
Nutrient transfer from soil nematodes to plants: a direct pathway provided by the mycorrhizal...
8
Plant, Cell and Environment (2001) 24, 1219–1226 © 2001 Blackwell Science Ltd 1219 Blackwell Science, LtdOxford, UK PCEPlant, Cell and Environment0016-8025Blackwell Science Ltd 2001 24 769 Nutrient mobilization by mycorrhizal mycelium J. Perez-Moreno & D. J. Read Original ArticleBEES SGML Correspondence: Professor D. J. Read. Fax: + 44 114 222 0002; e-mail: D.J.Read@Sheffield.ac.uk Nutrient transfer from soil nematodes to plants: a direct pathway provided by the mycorrhizal mycelial network J. PEREZ-MORENO* & D. J. READ Department of Animal and Plant Sciences, University of Sheffield, Sheffield, S10 2TN, UK ABSTRACT A pathway for the transfer of nutrients from dead nema- todes to mycorrhizal plants is described for the first time. Plants of Betula pendula were grown in transparent micro- cosms in the mycorrhizal (M) or non-mycorrhizal (NM) condition, either with or without nematode necromass of known nitrogen (N) and phosphorus (P) contents as the major potential source of these elements. Plants colonized by the mycorrhizal fungus Paxillus involutus produced greater yields and had larger N and P contents in the pres- ence of nematodes than did their NM counterparts. The symbiotic systems were shown to exploit the N and P orig- inally contained in necromass more effectively, and to transfer the nutrients to the plants in quantities approxi- mately double those seen in NM systems. Even so, NM plants obtained sufficient N and P from dead nematodes to enable some enhancement of growth. Our observations confirm that mycorrhizal fungi provide the potential for the recycling of nutrients contained in this quantitatively important component of the soil mesofauna and demon- strate that the symbiotic pathway is considerably more effective than that provided by saprotrophs alone. The con- sequences of this nutrient transfer pathway for nutrient recycling in temperate forest ecosystems are considered. Key-words: Birch; ectomycorrhizal mycelium; mesofauna; nitrogen; nutrient mobilization; organic substrates; phosphorus; soil animals. INTRODUCTION In recent times, there has been increasing recognition of the ability of ectomycorrhizal (ECM) fungi to mobilize plant nutrients contained in complex organic substrates (Leake & Read 1997). Although the potential for such activities was first revealed in studies using monoxenic cultures of fungi grown in the presence of pure compounds such as proteins (Abuzinadah & Read 1986a, b) and organic phos- phates (Dighton 1983; Hilger & Krause 1989; Antibus, Sin- sabaugh & Linkins 1992), emphasis has shifted, of late, to more realistic scenarios in which the fungi are grown in association with their hosts and are provided with sub- strates in the form in which they are likely to be encoun- tered in nature. These have included detrital material collected from the fermentation horizon (FH) of forest soils (Bending & Read 1995) and pollen, a relatively well- defined potential substrate that is deposited annually in large quantities onto the forest floor (Perez-Moreno and Read 2001). Among the other potential sources of nutrients for ECM plants are soil animals. Interestingly, whereas the meso- fauna of forest soils has been well characterized both in terms of the quantitative representation of the various taxa (Petersen 1982) and of their contribution to the dynamics of decomposer food webs (Verhoef & Brussaard 1990), rel- atively little attention has been paid to the processes whereby the nutrients contained in the animals themselves are recycled through these webs. In the context of mycor- rhizal systems, emphasis has been placed upon the possibil- ity that fungivorous components of the mesofauna may reduce the effectiveness of the mycelial network in soils (Gange 2000), while the possibility that the necromass of soil animals might constitute a significant source of nutri- ents for mycorrhizal fungi has been largely overlooked. However, Klironomos and Hart (2001) have recently shown that one ectomycorrhizal fungus, Laccaria bicolor (Maire) Orton, might act as a predator on collembolans, so extracting nitrogen from their bodies. Among the mesofaunal groups known to be present in very large numbers in temperate forest soils of the kind that support ECM trees are the nematodes. Petersen (1982) estimated that the biomass of these organisms in temperate forest ranged between 15 and 3040 mg dry weight (DW) m −2 , these values being equivalent to 370 × 10 3 and 29·8 × 10 6 individuals m −2 . As each individual nematode is a nutri- ent-enriched package that, on death of the organism, becomes a potential substrate for ECM fungi, it is of great interest to determine the extent to which mycorrhizal infec- tion enables these resources to be captured by the plant. Here, using birch seedlings grown in microcosms with the symbiotic fungus Paxillus involutus (Fr.) Fr., we test the hypothesis that a mycorrhizal symbiont has the ability to mobilize nutrients from dead nematodes. In addition, we evaluate the response of the colonized plant, in terms both of growth and nutrient contents, associated with the mobi- lization process. *Present address: Microbiología, Edafología-IRENAT, Colegio de Post- graduados, Montecillo, Texcoco, Edo. de México, CP56230, Mexico.
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Nutrient transfer from soil nematodes to plants: a direct pathway provided by the mycorrhizal...

Plant, Cell and Environment
(2001)
24,
1219–1226
©
2001 Blackwell Science Ltd
1219
Blackwell Science, LtdOxford, UKPCEPlant, Cell and Environment0016-8025Blackwell Science Ltd 200124769Nutrient mobilization by mycorrhizal myceliumJ. Perez-Moreno & D. J. ReadOriginal ArticleBEES SGML
Correspondence: Professor D. J. Read. Fax: +44
114
222
0002; e-mail: [email protected]
Nutrient transfer from soil nematodes to plants: a direct pathway provided by the mycorrhizal mycelial network
J.
PEREZ-MORENO
* & D. J.
READ
Department of Animal and Plant Sciences, University of Sheffield, Sheffield, S10 2TN, UK
ABSTRACT
A pathway for the transfer of nutrients from dead nema-todes to mycorrhizal plants is described for the first time.Plants of
Betula pendula
were grown in transparent micro-cosms in the mycorrhizal (M) or non-mycorrhizal (NM)condition, either with or without nematode necromass ofknown nitrogen (N) and phosphorus (P) contents as themajor potential source of these elements. Plants colonizedby the mycorrhizal fungus
Paxillus involutus
producedgreater yields and had larger N and P contents in the pres-ence of nematodes than did their NM counterparts. Thesymbiotic systems were shown to exploit the N and P orig-inally contained in necromass more effectively, and totransfer the nutrients to the plants in quantities approxi-mately double those seen in NM systems. Even so, NMplants obtained sufficient N and P from dead nematodes toenable some enhancement of growth. Our observationsconfirm that mycorrhizal fungi provide the potential for therecycling of nutrients contained in this quantitativelyimportant component of the soil mesofauna and demon-strate that the symbiotic pathway is considerably moreeffective than that provided by saprotrophs alone. The con-sequences of this nutrient transfer pathway for nutrientrecycling in temperate forest ecosystems are considered.
Key-words
: Birch; ectomycorrhizal mycelium; mesofauna;nitrogen; nutrient mobilization; organic substrates;phosphorus; soil animals.
INTRODUCTION
In recent times, there has been increasing recognition of theability of ectomycorrhizal (ECM) fungi to mobilize plantnutrients contained in complex organic substrates (Leake& Read 1997). Although the potential for such activitieswas first revealed in studies using monoxenic cultures offungi grown in the presence of pure compounds such asproteins (Abuzinadah & Read 1986a, b) and organic phos-phates (Dighton 1983; Hilger & Krause 1989; Antibus, Sin-
sabaugh & Linkins 1992), emphasis has shifted, of late, tomore realistic scenarios in which the fungi are grown inassociation with their hosts and are provided with sub-strates in the form in which they are likely to be encoun-tered in nature. These have included detrital materialcollected from the fermentation horizon (FH) of forest soils(Bending & Read 1995) and pollen, a relatively well-defined potential substrate that is deposited annually inlarge quantities onto the forest floor (Perez-Moreno andRead 2001).
Among the other potential sources of nutrients for ECMplants are soil animals. Interestingly, whereas the meso-fauna of forest soils has been well characterized both interms of the quantitative representation of the various taxa(Petersen 1982) and of their contribution to the dynamicsof decomposer food webs (Verhoef & Brussaard 1990), rel-atively little attention has been paid to the processeswhereby the nutrients contained in the animals themselvesare recycled through these webs. In the context of mycor-rhizal systems, emphasis has been placed upon the possibil-ity that fungivorous components of the mesofauna mayreduce the effectiveness of the mycelial network in soils(Gange 2000), while the possibility that the necromass ofsoil animals might constitute a significant source of nutri-ents for mycorrhizal fungi has been largely overlooked.However, Klironomos and Hart (2001) have recentlyshown that one ectomycorrhizal fungus,
Laccaria bicolor
(Maire) Orton, might act as a predator on collembolans, soextracting nitrogen from their bodies.
Among the mesofaunal groups known to be present invery large numbers in temperate forest soils of the kind thatsupport ECM trees are the nematodes. Petersen (1982)estimated that the biomass of these organisms in temperateforest ranged between 15 and 3040 mg dry weight (DW)m
−
2
, these values being equivalent to 370
×
10
3
and 29·8
×
10
6
individuals m
−
2
. As each individual nematode is a nutri-ent-enriched package that, on death of the organism,becomes a potential substrate for ECM fungi, it is of greatinterest to determine the extent to which mycorrhizal infec-tion enables these resources to be captured by the plant.
Here, using birch seedlings grown in microcosms withthe symbiotic fungus
Paxillus involutus
(Fr.) Fr., we test thehypothesis that a mycorrhizal symbiont has the ability tomobilize nutrients from dead nematodes. In addition, weevaluate the response of the colonized plant, in terms bothof growth and nutrient contents, associated with the mobi-lization process.
*
Present address: Microbiología, Edafología-IRENAT, Colegio de Post-graduados, Montecillo, Texcoco, Edo. de México, CP56230, Mexico.

1220
J. Perez-Moreno & D. J. Read
© 2001 Blackwell Science Ltd,
Plant, Cell and Environment
,
24
, 1219–1226
MATERIALS AND METHODS
Mycorrhizal synthesis
Mycorrhizas were synthesized on
Betula pendula
Roth.seedlings using the ectomycorrhizal fungus
Paxillus involu-tus
following a procedure adapted from Brun
et al
. (1995).Seeds of
B. pendula
were surface-sterilized by exposure toH
2
O
2
for 25 min, then rinsed with sterile distilled water andplaced to germinate on water agar in Petri dishes.
Paxillusinvolutus
(strain USPI97 isolated from Cropton Forest,north Yorkshire, UK) was grown on cellophane overlyingagar supplemented with modified Norkrans (1953) nutrientsolution (see Perez-Moreno & Read 2000). Once the myce-lium had covered approximately one-third of the area ofthe Petri dish, the birch seedlings were aseptically trans-ferred, their roots being placed in direct contact with themycorrhizal fungus. All lateral roots were colonized within15 d, at which time seedlings were transferred singly tomicrocosms.
Construction of microcosms
Microcosms were constructed from 15
×
15
×
1 cm sheets oftransparent plastic as described by Perez-Moreno & Read(2000). Over the surface of the lower plate, a film of wateragar was poured, onto which a thin layer of inert clay (leca)granules was distributed. Finally, a layer of
Sphagnum
peat(2·5 g DW per microcosm) was spread over the granules toprovide a nutrient-poor seminatural matrix for support ofthe seedlings and growth of the vegetative mycorrhizalmycelium. Single mycorrhizal (M) or non-mycorrhizal(NM) birch seedlings were transferred from the water agarplates to each of a series of such observation chambers towhich were clipped an upper plate forming a sandwich.Shoots of the plants projected into the atmosphere. Rootswere free to forage in the clay-peat substrate. The micro-cosms were wrapped in aluminium foil and incubated in acontrolled environment growth room under an 18 h photo-period (150
µ
mol m
2
s
−
1)
with a day : night temperature of15 : 10
°
C.
Extraction of nematodes and addition to the microcosms
The nematode selected for use,
Heterorhabditis megidis,
was obtained as a pure culture from a commercial source(Scarletts Plant Care, Colchester, UK). The cultures werestored at +5
°
C until required, then large aggregates ofnematodes, together with the clay carrier in which theywere embedded, were added to distilled water and centri-fuged at approximately 1000
¥
g
for 10 min in order toseparate the nematodes from the culture medium. The cen-trifugation produced a discrete layer of nematodes at thebase of the water column, overlying the clay carrier. Fromthis layer, pure suspensions of the nematodes could then becarefully removed using a 250
µ
L pipette. The nematodeswere placed onto sterile Petri dishes, which were thenexposed to direct sunlight to produce rapid dehydration.
The dry nematode aggregates were checked under themicroscope to ensure that only pure samples were used inthe experiments. They were then stored at +5
°
C untilrequired. Preliminary experiments in which these nema-todes were rehydrated following desiccation confirmed thatmortality was 100%.
When the vegetative ectomycorrhizal mycelium of
P.involutus
had grown sufficiently to cover most of the peat-granule surface, the trays containing dead nematodes wereadded to microcosms containing M or NM plants using amodification of the approach used for pollen by Perez-Moreno & Read (2001). There was an additional set of Mand NM chambers to which no nematodes were added.Dead air-dried nematodes were added in small (3 cm
×
3 cm) plastic trays to half the M and NM microcosms. Eachtray contained, as a support matrix, 2 g dry wt of clay (leca)granules in 2 mL of water agar (0·8%), onto the surface ofwhich the nematodes were spread. The nematodes wereadded in two aliquots, the first consisting of 15 mg added attime zero and the second of 35 mg, which was added 45 dlater. Two nematode-containing trays (i.e. 50 mg DW nem-atodes in total) were added per chamber together with asingle tray containing support matrix only, the latter beingused to provide a comparison of mycelial development intrays with and without nematodes.
The quantity of nematode material added to each micro-cosm when expressed on an area basis, was equivalent to2220 mg DW m
−
2
, and hence lay within the range 15 and3040 mg DW m
−
2
reported by Petersen (1982) as beingtypically found in temperate forest soils. There were threereplicate microcosms with nematodes (designated:+nematodes) and three without (designated: –nematodes)in both M or NM treatments.
All microcosms were incubated in a controlled environ-ment growth cabinet (day–night temperature 15–10
°
C, daylength 18 h, irradiance 150
µ
mol m
−
2
s
−
1
) for a further periodof 195 d after which time harvest was carried out. Duringthe incubation period the pattern of development of thevegetative mycelium of
P. involutus
over the trays contain-ing dead nematodes, was followed. Regular visual inspec-tion of all microcosms confirmed that roots neither enterednor contacted the trays themselves.
Nutrient analysis of plants and nematodes
At harvest, both M and NM plants grown in +or –nematode microcosms were partitioned into roots andshoots and oven-dried at 80
°
C before their dry weights andtissue N and P contents were determined in replicate sub-samples. The tissues were digested until clear in a concen-trated H
2
SO
4
(98%)–salicylic acid mix (33 g of salicylic aciddissolved in 1 litre of H
2
SO
4
) to which a catalyst was added(lithium sulphate : copper sulphate in a 10 : 1 ratio). Theresulting samples were allowed to stand until they werecold and then were diluted with water. The extracts werestored at
−
20
°
C before colorimetric analysis using a Teca-tor 5012 Analyser (Foss UK Ltd, Didcot, Oxfordshire, UK).N and P concentrations in the nematodes were determined
©
2001 Blackwell Science Ltd
,
Plant, Cell and Environment
,
24
, 1219–1226
Nutrient mobilization by mycorrhizal mycelium
1221
at time zero and at harvest by the same methods as thosedescribed for plant materials.
RESULTS
Growth responses of the plants
Over the course of the experiment, birch plants grown inboth the M and NM condition showed a progressiveincrease in vigour in the presence of dead nematodes. Incontrast, plants grown in the equivalent treatment withoutnematodes were of stunted appearance and leaf develop-ment was weak (Fig. 1). At harvest it was found that in thepresence of nematodes mycorrhizal colonization facilitateda significant increase of total yield of plants relative to thoseseen in NM treatments (476·1
±
18·0 versus 307·0
±
32·0;means
±
SE,
n
= 3; Student’s
t
-test
P
< 0·05) (Fig. 2). Both
shoots and roots of M plants had significantly greater bio-mass than did their NM counterparts (Fig. 2). Whole plant,shoot and root yields in the + nematode treatment were sig-nificantly greater than those in –nematode treatment inboth M and NM microcosms (Fig. 2).
Nutrient contents of the plants
Evaluation of nutrient contents of the plants showed a sim-ilar overall pattern to that revealed by analysis of theirgrowth. The total N and P contents of M plants grown inmicrocosms with nematodes were, respectively, 2·1 and 1·7times greater than those of their equivalent NM treatments,the differences between these treatments being highly sig-nificant (Student’s
t
-test,
P
< 0·01)(Table 1). The shoots androots of both M (Fig. 3a,c) and NM (Fig. 3b,d) plants grownwith nematodes contained significantly greater N and P
Figure
1.
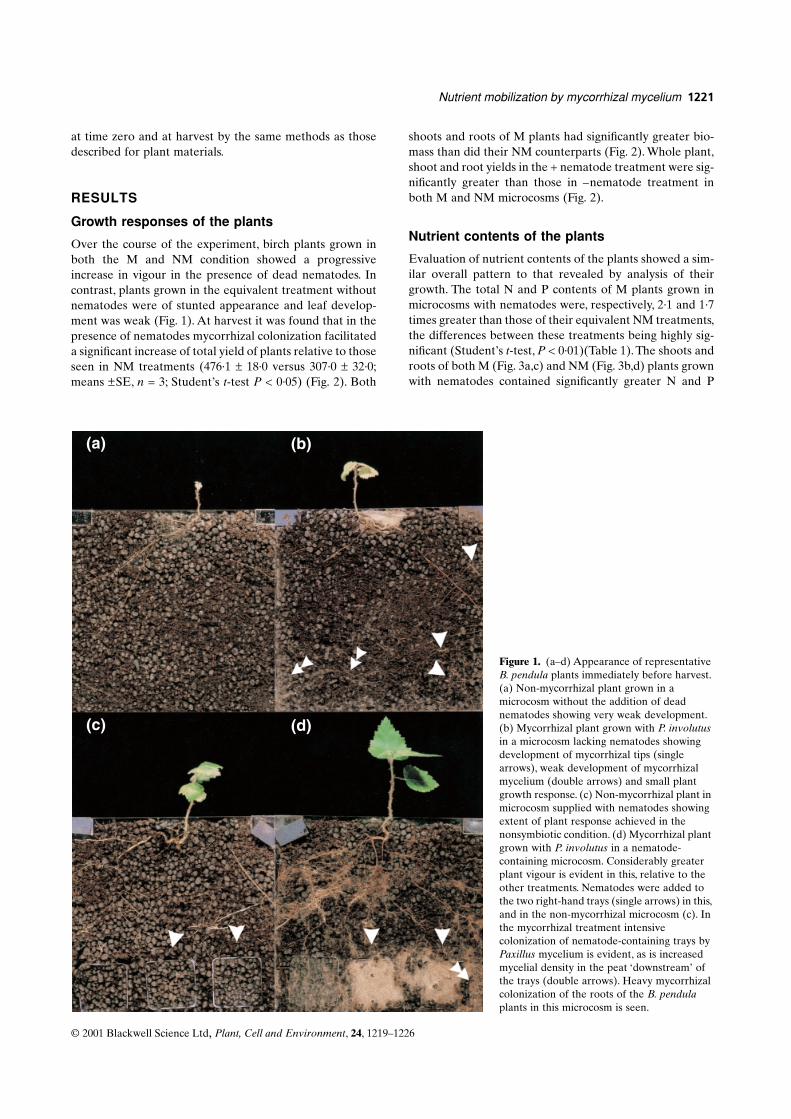
(a–d) Appearance of representative
B. pendula
plants immediately before harvest. (a) Non-mycorrhizal plant grown in a microcosm without the addition of dead nematodes showing very weak development. (b) Mycorrhizal plant grown with
P. involutus
in a microcosm lacking nematodes showing development of mycorrhizal tips (single arrows), weak development of mycorrhizal mycelium (double arrows) and small plant growth response. (c) Non-mycorrhizal plant in microcosm supplied with nematodes showing extent of plant response achieved in the nonsymbiotic condition. (d) Mycorrhizal plant grown with
P. involutus
in a nematode-containing microcosm. Considerably greater plant vigour is evident in this, relative to the other treatments. Nematodes were added to the two right-hand trays (single arrows) in this, and in the non-mycorrhizal microcosm (c). In the mycorrhizal treatment intensive colonization of nematode-containing trays by
Paxillus
mycelium is evident, as is increased mycelial density in the peat ‘downstream’ of the trays (double arrows). Heavy mycorrhizal colonization of the roots of the
B. pendula
plants in this microcosm is seen.
(a)
(c)
(b)
(d)

1222
J. Perez-Moreno & D. J. Read
© 2001 Blackwell Science Ltd,
Plant, Cell and Environment
,
24
, 1219–1226
contents than those of plants in microcosms without nem-atodes, the differences between + and –nematode treat-ments being significant in all cases except that of root Pcontents of NM plants (Fig. 3d).
The M plants had higher N and P contents than those inthe NM category. The N contents of shoots and roots of Mplants grown in microcosms with nematodes were 2·3 and1·9 times greater than those in the equivalent NM treat-ments, there being significant differences (Student’s
t
-test,
P
< 0·05) between both M and NM shoots (1642·5
±
203·0
versus 709·1
±
47·0; means
±
SE,
n
= 3) and roots (2434·8.
±
266·0 versus 1263·0
±
142·0; means
±
SE,
n
= 3) (Fig. 3a,b).Similarly in the case of P (Fig. 3c,d), the shoot and rootcontents were, respectively, 1·4 and 1·8 times greater in theM + nematode than those in the NM + nematode treat-ment. The whole-plant quantities of N and P were greaterin +nematode than in –nematode treatments in both M andNM plants with the exception of P in NM plants (Table 1).
Nutrient depletion in nematodes
The growth responses and gains of nutrients in M and NMplants grown in the presence of nematodes were paralleledby significant reductions of both N and P concentrations innematodes (Fig. 4). However, in the microcosms containingM plants larger amounts of nutrients were transferred fromthe nematodes than in those containing NM plants, therebeing significant differences between treatments (
P
< 0·05).The reductions in N and P concentrations of the nematodenecromass of microcosms containing M plants wereapproximately double those seen in microcosms supportingNM plants (Fig. 4). Colonization of dead nematodes by
P.involutus
was associated with a reduction of their total Nand P contents after 195 d of 68·2 and 65·7%, respectively.The differences between these and the NM systems, wherethe depletion of the same nutrients was only 37·1 and24·5%, respectively, were significant at
P
< 0·05 (Table 2).
Relation between nutrient gains in plants and their losses from nematodes
Comparisons of nutrient gain values in M and NM systemsindicate that the symbiosis facilitated significantly morenutrient transfer to the plants (Table 1). The total N and Ptransfer from nematodes in M plants was more than doublethat seen in their NM counterparts (Table 1). In the case ofP the gain by M plants amounted to 106·8
µ
g (Table 1)whereas the loss from the nematodes was 146·9
µ
g (Table2). Thus, only a portion, approximately 73%, of the Premoved from the dead nematodes was transferred from
Figure
2.
Dry weight yields of
B. pendula
plants grown with the ectomycorrhizal fungus
P. involutus
(above) or in the non-mycorrhizal condition (below) for 195
d in microcosms with (closed bars) and without (open bars) nematodes. Values are expressed as means
±
SE. Asterisks indicate significant differences within tissue category according to Student’s
t
-test: *
P
<
0·05; **
P
<
0·005.
(a)
(b)
Mycorrhizal
Non-mycorrhizal
480
360
240
120
0
480
360
240
120
0
DW
(m
g)
Shoot Root Whole plant
Table
1.
Total N and P contents of
B. pendula
grown in microcosms for 195
d, with or without dead nematodes as a potential nutrient source, either in the mycorrhizal condition with the fungus
P. involutus,
or as non-mycorrhizal plants
Treatment
Whole-plant contents (µg) Gain of nutrients by plants (µg) a
N P N P
Microcosms with non-mycorrhizal plants 1122·0 ± 31** 48·2 ± 11·0*With nematodes 1971·9 ± 148* 99·3 ± 5·8Without nematodes 849·9 ± 179 51·1 ± 15·0Microcosms with mycorrhizal plants 2810·5 ± 98 106·8 ± 8·0With nematodes 4077·3 ± 131** 166·1 ± 8·5**Without nematodes 1266·8 ± 188 59·3 ± 16·0
Values are means ± SE, n = 3.a Gain of nutrients by plants = (plant nutrient contents in microcosms with nematodes) − (plant nutrient contents in microcosms without nematodes). In this case, the ± SE were calculated by a postiori ranking of microcosms with mycorrhizal and non-mycorrhizal plants.Asterisks indicate significant differences between nutrients in microcosms with and without nematodes for whole-plant contents and between nutrients in microcosms with mycorrhizal and with non-mycorrhizal plants for gain of nutrients using t-tests at *P < 0·05 and **P < 0·01.
© 2001 Blackwell Science Ltd, Plant, Cell and Environment, 24, 1219–1226
Nutrient mobilization by mycorrhizal mycelium 1223
the fungus to the plant. The quantity (106·8 µg) of Pacquired by the M plants in the + nematode systems repre-sented 48% of the 223·5 µg originally present in the animaltissues. In contrast, in the NM systems, the P gain amountedonly to 48·2 µg (Table 1), this being half of that transferredin M systems, and representing only 22% of the total orig-inally present in nematodes.
A different overall pattern was seen in the case ofN, where the gain by M plants, amounting to 2810·5 µg(Table 1), was greater than the loss from the nematodes(2095·7 µg) (Table 2), there being significant differencesbetween these values (Student’s t-test, P < 0·05). The pos-sible basis of the apparent discrepancy between loss of Nfrom the putative source and gain by the plant is discussedbelow (Discussion). By contrast, in NM systems the N gainby plants of 1122·0 µg (Table 1) and the losses from nema-todes, 1140·0 µg (Table 2), were similar.
DISCUSSION
To our knowledge this is the first demonstration thatectomycorrhizal colonization can provide access to thenutrient pools contained in the bodies of soil nematodes.It is an important observation because nematode popula-tions are typically very large in the normally nutrient-impoverished forest soils that support ectomycorrhizalplant communities (Petersen 1982). The results indicatethat a significant proportion of the most important plantnutrients that are initially sequestered in the bodies ofsoil nematodes can be effectively recycled by mycorrhizalfungi to their autotrophic partners rather than beingimmobilized or transferred to other levels within the foodweb.
The ability of some saprotrophic soil fungi to use deadnematodes as nutrient sources has been previously recog-
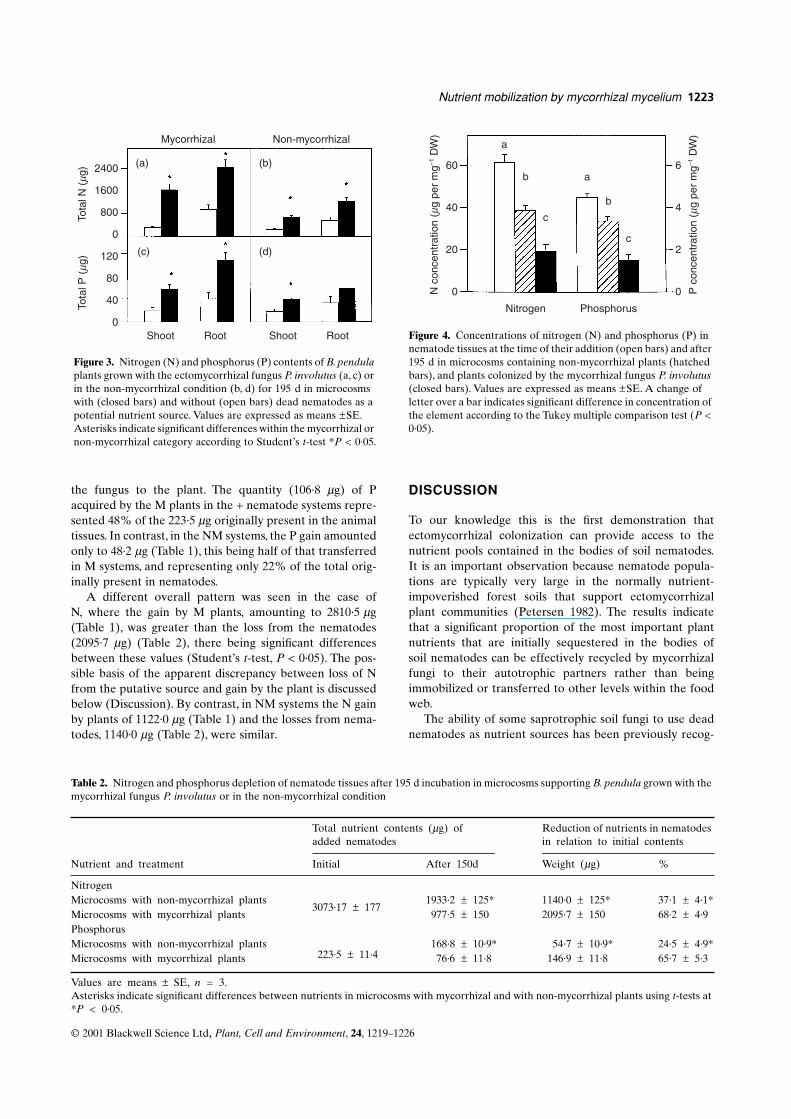
Figure 3. Nitrogen (N) and phosphorus (P) contents of B. pendula plants grown with the ectomycorrhizal fungus P. involutus (a, c) or in the non-mycorrhizal condition (b, d) for 195 d in microcosms with (closed bars) and without (open bars) dead nematodes as a potential nutrient source. Values are expressed as means ±SE. Asterisks indicate significant differences within the mycorrhizal or non-mycorrhizal category according to Student’s t-test *P < 0·05.
Mycorrhizal Non-mycorrhizal
(a) (b)
(d)(c)
2400
1600
800
0
120
80
40
0
Tota
l P (
mg)
Tota
l N (
mg)
Shoot Root Shoot Root Figure 4. Concentrations of nitrogen (N) and phosphorus (P) in nematode tissues at the time of their addition (open bars) and after 195 d in microcosms containing non-mycorrhizal plants (hatched bars), and plants colonized by the mycorrhizal fungus P. involutus (closed bars). Values are expressed as means ±SE. A change of letter over a bar indicates significant difference in concentration of the element according to the Tukey multiple comparison test (P < 0·05).
60
40
20
0
6
4
2
0
a
ab
c
b
c
Nitrogen Phosphorus
N c
once
ntra
tion
(mg
per
mg–1
DW
)
P c
once
ntra
tion
(mg
per
mg–1
DW
)
Table 2. Nitrogen and phosphorus depletion of nematode tissues after 195 d incubation in microcosms supporting B. pendula grown with the mycorrhizal fungus P. involutus or in the non-mycorrhizal condition
Total nutrient contents (µg) of added nematodes
Reduction of nutrients in nematodes in relation to initial contents
Nutrient and treatment Initial After 150d Weight (µg) %
NitrogenMicrocosms with non-mycorrhizal plants
3073·17 ± 1771933·2 ± 125* 1140·0 ± 125* 37·1 ± 4·1*
Microcosms with mycorrhizal plants 977·5 ± 150 2095·7 ± 150 68·2 ± 4·9PhosphorusMicrocosms with non-mycorrhizal plants
223·5 ± 11·4168·8 ± 10·9* 54·7 ± 10·9* 24·5 ± 4·9*
Microcosms with mycorrhizal plants 76·6 ± 11·8 146·9 ± 11·8 65·7 ± 5·3
Values are means ± SE, n = 3.Asterisks indicate significant differences between nutrients in microcosms with mycorrhizal and with non-mycorrhizal plants using t-tests at *P < 0·05.

1224 J. Perez-Moreno & D. J. Read
© 2001 Blackwell Science Ltd, Plant, Cell and Environment, 24, 1219–1226
nized. Dyer, Boddy & Preston-Meek (1992) supplied twobasidiomycetous wood decomposer fungi, Phanaerochaetevelutina (DC.Pers.) Parmasto and Stereum hirsutum(Willd.Fr.) S.F. Gray with necromass of the nematodePanagrellus redivivus (L.) as a sole nutrient source. In thepresence of this substrate they observed some increase inmean radial extension growth as well as of protease andacid phosphatase activity of P. velutina. Under the sameexperimental conditions increases in activity of thisenzymes were also observed in S. hirsutum. Since the ecto-mycorrhizal fungus P. involutus employed in the presentstudy, showed clear evidence of growth stimulation in andaround the nematode-containing trays and is known tohave the potential for release of these extracellularenzymes, it is possible to hypothesize that the improvedgrowth and nutrient acquisition seen in the colonized plantswas attributable to direct attack by the fungus upon thebodies of the nematodes. However, as considerable timeelapsed between placement of the necromass-containingtrays into the microcosms and their colonization by P. invo-lutus, it is more likely that the primary nutrient mobilizingevent were initiated by saprotrophs, the propagules ofwhich, be they fungal or bacterial, would be associated withthe bodies of the nematodes at the time of their death. Inthis context it is of interest that Tsuneda & Thorn (1995),while noting that the fungal saprotroph Pleurotus ostreatus(Jacq.Fr.) Kummer was able to colonize the corpses of deadnematodes, observed that decomposition was initiated bybacteria and that penetration by the fungus of the bacterialbiomass rather than direct attack appeared to facilitate itsaccess to nutrients.
If, as our observations suggest, the success of the symbi-osis in this case is based upon secondary exploitation ofnutrients released by earlier colonists the situation is clearlydifferent from that seen in the specialized group of nem-atophagous fungi that have the ability both to kill their preyand directly mobilize the nutrients contained in them bymeans of a variety of well-characterized enzymatic pro-cesses (Toth, Toth & Nordbring-Hertz 1980; Schenck et al.1980; Tunlid et al. 1994; Araujo et al. 1997).
The nutrient-foraging strategy deployed by ectomycor-rhizal fungi, which is dependent upon their access to carbo-hydrate supplies from the autotroph (Söderström & Read1987), is to produce an extensive mycelial system thatenables effective exploitation of the soil volume (Read1992). This ensures that while they may not be in direct con-tact with individual food depots such as the senescing bod-ies of nematodes, they are close enough to respond to anynearby enrichment of nutrient availability by localized pro-liferation of hyphal branching (Bending & Read 1995;Perez-Moreno & Read, 2000, 2001).
The present experiment provides a test of the relativeeffectiveness, for the plant, of mycorrhizal and saprotrophicfungal nutrient mobilization pathways, because in neithercase did roots make contact with the trays containing theanimal necromass.
As was shown when pollen was provided as the solepotential source of nutrients in mycorrhizal and non-
mycorhizal microcosms (Perez-Moreno & Read 2001),saprotrophic activities alone enabled sufficient mobiliza-tion of N and P to facilitate some growth stimulation andnutrient enrichment in the plant. However this process isrelatively ineffective presumably in part because saprotro-phs will transfer nutrients into the soil in all directions froma localized source rather than towards individual roots. Incontrast, the mycelium of the symbiont P. involutus, canallocate its resources directly to the interconnected mycor-rhizal roots through the mycelial network that is sustainedby carbon supplied by the host plant. The rapidity withwhich the symbiont mycelium proliferates after contactingthe substrate indicate that it will be able to respond effec-tively to short-term pulses of nutrient release.
The extent of N and P mobilization in mycorrhizal sys-tems, 68 and 65%, respectively, from dead nematodes wassomewhat less than that reported from pollen (Perez-Moreno & Read 2001) where the equivalent values were,respectively, 76 and 97%. However, the quantities of N andP mobilized from these animals were greater than thosereported from detrital material collected from the forestsoil FH (Entry, Rose & Cromack 1991; Bending & Read1995; Perez-Moreno & Read, 2000). Amounts of nutrientrelease from FH substrates ranged from 0 to 32% in thecase of N and from 3 to 40% in that of P, being in all casesmuch smaller than those from nematodes.
The nutrient analysis showed that in the case of M sys-tems, the N gain in the plants was greater than was the lossfrom nematodes. A possible explanation for the observeddiscrepancy could be the occurrence of N2-fixing activity inthe conspicuous ECM mycelial patches associated with thedead nematodes. Both associative N2-fixation and the pres-ence of bacterial species, e.g. Bacillus spp., known to haveN2-fixing potential have been previously found in the myc-orrhizosphere of forest ECM trees (Li & Hung 1987; Li,Massicote & Moore 1992; Nurmiaho-Lassila et al. 1997;Timonen et al. 1998). It has been pointed out by Perez-Moreno & Read (2000) and Sen (2000) that the vigorousgrowth of external vegetative ECM mycelium would lead,by enhancement of CO2 release, both to low oxygen tensionand high C-availability, providing therefore an ideal nichefor the development of associative N2-fixing bacteria. Theoccurrence of N2-fixation requires confirmation, preferablyby direct quantification using 15N labelling methods. Perez-Moreno & Read (2000) found a similar discrepancybetween N loss from FH materials and gains by mycorrhizalplants. In that study, the excess N observed in the plants waseven greater than that observed in the present study.
An additional possibility is that exploitation of the nutri-ent contents of the nematodes has a ‘priming’ effect uponthe fungus enabling it more effectively to scavenge for Nfrom the surrounding growth environment. It was observedthat formation of dense mycelial patches over traysenriched with dead nematodes, was followed by enhance-ment of mycelial growth in areas distal to the trays andalong lower lateral edges of the microcosms (Fig. 1b). Stim-ulation of hyphal growth in territories ‘downstream’ ofnutrient enriched patches has been observed in

© 2001 Blackwell Science Ltd, Plant, Cell and Environment, 24, 1219–1226
Nutrient mobilization by mycorrhizal mycelium 1225
saprotrophic fungi (Rayner 1996). An effect of this growthenhancement, which may be facilitated by release from Plimitation, could be to increase the ability of the fungus toscavenge for, or mineralize N from, the peat.
It is necessary to consider the extent to which and themechanisms whereby the processes observed in this studymight operate in the field. Clearly the distribution of nem-atodes is unlikely to be as clustered in nature as in themicrocosm systems. However there is evidence (De Goede,Verschoor & Georgieva 1993) that, under field conditions, asignificant proportion of the numerically large populationof these organisms is concentrated in that part of the soilprofile, the FH horizon, in which ECM mycelium prolifer-ates most intensively. Indeed this concentration may in partarise specifically because, for the fungivorous nematodes atleast, this horizon is the richest source of nutrient provision.It is not clear from the literature whether this group of nem-atodes does feed extensively upon mycorrhizal mycelia butthe size of its biomass indicates that major impacts uponsome components of the fungal system are likely.
Nematodes, be they detritivores, bacterivores or fungi-vores have nutrient-enriched bodies in which the proteincontents range from 50 to 80% (Lee & Atkinson 1976). Tothe extent that these organisms, except in the egg stage, donot produce structural nitrogenous polymers like chitin,they probably represent a more labile potential source ofnutrients than do some other mesofaunal groups, for exam-ple collembolans. This would lead to relatively rapid turn-over of the nutrients originally sequestered in thepopulation. Since Laccaria bicolor has been shown(Klironomos and Hart 2001) to attack the bodies of livingas well as dead collembolans, and to facilitate transfer ofnitrogen from the animals to its host plant, it appears thateven these more complex nutrient sources may be accessi-ble to some mycorrhizal associations.
Whether or not mycorrhizal fungi such as Paxillus, withtheir acknowledged abilities to release the extracellularenzyme protease (Abuzinadah & Read 1986a; Finlay,Frostegard & Sonnerfeldt 1992) and phosphatase (Hilger,Thomas & Krause 1986; Hilger & Krause 1989; Kielisze-wska-Rokicka 1992) are primary decomposers, it is clearthat the ability of their mycelia to occupy the FH and torapidly proliferate within it in response to localized nutri-tional cues will facilitate effective involvement in thedecomposer food web, to the direct advantage of the plant.Within this web, the key to the foraging success of theECM fungi, whether it involves primary degradation ofnematode corpses or secondary attack upon the first wavesof saprotrophs, is likely to be their freedom from carbonlimitation.
ACKNOWLEDGMENTS
We thank Ms Irene Johnson, Mr Glynn Woods and MrDavid Hollingworth for excellent technical assistance. Wealso thank the British Chevening Scholarships Programme,CONACyT (Consejo Nacional de Ciencia y Tecnología)
and Colegio de Postgraduados (Mexico) for provision offinancial support to J.P.-M.
REFERENCES
Abuzinadah R.A. & Read D.J. (1986a) The role of proteins inthe nitrogen nutrition of ectomycorrhizal plants I. Utilisation ofpeptides and proteins by ectomycorrhizal fungi. New Phytologist103, 481–493.
Abuzinadah R.A. & Read D.J. (1986b) The role of proteins inthe nitrogen nutrition of ectomycorrhizal plants II. Utilisation ofprotein by mycorrhizal plants of Pinus contorta. New Phytologist103, 495–506.
Antibus R.K., Sinsabaugh R.L. & Linkins A.E. (1992) Phos-phatase-activities and phosphorus uptake from inositol phos-phate by ectomycorrhizal fungi. Canadian Journal of Botany 70,794–801.
Araujo J.V., Junghans T.G., Alfenas A.C. & Gomes A.P.S.(1997) Isoenzyme analysis of Arthrobotrys, a nematode-trap-ping fungus. Brazilian Journal of Medical and BiologicalResearch 30, 1149–1152.
Bending G.D. & Read D.J. (1995) The structure and function ofthe vegetative mycelium of ectomycorrhizal plants VI. Activitiesof nutrient mobilising enzymes in birch litter colonised by Pax-illus involutus (Fr.) Fr. New Phytologist 130, 411–417.
Brun A., Chalot M., Finlay R.D. & Söderström B. (1995) Struc-ture and function of the ectomycorrhizal association betweenPaxillus involutus (Batsch) Fr. & Betula pendula Roth. I.Dynamics of mycorrhiza formation. New Phytologist 129, 487–493.
De Goede R.G.M., Verschoor B.C. & Georgieva S.S. (1993)Nematode distribution, trophic structure and biomass in a pri-mary succession of blown-out areas in a drift sand landscape.Fundamental and Applied Nematology 16, 525–538.
Dighton J. (1983) Phosphatase production by mycorrhizal fungi.Plant Soil 71, 455–462.
Dyer H.C., Boddy L. & Preston-Meek C.M. (1992) Effect of thenematode Panagrellus redivivus on growth and enzyme produc-tion by Phanerochaete velutina and Stereum hirsutum. Mycolog-ical Research 96, 1019–1028.
Entry J.A., Rose C.L. & Cromack K. Jr (1991) Litter decompo-sition and nutrient release in ectomycorrhizal mat soils of a Dou-glas fir ecosystem. Soil, Biology and Biochemistry 23, 285–290.
Finlay R.D., Frostegard A. & Sonnerfeldt A.M. (1992) Utiliza-tion of organic and inorganic nitrogen-sources by ectomycor-rhizal fungi in pure culture and in symbiosis with Pinus contortaDougl ex loud. New Phytologist 120, 105–115.
Gange A. (2000) Arbuscular mycorrhizal fungi, Collembola andplant growth. Trends in Ecology and Evolution 15, 369–372.
Hilger A.B., Thomas K.I. & Krause H.H. (1986) The effects ofseveral buffers on growth and phosphorus nutrition of selectedectomycorrhizal fungi. Soil Biology and Biochemistry 18, 61–67.
Hilger A.B. & Krause H.H. (1989) Growth characteristics ofLaccaria laccata and Paxillus involutus in liquid culture mediawith inorganic and organic phosphorus sources. Canadian Jour-nal of Botany 67, 1782–1789.
Kieliszewska-Rokicka B. (1992) Effect of nitrogen level on acid-phosphatase-activity of 8 isolates of ectomycorrhizal fungusPaxillus involutus cultured in vitro. Plant and Soil 139, 229–238.
Klironomos J.N. & Hart M.M. (2001) Animal nitrogen swap forplant carbon. Nature 410, 651–652.
Leake J.R. & Read D.J. (1997) Mycorrhizal fungi in terrestrialhabitats. In The Mycota IV: Environmental and Microbial Rela-

1226 J. Perez-Moreno & D. J. Read
© 2001 Blackwell Science Ltd, Plant, Cell and Environment, 24, 1219–1226
tionship (eds D.T. Wicklow & B. Söderström ), pp. 281–301.Springer-Verlag, Berlin.
Lee D.L. & Atkinson H.J. (1976) Physiology of Nematodes. Mac-millan Press, London.
Li C.Y. & Hung L.L. (1987) Nitrogen- fixing (acetylene-reduc-ing) bacteria associated with ectomycorrhiza of Douglas fir.Plant and Soil 98, 425–428.
Li C.Y., Massicote H.B. & Moore L.V.H. (1992) Nitrogen-fixingBacillus sp. associated with Douglas-fir tuberculate ectomycor-rhizae. Plant and Soil 140, 35–40.
Norkrans B. (1953) The effect of glutamic acid, aspartic acid, andrelated compounds on the growth of certain Tricholoma species.Physiologia Plantarum 6, 584–593.
Nurmiaho-Lassila E.-L., Timonen S., Haahtela K. & Sen R.(1997) Bacterial colonization patterns of intact Pinus sylvestrismycorrhizospheres in dry pine forest soil: an electron micros-copy study. Canadian Journal of Microbiology 43, 1017–1035.
Perez-Moreno J. & Read D.J. (2000) Mobilization and transferof nutrients from litter to tree seedlings via the vegetativemycelium of ectomycorrhizal plants. New Phytologist 145, 301–309.
Perez-Moreno J. & Read D.J. (2001) Exploitation of pollen bymycorrhizal mycelial systems with special reference to nutrientrecycling in boreal forests. Proceedings of the Royal Society ofLondon B 268, 1329–1335.
Petersen H. (1982) Structure and size of animal populations.Oikos 39, 306–329.
Rayner A.D.M. (1996) Has chaos theory a place in environmentalmycology? In Fungi and Environmental change (eds J.C. Frank-land, N. Magan & G.M. Gadd), pp. 317–341. Cambridge Uni-versity Press, Cambridge.
Read D.J. (1992) The mycorrhizal mycelium. In Mycorrhizal
Mycelium: an Integrative Plant Fungal Process (ed. M.J. Allen),pp. 102–133. Chapman & Hall, New York.
Schenck S., Chase T. Jr, Rosenzweig W.D. & Pramer D. (1980)Collagenase production by nematode trapping fungi. AppliedEnvironmental Microbiology 40, 567–570.
Sen R. (2000) Budgeting for the wood-wide web. New Phytologist145, 161–163.
Söderström B. & Read D.J. (1987) Respiratory activity of intactand excised ectomycorrhizal mycelial systems growing in unster-ilised soil. Soil Biology and Biochemistry 19, 231–236.
Timonen S., Jørgensen K.S., Haahtela K. & Sen R. (1998) Bac-terial community structure at defined locations of Pinus sylves-tris–Suillus bovinus and Pinus sylvestris–Paxillus involutusmycorrhizospheres in dry pine forest humus and nursery peat.Canadian Journal of Microbiology 44, 499–513.
Toth R., Toth D. & Nordbring-Hertz B. (1980) Cytochemicallocalization of acid phosphatase in the nematode-trapping fun-gus Arthrobotrys oligospora. Mycologia 72, 813–817.
Tsuneda A. & Thorn R.G. (1995) Interactions of wood decayfungi with other microorganisms, with emphasis on the degra-dation of cell-walls. Canadian Journal of Botany 73 (suppl.),1325–1333.
Tunlid A., Rosén S., Ek B. & Rask L. (1994) Purification andcharacterization of an extracellular serine protease from thenematode-trapping fungus Arthrobotrys oligospora. Microbiol-ogy 140, 1687–1695.
Verhoef H.A. & Brussaard L. (1990) Decomposition and nitro-gen mineralisation in natural and agro-ecosystems, the contribu-tion of soil animals. Biogeochemistry 11, 175–211.
Received 20 April 2001; received in revised form 28 June 2001; accepted for publication 28 June 2001




















