
Diabetes Mellitus and Endocrine, Nutritional and Metabolic ...

Oleic Acid Inhibits Endothelial ActivationA Direct Vascular Antiatherogenic Mechanism of a Nutritional
Component in the Mediterranean Diet
Maria Annunziata Carluccio, Marika Massaro, Cosima Bonfrate, Luisa Siculella, Michele Maffia,Giuseppe Nicolardi, Alessandro Distante, Carlo Storelli, Raffaele De Caterina
Abstract—Because oleic acid is implicated in the antiatherogenic effects attributed to the Mediterranean diet, weinvestigated whether this fatty acid can modulate endothelial activation, ie, the concerted expression of gene productsinvolved in leukocyte recruitment and early atherogenesis. We incubated sodium oleate with human umbilical veinendothelial cells for 0 to 72 hours, followed by coincubation of oleate with human recombinant tumor necrosis factor,interleukin (IL)-1a, IL-1b, IL-4, Escherichia colilipopolysaccharide (LPS), or phorbol 12-myristate 13-acetate for afurther 6 to 24 hours. The endothelial expression of vascular cell adhesion molecule-1 (VCAM-1), E-selectin, andintercellular adhesion molecule-1 was monitored by cell surface enzyme immunoassays or flow cytometry, andsteady-state levels of VCAM-1 mRNA were assessed by Northern blot analysis. At 10 to 100mmol/L for .24 hours,oleate inhibited the expression of all adhesion molecules tested. After a 72-hour incubation with oleate and a further16-hour incubation with oleate plus 1mg/mL LPS, VCAM-1 expression was reduced by.40% compared with control.Adhesion of monocytoid U937 cells to LPS-treated endothelial cells was reduced concomitantly. Oleate also produceda quantitatively similar reduction of VCAM-1 mRNA levels on Northern blot analysis and inhibited nuclear factor-kBactivation on electrophoretic mobility shift assays. Incubation of endothelial cells with oleate for 72 hours decreased therelative proportions of saturated (palmitic and stearic) acids in total cell lipids and increased the proportions of oleatein total cell lipids without significantly changing the relative proportions of polyunsaturated fatty acids. Although lesspotent than polyunsaturated fatty acids in inhibiting endothelial activation, oleic acid may contribute to the preventionof atherogenesis through selective displacement of saturated fatty acids in cell membrane phospholipids and aconsequent modulation of gene expression for molecules involved in monocyte recruitment.(Arterioscler Thromb VascBiol. 1999;19:220-228.)
Key Words: fatty acidsn endothelial activationn adhesion moleculesn atherogenesisn nuclear factor-kB
There is strong evidence for an association between aMediterranean-style diet, in which olive oil is the prin-
cipal source of fat, and protection from cardiovascular dis-ease.1–3 Although intervention studies with morbidity andmortality end points are still generally lacking, the associationhas been recognized as an established fact.4 There are amultitude of effects of components of the Mediterranean dietpotentially able to explain a causal association. Such dietslower total and LDL cholesterol compared with diets veryrich in saturated fatty acids,5–7 thus reducing a dominant riskfactor for the development of atherosclerosis. Additionalmechanisms may be favorable effects on other coronary riskfactors, such as hypertension,8,9 diabetes,10–12 and excessbody weight.13
Atherosclerosis is thought to be initiated at critical sites ofthe arterial vasculature by a process of monocyte adhesion tothe vessel wall, sustained by the occurrence of active func-
tional changes on the endothelial surface.14 Recent researchhas shown direct vascular effects of fatty acids, with possiblerelevance to atherogenesis. In particular, thev-3 fatty acidsdocosahexaenoate (DHA) and, to a lesser extent, eicosapen-taenoate, which have only minor effects on total and LDLcholesterol15 and yet also appear to be linked to protectionfrom atherosclerosis,16 may act by inhibiting early athero-genic events related to monocyte adhesion to endothelialcells.17–19This process occurs through inhibition of endothe-lial activation,17,18 ie, the concerted expression of cytokine-inducible endothelial leukocyte adhesion molecules and leu-kocyte chemoattractants affecting monocyte adhesion.Inhibition of a common signal-transduction pathway involv-ing the transcription factor nuclear factor-kB (NF-kB)20 wastherefore hypothesized.17 On the basis of a comparison of thebiological activity of several fatty acids, we have alsoreported preliminary evidence that an important structural
Received May 6, 1998; revision accepted June 3, 1998.From the CNR Institute of Clinical Physiology, Lecce (M.A.C, M. Massaro, C.B.) and Pisa (A.D., R.D.C.), and the Department of Biology, University
of Lecce (L.S., M. Maffia, G.N., C.S.), Lecce, Italy.Correspondence to Raffaele De Caterina, MD, PhD, Laboratory for Thrombosis and Vascular Research, CNR Institute of Clinical Physiology, Via Savi,
8, I-56126 Pisa, Italy. E-mail [email protected]© 1999 American Heart Association, Inc.
Arterioscler Thromb Vasc Biol.is available at http://www.atvbaha.org
220 by guest on January 18, 2016http://atvb.ahajournals.org/Downloaded from

feature for this effect appears to be the presence of doublebonds on the fatty acid backbone.21
We therefore anticipated that the monounsaturated fattyacid oleate, which is the predominant fatty acid in Mediter-ranean diets, also possesses some inhibitory activity onendothelial activation. In accordance, the present investiga-tion was undertaken aimed at (1) demonstrating the occur-rence, extent, and conditions for this effect; (2) showing itsrelationship to changes in fatty acid composition in endothe-lial cell lipids; and (3) offering mechanistic clues for itsbiological plausibility in explaining the beneficial effects ofdiets rich in oleate on atherogenesis.
MethodsEndothelial Cell CulturesHuman umbilical vein endothelial cells (HUVECs) were isolatedfrom umbilical cords harvested in the Obstetrics and GynecologyDivision of “Vito Fazzi” Hospital in Lecce, Italy. HUVECs wereharvested enzymatically with 1 mg/mL type IA collagenase asdescribed22 and maintained in medium 199 (Mascia Brunelli) con-taining 25 mmol/L HEPES, 50 U/mL heparin,23 50 mg/mL endothe-lial cell growth factor,24 2 mmol/L L-glutamine, antibiotics, and 15%FCS. Once grown to confluence, the cells were replated on 1.5%gelatin–coated flasks at 20 000 cells/cm2. HUVECs isolated by thesetechniques form a confluent monolayer of polygonal cells andexpress von Willebrand factor as determined by their content ofimmunoreactive protein. Cells were used at a time calibrated to allowthem reach confluence (60 000 to 80 000 cells/cm2) at the time ofstimulation.
ReagentsOleic acid (18:1 n-9cis), palmitic (16:0) and stearic (18:0) acids asnegative controls, and DHA (22:6 n-3 all-cis) as a positive controlfor inhibition of endothelial activation were obtained as.99% puresodium salts from Nu-Chek, dissolved in water at 10 mmol/L stockconcentration, divided into aliquots under an N2 stream, and stored at280°C for no longer than 6 months before the experiments. Forcontrols, the same fatty acids were also obtained from Sigma. At thetime of the experiments, fatty acids were further dissolved inserum-containing medium at the final desired concentration.
The following human recombinant cytokines were used at thereported final concentrations: interleukin (IL)-1a (Hoffmann–LaRoche, at 1 to 10 ng/mL), IL-1b (1 to 10 ng/mL), tumor necrosisfactor (TNF)-a (1 to 10 ng/mL), and IL-4 (50 to 100 ng/mL) (thelatter 3 all from Genzyme).Escherichia coli lipopolysaccharide(LPS, 10 to 10 000 ng/mL) was purchased from Sigma. Phorbol12-myristate 13-acetate (PMA, 10 nmol/L, equivalent to 6.3 ng/mL)as a stimulus for endothelial activation that bypasses membranereceptors was purchased from Sigma as were all other reagents,when not otherwise specified.
Detection of Cell Surface MoleculesAssay of cell surface molecules was carried out by either cell surfaceenzyme immunoassays (EIAs) or flow cytometry by using mouseanti-human monoclonal antibodies against vascular cell adhesionmolecule-1 (VCAM-1; Ab E1/6), E-selectin (Ab H18/7), intercellu-lar adhesion molecule-1 (ICAM-1; Ab HU5/3), major histocompat-ibility complex class I (MHC-I; W6/3225), or the monoclonalantibody E1/1, recognizing a constitutive and non–cytokine-inducible endothelial cell antigen.26 These last 2 endothelial surfacemolecules share a high level of basal constitutive expression,although MHC-I antigen, at variance from E1/1 antigen, shows somedegree of inducibility by cytokines. EIAs were carried out byincubating monolayers first with saturating concentrations of spe-cific monoclonal antibodies against the target molecule, then withbiotinylated goat anti-mouse IgG (Amersham Life Sciences, Amer-sham), and finally with streptavidin–alkaline phosphatase (Amer-sham). Layers were washed 3 times with PBS between eachincubation step, and the integrity of the monolayer was monitored by
phase-contrast microscopy. The surface expression of each proteinwas quantified spectrophotometrically by reading the optical densityof the wells at a wavelength of 450 nm 20 minutes after addition ofthe chromogenic substrate (3,39,5,59-tetramethylbenzidine) and stop-ping the reaction with 1N H2SO4.
For flow-cytometric analysis, the surface expression of adhesionmolecules was assessed by incubating HUVECs, suspended inHanks’ buffered saline solution with 3 mmol/L EDTA, with thespecific primary antibody for 30 minutes at 4°C and subsequentlywith goat anti-mouse F(ab9)2 from IgG (heavy and light) labeled withfluorescein isothiocyanate (Immunotech) at 4°C. After washing andfixation in 1% formaldehyde, the cell suspension was passed througha Becton-Dickinson FACScan analyzer. Results were plotted asintensity of fluorescence (arbitrary units, on a logarithmic scale asthe abscissa) versus cell number (on the ordinate; the total number ofcells counted for each condition was 104).
Assessment of Cell Number and ViabilityCell number was assessed by direct cell counting of adherent cells,after trypsin detachment, in a Neubauer hemocytometer (VWRScientific) and staining with trypan blue. The percentage of cellsexcluding trypan blue was taken as a measure of cell viability.
Assessment of Total Protein SynthesisHUVECs were cultured in 96-well plates in the presence or absenceof fatty acids for 0 to 72 hours and then for up to 24 hours in thepresence or absence of LPS or cytokines. After this time, the totalcell-associated protein content was assessed by the Bradford meth-od.27 In parallel experiments, the dye reaction was correlated linearly(R50.98) with cell number per well, which in turn was determinedby hemocytometric counting of trypsinized cells in parallel cultures.
Monocytoid Cell Adhesion AssaysMonocytoid U937 cells were obtained through American TypeCulture Collection (Manassas, Va) and grown in RPMI 1640medium (Gibco BRL, Life Technologies Italia srl) containing 10%FCS. U937 cells were concentrated by centrifugation to 106
cells/mL. For the adhesion assays, HUVECs were grown to conflu-ence in 6-well tissue culture plates, after which LPS (10 to 10 000ng/mL) or IL-1a (10 ng/mL) was added for an additional 16 hours toinduce the expression of VCAM-1 in the presence or absence ofoleate (10 to 100mmol/L). For controls, some monolayers weretreated with a blocking mouse anti-human monoclonal antibody(E1/6) against VCAM-1. The adhesion assay was performed byadding 1 mL of the concentrated U937 cell suspension to eachmonolayer under rotating conditions (63 rpm) at 21°C.28,29 After 10minutes, nonadhering cells were removed by gentle washing withmedium 199, and the monolayers were fixed with 1% paraformal-dehyde. The number of adherent cells was determined by counting 6different fields by using an ocular grid and a 203 objective(0.16 mm2/field). Fields for counting adherent leukocytes wererandomly located at a half-radius distance from the center of themonolayers.
Isolation of RNA and Northern Blot AnalysisTotal cellular RNA was isolated by a single extraction using an acidguanidinium thiocyanate-phenol-chloroform method.30 RNA con-centration and quality were determined from the absorbance at 260nm and the absorbance ratio 260 nm/280 nm, respectively. RNAquality was confirmed by gel electrophoresis before Northern blotanalysis. For this analysis, 20mg of cellular RNA was applied toeach lane, separated on a 1% agarose-formaldehyde gel, transferredto a nylon membrane (Amersham Hybond-N), and immobilized byshort-wave UV illumination. The membranes were prehybridized forat least 2 hours before hybridization with a32P-labeled DNA probefor VCAM-1 (labeled by random hexanucleotide priming; Pharma-cia) to specific activities.108 counts per minute permg DNA andautoradiographed. 18S and 28S rRNA fluorescence intensity of theethidium bromide–stained membranes served as a control of equalloading of different lanes in the Northern blot analyses.
Quantification of densities of autoradiographic bands for Northernblot hybridization was performed with the aid of the National
Carluccio et al February 1999 221
by guest on January 18, 2016http://atvb.ahajournals.org/Downloaded from

Institutes of Health (Bethesda, Md) Image 1.60 software on aMacIntosh Quadra 800 computer (Apple).
Preparation of Nuclear Extracts andElectrophoretic Mobility Shift Assay (EMSA)Confluent HUVECs were pretreated for 0 to 72 hours with50 mmol/L and 100mmol/L sodium oleate and then exposed to LPS(1 mg/mL) for 1 hour. Cells were scraped mechanically and collectedinto chilled microfuge tubes. Nuclear and cytosolic extracts wereprepared according to Dignam et al,31 with the additional step ofwashing the nuclear pellets in a low-salt buffer before high-saltextraction of nuclear proteins to remove any residual cytosoliccontamination. Aliquots were then assayed for protein concentra-tion.27 To minimize proteolysis, all buffers included 1.0 mmol/LPMSF, and extracts were stored at280°C until analysis. Thewild-type VCAM-1 promoter oligonucleotide was synthesized toencompass the 2 NF-kB consensus repeats (underlined) described atcoordinates277 and 263 of the human VCAM-1 promoter,32
according to the sequence: 59-CTGCCCTGGGTTTCCCCTTGA-AGGGATTTCCCTCC-39, together with its complementary oli-gonucleotide, 59-GAGGCGGAGGGAAATCCCTTCAAGGGG-AAACCCAGG-39.
The mutant VCAM-1 promoter oligonucleotide was identical tothe wild-type VCAM-1 promoter oligonucleotide except for 4nucleotide mutations (boldface) in each of the 2 NF-kB binding sites:59-CTGCCCTGAGTCACGCCTTGAAGAGACATCACTCC-39,together with the corresponding complementary oligonucleotide,59-GAGGCGGAGGGAAATCCCTTCAAGGGGAAACCCAGG-39. All oligonucleotides were synthesized by GIBCO-BRL LifeTechnologies, and double-stranded oligonucleotides were radiola-beled by Klenow filling-in with 50 mCi of [a-32P]dCTP (3000Ci/mmol) and unlabeled dATP, dGTP, and dTTP (each at 100mol/L). Unincorporated nucleotides were removed by column chro-matography over a Sephadex G-50 column. The DNA bindingreaction was performed at 30°C for 15 minutes in a volume of 20mLcontaining 8mg of nuclear extract, 225mg/mL BSA, 10 000 cpm of32P-labeled probe, 2mg poly(dI-dC) (Boehringer Mannheim Italia),15mL of binding buffer (12 mmol/L HEPES, pH 7.9, 4 mmol/L Tris,60 mmol/L KCl, 1 mol/L EDTA, 12% glycerol, 1 mol/L DTT, and1 mol/L PMSF) with or without a 100-fold molar excess of coldcompetitor. Samples were subjected to electrophoresis on native 5%0.53 Tris-borate-polyacrylamide gels.
Fatty Acid AnalysisRelative amounts of the individual chemical species of the mostabundant fatty acids present in cell lipids were quantified byintegration of peaks obtained by gas-liquid chromatographic separa-tion of fatty acids from HUVEC monolayers. After chloroform-methanol (2:1, vol/vol) extraction, total cell lipids were subjected toacidic trans-methylation,33 and fatty acid methyl esters were ana-lyzed by flame-ionization capillary gas-liquid chromatography on aHewlett-Packard GC system, HP 6890 series, using a SupelcoOmegawax 250 column (30 m30.25-mm ID, 0.25-mm film thick-
ness). The oven temperature was programmed at 150°C for 15minutes and then increased to 250°C at 4°C/min, then to 250°C for35 minutes. The H2 and air flow were 30 and 330 mL/min,respectively; the carrier (helium) pressure was 17 psi. Individualpeaks were integrated automatically with a Hewlett-Packard integra-tor and identified by comparison with retention times of fatty acidstandards.
Experimental DesignsCultured HUVECs were preincubated with oleate or other fatty acidsfor 0 to 72 hours, followed by stimulation with cytokines (IL-1a,IL-1b, TNF, IL-4), PMA, or LPS for an additional 16 to 24 hours. Atthe end of this period, the expression of endothelial surface mole-cules (cell surface EIA and/or flow cytometry) of VCAM-1 mRNA(Northern blot analysis) or of parameters of cell viability (morphol-ogy, number, trypan blue exclusion, total protein synthesis) wereassessed, or the adhesion assays were performed. Each experimentwas performed at least in triplicate. In EIAs, a minimum number of8 repeats was run in each experimental condition.
StatisticsMultiple comparisons were performed by 1-way ANOVA andindividual differences tested by Fisher’s protected least significancedifference test after demonstration of significant intergroup differ-ences by ANOVA. Two-group comparisons were performed byunpaired Student’st test. Comparisons of distribution of fluores-cence intensities for flow cytometry were performed withKolmogorov-Smirnov’s statistics with the aid of the Becton-Dickinson statistical software package.
ResultsToxicity Profile of Oleate Effects on HUVECsThe absence of any detectable toxic effect of sodium oleate atthe concentrations and times used on cell number or param-eters of cell viability is shown in Table 1. Sodium oleate didnot reduce cell counts and did not affect either trypan blueexclusion or the estimate of total cell-associated protein forconcentrations up to 100mmol/L and for preincubation timesup to 72 hours before addition of the stimulating cytokines,ruling out the possibility that the effects described belowcould be due to a general inhibition of cell metabolism or anygeneral “toxic” effect. Also, no change in MHC-I or E1/1antigen expression was detected under any circumstance upto 100mmol/L oleate concentrations (not shown).
Oleate Reduces LPS-Stimulated VCAM-1Expression in HUVECsDespite any general effect on total protein synthesis, prein-cubation of HUVECs with oleate for 72 hours and then for a
TABLE 1. Control of Oleate Toxicity on HUVECs
Control
Oleate, mmol/L
10 50 100
Cell counts, cells/1000 per cm2
0 Hours 3062 3062 3062 3062
24 Hours 2863 2764 3068 3164
48 Hours 4267 4261 44610 3765
72 Hours 6864 6168 7169 6668
Viability by trypan blue exclusion .95% .95% .95% .95%
Total cellular proteins, ng/1000 cells
48 Hours 590690 6606100 743680 680670
72 Hours 640690 7806100 6206100 670690
Values are mean6SD.
222 Oleic Acid Inhibits Endothelial Activation
by guest on January 18, 2016http://atvb.ahajournals.org/Downloaded from
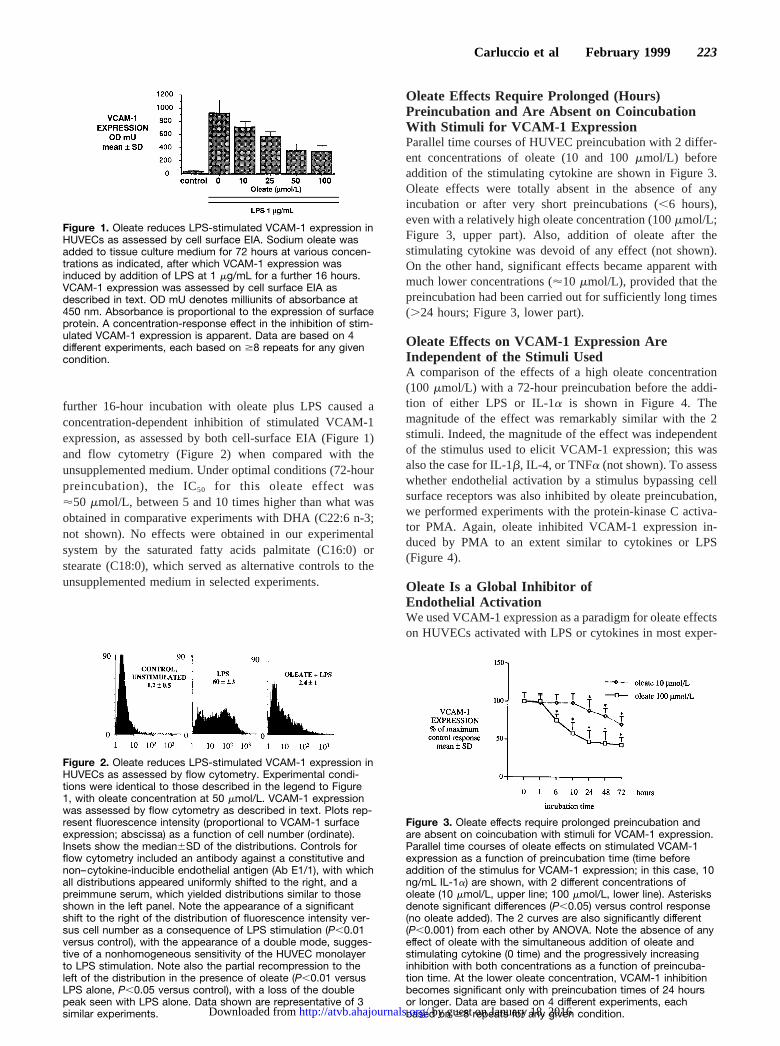
further 16-hour incubation with oleate plus LPS caused aconcentration-dependent inhibition of stimulated VCAM-1expression, as assessed by both cell-surface EIA (Figure 1)and flow cytometry (Figure 2) when compared with theunsupplemented medium. Under optimal conditions (72-hourpreincubation), the IC50 for this oleate effect was'50 mmol/L, between 5 and 10 times higher than what wasobtained in comparative experiments with DHA (C22:6 n-3;not shown). No effects were obtained in our experimentalsystem by the saturated fatty acids palmitate (C16:0) orstearate (C18:0), which served as alternative controls to theunsupplemented medium in selected experiments.
Oleate Effects Require Prolonged (Hours)Preincubation and Are Absent on CoincubationWith Stimuli for VCAM-1 ExpressionParallel time courses of HUVEC preincubation with 2 differ-ent concentrations of oleate (10 and 100mmol/L) beforeaddition of the stimulating cytokine are shown in Figure 3.Oleate effects were totally absent in the absence of anyincubation or after very short preincubations (,6 hours),even with a relatively high oleate concentration (100mmol/L;Figure 3, upper part). Also, addition of oleate after thestimulating cytokine was devoid of any effect (not shown).On the other hand, significant effects became apparent withmuch lower concentrations ('10mmol/L), provided that thepreincubation had been carried out for sufficiently long times(.24 hours; Figure 3, lower part).
Oleate Effects on VCAM-1 Expression AreIndependent of the Stimuli UsedA comparison of the effects of a high oleate concentration(100 mmol/L) with a 72-hour preincubation before the addi-tion of either LPS or IL-1a is shown in Figure 4. Themagnitude of the effect was remarkably similar with the 2stimuli. Indeed, the magnitude of the effect was independentof the stimulus used to elicit VCAM-1 expression; this wasalso the case for IL-1b, IL-4, or TNFa (not shown). To assesswhether endothelial activation by a stimulus bypassing cellsurface receptors was also inhibited by oleate preincubation,we performed experiments with the protein-kinase C activa-tor PMA. Again, oleate inhibited VCAM-1 expression in-duced by PMA to an extent similar to cytokines or LPS(Figure 4).
Oleate Is a Global Inhibitor ofEndothelial ActivationWe used VCAM-1 expression as a paradigm for oleate effectson HUVECs activated with LPS or cytokines in most exper-
Figure 1. Oleate reduces LPS-stimulated VCAM-1 expression inHUVECs as assessed by cell surface EIA. Sodium oleate wasadded to tissue culture medium for 72 hours at various concen-trations as indicated, after which VCAM-1 expression wasinduced by addition of LPS at 1 mg/mL for a further 16 hours.VCAM-1 expression was assessed by cell surface EIA asdescribed in text. OD mU denotes milliunits of absorbance at450 nm. Absorbance is proportional to the expression of surfaceprotein. A concentration-response effect in the inhibition of stim-ulated VCAM-1 expression is apparent. Data are based on 4different experiments, each based on $8 repeats for any givencondition.
Figure 2. Oleate reduces LPS-stimulated VCAM-1 expression inHUVECs as assessed by flow cytometry. Experimental condi-tions were identical to those described in the legend to Figure1, with oleate concentration at 50 mmol/L. VCAM-1 expressionwas assessed by flow cytometry as described in text. Plots rep-resent fluorescence intensity (proportional to VCAM-1 surfaceexpression; abscissa) as a function of cell number (ordinate).Insets show the median6SD of the distributions. Controls forflow cytometry included an antibody against a constitutive andnon–cytokine-inducible endothelial antigen (Ab E1/1), with whichall distributions appeared uniformly shifted to the right, and apreimmune serum, which yielded distributions similar to thoseshown in the left panel. Note the appearance of a significantshift to the right of the distribution of fluorescence intensity ver-sus cell number as a consequence of LPS stimulation (P,0.01versus control), with the appearance of a double mode, sugges-tive of a nonhomogeneous sensitivity of the HUVEC monolayerto LPS stimulation. Note also the partial recompression to theleft of the distribution in the presence of oleate (P,0.01 versusLPS alone, P,0.05 versus control), with a loss of the doublepeak seen with LPS alone. Data shown are representative of 3similar experiments.
Figure 3. Oleate effects require prolonged preincubation andare absent on coincubation with stimuli for VCAM-1 expression.Parallel time courses of oleate effects on stimulated VCAM-1expression as a function of preincubation time (time beforeaddition of the stimulus for VCAM-1 expression; in this case, 10ng/mL IL-1a) are shown, with 2 different concentrations ofoleate (10 mmol/L, upper line; 100 mmol/L, lower line). Asterisksdenote significant differences (P,0.05) versus control response(no oleate added). The 2 curves are also significantly different(P,0.001) from each other by ANOVA. Note the absence of anyeffect of oleate with the simultaneous addition of oleate andstimulating cytokine (0 time) and the progressively increasinginhibition with both concentrations as a function of preincuba-tion time. At the lower oleate concentration, VCAM-1 inhibitionbecomes significant only with preincubation times of 24 hoursor longer. Data are based on 4 different experiments, eachbased on $8 repeats for any given condition.
Carluccio et al February 1999 223
by guest on January 18, 2016http://atvb.ahajournals.org/Downloaded from
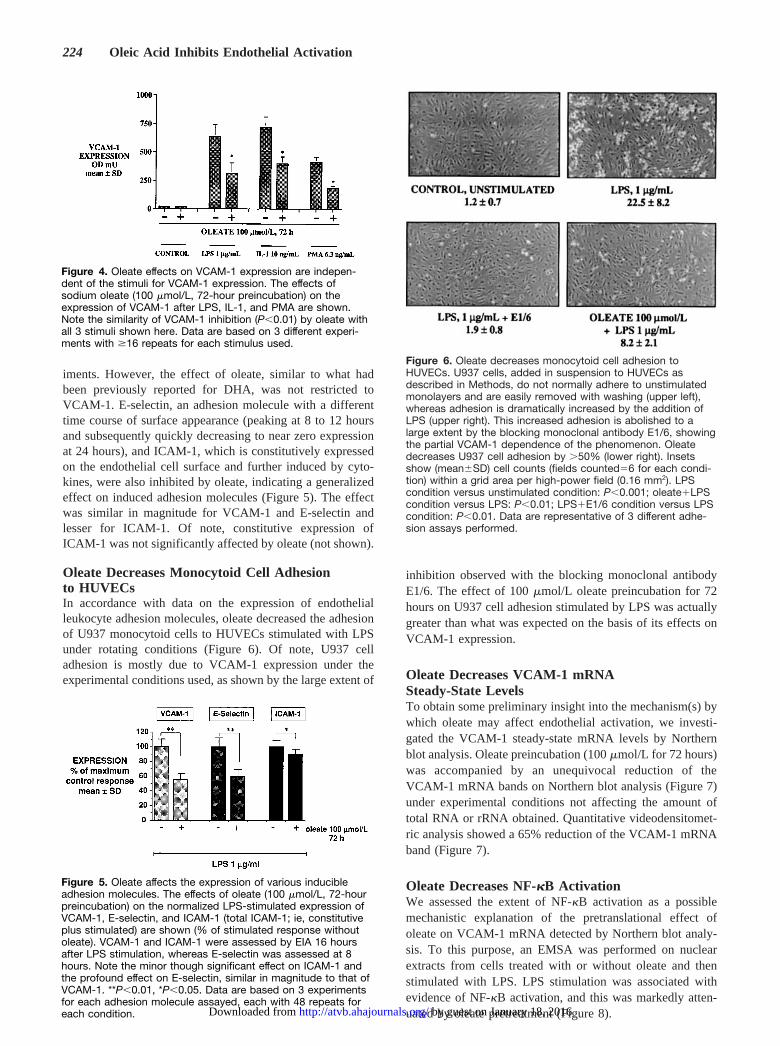
iments. However, the effect of oleate, similar to what hadbeen previously reported for DHA, was not restricted toVCAM-1. E-selectin, an adhesion molecule with a differenttime course of surface appearance (peaking at 8 to 12 hoursand subsequently quickly decreasing to near zero expressionat 24 hours), and ICAM-1, which is constitutively expressedon the endothelial cell surface and further induced by cyto-kines, were also inhibited by oleate, indicating a generalizedeffect on induced adhesion molecules (Figure 5). The effectwas similar in magnitude for VCAM-1 and E-selectin andlesser for ICAM-1. Of note, constitutive expression ofICAM-1 was not significantly affected by oleate (not shown).
Oleate Decreases Monocytoid Cell Adhesionto HUVECsIn accordance with data on the expression of endothelialleukocyte adhesion molecules, oleate decreased the adhesionof U937 monocytoid cells to HUVECs stimulated with LPSunder rotating conditions (Figure 6). Of note, U937 celladhesion is mostly due to VCAM-1 expression under theexperimental conditions used, as shown by the large extent of
inhibition observed with the blocking monoclonal antibodyE1/6. The effect of 100mmol/L oleate preincubation for 72hours on U937 cell adhesion stimulated by LPS was actuallygreater than what was expected on the basis of its effects onVCAM-1 expression.
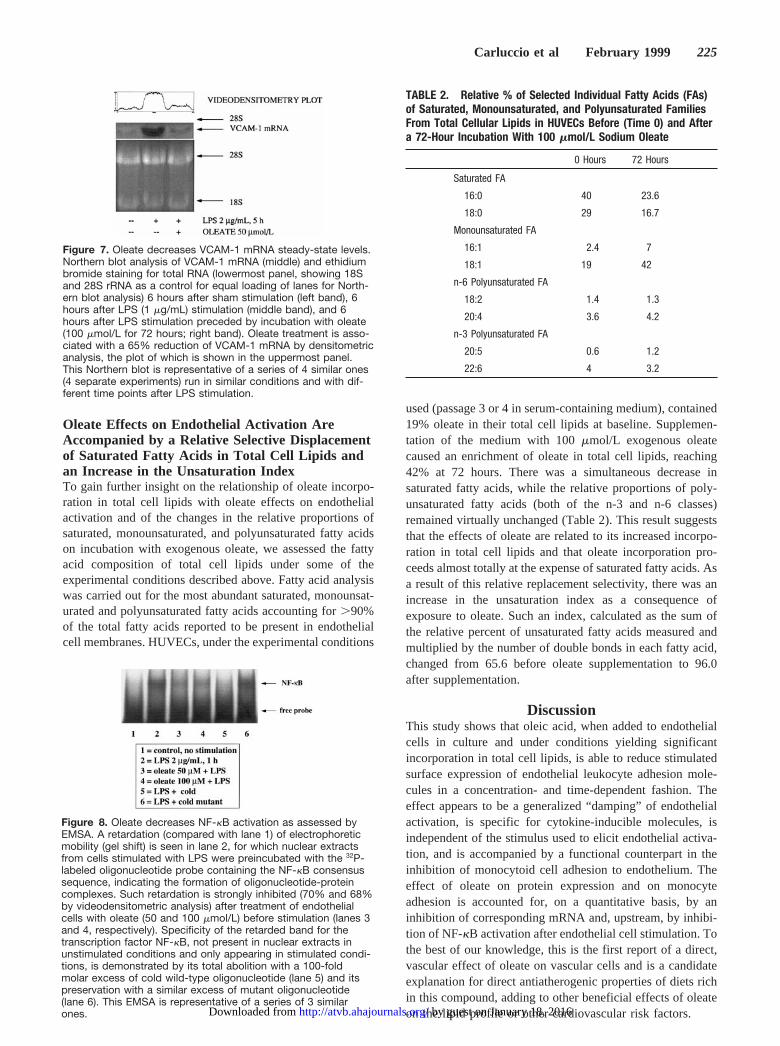
Oleate Decreases VCAM-1 mRNASteady-State LevelsTo obtain some preliminary insight into the mechanism(s) bywhich oleate may affect endothelial activation, we investi-gated the VCAM-1 steady-state mRNA levels by Northernblot analysis. Oleate preincubation (100mmol/L for 72 hours)was accompanied by an unequivocal reduction of theVCAM-1 mRNA bands on Northern blot analysis (Figure 7)under experimental conditions not affecting the amount oftotal RNA or rRNA obtained. Quantitative videodensitomet-ric analysis showed a 65% reduction of the VCAM-1 mRNAband (Figure 7).
Oleate Decreases NF-kB ActivationWe assessed the extent of NF-kB activation as a possiblemechanistic explanation of the pretranslational effect ofoleate on VCAM-1 mRNA detected by Northern blot analy-sis. To this purpose, an EMSA was performed on nuclearextracts from cells treated with or without oleate and thenstimulated with LPS. LPS stimulation was associated withevidence of NF-kB activation, and this was markedly atten-uated by oleate pretreatment (Figure 8).
Figure 4. Oleate effects on VCAM-1 expression are indepen-dent of the stimuli for VCAM-1 expression. The effects ofsodium oleate (100 mmol/L, 72-hour preincubation) on theexpression of VCAM-1 after LPS, IL-1, and PMA are shown.Note the similarity of VCAM-1 inhibition (P,0.01) by oleate withall 3 stimuli shown here. Data are based on 3 different experi-ments with $16 repeats for each stimulus used.
Figure 5. Oleate affects the expression of various inducibleadhesion molecules. The effects of oleate (100 mmol/L, 72-hourpreincubation) on the normalized LPS-stimulated expression ofVCAM-1, E-selectin, and ICAM-1 (total ICAM-1; ie, constitutiveplus stimulated) are shown (% of stimulated response withoutoleate). VCAM-1 and ICAM-1 were assessed by EIA 16 hoursafter LPS stimulation, whereas E-selectin was assessed at 8hours. Note the minor though significant effect on ICAM-1 andthe profound effect on E-selectin, similar in magnitude to that ofVCAM-1. **P,0.01, *P,0.05. Data are based on 3 experimentsfor each adhesion molecule assayed, each with 48 repeats foreach condition.
Figure 6. Oleate decreases monocytoid cell adhesion toHUVECs. U937 cells, added in suspension to HUVECs asdescribed in Methods, do not normally adhere to unstimulatedmonolayers and are easily removed with washing (upper left),whereas adhesion is dramatically increased by the addition ofLPS (upper right). This increased adhesion is abolished to alarge extent by the blocking monoclonal antibody E1/6, showingthe partial VCAM-1 dependence of the phenomenon. Oleatedecreases U937 cell adhesion by .50% (lower right). Insetsshow (mean6SD) cell counts (fields counted56 for each condi-tion) within a grid area per high-power field (0.16 mm2). LPScondition versus unstimulated condition: P,0.001; oleate1LPScondition versus LPS: P,0.01; LPS1E1/6 condition versus LPScondition: P,0.01. Data are representative of 3 different adhe-sion assays performed.
224 Oleic Acid Inhibits Endothelial Activation
by guest on January 18, 2016http://atvb.ahajournals.org/Downloaded from
Oleate Effects on Endothelial Activation AreAccompanied by a Relative Selective Displacementof Saturated Fatty Acids in Total Cell Lipids andan Increase in the Unsaturation IndexTo gain further insight on the relationship of oleate incorpo-ration in total cell lipids with oleate effects on endothelialactivation and of the changes in the relative proportions ofsaturated, monounsaturated, and polyunsaturated fatty acidson incubation with exogenous oleate, we assessed the fattyacid composition of total cell lipids under some of theexperimental conditions described above. Fatty acid analysiswas carried out for the most abundant saturated, monounsat-urated and polyunsaturated fatty acids accounting for.90%of the total fatty acids reported to be present in endothelialcell membranes. HUVECs, under the experimental conditions
used (passage 3 or 4 in serum-containing medium), contained19% oleate in their total cell lipids at baseline. Supplemen-tation of the medium with 100mmol/L exogenous oleatecaused an enrichment of oleate in total cell lipids, reaching42% at 72 hours. There was a simultaneous decrease insaturated fatty acids, while the relative proportions of poly-unsaturated fatty acids (both of the n-3 and n-6 classes)remained virtually unchanged (Table 2). This result suggeststhat the effects of oleate are related to its increased incorpo-ration in total cell lipids and that oleate incorporation pro-ceeds almost totally at the expense of saturated fatty acids. Asa result of this relative replacement selectivity, there was anincrease in the unsaturation index as a consequence ofexposure to oleate. Such an index, calculated as the sum ofthe relative percent of unsaturated fatty acids measured andmultiplied by the number of double bonds in each fatty acid,changed from 65.6 before oleate supplementation to 96.0after supplementation.
DiscussionThis study shows that oleic acid, when added to endothelialcells in culture and under conditions yielding significantincorporation in total cell lipids, is able to reduce stimulatedsurface expression of endothelial leukocyte adhesion mole-cules in a concentration- and time-dependent fashion. Theeffect appears to be a generalized “damping” of endothelialactivation, is specific for cytokine-inducible molecules, isindependent of the stimulus used to elicit endothelial activa-tion, and is accompanied by a functional counterpart in theinhibition of monocytoid cell adhesion to endothelium. Theeffect of oleate on protein expression and on monocyteadhesion is accounted for, on a quantitative basis, by aninhibition of corresponding mRNA and, upstream, by inhibi-tion of NF-kB activation after endothelial cell stimulation. Tothe best of our knowledge, this is the first report of a direct,vascular effect of oleate on vascular cells and is a candidateexplanation for direct antiatherogenic properties of diets richin this compound, adding to other beneficial effects of oleateon the lipid profile or other cardiovascular risk factors.
Figure 7. Oleate decreases VCAM-1 mRNA steady-state levels.Northern blot analysis of VCAM-1 mRNA (middle) and ethidiumbromide staining for total RNA (lowermost panel, showing 18Sand 28S rRNA as a control for equal loading of lanes for North-ern blot analysis) 6 hours after sham stimulation (left band), 6hours after LPS (1 mg/mL) stimulation (middle band), and 6hours after LPS stimulation preceded by incubation with oleate(100 mmol/L for 72 hours; right band). Oleate treatment is asso-ciated with a 65% reduction of VCAM-1 mRNA by densitometricanalysis, the plot of which is shown in the uppermost panel.This Northern blot is representative of a series of 4 similar ones(4 separate experiments) run in similar conditions and with dif-ferent time points after LPS stimulation.
Figure 8. Oleate decreases NF-kB activation as assessed byEMSA. A retardation (compared with lane 1) of electrophoreticmobility (gel shift) is seen in lane 2, for which nuclear extractsfrom cells stimulated with LPS were preincubated with the 32P-labeled oligonucleotide probe containing the NF-kB consensussequence, indicating the formation of oligonucleotide-proteincomplexes. Such retardation is strongly inhibited (70% and 68%by videodensitometric analysis) after treatment of endothelialcells with oleate (50 and 100 mmol/L) before stimulation (lanes 3and 4, respectively). Specificity of the retarded band for thetranscription factor NF-kB, not present in nuclear extracts inunstimulated conditions and only appearing in stimulated condi-tions, is demonstrated by its total abolition with a 100-foldmolar excess of cold wild-type oligonucleotide (lane 5) and itspreservation with a similar excess of mutant oligonucleotide(lane 6). This EMSA is representative of a series of 3 similarones.
TABLE 2. Relative % of Selected Individual Fatty Acids (FAs)of Saturated, Monounsaturated, and Polyunsaturated FamiliesFrom Total Cellular Lipids in HUVECs Before (Time 0) and Aftera 72-Hour Incubation With 100 mmol/L Sodium Oleate
0 Hours 72 Hours
Saturated FA
16:0 40 23.6
18:0 29 16.7
Monounsaturated FA
16:1 2.4 7
18:1 19 42
n-6 Polyunsaturated FA
18:2 1.4 1.3
20:4 3.6 4.2
n-3 Polyunsaturated FA
20:5 0.6 1.2
22:6 4 3.2
Carluccio et al February 1999 225
by guest on January 18, 2016http://atvb.ahajournals.org/Downloaded from

Oleic acid is a predominant component of olive oil and, inturn, of the Mediterranean diet,1–3 which is currently thoughtto exert atheroprotective effects, mostly through a lowering oftotal and LDL cholesterol5–7 or through a decrease in coro-nary risk factors such as hypertension,8,9 diabetes,10–12 orobesity.13 The data presented here make the alternative oradditional explanation plausible that at least some of oleate’sbeneficial effects are exerted through inhibition of the veryearly phenomena in atherogenesis, leading to monocyterecruitment and the development of the fatty streak. Fattyacids in general have recently emerged as possible physio-logical regulators of the endothelial responsiveness to acti-vating stimuli. In particular, the highly unsaturated n-3 fattyacid DHA, abundant in fish and fish-derived products, hasbeen shown by us17,18 as well as others19 to be a potentinhibitor of endothelial responses to cytokines, thus decreas-ing the expression of VCAM-1, E-selectin, and ICAM-1, aswell as of the soluble products of endothelial activation, suchas IL-6 and IL-8. Functional consequences of these biochem-ical effects have also been shown.17,18 In an effort to under-stand the structural component of the fatty acid structurerequired for these effects, we have recently compared severalfatty acids, differing in chain length and the number, position,and configuration of the double bonds. While saturated fattyacids were always devoid of any ability to interfere withendothelial activation, the presence of double bonds appearedto be the minimum essential and sufficient requirement forthe inhibition of endothelial activation, with no apparentrelevance of the position (n-3 versus n-6) and even of theconfiguration (cisversus trans) of the double bond. Theoriginal fatty acid investigated, DHA, appeared to be the mostpotent because of the high number of double bonds, thehighest able to be accommodated in the fatty acid chain ofthat length.21 This finding led to the hypothesis that themonounsaturated fatty acid oleate, in which the single doublebond is located in the n-9 position but which is much moreabundant nutritionally than many polyunsaturates, could alsopossess some of these properties. We therefore investigatedthe occurrence, extent, and conditions for this effect in detailand its relationship to changes in fatty acid composition inendothelial cell lipids and sought to reach conclusions on itsbiological plausibility in explaining its benefits on atherogen-esis reduction.
In line with our predictions, oleate exerted a concentration-dependent inhibitory effect on stimulated VCAM-1 expres-sion. The magnitude of inhibition, an ability to reduceVCAM-1 expression to'50% of the control response,depended on both the concentration and the duration ofincubation. At any given concentration, a plateau of responseto oleate was reached by prolonging the incubation time to 72hours before cytokine stimulation, whereas no inhibitionoccurred by coincubation or administration of oleate afterLPS or cytokines. This kinetics closely resembles that ofDHA.17 Also, similar to what had been previously shown forDHA, the effect is totally independent of the stimulus used:cytokines acting on totally different receptors, such as IL-1 (aor b), TNF-a, IL-4, or LPS, and inducing different responses,both in terms of selectivity (IL-4 selectively inducingVCAM-1 but not E-selectin on endothelial cells) and relativepotency (maximum response to IL-4 being, for example,,1⁄2that induced by a maximal concentration of LPS), had their
responses inhibited to the same extent by any given concen-tration of oleate administered with the same preincubationtime. Furthermore, oleate also inhibited VCAM-1 expressioninduced by PMA, an agent able to induce endothelial activa-tion that bypasses cell surface receptors and likely actingthrough direct activation of protein kinase C, possibly evenbypassing NF-kB activation.34 We also confirm, in line withwhat we reported for other more unsaturated fatty acids,17 butcontrary to what has been reported by others for DHA,19 thatthe effect of oleate is not restricted to VCAM-1 expressionbut also pertains to other inducible adhesion molecules,suggesting a mechanism on a common pathogenetic trans-ducer of endothelial activation. Together with the data on theinhibition of VCAM-1 mRNA accumulation obtained hereinby Northern blot analysis and the notion that the regulation ofadhesion molecule expression appears to be to largely tran-scriptional,20 we postulate transcriptional interference byoleate, possibly mediated by inhibition of the activation of thetranscription factor NF-kB. The same concentrations ofoleate that exerted effects on adhesion molecule expressionwere indeed also able to decrease the nuclear translocationand binding of NF-kB to its consensus sequences, showingfor the first time an effect of a monounsaturated fatty acid ona common pathway of cell activation by a variety of cytokineand noncytokine stimuli. Thus, the effect of oleate on NF-kBis at least a partial explanation for its pleiotropy, ie, effects onmultiple adhesion molecules and, possibly, on other media-tors of endothelial activation implicated in monocyte andmonocytoid cell adhesion (eg, monocyte chemoattractantprotein-135). This pleiotropy would also account for thenoteworthy inhibition of monocytoid cell adhesion, which weherein report being actually larger than expected on the basisof VCAM-1 inhibition data alone. Our results do not rule outadditional effects of oleate on adhesion molecule expressionindependent of NF-kB, as actually suspected on the basis ofproven effects on PMA- or IL-4–induced activation. Sucheffects may be modifications of specific receptor-agonistinteractions or on protein kinase C–dependent signal trans-duction pathways that have not been investigated further inthe current study. However, results on NF-kB activation mayaccount for most of the effects seen with the activatingcytokines used.
The biological relevance of the observations herein re-ported for oleate is mostly supported by 2 lines of reasoning.The first relates to concentrations of fatty acids required forthe reported effects. Although fatty acids mostly circulate inplasma while bound to plasma proteins, it is likely that asmall portion of free fatty acids in equilibrium with the boundfatty acid pool is responsible for most biological effects anddrives their incorporation into membrane phospholipids byenzymatic esterification mechanisms.36 Plasma concentra-tions of oleate under conditions of high olive oil consumptionare likely to be fully in the range of concentrations exertingbiological effects in our system, likely between 10 and100 mmol/L. We currently report biological effects demon-strable with oleate concentrations as low as 10mmol/L,provided that incubation is performed for times long enoughto ensure adequate incorporation. Of note, our biologicalsystem also contained binding proteins (albumin amongothers) because of the relatively high content of serum used(15% vol/vol). Therefore, the same binding to plasma pro-
226 Oleic Acid Inhibits Endothelial Activation
by guest on January 18, 2016http://atvb.ahajournals.org/Downloaded from

teins that occurs in vivo after intake of olive oil–containingmeals and the subsequent increase in the concentrations offree oleate in the free fatty acid plasma pool are likely tooccur and be reproduced by our in vitro experimental system.The second line of consideration, pointing to the plausibilityof the reported effect as an explanation for the benefits ofoleate on atherogenesis, is based on the relative selectivity ofoleate incorporation in total cellular lipid pools. On prolongedincubation, oleate selectively displaced saturated fatty acids,thereby decreasing their relative proportion to an extentvirtually equal to the magnitude of oleate incorporation.Contrary to this result, the polyunsaturated fatty acid pool,taken as the sum of the most abundant representatives of then-3 and n-6 fatty acid classes, was almost totally unaffectedby oleate. As an overall consequence of the selective increasein oleate concentration of the medium, the unsaturation index,calculated from the relative proportions of all main unsatur-ated and saturated fatty acids in the lipid pool and the numberof double bonds present in each fatty acid, increased. Thiscontrasts with what we previously reported for the incorpo-ration of the n-3 polyunsaturated fatty acid DHA, whichdisplaces both saturated and other polyunsaturated fatty acidsfrom membrane lipids.17 The nonrandom distribution ofoleate during its incorporation suggests the existence ofmetabolic mechanisms governing specific fatty acid incorpo-ration and allowing precise directioning of selected fatty acidspecies into relatively segregated pools. Implications of thisselectivity would be that the increase in oleate content in thediet would be reflected in a relatively selective replacementof saturated fatty acids, which are biologically “inert” withrespect to endothelial activation, with a biologically activeunsaturated fatty acid component. Also, this would imply thatcoadministration of oleate and of higher unsaturated fattyacid species could be additive and not mutually contrastingwith respect to the biological effects herein described. Re-search on the mechanisms of this selectivity of incorporationand analysis of specific fatty acid phospholipid pools whereoleate becomes mostly incorporated therefore appear war-ranted. In addition, research on the intimate molecular mech-anism by which the presence of a single double bond in thestructure of oleate may cause—directly or through an as-yet-unknown metabolite(s)—the reported attenuation of NF-kBactivation and of the endothelial responses to cytokines(possibly by interference with the generation of reactiveoxygen species) appears to be of considerable interest.
AcknowledgmentsThe entirety of this work was carried out at the Lecce Section of theCNR Institute of Clinical Physiology and at the Institute of Physi-ology at the University of Lecce, Italy. Minor support from theNational Research Council and Ministero dell’Universita e dellaRicerca Scientifica (60%) is acknowledged. The authors also expressgratitude to Fondazione Rico Semeraro—Lecce for fellowship sup-port to Dr M.A. Carluccio and to the International Association forPost-Graduate Education for the purchase of disposables. Theauthors also wish to express gratitude to the Department of Obstet-rics and Gynecology at the Vito Fazzi Hospital in Lecce for thesupply of umbilical cords; Dr Giovan Battista Lobreglio, Laboratoryof Clinical Chemistry, Vito Fazzi Hospital, Lecce, for the use offlow-cytometry apparatus; Dr Riccardo Guzzardi, Head, LecceSection of the CNR Institute of Clinical Physiology; and to thosemany who in general have supported the establishment of anAtherosclerosis Research Laboratory in Lecce. The authors are also
grateful to Dr Michael A. Gimbrone, Jr, Brigham and Women’sHospital, and Dr Arnold Freedman, Dana Farber Cancer Institute,Boston, Mass, for generous gifts of the antibodies used in thesestudies, and to Dr Peter Libby, Brigham and Women’s Hospital,Boston, Mass, for generous gifts of IL-1a.
References1. Keys A, Menotti A, Karvonen M, Aravanis C, Blackburn H, Buzina R,
Djordjevic B, Dontas A, Fidanza F, Keys M. The diet and 15-year deathrate in the Seven Countries Study.Am J Epidemiol. 1986;124:903–915.
2. Willett W. Diet and coronary heart disease.Monogr Epidemiol Biostat.1990;15:341–379.
3. World Health Organization. Diet, nutrition, and the prevention of chronicdiseases: report of WHO Study Group. Geneva, Switzerland: WorldHealth Organization; 1990:1–75.
4. Assmann G, de Backer G, Bagnara S, Betteridge J, Crepaldi G,Fernandez-Cruz A, Godfredsen J, Jacotot B, Paoletti R, Renaud S, RicciG, Rocha E, Trautwein E, Urbinati GC, Varela G. International consensusstatement: olive oil and the Mediterranean diet: implications for health inEurope.Proceedings of an International Meeting of the European Com-mission. Rome, Italy: April 11, 1997. 1997:1–3.
5. Mensink R, Katan M. Effect of dietary fatty acids on serum lipids andlipoproteins: a meta-analysis of 27 trials.Arterioscler Thromb. 1992;12:911–919.
6. Gardner C, Kraemer H. Monounsaturated versus polyunsaturated dietaryfat and serum lipids: a meta-analysis.Arterioscler Thromb Vasc Biol.1995;15:1917–1927.
7. Denke M. Cholesterol-lowering diets: a review of evidence.Arch InternMed. 1995;155:17–26.
8. Riemersma R, Wood D, Butler S, Elton R, Oliver M, Salo M, Nikkari T,Vartiainen E, Puska P, Gey F. Linoleic acid content in adipose tissue andcoronary heart disease.BMJ. 1986;292:1423–1427.
9. Espino-Montoro A, Lopez-Miranda J, Castro P, et al. Monounsaturatedfatty acid enriched diets lower plasma insulin levels and blood pressure inhealthy young men.Nutr Metab Cardiovasc Dis. 1996;6:147–154.
10. Garg A, Grundy S, Unger R. Comparison of effects of high and lowcarbohydrate diets on plasma lipoprotein and insulin sensitivity inpatients with mild NIDDM.Diabetes. 1992;41:1278–1285.
11. Griffin M, Dimitriadis E, Lenehan K, et al. Non-insulin-dependentdiabetes mellitus: dietary monounsaturated fatty acids and low-densitylipoprotein composition and function.Q J Med. 1996;89:211–216.
12. Hannah J, Howard B. Dietary fats, insulin resistance, and diabetes.J Car-diovasc Risk. 1994;1:31–37.
13. Bosello O, Armellini F, Zamboni M. Obesity. In: Spiller G, ed.TheMediterranean Diets in Health and Disease.New York, NY: VanNostrand Reinhold; 1991:256–276.
14. Gimbrone MA Jr, Kume N, Cybulsky MI. Vascular endothelial dys-function and the pathogenesis of atherosclerosis. In: Weber P, Leaf A,eds.Atherosclerosis Reviews. New York, NY: Raven Press; 1993:1–9.
15. Harris W. Fish oils and plasma lipid and lipoprotein metabolism inhumans: a critical review.J Lipid Res. 1989;30:785–807.
16. Newman WP, Middaugh JP, Propst MT, Rogers DR. Atherosclerosis inAlaska natives and non-natives.Lancet. 1993;341:1056–1057.
17. De Caterina R, Cybulsky MI, Clinton SK, Gimbrone MA Jr, Libby P. Thev-3 fatty acid docosahexaenoate reduces cytokine-induced expression ofpro-atherogenic and pro-inflammatory proteins in human endothelialcells.Arterioscler Thromb. 1994;14:1829–1836.
18. De Caterina R, Cybulsky MI, Clinton SK, Gimbrone MA Jr, Libby P.Omega-3 fatty acids and endothelial leukocyte adhesion molecules.Pros-taglandins Leukot Essent Fatty Acids. 1995;52:192–195.
19. Weber C, Erl W, Pietsch A, Danesch U, Weber PC. Docosahexaenoicacid selectively attenuates induction of vascular cell adhesion molecule-1and subsequent monocytic cell adhesion to human endothelial cells stim-ulated by tumor necrosis factor-a. Arterioscler Thromb Vasc Biol. 1995;15:622–628.
20. Collins T. Endothelial nuclear factor-kB and the initiation of the athero-sclerotic lesion.Lab Invest. 1993;68:499–508.
21. De Caterina R, Bernini W, Libby P. Fatty acid structural requirements forthe inhibition of endothelial activation.Circulation. 1996;94(suppl.I):I-102.
22. Gimbrone MA Jr. Culture of vascular endothelium. In: Spaet T, ed.Progress in Hemostasis and Thrombosis. Vol 3. New York, NY: Grune& Stratton; 1976:1–28.
23. Thornton S, Muller S, Levine E. Human endothelial cells: use of heparinin cloning and long term serial cultivation.Science. 1983;222:623–625.
Carluccio et al February 1999 227
by guest on January 18, 2016http://atvb.ahajournals.org/Downloaded from

24. Maciag T, Cerundolo J, Ilsley J, Kelley P, Forand R. An endothelial cellgrowth factor from bovine hypothalamus: identification and partial char-acterization.Proc Natl Acad Sci U S A. 1976;76:5674–5678.
25. Warner SJC, Friedman GB, Libby P. Regulation of major histocompati-bility gene expression in cultured human vascular smooth muscle cells.Arteriosclerosis. 1989;9:279–288.
26. Pober J, Gimbrone MA Jr, Lapierre L, Mendrick D, Fiers W, Rothlein R,Springer T. Overlapping patterns of activation of human endothelial cellsby interleukin 1, tumor necrosis factor, and immune interferon.J Immunol. 1986;137:1893–1896.
27. Bradford M. A rapid and sensitive method for the quantitation ofmicrogram quantities of proteins utilizing the principle of protein-dyebinding.Anal Biochem. 1976;72:248–254.
28. Spertini O, Luscinskas FW, Gimbrone MA Jr, Tedder T. Monocyteattachment to activated human vascular endothelium in vitro is mediatedby leukocyte adhesion molecule-1 (L-selectin) under nonstatic conditions.J Exp Med. 1992;175:1789–1792.
29. Schleimer R, Sterbinsky S, Kaiser J, Bickel C, Klunk D, Tomioka K,Newman W, Luscinskas FW, Gimbrone MA Jr, McIntyre W, Bochner B.Interleukin-4 induces adherence of human eosinophils and basophils butnot neutrophils to endothelium.J Immunol. 1992;148:1086–1092.
30. Chomczynski P, Sacchi N. Single-step method of RNA isolation by acidguanidinium thiocyanate-phenol-chloroform extraction.Anal Biochem.1986;162:156–159.
31. Dignam J, Lebovitz R, Roeder R. Accurate transcription initiation byRNA polymerase II in a soluble extract from isolated mammalian nuclei.Nucleic Acids Res. 1983;11:1475–1489.
32. Iademarco M, McQuillan J, Rose G, Dean D. Characterization of thepromoter for vascular cell adhesion molecule-1 (VCAM-1).J Biol Chem.1992;267:16323–16329.
33. Morrison W, Smith L. Preparation of fatty acid methyl esters and dim-ethylacetals from lipids with boron fluoride-methanol.J Lipid Res. 1964;5:600–608.
34. Deisher T, Haddix T, Montgomery K, Pohlman T, Kaushansky K,Harlan J. The role of protein kinase C in the induction of VCAM-1expression on human umbilical vein endothelial cells.FEBS Lett.1993;331:285–290.
35. Rollins BJ. JE/MCP-1: an early response gene encodes a monocyte-specific cytokine.Cancer Cells. 1991;3:517–524.
36. Vance DE, Vance JE.Biochemistry of Lipids and Membranes. New York,NY: Benjamin/Cummings; 1985:1–220.
228 Oleic Acid Inhibits Endothelial Activation
by guest on January 18, 2016http://atvb.ahajournals.org/Downloaded from

Maffia, Giuseppe Nicolardi, Alessandro Distante, Carlo Storelli and Raffaele De CaterinaMaria Annunziata Carluccio, Marika Massaro, Cosima Bonfrate, Luisa Siculella, Michele
of a Nutritional Component in the Mediterranean DietOleic Acid Inhibits Endothelial Activation: A Direct Vascular Antiatherogenic Mechanism
Print ISSN: 1079-5642. Online ISSN: 1524-4636 Copyright © 1999 American Heart Association, Inc. All rights reserved.
Greenville Avenue, Dallas, TX 75231is published by the American Heart Association, 7272Arteriosclerosis, Thrombosis, and Vascular Biology
doi: 10.1161/01.ATV.19.2.2201999;19:220-228Arterioscler Thromb Vasc Biol.
http://atvb.ahajournals.org/content/19/2/220World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://atvb.ahajournals.org//subscriptions/
at: is onlineArteriosclerosis, Thrombosis, and Vascular Biology Information about subscribing to Subscriptions:
http://www.lww.com/reprints
Information about reprints can be found online at: Reprints:
document. AnswerPermissions and Rights Question andunder Services. Further information about this process is available in the
permission is being requested is located, click Request Permissions in the middle column of the Web page whichCopyright Clearance Center, not the Editorial Office. Once the online version of the published article for
can be obtained via RightsLink, a service of theArteriosclerosis, Thrombosis, and Vascular Biologyin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on January 18, 2016http://atvb.ahajournals.org/Downloaded from
Copyright © 2022 FDOKUMEN




















