as hay or silage, andsorghum hay. Concentrate feedstuff can consist of acombination of the following fodder items: corn assilage, ground or as hominy; wheat as silage ormiddlings, or derived from other processing meth-ods; oats; barley; distillers dried grains, soybean,canola, cottonseed, sunflower meal (or other fatsources such beef tallow or pork lard); beet pulp;almond hulls; and others (e.g., Amaral-Philips andHemken, 1997; Drake et al., 2002; George et al.,2002; Getachew et al., 2004).
With the cattle droppings, undigested organiccompounds from the cattle fodder are continuouslyadded to the ground, leading to their overallenrichment in the top soil. Depending on theruminant diet, the organic composition in thesurface soil can vary, and with it, the compositionof fugitive dust from OLCCO. In general, the majororganic compounds found are derived from epicu-ticular waxes and internal (generally intracellular)
lipids of forages and grains. Some of thosecompounds are altered by microbial activity duringpassage through the digestive system of the cattle,and are discussed below. The concentration oforganic compounds in the surface soil samples fromthe dairies (FDCTD1 and FDCTD2) and feedlots(FDCTF1 and FDCTF2) are given in Table 2.
3.3. Aliphatic lipids
In the soil from open lot concentrated cattleoperations, undigested plant epicuticular wax con-stituents are comprised of n-alkanes XC27 with astrong odd carbon number predominance andcarbon number maximum (Cmax) at 31, n-alkanoicacids XC22 with a strong even carbon numberpredominance and Cmax at 30, and n-alkanols XC22
also with a strong even carbon number predomi-nance and Cmax at 30 (Table 2). An example of aGC-MS total ion current trace for a total methy-lated extract is shown in Fig. 1a. Epicuticular plantwax n-alkanes are chemically very stable and cantherefore be used to estimate the intake and dietcomposition for cattle and other herbivores (e.g.,Bugalho et al., 2004; Dove and Mayes, 1996).Comparing the content of undigested plant waxaliphatic lipids in the surface soils from the dairiesand feedlots, it is evident that forage is part of thecattle diet in both dairies (FDCTD1, FDCTD2) andalso at feedlot FDCTF2. In contrast, the soil dustsample from feedlot FDCTF1 contains markedlylower levels of plant derived epicuticular waxconstituents, suggesting that forage is only a minorcomponent of the cattle diet. Minor secondary lipidproducts are comprised of n-alkanals ranging frommainly C12 to C29 and n-alkan-2-ones ranging fromC11 to C35. From the vegetation feed, unaltered andhydrogenated intracellular lipids are found in thesurface soil dust samples and consist mainly of then-alkanoic and alkenoic acids oC20 with Cmax at18, steroids, triterpenols, chlorophyll breakdownproducts and various tocopherols (Table 2).
In forage- and cereal-based feed, as well as insupplemental fat sources such as canola, sunflower,soybean, and cottonseed, palmitic acid (C16) is themajor saturated fatty acid and therefore, commonlyfound at levels two to five times higher than stearicacid (C18) (e.g., Aurousseau et al., 2004; Chilliard etal., 2001; Collomb et al., 2004; Dhiman et al., 1999;Doreau and Poncet, 2000; Elgersma et al., 2003;French et al., 2003; Whiting et al., 2004). Incontrast, the sum of the unsaturated C18 fatty acids

ARTICLE IN PRESS
Table
2
Concentrationsoforganic
compoundsin
thesoilsamples(reported
asmg
g�1)a
Compounds
Form
ula
Dairies
Feedlots
CompdID
b
FDCTD1
FDCTD2
FDCTF1
FDCTF2
n-A
lkanes
n-Pentacosane
C25H
52
0.9370.03
0.3170.01
——
a
n-H
exacosane
C26H
54
0.2270.007
0.0670.002
——
a
n-H
eptacosane
C27H
56
5.470.23
1.970.08
0.2970.01
2.1070.09
a
n-O
ctacosane
C28H
58
1.270.03
0.4070.01
0.0770.003
0.6270.02
a
n-N
onacosane
C29H
60
35.170.99
11.670.33
1.670.05
12.570.36
a
n-Triacontane
C30H
62
2.970.09
1.170.03
0.1270.003
1.270.04
a
n-H
entriacontane
C31H
64
84.272.71
32.771.05
3.770.12
39.571.27
a
n-D
otriacontane
C32H
66
1.970.06
0.7370.02
0.1870.005
1.170.03
a
n-Tritriacontane
C33H
68
7.770.24
2.270.07
0.2470.008
3.370.11
a
n-A
lkanoic
aci
ds
n-O
ctanoic
acid
C8H
16O
20.5270.021
0.1070.004
0.0270.0008
0.0970.0037
a
n-N
onanoic
acid
C9H
18O
21.170.04
0.0470.0010
0.0170.0004
0.0970.0033
a
n-D
ecanoic
acid
C10H
20O
20.9070.034
0.0670.0020
0.0170.0004
0.1370.0049
a
n-U
ndecanoic
acid
C11H
22O
20.2570.009
0.0270.0007
0.0370.0010
0.0770.0024
a
n-D
odecanoic
acid(lauricacid)
C12H
24O
21.970.07
0.2470.0084
0.0870.0028
0.7470.026
a
n-Tridecanoic
acid
C13H
26O
21.170.04
0.170.0038
0.0270.0007
0.7670.029
a
n-Tetradecanoic
acid(m
yristic
acid)
C14H
28O
211.870.50
2.870.12
0.3770.0157
11.370.48
a
n-Pentadecanoic
acid
C15H
30O
29.170.43
2.770.13
0.2070.0095
8.970.42
a
n-H
exadecanoic
acid(palm
itic
acid)
C16H
32O
2101.074.9
83.974.1
4.570.22
134.576.5
a
n-H
eptadecanoic
acid
C17H
34O
29.770.52
6.570.35
0.4070.0215
16.770.90
a
n-O
ctadecanoic
acid(stearicacid)
C18H
36O
2219.6712.0
328.0717.9
13.970.76
391.0721.3
a
n-N
onadecanoic
acid
C19H
38O
21.170.07
0.9770.06
0.0970.0058
2.070.13
a
n-Eicosanoic
acid
C20H
40O
210.370.66
9.970.63
0.6070.0381
12.470.79
a
n-H
eneicosanoic
acid
C21H
42O
21.270.10
0.8670.07
0.1770.0147
1.570.13
a
n-D
ocosanoic
acid
C22H
44O
212.270.86
8.970.63
1.470.10
10.470.73
a
n-Tricosanoic
acid
C23H
46O
23.370.28
3.070.25
0.6770.056
4.770.39
a
n-Tetracosanoic
acid
C24H
48O
214.871.37
12.671.16
2.970.27
16.371.51
a
n-Pentacosanoic
acid
C25H
50O
21.770.20
1.870.21
0.3870.045
1.670.19
a
n-H
exacosanoic
acid
C26H
52O
28.671.06
7.270.89
2.370.28
8.170.99
a
n-H
eptacosanoic
acid
C27H
54O
21.370.19
0.8470.12
0.2670.037
0.6570.093
a
n-O
ctacosanoic
acid
C28H
56O
212.971.90
10.171.49
3.570.52
7.771.14
a
n-N
onacosanoic
acid
C29H
58O
25.670.79
4.170.58
1.070.14
2.370.32
a
n-Triacontanoic
acid
C30H
60O
256.678.1
57.578.2
12.371.76
32.174.6
a
n-H
entriacontanoic
acid
C31H
62O
22.670.35
1.970.25
0.5070.067
1.270.16
a
n-D
otriacontanoic
acid
C32H
64O
213.171.81
10.471.44
3.170.43
7.371.01
a
n-Tritiacontanoic
acid
C33H
66O
20.5570.08
0.3470.048
0.1570.021
0.2770.038
a
n-Tetratriacontanoic
acid
C34H
68O
21.770.24
1.470.19
0.5970.084
1.670.23
a
n-A
lken
oic
aci
ds
Octadecenoic
acids
C18H
34O
256.473.1
63.073.4
2.670.14
123.476.7
a
Octadecadienoic
acid
C18H
32O
224.071.3
27.271.5
0.6270.034
51.172.8
a
W.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–4932

ARTICLE IN PRESSn-A
lkanols
n-O
ctadecanol
C18H
38O
2.670.09
0.8270.030
——
b
n-D
ocosanol
C22H
46O
5.670.21
2.270.081
0.4870.018
1.970.07
a
n-Tetracosanol
C24H
50O
13.270.69
4.170.21
1.270.062
6.570.34
b
n-H
exacosanol
C26H
54O
2.4170.13
1.970.10
1.470.073
2.770.14
b
n-O
ctacosanol
C28H
58O
6.3170.74
4.870.57
2.570.29
6.970.81
b
n-Triacontanol
C30H
62O
10.971.36
9.671.19
4.970.61
9.271.15
b
n-D
otriacontanol
C32H
66O
——
2.670.34
3.970.51
b
n-A
lkanals
n-D
odecanal
C12H
24O
0.1970.005
—0.1270.003
0.3370.009
a
n-Tridecanal
C13H
26O
0.3570.016
0.2170.097
0.1070.004
0.3470.016
a
n-Tetradecanal
C14H
28O
1.970.05
1.670.04
0.4270.012
3.070.08
a
n-Pentadecanal
C15H
30O
2.170.04
1.570.03
0.5670.011
3.570.07
b
n-H
exadecanal
C16H
32O
2.470.17
2.070.14
0.6270.044
2.470.17
b
n-H
eptadecanal
C17H
34O
0.5570.020
0.5270.0191
0.2070.007
0.5070.018
b
n-O
ctadecanal
C18H
36O
1.870.066
3.970.14
2.270.08
4.270.15
b
n-Eicosanal
C20H
40O
0.9670.049
3.370.17
1.670.08
2.570.13
b
n-H
eneicosanal
C21H
42O
——
0.1070.005
—b
n-D
ocosanal
C22H
44O
1.170.13
0.7770.090
0.6070.070
1.370.15
b
n-Tricosanal
C23H
46O
0.9770.12
0.2970.036
0.5070.062
0.8770.108
b
n-Tetracosanal
C24H
48O
1.970.25
0.8770.115
0.7470.098
1.770.22
b
n-Pentacosanal
C25H
50O
0.6070.081
0.2370.031
0.1070.014
0.4270.057
b
n-H
exacosanal
C26H
52O
1.570.21
0.6470.089
0.4870.067
0.7070.097
b
n-H
eptacosanal
C27H
54O
0.6270.088
0.6270.088
0.2770.038
—b
n-O
ctacosanal
C28H
56O
3.570.50
2.570.36
3.470.49
2.470.35
b
n-N
onacosanal
C29H
58O
0.7470.11
1.170.16
0.8070.116
—b
n-A
lkan-2
-ones
n-D
odecan-2-one
C12H
24O
——
—0.0370.0008
a
n-Tridecan-2-one
C13H
26O
0.0570.0013
0.0270.0052
0.0170.0003
0.2770.0070
a
n-Tetradecan-2-one
C14H
28O
0.0470.0014
0.0270.0068
—0.0970.0031
a
n-Pentadecan-2-one
C15H
30O
0.1370.0044
0.1070.0034
0.0170.0003
0.5070.017
a
n-H
exadecan-2-one
C16H
32O
0.0670.0016
0.0570.0014
—0.3870.010
b
n-H
eptadecan-2-one
C17H
34O
0.1270.0033
0.0870.0022
—0.2270.0061
b
n-H
eneicosan-2-one
C21H
42O
—0.0370.0092
——
b
n-Tricosan-2-one
C23H
46O
—0.0270.0006
——
b
n-Tetracosan-2-one
C24H
48O
—0.0270.0007
——
b
n-Pentacosan-2-one
C25H
50O
0.0670.0020
0.0370.0010
——
b
n-H
exacosan-2-one
C26H
52O
0.0370.0011
0.0270.0007
——
b
n-H
eptacosan-2-one
C27H
54O
0.2270.008
0.1270.0044
——
b
n-N
onacosan-2-one
C29H
58O
0.6370.024
0.3670.014
——
b
n-H
entriacontan-2-one
C31H
62O
1.770.07
1.470.05
——
b
n-D
otriacontan-2-one
C32H
64O
0.4470.017
0.4070.016
——
b
n-Tritriacontan-2-one
C33H
66O
1.270.05
1.070.042
——
b
n-Pentatriacontan-2-one
C35H
70O
0.2570.011
0.1970.0084
——
b
Ste
roid
s
Cholesterol(cholest-5-en-3b-ol)
C27H
46O
158.4718.2
143.8716.5
119.8713.7
458.0752.5
a
Cholestanol(5a-cholestan-3b-ol)
C27H
48O
44.975.4
25.573.1
16.171.9
55.076.6
a
Coprostanol(5b-cholestan-3b-ol)
C27H
48O
62.477.1
80.279.2
86.179.9
260.0729.8
a
epi-Coprostanol(5b-cholestan-3a-ol)
C27H
48O
84.179.9
120.1714.2
142.8716.9
341.7740.4
a
Ergosterol
C28H
44O
0.9170.109
0.7270.086
0.4670.055
0.3670.043
a
W.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–49 33

ARTICLE IN PRESSTable
2(c
onti
nued
)
Compounds
Form
ula
Dairies
Feedlots
CompdID
b
FDCTD1
FDCTD2
FDCTF1
FDCTF2
Brassicasterol
C28H
46O
1.570.20
0.8470.110
0.1570.0197
0.8170.11
b
Ergostanol(24a-methyl-5a-cholestan-3b�
ol)
C28H
50O
53.777.7
36.975.3
27.273.9
113.1716.3
b
Campesterol
C28H
48O
40.675.0
34.674.3
29.173.6
79.479.9
a
5b-Campestanol(24a-methyl-5b-cholestan-3b-ol)c
C28H
52O
48.675.7
14.671.7
12.271.4
155.9718.4
b
epi-5b-Campestanol(24a-methyl-5b-cholestan-3a-ol)d
C28H
50O
64.177.9
24.773.1
40.575.0
138.1717.1
b
Stigmasterol(stigmasta-5,22-dien-3b-ol)
C29H
48O
16.872.1
19.272.4
17.372.2
33.374.2
a
Sitosterol
C29H
50O
75.4710.4
43.776.0
30.074.1
196.3727.1
a
5b-Stigmastanol(24b-ethyl-5b-cholestan-3b-ol)e
C29H
52O
71.2710.3
53.577.7
68.779.9
223.4732.3
b
epi-5b-Stigmastanol(24b-ethyl-5b-cholestan-3a-ol)f
C29H
52O
301.7744.3
225.8733.2
220.8732.4
722.37106.1
b
Stigmasta-7,16-dien-3b�
ol
C29H
48O
286.6740.3
57.578.1
52.977.4
334.4747.0
b
Stigmastanol(sitostanol)g
C29H
52O
165.9722.3
55.977.5
81.1710.9
225.4730.3
b
D7-Stigmasterol(5a-stigmast-7-en-3b-ol)
C29H
50O
98.4715.7
66.1710.6
31.175.0
38.176.1
b
Stigmasta-3,5-dien-7-one
C29H
46O
3.470.61
3.170.56
2.570.45
3.670.65
b
5b-Stigmastan-3-one
C29H
50O
12.871.9
7.471.1
5.870.85
11.071.6
b
5a-Stigmastan-3-one
C29H
50O
6.270.86
3.970.54
2.370.32
9.071.2
b
Tri
terp
enoid
s
a-Amyrin
C30H
50O
16.571.9
11.071.2
3.970.44
19.172.1
a
b-Amyrin
C30H
50O
8.971.01
6.970.78
2.370.26
10.271.2
a
Oleanonoic
acid(3-oxo-olean-12-en-28-oic
acid)
C30H
46O
31.970.26
6.070.83
--
b
Cycloartenol
C30H
50O
2.870.44
2.370.36
1.170.17
3.470.53
a
24-M
ethylenecycloartanol
C31H
52O
9.271.38
7.671.14
7.671.14
17.172.6
b
Isopre
noid
sand
Der
ivati
ves
Phytol
C20H
40O
26.072.2
25.372.1
1.570.13
23.972.0
a
Phytone
C18H
36O
2.770.23
0.7570.063
0.1670.013
1.070.08
a
Squalene
C30H
50
1.770.13
7.770.57
0.3870.028
4.570.33
a
Toco
pher
ols
and
der
ivati
ves
Vitamin
E(a-tocopherol)
C29H
50O
215.271.6
18.972.0
5.270.54
43.974.6
a
a-Tocopherylacetate
C31H
52O
36.970.76
8.170.89
0.1370.011
58.976.5
b
g-Tocopherol
C28H
48O
26.170.63
4.570.46
1.770.17
11.071.13
b
a-Tocopherolhydroquinone
C29H
52O
39.170.91
9.971.00
0.6570.69
35.673.6
b
aValues
notreported:below
detection.
bCompoundidentification:a,authenticstandard
verificationandquantification;b,MS-spectrum
verificationandquantified
usingauthenticstandard
withsimilarstructure,polarity,
andvolatility.Theindividualsample
resultsare
foundin
Rogge(2000).
c5b-Campestanol(24a-methyl-5b-cholestan-3b-ol)alsocalled
24-m
ethylcoprostanol.
depi-5b-Campestanol(24a-methyl-5b-cholestan-3a-ol)alsocalled
24-m
ethyl-epi-coprostanol.
e5b-Stigmastanol(24b-ethyl-5b-cholestan-3b-ol)alsocalled
5b-sitostanolor24-ethylcoprostanol.
f epi-5b-Stigmastanol(24b-ethyl-5b-cholestan-3a-ol)alsocalled
epi-5b-sitostanolor24-ethyl-epi-coprostanol.
gStigmastanol(sitostanol)alsocalled
24b-ethyl-5a-cholestan-3b-ol.
W.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–4934

ARTICLE IN PRESS
Fig. 1. GC–MS total ion current (TIC) traces for feedlot soil extracts: (a) total methylated extract (FDCTF2); and (b) sterol range for
total silylated extract (FDCTF2) (numbers refer to carbon chain length of homologous compound series, n ¼ normal alkanes, � ¼ methyl
alkanoate, TAc ¼ a-tocopheryl acetate, THQ ¼ a-tocopherol hydroquinone, a- and b-A ¼ a- and b-amyrin, other compounds are
labeled).
W.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–49 35
(oleic acid, C18:1; linoleic acid, C18:2; linolenic acid,C18:3) is 10 to 20 times higher than the saturated C18
homologue (stearic acid) and also 5–10 times higherthan the respective C16 unsaturated fatty acid(palmitoleic acid, C16:1). Upon ingestion, forage-and cereal-based dietary fat as triglycerides is firstenzymatically lipolysed and a large portion of theunsaturated fatty acids are then hydrogenated bygut microorganisms in the forestomach of rumi-nants to the respective saturated analogues (Doreauand Ferlay, 1994; Moate et al., 2004; Polan et al.,1964). Because of the conversion of the unsaturatedto the saturated fatty acids, it is not surprising tofind that the major fatty acid in the surface soil
samples from OLCCO is stearic acid (C18), with aconcentration three times higher (3.070.7) thanpalmitic acid (C16). This is quite in contrast to thereversed higher C16 vs. C18 fatty acid ratio found inagricultural soils, plants, and other sources that mayrelease these two major fatty acids. Despite thehydrogenation of double bonds, mono- and diun-saturated C18 fatty acids are still detectable, butonly at about half the levels as found for stearicacid.
Isoprenoid derivatives, such as phytol (3,7,11,15-tetramethylhexadec-2-en-1-ol) and its oxidation pro-duct phytone (6,10,14-trimethylpentadecan-2-one) aredetectable as digestion products from the chlorophyll

ARTICLE IN PRESSW.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–4936
in the green forage feed. These chlorophyll derivativesshow a similar trend as the epicuticular plant waxcompounds, with lowest concentrations as well forfeedlot FDCTF1. Squalene (2,6,10,15,19,23-hexam-ethyltetracosa-2,6,10,14,18,22-hexaene), the precursorin steroid biosynthesis, is present as a trace component(Table 2).
3.4. Tocopherols
Vitamin E, a natural antioxidant, is essential forgrowth, disease prevention, tissue integrity andreproduction in animal husbandry. Natural vitaminE, as it occurs in vegetation, consists primarily of a-tocopherol (I, R ¼ H, all chemical structures citedare given in Appendix I) with minor amounts of g-tocopherol (II) (e.g., Wyatt et al., 1998). Processedgrain-based cereal feed, hay, and ensiled forages aredeficient in vitamin E (McDowell et al., 1996; Yanget al., 2002). Thus, in contrast to pasture grazingcattle, concentrated cattle operations commonlysupplement the stock diets with vitamin E (e.g.,Arnold et al., 1992; Jensen et al., 1995; Lynch et al.,1999; McDowell et al., 1996; Wen et al., 1996). Thisresults in higher levels of a-tocopherol in animaltissues, which in turn increases tissue stabilitytoward lipid oxidation. Delaying lipid breakdownin cut beef also retards muscle-pigment oxidationand meat discoloration, and therefore, increases theshelf-life of fresh, frozen and vacuum-packed beef(e.g., Arnold et al., 1992; Jensen et al., 1995; Lynchet al., 1999; McDowell et al., 1996; Wen et al.,1996). Likewise, in order to maintain milk freshnessand pleasant flavor, vitamin E addition to thefeedstuff of milking cows in dairy farms is commonpractice (Focant et al., 1998; Havemose et al., 2004).Animal feed in concentrated cattle operations isnormally supplemented with a-tocopheryl acetate (I,R ¼ Ac), which is very stable to in vitro oxidationand is promptly hydrolyzed in the animal gut to a-tocopherol (e.g., Arnold et al., 1992; Jensen et al.,1995; Lynch et al., 1999; McDowell et al., 1996;Wen et al., 1996).
The dual origin of the tocopherols in these soilsamples is evident from the data shown in the keyion plot (Fig. 2a). The g-tocopherol is derivedexclusively from the vegetation feed, while a-tocopherol is derived from vegetation fodder andalso from the hydrolysis of the tocopheryl acetatefeed supplement. Excess a-tocopheryl acetate, nothydrogenated in the animal gut is detected as well inthe soil samples, varying from being the major
tocopheryl analog (feedlot FDCTF2, 58.9 mg g�1) tobeing the minor analog in soil dust from feedlotFDCTF1 (0.10 mg g�1), see Table 2. Because a-tocopheryl acetate is relatively stable, its presence inthe soil dust samples indicates the degree of usage ofa-tocopheryl acetate as feed supplement. A signifi-cant metabolite of a-tocopherol elutes after theacetate and the mass spectrum is shown in Fig. 2b.The molecular weight of 448 Dalton fits with thecomposition C29H52O3 and the fragmentation pat-tern is interpreted to be the hydration product of thechroman ring of a-tocopherol, namely 1-(20,50-dihydroxy-30,40,60-trimethylphenyl)-3-hydroxy-3,7,11,15-tetramethylhexadecane (a-tocopherol hy-droquinone, III). a-Tocopherol conversion anddegradation is complex. One route leads via hepaticoxidation to chroman ring opening, yielding a-tocopherol quinone which can then be reduced toa-tocopherol hydroquinone by microsomal andmitochondrial enzymes (Shi et al., 1999;Schmandke, 1964). In biological systems, a-toco-pherol hydroquinone is the most active antioxidantof the tocopherols (Liebler and Burr, 2000; Shiet al., 1999). Here, the highest a-tocopherol hydro-quinone concentration was found for feedlotFDCTF2, which also shows the highest levels ofall tocopherols and derivatives identified.
3.5. Steroids
What cholesterol (IV, R ¼ H) is for the animalkingdom, phytosterols are for plants. Today, morethan 100 different phytosterols are known (e.g.,Moreau et al., 2002). The most common phytosterolin terrestrial plants is sitosterol (IV, R ¼ bC2H5),followed by campesterol (IV, R ¼ aCH3), stigmas-terol (V, R ¼ bC2H5), brassicasterol (V, R ¼ bCH3)and others (e.g., Moreau et al., 2002; Hartmann,1998). Phytosterols are biosynthesized by themevalonate pathway, similar to cholesterol inanimals, and the synthesis proceeds via precursorsthat include squalene and cycloartenol (e.g.,Moreau et al., 2002; Venkatramesh et al., 2003).Phytosterols (carbon numbers: 28 or 29) are similarin structure to cholesterol (carbon number: 27).They have like most 4-desmethylsterols a doublebond between carbon 5 and 6 of the ring system andare thus called D5-sterols. Unlike cholesterol,phytosterols have an additional methyl or ethylgroup in the alkyl side chain at the C-24 positionand some (e.g., stigmasterol) a second double bondalso in the side chain.

ARTICLE IN PRESS
Fig. 2. (a) Key ion fragmentogram for m/z 416 and 430 for the tocopherols, and (b) mass spectrum of a-tocopherol hydroquinone.
W.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–49 37
In the animal gut, D5-sterols are biohydrogenatedby the gut microflora to their saturated stanolequivalents with possible configurational changes atthe C-5 and C-3 positions resulting in three isomers:stanols (5aH, 3bOH, VI), coprostanols (5bH,3bOH, VII) and epi-coprostanols (5bH, 3aOH,VIII) (Smith and Gouron, 1969; Grimalt et al.,1990; Leeming et al., 1997, 1998; Skurikhin, 1984;Snog-Kjaer et al., 1956). When excreted, stanols insoil are rather stable (Bull et al., 2000; Marvin et al.,2001); and therefore, used to identify ancient animalfarms (Evershed et al., 1997). The specificity of fecalsterols and stanols depends on the diet, sterolbiosynthesis by animals, and ability to biohydro-genate sterols to stanols by anaerobic bacteria in the
digestive tract (e.g., Leeming et al., 1996; Truong etal., 2003). Depending on the animals being carni-vores, herbivores, or omnivores, the sterol intakewith the food includes: (1) mainly cholesterol; or(2) mainly phytosterols; or (3) both, cholesterol andphytosterols. Consequently, the feces of carnivorousanimals include cholesterol, the 5b-derivativescoprostanol, epi-coprostanol and related isomers.Instead, the fecal steroids of herbivorous animalsinclude in addition to its own biosynthesizedcholesterol with the biohydrogenated derivatives,mainly 5a-phytosterols and 5b-phytostanols. Sitos-terol, the most common and stigmasterol the thirdmost common phytosterol, are both transformed bybiohydrogenation mainly to 5b-stigmastanol and

ARTICLE IN PRESSW.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–4938
epi-5b-stigmastanol. Similarly, campesterol, thesecond most common phytosterol is transformedto 5b-campestanol and epi-5b-campestanol. Yeastsand many fungi also produce a 4-desmethylsterol,called ergosterol (IX, R ¼ bCH3) (Moreau et al.,2002), which is commonly associated with soilorganic matter and consequently with feedstuff.Ingested, ergosterol is also biohydrogenated to itsrespective 5b-stanol, ergostanol. Brassicasterol, an-other phytosterol, when biohydrogenated, is trans-formed as well in part to ergostanol (Moreau et al.,2002).
Large ruminant livestock such as cattle, consumea substantial amount of feed every day and produce
Fig. 3. GC–MS TIC traces for dairy farm soil extracts: (a) sterol rang
extract (FDCTD1) (numbers and abbreviations as in Fig. 1).
equally large amounts of dung. Thus, it is notsurprising, that the steroid mixture in these soil dustsamples from OLCCO is comprised of the originaland altered steroids ranging from C27 to C29
(Fig. 3a). Of all compound classes quantified, thesteroids are the most abundant compound group inthese soil samples, which is obvious in the examplesof the GC–MS chromatograms for the total extracts(e.g., Figs. 1a and 3b) and abundance (Table 2).Some of the isomers coelute in the underivatizedGC–MS analyses and thus are better resolved (alsowith better response factors) as the silylatedderivatives (Figs. 1b and 3b). The Kovats retentionindices are given in Table 3 for the free and silylated
e for total methylated extract (FDCTD1) and (b) total silylated

ARTICLE IN PRESSW.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–49 39
compounds. The primary (or precursor) steroids(e.g., Fig. 3a) are minor compared to the hydro-genated derivatives. Cholesterol ranges from 120 to458 mg g�1, campesterol from 29 to 80 mg g�1 andsitosterol from 44 to 196 mg g�1 (Table 2). Thesevalues are very high compared to uncontaminatednatural sediments and crop fields, where cholesterolor sitosterol are generally o1 mg g�1 (e.g., Chan etal., 1998; Rogge et al., 2006; Seguel et al., 2001).
The coprostanols and epi-coprostanols are thedominant isomers, with the stanols at lower levels(e.g., Figs. 1 and 2). The coprostanols and epi-coprostanols are poorly resolved by GC as theunderivatized compounds but resolve as the TMSethers (Table 3). The major compound is the C29
epi-stanol and therefore the mass spectrum of epi-5b-stigmastanol (VIII, R ¼ bC2H5, 24-ethyl-epi-coprostanol) is given in Fig. 4a. Its molecularweight is at m/z 416 and fragmentation occurs byloss of CH3 to m/z 401, loss of H2O to m/z 398 anda base peak at m/z 215. This 5b-series, as well as the5a-stanols (VI, 5aH), is illustrated in Fig. 4c usingthe base peak as key ion. Epi-5b-stigmastanol hasthe highest concentrations, ranging from 221 to722 mg g�1, with cholesterol (120–458 mg g�1) andepi-coprostanol (84–342 mg g�1) as next most abun-dant (Table 2). The mass spectrum of epi-5b-stigmastanol trimethylsilyl ether (VIII, R ¼ bC2H5,OTMS) is shown in Fig. 4b. Its molecular weight isat m/z 488 and fragments by loss of CH3 to m/z
473, loss of trimethylsilanol to the base peak m/z
Table 3
Kovats retention indices for C27–C29 sterols
Compound Co
Coprostanone (5b-cholestan-3-one)a C27
Coprostanol (5b-cholestan-3b-ol)a C27
epi-Coprostanol (5b-cholestan-3a-ol)a C27
Cholesterol (cholest-5-en-3b-ol)a C27
epi-Cholestanol (5a-cholestan-3a-ol)a C27
Cholestanol (5a-cholestan-3b-ol)a C27
5b-Campestanol (24a-methyl-5b-cholestan-3b-ol) C28
epi-5b-Campestanol (24a-methyl-5b-cholestan-3a-ol) C28
Campesterol (24a-methylcholest-5-en-3b-ol)a C28
5b-Stigmastanol (24b-ethyl-5b-cholestan-3b-ol) C29
epi-5b-Stigmastanol (24b-ethyl-5b-cholestan-3a-ol) C29
Ergostanol (24a-methyl-5a-cholestan-3b-ol) C28
Sitosterol (24b-ethylcholest-5-en-3b-ol)a C29
Stigmastanol (24b-ethyl-5a-cholestan-3b-ol) C29
aConfirmed with authentic standard.
398 and further fragmentation to yield the key ionat m/z 215. The trimethylsilyl ether of 24-ethylco-prostanol (VII, R ¼ bC2H5) elutes just prior to epi-5b-stigmastanol-TMS and also has a key ion at m/z
215. Therefore, the series is shown again in Fig. 4das the TMS derivatives and the isomers are labeled.
Stanones (X) and coprostanones (XI) are minorcomponents (Table 2), comprised maily of 5a-stigma-stan-3-one and 5b-stigmastan-3-one. In the case ofdomestic sewage outfalls, coprostanone (5a(H)-chole-stan-3-one) has been shown to be the major fecalsteroid (Grimalt et al., 1990). This is, however, not thecase for raw sewage or soil from feedlots and dairieswhere the coprostanones are minor.
3.6. Triterpenoids
The triterpenoids found in these surface soilsfrom open lot dairies and feedlots are minor inconcentration and consist of the unaltered naturalproducts derived from the feed. There are twocompound groups associated with triterpenoids.The first consisting of amyrins, a-amyrin (urs-12-en-3b-ol, XII), b-amyrin (olean-12-en-3b-ol, XIII)and a trace amount of oleanonoic acid (3-oxo-olean-12-en-28-oic acid), and the second of 24-methylenecycloartanol (XIV) and cycloartenol (XV)(Table 2). These compounds are quite common inmost vegetation (Simoneit, 1986, 1989) and thus arenot source specific for fugitive dust from animalhusbandry.
mposition Kovats index (DB-5)
Free compound As TMS ether
H46O 3057 —
H48O 3193 3002
H48O 3203 3016
H46O 3230 3098
H48O 3235 3010
H48O 3250 3104
H50O 3270 3107
H50O 3284 3113
H48O 3309 3204
H52O 3366 3196
H52O 3372 3206
H50O 3362 3216
H50O 3392 3296
H52O 3415 3305

ARTICLE IN PRESS
Fig. 4. Mass spectra of (a) epi-5b-stigmastanol and (b) epi-5b-stigmastanol trimethylsilyl ether, and key ion fragmentograms of m/z 215
for (c) the stanol range of the total methylated extract of FDCTD2 and (d) the same range for the total silylated extract.
W.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–4940
4. Organic marker compounds
Determining the contribution of fugitive dustfrom OLCCO to the atmospheric particle burdenhas at best been speculative, especially whenconsidering the daily and seasonal variations indust emissions, controlled by a multitude of factors.If on the other hand suitable organic markercompounds were available for fugitive dust emis-sions from OLCCO, then it would be possible todetermine the actual contributions to a given site ofinterest. For an organic compound to be suitable asmarker: (1) the emissions should be high enough sothat it can be measured in the atmosphere overappreciable distances; (2) it should be chemicallystable so that it is not depleted significantly duringatmospheric transport from the source to thereceptor site; (3) it should be unique for the sourceof interest, if not unique; then (4) a set ofcompounds should reveal a source specific concen-tration pattern that is unique; and (5) it should beassociated with the particle size fraction underconsideration. Based on these conditions, possiblecandidates for organic markers of fugitive dustassociated with surface soil from OLCCO are: the
ratio of C18/C16 n-alkanoic acids, tocopherols andderivatives, phytosterols and phytostanols. Theseare examined next to assess their source specificityand suitability for source/receptor apportionmentstudies.
For a given diet and large number of animals,cattle dung is a more or less homogeneous material.Because dried cattle dung is pulverized by themilling action of cattle, fine as well as coarseparticulate matter contains the same organic andinorganic matter. Therefore, potential organicmarkers, derived here from surface soil samplescontaining dried cattle feces, are likely present inairborne fugitive dust from OLCCO.
4.1. Aliphatic lipids
The aliphatic lipids that are associated with thesurface soils at OLCCO are derived from undigestedepicuticular plant waxes and intracellular plantmatter. Lipids associated with epicuticular plantwaxes are also released directly to the atmosphereby abrasion of waxy protrusions from leaves (Roggeet al., 1993c), as well as from windblown dust fromagricultural fields (Rogge, 1999; Rogge et al., 2006).

ARTICLE IN PRESSW.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–49 41
The most prominent intracellular aliphatic lipids arepalmitic acid (C16), stearic acid (C18), and theirunsaturated homologues. As discussed before, theratio of stearic acid (C18) to palmitic acid (C16) isunique. In Fig. 5, the C18/C16 fatty acid ratio isshown for the surface soils from the dairy farms andfeedlots investigated here, and for other importantsources of particulate matter. The surface dustsfrom OLCCO show an average C18/C16 fatty acidratio of 3.0, while all other sources for particulatematter have C18/C16 ratios ranging from 0.17 to0.71, depending on source type. Consequently, atleast in the vicinity of OLCCO, the ratio for C18/C16
fatty acids can be used, both as a qualitative andquantitative tool to determine the spread of thefugitive dust plume, fall-out of fugitive dust, as wellas for source/receptor modeling studies. In countrieswhere cattle dung is used for cooking purposes, fineparticulate smoke from the combustion of driedcattle dung also has the characteristically elevatedC18/C16 fatty acid ratio, see Fig. 5 (Sheesley et al.,2003). Consequently, the ratio of C18/C16 fatty acidcan also be used as marker and indicator in areaswere cattle dung is burned for domestic purposes.
Fat
ty A
cids
Rat
io =
C18
/C16
0.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0
Dai
ries
& F
eedl
ots
Cot
ton
Fie
lds
(a)
Saf
flow
er F
ield
s (a
)
Tom
ato
Fie
lds
(a)
Alm
ond
Orc
hard
s (a
)
Vin
eyar
ds (
a)
Pav
ed R
oad
Dus
t (S
JV)
(a)
Unp
aved
Roa
d D
ust (
SJV
) (a
)
Cat
tle D
ung
Sm
oke
(b)
Pav
ed R
oad
Dus
t (LA
) (c
)
Surface Soil and Dust Samples
Fig. 5. Fatty acid ratio ¼ C18/C16 in surface soils of open lot dairies
unpaved roads, and in PM2.5 of many urban and rural sources. (a) Rog
(d) Hays et al. (2002); (e) Rogge et al. (1993c); (f) Rogge et al. (1991); (g
(j) Rogge et al. (1998); (k) Schauer et al. (2001); (m) Rogge et al. (1993
4.2. Tocopherols
Natural tocopherols have also been found insurface soils from fields growing different crops(Rogge, 1999; Rogge et al., 2006). Because toco-pherols are antioxidants they can degrade quickly.However, excess synthetic a-tocopheryl acetate thatis fed to cattle has not been found in other sourcesfor fugitive dust. But the tocopherol concentrationsin surface soils of OLCCO are typically low andvariable. Thus, only the more stable a-tocopherylacetate may be suitable to examine the spread andout-fall of fugitive dust from OLCCO.
4.3. Sterols and stanols
Sterols and especially their hydrogenated stanolsare chemically quite stable and therefore, arefrequently used as markers to differentiate sewagespills into surface waters from human sources(mainly coprostanol) and herbivores (mainly 5b-stigmastanol and epi-5b-stigmastanol) (e.g., Elhm-mali et al., 2000; Isobe et al., 2002; Leeming et al.,1996; Standley et al., 2000), in archeology (Evershed
Fol
iar
Veg
etat
ion
Sm
oke
(d)
Wax
y Le
af S
urfa
ce A
bras
ions
(e)
Ham
burg
er C
harb
roili
ng (
f)
Ham
burg
er C
harb
roili
ng (
g)
Stir
fryi
ng V
eget
able
s in
Oils
(h)
Stir
fryi
ng V
eget
able
s in
Oils
(i)
Woo
d S
mok
e (j)
Woo
d S
mok
e (k
)
Car
Exh
aust
(l)
Die
sel T
ruck
s E
xhau
st (
l)PM 2.5
and beef cattle feedlots, agricultural fields, dust from paved and
ge et al. (2006); (b) Sheesley et al. (2003); (c) Rogge et al. (1993b);
) Schauer et al. (1999), (h) Rogge (1999); (i) Schauer et al. (2002);
a).
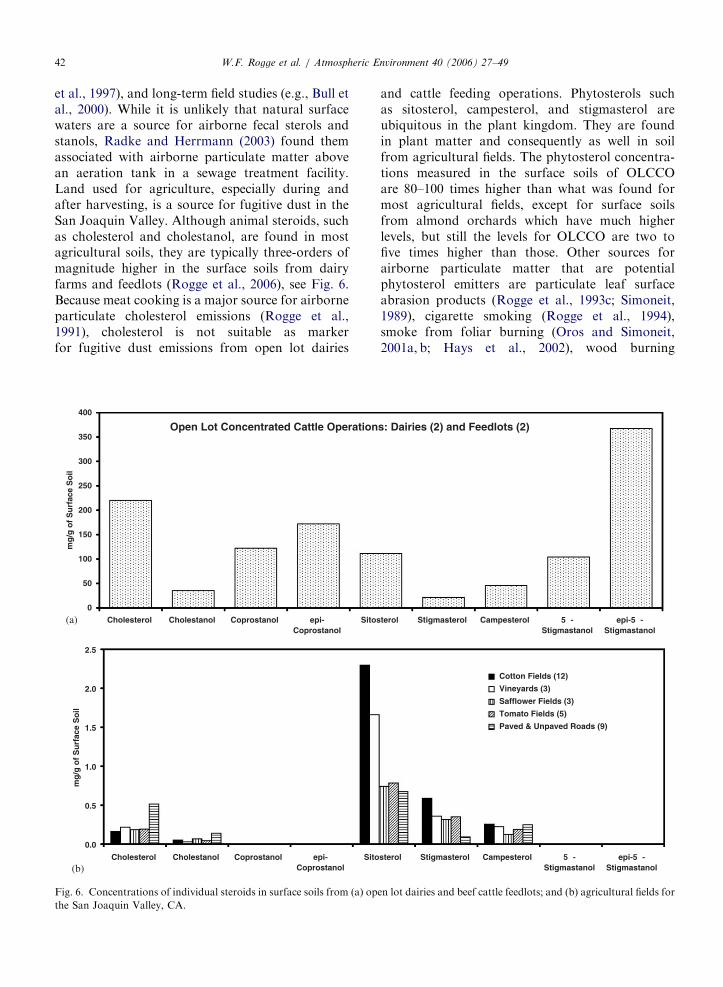
ARTICLE IN PRESSW.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–4942
et al., 1997), and long-term field studies (e.g., Bull etal., 2000). While it is unlikely that natural surfacewaters are a source for airborne fecal sterols andstanols, Radke and Herrmann (2003) found themassociated with airborne particulate matter abovean aeration tank in a sewage treatment facility.Land used for agriculture, especially during andafter harvesting, is a source for fugitive dust in theSan Joaquin Valley. Although animal steroids, suchas cholesterol and cholestanol, are found in mostagricultural soils, they are typically three-orders ofmagnitude higher in the surface soils from dairyfarms and feedlots (Rogge et al., 2006), see Fig. 6.Because meat cooking is a major source for airborneparticulate cholesterol emissions (Rogge et al.,1991), cholesterol is not suitable as markerfor fugitive dust emissions from open lot dairies
(a)
(b)
0.0
0.5
1.0
1.5
2.0
2.5
Cholesterol Cholestanol Coprostanol epi-Coprostanol
Sito
mg
/g o
f S
urf
ace
So
il
Open Lot Concentrated Cattle Operation
0
50
100
150
200
250
300
350
400
Cholesterol Cholestanol Coprostanol epi-Coprostanol
Sitos
mg
/g o
f S
urf
ace
So
il
Fig. 6. Concentrations of individual steroids in surface soils from (a) op
the San Joaquin Valley, CA.
and cattle feeding operations. Phytosterols suchas sitosterol, campesterol, and stigmasterol areubiquitous in the plant kingdom. They are foundin plant matter and consequently as well in soilfrom agricultural fields. The phytosterol concentra-tions measured in the surface soils of OLCCOare 80–100 times higher than what was found formost agricultural fields, except for surface soilsfrom almond orchards which have much higherlevels, but still the levels for OLCCO are two tofive times higher than those. Other sources forairborne particulate matter that are potentialphytosterol emitters are particulate leaf surfaceabrasion products (Rogge et al., 1993c; Simoneit,1989), cigarette smoking (Rogge et al., 1994),smoke from foliar burning (Oros and Simoneit,2001a, b; Hays et al., 2002), wood burning
sterol Stigmasterol Campesterol 5 -Stigmastanol
epi-5 -Stigmastanol
Cotton Fields (12)
Vineyards (3)
Safflower Fields (3)
Tomato Fields (5)
Paved & Unpaved Roads (9)
s: Dairies (2) and Feedlots (2)
terol Stigmasterol Campesterol 5 -Stigmastanol
epi-5 -Stigmastanol
en lot dairies and beef cattle feedlots; and (b) agricultural fields for

ARTICLE IN PRESSW.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–49 43
smoke (Rogge et al., 1998; Schauer et al., 2001),as well as wild fires. Consequently, by itself,phytosterols are not source specific for fugitivedust from OLCCO but may still be usable fornear source fall-out and plume dispersiondetermination.
In contrast, phytostanols such as 5b-stigmastanol(VII, R ¼ bC2H5) and epi-5b-stigmastanol (VIII,R ¼ bC2H5), the compounds with the highestconcentrations found in surface soils from OLCCO,are not detectable in surface soils from agriculturalfields (see Fig. 6) or any other source type thatrelates to biomass and emits particulate matter tothe atmosphere. These 5b-phytostanols are found insurface waters contaminated with waste fromanimal farms (Leeming et al., 1996; Standleyet al., 2000). Moreover, phytostanols are chemicallyeven more stable than their parent phytosterols(Bull et al., 2000). Consequently, 5b-stigmastanoland epi-5b-stigmastanol are the best potentialorganic marker compounds for fugitive dust emis-sions from OLCCO. Furthermore, smoke fromburning of dried cattle dung has also been foundto contain 5b-phytostanols (Sheesley et al., 2003). InAfrica and South Asia, where dried cattle dung isused as fuel, and where OLCCO are not typical,phytostanols can be used as organic markers forsmoke from burning dried dung.
Currently only limited information is available onthe contributions of particulate matter from aera-tion tanks of wastewater treatment tanks (Radkeand Herrmann, 2003). Therefore, should aerosolemissions occur from sewage treatment aerationtanks near a site considered for source apportion-ment of airborne particulate matter, then 5b-stigmastanol and epi-5b-stigmastanol should beused in conjunction with the following additionalsteroids: coprostanol (VII, R ¼ H), epi-coprostanol(VIII, R ¼ H), cholesterol (IV, R ¼ H), cholestanol(VI, R ¼ H), stigmastanol (VI, R ¼ bC2H5), andsitosterol (IV, R ¼ bC2H5).
5. Conclusions
Open lot dairy farms and beef cattle feedlots areconsidered to be an important source for fugitive
dust emissions in the San Joaquin Valley ofCalifornia. The organic compound composition ofsurface soils from two open lot dairy farms and twobeef feedlots has been studied by GC–MS. Morethan 100 individual compounds were quantified,including n-alkanes, n-alkanoic acids, n-alkenoicacids, n-alkanols, n-alkanals, n-alkan-2-ones, ster-oids, triterpenoids, isoprenoids, tocopherols, andothers. Some of the more abundant compoundsidentified in the surface soils include sterols, longchain n-alkanes and n-alkanoic acids from plantwax, and palmitic and stearic acid. Of particularinterest are organic compounds derived from fodderthat are biohydrogenated in the rumen of cattle. Themost promising marker compounds for fugitive dustfrom open lot dairy farms and cattle feedlots arederived from the biohydrogenation of phytosterolsin feed. Here, suitable marker compounds have beenidentified and verified in relation to other sourceprofiles. It has been found, that 5b-stigmastanol andepi-5b-stigmastanol are unique markers. In addi-tion, the ratio of C18/C16 fatty acids in surface soilsfrom open lot concentrated cattle operations ismarkedly higher (3.070.7) than for all otherpotential sources (average: 0.17–0.71). Further-more, antioxidant feed supplements (vitamin Eproducts) are also detectable in the surface soilsand the more stable synthetic a-tocopheryl acetate isa supporting source marker were supplementalvitamin E in form of a-tocopheryl acetate is fed tocattle.
Acknowledgements
This research was supported by the California AirResources Board under Agreement 98-3PM.P.M.M. thanks CNPq (Brazilian National ResearchCouncil) for financial support (No. 200330/01-2).
Appendix I
Chemical structures cited in the text are given asfollows:

ARTICLE IN PRESSW.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–4944

ARTICLE IN PRESSW.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–49 45
References
Algeo, J.W., Elam, C.J., Martinez, A., Westing, T., 1972. Feedlot
Air, Water and Soil Analysis: Bulletin D, How to Control
Feedlot Pollution. California Cattle Feeders Association,
Bakersfield, CA.
Amaral-Philips, D.M., Hemken, R.W., 1997. Using byproducts
to feed cattle. Cooperative extension service, University of
Kentucky. Publication: ASC-136. http://www.ca.uky.edu/
agc/pubs/asc/asc136/asc136.pdf.
ARB, 2003. Proposed Update of ARB Particulate Matter (PM)
Chemical Speciation and Size Fraction Data, www.arb.
ca.gov/EMISINV/speciate/pmtbl.htm.
Arnold, R.N., Scheller, K.K., Arp, S.C., Williams, S.N., Buege,
D.R., Schaefer, D.M., 1992. Effect of long- or short-term
feeding of a-tocopheryl acetate to Holstein and crossbred beef
steers on performance, carcass characteristics and beef color
stability. Journal of Animal Science 70, 3055–3067.
Ashbaugh, L.L., Carvacho, O.F., Brown, M.S., Chow, J.C.,
Watson, J.G., Magliano, K.C., 2003. Soil sample collection
and analysis for the fugitive dust characterization study.
Atmospheric Environment 37, 1163–1173.
Aurousseau, B., Bauchart, D., Calichon, E., Micol, D., Priolo, A.,
2004. Effect of grass or concentrate feeding systems and rate of
growth on triglyceride and phospholipid and their fatty acids in
the M. lonissimus thoracis of lambs. Meat Science 66, 531–541.

ARTICLE IN PRESSW.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–4946
Bessot, J.C., Blaumeister, M., Kopferschmitt, M.C., Pauli, G.,
1996. Occupational asthma in a farming environment. Revue
des Maladies Respiratoires 13, 205–215.
Bugalho, M.N., Dove, H., Kelman, W., Wood, J.T., Mayes,
R.W., 2004. Plant wax alkanes and alcohols as herbivore diet
composition markers. Journal of Range Management 57,
259–268.
Bull, I.D., Bergen, P.F., Nott, C.J., Poulton, P.R., Evershed,
R.P., 2000. Organic geochemical studies of soils from the
Rothamsted classical experiments. The fate of lipids in
different long-term experiments. Organic Geochemistry 31,
389–408.
Carvacho, O.F., Ashbaugh, L.L., Brown, M.S., Flocchini, R.G.,
2001. Relationship between San Joaquin Valley soil texture
and PM10 emission potential using the UC Davis dust
resuspension test chamber. Transaction of the ASAE 44,
1603–1608.
Cathomas, R.L., Bruesch, H., Fehr, R., Reinhart, W.H., Kuhn,
M., 2002. Organic dust exposure in dairy farmers in an alpine
region. Swiss Medical Weekly 132, 174–178.
Chan, K.-H., Lam, M.H.W., Poon, K.-F., Yeung, H.-Y., Tony,
K.T., 1998. Application of sedimentary fecal stanols and
sterols in tracing sewage pollution in coastal waters. Water
Research 32, 225–235.
Chilliard, Y., Ferlay, A., Doreau, M., 2001. Effect of different
types of forages, animal fat or marine oils in cow’s diet on
milk fat secretion and composition, especially conjugated
linoleic acid (CLA) and polyunsaturated fatty acids. Live-
stock Product Science 70, 31–48.
Chow, J.C., Watson, J.G., Lowenthal, D.H., Solomon, P.A.,
Magliano, K.L., Ziman, S.D., Richards, L.W., 1992. PM10
source apportionment in California’s San Joaquin Valley.
Atmospheric Environment 26A, 3335–3354.
Chow, J.C., Watson, J.G., Lowenthal, D.H., Solomon, P.A.,
Magliano, K.L., Ziman, S.D., Richards, L.W., 1993. PM10
and PM2.5 compositions in California’s San Joaquin Valley.
Aerosol Science and Technology 18, 105–128.
Chow, J.C., Watson, J.G., Ashbaugh, L.L., Magliano, K.L.,
2003. Similarities and differences in PM10 chemical source
profiles for geological dust from the San Joaquin Valley,
California. Atmospheric Environment 37, 1317–1340.
Clausnitzer, H., Singer, M.J., 2000. Environmental influence on
respirable dust production from agricultural operations in
California. Atmospheric Environment 34, 1739–1745.
Collomb, M., Sollberger, H., Butikofer, U., Sieber, R., Stoll, W.,
Schaeren, W., 2004. Impact of basal diet of hay and fodder
beet supplemented with rapeseed, linseed and sunflowerseed
on the fatty acid composition of milk fat. International Dairy
Journal 14, 549–559.
Dhiman, T.R., Helmink, E.D., McMahon, D.J., Fife, R.L.,
Pariza, M.W., 1999. Conjugated linoleic acid content of milk
and cheese from cows fed extruded oilseeds. Journal of Dairy
Science 82, 412–419.
Doreau, M., Ferlay, A., 1994. Digestion and utilization of fatty
acids by ruminants. Animal Feed Science and Technology 45,
379–396.
Doreau, M., Poncet, C., 2000. Ruminal biohydrogenation of
fatty acids originating from fresh or persevered grass.
Reproduction and Nutrition Development 40, 201.
Dove, H., Mayes, R.W., 1996. Plant wax components: a new
approach to estimating intake and diet composition in
herbivores. Journal of Nutrition 126, 13–26.
Drake, D.J., Nader, G., Forero, L., 2002. Feeding Rice Straw to
Cattle. University of California, Division of Agriculture and
Natural Resources. Publication: 8079. http://anrcatalog.uc-
davis.edu.
Elam, C.J., Algeo, J.W., Westing, T., Hoki, L., 1971. Measure-
ment and Control of Feedlot Particulate Matter: Bulletin C.
How to Control Feedlot Pollution. California Cattle Feeders
Association, Bakersfield, CA.
Elgersma, A., Ellen, G., van der Horst, H., Muuse, B.G., Boer,
H., Tammonga, S., 2003. Comparison of fatty acid composi-
tion of fresh and ensiled perennial ryegrass (Lolium perenne
L.), affected by cultivar and regrowth interval. Animal Feed
Science and Technology 1008, 191–205.
Elhmmali, M.M., Roberst, D.J., Evershed, R.P., 2000. Combined
analysis of bile acids and sterols/stanols from riverine
particulates to assess sewage discharge and other fecal
sources. Environmental Science and Technology 34, 39–46.
Evershed, R.P., Bethell, P.H., Reynolds, P.J., Welch, N.J., 1997.
5b-Stigmastanol and related 5b-stanols as biomarkers of
manuring: analysis of modern experimental material and
assessment of the archaeological potential. Journal of
Archaeological Science 24, 485–495.
Focant, M., Mignolet, E., Marique, M., Clabots, F., Breyne, T.,
Dalemans, D., Larondelle, Y., 1998. The effect of vitamin E
supplementation of cow diets containing rapseed and linseed
on the prevention of milk fat oxidation. Journal of Dairy
Science 81, 1095–1101.
French, P., O’Riordan, E.G., Monahan, F.J., Caffrey, P.J.,
Moloney, A.P., 2003. Fatty acid composition of intra-
muscular triacylglycerols of steers fed autumn grass and
concentrations. Livestock Production Science 81, 307–317.
George, M.R., Kearney, T.E., Schoner, C.A., 2002. Oat hay and
silage production. Division of agricultural science, University
of California Davis, Publication No. 21265e. http://anrcata-
log.ucdavis.edu.
Getachew, G., Robinson, P.H., DePeters, E.J., Taylor, S.J., 2004.
Relationships between chemical composition, dry matter
degradation and in vitro gas production of several ruminant
feeds. Animal Feed Science and Technology 111, 57–71.
Ginting, D., Kessavalou, A., Eghball, B., Doran, J.W., 2003.
Greenhouse gas emissions and soil indicators four years after
manure and compost applications. Journal of Environmental
Quality 32, 23–32.
Goss, M.J., Barry, D.A.J., Rudolph, D.L., 1998. Contamination
in Ontario farmstead domestic wells and its association with
agriculture: 1. Results from drinking water wells. Journal of
Contaminant Hydrology 32, 267–293.
Gregory, N.G., 2000. Intensive farming of animals in 2020.
Outlook on Agriculture 29, 15–23.
Grimalt, J.O., Fernandez, P., Bayona, J.M., Albaiges, J., 1990.
Assessment of fecal sterols and ketones as indicators of urban
sewage inputs to coastal waters. Environmental Science and
Technology 24, 357–363.
Hanselman, T.A., Grartz, D.A., Wilke, A.C., 2003. Manure-
borne estrogens as potential environmental contaminants: a
review. Environmental Science and Technology 37,
5411–5478.
Hartmann, M.A., 1998. Plant sterols and the membrane
environment. Trends in Plant Science 3, 170–175.
Hartung, J., 1998. Nature and amount of aerial pollutants from
livestock buildings. Deutsche Tierarztliche Wochenschrift
105, 213–216.

ARTICLE IN PRESSW.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–49 47
Havemose, M.S., Weisbjerg, M.R., Bredie, W.L.P., Nielson, J.H.,
2004. Influence of feeding different types of roughage on the
oxidative stability of milk. International Dairy Journal 14,
563–570.
Hays, M.D., Geron, C.D., Linna, K.J., Smith, N.D., Schauer,
J.J., 2002. Speciation of gas-phase and fine particle emissions
from burning of foliar fuels. Environmental Science and
Technology 36, 2281–2295.
Hinrichs, C.C., Welsh, R., 2003. The effect of the industrializa-
tion of US livestock agriculture on promoting sustainable
production practices. Agriculture and Human Values 20,
125–141.
Hintz, T., Linke, S., 1998. A comprehensive experimental study
of aerial pollutants in and emissions from livestock buildings.
Part 2: results. Journal of Agricultural Engineering Research
70, 119–129.
Isobe, K.O., Tarao, M., Zakaria, M.P., Chiem, N.H., Minh,
L.Y., Takada, H., 2002. Quantitative application of fecal
sterols using gas chromatography–mass spectrometry to
investigate fecal pollution in tropical waters: Western
Malaysia and Mekong Delta, Vietnam. Environmental
Science Technology 36, 4497–4507.
Jensen, C., Skibsted, L.H., Jakobsen, K., Bertelsen, G., 1995.
Supplementation of broiler diets will all-rac-a- or a mixture of
natural source RRR-a-, g-, d-tocopheryl acetate. 2. Effect onthe oxidative stability of raw and precooked broiler meat
products. Poultry Science 74, 2048–2056.
Jjemba, P., 2002. The potential impact of veterinary and human
therapeutic agents in manure and biosolids on plants grown
on arable land: a review. Agriculture, Ecosystems and
Environment 93, 267–278.
Kauppinen, T., Riala, R., Seitsamo, J., Hernberg, S., 1992.
Primary liver-cancer and occupational exposure. Scandina-
vian Journal of Work Environment and Health 18, 18–25.
Lammel, G., Schneider, F., Bruggemann, E., Gnauk, T., Rohrl,
A., Wieser, P., 2004. Aerosols emitted from a livestock farm
in Southern Germany. Water, Air, and Soil Pollution 154,
313–330.
Larney, F.J., Yanke, L.J., Miller, J.J., McAllister, T.A., 2003.
Fate of coliform bacteria in composted beef cattle feedlot
manure. Journal of Environmental Quality 32, 1508–1515.
Leeming, R., Ball, A., Ashbolt, N., Nichols, P., 1996. Using
faecal steroids from humans and animals to distinguish faecal
pollution in receiving waters. Water Research 30, 2893–2900.
Leeming, R., Latham, V., Rayner, M., Nichols, P., 1997.
Detecting and distinguishing sources of sewage pollution in
Australian inland and coastal waters and sediments. In:
Eganhouse, R. (Ed.), Molecular Markers in Environmental
Geochemistry, vol. 671. American Chemical Society Sympo-
sium Series, pp. 306–319.
Leeming, R., Bate, N., Hewlett, R., Nichols, P.D., 1998.
Discriminating fecal pollution: a case study of stormwater
entering Port Phillip Bay, Australia. Water Science and
Technology 38, 15–22.
Liebler, D.C., Burr, J.A., 2000. Antioxidant reactions of
a–tocopherol-hydroquinone. Lipids 35, 1045–1047.Lynch, M.P., Kerry, J.P., Buckley, D.J., Faustman, C., Morris-
sey, P.A., 1999. Effect of dietary vitamin E supplementation
on the colour and lipid stability of fresh, frozen and vacuum-
packed beef. Meat Science 52, 95–99.
Marvin, C., Coakley, J., Mayer, T., Brown, M., Thiessen, L.,
2001. Application of fecal sterol ratios in sediments and
effluents as source tracers. Water Quality Research Journal of
Canada 36, 781–792.
McDowell, L.R., Williams, S.N., Hidiroglou, N., Njeru, C.A.,
Hill, G.M., Ochoa, L., Wilkinson, N.S., 1996. Vitamin E
supplementation for the ruminant. Animal Feed Science
Technology 60, 273–296.
McGinn, S.M., Koenig, K.M., Coates, T., 2002. Effect of diet on
odorant emissions from cattle manure. Canadian Journal of
Animal Science 82, 435–444.
McGinn, S.M., Janzen, H.H., Coates, T., 2003. Atmospheric
ammonia, volatile fatty acids, and other odorants near beef
feedlots. Journal of Environmental Quality 32, 1173–1182.
McVean, D.W., Franzen, D.K., Keefe, T.J., Bennett, B.W., 1986.
Airborne particle concentration and meteorological condi-
tions associated with incidence in feedlot cattle. American
Journal of Veterinary Research 47, 2676–2682.
Moate, P.J., Chalupa, W., Jenkins, T.C., Boston, R.C., 2004. A
model to describe ruminal metabolism and intestinal absorp-
tion of long chain fatty acids. Animal Feed Science and
Technology 112, 79–105.
Molocznik, A., 2002. Qualitative and quantitative analysis of
agricultural dust in working environment. Annals of Agri-
cultural and Environmental Medicine 9, 71–78.
Molocznik, A., Zagorski, J., 2001. Exposure of farmers to dust
on private farms of various production profiles. Annals of
Agricultural and Environmental Medicine 8, 151–161.
Moreau, R.A., Whitaker, B.D., Hicks, K.B., 2002. Phytosterols,
phytostanols, and their conjugates in foods: structural
diversity, quantitative analysis, and health-promoting uses.
Progress in Lipid Research 41, 457–500.
NRC (National Research Council), 2003. Final Report: Air
emissions from Animal Feeding Operations: Current Knowl-
edge, Future Needs. National Academic Press, Washington,
DC (264pp).
OMNI, 1989. Determination of particle size distribution and
chemical composition of particulate matter from selected
sources in California. In: Houck, J. E., et al. (Eds.), OMNI
Environmental Services, Inc. and Desert Research Institute,
ARB contract #a6-175-32. Final Report, June 30.
Oros, D.R., Simoneit, B.R.T., 2001a. Identification and emission
factors of molecular tracers in organic aerosols from biomass
burning: Part 1. Temperate climate conifers. Applied Geo-
chemistry 16, 1513–1544.
Oros, D.R., Simoneit, B.R.T., 2001b. Identification and emission
factors of molecular tracers in organic aerosols from biomass
burning: Part 2. Deciduous trees. Applied Geochemistry 16,
1545–1565.
Phetteplace, H.W., Johnson, D.E., Seidl, A.F., 2001. Greenhouse
gas emissions from simulated beef and dairy livestock systems
in the United States. Nutrient Cycling in Agroecosystems 60,
99–102.
Polan, C.E., McNeill, J.J., Tove, S.B., 1964. Biohydrogenation of
unsaturated fatty acids by rumen bacteria. Journal of
Bacteriology 88, 1056–1064.
Purdy, C.W., Straus, D.C., Parker, D.B., Wilson, S.C., Clark,
R.N., 2004. Comparison of the type and number of
microorganisms and concentration of endotoxin in the air
of feedyards in the Southern High Plains. American Journal
of Veterinary Research 65, 45–52.
Radke, M., Herrmann, R., 2003. Aerosol-bound emissions of
aromatic hydrocarbons and sterols from aeration tanks of

ARTICLE IN PRESSW.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–4948
municipal waste water treatment plants. Environmental
Science and Technology 37, 2109–2113.
Radon, K., Danuser, B., Iversen, M., Jorres, R., Monso, E.,
Opravil, U., Weber, C., Donham, K.J., Nowak, D., 2001.
Respiratory symptoms in European animal farmers. Eur-
opean Respiratory Journal 17, 747–754.
Rautiainen, M., Virtanen, T., Ruoppi, P., Nuutinen, J.,
Mantyjarvi, R., 1997. Humoral responses to bovine dust in
dairy farmers with allergic rhinitis. Acta Oto-Laryngologica,
169–172.
Rogge, W.F., 1999. Investigating indoor air quality by using a
dynamic and modular source testing approach for indoor
cooking. Final Report to the Center for Indoor Air Research,
333pp.
Rogge, W.F., 2000. Development of profiles for distinguishing
dust emission sources using organic tracers, Final Report to
the California Air Resources Board, Contract No. 98-3PM,
158pp.
Rogge, W.F., Mazurek, M.A., Hildemann, L.M., Cass, G.R.,
Simoneit, B.R.T., 1991. Sources of fine organic aerosol: 1.
Charbroilers and meat cooking operations. Environmental
Science and Technology 25, 1112–1125.
Rogge, W.F., Mazurek, M.A., Hildemann, L.M., Cass, G.R.,
Simoneit, B.R.T., 1993a. Sources of fine organic aerosol: 2.
Noncatalyst and catalyst-equipped automobiles and heavy-
duty diesel trucks. Environmental Science and Technology 27,
1892–1904.
Rogge, W.F., Hildemann, L.M., Mazurek, M.A., Cass, G.R.,
Simoneit, B.R.T., 1993b. Sources of fine organic aerosol: 3.
Road dust, tire debris, and organometallic brake lining dust.
Environmental Science and Technology 27, 1892–1904.
Rogge, W.F., Hildemann, L.M., Mazurek, M.A., Cass, G.R.,
Simoneit, B.R.T., 1993c. Sources of fine organic aerosol: 4.
Particulate abrasion products from leaf surfaces of urban
plants. Environmental Science and Technology 27,
2700–2711.
Rogge, W.F., Hildemann, L.M., Mazurek, M.A., Cass, G.R.,
Simoneit, B.R.T., 1994. Sources of fine organic aerosol: 6.
Cigarette smoke in the urban atmosphere. Environmental
Science and Technology 28, 1375–1388.
Rogge, W.F., Hildemann, L.M., Mazurek, M.A., Cass, G.R.,
Simoneit, B.R.T., 1998. Sources of fine organic aerosol: 9.
Pine, oak, and synthetic log combustion in residential
fireplaces. Environmental Science and Technology 32, 13–22.
Rogge, W.F., Medeiros, P.M., Simoneit, B.R.T., 2006. Organic
tracers in surface soils of crop fields from the San Joaquin
valley fugitive dust characterization study. Atmospheric
Environment, in preparation.
Saintfort, R., Raina, R.M., Prescott, B., 1995. Subsurface
environmental-quality under a cattle feedlot and adjacent
cropfield. Journal of Environmental Science and Health Part
A—Environmental Science and Engineering &Toxic and
Hazardous Substance Control 30, 637–650.
Schauer, J.J., Kleeman, M.J., Cass, G.R., Simoneit, B.R.T., 1999.
Measurement of emissions from air pollution sources: 1.
C1–C29 organic compounds from meat charbroiling. Environ-
mental Science and Technology 33, 1566–1577.
Schauer, J.J., Kleeman, M.J., Cass, G.R., Simoneit, B.R.T., 2001.
Measurement of emissions from air pollution sources: 3.
C1–C29 organic compounds from fireplace combustion of
wood. Environmental Science and Technology 35, 1716–1728.
Schauer, J.J., Kleeman, M.J., Cass, G.R., Simoneit, B.R.T., 2002.
Measurement of emissions from air pollution sources: 3.
C1–C27 organic compounds from cooking with seed oils.
Environmental Science and Technology 36, 567–575.
Schmandke, H., 1964. Fate of the tocopherols in the animal
organism. Nahrung 8, 340–350.
Scott, H.M., Soskolne, C.L., Martin, S.W., Shoukri, M.M.,
Lissemore, K.D., Coppock, R.W., Guidotti, T.L., 2003. Air
emissions from sour-gas producing plants and dairy-cattle
reproduction in Alberta, Canada. Preventive Veterinary
Medicine 57, 69–95.
Seguel, C.G., Mudge, S.M., Salgado, C., Toledo, M., 2001.
Tracing sewage in the marine environment: altered signatures
in Concepcion Bay, Chile. Water Research 35, 4166–4174.
Shaw, B.W., Buharivala, P.P., Parnell, C.B., Demny, M.A., 1998.
Emission factors for grain receiving and feed loading
operations at feed mills. Transactions of the ASAE 41,
757–765.
Sheesley, R.J., Schauer, J.J., Chowdhury, Z., Cass, G.R.,
Simoneit, B.R.T., 2003. Characterization of organic aerosols
emitted from the combustion of biomass indigenous to South
Asia. Journal of Geophysical Research 108 Art. No. 4285.
Shi, H.-L., Noguchi, N., Niki, E., 1999. Comparative study on
dynamics of antioxidative action of a–tocopheryl hydroqui-none, ubiquinol, and a–tocopherol against lipid peroxidation.
Free Radical Biology and Medicine 27, 334–346.
Simoneit, B.R.T., 1986. Cyclic terpenoids in the geosphere. In:
Johns, R.B. (Ed.), Biological Markers in the Sedimentary
Record. Elsevier Science Publishers, Amsterdam, pp. 43–99.
Simoneit, B.R.T., 1989. Organic matter of the troposphere. V.
Application of molecular marker analysis to biogenic emis-
sions into the troposphere for source reconciliations. Journal
of Atmospheric Chemistry 8, 251–275.
Skurikhin, V.N., 1984. Identification of neutral steroids in the
diet, chyme and feces of heifers by chromatography and mass
spectrometry. Doklady Vsesoyuznoi Akademii Sel’skokho-
zyaistvennykh Nauk 7, 42–44.
Smith, L.L., Gouron, R.E., 1969. Sterol metabolism. VI.
Detection of 5b-cholestan-3b-ol in polluted waters. Water
Research 3, 141–148.
Snog-Kjaer, A., Prange, I., Dam, H., 1956. Conversion of
cholesterol into coprosterol by bacteria in vitro. Journal of
General Microbiology 14, 256–260.
Sorenson, W.G., Siegel, P.D., Olenchock, S.A., May, J.J., Pratt,
D.S., 1992. Fungi in aerosols of hay associated with
respiratory-distress in dairy-cattle. International Biodeter-
ioration & Biodegradation 30, 353–363.
Standley, L.J., Kaplan, L.A., Smith, D., 2000. Molecular tracers
of organic matter sources to surface water resources.
Environmental Science and Technology 34, 3124–3130.
Subak, S., 1999. Global environmental costs of beef production.
Ecological Economics 30, 79–91.
Sweeten, J.M., Miner, J.R., 1993. Odor intensities at cattle
feedlots in nuisance litigation. Bioresource Technology 45,
177–188.
Sweeten, J.M., Parnell, C.B., Shaw, B.W., Auvermann, B.W.,
1998. Particle size distribution of cattle feedlot dust emission.
Transactions of the ASAE 41, 1477–1481.
Takai, H., Pedersen, S., Johnsen, J.O., Metz, J.H.M., Koerkamp,
P.W.G.G., Uenk, G.H., Phillips, V.R., Holden, M.R.,
Sneath, R.W., Short, J.L., Whilte, R.P., Hartung, J., Seedorf,
J., Schroder, M., Linkert, K.H., Wathes, C.M., 1998.

ARTICLE IN PRESSW.F. Rogge et al. / Atmospheric Environment 40 (2006) 27–49 49
Concentrations and emissions of airborne dust in livestock
buildings in Northern Europe. Journal of Agricultural
Engineering Research 70, 59–77.
Todd, R.W., Guo, W., Steward, B.A., Robinson, C., 2004.
Vegetation, phosphorus, and dust gradients downwind from a
cattle feedyard. Journal of Range Management 57, 291–299.
Truong, T.T., Marriott, P.J., Porter, N.A., Leeming, R., 2003.
Application of comprehensive two-dimensional gas chroma-
tography to the quantification of overlapping faecal sterols.
Journal of Chromatography A 1019, 197–210.
USDA (U.S. Department of Agriculture), 2000. Confined
livestock air quality subcommittee. In: Sweetem, J.M. (Ed.),
Chair. Air Quality Research and Technology Transfer
Program for Concentrated Animal Feeding Operations.
USDA Agricultural Task Force (AAQTF) Meeting, Wa-
shington, DC.
USDA (US Department of Agriculture), 2004. USDA-National
Agricultural Statistics Service (NASS). www.usda.gov/nass/.
USEPA (US Environmental Protection Agency), 1986. Supple-
ment A to compilation of air pollution emission factors,
Section 6.15 Beef Cattle Feedlots (Stationary point and area
sources, Vol. 1) AP-42. Office of Air Quality Planning and
Standards, Research Triangle Park, NC.
Venkatramesh, M., Karunanandaa, B., Sun, B., Gunter, C.A.,
Ganesh, S.B., Kishore, G.M., 2003. Expression of a
Streptomyces 3-hydroxysteroid oxidase gene in oilseeds for
converting phytosterols to phytostanols. Phytochemistry 62,
39–46.
Wen, J., Morrissey, P.A., Buckley, D.J., Sheehy, P.J., 1996.
Oxidative stability and a-tocopherol retention in turkey
burgers during refrigerated and frozen storage as influenced
by dietary a-tocopheryl acetate. British Poultry Science 37,
787–795.
Whiting, TC.M., Mutsvangwa, T., Walton, J.P., Cant, J.P.,
McBride, B.W., 2004. Effects of feeding either fresh alfalfa or
alfalfa silage on milk fatty acid content in Holstein dairy
cows. Animal Feed Science and Technology 113, 27–37.
Wing, S., Wolf, S., 2000. Intensive livestock operations, health,
and quality of life among eastern North Carolina residents.
Environmental Health Perspectives 108, 233–238.
Woolums, A.R., McAllister, T.A., Loneragan, G.H., 2001.
Interstitial pneumonia in feedlot cattle: noninfectious causes.
Compendium on Continuing Education for the Practicing
Veterinarian 23, S86–S93.
Wyatt, C.J., Carballido, S.P., Mendez, R.O., 1998. Alpha- and
gamma-tocopherol content of selected foods in the Mexican
diet: effect of cooking losses. Journal of Agricultural and
Food Chemistry 46, 4657–4661.
Yang, A., Brewster, M.J., Lanari, M.C., Tume, R.K., 2002.
Effect of vitamin E supplementation on a-tocopherol and b-carotene concentrations in tissues from pasture- and grain-fed
cattle. Meat Science 60, 35–40.




















