
NOVAS OCORRÊNCIAS DE CERAMIACEAE EPÍFITAS PARA A PLATAFORMA CONTINENTAL DOS ESTADOS DO ESPÍRITO...
24
NOVAS OCORRÊNCIAS DE CERAMIACEAE EPÍFITAS NOVAS OCORRÊNCIAS DE CERAMIACEAE EPÍFITAS NOVAS OCORRÊNCIAS DE CERAMIACEAE EPÍFITAS NOVAS OCORRÊNCIAS DE CERAMIACEAE EPÍFITAS NOVAS OCORRÊNCIAS DE CERAMIACEAE EPÍFITAS PARA A PLATAFORMA CONTINENTAL DOS ESTADOS PARA A PLATAFORMA CONTINENTAL DOS ESTADOS PARA A PLATAFORMA CONTINENTAL DOS ESTADOS PARA A PLATAFORMA CONTINENTAL DOS ESTADOS PARA A PLATAFORMA CONTINENTAL DOS ESTADOS DO ESPÍRITO SANTO E RIO DE JANEIRO, BRASIL DO ESPÍRITO SANTO E RIO DE JANEIRO, BRASIL DO ESPÍRITO SANTO E RIO DE JANEIRO, BRASIL DO ESPÍRITO SANTO E RIO DE JANEIRO, BRASIL DO ESPÍRITO SANTO E RIO DE JANEIRO, BRASIL Juliana Torres, Lísia Mônica de Souza Gestinari & Yocie Yoneshigue-Valentin RESUMO: A implementação do Programa REVIZEE permitiu avançar, significativamente, com os conhecimentos sobre as macroalgas da Plataforma Continental do Brasil. Para o presente estudo, foram analisadas amostras coletadas, através de draga retangular, em cinco estações nos estados do Espírito Santo e Rio de Janeiro, em profundidades que variaram entre 36 e 124 m (20,352ºS - 22,883ºS; 037,620ºW - 041,154ºW). No laboratório, constatou-se a presença das Ceramiaceae epífitas nas seguintes espécies de algas hospedeiras: Dictyopteris jamaicensis W. R. Taylor, 1960, Zonaria tournefortii (J. V. Lamour.) Mont., 1846, Pterocladiella capillacea (S. G. Gmel.) Santel. & Hommers., 1997, Cryptonemia seminervis (C. Agardh) J. Agardh, 1846 e Plocamium brasiliense (Grev. in J. St.-Hil.) M. Howe & W. R. Taylor, 1931. Foram registradas 11 espécies de Ceramiaceae: Aglaothamnion cf. priceanum Maggs, Guiry & Rueness, 1991, Antithamnionella atlantica (E. C. Oliveira) C. W. Schneid., 1984, A. bermudica C. W. Schneid. in Schneider & Searles, 1997, A. breviramosa (Dawson) Wollaston in Womersley & Bailey, 1970, Callithamniella tingitana (Schousb. ex Bornet) Feldm.-Maz., 1940, Compsothamnion thuyoides (Sm.) F. Schmitz, 1897, Diplothamnion tetrastichum A. B. Joly & Yamaguishi in Joly et al., 1965b, Pleonosporium boergesenii (A. B. Joly) R. E. Norris, 1985, Pterothamnion heteromorphum (J. Agardh) Athanas. & Kraft, 1994, Spermothamnion repens (Dillwyn) Rosenv., 1923-1924 e Tiffaniella snyderiae? (Farl.) I. A. Abbott., 1971. Do total das 11 espécies encontradas, sete ocorreram no Estado do Espírito Santo (Aglaothamnion cf. priceanum, Antithamnionella atlantica, A. bermudica, A. breviramosa, Pleonosporium boergesenii , Spermothamnion repens e Tiffaniella snyderiae?) e quatro ocorreram no Estado do Rio de Janeiro (Callithamniella tingitana, Compsothamnion thuyoides , Diplothamnion tetrastichum e Pterothamnion heteromorphum). Algumas dessas espécies ocorrem comumente em mares do Hemisfério Norte e constituem novas citações para as águas do Atlântico Meridional (Aglaothamnion cf. priceanum, A. bermudica, Pterothamnion heteromorphum e Tiffaniella snyderiae?). PALAVRAS-CHAVE: Ceramiaceae. Águas profundas. Espírito Santo. Rio de Janeiro. Brasil. ABSTRACT: New records of epiphytes Ceramiaceae from the Continental Shelf of Espírito Santo and Rio de Janeiro States, Brazil. The implementation of the REVIZEE Program significantly allowed advancing in the knowledge of the seaweeds from the Continental Shelf of Brazil. For the present study, samples collected by a rectangular dredge in five stations of Espírito Santo and Rio de Janeiro States, at depths of 36 to 124 m (20,352ºS - 22,883ºS; 037,620ºW - 041,154ºW) were analyzed. In the laboratory, the epiphytes Ceramiaceae were observed in the following host species: Dictyopteris jamaicensis W. R. Taylor, 1960, Zonaria tournefortii (J. V. Lamour.) Mont., 1846, Pterocladiella capillacea (S. G. Gmel.) Santel. & Hommers., 1997, Cryptonemia seminervis (C. Agardh) J. Agardh, 1846 and Plocamium brasiliense (Grev. in J. St.-Hil.) M. Howe & W. R. Taylor, 1931. Eleven Ceramiaceae species were recorded: Aglaothamnion cf. priceanum Maggs, Guiry & Rueness, 1991, Antithamnionella atlantica (E. C. Oliveira) C. W. Schneid., 1984, A. bermudica C. W. Schneid. in Schneider & Searles, 1997, A. breviramosa (Dawson) Wollaston in Womersley & Bailey, 1970, Callithamniella tingitana (Schousb. ex Bornet)
Transcript of NOVAS OCORRÊNCIAS DE CERAMIACEAE EPÍFITAS PARA A PLATAFORMA CONTINENTAL DOS ESTADOS DO ESPÍRITO...

NOVAS OCORRÊNCIAS DE CERAMIACEAE EPÍFITASNOVAS OCORRÊNCIAS DE CERAMIACEAE EPÍFITASNOVAS OCORRÊNCIAS DE CERAMIACEAE EPÍFITASNOVAS OCORRÊNCIAS DE CERAMIACEAE EPÍFITASNOVAS OCORRÊNCIAS DE CERAMIACEAE EPÍFITAS
PARA A PLATAFORMA CONTINENTAL DOS ESTADOSPARA A PLATAFORMA CONTINENTAL DOS ESTADOSPARA A PLATAFORMA CONTINENTAL DOS ESTADOSPARA A PLATAFORMA CONTINENTAL DOS ESTADOSPARA A PLATAFORMA CONTINENTAL DOS ESTADOS
DO ESPÍRITO SANTO E RIO DE JANEIRO, BRASILDO ESPÍRITO SANTO E RIO DE JANEIRO, BRASILDO ESPÍRITO SANTO E RIO DE JANEIRO, BRASILDO ESPÍRITO SANTO E RIO DE JANEIRO, BRASILDO ESPÍRITO SANTO E RIO DE JANEIRO, BRASIL
Juliana Torres, Lísia Mônica de Souza Gestinari
& Yocie Yoneshigue-Valentin
RESUMO: A implementação do Programa REVIZEE permitiu avançar, significativamente,
com os conhecimentos sobre as macroalgas da Plataforma Continental do Brasil. Para o
presente estudo, foram analisadas amostras coletadas, através de draga retangular, em
cinco estações nos estados do Espírito Santo e Rio de Janeiro, em profundidades que
variaram entre 36 e 124 m (20,352ºS - 22,883ºS; 037,620ºW - 041,154ºW). No
laboratório, constatou-se a presença das Ceramiaceae epífitas nas seguintes espécies de
algas hospedeiras: Dictyopteris jamaicensis W. R. Taylor, 1960, Zonaria tournefortii (J.
V. Lamour.) Mont., 1846, Pterocladiella capillacea (S. G. Gmel.) Santel. & Hommers.,
1997, Cryptonemia seminervis (C. Agardh) J. Agardh, 1846 e Plocamium brasiliense
(Grev. in J. St.-Hil.) M. Howe & W. R. Taylor, 1931. Foram registradas 11 espécies de
Ceramiaceae: Aglaothamnion cf. priceanum Maggs, Guiry & Rueness, 1991,
Antithamnionella atlantica (E. C. Oliveira) C. W. Schneid., 1984, A. bermudica C. W.
Schneid. in Schneider & Searles, 1997, A. breviramosa (Dawson) Wollaston in Womersley
& Bailey, 1970, Callithamniella tingitana (Schousb. ex Bornet) Feldm.-Maz., 1940,
Compsothamnion thuyoides (Sm.) F. Schmitz, 1897, Diplothamnion tetrastichum A. B.
Joly & Yamaguishi in Joly et al., 1965b, Pleonosporium boergesenii (A. B. Joly) R. E.
Norris, 1985, Pterothamnion heteromorphum (J. Agardh) Athanas. & Kraft, 1994,
Spermothamnion repens (Dillwyn) Rosenv., 1923-1924 e Tiffaniella snyderiae? (Farl.) I.
A. Abbott., 1971. Do total das 11 espécies encontradas, sete ocorreram no Estado do
Espírito Santo (Aglaothamnion cf. priceanum, Antithamnionella atlantica, A. bermudica,
A. breviramosa, Pleonosporium boergesenii, Spermothamnion repens e Tiffaniella
snyderiae?) e quatro ocorreram no Estado do Rio de Janeiro (Callithamniella tingitana,
Compsothamnion thuyoides, Diplothamnion tetrastichum e Pterothamnion
heteromorphum). Algumas dessas espécies ocorrem comumente em mares do Hemisfério
Norte e constituem novas citações para as águas do Atlântico Meridional (Aglaothamnion
cf. priceanum, A. bermudica, Pterothamnion heteromorphum e Tiffaniella snyderiae?).
PALAVRAS-CHAVE: Ceramiaceae. Águas profundas. Espírito Santo. Rio de Janeiro. Brasil.
ABSTRACT: New records of epiphytes Ceramiaceae from the Continental Shelf of Espírito
Santo and Rio de Janeiro States, Brazil.
The implementation of the REVIZEE Program significantly allowed advancing in the
knowledge of the seaweeds from the Continental Shelf of Brazil. For the present
study, samples collected by a rectangular dredge in five stations of Espírito Santo and
Rio de Janeiro States, at depths of 36 to 124 m (20,352ºS - 22,883ºS; 037,620ºW -
041,154ºW) were analyzed. In the laboratory, the epiphytes Ceramiaceae were
observed in the following host species: Dictyopteris jamaicensis W. R. Taylor, 1960,
Zonaria tournefortii (J. V. Lamour.) Mont., 1846, Pterocladiella capillacea (S. G. Gmel.)
Santel. & Hommers., 1997, Cryptonemia seminervis (C. Agardh) J. Agardh, 1846 and
Plocamium brasiliense (Grev. in J. St.-Hil.) M. Howe & W. R. Taylor, 1931. Eleven
Ceramiaceae species were recorded: Aglaothamnion cf. priceanum Maggs, Guiry &
Rueness, 1991, Antithamnionella atlantica (E. C. Oliveira) C. W. Schneid., 1984, A.
bermudica C. W. Schneid. in Schneider & Searles, 1997, A. breviramosa (Dawson)
Wollaston in Womersley & Bailey, 1970, Callithamniella tingitana (Schousb. ex Bornet)

8080808080 J. Torres, L. M. S. Gestinari & Y. Yoneshigue-Valentin
Feldm.-Maz., 1940, Compsothamnion thuyoides (Sm.) F. Schmitz, 1897, Diplothamnion
tetrastichum A. B. Joly & Yamaguishi in Joly et al., 1965b, Pleonosporium boergesenii
(A. B. Joly) R. E. Norris, 1985, Pterothamnion heteromorphum (J. Agardh) Athanas. &
Kraft, 1994, Spermothamnion repens (Dillwyn) Rosenv., 1923-1924 and Tiffaniella
snyderiae? (Farl.) I. A. Abbott, 1971. Seven of these species were found in Espírito
Santo State (Aglaothamnion cf. priceanum, Antithamnionella atlantica, A. bermudica,
A. breviramosa, Pleonosporium boergesenii, Spermothamnion repens and Tiffaniella
snyderiae?) and four in the Rio de Janeiro State (Callithamniella tingitana,
Compsothamnion thuyoides, Diplothamnion tetrastichum and Pterothamnion
heteromorphum). Some of these 11species are usually found in the North Hemisphere
seas and represent new records for the Southern Atlantic waters (Aglaothamnion cf.
priceanum, A. bermudica, Pterothamnion heteromorphum and Tiffaniella snyderiae?).
KEY WORDS: Ceramiaceae. Deep-water. Espírito Santo. Rio de Janeiro. Brazil.
IIIIINTRODUÇÃONTRODUÇÃONTRODUÇÃONTRODUÇÃONTRODUÇÃO
A presença da Água Central do Atlântico Sul (ACAS) na região da Plataforma
Continental dos estados do Espírito Santo e Rio de Janeiro (Valentin & Monteiro-
Ribas, 1993) é considerada um fator determinante na estrutura das comunidades
algáceas ali encontradas, em razão das baixas temperaturas e altos teores em
nutrientes (Yoneshigue-Valentin et al., 2006). Em contraposição, a presença de
águas quentes, e com baixos teores em nutrientes, provenientes da Corrente do
Brasil também estabelece um fator importante para as comunidades de algas de
afinidades tropicais.
Nesses Estados, são poucos os trabalhos publicados sobre as macroalgas de ecossistemas
profundos (Howe & Taylor, 1931; Taylor, 1931; Joly & Yoneshigue-Braga, 1966; Joly &
Oliveira Filho, 1967; Joly et al., 1978; Teixeira et al., 1985; Marcot-Coqueuniot, 1988).
Entretanto, estudos recentes têm revelado a existência de uma diversidade de
populações de macroalgas nos ecossistemas profundos, ainda pouco conhecidas devido
às dificuldades de coleta (Haroun et al., 1993; Yoneshigue-Valentin et al.,1995; Bravin
et al., 1999; Yoneshigue-Valentin et al., 2006).
Com a implementação do Programa REVIZEE, que visa inventariar os recursos vivos
da Zona Econômica Exclusiva, foi possível avançar, significativamente, com os
conhecimentos sobre as algas multicelulares da Plataforma Continental do Brasil,
principalmente aquelas com talos de pequeno porte e de consistência delicada,
crescendo sobre as frondes de outras macroalgas. Desta forma, o conhecimento da
composição específica das comunidades profundas torna o estudo de alta relevância
no contexto científico nacional e internacional pela descoberta de espécies ainda
não referidas para o litoral brasileiro ampliando, assim, sua área de distribuição
geográfica.
Tendo em vista os poucos estudos realizados sobre a Família Ceramiaceae em
ecossistemas profundos, o presente trabalho tem como objetivo contribuir para o
melhor conhecimento da composição e distribuição dos representantes dessa Família
não só para os estados do Espírito Santo e Rio de Janeiro como, também, para o
litoral brasileiro.
8181818181Novas ocorrências de Ceramiaceae epífitas para a plataforma continental do ES e do RJ, Brasil
MMMMMATERIALATERIALATERIALATERIALATERIAL EEEEE M M M M MÉTODOSÉTODOSÉTODOSÉTODOSÉTODOS
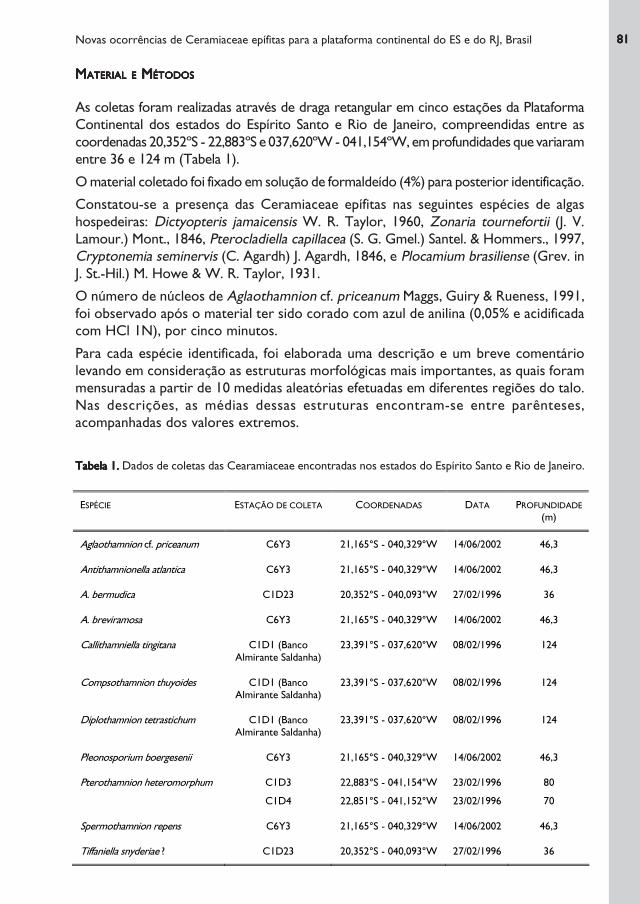
As coletas foram realizadas através de draga retangular em cinco estações da Plataforma
Continental dos estados do Espírito Santo e Rio de Janeiro, compreendidas entre as
coordenadas 20,352ºS - 22,883ºS e 037,620ºW - 041,154ºW, em profundidades que variaram
entre 36 e 124 m (Tabela 1).
O material coletado foi fixado em solução de formaldeído (4%) para posterior identificação.
Constatou-se a presença das Ceramiaceae epífitas nas seguintes espécies de algas
hospedeiras: Dictyopteris jamaicensis W. R. Taylor, 1960, Zonaria tournefortii (J. V.
Lamour.) Mont., 1846, Pterocladiella capillacea (S. G. Gmel.) Santel. & Hommers., 1997,
Cryptonemia seminervis (C. Agardh) J. Agardh, 1846, e Plocamium brasiliense (Grev. in
J. St.-Hil.) M. Howe & W. R. Taylor, 1931.
O número de núcleos de Aglaothamnion cf. priceanum Maggs, Guiry & Rueness, 1991,
foi observado após o material ter sido corado com azul de anilina (0,05% e acidificada
com HCl 1N), por cinco minutos.
Para cada espécie identificada, foi elaborada uma descrição e um breve comentário
levando em consideração as estruturas morfológicas mais importantes, as quais foram
mensuradas a partir de 10 medidas aleatórias efetuadas em diferentes regiões do talo.
Nas descrições, as médias dessas estruturas encontram-se entre parênteses,
acompanhadas dos valores extremos.
Tabela 1.Tabela 1.Tabela 1.Tabela 1.Tabela 1. Dados de coletas das Cearamiaceae encontradas nos estados do Espírito Santo e Rio de Janeiro.
ESPÉCIE ESTAÇÃO DE COLETA COORDENADAS DATA PROFUNDIDADE
(m)
Aglaothamnion cf. priceanum C6Y3 21,165°S - 040,329°W 14/06/2002 46,3
Antithamnionella atlantica C6Y3 21,165°S - 040,329°W 14/06/2002 46,3
A. bermudica C1D23 20,352°S - 040,093°W 27/02/1996 36
A. breviramosa C6Y3 21,165°S - 040,329°W 14/06/2002 46,3
Callithamniella tingitana C1D1 (Banco
Almirante Saldanha)
23,391°S - 037,620°W 08/02/1996 124
Compsothamnion thuyoides C1D1 (Banco
Almirante Saldanha)
23,391°S - 037,620°W 08/02/1996 124
Diplothamnion tetrastichum C1D1 (Banco
Almirante Saldanha)
23,391°S - 037,620°W 08/02/1996 124
Pleonosporium boergesenii C6Y3 21,165°S - 040,329°W 14/06/2002 46,3
Pterothamnion heteromorphum C1D3
C1D4
22,883°S - 041,154°W
22,851°S - 041,152°W
23/02/1996
23/02/1996
80
70
Spermothamnion repens C6Y3 21,165°S - 040,329°W 14/06/2002 46,3
Tiffaniella snyderiae ? C1D23 20,352°S - 040,093°W 27/02/1996 36

8282828282 J. Torres, L. M. S. Gestinari & Y. Yoneshigue-Valentin
Os táxons são apresentados de acordo com a revisão taxonômica de Wynne (2005).
Tanto o hábito geral quanto as estruturas mais importantes foram ilustrados por meio
de câmara clara acoplada a microscópio ótico.
As lâminas confeccionadas encontram-se depositadas no Herbário da UFRJ (RFA).
As novas ocorrências para o Espírito Santo são precedidas por um asterisco (*); aquelas
para o Rio de Janeiro por dois asteriscos (**) e aquelas para o Atlântico Meridional por
um sinal de adição (+).
RRRRRESULTADOSESULTADOSESULTADOSESULTADOSESULTADOS
Foram registradas 11 espécies de Ceramiaceae: Aglaothamnion cf. priceanum
Maggs, Guiry & Rueness, 1991, Antithamnionella atlantica (E. C. Oliveira) C. W.
Schneid., 1984, A. bermudica C. W. Schneid. in Schneider & Searles, 1997, A.
breviramosa (Dawson) Wollaston in Womersley & Bailey, 1970, Callithamniella
tingitana (Schousb. ex Bornet) Feldm.-Maz., 1940, Compsothamnion thuyoides
(Sm.) F. Schmitz, 1897, Diplothamnion tetrastichum A. B. Joly & Yamaguishi in
Joly et al., 1965b, Pleonosporium boergesenii (A. B. Joly) R. E. Norris, 1985,
Pterothamnion heteromorphum ( J . Agardh) Athanas . & Kraf t , 1994,
Spermothamnion repens (Dillwyn) Rosenv., 1923-24, e Tiffaniella snyderiae? (Farl.)
I. A. Abbott, 1971.
*+ Aglaothamnion cf. priceanum Maggs, Guiry & Rueness
(Figuras 1-2)
Brit. Phycol. J. 26: 343. 1991.
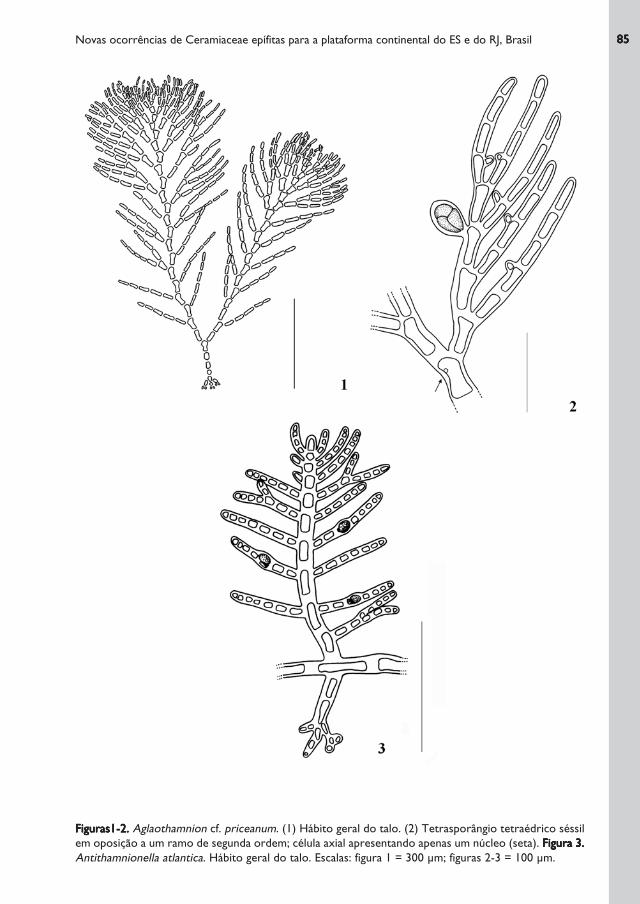
Talo formado por pequenos grupos de eixos eretos aderidos ao substrato por rizóides
filamentosos que terminam em apressórios discóides. Eixos eretos medindo 0,1-
0,2 cm de altura, complanados, com um único eixo principal linear ou irregularmente
flabelado, de cor rósea, delicado, mas não flácido. Células apicais medindo 7-(8)-13
μm de largura; células axiais medindo 20-(25)-33 μm de diâmetro, sendo mais longas
que largas na porção mediana do talo. Células dos eixos principais e laterais
apresentando apenas um núcleo. Ramos laterais de primeira ordem seguindo um
padrão de ramificação regularmente alternado-dístico. Ramos laterais de segunda e
terceira ordem seguindo um padrão irregular de ramificação. Presença de
tetrasporângios tetraédricos sésseis, medindo 57-(67)-75 x 38-(49)-57 μm, em
oposição a um dos ramos de segunda ordem, do lado adaxial.
Habitat: Crescendo sobre talos de Plocamium brasiliense (Tabela1).
Material Examinado: BRASIL, ESPÍRITO SANTO, estação C6Y3, 21,165°S - 040,329°W,
RFA 33443, RFA 33444, C. F. D. Gurgel & A. Taouil s.n. 14/VI/2002.
Comentários: Feldmann-Mazoyer (1940) propôs o gênero Aglaothamnion Feldmann-
Mazoyer, 1940, caracterizando-o por apresentar células vegetativas uninucleadas, ramos
carpogoniais em ziguezague ou em forma de “u” e gonimolobos lobados. Porém, alguns
autores como Dixon & Price (1981) e Womersley (1998) não reconhecem

8383838383Novas ocorrências de Ceramiaceae epífitas para a plataforma continental do ES e do RJ, Brasil
completamente o gênero Aglaothamnion por notarem que todas as características
listadas por Feldmann-Mazoyer (1940) também haviam sido observadas em espécies
de Callithamnion Lyngb., 1819, e são variáveis dentro de Aglaothamnion. Entretanto,
McIvor et al. (2002), através de análises filogenéticas, afirmaram que transferir todo
o clado Callithamnion / Aglaothamnion para um único gênero (Callithamnion) não
seria a melhor alternativa, uma vez que o nível de divergência nas seqüências é alto.
A presença de células uninucleadas em todas as frondes do material estudado confirma
o gênero Aglaothamnion (Aponte et al., 1994). Além disso, as plantas analisadas
neste estudo correspondem, em parte, à descrição de Maggs et al. (1991) para A.
priceanum, uma vez que algumas diferenças são encontradas nos exemplares
estudados: os ramos laterais de segunda e terceira ordem não seguem o mesmo
padrão de ramificação dos ramos de primeira ordem; as plantas não apresentam as
duas primeiras células dos ramos laterais de primeira ordem portando um râmulo
adaxial cada e, a terceira, um râmulo abaxial.
*Antithamnionella atlantica (E. C. Oliveira) C. W. Schneid.
(Figura 3)
Phycologia 23: 455. 1984.
Basiônimo: Antithamnion atlanticum E. C. Oliveira, Bol. Fac. Fil., Ciênc. Letr., Univ. S.
Paulo, Série Botânica 26: 37. 1969.
Talo delicado, medindo 482-(489)-495 μm de altura, de cor rósea, formado por eixo
prostrado, fixo por rizóides pluricelulares, a partir do qual nascem ramos eretos,
opostos, de crescimento determinado ou indeterminado, podendo cada par ser
formado por dois râmulos ou por um ramo de crescimento indeterminado. Eixo
prostrado com células medindo 40-(51)-59 x 7-(12)-13 μm. Ramos eretos medindo
73-(204)-495 μm de altura, sendo formados por um eixo central, dissecado por
râmulos opostos dois a dois, com disposição dística, saindo da porção superior das
células dos eixos principais. Râmulos laterais recurvados para cima, gradualmente
menores em direção ao ápice, medindo os mais inferiores 7-(12)-13 μm de diâmetro,
sendo simples ou ramificados 1 - 3 vezes; células basais com mesmo tamanho das
células adjacentes. Células glandulares elípticas localizadas sobre a terceira célula, da
base para o ápice, dos râmulos ramificados ou simples.
Habitat: Crescendo sobre talos de Zonaria tournefortii e Cryptonemia seminervis
(Tabela 1).
Material Examinado: BRASIL, ESPÍRITO SANTO, estação C6Y3, 21,165°S - 040,329°W,
RFA 33445, C. F. D. Gurgel & A. Taouil s.n., 14/VI/2002.
Comentários: Wollaston (1968, 1971a, b) separou Antithamnion Nägeli, 1847, e gêneros
intimamente relacionados utilizando caracteres como: formação dos ramos, posição
da célula glandular e forma e divisão do tetrasporângio. Posteriormente, esta autora
retomou o gênero Antithamnionella Lyle, 1922, que, segundo ela, distingue-se de
Antithamnion pelos seguintes caracteres: forma recurvada do ápice do eixo ereto
principal, desenvolvimento em seqüência irregular dos verticilos de ramos curtos,
verticilos de 1 - 4 ramos curtos, posição lateral de células glandulares sobre uma célula
do ramo curto (quando encontradas na maioria das espécies de Antithamnionella),

8484848484 J. Torres, L. M. S. Gestinari & Y. Yoneshigue-Valentin
tetrasporângios quase esféricos, com divisão freqüentemente tetraédrica,
espermatângios encontrados mais nas laterais dos ramos que nos ápices e somente 1-
3 procarpos produzidos por ramo apical, cada um presente em um ramo determinado
reduzido. Com base nestes caracteres, Schneider (1984) transferiu Antithamnion
atlanticum E. C. Oliveira, 1969, originalmente descrita para o Estado do Espírito Santo,
para a espécie Antithamnionella atlantica. Segundo Oliveira Filho (1969), esta espécie
difere de todas as outras com râmulos opostos dísticos citadas na literatura
aproximando-se apenas de Antithamnion basisporum Tokida & Inaba, 1950, da qual
pode ser separada por apresentar esporângios tetraédricos e apenas um por râmulo;
râmulos com apenas 2 - 7 células e freqüentemente ramificados, enquanto A. basisporum
apresenta tetrasporângios cruciados e 1 - 2 por râmulo; râmulos com 7 - 16 células e
raramente ramificados. O material estudado concorda com as descrições de Oliveira
Filho (1969); Ugadim et al. (1986); Schneider & Searles (1991) e Athanasiadis (1996).
*+Antithamnionella bermudica C. W. Schneid. in Schneider & Searles
(Figuras 4-5)
Phycologia 36: 15. 1997.
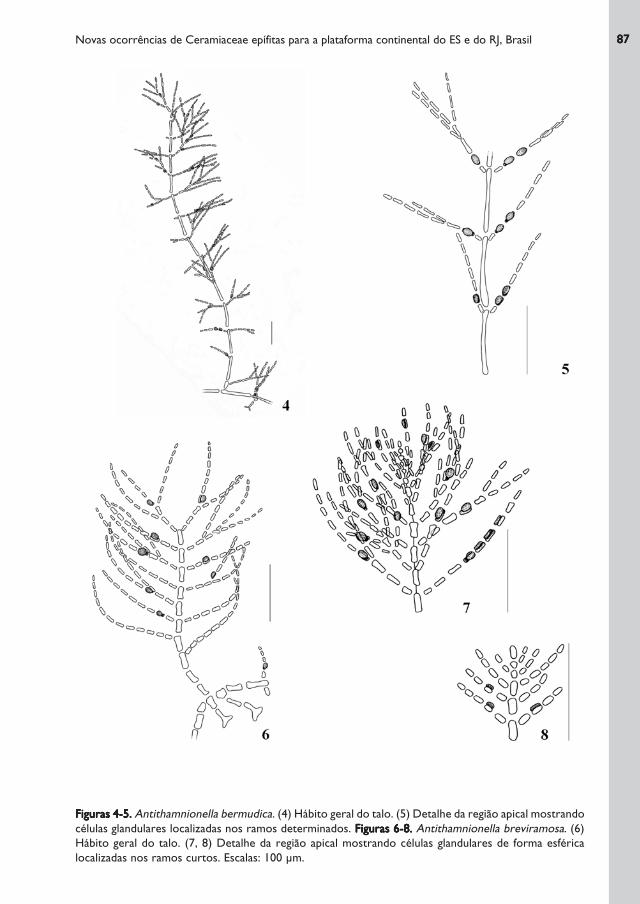
Talo medindo até 2 mm de altura, de cor rósea, formado por eixo prostrado, fixo por
rizóides pluricelulares com, ou sem, apressórios digitados; rizóides medindo 38-(63)-75
μm de diâmetro, originados de células periaxiais (células basais das ramificações), substituindo
um ramo lateral determinado oposto a outro, ou a um ramo lateral indeterminado; células
medianas dos eixos indeterminados medindo 113-(128)-189 x 19 μm; eixos indeterminados
ramificados disticamente, a partir de cada célula axial, sendo cada par constituido por dois
ramos laterais determinados ou, raramente, por um ramo lateral determinado e outro
indeterminado; ápices dos ramos indeterminados formados por ramos determinados
parcialmente desenvolvidos; ramos determinados simples, ou unilateralmente ramificados
uma ou duas vezes, com 5 - 8 células medindo 19 μm de diâmetro, formando ângulos de 65
- 90° com as porções superiores das células axiais, ou de 30 - 45º com o eixo prostrado.
Células glandulares localizadas sobre a segunda e/ou terceira célula, da base para o ápice,
dos ramos determinados e râmulos, raramente na periaxial ou na quarta célula.
Habitat: Crescendo sobre talos de Cryptonemia seminervis (Tabela1).
Material Examinado: BRASIL, ESPÍRITO SANTO, estação C1D23, 20,352°S - 040,093°W,
RFA 33446, C. F. D. Gurgel & A. Taouil s.n., 27/II/1996.
Comentários: Nenhuma das outras espécies conhecidas de Antithamnionella com ramos
determinados curtos (menos de 10 células) apresenta ramos determinados dísticos
estritamente opostos, ou um padrão de ramificação dos ramos determinados semelhante
à A. bermudica (Schneider & Searles, 1997). Além de A. bermudica, A. ternifolia (Hook.
& Harv.) Lyle, 1922, e Pterothamnion nodiferum (J. Agardh) Athanas. & Kraft, 1994,
ocasionalmente apresentam uma célula glandular substituindo um râmulo primário
(Wollaston, 1968). O material estudado concorda com a descrição original de Schneider
& Searles (1997), embora células glandulares tenham sido encontradas sobre a segunda
e terceira células de um mesmo ramo, como foi verificado, também, por Feldmann-
Mazoyer (1940), em Pterothamnion plumula (J. Ellis) Nägeli in Nägeli & Cramer, 1882
(como Antithamnion plumula J. Ellis, 1863).
8585858585Novas ocorrências de Ceramiaceae epífitas para a plataforma continental do ES e do RJ, Brasil
Figuras1-2.Figuras1-2.Figuras1-2.Figuras1-2.Figuras1-2. Aglaothamnion cf. priceanum. (1) Hábito geral do talo. (2) Tetrasporângio tetraédrico séssil
em oposição a um ramo de segunda ordem; célula axial apresentando apenas um núcleo (seta). Figura 3.Figura 3.Figura 3.Figura 3.Figura 3.
Antithamnionella atlantica. Hábito geral do talo. Escalas: figura 1 = 300 μm; figuras 2-3 = 100 μm.

8686868686 J. Torres, L. M. S. Gestinari & Y. Yoneshigue-Valentin
*Antithamnionella breviramosa (Dawson) Wollaston in Womersley & Bailey
(Figuras 6-8)
Phil. Trans. R. Soc. 259b: 322. 1970.
Basiônimo: Antithamnion breviramosus Dawson, Allan Hancock Foundation Publications,
Occasional Paper 8: 14. 1949.
Talo delicado, medindo até 1mm de altura, de cor rósea, formado por filamentos rastejantes
longos apresentando 18-(21)-22 μm de diâmetro, com células medindo 59-(72,5)-79 x 7-
(12)-13 μm, fixo ao substrato por rizóides unicelulares que medem 41-(60)-74 x 13-(16)-
19 μm e nascem das pequenas células basais dos ramos laterais curtos daqueles filamentos,
terminando por discos lobados. Ramos eretos com 7-20 células axiais medindo 20-(40)-66
x 7-(12)-13 μm. Destes ramos de crescimento indeterminado saem, na porção distal de
cada célula, 1 ou 2 ramos curtos, dispostos verticiladamente e de maneira espiralada, atingindo
73-(140)-211 μm de comprimento, sendo os maiores, em geral, com 1 - 2 divisões; ramos
curtos medindo 7-(8)-13 μm de diâmetro, com 4 - 12 células que medem 13-(16)-20 x 9-
(10)-12 μm. Estes ramos são constituídos por râmulos com 6-(7)-9 μm de diâmetro. Células
glandulares refringentes abundantes, de forma esférica, medindo 13-(18)-20 μm de diâmetro,
localizadas, em geral, sobre a célula basal da primeira bifurcação, às vezes nas células basais
das outras bifurcações e, mais raramente, em outras células dos râmulos. Cada ramo curto
de primeira ordem transporta, em geral, 1 - 2 células glandulares.
Habitat: Crescendo sobre talos de Cryptonemia seminervis (Tabela1).
Material examinado: BRASIL, ESPÍRITO SANTO, estação C6Y3, 21,165°S - 040,329°W,
RFA 33447, C. F. D. Gurgel & A. Taouil s.n., 14-VI-2002.
Comentários: Nossos exemplares apresentam medidas inferiores para diâmetro
dos filamentos rastejantes e suas células rizoidais, células do eixo indeterminado e
dos ramos curtos, quando comparados com os dos autores brasileiros [Joly (1965),
como Antithamnion ternatum A. B. Joly & Cord., 1965a, e Antithamnion tristicum
A. B. Joly & Yamaguishi, 1963a, que, mais tarde, foram colocadas em sinonímia -
Antithamnion tristicum - por Oliveira Filho (1969); Yoneshigue (1985), como
Antithamnionella elegans (Berthold) Boudour. & Verlaque, 1976, e Ugadim et al. (1986),
Horta (2000), Torres et al. (2004), como Antithamnionella breviramosa (Dawson)
Wollaston in Womersley & Bailey, 1970] e estrangeiros, como Börgesen (1930) e
Feldmann-Mazoyer (1940), como Antithamnion elegans Berthold, 1882, Dawson
(1949), como Antithamnion breviramosus Dawson, 1949, Wollaston (1968, 1971a)
e Womersley & Bailey (1970), como Antithamnionella breviramosa, Schneider (1984),
como Antithamnion tristicum e Yoneshigue (1985), como Antithamnionella elegans.
Entretanto, as células glandulares apresentam medidas maiores que as dos autores
citados anteriormente. Supõe-se que essas medidas dos talos do presente estudo
sejam diferentes dos dados da literatura, provavelmente pelo fato de terem sido
encontrados à uma maior profundidade (46,3 m – Tabela 1). Assim, preferiu-se basear
a identificação da espécie no número de ramos curtos por célula axial, mesmo havendo
a variação de 1 - 5 desse caracter na espécie (Börgesen, 1930; Feldmann-Mazoyer,
1940; Dawson, 1949; Wollaston, 1968, 1971a; Womersley & Bailey, 1970; Abbott
& Hollenberg, 1976; Schneider, 1984; Yoneshigue, 1985; Schneider & Searles, 1991,
1997; Athanasiadis, 1996; Horta, 2000; Littler & Littler, 2000; Torres et al., 2004).
8787878787Novas ocorrências de Ceramiaceae epífitas para a plataforma continental do ES e do RJ, Brasil
Figuras 4-5.Figuras 4-5.Figuras 4-5.Figuras 4-5.Figuras 4-5. Antithamnionella bermudica. (4) Hábito geral do talo. (5) Detalhe da região apical mostrando
células glandulares localizadas nos ramos determinados. Figuras 6-8.Figuras 6-8.Figuras 6-8.Figuras 6-8.Figuras 6-8. Antithamnionella breviramosa. (6)
Hábito geral do talo. (7, 8) Detalhe da região apical mostrando células glandulares de forma esférica
localizadas nos ramos curtos. Escalas: 100 μm.

8888888888 J. Torres, L. M. S. Gestinari & Y. Yoneshigue-Valentin
Feldmann-Mazoyer (1940), Wollaston (1968, 1971a) e Ugadim et al. (1986) consideram
a presença de três ramos curtos por célula axial como sendo um caracter freqüente e
típico para as espécies Antithamnionella breviramosa e Antithamnionella elegans
(Yoneshigue, 1985). Desse modo, a espécie estudada difere da maioria dos autores
pela presença de 1 - 2 ramos curtos por célula axial. Contudo, concordam com a
descrição de Womersley & Bailey (1970) para as Ilhas Salomão e as frondes observadas
por Dawson (1949) e Schneider & Searles (1991), que também mencionam a presença
de 1 - 2 ramos curtos por célula axial. Dessa maneira, o número de ramos curtos por
célula axial e medidas dos caracteres morfológicos não estão sendo considerados como
critérios estáveis neste estudo.
**Callithamniella tingitana (Schousb. ex Bornet) Feldm.-Maz.
(Figuras 9-14)
Imprimerie Minerva, p. 429. 1940.
Basiônimo: Callithamnion tingitanum Schousb. ex Bornet, Mém. Soc. Sci. Nat.
Cherbourg 28: 329. 1892.
Talo filamentoso, de cor vermelha, medindo de 2-(5)-7 mm de altura, fixo ao substrato
por rizóides pluricelulares, cujas extremidades formam apressórios mais ou menos
digitados. Talo constituído por um eixo decumbente e por eixos eretos de crescimento
definido e indefinido. Cada segmento, tanto do eixo rastejante como do ereto, porta um
ramo curto ou um ramo longo, substituindo o ramo curto. Ramos eretos, partindo do
eixo decumbente, dissecados por ramos curtos, dispostos em uma espiral de dupla hélice,
inseridos na divergência de 1/4, dando a impressão de estarem alternados, um por
segmento, às vezes apresentando células apicais portando pêlos. Eixos rastejantes medindo
95-(158)-215 μm de comprimento e 36-(38)-46 μm de diâmetro. Ramos curtos que
revestem o eixo decumbente constituidos por 9-(12)-16 células, medindo 32-(43)-57
μm de comprimento e 13 μm de diâmetro na base; até 296 μm de comprimento e ao
redor de 6 μm de diâmetro no ápice. Ramos eretos de crescimento indefinido medindo
490 μm de comprimento. Tetrasporângios ovais, cruciadamente divididos, inseridos por
um curto pedúnculo (unicelular) sobre o eixo principal, substituindo um dos ramos
curtos, medindo 32-(62)-89 μm de altura e 25-(43)-57 μm de diâmetro.
Habitat: Crescendo sobre talos de Dictyopteris jamaicensis, dividindo o substrato com
Diplothamnion tetrastichum (Tabela 1).
Material examinado: BRASIL, RIO DE JANEIRO, estação C1D1 (Banco Almirante
Saldanha), 23,391°S - 037,620°W, RFA 33448, C. F. D. Gurgel & A. Taouil s.n.,
08/II/1996.
Comentários: Feldmann-Mazoyer (1938) descreveu o gênero Callithamniella baseada
em Callithamniella tingitana. No Brasil, são descritas duas espécies de Callithamniella
para águas superficiais (C. tingitana, por Joly, 1965, e C. flexilis Baardseth, 1941, por
Yoneshigue, 1985), as quais distinguem-se pelos ramos curtos freqüentemente
bifurcados, em C. tingitana, e pelos ramos curtos simples e alongados portando, muitas
vezes, espermatângios oriundos da transformação de um ramo curto adicional, como
observado em C. flexilis. Além disso, a divergência na filotaxia de um mesmo eixo é de
1/4 a 1/6 em C. tingitana (Simons, 1970) e de 1/4 a 1/8 em C. flexilis, variando entre

8989898989Novas ocorrências de Ceramiaceae epífitas para a plataforma continental do ES e do RJ, Brasil
indivíduos ou dentro de uma mesma população (Yoneshigue, 1985). Abbott (1976)
transferiu as duas espécies conhecidas deste gênero (C. tingitana e C. flexilis) para
Grallatoria M. Howe, 1920, com base em plantas tetraspóricas e masculinas coletadas
nas Carolinas. Entretanto, existem diferenças bem marcantes entre os dois gêneros.
Em Callithamniella, cada segmento do eixo rastejante porta um ramo curto ou um
ramo longo, inserido de maneira espiralada enquanto, em Grallatoria, cada segmento
porta um ramo curto e um ramo longo, inseridos de maneira dorsiventral. O número
e a inserção dos rizóides em cada segmento desse eixo também apresentam diferenças
entre os dois gêneros: um rizóide na célula basal de um ramo curto, ou na primeira
célula de um ramo longo, em Callithamniella, e 1 - 2 rizóides na face ventral do eixo
rastejante, em Grallatoria. A filotaxia também é bem marcante para os dois gêneros:
dupla hélice, em Callithamniella; uma única hélice em direção à base e pseudodicotomias
em direção ao ápice, em Grallatoria. Esses dois gêneros distinguem-se, também, quanto
ao plano de divisão dos tetrasporângios: geralmente cruciados em Callithamniella e
sempre tetraédricos em Grallatoria. Baseado nos diferentes caracteres vegetativos e
reprodutivos de Grallatoria reptans M. Howe, 1920, Schneider (1984) considerou
Callithamniella como um gênero distinto, sendo seguido por Wynne & Ballantine (1985).
No Brasil, essa espécie foi descrita pela primeira vez por Joly et al. (1965a) para as
águas superficiais da Ilha Anchieta, São Paulo. Logo após, foi descrita para o habitat
raso, na região de Arraial do Cabo por Villaça (1988) e na região de Saquarema por
Amado Filho (1991). Através dos caracteres morfológicos observados, conduziu-se à
identificação dos espécimes como C. tingitana. Entretanto, o material estudado apresenta
somente rizóides pluricelulares, ao contrário do encontrado por Sansón (1994), cujas
plantas possuíam somente rizóides unicelulares. Todas as demais descrições, incluindo
Schneider & Searles (1991), mencionam rizóides pluricelulares e unicelulares em um
mesmo eixo rastejante.
**Compsothamnion thuyoides (Sm.) F. Schmitz
(Figuras 15-20)
Die Natürlichen Pflanzenfamilien, p. 491. 1897.
Basiônimo: Conferva thuyoides Sm., English Botany, pl. 2205. 1810.
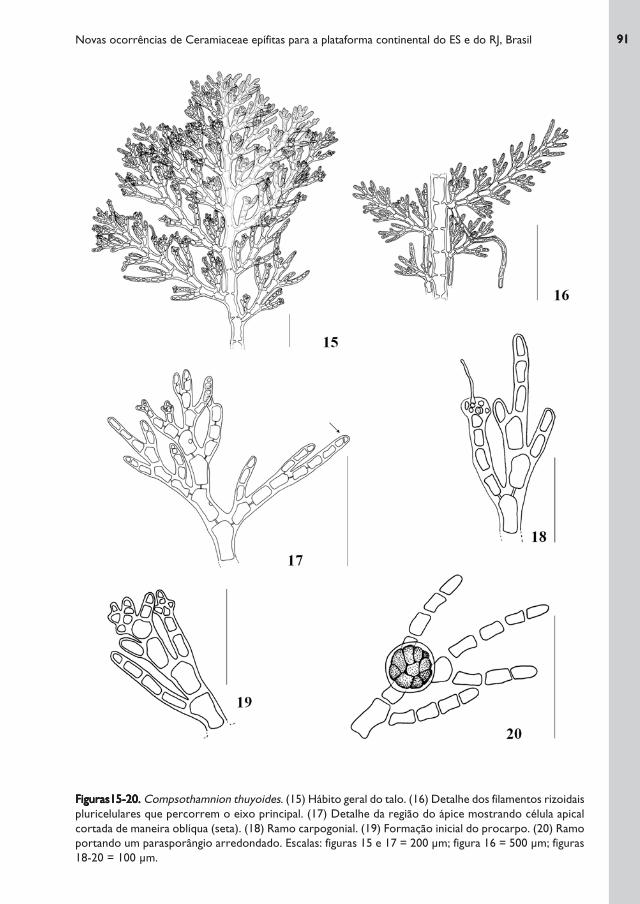
Talo filamentoso, ereto, de cor rósea, de hábito triangular, delicado e flácido, atingindo até
1,5 cm de altura. Fixo ao substrato por numerosos filamentos rizoidais pluricelulares
ramificados, ou não, originados na porção basal dos filamentos, que percorrem o eixo
principal através de ligações secundárias com as células dos râmulos adjacentes (formando
uma corticação parcial), e por um apressório filamentoso e discóide na região basal. Esses
filamentos são longos, medindo 35 μm de diâmetro, originando-se da primeira célula basal
da ramificação lateral. Ramificação de primeira ordem alterna em um plano onde, da parte
superior de cada célula axial, originam-se ramos curtos que, por sua vez, ramificam-se
novamente seguindo o mesmo padrão de ramificação dos ramos de primeira ordem
(originando os ramos de segunda ordem), sendo que as divisões celulares proporcionam
uma aparência de ziguezague ao talo, em função da orientação oblíqua dos núcleos. Os
ramos curtos originados a partir da célula basal da ramificação de primeira ordem estão
sempre na posição adaxial. Talo com crescimento apical a partir do eixo principal, onde a
célula apical – 15-(15,5)-16 μm de comprimento - é cortada de maneira oblíqua.

9090909090 J. Torres, L. M. S. Gestinari & Y. Yoneshigue-Valentin
Figuras 9-14.Figuras 9-14.Figuras 9-14.Figuras 9-14.Figuras 9-14. Callithamniella tingitana. (9) Hábito geral do talo. (10) Detalhe de um ramo longo dissecado
por ramos curtos apresentando células apicais portando pêlos (setas). (11) Detalhe de um rizóide. (12,
13) Detalhe de um ramo de crescimento definido partindo de um rizóide. As setas indicam a direção
apical do filamento prostrado. (14) Ramo portando um tetrasporângio. Escalas: figura 9 = 300 μm; figura
10 = 200 μm; figuras 11-14 = 100 μm.
9191919191Novas ocorrências de Ceramiaceae epífitas para a plataforma continental do ES e do RJ, Brasil
Figuras15-20.Figuras15-20.Figuras15-20.Figuras15-20.Figuras15-20. Compsothamnion thuyoides. (15) Hábito geral do talo. (16) Detalhe dos filamentos rizoidais
pluricelulares que percorrem o eixo principal. (17) Detalhe da região do ápice mostrando célula apical
cortada de maneira oblíqua (seta). (18) Ramo carpogonial. (19) Formação inicial do procarpo. (20) Ramo
portando um parasporângio arredondado. Escalas: figuras 15 e 17 = 200 μm; figura 16 = 500 μm; figuras
18-20 = 100 μm.

9292929292 J. Torres, L. M. S. Gestinari & Y. Yoneshigue-Valentin
Células vegetativas multinucleadas, medindo 42 - 160 μm de comprimento e 24 - 104
μm de largura. Os exemplares estudados apresentam vários procarpos solitários e
terminais nos râmulos, em diversos estágios de desenvolvimento até o cistocarpo
arredondado, mostrando que o carpogônio já havia sido fecundado. Foram encontrados
parasporângios arredondados, medindo 53 x 53 μm de diâmetro.
Habitat: Crescendo sobre talos de Pterocladiella capillacea (Tabela 1).
Material examinado: BRASIL, RIO DE JANEIRO, estação C1D1 (Banco Almirante Saldanha),
23,391°S - 037,620°W, RFA 33449, RFA 33450, C. F. D. Gurgel & A. Taouil s.n., 08-II-1996.
Comentários: Algumas espécies do gênero Compsothamnion (Nägeli) F. Schmitz, 1862,
podem ser confundidas com espécies do gênero Aglaothamnion em função do padrão de
ramificação. Entretanto, este último não apresenta células multinucleadas, além dos dois gêneros
apresentarem diferenças quanto aos tetrasporângios. Estas estruturas são sésseis e solitárias
em frondes de Compsothamnion, enquanto em Aglaothamnion podem ser sésseis ou
pediceladas e solitárias ou gregárias em número de até três por célula. O material estudado
concorda com as descrições de Feldmann-Mazoyer (1940), Schneider & Searles (1991),
Maggs & Hommersand (1993), Horta (2000), Littler & Littler (2000) e Torres et al. (2004).
**Diplothamnion tetrastichum A. B. Joly & Yamaguishi in Joly et al.
(Figuras 21-23)
Rickia 2: 169. 1965b.
Talo filamentoso, segmentado, de cor vermelha, medindo até 1cm de altura, formado
por eixos decumbentes, dos quais originam-se os eretos. Eixos decumbentes medindo
120 - 165 μm de diâmetro com segmentos que medem 330 - 700 μm de comprimento;
de cada segmento podem-se originar os rizóides, unicelulares, geralmente digitados nas
extremidades, medindo 60 - 150 μm de diâmetro. Eixos eretos principais transportando
em cada segmento, na porção superior, dois ramos laterais opostos, de tal sorte que
segmentos sucessivos transportam ramos a 90° sendo, então, a sua disposição oposta
cruzada. Esses eixos são formados por segmentos grandes, medindo 225 - 270 μm de
comprimento e 135 - 165 μm de diâmetro, onde os ramos laterais curtos são formados
por uma única célula que se bifurca no ápice. Células do ramo lateral medindo 240 - 375 μm
de comprimento e 60 - 105 μm de diâmetro.
Habitat: Crescendo esparsamente junto aos talos de Callithamniella tingitana, sobre
talos de Dictyopteris jamaicensis (Tabela 1).
Material examinado: BRASIL, RIO DE JANEIRO, estação C1D1 (Banco Almirante
Saldanha), 23,391°S - 037,620°W, RFA 33451, C. F. D. Gurgel & A. Taouil s.n.,
08-II-1996.
Comentários: Esta espécie foi descrita pela primeira vez para a ciência a partir do material
coletado como epífita de algas maiores nos costões rochosos do litoral norte do Estado
de São Paulo por Joly et al. (1965b). Foi observada também por Oliveira Filho (1969)
sobre talos de macroalgas que crescem nos costões expostos do litoral do Estado do
Espírito Santo. Pela disposição dos ramos laterais cruzados nos segmentos sucessivos,
perfazendo um ângulo de 90º entre esses ramos, os exemplares estudados não deixam
dúvida quanto sua identificação.

9393939393Novas ocorrências de Ceramiaceae epífitas para a plataforma continental do ES e do RJ, Brasil
*Pleonosporium boergesenii (A. B. Joly) R. E. Norris
(Figuras 24-26)
Brit. Phycol. J. 20: 61. 1985.
Basiônimo: Mesothamnion boergesenii A. B. Joly, Bol. Fac. Fil., Ciênc. Letr., Univ. S.
Paulo, Série Botânica 4: 142. 1957.
Talo ereto, filamentoso, delicado, medindo até 0,9 cm de altura, de cor avermelhada. Fixa-
se ao substrato por numerosos rizóides ramificados, multicelulares. Ramificação alterna a
pseudodicotômica. Ramos eretos cilindricos, medindo 75-(96)-113 μm de diâmetro. Ramos
laterais menores que o eixo principal, recurvados para dentro e com ápices obtusos; células
cilindricas, medindo 94-(102)-113 x 38-(46)-57 μm. Polisporângios sésseis, únicos e globosos,
medindo 57-(70)-94 μm de diâmetro e divididos em 4-(6)-8 esporos.
Habitat: Crescendo sobre talos de Zonaria tournefortii (Tabela 1).
Material examinado: BRASIL, ESPÍRITO SANTO, estação C6Y3, 21,165°S - 040,329°W,
RFA 33452, C. F. D. Gurgel & A. Taouil s.n., 14-VI-2002.
Comentários: O material estudado concorda com as descrições de Schneider & Searles
(1991) e Littler & Littler (2000).
**Pterothamnion heteromorphum (J. Agardh) Athanas. & Kraft
(Figuras 27-34)
Eur. J. Phycol. 29: 130. 1994.
Basiônimo: Platythamnion heteromorphum (J. Agardh) Setchell & N. L. Gardner, Univ.
Calif. Publ. Bot. 1: 344. 1903.
Talo filamentoso unisseriado, de cor vermelha, alcançando até 4 cm de altura.
Fixa-se ao substrato por inúmeros rizóides multicelulares, que apresentam 20-
(28)-40 μm de largura e são formados por células grandes pigmentadas, os quais
substituem um dos ramos dísticos do talo, em parte cespitoso. O eixo ereto
apresenta uma organização uniaxial com o eixo principal e laterais de crescimento
indeterminado. Células do eixo principal medindo 140-(252)-400 μm de
comprimento e 90-(174)-240 μm de largura; células dos eixos laterais quadráticas
medindo 30-(42)-60 μm de tamanho. Os ápices destes eixos apresentam um aspecto
sinusoidal e são recobertos pela superposição dos râmulos laterais. De cada porção
distal da célula axial do eixo indeterminado origina-se um verticilo de quatro
ramificações opostas de crescimento determinado. Duas destas ramificações são
dísticas e desiguais em um plano e as outras duas, de pouco desenvolvimento,
localizam-se perpendicularmente às duas primeiras. Nas extremidades dos ramos
dísticos, as últimas células são substituídas por uma estrutura estrelada. Além disso,
estes ramos portam células glandulares no lado adaxial, medindo 18 - 20 μm de
comprimento e 13 - 15 μm de diâmetro. Os ramos carpogoniais, compostos por
quatro células (incluindo a tricogine), estão dispostos nos ápices dos eixos
indeterminados. Tetrasporângios localizados no lado adaxial dos ramos
determinados, medindo 40-(44)-50 μm de comprimento e 30-(40)-50 μm de
diâmetro.
9494949494 J. Torres, L. M. S. Gestinari & Y. Yoneshigue-Valentin
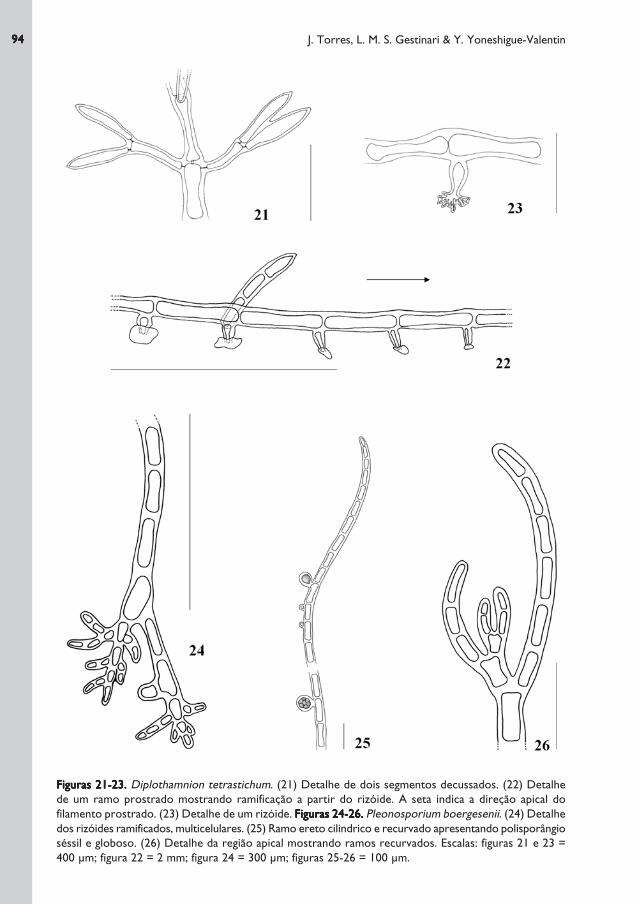
Figuras 21-23.Figuras 21-23.Figuras 21-23.Figuras 21-23.Figuras 21-23. Diplothamnion tetrastichum. (21) Detalhe de dois segmentos decussados. (22) Detalhe
de um ramo prostrado mostrando ramificação a partir do rizóide. A seta indica a direção apical do
filamento prostrado. (23) Detalhe de um rizóide. Figuras 24-26.Figuras 24-26.Figuras 24-26.Figuras 24-26.Figuras 24-26. Pleonosporium boergesenii. (24) Detalhe
dos rizóides ramificados, multicelulares. (25) Ramo ereto cilindrico e recurvado apresentando polisporângio
séssil e globoso. (26) Detalhe da região apical mostrando ramos recurvados. Escalas: figuras 21 e 23 =
400 μm; figura 22 = 2 mm; figura 24 = 300 μm; figuras 25-26 = 100 μm.

9595959595Novas ocorrências de Ceramiaceae epífitas para a plataforma continental do ES e do RJ, Brasil
Figuras 27-34.Figuras 27-34.Figuras 27-34.Figuras 27-34.Figuras 27-34. Pterothamnion heteromorphum. (27) Hábito geral do talo. (28) Detalhe da célula axial do
eixo indeterminado portando o verticilo de quatro ramificações opostas de crescimento determinado. (29)
Ramos portando células glandulares no lado adaxial. 30. Rizóide multicelular substituindo um dos ramos
dísticos do talo. (31, 32) Ápices dos eixos laterais com aspecto sinusoidal e recobertos pela superposição
dos râmulos laterais; presença de células quadráticas. (33) Tetrasporângios localizados no lado adaxial dos
ramos determinados. (34) Extremidades dos ramos dísticos com as últimas células substituídas por uma
estrutura estrelada. Escalas: figura 27 = 1 mm; figuras 28, 30 e 33 = 200 μm; figuras 29, 31-32 e 34 = 100 μm.

9696969696 J. Torres, L. M. S. Gestinari & Y. Yoneshigue-Valentin
Habitat: Crescendo sobre talos de Zonaria tournefortii (Tabela 1).
Material examinado: BRASIL, RIO DE JANEIRO, estações C1D3 (22,883°S - 041,154°W)
e C1D4 (22,851°S - 041,152°W), RFA 33453, RFA 33454, RFA 33455, RFA 33456, RFA
33457, C. F. D. Gurgel & A. Taouil s.n., 23-II-1996.
Comentários: Daines (1913) e Feldmann-Mazoyer (1940) estudaram as estruturas reprodutivas
femininas do gênero Platythamnion J. Agardh, 1892, e reconheceram que o desenvolvimento
do ramo carpogonial e do gonimoblasto, tanto em Platythamnion como em Antithamnion
Nägeli, 1847, era praticamente idêntico e as diferenças nas estruturas vegetativas entre estes
dois gêneros não eram suficientes para a separação genérica. Dawson (1962) considerou
Platythamnion como um gênero artificial fundido com Antithamnion. Entretanto, Kylin (1925)
e Wollaston (1968, 1971b) ainda reconhecem Platythamnion como um gênero distinto. O
gênero Pterothamnion Nägeli in Nägeli & Cramer, 1855, apresenta um padrão de ramificação
similar ao de Platythamnion, embora Pterothamnion desenvolva um par de ramos dísticos de
crescimento determinado na sua porção basal e eixos axiais com a ausência de filamentos
rizoidais. Além disso, da porção distal de cada célula axial origina-se um verticilo de quatro
ramos determinados, sendo que um par de ramos dísticos maiores encontra-se situado em
um plano e o outro, de ramos pouco desenvolvidos, disposto perpendicularmente ao outro.
Por outro lado, a única característica uniforme que distingue o gênero Platythamnion é a
presença de quatro ramos verticilados (Wollaston, 1972; Athanasiadis & Kraft, 1994). Além
das referências anteriores, o material estudado concorda com a descrição de Abbott &
Hollenberg (1976) e com os comentários de Itono (1977) para Platythamnion heteromorphum.
*Spermothamnion repens (Dillwyn) Rosenv.
(Figuras 35-38)
K. danske Vidensk. Selsk. Skr.: 298. 1923-24.
Basiônimo: Conferva repens Dillwyn, British Confervae: 18. 1802.
Talo fixo ao substrato por eixos prostrados que portam numerosos eixos eretos, de cor
rósea, delicado e flácido, medindo 245-(562)-793 μm de altura. Eixos rastejantes com 33-
(35)-40 μm de diâmetro, compostos por células cilíndricas medindo 94-(126)-151 x 19 μm,
dando origem a outros ramos prostrados lateralmente em intervalos irregulares, às vezes aos
pares e, também, a eixos eretos em uma posição anterior nas células dos eixos prostrados,
ou em intervalos irregulares, ou a partir de cada célula. Rizóides unicelulares formados próximo
à porção posterior das células dos eixos prostrados, medindo 19 μm de largura, terminando
em apressórios digitados. Eixos eretos com 205-(428)-726 x 26-(29)-33 μm, alargando-se em
direção ao ápice, cujas células são arredondadas e medem 38-(64)-75 x 13-(16)-19 μm.
Ramificação muito irregular; às vezes sem ramificação nas porções inferiores, às vezes portando
um ramo em cada célula. O mesmo padrão de ramificação pode ser verificado para os eixos
laterais. Presença de procarpos medindo 57-(70)-75 x 57-(66)-75 μm.
Habitat: Crescendo sobre talos de Cryptonemia seminervis (Tabela 1).
Material examinado: BRASIL, ESPÍRITO SANTO, estação C6Y3, 21,165°S - 040,329°W,
RFA 33458, C. F. D. Gurgel & A. Taouil s.n., 14-VI-2002.
Comentários: Os gêneros Ptilothamnion Thuret in LeJolis, 1863, e Spermothamnion Aresch.,
1847, são muito semelhantes quanto à morfologia vegetativa. Porém, diferem-se pelo

9797979797Novas ocorrências de Ceramiaceae epífitas para a plataforma continental do ES e do RJ, Brasil
desenvolvimento do carposporófito (Ballantine & Wynne, 1998; Guimarães & Fujii, 2001).
Pelo hábito geral do talo e medidas dos caracteres, os exemplares estudados são muito
semelhantes à espécie Spermothamnion repens descrita por Feldmann-Mazoyer (1940) para o
mar Mediterrâneo. Do ponto de vista de sua morfologia e organização do talo, os espécimes
deste estudo apresentam um elevado grau de variabilidade no padrão de ramificação. Em algumas
regiões do talo, podem ocorrer ramificações pareadas e ramificações alternadas ou secundas,
concordando com a descrição de Maggs & Hommersand (1993) para as Ilhas Britânicas. Alguns
autores consideraram as várias formas de crescimento de S. repens como espécies distintas.
Segundo Schneider & Searles (1991), exemplares de S. repens podem ser confundidos com
Lejolisia exposita C. W. Schneid. & Searles in Searles & Schneider, 1989, pela superposição das
medidas de largura das células nessas duas espécies. Para o litoral brasileiro são citadas S.
nonatoi A. B. Joly, 1957, caracterizada pela presença de uma conspícua célula apical e filamentos
eretos ao redor de 108 μm de diâmetro, segundo Joly (1965), e S. investiens (P. Crouan
& H. Crouan in Schramm & Mazé) Vickers, 1905, que apresenta filamentos eretos com
diâmetro entre 12 e 14 μm para o material do Espírito Santo (Oliveira Filho, 1969).
*+Tiffaniella snyderiae? (Farl.) I. A. Abbott.
(Figura 39)
Pacific Science 25: 349. 1971.
Basiônimo: Spermothamnion snyderiae Farl., Erythea 7: 77. 1899.
Talo de cor rósea, medindo até 2,1cm de altura, fixo ao substrato por rizóides digitados
localizados nos eixos prostrados que medem 113-(173)-283 μm de diâmetro. Filamentos
eretos unisseriados, medindo 1,1-(1,2)-1,5 cm de altura x 57-(64)-113 μm de diâmetro no
ápice e 113-(169)-245 μm de diâmetro na porção mediana. Estes filamentos são unilateral
ou irregularmente ramificados, raramente com mais de três ordens de ramificação, com
células medianas medindo 94-(130)-151 μm de diâmetro.
Habitat: Crescendo sobre talos de Cryptonemia seminervis (Tabela 1).
Material examinado: BRASIL, ESPÍRITO SANTO, estação C1D23, 20,352°S - 040,093°W,
RFA 33459, C. F. D. Gurgel & A. Taouil s.n., 27-II-1996.
Comentários: Segundo Itono (1977), Abbott (1971) sugeriu a nova combinação, baseada
na presença de fusão celular conspícua no cistocarpo, ausência de filamento involucral,
também no cistocarpo, e presença de polisporângios, embora seus exemplares não
possuissem rizóides bulbosos. A presença destes rizóides é considerada a característica
determinante do gênero Tiffaniella Doty & Meñez, 1960 (Doty & Meñez, 1960). A
identificação de T. snyderiae realizada somente através do hábito geral pode confundir
esta espécie com T. cymodoceae (Börgesen) Gordon, 1972, T. gorgonea (Mont.) Doty
& Meñez, 1960, e T. phycophilum (W. R. Taylor) Gordon, 1972. Entretanto, T.
cymodoceae apresenta menor diâmetro do eixo prostrado (40-90 μm), menor altura
dos filamentos eretos (3-7 mm) e ramificação subdicotômica, enquanto T. snyderiae
apresenta ramificação unilateral ou irregular. Assim como T. snyderiae, T. gorgonea
apresenta rizóides multicelulares, mas estes são como os de T. saccorhiza (Setchell &
N. L. Gardner) Doty & Meñez, 1960, fixos entre os utrículos de Codium Stackh., 1797.
T. phycophilum diferencia-se das demais espécies por apresentar rizóides unicelulares.
9898989898 J. Torres, L. M. S. Gestinari & Y. Yoneshigue-Valentin
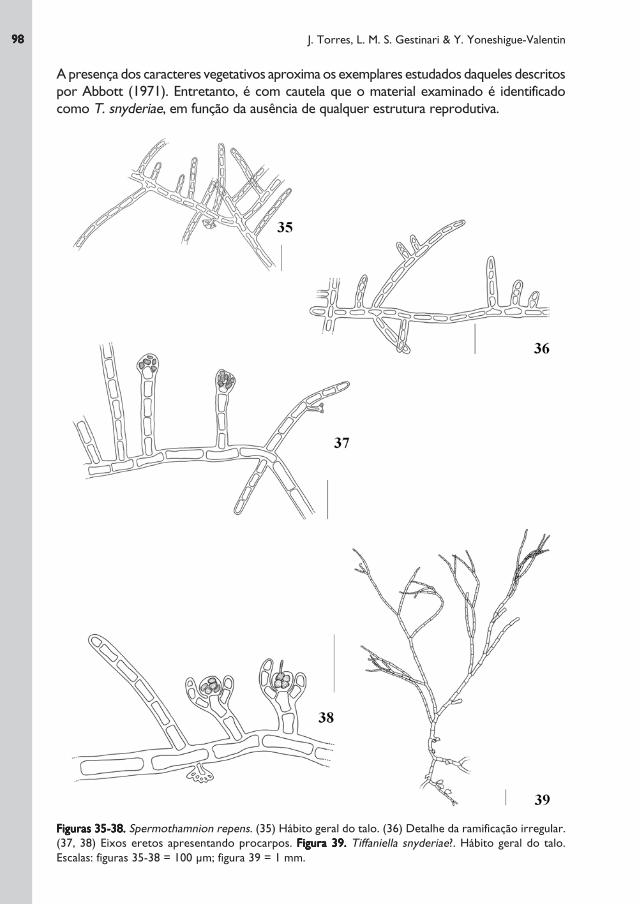
A presença dos caracteres vegetativos aproxima os exemplares estudados daqueles descritos
por Abbott (1971). Entretanto, é com cautela que o material examinado é identificado
como T. snyderiae, em função da ausência de qualquer estrutura reprodutiva.
Figuras 35-38.Figuras 35-38.Figuras 35-38.Figuras 35-38.Figuras 35-38. Spermothamnion repens. (35) Hábito geral do talo. (36) Detalhe da ramificação irregular.
(37, 38) Eixos eretos apresentando procarpos. Figura 39.Figura 39.Figura 39.Figura 39.Figura 39. Tiffaniella snyderiae?. Hábito geral do talo.
Escalas: figuras 35-38 = 100 μm; figura 39 = 1 mm.

9999999999Novas ocorrências de Ceramiaceae epífitas para a plataforma continental do ES e do RJ, Brasil
CCCCCONSIDERAÇÕESONSIDERAÇÕESONSIDERAÇÕESONSIDERAÇÕESONSIDERAÇÕES F F F F FINAISINAISINAISINAISINAIS
A área estudada compreende os estados do Espírito Santo e Rio de Janeiro, inserida na
zona fitogeográfica nordeste-oriental proposta por Oliveira Filho (1977) e reafirmada por
Oliveira et al. (2004). Esta zona destaca-se pelos diversos tipos de substrato, oferecendo
ambiente propício ao estabelecimento de diversas populações algáceas. Além disso, este
trecho do litoral é particularmente banhado por massas d’água diferentes, como as águas
quentes e oligotróficas da Corrente do Brasil, que se desloca de norte para sul, e as Águas
Centrais do Atlântico Sul (ACAS), com origem na Corrente das Malvinas, de temperatura
mais baixa, inferior a 18°C, e ricas em nutrientes, percorrendo a costa de sul para norte.
Do total das 11 espécies encontradas, sete ocorreram no Estado do Espírito Santo
(Aglaothamnion cf. priceanum, Antithamnionella atlantica, A. bermudica, A. breviramosa,
Pleonosporium boergesenii, Spermothamnion repens e Tiffaniella snyderiae?) e quatro
ocorreram no Estado do Rio de Janeiro (Callithamniella tingitana, Compsothamnion
thuyoides, Diplothamnion tetrastichum e Pterothamnion heteromorphum). Algumas destas
espécies ocorrem comumente em mares do Hemisfério Norte e constituem novas citações
para as águas do Atlântico Meridional (Aglaothamnion cf. priceanum, A. bermudica,
Pterothamnion heteromorphum e Tiffaniella snyderiae?), ampliando suas áreas de
distribuição latitudinal.
RRRRREFERÊNCIASEFERÊNCIASEFERÊNCIASEFERÊNCIASEFERÊNCIAS
ABBOTT, I. A., 1971. On some Ceramiaceae (Rhodophyta) from California. Pacific SciencePacific SciencePacific SciencePacific SciencePacific Science, 2525252525: 349-356.
ABBOTT, I. A., 1976. On the red algal genera Grallatoria Howe and Callithamniella Feldmann-
Mazoyer (Ceramiales). British Phycological JournalBritish Phycological JournalBritish Phycological JournalBritish Phycological JournalBritish Phycological Journal, 1111111111: 143-149.
ABBOTT, I. A. & HOLLENBERG, G. J., 1976. Marine algae of California. Marine algae of California. Marine algae of California. Marine algae of California. Marine algae of California. Stanford: Stanford University
Press. 829p.
AMADO FILHO, G. M., 1991. Algas marinhas bentônicas do litoral de Saquarema à ItacoatiaraAlgas marinhas bentônicas do litoral de Saquarema à ItacoatiaraAlgas marinhas bentônicas do litoral de Saquarema à ItacoatiaraAlgas marinhas bentônicas do litoral de Saquarema à ItacoatiaraAlgas marinhas bentônicas do litoral de Saquarema à Itacoatiara
(RJ)(RJ)(RJ)(RJ)(RJ). 323p. Dissertação (Mestrado em Botânica) - Programa de Pós-Graduação em Botânica,
Museu Nacional, Universidade Federal do Rio de Janeiro, Rio de Janeiro.
APONTE, N. E.; BALLANTINE, D. L. & NORRIS, J. N., 1994. Culture studies on the morphology and
life history of Aglaothamnion herveyi (Howe) comb. nov., with notes on A. felipponei (Howe)
comb. nov. (Ceramiaceae, Rhodophyta). PhycologiaPhycologiaPhycologiaPhycologiaPhycologia, 3333333333: 231-238.
ATHANASIADIS, A., 1996. Morphology and classification of the Ceramioideae (Rhodophyta) based
on phylogenetic principles. Opera BotanicaOpera BotanicaOpera BotanicaOpera BotanicaOpera Botanica, 128128128128128: 3-216.
ATHANASIADIS, A. & KRAFT, G., 1994. Description of Pterothamnion squarrulosum (Harvey) comb.
nov. from south-eastern Australia and southern New Zealand, with a taxonomic re-assessment
of the genera Pterothamnion, Platythamnion and Glandothamnus (Ceramiaceae, Rhodophyta).
European Journal of PhycologyEuropean Journal of PhycologyEuropean Journal of PhycologyEuropean Journal of PhycologyEuropean Journal of Phycology, 2929292929: 119-133.
Ballantine, D. L. & Wynne, M. J., 1998. Ptilothamnion speluncarum (Collins & Herv.) comb. nov.
(Ceramiaceae, Rhodophyta) from Puerto Rico. Cryptogamie, AlgologieCryptogamie, AlgologieCryptogamie, AlgologieCryptogamie, AlgologieCryptogamie, Algologie, 1919191919: 223-229.
BÖRGESEN, F., 1930. Marine algae from the Canary Islands, especially from Teneriffe and Gran
Canaria. III Rhodophyceae, Part III Ceramiales. Biologiske Meddelelser Kongelige DanskBiologiske Meddelelser Kongelige DanskBiologiske Meddelelser Kongelige DanskBiologiske Meddelelser Kongelige DanskBiologiske Meddelelser Kongelige Dansk
Videnskabernes SelskabVidenskabernes SelskabVidenskabernes SelskabVidenskabernes SelskabVidenskabernes Selskab, 99999: 1-159.

100100100100100 J. Torres, L. M. S. Gestinari & Y. Yoneshigue-Valentin
BORNET, E., 1892. Les algues de P.K.A. Schousboe, récoltées au Maroc e dans la Méditerranée
de 1815 à 1829 et déterminées par M. Edouard Bornet. Mémoires de la Société des SciencesMémoires de la Société des SciencesMémoires de la Société des SciencesMémoires de la Société des SciencesMémoires de la Société des Sciences
Naturelles de Cherbourg, 28Naturelles de Cherbourg, 28Naturelles de Cherbourg, 28Naturelles de Cherbourg, 28Naturelles de Cherbourg, 28: 165-376.
BRAVIN, I. C.; TORRES, J.; GURGEL, C. F. D. & YONESHIGUE-VALENTIN, Y., 1999. Novas ocorrências de
Clorofíceas marinhas de profundidade para o Brasil. HoehneaHoehneaHoehneaHoehneaHoehnea, 2626262626: 121-133.
COLLINS, F. S. & HERVEY, A. B., 1917. The algae of Bermuda. Proceedings of the AmericanProceedings of the AmericanProceedings of the AmericanProceedings of the AmericanProceedings of the American
Academy of Arts and Science, 53:Academy of Arts and Science, 53:Academy of Arts and Science, 53:Academy of Arts and Science, 53:Academy of Arts and Science, 53: 1-195.
DAINES, L. L., 1913. Comparative development of the cystocarps of Antithamnion and Prionitis.
University of California Publications in BotanyUniversity of California Publications in BotanyUniversity of California Publications in BotanyUniversity of California Publications in BotanyUniversity of California Publications in Botany, 44444: 283-302.
DAWSON, E. Y., 1949. Contributions toward a marine flora of the southern California Channel
Islands, I-III. Allan Hancock Foundation Publications. Occasional Paper Allan Hancock Foundation Publications. Occasional Paper Allan Hancock Foundation Publications. Occasional Paper Allan Hancock Foundation Publications. Occasional Paper Allan Hancock Foundation Publications. Occasional Paper, 88888: 1-57.
DAWSON, E. Y., 1962. Marine red algae of Pacific Mexico. Pt. 7 Ceramiales: Ceramiaceae,
Delesseriaceae. Allan Hancock Pacific ExpeditionAllan Hancock Pacific ExpeditionAllan Hancock Pacific ExpeditionAllan Hancock Pacific ExpeditionAllan Hancock Pacific Expedition, 26 26 26 26 26: 1-207.
DILLWYN, L. W., 1802. British Confervae.British Confervae.British Confervae.British Confervae.British Confervae. London. Pls. 1-20.
DIXON, P. S. & PRICE, J. H., 1981. The genus Callithamnion (Rhodophyta: Ceramiaceae) in the
British Isles. Bulletin of the British Museum (Natural History), BotanyBulletin of the British Museum (Natural History), BotanyBulletin of the British Museum (Natural History), BotanyBulletin of the British Museum (Natural History), BotanyBulletin of the British Museum (Natural History), Botany, 99999: 99-141.
DOTY, M. S. & MEÑEZ, E. G., 1960. Tiffaniella, a new genus in the Ceramiales. TransactionsTransactionsTransactionsTransactionsTransactions
American Microscopical SocietyAmerican Microscopical SocietyAmerican Microscopical SocietyAmerican Microscopical SocietyAmerican Microscopical Society, 7979797979: 135-144.
FELDMANN-MAZOYER, G., 1938. Un nouveau genre de Céramiacée: Callithamniella. Bulletin des TravauxBulletin des TravauxBulletin des TravauxBulletin des TravauxBulletin des Travaux
Publiés par la Station d’Aquiculture et de Pêches de Castiglione. Annales, 1937Publiés par la Station d’Aquiculture et de Pêches de Castiglione. Annales, 1937Publiés par la Station d’Aquiculture et de Pêches de Castiglione. Annales, 1937Publiés par la Station d’Aquiculture et de Pêches de Castiglione. Annales, 1937Publiés par la Station d’Aquiculture et de Pêches de Castiglione. Annales, 1937: 91-102.
FELDMANN-MAZOYER, G., 1940. Recherches sur les Ceramiacée de la Mediterranée OccidentaleRecherches sur les Ceramiacée de la Mediterranée OccidentaleRecherches sur les Ceramiacée de la Mediterranée OccidentaleRecherches sur les Ceramiacée de la Mediterranée OccidentaleRecherches sur les Ceramiacée de la Mediterranée Occidentale.
Argel: Imprimerie Minerva. 510p.
GUIMARÃES, S. M. P. B. & FUJII, M. T., 2001. Ptilothamnion speluncarum (Ceramiales, Rhodophyta):
estudo das estruturas reprodutivas femininas confirma a presença do gênero no Brasil. HoehneaHoehneaHoehneaHoehneaHoehnea,
2828282828: 297-305.
HAROUN, R. J.; VAN REINE, W. F. P.; MÜLLER, D. G.; SERRÃO, E. & HERRERA, R., 1993. Deep-water
macroalgae from the Canary Islands, new records and biogeographical relationships. HelgoländerHelgoländerHelgoländerHelgoländerHelgoländer
MeeresuntersMeeresuntersMeeresuntersMeeresuntersMeeresunters, 4747474747: 125-143.
HORTA, P. A., 2000. Macroalgas do infralitoral do sul e sudeste do Brasil: taxonomia eMacroalgas do infralitoral do sul e sudeste do Brasil: taxonomia eMacroalgas do infralitoral do sul e sudeste do Brasil: taxonomia eMacroalgas do infralitoral do sul e sudeste do Brasil: taxonomia eMacroalgas do infralitoral do sul e sudeste do Brasil: taxonomia e
biogeografia.biogeografia.biogeografia.biogeografia.biogeografia. 301p. Tese (Doutorado em Botânica) - Programa de Pós-Graduação em Botânica,
Instituto de Biociências, Universidade de São Paulo, São Paulo.
HOWE, M. A. & TAYLOR, W. R., 1931. Notes on new or little know marine algae from Brazil.
BrittoniaBrittoniaBrittoniaBrittoniaBrittonia, 11111: 7-33.
ITONO, H., 1977. Studies on the Ceramiaceous algae (Rhodophyta) from Southern parts of
Japan. Bibliotheca PhycologicaBibliotheca PhycologicaBibliotheca PhycologicaBibliotheca PhycologicaBibliotheca Phycologica, 3535353535: 1-499.
JOLY, A. B., 1957. Contribuição ao conhecimento da flora ficológica marinha da Baía de Santos
e arredores. Boletim da Faculdade de Filosofia, Ciências e Letras da Universidade de SãoBoletim da Faculdade de Filosofia, Ciências e Letras da Universidade de SãoBoletim da Faculdade de Filosofia, Ciências e Letras da Universidade de SãoBoletim da Faculdade de Filosofia, Ciências e Letras da Universidade de SãoBoletim da Faculdade de Filosofia, Ciências e Letras da Universidade de São
Paulo, BotânicaPaulo, BotânicaPaulo, BotânicaPaulo, BotânicaPaulo, Botânica, 14 14 14 14 14: 3-199.
JOLY, A. B., 1965. Flora marinha do litoral norte do Estado de São Paulo e regiôes circunvizinhas.
Boletim da Faculdade de Filosofia, Ciências e Letras da Universidade de São Paulo, Série 294Boletim da Faculdade de Filosofia, Ciências e Letras da Universidade de São Paulo, Série 294Boletim da Faculdade de Filosofia, Ciências e Letras da Universidade de São Paulo, Série 294Boletim da Faculdade de Filosofia, Ciências e Letras da Universidade de São Paulo, Série 294Boletim da Faculdade de Filosofia, Ciências e Letras da Universidade de São Paulo, Série 294
(Botânica)(Botânica)(Botânica)(Botânica)(Botânica), 21 21 21 21 21: 1-393.
JOLY, A. B.; CORDEIRO, M.; YAMAGUISHI, N. & UGADIM, Y., 1965a. Additions to the marine flora of
Brasil IV. RickiaRickiaRickiaRickiaRickia, 22222: 129-145.

101101101101101Novas ocorrências de Ceramiaceae epífitas para a plataforma continental do ES e do RJ, Brasil
JOLY, A. B.; CORDEIRO, M.; YAMAGUISHI, N. & UGADIM, Y., 1965b. New marine algae from Southern
Brasil. RickiaRickiaRickiaRickiaRickia, 22222: 159-181.
JOLY, A. B. & OLIVEIRA FILHO, E. C., 1967. Two Brazilian Laminaria. Publicações do Instituto dePublicações do Instituto dePublicações do Instituto dePublicações do Instituto dePublicações do Instituto de
Pesquisa da MarinhaPesquisa da MarinhaPesquisa da MarinhaPesquisa da MarinhaPesquisa da Marinha, 44444: 1-13.
JOLY A. B.; SEMIR, J. & WAKABARA, Y., 1978. Brazilian deep-water marine algae. Additions to the
Brazilian flora V. Revista Brasileira de Botânica,Revista Brasileira de Botânica,Revista Brasileira de Botânica,Revista Brasileira de Botânica,Revista Brasileira de Botânica, 11111: 125-129.
JOLY, A. B. & YONESHIGUE-BRAGA, Y., 1966. Primeira nota sobre algas marinhas durante as viagens do
navio oceanográfico Almirante Saldanha. Publicações do Instituto de Pesquisa da MarinhaPublicações do Instituto de Pesquisa da MarinhaPublicações do Instituto de Pesquisa da MarinhaPublicações do Instituto de Pesquisa da MarinhaPublicações do Instituto de Pesquisa da Marinha, 3434343434: 1-32.
KYLIN, H., 1925. The marine algae in the Biological Station at Friday Harbor, Wash. LundsLundsLundsLundsLunds
Universitets Arsskrift, N. F. Avd.Universitets Arsskrift, N. F. Avd.Universitets Arsskrift, N. F. Avd.Universitets Arsskrift, N. F. Avd.Universitets Arsskrift, N. F. Avd., 2121212121: 1-87.
LITTLER, M. M. & LITTLER, D. S., 2000. Caribbean Reef Plants. Caribbean Reef Plants. Caribbean Reef Plants. Caribbean Reef Plants. Caribbean Reef Plants. Washington, D. C.: Off Shore Graphics, Inc. 542p.
MAGGS, C. A.; GUIRY, M. D. & RUENESS, J., 1991. Aglaothamnion priceanum sp. nov. (Ceramiaceae,
Rhodophyta) from the North-eastern Atlantic: morphology and life history of parasporangial
plants. British Phycological JournalBritish Phycological JournalBritish Phycological JournalBritish Phycological JournalBritish Phycological Journal, 2626262626: 343-352.
MAGGS, C. A. & HOMMERSAND, M. H., 1993. Seaweeds of the British IslesSeaweeds of the British IslesSeaweeds of the British IslesSeaweeds of the British IslesSeaweeds of the British Isles. Volume 1: Rhodophyta.
Part 3A: Ceramiales. London: The Natural History Museum. 444p.
MARCOT-COQUEUNIOT, J., 1988. Sur quelques Peyssoneliacee (Rhodophyceae) récoltées en
profondeur au large des côtes du Brésil. Vie et MilieuVie et MilieuVie et MilieuVie et MilieuVie et Milieu, 3838383838: 299-308.
MCIVOR, L.; MAGGS, C. A. & STANHOPE, M. J., 2002. Rbcl sequences indicate a single evolutionary
origin of multinucleate cells in the red algal tribe Callithamnieae. Molecular Phylogenetics andMolecular Phylogenetics andMolecular Phylogenetics andMolecular Phylogenetics andMolecular Phylogenetics and
EvolutionEvolutionEvolutionEvolutionEvolution, 2323232323: 433-446.
NORRIS, R. E., 1985. Studies on Pleonosporium and Mesothamnion (Ceramiaceae, Rhodophyta)
with a description of a new species from Natal. British Phycological Journal, 20British Phycological Journal, 20British Phycological Journal, 20British Phycological Journal, 20British Phycological Journal, 20: 59-68.
OLIVEIRA, E. C.; HORTA, P. A.; AMANCIO, C. E. & SANT’ANNA, C. L. 2004. Evaluation and priorityEvaluation and priorityEvaluation and priorityEvaluation and priorityEvaluation and priority
actions for conservation of the biodiversity of the marine and coastal zone actions for conservation of the biodiversity of the marine and coastal zone actions for conservation of the biodiversity of the marine and coastal zone actions for conservation of the biodiversity of the marine and coastal zone actions for conservation of the biodiversity of the marine and coastal zone - Marine
angiosperms and algae from the Brazilian coast. Disponível em: <www.bdt.fat.org.br-workshop-
costa-algas>. Acesso em: 30 ago. 2006.
OLIVEIRA FILHO, E. C., 1969. Algas marinhas do sul do Estado do Espírito Santo (Brasil). I.
Ceramiales. Boletim de Botânica da Universidade de São PauloBoletim de Botânica da Universidade de São PauloBoletim de Botânica da Universidade de São PauloBoletim de Botânica da Universidade de São PauloBoletim de Botânica da Universidade de São Paulo, 2626262626:1-277.
OLIVEIRA FILHO, E. C., 1977. Algas marinhas bentônicas do Brasil.Algas marinhas bentônicas do Brasil.Algas marinhas bentônicas do Brasil.Algas marinhas bentônicas do Brasil.Algas marinhas bentônicas do Brasil. 407p. Tese (Livre-Docência).
Instituto de Biociências, Universidade de São Paulo, São Paulo.
ROSENVINGE, L. K., 1923-1924. The marine algae of Denmark: Contributions to their natural
history. Part III. Rhodophyceae III (Ceramiales). Kongelige Danske Videnskabernes SelskabsKongelige Danske Videnskabernes SelskabsKongelige Danske Videnskabernes SelskabsKongelige Danske Videnskabernes SelskabsKongelige Danske Videnskabernes Selskabs
Skrifter, 7. Række, Naturvidenskabelig og Mathematisk Afdeling 7Skrifter, 7. Række, Naturvidenskabelig og Mathematisk Afdeling 7Skrifter, 7. Række, Naturvidenskabelig og Mathematisk Afdeling 7Skrifter, 7. Række, Naturvidenskabelig og Mathematisk Afdeling 7Skrifter, 7. Række, Naturvidenskabelig og Mathematisk Afdeling 7: 285-487.
SANSÓN, M., 1994. Notes on Ceramiaceae (Rhodophyta) from the Canary Islands: new records
and observations on morphology and geographical distribution. Botanica MarinaBotanica MarinaBotanica MarinaBotanica MarinaBotanica Marina, 3737373737: 347-356.
SCHNEIDER, C. W., 1984. Studies on Antithamnionella, Callithamniella and Calloseris (Rhodophyta,
Ceramiales) from North Carolina, USA. PhycologiaPhycologiaPhycologiaPhycologiaPhycologia, 2323232323: 455-464.
SCHNEIDER, C. W. & SEARLES, R. B., 1991. Seaweeds of the Southeastern United StatesSeaweeds of the Southeastern United StatesSeaweeds of the Southeastern United StatesSeaweeds of the Southeastern United StatesSeaweeds of the Southeastern United States – Cape
Hatteras to Cape Canaveral. Durham: Duke University Press. 553p.
SCHNEIDER, C. W. & SEARLES, R. B., 1997. Notes on the marine algae of the Bermudas. 1. New
records of Antithamnieae and Dohrnielleae (Ceramiaceae, Rhodophyta), including Antithamnionella
bermudica sp. nov. PhycologiaPhycologiaPhycologiaPhycologiaPhycologia, 3636363636: 12-23.

102102102102102 J. Torres, L. M. S. Gestinari & Y. Yoneshigue-Valentin
SETCHELL, W. A. & GARDNER, N. L., 1903. Algae of Northwestern America. University of CaliforniaUniversity of CaliforniaUniversity of CaliforniaUniversity of CaliforniaUniversity of California
Publications in Botany, 1Publications in Botany, 1Publications in Botany, 1Publications in Botany, 1Publications in Botany, 1: 165-419.
SIMONS, R. H., 1970. Marine algae from southern Africa I. Six new species from the inter and
infratidal zones. Division of Sea Fisheries, Investigation Report, South Africa, 88Division of Sea Fisheries, Investigation Report, South Africa, 88Division of Sea Fisheries, Investigation Report, South Africa, 88Division of Sea Fisheries, Investigation Report, South Africa, 88Division of Sea Fisheries, Investigation Report, South Africa, 88: 13p.
SMITH, J. E., 1810. English Botany.English Botany.English Botany.English Botany.English Botany. London. Pl. 2205.
TAYLOR, W. R., 1931. Synopsis of the marine algae of Brazil. Revue AlgologiqueRevue AlgologiqueRevue AlgologiqueRevue AlgologiqueRevue Algologique, 5 5 5 5 5: 279-313.
TEIXEIRA, V. L.; PEREIRA, R. C.; MUNIZ, J. A. & SILVA, L. F., 1985. Contribuição ao estudo das algas
de profundidade da costa sudeste do Brasil. Ciência e CulturaCiência e CulturaCiência e CulturaCiência e CulturaCiência e Cultura, 3737373737: 809-815.
TORRES, J.; PEREIRA, S. M. B. & YONESHIGUE-VALENTIN, Y., 2004. Ceramiaceae (Rhodophyta) de
áreas recifais do estado de Pernambuco, Brasil. HoehneaHoehneaHoehneaHoehneaHoehnea, 3131313131: 119-149.
UGADIM, Y.; GUIMARÃES, S. M. P. B. & KANAGAWA, A. I., 1986. Estudos em Acrothamnion,
Antithamnion e Antithamnionella (Rhodophyta, Ceramiales) do Brasil. RickiaRickiaRickiaRickiaRickia, 1313131313: 35-47.
VALENTIN, J. L. & MONTEIRO-RIBAS, W. M., 1993. Zooplankton community structure on the east-
southeast Brazilian Continental Shelf (18° - 23° latitude). Continental Shelf ResearchContinental Shelf ResearchContinental Shelf ResearchContinental Shelf ResearchContinental Shelf Research, 1313131313: 407-424.
VILLAÇA, R. C., 1988. Le phytobenthos infralittoral des biotopes sciaphiles dans la régionLe phytobenthos infralittoral des biotopes sciaphiles dans la régionLe phytobenthos infralittoral des biotopes sciaphiles dans la régionLe phytobenthos infralittoral des biotopes sciaphiles dans la régionLe phytobenthos infralittoral des biotopes sciaphiles dans la région
d’upwelling de Cabo Frio (Brésil).d’upwelling de Cabo Frio (Brésil).d’upwelling de Cabo Frio (Brésil).d’upwelling de Cabo Frio (Brésil).d’upwelling de Cabo Frio (Brésil). 217p. Thèse (Docteur) - Faculté des Sciences de Luminy,
Université d’Aix-Marseille II, France.
WOLLASTON, E. M., 1968. Morphology and taxonomy of southern Australian genera of Crouanieae
Schmitz (Ceramiaceae, Rhodophyta). Australian Journal of BotanyAustralian Journal of BotanyAustralian Journal of BotanyAustralian Journal of BotanyAustralian Journal of Botany, 1616161616: 217-417.
WOLLASTON, E. M., 1971a. Antithamnion and related genera occurring on the Pacific coast of
North America. SyesisSyesisSyesisSyesisSyesis, 44444: 73-92.
WOLLASTON, E. M., 1971b. Generic features of Antithamnion (Ceramiaceae, Rhodophyta) in the
Pacific region. Proceedings of the International Seaweed SymposiumProceedings of the International Seaweed SymposiumProceedings of the International Seaweed SymposiumProceedings of the International Seaweed SymposiumProceedings of the International Seaweed Symposium, 77777: 142-145.
WOLLASTON, E. M., 1972. The genus Platythamnion J.Ag. (Ceramiaceae, Rhodophyta) on the Pacific Coast
of North America between Vancouver, British Columbia, and southern California. SyesisSyesisSyesisSyesisSyesis, 5 5 5 5 5: 43-53.
WOMERSLEY, H. B. S., 1998. The marine benthic flora of southern AustraliaThe marine benthic flora of southern AustraliaThe marine benthic flora of southern AustraliaThe marine benthic flora of southern AustraliaThe marine benthic flora of southern Australia: Rhodophyta – Part
III C. Ceramiales – Ceramiaceae, Dasyaceae. South Australia: Graphic Print Group. 535p.
WOMERSLEY, H. B. S. & BAILEY, A., 1970. Marine algae of the Solomon Islands. PhilosophicalPhilosophicalPhilosophicalPhilosophicalPhilosophical
Transactions of the Royal SocietyTransactions of the Royal SocietyTransactions of the Royal SocietyTransactions of the Royal SocietyTransactions of the Royal Society, 259b259b259b259b259b: 257-352.
WYNNE, M. J., 2005. A checklist of benthic marine algae of the tropical and subtropical western
Atlantic: second revision. Nova HedwigiaNova HedwigiaNova HedwigiaNova HedwigiaNova Hedwigia, 129129129129129: 1-152.
WYNNE, M. J. & BALLANTINE, D. L., 1985. Notes on the marine algae of Puerto Rico. IV. The taxonomic
placement of Grallatoria (Ceramiaceae, Rhodophyta). Cryptogamie, AlgologieCryptogamie, AlgologieCryptogamie, AlgologieCryptogamie, AlgologieCryptogamie, Algologie, 66666: 219-229.
YONESHIGUE, Y., 1985. Taxonomie et ecologie des algues marines de la région de Cabo FrioTaxonomie et ecologie des algues marines de la région de Cabo FrioTaxonomie et ecologie des algues marines de la région de Cabo FrioTaxonomie et ecologie des algues marines de la région de Cabo FrioTaxonomie et ecologie des algues marines de la région de Cabo Frio
(Rio de Janeiro, Brésil)(Rio de Janeiro, Brésil)(Rio de Janeiro, Brésil)(Rio de Janeiro, Brésil)(Rio de Janeiro, Brésil). 454p. Thèse (Docteur d’État-Sciences) - Faculté des Sciences de Luminy,
Université d’Aix-Marseille II, France.
YONESHIGUE-VALENTIN, Y.; GESTINARI, L. M. S. & FERNANDES, D. R. P., 2006. Capítulo 2. Macroalgas.
In: LAVRADO, H. P. & IGNACIO, B. L. (Eds.) Biodiversidade bentônica da região central da ZonaBiodiversidade bentônica da região central da ZonaBiodiversidade bentônica da região central da ZonaBiodiversidade bentônica da região central da ZonaBiodiversidade bentônica da região central da Zona
Econômica Exclusiva brasileira.Econômica Exclusiva brasileira.Econômica Exclusiva brasileira.Econômica Exclusiva brasileira.Econômica Exclusiva brasileira. Rio de Janeiro: Museu Nacional. p.67-105 (Série Livros n. 18).
YONESHIGUE-VALENTIN, Y.; MITCHELL, G. J. P. & GURGEL, C. F. D., 1995. Quelques observations
préliminaires sur les macroalgues de la plate-forme continentale du sud-est brésilien. ActaActaActaActaActa
Botanica GallicaBotanica GallicaBotanica GallicaBotanica GallicaBotanica Gallica, 142142142142142: 161-165.




















