
Nose-to-brain peptide delivery – the potential of nanotechnology
44
Nose-to-brain peptide delivery – the potential of nanotechnology Eleni Samaridou 1 and Maria José Alonso 1,2* 1 Center for Research in Molecular Medicine and Chronic Diseases (CIMUS), Av. Barcelona s/n, Campus Vida, Universidade de Santiago de Compostela, 15782 Santiago de Compostela, Spain. 2 Department of Pharmacy and Pharmaceutical Technology, School of Pharmacy, Universidade de Santiago de Compostela, 15782 Santiago de Compostela, Spain. * Corresponding Author: Prof. María José Alonso. Department of Pharmacy and Pharmaceutical Technology, Center for Research in Molecular Medicine and Chronic Diseases, IDIS research Institute, Universidade de Santiago de Compostela, 15782, Santiago de Compostela, Spain. E-mail address: [email protected] (M.J. Alonso)
-
Upload
khangminh22 -
Category
Documents
-
view
0 -
download
0
Transcript of Nose-to-brain peptide delivery – the potential of nanotechnology

Nose-to-brain peptide delivery – the potential of nanotechnology
Eleni Samaridou1 and Maria José Alonso
1,2*
1Center for Research in Molecular Medicine and Chronic Diseases (CIMUS), Av. Barcelona s/n,
Campus Vida, Universidade de Santiago de Compostela, 15782 Santiago de Compostela, Spain.
2Department of Pharmacy and Pharmaceutical Technology, School of Pharmacy, Universidade de
Santiago de Compostela, 15782 Santiago de Compostela, Spain.
*Corresponding Author: Prof. María José Alonso.
Department of Pharmacy and Pharmaceutical Technology, Center for Research in Molecular Medicine
and Chronic Diseases, IDIS research Institute, Universidade de Santiago de Compostela, 15782,
Santiago de Compostela, Spain.
E-mail address: [email protected] (M.J. Alonso)
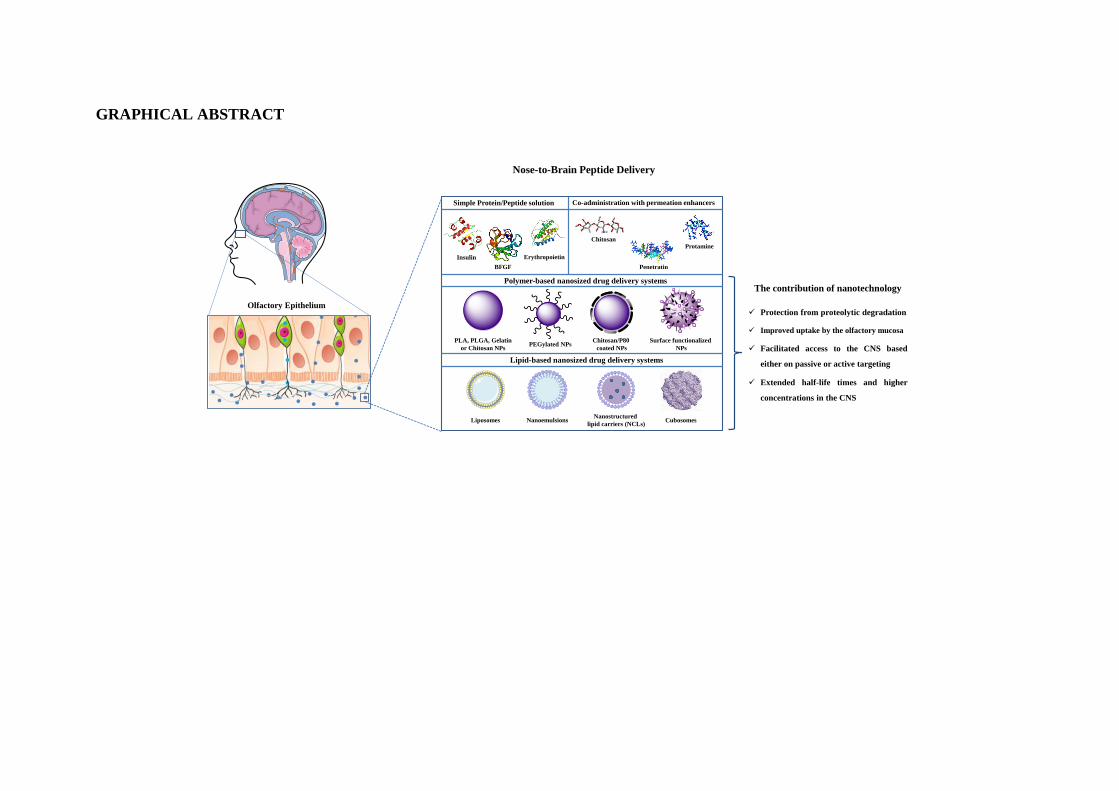
GRAPHICAL ABSTRACT
Olfactory Epithelium
Nose-to-Brain Peptide Delivery
Simple Protein/Peptide solution Co-administration with permeation enhancers
Polymer-based nanosized drug delivery systems
Insulin Erythropoietin
BFGF
Chitosan
Penetratin
PLA, PLGA, Gelatin
or Chitosan NPsPEGylated NPs
Chitosan/P80
coated NPs
Surface functionalized
NPs
Protamine
Protection from proteolytic degradation
Improved uptake by the olfactory mucosa
Facilitated access to the CNS based
either on passive or active targeting
Extended half-life times and higher
concentrations in the CNS
The contribution of nanotechnology
Lipid-based nanosized drug delivery systems
NanoemulsionsNanostructured
lipid carriers (NCLs)Liposomes Cubosomes

Abstract
Nose-to-Brain (N-to-B) delivery offers to protein and peptide drugs the possibility to reach the brain
in a non-invasive way. This article is a comprehensive review of the state-of-the-art of this emerging
peptide delivery route, as well as of the challenges associated to it. Emphasis is given on the
potential of nanosized drug delivery carriers to enhance the direct N-to-B transport of protein or
peptide drugs. In particular, polymer- and lipid- based nanocarriers are comparatively analyzed in
terms of the influence of their physicochemical characteristics and composition on their in vivo fate
and efficacy. The use of biorecognitive ligands and permeation enhancers in order to enhance their
brain targeting efficiency is also discussed. The article concludes highlighting the early stage of this
research field and its still unveiled potential. The final message is that more explicatory PK/PD
studies are required in order to achieve the translation from preclinical to the clinical development
phase.
Keywords:
Nose-to-brain delivery; intranasal drug administration; therapeutic peptides; protein drugs; olfactory;
nanomedicine; nanoparticles; polymer-based nanocarriers; lipid-based nanocarriers.
1. Introduction
Neurological disorders, such as Alzheimer’s disease, Parkinson’s disease, Multiple sclerosis etc., but
also diseases like obesity, behavior disorders and sexual dysfunction have been directly associated to
different modalities of brain dysfunction. These debilitating diseases are nowadays continuously
growing, affecting more and more people worldwide. In addition to the social burden and the
individual suffering that they cause, the treatment of these diseases is also associated with very high
costs.1 Up to date, most of the drugs intended to treat CNS disorders and other brain related diseases
are administered systemically and, for this, a prerequisite is that they are able to cross the blood–
brain barrier (BBB).2,3
Unfortunately, there is a significant number of drugs, notable peptide and
protein drugs which could potentially have a powerful effect in the CNS provided that they could
overcome the BBB or acquire other routes of access to the brain.
Nose-to-brain (N-to-B) delivery may represent a non-invasive method that enables the delivery of
complex drugs to the CNS, while avoiding the BBB. This route is based on the principle that drugs
can access the CNS following a “shortcut” from the nose directly to the brain along the trigeminal or
olfactory nerves, located at the upper part of the nasal cavity. The increasing numbers of peptide and
protein drugs which may be of interest to treat chronic CNS diseases and the recent identification of
important brain functions have stimulated research in the nose-to-brain delivery field. Within this
field, the level of evidence of the value of nanotechnology for the direct CNS targeted peptide

delivery is still limited, however the knowledge generated over the last decade about this specific
topic has raised some expectancies.4,5
Based on this background information, the main objective of this article is to focus on the potential
of nanotechnology-mediated peptide delivery to the brain via the nose. More precisely, this review
will provide the reader with a view of the challenges associated to this modality of administration,
followed by the current status of the nose-to-brain peptide transport, and it will end with a critical
analysis of the value of nanotechnology as compared to that of penetration enhancers for helping
peptide drugs to reach brain targets.
2. Challenges and barriers to N-to-B peptide delivery
In early 1937, Faber reported for the first time the possibility of a direct passage from the nose to the
brain, after administering a dye in the nostrils of rabbits.6 Still, it was only in the late 90s, that the
growing interest in the field of brain delivery motivated the scientific community to start exploring
this alternative route.7–13
Mechanistic studies in animal models have proven that N-to-B drug
transport takes place either by extracellular or transcellular transport mechanisms along the olfactory
epithelium or via the trigeminal nerve, after administration of the drug into the nasal cavity (Fig.
1).14–19
Despite this increasing interest, the mechanisms underlying this direct N-to-B pathway are
not fully elucidated yet.
Pathways related to the transport of peptides from the nose directly to the brain
The olfactory region is located at the top part of the nasal cavity under the cribiform plate in close
proximity to the olfactory bulb, interlocking the nose with the brain (Fig. 1). More specifically, the
olfactory epithelium consists of three types of cells, namely the basal epithelial cells, sustentacular
cells, and the olfactory neurons with their cilia extending towards the nasal cavity (Fig. 2).16,20
More
detailed information about nasal physiology can be found in previous reviews.16,18,19,21
Figure 1. Olfactory and Trigeminal nerve position in the nasal cavity
Trigeminal
Nerve
Olfactory
Nerve
Brain
Nose
Nasal
Epithelium
Olfactory
Epithelium

After administration of the drug into the nasal cavity, N-to-B drug transport may occur through the
olfactory epithelium, either (i) by axonal transport after internalization into the neurons, (ii) by
paracellular transport across the spaces between cells and, notably across the channels next to the
olfactory nerves, or (iii) by transcellular transport across the basal epithelial cells (Fig. 2).14–17,22,23
The paracellular pathway is considered to be the dominant transport mechanism based on animal
studies, and it allows a more rapid drug transport (usually <30 minutes) than the others, which can
last from a few hours up to days.16,24
This could be explained by the slow regeneration of the
olfactory neurons (every ~1 month) and the coexistence of mature and newly formed neurons,
resulting in the absence of tight junctions in some parts of the olfactory epithelium.24
This leakiness,
in combination with the bulk flow of the cerebrospinal fluid (CSF) into the brain, enables the
transport of the intranasally administered drugs to the CNS.16
However, the predominance of one
specific transport mechanism vs. the others depends on the properties of the drug or the delivery
system used.25
Depending on the pathway, the drug may reach the olfactory bulb by intraneuronal
uptake and, from there, it may go into the brain regions connected to the olfactory tract (i.e., the
piriform cortex, hypothalamus, amygdala) and finally disseminate through the CNS, and/or it may
diffuse directly from the CSF into the whole CNS (Fig. 2).26
Figure 2. Schematic representation of the olfactory pathway and possible uptake mechanisms
involved in the transport of peptides from the nose directly to the brain. (CSF: cerebrospinal fluid;
DDS: drug delivery system)
Olfactory Pathway
Thalamus
Hippocampus
Hypothalamus
Piriform cortex
Olfactory
Neuron
Drug
or DDS
Olfactory
Bulb
Cribriform
Plate
Mucus
Layer
Olfactory
Epithelium
Intra-
neuronal
Trans-
cellular
Para-
cellular
CSF
Extra-
neuronal
Uptake Mechanisms
Amygdala

Based on studies in different animal models, some authors have proven that direct N-to-B drug
delivery can also take place along the less explored trigeminal nerve, parts of which extend from the
brainstem through the nasal respiratory epithelium, and provide thus a direct passage to the caudal
and the rostral parts of the brain (Fig. 3).24,25,27–30
Still, the contribution of the trigeminal pathway is
not fully understood and is considered to be less relevant than the olfactory track.31
Figure 3. Schematic representation of the Trigeminal nerve pathway. (CSF: cerebrospinal fluid)
Challenges encountered in the transport of peptides from the nose directly to the brain
Despite the potential of this patient-friendly drug delivery route to the CNS, there are significant
challenges associated to this modality of administration. Nose-to-brain transport is significantly
affected by the surface and structural properties of the administered biomolecules (e.g., size and
lipophilicity, degree of ionization). Proteins, because of their larger size (>1000 Da) and
hydrophilicity, are transported in a far less extent than smaller lipophilic molecules. Another
important factor is the presence of metabolic enzymes (cytochrome P450, esterases and transferases)
in the mammalian olfactory mucosa.15,32–35
On the other hand, from an anatomical point of view, the
localization of the olfactory epithelium in the roof of the nasal cavity makes it difficult for drugs to
gain access to the targeted region.36,37
To address these pitfalls and enhance the bioavailability of the
protein molecules, different approaches have been suggested, such as the use of permeation
enhancers, cell penetrating molecules, mucoadhesives or nano-based drug delivery systems.4,5,38–48
The last ones have the additional advantage of protecting the therapeutic load, while improving its
interaction with the olfactory region. However, a limitation of this route, is related to the low
volumes that can be administered (maximal dosing volume in humans is 0.4 mL), which implies the
need of designing nanocarriers with a high drug loading capacity.49
Lastly, it is worth mentioning
Trigeminal Pathway
Brain
StemTrigeminal
Nerve
CSFBulk Flow
Nasal
Epithelium

that so far, the mechanistic studies have been mainly performed in animals, whereas the studies in
humans have focused on the evaluation of the drugs therapeutic effects, or, in some exceptional
situations the measurement of the drug concentrations in the CSF, or the use of positron emission
tomography (PET) scanning.22,50–52
Despite the above listed limitations, the potential benefits of this
pain-free and direct approach are clear and, hence, N-to-B transport of peptides is becoming a
promising alternative to the other established methods for CNS delivery.
3. Direct transport of P/P drugs from nose-to-brain
Owing to the aforementioned challenges, there has been much discussion during the last decades
about whether peptides can efficiently employ the nose-to-brain pathway.53
Nowadays, after
numerous studies published and patents filed, we can say that peptides and proteins can be
transported to the CNS directly through the nose. Fig. 4 depicts the growing interest in this scientific
area, as found in a PubMed database search that selected exclusively articles reporting in vivo studies
(172 articles).54
Interestingly, almost one third of the registered studies were published in the last 3
years.
The first study describing the direct delivery of a protein molecule to the brain through the olfactory
nerve pathway was published by Frey et al. in 1995.8,55
This affirmation was based on the observed
accumulation of the radiolabeled nerve growth factor (NGF) in the olfactory bulb shortly after its
intranasal administration in rats.8 Years later, a breakthrough study was published, in which the
authors showed that insulin and other peptides like melanocortin (4–10) could efficiently reach the
CSF, with undetectable serum levels, after their intranasal administration in 36 healthy human
volunteers.56
Their study was considered to be the first proof of the existence of the olfactory track in
humans, even if Merkus and van den Berg pointed out the necessity of comparing the nose-to-brain
and the intravenous routes in order to have a clear understanding of the distribution of drugs into the
brain.50
Figure 4. Number of publications on nose-to-brain peptide delivery, reporting in vivo studies
[PubMed database].
0
10
20
30
40
50
60
70
1995-1999 2000-2004 2005-2009 2010-2014 2015-present
Nu
mb
er o
f P
ub
lica
tio
ns
Year
in vivo studies

Table 1 illustrates the studies that reported in vivo data showing the nose-to-brain transport of
protein or peptide molecules, administered either as simple aqueous solution, with permeation
enhancers, or in the form of drug delivery systems. Overall, the most explored protein delivered to
the brain via this direct route is insulin, followed by oxytocin and hypocretin-1. These well
characterized peptides have crucial regulating functions in the CNS, owing to the wide distribution
of their receptors in the CNS, and numerous clinical trials have shown their potential in the treatment
against several disorders, such as cognitive and behavior disorders, narcolepsy, and other important
neurological disorders, i.e. Alzheimer’s disease and Parkinson’s disease.57–67
With regard to the
preclinical data, the limited pharmacokinetic data (37 publications) make very difficult the drawing
of clear conclusions regarding the bioavailability of the different peptides in the brain after their
intranasal administration. Still, while most of the studies have reported enhanced brain delivery of
the protein molecules after their intranasal administration in comparison to their intravenous
administration, the bioavailability of these molecules remained low, generally lower than 1%.55,68
Intranasal co-administration or conjugation with permeation enhancers, such as cell permeating
peptides (i.e. penetratin, low molecular weight protamine, Pz-peptide, Tat), polyethylenimine,
chitosan, lauroyl carnitine, cyclodextrins, Pluronic P85 or peppermint oil, has been a frequent
strategy to enhance the bioavailability of the intranasally (i.n.) administered protein molecules.39,40,42–
48,69–80 For example, some authors reported that the co-administration of chitosan (0.25%) with NGF
led to an increase of up to 13 times in the brain bioavailability of the intranasally administered NGF,
relative to the i.n. free peptide solution.39,42
Similarly, other authors found a 60% enhanced olfactory
bulb uptake of exendin after its co-administration with cyclodextrin, compared to the intravenously
administered free peptide.70
On the other hand, there have been attempts to obtain direct visual evidence of the route that the
proteins follow after their intranasal administration, employing mainly radioactivity based imaging
techniques.8,29,43,44,46,48,69,70,75,77,81–95
These studies provided undeniable evidence of the peptides’
ability to reach the CNS following the olfactory and/or trigeminal pathway.
Table 1. Peptide/protein drugs transported directly from the nose to the brain
P/P drug MW
(Da)
Disease Animal Model Ref.
Insulin 5,800 AD Mice, rats, rabbits,
sheep
46,47,75,94,96–
104
Oxytocin 1,000 CBD, ASD,
PTSD, SD, SCZD
Rats, pigs, vampire
bats, monkeys,
macaques
105–109
Erythropoietin 30,400 AD, CI, Epilepsy Mice, rats 85,88,93,110–
114
Human nerve growth factor 26,500 AD Mice, rats 8–10,39,73,115–
120
Basic fibroblast growth factor 18,000 PD Rats 74,121–124

NAP neuropeptide
(NAPVSIPQ)
825 AD, MCI, SCZD,
FTD
Mice, rats 82,125–130
Vasoactive intestinal peptide 2,800 AD Mice, rats 69,81,131,132
Insulin-like growth factor I 7,650 AD, HD, CI Mice, rats 28,85,133–136
Glucagon- like peptide I 4,100 OB Mice 70,71,93,137
Exendin (9-39) 3,400 CHI Mice, rats 43,70,71,137
Leptin 16,000 OB Rats 48,84,138
Interferon – β1Β 18,500 MS Rats, monkeys 29,71,83
Brain derived neurotrophic
factor
26,900 AD, HD, ASD Mice, rats 42,88,139
Neurotoxin I 6,900 Pain management Mice, rats 140–142
S14G-HN (humanin
derivative)
- AD Mice, rats 90,143
Hypocretin-I (orexin A) 3,500 Narcolepsy Rats, monkeys 86,95,144
Glial cell-derived neurotrophic
factor
15,000 PD Mice, rats 145–147
Bovine serum albumin 66,500 - Mice 76,92
Transforming growth factor β1 25,000 PD, CI Mice, rats 121,148
Vascular endothelial growth
factor
38,200 AD Rats 87,149
Pituitary adenylate cyclase-
activating peptide
4,500 PD, CI Mice 44,91
Thyrotropin-releasing hormone 362 Epilepsy Rats 150,151
Substance P 1,347 PD Rats 152,153
NEMO-binding domain
peptide
2,841 HIE Rats 45,154
Osteopontin 35,423 CI Rats 155,156
Arginine-vasopressin 1,080 CBD, pro-social
effects
Rats, monkeys 157,158
Glucagon-like peptide II 3,766 Depression Mice 79
Ovalbumin 45,000 - Rats 159
Leucine-enkephalin 555 Pain management Mice 160
Neurotrophin-4 22,400 MS Rats 88
interleukin-1 receptor
antagonist
17,000 CI Rats 161
Ciliary neurotrophic factor 22,700 AD, HD Rats 88
Transforming growth factor –a 5,000-
35,000
CI Rats 162
Growth differentiation factor 5 27,400 PD Rats 163
Galamin-like peptide 6,500 OB Mice 164
Calcitonin gene-related peptide 3,800 Migraine Rats 165
Exendin-4 4,186 CI Mice 166
Urocortin 4,700 PD Rats 167
Hexarelin 887 GHD Rabbits 89
Neuropeptide Y 4,253 PTSD,
Depression
Rats 168

Neuropeptide S 2,206 Anxiety disorder Mice 169
Cystatin C-peptide 13,000 AD Mice 170
H102 Peptide - AD Rats 171
D1–D2 interfering peptide - MDD Rats 172
Apelin-13 1,550 CI Mice 173
Human b-Amyloid Peptide 4,514 AD Rats 174
Cyclosporine-A 1,200 AD, PD Rats 175
TNF-alpha inhibitory single-
chain antibody fragment
ESBA105
26,300 AD, PD, MS Mice 72
Horseradish peroxidase 40,000 - Rats 55
V24P(10–40) - AD Mice 77
R8-Ab(25–35) - AD Mice 80
Elastin-like polypeptide - - Mice 78
AD: Alzheimer’s Disease, ASD: Autism Spectrum Disorder, CBD: Cognitive and behavior disorders, CHI:
Congenital Hyperinsulinism, CI: Cerebral ischemia, FTD: Fronto-temporal dementia, GHD: Growth
hormone deficiency, HD: Huntington’s disease, HIE: Hypoxic-ischemic encephalopathy, MCI: Mild
cognitive impairment, MDD: Major depressive disorder, MS: Multiple sclerosis, OB: Obesity, PD:
Parkinson’s disease PTSD: Post-traumatic Stress Disorder, SCZD: Schizophrenia, SD: Sexual dysfunction
Given the still limited number of studies in animal models, it is not surprising that the clinical
development of the peptide nose-to-brain formulations is still at an early stage. Nevertheless, as
shown in Table 2, it is encouraging to realize that several peptides are making their way along the
clinical development path, with some of them, i.e. insulin and oxytocin, having reached Phase IV
clinical studies.59,67,176–178
In the case of insulin, the indication selected in the phase IV clinical
studies focused on its role in energy metabolism.178
So far the disclosed studies have shown that a
single dose of intranasal insulin can increase the peripheral insulin sensitivity and reduce the hepatic
fat in healthy humans.179,180
Apart from this, intranasal insulin is currently being evaluated for its
memory-ameliorating effect in Alzheimer’s suffering patients, as well as its action against different
CNS disorders in humans, like Parkinson's disease, multiple system atrophy and psychiatric
conditions such as schizophrenia and major depressive disorder.64
With regard to oxytocin, it is
known that it works as a brain neurotransmitter, thereby influencing the social behavior and
emotional function in different species. Oxytocin has been tested against several disorders, such as
stress disorders, social dysfunction, cognitive and behavior disorders, Autism Spectrum Disorder
(ASD) and sexual dysfunction in clinical studies.65,181
The most advanced indication (Phase IV
clinical studies) is the control of satiety in people with schizophrenia, as well as its effect on pain
threshold.182,183
The first results of these on-going studies have shown that a single dose of intranasal
oxytocin can significantly increase the satiety feeling and, subsequently, reduce food consumption in
schizophrenia patients. No results have been posted yet on its effect on pain sensitivity.
Table 2. P/P drugs under clinical studies for N-to-B delivery

P/P drug Disease Stage of
development
Ref.
Insulin AD, OB, PD,
MS, SCZD,
MDD
Phase I 56,59,184–192
Phase II 52,184,186,188,193–199
Phase III 191,198,200
Phase IV 178
Oxytocin CBD, ASD,
PTSD, SD,
SCZD, Pain
management
Phase I 37,201–211
Phase II 212–217
Phase III 218–220
Phase IV 182,183
Arginine-vasopressin CBD, pro-
social effects
Phase I 56,97,203,206,221,222
Melanocortin (4–10) OB Phase I 56,184,223,224
Phase II 184
Cholecystokinin OB, AD, CBD Phase I 221,225,226
Phase II 227
NAP neuropeptide AD, SCZD Phase I 228
Phase II 229,230
Hypocretin-I (orexin A) OB, AD, PD,
narcolepsy
Phase I 231–233
Hexarelin GHD Phase I 234
Neuropeptide Y OB Phase I 184
Insulin-like growth
factor-I
DB, OB Phase I 195
AD: Alzheimer’s Disease, ASD: Autism Spectrum Disorder, CBD: Cognitive and behavior
disorders, DB: Diabetes, GHD: Growth hormone deficiency, MDD: Major depressive
disorder, OB: Obesity, PD: Parkinson’s disease PTSD: Post-traumatic Stress Disorder,
SCZD: Schizophrenia, SD: Sexual dysfunction
Another interesting point to note is that most of the peptide drugs presented in Table 2, have already
been tested in numerous clinical trials, using systemic administration routes and, thus, their safety
and pharmacological/toxicological effects have been assessed. Still, a new evaluation of the toxicity
and safety of these drugs would be required in the case of intranasal delivery.235
These studies may,
additionally, include histological assessments of local tissues and potentially affected brain areas.
Apart from this, the majority of the P/P drugs in Table 2 have been administered intranasally to
humans with the help of liquid delivery devices, such as spray pumps or nebulizers/atomizers.
Detailed description of the principles and the types of these devices can be found elsewhere.14
This
fact introduces additional regulatory requirements for the development of a intranasal product, such
as droplet size distribution, dose uniformity, plume geometry and spray pattern studies.236,237
4. The potential of nanotechnology for nose-to-brain peptide delivery
The contribution of nanotechnology to this field is crucial since not only it allows the protection of
the delicate therapeutic cargo from degradation, but most importantly it improves the uptake by the
olfactory mucosa and the access to the CNS, based either on passive or active targeting. As a result,
the use of drug delivery nanocarriers has led to enhanced drug concentrations and extended half-life

times with the subsequent improved therapeutic effect of the delivered molecules. According to our
records, Gao et al. was the first group to prove in 2007 that poly(ethylene glycol)-poly(lactic acid)
nanoparticles (PEG-PLA NPs) functionalized with wheat germ agglutinin (WGA) could deliver a
neuropeptide against Alzheimer’s disease directly from the nose to the brain.131
Since then, a variety
of nano-based drug delivery systems have been developed with the objective of improving the nose-
to-brain delivery of a variety of peptide drugs, most of them summarized in Tables 3 and 4. These
delivery carriers made of either polymers (Table 3), mainly polyesters and chitosan, or lipids (Table
4), were engineered to promote the transport across the neural pathway of different drug molecules.
Nevertheless, the number of research articles in this field is limited, a fact that explains the lack of
nose-to-brain potential nanomedicines in clinical development.
We have analyzed the tendency in the nose-to-brain nanomedicine field, and the results are
summarized in Figure 5. It should be noted that 87 % of the 172 publications analyzed, refer to the
administration of drugs in the form of a simple solution or after co-administration with permeation
enhancers to facilitate the drugs transport across the olfactory epithelium. Nevertheless, the trend is
changing now and the number of nano-based drug delivery systems explored to nose-to-brain
peptide/protein delivery increases year by year. The majority of these delivery carriers are either
polymeric nanoparticles, mainly made of PLA and derivatives, and lipid-based nanocarriers, with a
predominance of liposomes and lipid nanoparticles. A tendency is also observed towards the
functionalization of the nanocarriers.
Figure 5. Representation of the publication trends in N-to-B peptide delivery. (NPs: nanoparticles,
NLCs: nanostructured lipid carries, GNLs: gelatin core lipid carriers)
NPs
50.0%
GNLs
18.3%
NCLs
13.6%
Liposomes
9.0%
Emulsions
4.55%
Cubosomes
4.55%
Peptide
Solution
87.0%
Nanocarriers
13.0%
Type
Penetration Enhancement
172 publications in total
Subtype
Polymer Type
without
permeation
enhancers
87.0%
with
permeation
enhancers
13.0%
PEG-PCL
9.1%
PEG-
PLGA
18.2%
PEG-
PLA
36.3%
PLA
18.2%
Gelatin
9.1%
Chitosan
9.1%

4.1 Importance of the chemical and physicochemical characteristics of the nanocarriers
Although it has been generally accepted that the particle size is a key factor in the capacity of
nanocarriers to overcome mucus barriers and well-organized epithelia, the recent work by Ahmad et
al. provided a good illustration of this effect. The authors studied the biodistribution of
nanoemulsions of different sizes after intranasal administration in rats, by fluorescence imaging, and
concluded that nanocarriers with a particle size around 100 nm were able to be transported along the
olfactory or trigeminal route, whereas nanoemulsions with a larger droplet size were not able to
follow the olfactory pathway.238
Apart from this, a number of studies have also reported the
efficiency of nanocarriers with a size up to 200 nm in terms of facilitating the transport of molecules
in animal models.16,129,239–242
These results are aligned with the reported morphological studies of the olfactory epithelium of
different species, which have revealed that the average diameter of olfactory axons is around 200
nm, with many axons to have diameters even less than 100 nm, while in humans this range goes
from 100 nm up to 700 nm.16,243,244
This fact alone sets a size constraint for the effective transcellular
transport of the nanocarriers via the olfactory axons to the brain. In agreement with this
morphological constraint, the size of the majority of the nanocarriers developed for the peptide
delivery through the intranasal route is in the range of 70-150 nm (Table 3&4).
On the other hand, the effect of the zeta potential on the performance of the nanocarriers has not
been clearly elucidated yet. Apart from the already known fact that the use of positively charged
nanocarriers will likely adhere to the mucus layer, due to the presence of negatively charged mucus
proteins in the region, the potential benefit of this adhesive/retention behavior remains to be
clarified.41,245
In a recent study, the authors sought to describe this effect by tracking the localization
of fluorescently labeled poly(lactic-co-glycolic acid) nanoparticles (negative) and chitosan-coated
PLGA NPs (positive) in different brain areas, following their intranasal administration in rats.246
The
conclusion of this study was that while both type of carriers were found in the brain, the surface
charge and composition of the nanocarriers had a significant effect on their transport pathway, with
negative NPs showing a preference for the olfactory pathway, while the positive nanoparticles for
the trigeminal pathway. While these conclusions remain to be confirmed, it should be highlighted
that the comparison of these nanocarriers was not done in a rigorous manner as they had a different
particle size (118 nm for PLGA nanoparticles and 213 nm, for chitosan-coated PLGA nanoparticles).
Irrespective of the differences in the surface charge, a critical parameter that determines the fate of
the nanocarriers after their intranasal administration is their composition.16
For example, in a study
gelatin-nanostructured lipid carriers (GNLs) stabilized with Poloxamer 188 (size of ~170 nm and
zeta potential of –27 mV) were compared with gelatin nanoparticles (GNPs), (size of ~120 nm and
zeta potential of –17 mV), with regard to their capacity for the nose-to-brain transport of bFGF in

hemiparkinsonian rats.123
The results showed the superiority of the Poloxamer 188-containing
formulation, which was attributed to the known permeation and mucodiffusive properties of the
surfactant. Other studies have evaluated the effect of various coatings, i.e. chitosan and polysorbate
80 around the nanoparticles on their olfactory transport.16,136,141,142,247,248
The results showed that the
presence of polysorbate-80 or chitosan could potentially enhance the nanocarriers’ interaction with
the olfactory mucosa. As in the case of poloxamer, this positive behavior of polysorbate 80 was
attributed to its permeation and mucodiffusive properties, whereas chitosan apart from increasing the
hydrophilicity on the surface of the carrier, can significantly prolong also its retention time in the
olfactory area.239
However, these mechanistic behaviors remain to be elucidated.
4.2 Polymer-based nanocarriers
Polymeric nanocarriers, made either of natural or synthetic polymers, have recently attracted
significant research attention for N-to-B peptide delivery, since they offer a plethora of advantages,
like enhanced stability, the ability to protect and control the release of their therapeutic payload and
multiple possibilities for surface modification. However, so far only few examples have been
reported in the literature, including polylactic/glycolic acid (PLGA) nanoparticles, PEG-PLGA
nanoparticles, PLA NPs modified with chitosan or polysorbate 80, PEG-poly (ε-caprolactone) (PEG-
PCL) nanoparticles, chitosan nanoparticles, and gelatin nanoparticles. Table 3 summarizes the
physicochemical characteristics and the PK/PD behavior of the different polymer-based carriers
employed so far in the literature. The performance of these nanocarriers, after their i.n.
administration, was further enhanced in some cases via their surface modification with targeting
moieties, which will be discussed below.
The advantages of using polymer-based nanocarriers for the N-to-B delivery of small hydrophobic
drugs have been explored by different authors since 2000.249–251
This know-how was then transferred
to N-to-B peptide delivery, with Gao et al. being the first to prove the capacity of PEG-PLA NPs
functionalized with WGA, to facilitate the transport of a peptide drug, the vasoactive intestinal
peptide, to the brain in 2007. In their study, an almost 4-fold increase in the concentration of the
peptide in the brain was reported, when delivered associated to PEG-PLA NPs, reaching a 7-fold
increase with the help of the active targeting functionalization, in comparison to the peptide solution
after its intranasal administration in mice.131
Following this pioneering study, several authors have
investigated the effect of the surface composition of PLA/PLGA particles in their capacity to
overcome the N-to-B barriers. Interestingly, the potential of PLA or PLGA nanoparticles for
peptide/protein delivery, as well as the effect of the particle size and the PEGylation on the transport
of nanoparticles across the nasal mucosa was first reported by our group almost 2 decades ago.252,253
Years later, in 2009, the effect of the particle size on the performance of PLA particles for N-to-B
peptide delivery for the N-to-B transport of Thyrotropin-releasing hormone (TRH) was also
reported.151
The evaluation of the uptake of the particles by fluorescent microscopy led to the

conclusion that 100 nm PLA nanoparticles could gain access to different brain regions through the
olfactory epithelium, whereas the larger ones (~560 nm) could not. This explained the enhanced
therapeutic effect of the neuropeptide in terms of suppressing stage IV seizures in epileptic rats.150,151
Moreover, different authors employed PEG-PLGA of a 120 nm size in order to protect Urocortin
(UCN) and basic fibroblast growth factor (bFGF) respectively, from degradation and facilitate their
intranasal delivery to the CNS.122,167
Additionally, Zhang et al. presented PK results, showing an
almost 1.5-fold increase of the AUCs of bFGF in different brain areas in the case of the i.n.
administration of the bFGF-loaded NPs, relative to i.n. administered bFGF solution in rats.122
This
increase reached up to 3 times more in the case of the i.n. administration of lectin modified bFGF-
NPs, and will be further discussed below.
Another tendency observed is the surface modification of PLA NPs with either PEG-containing
surfactants, like polysorbate 80 (P80), or chitosan. This trend, in the case of P80, originated from the
reported improved performance of the P80-coated nanocarriers in the CNS drug delivery, which was
later adopted in N-to-B peptide delivery.254–256
Accordingly, P80-coated PLA NPs with a particle
size of 60 nm were employed for the nose to brain administration of Neurotoxin-I in rats, resulting in
an 1.5/1.8-fold increase in the peptide bioavailability relative to i.v. administration of the same
nanoparticles and peptide solution, respectively.140
In a similar study, the N-to-B transport of the
same peptide was evaluated upon its incorporation into P80-coated PLA nanoparticles (size was not
mentioned in the report) and the brain uptake showed a 2.2/3 times higher drug transport in
comparison to the i.v. administration of the same particles and the i.n. administration of the free
peptide in mice, respectively. This enhanced delivery could potentially be attributed to the PEG-
containing surface coating of the NPs that facilitates their diffusion through the olfactory mucosa
since it raises the hydrophilicity of the carrier, or to its general surfactant-like properties and its
interaction with the endothelial cells.141
Chitosan, on the other hand, is considered to be an attractive polymer due to its biocompatibility and
bioadhesion and penetration enhancing properties.257
It is worth mentioning that our group pioneered
in the development of CS nanoparticles and nanocapsules for nasal drug delivery in the mid-
90s.258,259
Regarding its use in N-to-B peptide drug delivery, chitosan-coated PLA nanoparticles,
having a particle size of 140 nm and a positive surface charge of +34 mV, were shown to
significantly enhance the brain uptake of the neurotoxin-I peptide (~1.8-fold relative to the uncoated
NPs), an effect that was attributed to the mucoadhesive and cell permeating properties of chitosan.142
In another study, Kumar et al. investigated the performance of trimethyl chitosan nanoparticles
(TMC NPs) with a mean size of 440 nm and a positive zeta potential of +15 mV for the intranasal
delivery of Leucine-enkephalin (Leu-Enk). The authors reported a high peptide uptake in different
brain regions, however this conclusion was simply based on qualitatively microscopic
observation.160
Both of the studies mentioned did not provide a systemic comparison though, and

thereby proof that the delivery of the respective peptides is only due to direct passage of the
therapeutic molecules via the olfactory or trigeminal nerves to the brain and not through the nasal
epithelium also.
Another biodegradable and FDA approved biomaterial that has received minor attention is gelatin.
Joachim et al. used gelatin nanoparticles (GNPs) to study their ability to transfer Osteopontin (OPN)
to the brain after their intranasal application. The selection of gelatin was based on the authors claim
that this protein can passively target several brain areas in case of ischemic stroke.155,260
According to
the expectations, the results of this work showed an enhanced response for the peptide associated to
the nanoparticles as compared to the free peptide administered intranasally.
Moreover, nanoparticles made of PEG-poly (ε-caprolactone) and surface-modified with lactoferrin,
with mean diameter less than 90 nm, have been used to study the delivery of NAP, a model
octapeptide, from the nose directly to the brain of Altzheimer’s disease animal models. Throughout
this study, a rapid accumulation of the NPs in various brain areas was showed based on fluorescent
microscopy, whereas the enhanced memory amelioration effect of the Lf-NPs was further proven by
a Morris water maze experiment.129
Depending on the polymer chosen and the composition of the nanocarriers, different properties can
be attributed to the systems. A general characteristic of the polymer-based carriers employed so far
for the N-to-B peptide delivery is that their diameter is within the range of 70-200 nm. Overall, the
surface functionalization of the carriers either with PEG, pegylated surfactants, chitosan and/or
targeting moieties makes a significant contribution to the enhancement of the peptide bioavailability
in the brain. However, a strict comparison among these nanocarriers is difficult to make due to the
different nature of the studies performed to evaluate them.
Table 3. Overview of polymer-based nanocarrier systems for N-to-B peptide delivery.
Nanocarrier Drug Size
(nm)
Z-pot
(mv)
Targeting
molecule
Animal Pharmacokinetics
outcome/
Therapeutic effect
Ref.
D,L- PLA NPs Thyrotropin-
releasing
hormone
~100 - - Rats Significant suppressed
seizures in epileptic rats
Enhancement of
neuroprotective effect
150,151
PLA NPs
coated with
polysorbate 80
NT-I 65 -29 - Rats 1.8-fold increased
bioavailability compared
to i.v. peptide solution
140
PLA NPs
coated with
polysorbate 80
NT-I - - - Mice 3-fold increased brain
concentration of NT-I
relative to the i.n. solution
141
PLA NPs
modified with
chitosan
NT-I 140 +34 - Rats 9.5-fold increased brain
concentration relative to
the i.n. solution and 1.8-
fold to unmodified PLA-
NPs
142
PEG-PLA NPs Vasoactive
intestinal
peptide
100 -
120
- WGA Mice ~7-fold increased
bioavailability relative to
i.n. peptide solution
131
PEG–PLGA
NPs
Urocortin 120 -20 OL Rats ~6-fold enhancement of
the therapeutic effect
relative to i.n. peptide
solution and 3 fold relative
to the unmodified NPs
167
PEG-PLGA
NPs
bFGF 120 -32 STL Rats 1.6- to 3.2-fold higher
AUCs in olfactory bulb,
cerebrum and cerebellum
of rats relative to i.n.
bFGF solution
122
N-trimethyl
chitosan NPs
Leucine –
encephalin
443 +15 - Mice Significant brain uptake
relative to i.n. solution
Significant enhancement
of the therapeutic effect
160
Gelatin NPs Osteopontin 183 - - Rats Enhancement of
therapeutic effect
155
PEG-co-PCL
NPs
NAP peptide 70 -
90
-24 Lf Mice
/Rats
~2.5- fold increased AUC
of coumarin in brain areas
relative to the i.n.
unmodified NPs
Enhanced memory
amelioration effect
129
PEG: Poly(ethylene glycol), PLA: polylactic acid, NPs: Nanoparticles, PLGA: poly(lactic-co-glycolic acid),
PEG-co-PCL: PEG-poly (ε-caprolactone), WGA: Wheat germ agglutinin, OL: Odorranalectin, Lf: Lactoferrin,
STL: Solanum tuberosum lectin, NT-I: Neurotoxin-I, bFGF: basic fibroblast growth factor
4.3 Lipid-based nanocarriers
Lipid-based nanocarriers describe a broad category of drug delivery systems that includes liposomes,
nanoemulsions and solid lipid nanoparticles. From our knowledge, Migliore et al. were the first, in
2009, to explore the use of lipid-based nanocarriers for the N-to-B delivery of protein drugs.159
Following this study, nanoemulsions, liposomes, cubosomes, as well as mixed systems like lipid
nanoparticles with solid matrix or nanostructured lipid carriers (NLC) have been successfully
employed for the N-to-B peptide delivery and analyzed below (Table 4).
In the pioneering work by Migliore et al., the authors investigated the potential of cationic liposomes
with a size of 300 nm, consisting of dioleoylphosphatidylcholine (DOPC), cholesterol and
stearylamine for the N-to-B transport of ovalbumin − a model protein – in rats.159
The i.n.
administered liposomal formulation exhibited increased residence time at the olfactory mucosa and
this was translated into a 10-fold protein concentration in the brain, as compared to the protein
solution administered by the same route. The authors attributed these positive results to the
electrostatic interactions of the positively charged carrier with the negatively charged mucus
proteins. Other authors reported the fact that pegylated liposomes with a size of 110 nm and a neutral
charge facilitated the direct transport of the H102 Peptide to the brain after intranasal administration
to a Alzheimer’s disease rat model.171
In this study, the performance of the liposomal formulation

was compared with that of the free peptide administered intranasally in the presence of chitosan. The
results showed that the i.n. liposomal formulation was significantly more effective than the chitosan
solution in terms of enhancing the access of the H102 peptide to the olfactory bulb, cerebellum,
cerebrum and hippocampus.
Cubosomes are a nanoparticulate system, consisting of amphiphilic lipids and surfactants that are
organized in a cubic nanostructure. These systems contain liquid-crystal phases that may facilitate
the dissolution of hydrosoluble peptides. Wu et al. explored the efficacy of pegylated cubosomes,
consisting of 1-Monoolein (Glycerol Monooleate), Poloxamer 407 and maleimide–PEG–oleate,
functionalized with a lectin, for the N-to-B delivery of the S14G-HN peptide, a novel peptide against
Alzheimer’s disease and Cerebral ischemia.143
The results showed a concentration dependent
enhancement of the neuroprotective effect of the S14G-HN peptide, following their intranasal
administration in rats.
Although a number of reports have described the possibility to deliver small hydrophobic molecules
from the nose to the brain using nanoemulsions, the only work describing a peptide-based
nanoemulsion formulation was applied to Cyclosporine A (CsA).175,261,262
The developed
nanoemulsions in this study consisted of flax-seed oil as the oil phase, known for its neuronal
regulating properties, egg phosphatidylcholine, polysorbate 80 and stearylamine. Their size was 270
nm and their zeta potential was +57 mV. The i.n. administration of CsA-loaded nanoemulsion
resulted in a significantly higher (6 to 18 times) drug concentration in the olfactory bulb and also in
the brain compared to the levels obtained following i.n. and i.v. administration of the free peptide, as
well as i.v. of the peptide loaded nanoemulsion.
Another lipid based system used for the N-to-B peptide delivery is the so called nanostructured lipid
carriers (NLCs), which consist of a combination of solid and liquid lipids. Gartziandia et al.
developed NLCs coated with chitosan, with a size of 114 nm and a positive zeta potential of +28 mV
for the delivery of human insulin-like growth factor-I (hIGF-I).136
The authors reported an enhanced
residence time of the nanocarriers in the nasal epithelium owing to the mucoadhesive properties of
chitosan and a high brain accumulation based on the results obtained using fluorescence imaging
techniques. In a later study, this system was also used by the same group for the N-to-B delivery of
the glial cell-derived neurotrophic factor (GDNF), achieving a significant improvement in terms of
behavior (movement recovery) and neuroprotection, in comparison to the GDNF solution, in a
hemiparkinsonian rat model.146
A different type of lipid nanoparticles, is the one containing a gelatin core (GNLs) surrounded by a
shell of phospholipids, cholesterol and Poloxamer 188.123,124,152,153
These nanoparticles, with a
diameter of 170 nm and a surface charge of -30 mV, were tested for their capacity to enhance the N-
to-B transport of the Substance P peptide, intended to treat hemiparkinsonian rats.152,153
The results

showed an enhanced response of the peptide when delivered through the nose. The same delivery
vehicle was also tested for the N-to-B delivery of bFGF in order to treat cerebral ischemia.124
The
results indicated that the intranasal administration of this formulation led to a significant increase
(~1.5-fold) in the concentrations of the peptide in different brain regions (pallium, hippocampus,
striatum, olfactory bulb), whereas no increase was observed after the i.v. administration of the bFGF
solution or the bFGF loaded GNLs. Furthermore the therapeutic effect of the i.n. administered bFGF
GNLs was verified by the reported improvement in the neurological deficit score and locomotor
activity of rats.
Overall, from these studies it could be concluded that the efficient N-to-B peptide delivery does not
depend solely on the nanocarrier size, but also on its composition. Compared to the other N-to-B
lipid-based peptide delivery technologies, the highest rise of peptide concentration in the brain,
relative to the i.n administration of the peptide solution, was reported for cationic liposomes (10
times higher) followed by cationic nanoemulsions (6 times higher) with diameters bigger than 200
nm (Table 4). This result could be attributed to the increased residence time achieved by these
positively charged formulations in the olfactory region and more importantly to the presence of
surfactants like polysorbate 80, known for its cell permeation enhancing effects. In conclusion,
proper selection of lipids and surfactants can positively affect the peptide delivery through the
olfactory route. Still, the limited number of studies performed so far does not allow us to draw
further conclusions. Focused mechanistic studies are needed to shed light on the complex
interactions of the different nano-based systems with the olfactory epithelium, the importance of the
surface characteristics of the carriers and their effect on the in vivo efficacy of the systems.
Table 4. Overview of lipid-based nanocarrier systems for N-to-B peptide delivery.
Nanocarrier Drug Size
(nm)
Z-pot
(mv)
Targeting
molecule
Animal Pharmacokinetics
outcome/
Therapeutic effect
Ref.
Cationic
liposomes
OVA 299 +19 - Rats 10-fold brain levels &
>4-fold AUCbrain/AUCblood
relative to i.n. peptide
solution
159
Liposomes H102
Peptide
110 -3 - Rats 1.6-3-fold higher AUC
values in specific brain
areas than the i.n. solution
(no drug in the brain after
i.v. admin.)
171
Pegylated
Cubosomes
S14G-HN
(humanin
derivative)
100 -
120
-14 OL Rats 1.7-3.5-fold increase of the
coumarin distribution in
brain regions relative to the
unmodified cubosomes
Significant enhancement of
therapeutic effect
143
Oil-in-Water
Nanoemulsion
CsA 270 +57 - Rats 6-fold increased brain
concentration relative to i.n
CsA solution
175

Gelatin
NCL
bFGF 170 -27 - Rats 1.5-fold enhancement of
peptide brain concentration
and its therapeutic effect
relative to i.n. peptide
solution (no transport)
123
Gelatin
NCL
SP 172 -30 - Rats Significant enhancement of
therapeutic effect
152,153
Gelatin
NCL
bFGF 128 -15 - Rats Increased peptide
concentration in brain
relative to i.n. peptide
solution
Enhanced response
124
Chitosan
coated NCL
hIGF-I 114 +28 - Mice Increased residence time in
the nasal epithelium and
brain-accumulation
136
Chitosan
coated NCL
GDNF 137 +30 - Rat Significant enhancement of
neuroprotective effect
146
OVA: Ovalbumin, bFGF: basic fibroblast growth factor, SP: Substance P, CsA: Cyclosporine-A, hIGF-I:
Human insulin-like growthfactor-I, GDNF: glial cell-derived neurotrophic factor, NCL: nanostructured lipid
carriers
5. Strategies to enhance N-to-B drug transport
5.1 Olfactory targeting strategies
As mentioned above, several studies have explored the use of covalently linked biorecognitive
ligands in order to enhance the N-to-B transport of the nanosized drug delivery carriers. Table 5
summarizes the targeting moieties that have been described in the literature for targeting the
olfactory region. The most commonly used targeting ligands have been proteins that have receptors
in the olfactory region, i.e. lactoferrin, or glycoproteins, i.e. lectins. Lactoferrin (Lf) has received a
significant attention due to the high expression of its receptor (LfR) in the brain endothelial cells and
neurons.49,129,263
For example, Liu et al., studied the N-to-B transport of lactoferrin conjugated PEG-
PCL NPs (< 90 nm), loaded with a neuroprotective octapeptide (NAP), in an Alzheimer’s disease
animal model, and observed a significant neuroprotective and memory ameliorating effect.129
According to the in vivo biodistribution study performed with fluorescent nanoparticles in rats, Lf-
functionalized NPs exhibited a rapid accumulation in various brain areas, in comparison to the
unmodified NPs. Other authors showed the capacity of the same type of nanoparticles (120 nm) for
the N-to-B delivery of rotigotine, a dopamine agonist used to treat Parkinson’s disease, in a mice
model. The results showed a two times higher drug concentration in different brain areas, relative to
the ones achieved with the unmodified NPs.264
Following the study by Ferrari et al. showing the affinity of several lectins for the N-acetyl-D-
glucosamine and sialic acid residues in the olfactory mucosa, a number of works has been oriented
to show that the functionalization of peptide-loaded nanoparticles with specific lectins, such as
wheat germ agglutinin (WGA), solanum tuberosum lectin (STL), ulex europeus agglutinin I (UEA-
1), is a useful approach to N-to-B delivery.41,131,265,266
For example, the functionalization of PEG-
PLA nanoparticles with WGA was reported to double the N-to-B transport of the Vasoactive
intestinal peptide associated to them, in comparison to the transport observed for the unmodified
nanoparticles.131
In a different study, the authors studied the brain distribution of the labeled (with
125I and coumarin) WGA-functionalized PLGA nanoparticles after nasal administration to rats.
267
The conclusion was that the WGA functionalized PEG-PLA NPs could move through the olfactory
epithelium and reach the olfactory bulb in 5 minutes, suggesting their transport through the
extraneuronal pathway via the olfactory epithelium. In addition, a strong radioactivity signal was
found in deeper brain areas (e.g., striatum, hippocampus, medulla, cerebellum, pons) at 30 min post-
administration.
PEG-PLGA nanoparticles have also been functionalized with STL.122,266,268
The resulting
nanoparticles showed an almost 2.5 times higher brain targeting efficiency (AUCBrain/AUCBlood) for
the STL functionalized PEG-PLGA NPs relative to the values attained for the unmodified NPs after
their i.n. administration in rats.266
Some authors have argued about the potential immunotoxicity of lectins and suggested the use of
smaller peptides with lectin-like function, like Odorranalectin (OL), a small peptide (1.7 KDa) from
frog skin secretions.269
This lectin recognizes and specifically binds to L-fucose, which is highly
expressed in the olfactory mucosa. In vivo fluorescence imaging studies showed that the interaction
of fluorescent PEG-PLGA nanoparticles (95 nm), as well as that of pegylated cubosomes (100-200
nm) functionalized with OL, with the olfactory epithelium was significantly improved (up to a 3.5-
fold increase) as compared to the one observed for the unmodified NPs.143,167
Table 5. Examples of targeting ligands and nanocarriers used for Nose-to-Brain drug delivery
Nanosystem Targeting ligands Animal
model
Ref.
PEG-PCL NPs Lactoferrin Rats/Mice 129
PEG-PLGA NPs Lactoferrin Mice 264
PEG-PLGA NPs STL Rats 122,266,268
PEG-PLA NPs WGA Rats/Mice 131,251,267,270
PEG-PLA NPs UEA I Rats 271
PEG-PLGA NPs OL Rats/Mice 167
Pegylated Cubosomes OL Rats 143
PEG: Poly(ethylene glycol), PEG-PCL: PEG-poly (ε-caprolactone), PLA: polylactic acid, NPs: Nanoparticles, PLGA: poly(lactic-glycolic acid), STL:
Solanum tuberosum lectin, WGA: Wheat germ agglutinin, UEA I: Ulex
europeus agglutinin I, OL: Odorranalectin
5.2 Cell-penetrating peptides (CPPs)

CPPs are traditionally positively charged oligopeptides, containing arginine and lysine residues, that
have been long recognized for their ability to interact with biological membranes and enable the
efficient cellular uptake of biomacromolecules, via different endocytosis pathways or direct
cytoplasmic translocation.272
Recent research on N-to-B drug delivery has proven the ability of these
peptides to significantly increase the permeation of several nanosystems through the olfactory
epithelium (Table 6). Among them, the human immunodeficiency virus Tat peptide
(GRKKRRQRRRPQ) is the most commonly used so far.273–277
Tat peptide’s cell penetrating
properties have been directly associated to the presence of the guanidinium groups of arginine,
which can induce both electrostatic and hydrogen bonding with the cell-surface.278
In a recent study,
the authors described a nanostructured lipid carrier (particle size of 206 nm and surface of +22 mV)
coated with Tat-conjugated chitosan, for the intranasal delivery of glial cell-derived neurotrophic
factor (GDNF).147
According to the results, the surface modification of the system with the Tat
peptide led to a significant enhancement of the therapeutic effect of the administered protein, in
terms of behavior, histological evaluation and microgliosis reduction in a mouse model of
Parkinson’s disease. On the other hand, insulin-loaded PLGA NPs, surface modified with the
cationic Tat peptide (~200 nm, +11 mV), were able to accumulate nearly 6.5 times more in the
olfactory bulb in a matter of a few hours, in contrast to the non-modified NPs, that exhibited poor
transport from the nose to the brain.276
Other studies have been oriented to analyze the potential of the Tat peptide to improve the N-to-B
transport of surface-conjugated MPEG-PCL nanomicelles with a size <100 nm and a slightly
positive surface charge.273
The same group employed this carrier for the nose-to-brain delivery of an
anti-tumor drug (camptothecin) and a small interfering ribonucleic acid.274,275,277
In a follow up
study, a similar system (50-80 nm, 10-15 mV) was employed, where MPEG-PCL micelles,
conjugated with a different arginine-based CPP, were tested for their ability to improve the N-to-B
delivery of macromolecules, using dextran as a model drug.279
A comparison was also made with a
system containing a hydrophobic derivative of the CPP (stearyl modification, STR) (100 nm, 20
mV), proving the possibility of targeting different brain areas, depending on the hydrophilicity of the
CPP. Using in vivo fluorescence imaging, it was found that both systems reached the olfactory bulb
in the short period of 15 minutes, in contrast to the labeled dextran control for which almost no
fluorescence was observed in the brain. More interestingly, the more hydrophobic complex
accumulated mostly in the forebrain, whereas the regular CPP complex could be found throughout
the brain. Still, the mechanisms involved throughout this brain translocation remain unknown.
Another CPP type that has attracted attention for nose-to-brain delivery are short peptide fragments
of the well-known CPP protamine, produced by enzymatic degradation, called low molecular weight
protamine (LMWP, CVSRRRRRRGGRRRR). The advantage of these chain-shortened peptides is
their significant lower toxicity and immunogenicity than their parent protein. For example, it has

been reported that the attachment of LMWP onto the surface of PEG-PLGA nanoparticles led to a
significant enhancement of the N-to-B delivery of coumarin.280
Taking all these results into account,
we can conclude that the CPPs hold a potential in terms of facilitating the nanoparticle-mediated
transport of macromolecules from the nose directly to the brain.
Table 6. Examples of nanocarriers surface-modified with CPPs for Nose-to-Brain drug delivery
Nanosystem Surface modification Animal
model
Ref.
Chitosan coated NCL Tat Mice 147
PLGA NPs Tat Mice 276
Nanomicelles MPEG- PCL Tat Rats 273–275,277
Nanomicelles CH2R4H2C Rats 279
PEG-PLA NPs LMWP Rats 280
NCL: nanostructured lipid carriers, PLGA: poly(lactic-glycolic acid), PEG-PCL: PEG-
poly (ε-caprolactone), PLA: polylactic acid, LMWP: low molecular weight protamine
6. Comparative analysis of in vivo studies
In a review by Merkus and van den Berg, the authors defined the principles for a reliable animal
study aiming to prove the direct N-to-B transport. These principles are: (i) the selection of realistic
administration volumes and concentrations (ii) the comparison of the data with those obtained upon
intravenous administration of the same formulation, (iii) the pharmacokinetic analysis in plasma and
CNS and (iv) studying the brain biodistribution of the administered formulations.50
Other authors
defined specific parameters to express the efficacy of different N-to-B delivery approaches, i.e. the
drug targeting efficiency % (DTE %), which represents the efficiency of the drug to reach the brain
compared to the blood, following intranasal versus parenteral administration, and the direct transport
percentage % (DTP %), which represents the percentage of the drug that reaches the brain due to
direct N-to-B transport, with respect to the total amount of drug found in the brain%), as described
by the following equations:13,242,281,282
Drug targeting efficiency % (DTE%)
𝐷𝑇𝐸% =(𝐴𝑈𝐶𝑏𝑟𝑎𝑖𝑛 𝐴𝑈𝐶𝑏𝑙𝑜𝑜𝑑⁄ )𝑖𝑛𝑡𝑟𝑎𝑛𝑎𝑠𝑎𝑙
(𝐴𝑈𝐶𝑏𝑟𝑎𝑖𝑛 𝐴𝑈𝐶𝑏𝑙𝑜𝑜𝑑⁄ )𝑝𝑎𝑟𝑒𝑛𝑡𝑒𝑟𝑎𝑙∙ 100%,
where AUCbrain and AUCblood are the area under the curve of the drug in the brain or blood
respectively, estimated each time for the corresponded route and
Direct transport percentage % (DTP%)
𝐷𝑇𝑃% =(𝐵𝑖𝑛−𝐵𝑥)
𝐵𝑖𝑛∙ 100%,

where Bin is the brain AUC after the intranasal administration, Bx is the brain AUC fraction from the
blood after intranasal administration and equals to Bx = (Bin/Ppar)·Pin, where Ppar is the blood AUC
following parenteral administration, and Pin is the blood AUC following intranasal administration.
The variety of pharmacokinetics/pharmacodynamics parameters analyzed in the different literature
reports (Table 3&4) renders the comparison of the in vivo performance of nanocarriers very difficult.
After a thorough screening of the selected publications, it is apparent that not all of them provide
adequate information to substantiate the direct N-to-B peptide transport, according to the criteria set
by Merkus and van den Berg.50
In fact, most of the reports do not disclose accurate
pK/biodistribution data, but rather indirect efficacy or semiquantitative biodistribution results (Fig.
6-7).124,129,136,143,150–153,155,160,167
Figure 6. (A) Mean concentration of cyclosporine-A (CsA) in the brain and (B) Brain-to-blood
concentration ratio of cyclosporine-A (CsA) after intranasal (IN) or intravenous (IV) administration
of CsA loaded nanoemulsion (CsA-NE) or CsA-solution (CsA-S) to rats. (*p < 0.05 or *p < 0.01 vs
control groups). Adapted with permission from [175].
[B]
[A]

In addition, some studies lack of proper controls, such as a systemic comparison or biodistribution
data, a fact that makes it inappropriate to conclude about the direct N-to-B transport
mechanism.152,155,160
It is, however, important to highlight that a significant number of studies have
reported direct evidence of the ability of nanocarriers to travel through the olfactory pathway and
deliver their payload to the different brain areas, employing fluorescence and radioactivity-based
imaging techniques (Fig. 7).122,129,131,136,140,142,143,151,159,160,167,276
Nevertheless, quantitative information
on the N-to-B transport of the nanocarriers is still missing in the literature.
[A]
Figure 7. Brain distribution study of PEG-PLGA NPs (Odorranalectin-functionalized (OL-NP) and
non-functionalized (NP)) in mice by in vivo imaging system. (A) Fluorescence images of animal
following i.n. administration at different time points; (B) Fluorescence intensity in the brain versus
time. (*p<0.05) Adapted with permission from [167].
Among the studies analyzed throughout this review, high nose-to-brain delivery efficiency was
reported for both, lipid based and polymer-based systems. Comparatively, the highest performance
of a nanocarrier in terms of enhanced transport of the associated peptide was reported for cationic
liposomes of 300 nm, which resulted in 10 times enhanced bioavailability of OVA in the brain.159
Similarly, PLA nanoparticles coated with chitosan (<150 nm) were found to lead to a 9.5-fold
enhanced concentration of Neurotoxin-I, in the brain.142
Another example, describes a 7-fold peptide
(vasoactive intestinal peptide) transport increase following the i.n. administration of WGA
[B]

functionalized PEG-PLGA nanoparticles as compared to the i.n. administered free peptide
solution.131
Similar transport efficiency was observed for the CsA administered in the form of a
cationic oil-in-water nanoemulsion (~270 nm).175
Among the discussed studies, the one from Zhang et al. is considered to be particularly rigorous as it
complies with all the criteria mentioned above.122
The group employed PEG-PLGA NPs surface
modified with STL, with a particle size of 120 nm and a negative surface charge of -32 mV, for the
N-to-B delivery of bFGF. Radioisotopic tracing method enabled the tracking of the radiolabeled
protein molecule after its i.v. and i.n. administration with or without drug delivery system, proving
the superiority of the i.n. administered nanoformulation. DTE% and DTP% values provided
additional evidence of the ability of the carrier to enhance the nose to brain peptide delivery (DTE%
= 1050% and DTP%=90.44% for the olfactory bulb). The enhanced therapeutic effect was later
verified in rats after the i.n. administration of the peptide loaded nanocarriers.
A general conclusion from these studies is that despite the recognized importance of the particle
size, the composition of the carrier plays a critical role in the fate of the intranasally administered
nanocarriers. Overall, the tendency is to observe that cationic or functionalized nanocarriers perform
better than plain negatively charged nanocarriers. Nevertheless, the variability in the parameters and
criteria, as well as the different animal models employed, do not allow an in-depth comparison
among the different systems. Therefore, more exhaustive comparative studies are required in order
to distinguish among the different nano-based N-to-B peptide delivery systems.
Comparative analysis of the in vivo efficacy of the nano-based DDS vs. permeation enhancers
As mentioned above, various authors have proven the ability of non-covalent permeation enhancers
to facilitate the N-to-B delivery of the co-administered peptide drugs through the olfactory
epithelium. One of the most popular permeation enhancers used for the N-to-B peptide delivery is
chitosan, followed by various cell permeating peptides (i.e. penetratin, low molecular weight
protamine, Pz-peptide). In terms of the peptide bioavailability in the brain, the most promising
results so far have been reported by Morishita’s group who presented up to 20 times increased levels
of insulin in different brain areas after its co-administration with L-penetratin (0.5 mM) as compared
to the i.n. administered free protein solution.46
Following these studies, Vaka and co-workers showed
a 13−14-fold increase in the brain bioavailability of NGF and BDNF respectively, when i.n. co-
administered with chitosan, in the form of a solution.39,42
The same authors showed the ability of
peppermint oil to increase around 8 times the brain concentration of NGF, after their i.n. co-
administration.73
Peppermint oil is also believed to have a tight junctions’ opening effect, promoting
thus the paracellular uptake in the olfactory epithelium.
Unfortunately, we have identified only one report showing the performance of penetration enhancers
vs. nanocarriers using the same peptide drug. In particular, in 2015, Zheng et al. compared the

efficiency of the permeation enhancer chitosan, and a formulation of PEGylated liposomes, with a
particle diameter of 110 nm, for the N-to-B delivery of the H102 peptide. The results showed
significantly higher (1.6–3-fold) values of the drug concentration over the time in different brain
areas for the liposomal formulation, as compared to the AUC values observed for the chitosan
solution. According to the authors, this significant difference in performance was attributed to the
ability of the nanoformulation to provide adequate protection to the peptide from enzymatic
degradation, something that a simple co-administration with a permeation enhancer failed to
provide.171
In another study, the authors explored the use of chitosan as a permeation enhancer for
the i.n. administration of bFGF, achieving up to 1.95 times increased concentrations in different
brain areas as compared to the i.n. administered free peptide in rats.74
Similar results though were
reported by later studies, using gelatin based nanocarriers for the N-to-B transport of the same
peptide.123,124
Overall, it is clear that, more comparative studies based on the same peptide drugs are required in
order to make a clear conclusion on the comparison between nano-based drug delivery systems and
permeation enhancers.
7. Conclusions and Future Perspectives
The increasing number of the in vivo studies published over the last years on the direct nose-to-brain
transport of protein and peptide drugs indicates the growing interest in this route. This modality of
administration is particularly attractive for peptides and proteins as they have the chance to reach the
CNS directly, avoiding the disadvantages and limitations of the systemic route. Several studies have
already proven the ability of peptide and protein drugs to be transported through this route to
different brain areas in animal models, while several of them are already undergoing clinical studies.
However, the bioavailability of these peptides remains quite low. The contribution of
nanotechnology in this field is becoming more and more evident. Nanosized drug delivery carriers
provide a versatile platform with great potential in overcoming the underlying challenges of this
route. In general, the particle size and the composition of the carrier are considered to be important
factors that may influence the fate of the nanocarriers after their intranasal administration.
Specifically, a particle size in the range of 100-200 nm has been explored with the conclusion that
the importance of this size range is also connected with the composition of the nanocarrier.
Regarding the composition, both muco-permeating agents (i.e, PEG and pegylated surfactants) and
cell permeation enhancing agents (i.e, chitosan) were reported to lead to some positive effects.
However, the mechanism underlying this positive outcome has not been elucidated. In addition, a
tendency is observed towards the surface functionalization of the nanocarriers with biorecognitive
ligands. Still, no nanosystem has reached the clinical development phase up to now. The limited
number of in vivo studies in non-primate animals and the differences in nasal anatomy between

human and animals, do not allow us to draw clear conclusions. In conclusion, the field of nano-based
N-to-B peptide delivery is still at an early development stage and its potential is not fully unveiled.
To translate this research from the bench to the clinic, more conclusive studies involving
pharmacokinetic and pharmacodynamic data are required to shed light on the brain targeting
efficiency of the nanosystems and the underlying mechanisms.
Acknowledgements
The work was supported by the European B-Smart Consortium, which received funding from the
European Union’s Horizon 2020 research and innovation programme under grant agreement No
721058. Images were adopted from Servier Medical Art by Servier
(http://www.servier.com/Powerpoint-image-bank) and modified by the authors under the following
terms: CREATIVE COMMONS Attribution 3.0 Unported (CC BY 3.0).
References
1. Why Choose Neurodegenerative Diseases? JPND research.
http://www.neurodegenerationresearch.eu/about/why/. Published 2014. Accessed March 17,
2017.
2. Begley DJ. Delivery of therapeutic agents to the central nervous system: the problems and the
possibilities. Pharmacol Ther. 2004;104(1):29-45.
3. Obermeier B, Daneman R, Ransohoff RM. Development, maintenance and disruption of the
blood-brain barrier. Nat Med. 2013;19(12):1584-1596.
4. Fonseca-Santos B, Daflon Gremião MP, Chorilli M. Nanotechnology-based drug delivery
systems for the treatment of Alzheimer’s disease. Int J Nanomedicine. 2015;10:4981-5003.
5. Kulkarni AD, Vanjari YH, Sancheti KH, Belgamwar VS, Surana SJ, Pardeshi C V.
Nanotechnology-mediated nose to brain drug delivery for Parkinson’s disease: a mini review.
J Drug Target. 2015;4(7):522-537.
6. Faber WM. The nasal mucosa and the subarachnoid space. Am J Anat. 1937;62(1):121-148.
7. Sakane T, Akizuki M, Yamashita S, Nadai T, Hashida M, Sezaki H. Transport of cephalexin
to the cerebrospinal fluid directly from the nasal cavity. J Pharm Pharmacol. 1991;43:449-
451.
8. Frey II WH, Liu J, Chen X, et al. Intranasal delivery of 125 I-NGF to the Brain via the
Olfactory Route. In: Iqbal, K., Mortimer, J. A., Winblad, B., Wisniewski, H. M. (Eds)
Research Advances in Alzheimer’s Disease and Related Disorders. John Wiley, New York. ;
1995:329-335.
9. Frey II WH, Liu J, Chen X, et al. Delivery of 125 I-NGF to the Brain via the Olfactory
Route. Drug Deliv. 1997;4(2):87-92.
10. Chen X-Q, Fawcett JR, Rahman Y-E, Ala T, Frey II WH. Delivery of Nerve Growth Factor
to the Brain via the Olfactory Pathway. J Alzheimers Dis. 1998;1(1):35-44.
11. Wang Y, Aun R, Tse FLS. Brain uptake of dihydroergotamine after intravenous and nasal
administration in the rat. Biopharm Drug Dispos. 1998;19(9):571-575.
12. Chou KJ, Donovan MD. Lidocaine distribution into the CNS following nasal and arterial
delivery: A comparison of local sampling and microdialysis techniques. Int J Pharm.

1998;171(1):53-61.
13. Chow HHS, Chen Z, Matsuura GT. Direct transport of cocaine from the nasal cavity to the
brain following intranasal cocaine administration in rats. J Pharm Sci. 1999;88(8):754-758.
14. Djupesland PG, Messina JC, Mahmoud RA. The nasal approach to delivering treatment for
brain diseases: an anatomic, physiologic, and delivery technology overview. Ther Deliv.
2014;5(6):709-733.
15. Wu H, Hu K, Jiang X. From nose to brain: understanding transport capacity and transport
rate of drugs. Expert Opin Drug Deliv. 2008;5(10):1159-1168.
16. Mistry A, Stolnik S, Illum L. Nanoparticles for direct nose-to-brain delivery of drugs. Int J
Pharm. 2009;379(1-2):146-157.
17. Hanson LR, Frey WH. Intranasal delivery bypasses the blood-brain barrier to target
therapeutic agents to the central nervous system and treat neurodegenerative disease. BMC
Neurosci. 2008;9 Suppl 3(3):S5.
18. Malerba F, Paoletti F, Capsoni S, Cattaneo A. Intranasal delivery of therapeutic proteins for
neurological diseases. Expert Opin Drug Deliv. 2011;8:1277-1296.
19. Djupesland PG, Mahmoud RA, Messina JC. Accessing the brain: the nose may know the
way. J Cereb Blood Flow Metab. 2013;33(5):793-794.
20. Moran DT, Rowley JC, Jafek BW, Lovell MA. The fine structure of the olfactory mucosa in
man. J Neurocytol. 1982;11(5):721-746.
21. Pires A, Fortuna A, Alves G, Falcão A. Intranasal drug delivery: How, why and what for? J
Pharm Pharm Sci. 2009;12(3):288-311.
22. Illum L. Is nose-to-brain transport of drugs in man a reality? J Pharm Pharmacol. 2004;56:3-
17.
23. Pardeshi CV, Belgamwar VS. Direct nose to brain drug delivery via integrated nerve
pathways bypassing the blood-brain barrier: an excellent platform for brain targeting. Expert
Opin Drug Deliv. 2013;10(7):957-972.
24. Ruigrok MJR, de Lange ECM. Emerging Insights for Translational Pharmacokinetic and
Pharmacokinetic-Pharmacodynamic Studies: Towards Prediction of Nose-to-Brain Transport
in Humans. AAPS J. 2015;17(3):493-505.
25. Dhuria S V., Hanson LR, Frey II WH. Intranasal Delivery to the Central Nervous System:
Mechanisms and Experimental Considerations. J Pharm Sci. 2010;99(4):26-33.
26. Carmichael ST, Clugnet M-C, Price JL. Central olfactory connections in the Macaque
monkey. J Comp Neurol. 1994;346:403-434.
27. Walker HK. Cranial Nerve V: The Trigeminal Nerve. Clin Methods Hist Phys Lab Exam.
1990:318-321.
28. Thorne RG, Pronk GJ, Padmanabhan V, Frey WH. Delivery of insulin-like growth factor-I to
the rat brain and spinal cord along olfactory and trigeminal pathways following intranasal
administration. Neuroscience. 2004;127(2):481-496.
29. Thorne RG, Hanson LR, Ross TM, Tung D, Frey WH. Delivery of interferon-1b to the
monkey nervous system following intranasal administration. Neuroscience. 2008;152(3):785-
797.
30. Silver WL, Finger TE. The anatomical and electrophysiological basis of peripheral nasal
trigeminal chemoreception. Ann N Y Acad Sci. 2009;1170:202-205.
31. Mustafa G, Alrohaimi AH, Bhatnagar A, Baboota S, Ali J, Ahuja A. Brain targeting by
intranasal drug delivery ( INDD ): a combined effect of trans-neural and para-neuronal
pathway. Drug Deliv. 2014;23(3):1-7.

32. Cuschieri A. Enzyme histochemistry of the olfactory mucosa and vomeronasal organ in the
mouse. J Anat. 1974;118(Pt 3):477-489.
33. Thiebaud N, Veloso Da Silva S, Jakob I, et al. Odorant Metabolism Catalyzed by Olfactory
Mucosal Enzymes Influences Peripheral Olfactory Responses in Rats. PLoS One. 2013;8(3).
34. Hu J, Sheng L, Li L, et al. Essential role of the cytochrome P450 enzyme CYP2A5 in
olfactory mucosal toxicity of naphthalene. Drug Metab Dispos. 2014;42(1):23-27.
35. Ding X, Xie F. Olfactory Mucosa: Composition, Enzymatic Localization, and Metabolism.
In: Richard L. Doty., ed. Handbook of Olfaction and Gustation. 3rd ed. John Wiley & Sons,
Inc.; 2015:63-92.
36. Quintana DS, Guastella AJ, Westlye LT, Andreassen OA. The promise and pitfalls of
intranasally administering psychopharmacological agents for the treatment of psychiatric
disorders. Mol Psychiatry. 2016;21(1):29-38.
37. Quintana DS, Westlye LT, Alnaes D, et al. Low dose intranasal oxytocin delivered with
Breath Powered device dampens amygdala response to emotional stimuli: A peripheral
effect-controlled within-subjects randomized dose-response fMRI trial.
Psychoneuroendocrinology. 2016;69:180-188.
38. Charlton ST, Davis SS, Illum L. Evaluation of bioadhesive polymers as delivery systems for
nose to brain delivery: In vitro characterisation studies. J Control Release. 2007;118(2):225-
234.
39. Vaka SRK, Sammeta SM, Day LB, Murthy SN. Delivery of Nerve Growth Factor to brain
via intranasal administration and enhancement of brain uptake. J Pharm Sci.
2009;98(10):3640–3646.
40. Feng C, Shao X, Zhang C, Liu Q, Chen J, Shen Y. Effect of absorption enhancers on nasal
delivery of basic fibroblast growth factor. African J Pharm Pharmacol.
2011;5(August):1070-1079.
41. Wen MM. Olfactory Targeting Through Intranasal Delivery of Biopharmaceutical Drugs to
the Brain: Current Development. Discov Med. 2011;11(61):497-503.
42. Vaka SRK, Murthy SN, Balaji A, Repka MA. Delivery of brain-derived neurotrophic factor
via nose-to-brain pathway. Pharm Res. 2012;29(2):441-447.
43. Kim TH, Park CW, Kim HY, et al. Low molecular weight (1 kDa) polyethylene glycol
conjugation markedly enhances the hypoglycemic effects of intranasally administered
exendin-4 in type 2 diabetic db/db mice. Biol Pharm Bull. 2012;35(7):1076-1083.
44. Nonaka N, Farr SA, Nakamachi T, Morley JE, Nakamura M, Shioda S BW. Intranasal
Administration of PACAP: Uptake by Brain and Brain Region Targeting with Cyclodextrins.
Peptides. 2012;36(2):168-175.
45. Yang D, Sun YY, Lin X, et al. Intranasal delivery of cell-penetrating anti-NF-κB peptides
(Tat-NBD) alleviates infection-sensitized hypoxic-ischemic brain injury. Exp Neurol.
2013;247:447-455.
46. Kamei N, Shingaki T, Kanayama Y, et al. Visualization and Quantitative Assessment of the
Brain Distribution of Insulin through Nose-to-Brain Delivery Based on the Cell-Penetrating
Peptide Noncovalent Strategy. Mol Pharm. 2016;13(3):1004-1011.
47. Kamei N, Tanaka M, Choi H, et al. Effect of an enhanced nose-to-brain delivery of insulin on
mild and progressive memory loss in the senescence-accelerated mouse. Mol Pharm.
2017;14(3):acs.molpharmaceut.6b01134.
48. Yuan D, Yi X, Zhao Y, et al. Intranasal delivery of N-terminal modified leptin-pluronic
conjugate for treatment of obesity. J Control Release. 2017;In press.
49. van Woensel M, Wauthoz N, Rosière R, et al. Formulations for intranasal delivery of

pharmacological agents to combat brain disease: A new opportunity to tackle GBM? Cancers
(Basel). 2013;5(3):1020-1048.
50. Merkus FWHM, van Den Berg MP. Can Nasal Drug Delivery Bypass the Blood-Brain
Barrier? Questioning the Direct Transport Theory. Drugs R D. 2007;8(3):133-144.
51. Schiöth HB, Craft S, Brooks SJ, Frey WH, Benedict C. Brain insulin signaling and
Alzheimer’s disease: Current evidence and future directions. Mol Neurobiol. 2012;46(1):4-
10.
52. Craft S, Baker LD, Montine TJ, et al. Intranasal Insulin Therapy for Alzheimer Disease and
Amnestic Mild Cognitive Impairment. Arch Neurol. 2012;69(1):29-38.
53. Gomez D, Martinez JA, Hanson LR, Frey II WH, Toth CC. Intranasal treatment of
neurodegenerative diseases and stroke. Front Biosci (Schol Ed). 2012;4:74-89.
54. PubMed Database. US National Library of Medicine, National Institutes of Health.
http://www.ncbi.nlm.nih.gov/pubmed/. Accessed March 1, 2017.
55. Thorne RG, Emory CR, Ala TA, Frey WH. Quantitative analysis of the olfactory pathway for
drug delivery to the brain. Brain Res. 1995;692(1-2):278-282.
56. Born J, Lange T, Kern W, McGregor GP, Bickel U, Fehm HL. Sniffing neuropeptides: a
transnasal approach to the human brain. Nat Neurosci. 2002;5(6):514-516.
57. Stockhorst U, De Fries D, Steingrueber HJ, Scherbaum WA. Insulin and the CNS: Effects on
food intake, memory, and endocrine parameters and the role of intranasal insulin
administration in humans. Physiol Behav. 2004;83(1 SPEC. ISS.):47-54.
58. de la Monte SM. Intranasal Insulin Therapy for Cognitive Impairment and
Neurodegeneration: Current State of the Art. Expert Opin Drug Deliv. 2013;10(12):1699-
1709.
59. Freiherr J, Hallschmid M, Frey WH, et al. Intranasal insulin as a treatment for alzheimer’s
disease: A review of basic research and clinical evidence. CNS Drugs. 2013;27(7):505-514.
60. Pedersen C a, Prange AJ. Induction of maternal behavior in virgin rats after
intracerebroventricular administration of oxytocin. Proc Natl Acad Sci U S A.
1979;76(12):6661-6665.
61. Liblau RS, Vassalli A, Seifinejad A, Tafti M. Hypocretin (orexin) biology and the
pathophysiology of narcolepsy with cataplexy. Lancet Neurol. 2015;14(3):318-328.
62. Roh JH, Jiang H, Finn MB, et al. Potential role of orexin and sleep modulation in the
pathogenesis of Alzheimer’s disease. J Exp Med. 2014;211(13):2487-2496.
63. Thannickal TC, Lai YY, Siegel JM. Hypocretin (orexin) and melanin concentrating hormone
loss and the symptoms of Parkinson’s disease [1]. Brain. 2008;131(1):2007-2008.
64. ClinicalTrials.gov. A service of the U.S. National Institutes of Health.
https://clinicaltrials.gov/.
65. Heinrichs M, Domes G. Neuropeptides and social behaviour: effects of oxytocin and
vasopressin in humans. Prog Brain Res. 2008;170(8):337-350.
66. Benedict C, Frey WH, Schiöth HB, Schultes B, Born J, Hallschmid M. Intranasal insulin as a
therapeutic option in the treatment of cognitive impairments. Exp Gerontol. 2011;46(2-
3):112-115.
67. Meyer-Lindenberg A, Domes G, Kirsch P, Heinrichs M. Oxytocin and vasopressin in the
human brain: social neuropeptides for translational medicine. Nat Rev Neurosci.
2011;12(9):524-538.
68. Thorne RG, Frey WH. Delivery of neurotrophic factors to the central nervous system:
pharmacokinetic considerations. Clin Pharmacokinet. 2001;40(12):907-946.

69. Dufes C, Olivier JC, Gaillard F, Gaillard A, Couet W, Muller JM. Brain delivery of
vasoactive intestinal peptide (VIP) following nasal administration to rats. Int J Pharm.
2003;255(1-2):87-97.
70. Banks WA, During MJ, Niehoff ML. Brain uptake of the Glucagon-Like Peptide-1
Antagonist Exendin (9-39) after Intranasal Administration. J Pharmacol Exp Ther.
2004;309(2):469-475.
71. Khafagy ES, Morishita M, Kamei N, Eda Y, Ikeno Y, Takayama K. Efficiency of cell-
penetrating peptides on the nasal and intestinal absorption of therapeutic peptides and
proteins. Int J Pharm. 2009;381(1):49-55.
72. Furrer E, Hulmann V, Urech DM. Intranasal delivery of ESBA105, a TNF-alpha-inhibitory
scFv antibody fragment to the brain. J Neuroimmunol. 2009;215(1-2):65-72.
73. Vaka SRK, Murthy SN. Enhancement of nose-brain delivery of therapeutic agents for
treating neurodegenerative diseases using peppermint oil. Pharmazie. 2010;65(9):690-692.
74. Feng C, Zhang C, Shao X, et al. Enhancement of nose-to-brain delivery of basic fibroblast
growth factor for improving rat memory impairments induced by co-injection of β-amyloid
and ibotenic acid into the bilateral hippocampus. Int J Pharm. 2012;423(2):226-234.
75. Kamei N, Takeda-Morishita M. Brain delivery of insulin boosted by intranasal
coadministration with cell-penetrating peptides. J Control Release. 2015;197:106-110.
76. Lin T, Liu E, He H, et al. Nose-to-brain delivery of macromolecules mediated by cell-
penetrating peptides. Acta Pharm Sin B. 2016;6(4):352-358.
77. Lin CY, Cheng YS, Liao TY, et al. Intranasal Administration of a Polyethylenimine-
Conjugated Scavenger Peptide Reduces Amyloid-β Accumulation in a Mouse Model of
Alzheimer’s Disease. J Alzheimer’s Dis. 2016;53(3):1053-1067.
78. McGowan JWD, Shao Q, Vig PJS, Bidwell GL. Intranasal administration of elastin-like
polypeptide for therapeutic delivery to the central nervous system. Drug Des Devel Ther.
2016;10:2803-2813.
79. Sasaki-Hamada S, Nakamura R, Nakao Y, et al. Antidepressant-like effects exerted by the
intranasal administration of a glucagon-like peptide-2 derivative containing cell-penetrating
peptides and a penetration-accelerating sequence in mice. Peptides. 2017;87:64-70.
80. Cheng Y, Chen Z, Liao T, et al. An intranasally delivered peptide drug ameliorates cognitive
decline in Alzheimer transgenic mice. EMBO Mol Med. 2017;9(5):703-715.
81. Gozes I, Bardea A, Reshef A, et al. Neuroprotective strategy for Alzheimer disease:
intranasal administration of a fatty neuropeptide. Proc Natl Acad Sci U S A. 1996;93(1):427-
432.
82. Gozes I, Giladi E, Pinhasov A, Bardea A, Brenneman DE. Activity-dependent neurotrophic
factor: intranasal administration of femtomolar-acting peptides improve performance in a
water maze. J Pharmacol Exp Ther. 2000;293(3):1091-1098.
83. Ross TM, Martinez PM, Renner JC, Thorne RG, Hanson LR, Frey WH. Intranasal
administration of interferon beta bypasses the blood-brain barrier to target the central nervous
system and cervical lymph nodes: A non-invasive treatment strategy for multiple sclerosis. J
Neuroimmunol. 2004;151(1-2):66-77.
84. Fliedner S, Schulz C, Lehnert H. Brain uptake of intranasally applied radioiodinated leptin in
Wistar rats. Endocrinology. 2006;147(5):2088-2094.
85. Fletcher L, Kohli S, Sprague SM, et al. Intranasal delivery of erythropoietin plus insulin-like
growth factor-I for acute neuroprotection in stroke. Laboratory investigation. J Neurosurg.
2009;111(July):164-170.
86. Dhuria S V., Hanson LR, Frey II WH. Intranasal Drug Targeting of Hypocretin-1 (Orexin-A)

to the Central Nervous System. J Pharm Sci. 2009;98(7):2501-2515.
87. Yang JP, Liu HJ, Cheng SM, et al. Direct transport of VEGF from the nasal cavity to brain.
Neurosci Lett. 2009;449(2):108-111.
88. Alcalá-Barraza SR, Lee MS, Hanson LR, et al. Intranasal delivery of neurotrophic factors
BDNF, CNTF, EPO, and NT-4 to the CNS. Drug Target. 2010;18(3):179-190.
89. Yu H, Kim K. Direct nose-to-brain transfer of a growth hormone releasing neuropeptide,
hexarelin after intranasal administration to rabbits. Int J Pharm. 2009;378(1-2):73-79.
90. Niikura T, Sidahmed E, Hirata-Fukae C, Aisen PS, Matsuoka Y. A humanin derivative
reduces amyloid beta accumulation and ameliorates memory deficit in triple transgenic mice.
PLoS One. 2011;6(1):1-12.
91. Rat D, Schmitt U, Tippmann F, et al. Neuropeptide pituitary adenylate cyclase-activating
polypeptide (PACAP) slows down Alzheimer’s disease-like pathology in amyloid precursor
protein-transgenic mice. FASEB J. 2011;25(9):3208-3218.
92. Falcone J a, Salameh TS, Yi X, et al. Intranasal administration as a route for drug delivery to
the brain: Evidence for a unique pathway for albumin. J Pharmacol Exp Ther.
2014;98108(October):54-60.
93. Chauhan MB, Chauhan NB. Brain Uptake of Neurotherapeutics after Intranasal versus
Intraperitoneal Delivery in Mice. J Neurol Neurosurg. 2015;2(1):2-9.
94. Salameh TS, Bullock KM, Hujoel IA, et al. Central Nervous System Delivery of Intranasal
Insulin: Mechanisms of Uptake and Effects on Cognition. J Alzheimer’s Dis. 2015;47(3):715-
728.
95. Dhuria S V., Fine JM, Bingham D, et al. Food consumption and activity levels increase in
rats following intranasal Hypocretin-1. Neurosci Lett. 2016;627:155-159.
96. Marks DR, Tucker K, Cavallin MA, Mast TG, Fadool DA. Awake Intranasal Insulin
Delivery Modifies Protein Complexes and Alters Memory, Anxiety, and Olfactory
Behaviors. J Neurosci. 2009;29(20):6734-6751.
97. Yang J, Lu L, Wang HC, et al. Effect of intranasal arginine vasopressin on human headache.
Peptides. 2012;38(1):100-104.
98. Yang Y, Ma D, Wang Y, et al. Intranasal insulin ameliorates tau hyperphosphorylation in a
rat model of type 2 diabetes. J Alzheimer’s Dis. 2013;33(2):329-338.
99. Maimaiti S, Anderson KL, DeMoll C, et al. Intranasal Insulin Improves Age-Related
Cognitive Deficits and Reverses Electrophysiological Correlates of Brain Aging. Journals
Gerontol - Ser A Biol Sci Med Sci. 2015;71(1):30-39.
100. Zhang Y, Dai C-L, Chen Y, Iqbal K, Liu F, Gong C-X. Intranasal Insulin Prevents
Anesthesia-Induced Spatial Learning and Memory Deficit in Mice. Sci Rep.
2016;6(November 2015):21186.
101. Rajasekar N, Nath C, Hanif K, Shukla R. Intranasal insulin exerts beneficial effects by
improving cerebral blood flow, Nrf-2 expression and BDNF in STZ (ICV) induced memory-
impaired rats. Life Sci. 2016;173:1-10.
102. Pang Y, Lin S, Wright C, et al. Intranasal insulin protects against substantia nigra
dopaminergic neuronal loss and alleviates motor deficits induced by 6-OHDA in rats.
Neuroscience. 2016;318:157-165.
103. Rhea EM, Humann SR, Nirkhe S, Farr SA, Morley JE, Banks WA. Intranasal Insulin
Transport is Preserved in Aged SAMP8 Mice and is Altered by Albumin and Insulin
Receptor Inhibition. J Alzheimer’s Dis. 2017;57(1):241-252.
104. Guo Z, Chen Y, Mao Y-F, et al. Long-term treatment with intranasal insulin ameliorates

cognitive impairment, tau hyperphosphorylation, and microglial activation in a
streptozotocin-induced Alzheimer’s rat model. Sci Rep. 2017;7:45971.
105. Carter GG, Wilkinson GS. Intranasal oxytocin increases social grooming and food sharing in
the common vampire bat Desmodus rotundus. Horm Behav. 2015;75:150-153.
106. Kent P, Awadia A, Zhao L, et al. Effects of intranasal and peripheral oxytocin or gastrin-
releasing peptide administration on social interaction and corticosterone levels in rats.
Psychoneuroendocrinology. 2016;64:123-130.
107. Hess L, Votava M, Málek J, Kurzová A, Slíva J. Sedative Effects of Intranasal Oxytocin in
Rabbits and Rhesus Monkeys. Physiol Res. 2016;65:473-480.
108. Camerlink I, Reimert I, Bolhuis JE. Intranasal oxytocin administration in relationship to
social behaviour in domestic pigs. Physiol Behav. 2016;163:51-55.
109. Lee MR, Scheidweiler KB, Diao XX, et al. Oxytocin by intranasal and intravenous routes
reaches the cerebrospinal fluid in rhesus macaques: determination using a novel oxytocin
assay. Mol Psychiatry. 2017;(September 2016):1-8.
110. Yu YP, Xu QQ, Zhang Q, Zhang WP, Zhang LH, Wei EQ. Intranasal recombinant human
erythropoietin protects rats against focal cerebral ischemia. Neurosci Lett. 2005;387(1):5-10.
111. Merelli A, Caltana L, Lazarowski A, Brusco A. Experimental evidence of the potential use of
erythropoietin by intranasal administration as a neuroprotective agent in cerebral hypoxia.
Drug Metabol Drug Interact. 2011;26(2):65-69.
112. Gao Y, Mengana Y, Cruz YR, et al. Different expression patterns of Ngb and EPOR in the
cerebral cortex and hippocampus revealed distinctive therapeutic effects of intranasal
delivery of Neuro-EPO for ischemic insults to the gerbil brain. J Histochem Cytochem.
2011;59(2):214-227.
113. Maurice T, Mustafa M-H, Desrumaux C, et al. Intranasal formulation of erythropoietin
(EPO) showed potent protective activity against amyloid toxicity in the Aβ₂₅₋₃₅ non-
transgenic mouse model of Alzheimer’s disease. J Psychopharmacol. 2013;27(11):1044-
1057.
114. Rodriguez Cruz Y, Strehaiano M, Rodriguez Obaya T, Garcia Rodriguez JC, Maurice T. An
Intranasal Formulation of Erythropoietin (Neuro-EPO) Prevents Memory Deficits and
Amyloid Toxicity in the APP(Swe) Transgenic Mouse Model of Alzheimer’s Disease. J
Alzheimers Dis. 2017;55(1):231-248.
115. De Rosa R, Garcia AA, Braschi C, et al. Intranasal administration of nerve growth factor
(NGF) rescues recognition memory deficits in AD11 anti-NGF transgenic mice. Proc Natl
Acad Sci. 2005;102(10):3811-3816.
116. Covaceuszach S, Capsoni S, Ugolini G, Spirito F, Vignone D, Cattaneo A. Development of a
Non Invasive NGF-Based Therapy for Alzheimer ’ s Disease. Curr Alzheimer Res.
2009;6:158-170.
117. Zhu W, Cheng S, Xu G, et al. Intranasal nerve growth factor enhances striatal neurogenesis in
adult rats with focal cerebral ischemia. Drug Deliv. 2011;18(January):338-343.
118. Tian L, Guo R, Yue X, et al. Intranasal administration of nerve growth factor ameliorate β-
amyloid deposition after traumatic brain injury in rats. Brain Res. 2012;1440:47-55.
119. Capsoni S, Marinelli S, Ceci M, et al. Intranasal “painless” human nerve growth factors
slows amyloid neurodegeneration and prevents memory deficits in app x PS1 mice. PLoS
One. 2012;7(5).
120. Lv Q, Fan X, Xu G, et al. Intranasal delivery of nerve growth factor attenuates aquaporins-4-
induced edema following traumatic brain injury in rats. Brain Res. 2013;1493:80-89.
121. Ma Y-P, Ma M-M, Cheng S-M, et al. Intranasal bFGF-induced progenitor cell proliferation

and neuroprotection after transient focal cerebral ischemia. Neurosci Lett. 2008;437(2):93-97.
122. Zhang C, Chen J, Feng C, et al. Intranasal nanoparticles of basic fibroblast growth factor for
brain delivery to treat Alzheimer’s disease. Int J Pharm. 2014;461(1-2):192-202.
123. Zhao YZ, Li X, Lu CT, et al. Gelatin nanostructured lipid carriers-mediated intranasal
delivery of basic fibroblast growth factor enhances functional recovery in hemiparkinsonian
rats. Nanomedicine Nanotechnology, Biol Med. 2014;10(4):755-764.
124. Zhao YZ, Lin M, Lin Q, et al. Intranasal delivery of bFGF with nanoliposomes enhances in
vivo neuroprotection and neural injury recovery in a rodent stroke model. J Control Release.
2016;224:165-175.
125. Alcalay RN, Giladi E, Pick CG, Gozes I. Intranasal administration of NAP, a neuroprotective
peptide, decreases anxiety-like behavior in aging mice in the elevated plus maze. Neurosci
Lett. 2004;361(1-3):128-131.
126. Matsuoka Y, Gray AJ, Hirata-Fukae C, et al. Intranasal NAP administration reduces
accumulation of amyloid peptide and tau hyperphosphorylation in a transgenic mouse model
of Alzheimer’s disease at early pathological stage. J Mol Neurosci. 2007;31(2):165-170.
127. Matsuoka Y, Jouroukhin Y, Gray AJ, et al. A Neuronal Microtubule-Interacting Agent,
NAPVSIPQ, Reduces Tau Pathology and Enhances Cognitive Function in a Mouse Model of
Alzheimer’s Disease. J Pharmacol Exp Ther. 2008;325(1):146-153.
128. Sharma NK, Sethy NK, Meena RN, Ilavazhagan G, Das M, Bhargava K. Activity-dependent
neuroprotective protein (ADNP)-derived peptide (NAP) ameliorates hypobaric hypoxia
induced oxidative stress in rat brain. Peptides. 2011;32(6):1217-1224.
129. Liu Z, Jiang M, Kang T, et al. Lactoferrin-modified PEG-co-PCL nanoparticles for enhanced
brain delivery of NAP peptide following intranasal administration. Biomaterials.
2013;34(15):3870-3881.
130. Magen I, Ostritsky R, Richter F, et al. Intranasal NAP (davunetide) decreases tau
hyperphosphorylation and moderately improves behavioral deficits in mice overexpressing α-
synuclein. Pharmacol Res Perspect. 2014;2(5):e00065.
131. Gao X, Wu B, Zhang Q, et al. Brain delivery of vasoactive intestinal peptide enhanced with
the nanoparticles conjugated with wheat germ agglutinin following intranasal administration.
J Control Release. 2007;121(3):156-167.
132. Cui X, Cao DY, Wang ZM, Zheng AP. Pharmacodynamics and toxicity of vasoactive
intestinal peptide for intranasal administration. Pharmazie. 2013;68(1):69-74.
133. Liu X-F, Fawcett JR, Hanson LR, Frey WH. The window of opportunity for treatment of
focal cerebral ischemic damage with noninvasive intranasal insulin-like growth factor-I in
rats. J Stroke Cerebrovasc Dis. 2004;13(1):16-23.
134. Cai Z, Fan L-W, Lin S, Pang Y, Rhodes PG. Intranasal Administration of IGF-1 Protects
against Lipopolysaccharide-induced Injury in the Developing Rat Brain. Neuroscience.
2011;194:195–207.
135. Lopes C, Ribeiro M, Duarte AI, et al. IGF-1 intranasal administration rescues Huntington’s
disease phenotypes in YAC128 mice. Mol Neurobiol. 2014;49(3):1126-1142.
136. Gartziandia O, Herran E, Pedraz JL, Carro E, Igartua M, Hernandez RM. Chitosan coated
nanostructured lipid carriers for brain delivery of proteins by intranasal administration.
Colloids Surfaces B Biointerfaces. 2015;134:304-313.
137. During MJ, Cao L, Zuzga DS, et al. Glucagon-like peptide-1 receptor is involved in learning
and neuroprotection. Nat Med. 2003;9(9):1173-1179.
138. Schulz C, Paulus K, Lehnert H. Central nervous and metabolic effects of intranasally applied
leptin. Endocrinology. 2004;145(6):2696-2701.

139. Chen H, Yang GZX, Getachew H, Acosta C, Sierra Sánchez C, Konofagou EE. Focused
ultrasound-enhanced intranasal brain delivery of brain-derived neurotrophic factor. Sci Rep.
2016;6(February):28599.
140. Cheng Q, Feng J, Chen J, Zhu X, Li F. Brain Transport of Neurotoxin-I with PLA
Nanoparticles through Intranasal Administration in Rats: A Microdialysis Study. Biopharm
Drug Dispos. 2008;29:431–439.
141. Ruan Y, Yao L, Zhang B, Zhang S, Guo J. Antinociceptive properties of nasal delivery of
Neurotoxin-loaded nanoparticles coated with polysorbate-80. Peptides. 2011;32(7):1526-
1529.
142. Zhang X, Liu L, Chai G, Zhang X, Li F. Brain pharmacokinetics of neurotoxin-loaded PLA
nanoparticles modified with chitosan after intranasal administration in awake rats. Drug Dev
Ind Pharm. 2013;39(September 2011):1618-1624.
143. Wu H, Li J, Zhang Q, et al. A novel small Odorranalectin-bearing cubosomes: Preparation,
brain delivery and pharmacodynamic study on amyloid-β 25-35-treated rats following
intranasal administration. Eur J Pharm Biopharm. 2012;80(2):368-378.
144. Deadwyler SA, Porrino L, Siegel JM, Hampson RE. Systemic and Nasal Delivery of Orexin-
A (Hypocretin-1) Reduces the Effects of Sleep Deprivation on Cognitive Performance in
Nonhuman Primates. J Neurosci. 2007;27(52):14239-14247.
145. Migliore MM, Ortiz R, Dye S, Campbell RB, Amiji MM, Waszczak BL. Neurotrophic and
neuroprotective efficacy of intranasal GDNF in a rat model of Parkinson’s disease.
Neuroscience. 2014;274:11-23.
146. Gartziandia O, Herrán E, Ruiz-Ortega JA, et al. Intranasal administration of chitosan-coated
nanostructured lipid carriers loaded with GDNF improves behavioral and histological
recovery in a partial lesion model of Parkinson’s disease. J Biomed Nanotechnol.
2016;12(12):2220-2230.
147. Hernando S, Herran E, Figueiro-Silva J, et al. Intranasal Administration of TAT-Conjugated
Lipid Nanocarriers Loading GDNF for Parkinson’s Disease. Mol Neurobiol. 2017:1-11.
148. Ma YP, Ma MM, Ge S, et al. Intranasally delivered TGF-β1 enters brain and regulates gene
expressions of its receptors in rats. Brain Res Bull. 2007;74(4):271-277.
149. Yang JP, Liu HJ, Wang ZL, et al. The dose-effectiveness of intranasal VEGF in treatment of
experimental stroke. Neurosci Lett. 2009;461(3):212-216.
150. Veronesi MC, Aldouby Y, Domb AJ, Kubek MJ. Thyrotropin-releasing hormone d,l
polylactide nanoparticles (TRH-NPs) protect against glutamate toxicity in vitro and kindling
development in vivo. Brain Res. 2009;1303:151-160.
151. Kubek MJ, Domb AJ, Veronesi MC. Attenuation of Kindled Seizures by Intranasal Delivery
of Neuropeptide-Loaded Nanoparticles. Neurotherapeutics. 2009;6(2):359-371.
152. Lu CT, Jin RR, Jiang YN, et al. Gelatin nanoparticle-mediated intranasal delivery of
substance P protects against 6-hydroxydopamine-induced apoptosis: An in vitro and in vivo
study. Drug Des Devel Ther. 2015;9:1955-1962.
153. Zhao YZ, Jin RR, Yang W, et al. Using gelatin nanoparticle mediated intranasal delivery of
neuropeptide substance P to enhance neuro-recovery in hemiparkinsonian rats. PLoS One.
2016;11(2):1-18.
154. Rangasamy SB, Corbett GT, Roy A, et al. Intranasal Delivery of NEMO-Binding Domain
Peptide Prevents Memory Loss in a Mouse Model of Alzheimer’s Disease. J Alzheimers Dis.
2015;47(2):385-402.
155. Joachim E, Kim ID, Jin Y, Kim K (Kevin), Lee JK, Choi H. Gelatin nanoparticles enhance
the neuroprotective effects of intranasally administered osteopontin in rat ischemic stroke

model. Drug Deliv Transl Res. 2014;4(5-6):395-399.
156. Jin YC, Lee H, Kim SW, et al. Intranasal Delivery of RGD Motif-Containing Osteopontin
Icosamer Confers Neuroprotection in the Postischemic Brain via αvβ3 Integrin Binding. Mol
Neurobiol. 2016;53(8):5652-5663.
157. Jarcho MR, Mendoza SP, Mason WA, Yang X, Bales KL. Intranasal Vasopressin Affects
Pair-Bonding And Peripheral Gene Expression In Male Callicebus Cupreus. Genes Brain
Behav. 2011;10(3):375-383.
158. Ludwig M, Tobin VA, Callahan MF, et al. Intranasal application of vasopressin fails to elicit
changes in brain immediate early gene expression, neural activity and behavioral
performance of rats. J Neuroendocr. 2014;25(7):655-667.
159. Migliore MM, Vyas TK, Cambell RB, Amiji MM, Waszczak BL. Brain Delivery of Proteins
by the Intranasal Route of Administration: A Comparison of Cationic Liposomes Versus
Aqueous Solution Formulations. J Pharm Sci. 2009;99(4):1745–1761.
160. Kumar M, Pandey RS, Patra KC, et al. Evaluation of neuropeptide loaded trimethyl chitosan
nanoparticles for nose to brain delivery. Int J Biol Macromol. 2013;61:189-195.
161. Lee JH, Kam EH, Kim JM, et al. Intranasal Administration of Interleukin-1 Receptor
Antagonist in a Transient Focal Cerebral Ischemia Rat Model. Biomol Ther. 2017;25(2):149-
157.
162. Guerra-Crespo M, Sistos A, Gleason D, Fallon JH. Intranasal administration of PEGylated
transforming growth factor-alpha improves behavioral deficits in a chronic stroke model. J
stroke Cerebrovasc Dis. 2010;19(1):3-9.
163. Hanson LR, Fine JM, Hoekman JD, et al. Intranasal delivery of growth differentiation factor
5 to the central nervous system. Drug Deliv. 2012;19(January):149-154.
164. Nonaka N, Farr SA, Kageyama H, Shioda S, Banks WA. Delivery of galanin-like peptide to
the brain: targeting with intranasal delivery and cyclodextrins. J Pharmacol Exp Ther.
2008;325(2):513-519.
165. Sun BL, Shen FP, Wu QJ, et al. Intranasal delivery of calcitonin gene-related peptide reduces
cerebral vasospasm in rats. Front Biosci (Elite Ed). 2010;2:1502-1513.
166. Zhang H, Meng J, Zhou S, et al. Intranasal Delivery of Exendin-4 Confers Neuroprotective
Effect Against Cerebral Ischemia in Mice. AAPS J. 2016;18(2):385-394.
167. Wen Z, Yan Z, Hu K, et al. Odorranalectin-conjugated nanoparticles: Preparation, brain
delivery and pharmacodynamic study on Parkinson’s disease following intranasal
administration. J Control Release. 2011;151(2):131-138.
168. Serova LI, Laukova M, Alaluf LG, Pucillo L, Sabban EL. Intranasal neuropeptide Y reverses
anxiety and depressive-like behavior impaired by single prolonged stress PTSD model. Eur
Neuropsychopharmacol. 2014;24(1):142-147.
169. Dine J, Ionescu IA, Avrabos C, et al. Intranasally applied neuropeptide S shifts a high-
anxiety electrophysiological endophenotype in the ventral hippocampus towards a “normal”-
anxiety one. PLoS One. 2015;10(4):1-10.
170. Shaw P, Zhang X. Intranasal Delivery of a Cystatin C-peptide as Therapy for Alzheimer’s
Disease. J Fed Am Soc Exp Biol. 2013;1(Supplement):533.1.
171. Zheng X, Shao X, Zhang C, et al. Intranasal H102 Peptide-Loaded Liposomes for Brain
Delivery to Treat Alzheimer’s Disease. Pharm Res. 2015;32(12):3837-3849.
172. Brown V, Liu F. Intranasal delivery of a Peptide with antidepressant-like effect.
Neuropsychopharmacology. 2014;39(9):2131-2141.
173. Chen D, Lee J, Gu X, Wei L, Yu SP. Intranasal Delivery of Apelin-13 Is Neuroprotective and

Promotes Angiogenesis After Ischemic Stroke in Mice. ASN Neuro. 2015;7(5):1-15.
174. Sipos E, Kurunczi A, Fehér A, et al. Intranasal delivery of human b-amyloid peptide in rats:
Effective brain targeting. Cell Mol Neurobiol. 2010;30(3):405-413.
175. Yadav S, Gattacceca F, Panicucci R, Amiji MM. Comparative biodistribution and
pharmacokinetic analysis of cyclosporine-A in the brain upon intranasal or intravenous
administration in an oil-in-water nanoemulsion formulation. Mol Pharm. 2015;12(5):1523-
1533.
176. Anagnostou E, Soorya L, Brian J, et al. Intranasal oxytocin in the treatment of autism
spectrum disorders: A review of literature and early safety and efficacy data in youth. Brain
Res. 2014;1580:188-198.
177. Chapman CD, Frey WH, Craft S, et al. Intranasal treatment of central nervous system
dysfunction in humans. Pharm Res. 2013;30(10):2475-2484.
178. German Diabetes Center. The Effect of Central Insulin on Insulin Sensitivity and Energy
Metabolism (Centrin). ClinicalTrials.gov Identifier: NCT01479075.
https://clinicaltrials.gov/ct2/show/NCT01479075. Published 2011.
179. Heni M, Wagner R, Kullmann S, et al. Central insulin administration improves whole-body
insulin sensitivity via hypothalamus and parasympathetic outputs in men. Diabetes.
2014;63(12):4083-4088.
180. Gancheva S, Koliaki C, Bierwagen A, et al. Effects of intranasal insulin on hepatic fat
accumulation and energy metabolism in humans. Diabetes. 2015;64(6):1966-1975.
181. Bakermans-Kranenburg MJ, van IJzendoorn MH. Sniffing around oxytocin: review and
meta-analyses of trials in healthy and clinical groups with implications for pharmacotherapy.
Transl Psychiatry. 2013;3(5):e258.
182. Warren K. Effects of Intranasal Oxytocin on Satiety Signaling in People With Schizophrenia.
ClinicalTrials.gov Identifier: NCT01614093.
https://clinicaltrials.gov/ct2/show/NCT01614093. Published 2012.
183. University N. The Effect of Intranasal Oxytocin on Pain Sensitivity and Threshold.
ClinicalTrials.gov Identifier: NCT02550093.
https://clinicaltrials.gov/ct2/show/NCT02550093?term=nasal+and+peptid. Published 2015.
184. Hallschmid M, Benedict C, Born J, Fehm HL, Kern W. Manipulating central nervous
mechanisms of food intake and body weight regulation by intranasal administration of
neuropeptides in man. Physiol Behav. 2004;83(1 SPEC. ISS.):55-64.
185. Benedict C, Hallschmid M, Hatke A, et al. Intranasal insulin improves memory in humans.
Psychoneuroendocrinology. 2004;29(10):1326-1334.
186. Reger MA, Watson GS, Frey WH, et al. Effects of intranasal insulin on cognition in memory-
impaired older adults: Modulation by APOE genotype. Neurobiol Aging. 2006;27(3):451-
458.
187. Benedict C, Kern W, Schultes B, Born J, Hallschmid M. Differential sensitivity of men and
women to anorexigenic and memory-improving effects of intranasal insulin. J Clin
Endocrinol Metab. 2008;93(4):1339-1344.
188. Bohringer A, Schwabe L, Richter S, Schachinger H. Intranasal insulin attenuates the
hypothalamic-pituitary-adrenal axis response to psychosocial stress.
Psychoneuroendocrinology. 2008;33(10):1394-1400.
189. Fan X, Copeland PM, Liu EY, et al. No effect of single-dose intranasal insulin treatment on
verbal memory and sustained attention in patients with schizophrenia. J Clin
Psychopharmacol. 2011;31(2):231-234.
190. Schöpf V, Kollndorfer K, Pollak M, Mueller CA, Freiherr J. Intranasal insulin influences the

olfactory performance of patients with smell loss, dependent on the body mass index: A pilot
study. Rhinology. 2015;53(4):371-378.
191. Hamidovic A. Position on zinc delivery to olfactory nerves in intranasal insulin phase I-III
clinical trials. Contemp Clin Trials. 2015;45:277-280.
192. Dash S, Xiao C, Morgantini C, Koulajian K, Lewis GF. Intranasal insulin suppresses
endogenous glucose production in humans compared with placebo in the presence of similar
venous insulin concentrations. Diabetes. 2015;64(3):766-774.
193. Kern W, Born J, Schreiber H, Fehm HL. Central nervous system effects of intranasally
administered insulin during euglycemia in men. Diabetes. 1999;48(3):557-563.
194. Benedict C, Hallschmid M, Schmitz K, et al. Intranasal insulin improves memory in humans:
superiority of insulin aspart. Neuropsychopharmacology. 2007;32(1):239-243.
195. Henkin RI. Intranasal insulin: From nose to brain. Nutrition. 2010;26(6):624-633.
196. Hallschmid M, Higgs S, Thienel M, Ott V, Lehnert H. Postprandial administration of
intranasal insulin intensifies satiety and reduces intake of palatable snacks in women.
Diabetes. 2012;61(4):782-789.
197. Novak P. Treatment of Parkinson Disease and Multiple System Atrophy Using Intranasal
Insulin. ClinicalTrials.gov Identifier: NCT02064166.
https://clinicaltrials.gov/ct2/show/record/NCT02064166. Published 2014.
198. Craft S, Aisen P. The Study of Nasal Insulin in the Fight Against Forgetfulness (SNIFF).
ClinicalTrials.gov Identifier: NCT01767909.
https://clinicaltrials.gov/ct2/show/NCT01767909. Published 2014.
199. Zhang H, Hao Y, Manor B, et al. Intranasal insulin enhanced resting-state functional
connectivity of hippocampal regions in type 2 diabetes. Diabetes. 2015;64(3):1025-1034.
200. Reger MA, Watson GS, Green PS, et al. Intranasal Insulin Administration Dose-Dependently
Modulates Verbal Memory and Plasma β-Amyloid in Memory-Impaired Older Adults. J
Alzheimers Dis. 2008;13(3):323-331.
201. Kosfeld M, Heinrichs M, Zak PJ, Fischbacher U, Fehr E. Oxytocin increases trust in humans.
Nature. 2005;435(7042):673-677.
202. Pedersen CA, Gibson CM, Rau SW, et al. Intranasal oxytocin reduces psychotic symptoms
and improves Theory of Mind and social perception in schizophrenia. Schizophr Res.
2011;132(1):50-53.
203. Rilling J, DeMarco A. Effects of intranasal oxytocin and vasopressin on cooperative behavior
and associated brain activity in men. Psychoneuroendocrinology. 2012;37(4):447-461.
204. Pedersen CA, Smedley KL, Leserman J, et al. Intranasal Oxytocin Blocks Alcohol
Withdrawal in Human Subjects. Alcohol Clin Exp Res. 2013;37(3):484-489.
205. Tachibana M, Kagitani-Shimono K, Mohri I, et al. Long-term administration of intranasal
oxytocin is a safe and promising therapy for early adolescent boys with autism spectrum
disorders. J Child Adolesc Psychopharmacol. 2013;23(2):123-127.
206. Murgatroyd CAD rer. nat, Hicks-Nelson A, Fink A, et al. Effects of chronic social stress and
maternal intranasal oxytocin and vasopressin on offspring interferon γ and behavior. Front
Endocrinol (Lausanne). 2016;7(December):155.
207. Leppanen J, Cardi V, Ng KW, et al. Effects of intranasal oxytocin on interpretation and
expression of emotions in anorexia nervosa. J Neuroendocrinol. 2017;(6):1-13.
208. Jarskog LF, Pedersen CA, Johnson JL, et al. A 12-week randomized controlled trial of twice-
daily intranasal oxytocin for social cognitive deficits in people with schizophrenia. Schizophr
Res. 2017;pii: S0920(17):30007-5.

209. Sack M, Spieler D, Wizelman L, et al. Intranasal oxytocin reduces provoked symptoms in
female patients with posttraumatic stress disorder despite exerting sympathomimetic and
positive chronotropic effects in a randomized controlled trial. BMC Med. 2017;15(1):40.
210. Li T, Chen X, Mascaro J, Haroon E, Rilling JK. Intranasal oxytocin, but not vasopressin,
augments neural responses to toddlers in human fathers. Horm Behav. 2017;S0018-
506X(16):30445-30447.
211. Williams DR, Bürkner PC. Effects of intranasal oxytocin on symptoms of schizophrenia: A
multivariate Bayesian meta-analysis. Psychoneuroendocrinology. 2017;75:141-151.
212. Domes G, Heinrichs M, Michel A, Berger C, Herpertz SC. Oxytocin Improves “Mind-
Reading” in Humans. Biol Psychiatry. 2007;61(6):731-733.
213. Guastella AJ, Howard AL, Dadds MR, Mitchell P, Carson DS. A randomized controlled trial
of intranasal oxytocin as an adjunct to exposure therapy for social anxiety disorder.
Psychoneuroendocrinology. 2009;34(6):917-923.
214. Guastella AJ, Einfeld SL, Gray KM, et al. Intranasal Oxytocin Improves Emotion
Recognition for Youth with Autism Spectrum Disorders. Biol Psychiatry. 2010;67(7):692-
694.
215. Woolley JD, Lam O, Chuang B, Ford JM, Mathalon DH, Vinogradov S. Oxytocin
administration selectively improves olfactory detection thresholds for lyral in patients with
schizophrenia. Psychoneuroendocrinology. 2015;53:217-222.
216. Kosaka H, Okamoto Y, Munesue T, et al. Oxytocin efficacy is modulated by dosage and
oxytocin receptor genotype in young adults with high-functioning autism: a 24-week
randomized clinical trial. Transl Psychiatry. 2016;6(8):e872.
217. Muin DA, Sheikh Rezaei S, Tremmel-Scheinost M, et al. Men’s sexual response to female
partner’s intranasal oxytocin administration for hypoactive sexual desire disorder: an open
prospective cohort study. Fertil Steril. 2017;107(3):781-787.
218. Guastella AJ, Gray KM, Rinehart NJ, et al. The effects of a course of intranasal oxytocin on
social behaviors in youth diagnosed with autism spectrum disorders: A randomized
controlled trial. J Child Psychol Psychiatry Allied Discip. 2015;56(4):444-452.
219. van Zuiden M, Frijling JL, Nawijn L, et al. The efficacy of intranasal oxytocin administration
at preventing PTSD in trauma-exposed emergency department patients: a randomized
controlled trial. Biol Psychiatry. 2016;S0006-3223(16):33067-33070.
220. Gordon I, Jack A, Pretzsch CM, et al. Intranasal oxytocin enhances connectivity in the neural
circuitry supporting social motivation and social perception in children with autism. Sci Rep.
2016;6(February):35054.
221. Pietrowsky R, Strüben C, Mölle M, Fehm HL, Born J. Brain potential changes after
intranasal vs. intravenous administration of vasopressin: Evidence for a direct nose-brain
pathway for peptide effects in humans. Biol Psychiatry. 1996;39(5):332-340.
222. Guastella AJ, Kenyon AR, Alvares GA, Carson DS, Hickie IB. Intranasal Arginine
Vasopressin Enhances the Encoding of Happy and Angry Faces in Humans. Biol Psychiatry.
2010;67(12):1220-1222.
223. Smolnik R, Perras B, Molle M, Fehm H, Born J. Event-related brain potentials and working
memory function in healthy humans after single-dose and prolonged intranasal administration
of adrenocorticotropin 4-10 and desacetyl-alpha-melanocyte stimulating hormone. J Clin
Psychopharmacol. 2000;20(4):445-454.
224. Wellhöner P, Hörster R, Jacobs F, Sayk F, Lehnert H, Dodt C. Intranasal application of the
melanocortin 4 receptor agonist MSH/ACTH(4-10) in humans causes lipolysis in white
adipose tissue. Int J Obes (Lond). 2012;36(5):703-708.

225. Denecke H, Czehak N, Pietrowsky R. Dose-response relationships of intranasal
cholecystokinin and the P300 event-related brain potential. Pharmacol Biochem Behav.
2002;73(3):593-600.
226. Denecke H, Meyer F, Feldkamp J, Fritzen R, Pietrowsky R. Repetitive intranasal
administration of cholecystokinin potentiates its central nervous effects. Physiol Behav.
2004;83(1 SPEC. ISS.):39-45.
227. Pietrowsky R, Claassen L, Frercks H, Fehm HL, Born J. Time course of intranasally
administered cholecystokinin-8 on central nervous effects. Neuropsychobiology.
2001;43(4):254-259.
228. Gozes I, Morimoto BH, Tiong J, et al. NAP: Research and Development of a Peptide Derived
from Activity-Dependent Neuroprotective Protein (ADNP). CNS Drug Rev. 2005;11(4):353-
368.
229. University of California LA. A Multicenter Study of NAP (AL-108) in Schizophrenia (AL-
108). ClinicalTrials.gov Identifier: NCT00505765.
https://clinicaltrials.gov/ct2/show/results/NCT00505765?term=davunetide&rank=3.
Published 2007.
230. Therapeutics A. Safety, Tolerability and Efficacy Study to Evaluate Subjects With Mild
Cognitive Impairment. ClinicalTrials.gov Identifier: NCT00422981.
https://clinicaltrials.gov/ct2/show/NCT00422981?term=davunetide&rank=5. Published 2007.
231. Baier PC, Weinhold SL, Huth V, Gottwald B, Ferstl R, Hinze-Selch D. Olfactory dysfunction
in patients with narcolepsy with cataplexy is restored by intranasal Orexin A (Hypocretin-1).
Brain. 2008;131(10):2734-2741.
232. Baier PC, Hallschmid M, Seeck-Hirschner M, et al. Effects of intranasal hypocretin-1 (orexin
A) on sleep in narcolepsy with cataplexy. Sleep Med. 2011;12(10):941-946.
233. Weinhold SL, Seeck-Hirschner M, Nowak A, Hallschmid M, Göder R, Baier PC. The effect
of intranasal orexin-A (hypocretin-1) on sleep, wakefulness and attention in narcolepsy with
cataplexy. Behav Brain Res. 2014;262:8-13.
234. Ghigo E, Arvat E, Gianotti L, et al. Growth hormone-releasing activity of hexarelin, a new
synthetic hexapeptide, after intravenous, subcutaneous, intranasal, and oral administration in
man. J Clin Endocrinol Metab. 1994;78(3):693-698.
235. FDA. Nonclinical Safety Evaluation of Reformulated Drug Products and Products Intended
for Administration by an Alternate Route - Guidance for Industry and Review Staff Good
Review Practice. Pharmacology/Toxicology.
https://www.fda.gov/downloads/drugs/guidancecomplianceregulatoryinformation/guidances/
ucm079245.pdf. Published 2015.
236. Trows S, Wuchner K, Spycher R, Steckel H. Analytical challenges and regulatory
requirements for nasal drug products in Europe and the U.S. Pharmaceutics. 2014;6(2):195-
219.
237. Hochhaus G, Davis-Cutting C, Oliver M, Lee SL, Lyapustina S. Current Scientific and
Regulatory Approaches for Development of Orally Inhaled and Nasal Drug Products:
Overview of the IPAC-RS/University of Florida Orlando Inhalation Conference. AAPS J.
2015;17(5):1305-1311.
238. Ahmad E, Feng Y, Qi J, et al. Evidence of nose-to-brain delivery of nanoemulsions: cargoes
but not vehicles. Nanoscale. 2017;9:1174-1183.
239. Mistry A, Glud SZ, Kjems J, et al. Effect of physicochemical properties on intranasal
nanoparticle transit into murine olfactory epithelium. J Drug Target. 2009;17(7):543-552.
240. Bhavna, Sharma V, Ali M, Baboota S, Ali J. Preparation and characterization of chitosan
nanoparticles for nose to brain delivery of a cholinesterase inhibitor. Indian J Pharm Sci.

2007;69(5):712-713.
241. Bshara H, Osman R, Mansour S, El-Shamy AEHA. Chitosan and cyclodextrin in intranasal
microemulsion for improved brain buspirone hydrochloride pharmacokinetics in rats.
Carbohydr Polym. 2014;99:297-305.
242. Kozlovskaya L, Abou-Kaoud M, Stepensky D. Quantitative analysis of drug delivery to the
brain via nasal route. J Control Release. 2014;189:133-140.
243. De Lorenzo AJ. Electron microscopy of the olfactory and gustatory pathways. Ann Otol
Rhinol Laryngol. 1960;69:410-420.
244. Morrison EE, Costanzo RM. Morphology of olfactory epithelium in humans and other
vertebrates. Microsc Res Tech. 1992;23(1):49-61.
245. Hanafy AS, Farid RM, ElGamal SS. Complexation as an approach to entrap cationic drugs
into cationic nanoparticles administered intranasally for Alzheimer’s disease management:
preparation and detection in rat brain. Drug Dev Ind Pharm. 2015;9045(March 2016):1-14.
246. Bonaccorso A, Musumeci T, Serapide MF, Pellitteri R, Uchegbu IF, Puglisi G. Nose to brain
delivery in rats: Effect of surface charge of rhodamine B labeled nanocarriers on brain
subregion localization. Colloids Surfaces B Biointerfaces. 2017;154:297-306.
247. Mistry A, Stolnik S, Illum L. Nose-to-Brain Delivery: Investigation of the Transport of
Nanoparticles with Different Surface Characteristics and Sizes in Excised Porcine Olfactory
Epithelium. Mol Pharm. 2015;12(8):2755-2766.
248. Gartziandia O, Egusquiaguirre SP, Bianco J, et al. Nanoparticle transport across in vitro
olfactory cell monolayers. Int J Pharm. 2016;499(1-2):81-89.
249. Betbeder D, Sperandio S, Latapie JP, de Nadaí J, Zajac J, Etienne E, Francés B. Biovector TM
nanoparticles improve antinociceptive effi cacy of nasal morphine. Pharm Res. 2000;17:743-
748.
250. Zhang Q-Z, Zha L-S, Zhang Y, et al. The brain targeting efficiency following nasally applied
MPEG-PLA nanoparticles in rats. J Drug Target. 2006;14(5):281-290.
251. Gao X, Tao W, Lu W, et al. Lectin-conjugated PEG-PLA nanoparticles: Preparation and
brain delivery after intranasal administration. Biomaterials. 2006;27(18):3482-3490.
252. Tobío M, Gref T, Sánchez A, Langer R, Alonso MJ. Stealth PLA-PEG Nanoparticles as
Protein Carriers for Nasal Administration. Pharm Res. 1998;15(2):270-275.
253. Tobı́o M, Sánchez A, Vila A, et al. The role of PEG on the stability in digestive fluids and in
vivo fate of PEG-PLA nanoparticles following oral administration. Colloids Surfaces B
Biointerfaces. 2000;18(3-4):315-323.
254. Gulyaev A, Gelperina S, Skidan I, Antropov A, Kivman G, Kreuter J. Significant transport of
doxorubicin into the brain with polysorbate 80-coated nanoparticles. Pharm Res.
1999;16(10):1564-1569.
255. Sun W, Xie C, Wang H, Hu Y. Specific role of polysorbate 80 coating on the targeting of
nanoparticles to the brain. Biomaterials. 2004;25(15):3065-3071.
256. Gao K, Jiang X. Influence of particle size on transport of methotrexate across blood brain
barrier by polysorbate 80-coated polybutylcyanoacrylate nanoparticles. Int J Pharm.
2006;310(1-2):213-219.
257. Garcia-Fuentes M, Alonso MJ. Chitosan-based drug nanocarriers: Where do we stand? J
Control Release. 2012;161(2):496-504.
258. Calvo P, Remunan-Lopez C, Vila-Jato JL, Alonso MJ. Development of positively charged
colloidal drug carriers: Chitosan-coated polyester nanocapsules and submicron emulsions.
Colloid Polym Sci. 1997;275(1):46-53.

259. Teijeiro-Osorio D, Remuñán-López C, Alonso MJ. New generation of hybrid
poly/oligosaccharide nanoparticles as carriers for the nasal delivery of macromolecules.
Biomacromolecules. 2009;10(2):243-249.
260. Meller R, Stevens SL, Minami M, et al. Neuroprotection by osteopontin in stroke. J Cereb
Blood Flow Metab. 2005;25(2):217-225.
261. Kumar M, Misra A, Babbar AK, Mishra AK, Mishra P, Pathak K. Intranasal nanoemulsion
based brain targeting drug delivery system of risperidone. Int J Pharm. 2008;358(1-2):285-
291.
262. Kumar M, Misra A, Mishra AK, Mishra P, Pathak K. Mucoadhesive nanoemulsion-based
intranasal drug delivery system of olanzapine for brain targeting. J Drug Target.
2008;16(10):806-814.
263. Patel A, Patel M, Yang X, Mitra A. Recent advances in protein and peptide drug delivery: a
special emphasis on polymeric nanoparticles. Protein Pept Lett. 2014;21(11):1102-1120.
264. Bi C. Intranasal delivery of rotigotine to the brain with lactoferrin-modified PEG-PLGA
nanoparticles for Parkinson ’ s disease treatment. Int J Nanomedicine. 2016;11:6547-6559.
265. Ferrari CC, Carmanchahi PD, Aldana Marcos HJ, Mugnaini MT, Affanni JM, Paz D a.
Identification and localisation of glycoconjugates in the olfactory mucosa of the armadillo
Chaetophractus villosus. J Anat. 1999;194 ( Pt 3(3):395-405.
266. Chen J, Zhang C, Liu Q, et al. Solanum tuberosum lectin-conjugated PLGA nanoparticles for
nose-to-brain delivery: in vivo and in vitro evaluations. J Drug Target. 2012;20(2):174-184.
267. Liu Q, Shen Y, Chen J, et al. Nose-to-brain transport pathways of wheat germ agglutinin
conjugated PEG-PLA nanoparticles. Pharm Res. 2012;29(2):546-558.
268. Piazza J, Hoare T, Molinaro L, et al. Haloperidol-loaded intranasally administered lectin
functionalized poly(ethylene glycol)-block-poly(D,L)-lactic-co-glycolic acid (PEG-PLGA)
nanoparticles for the treatment of schizophrenia. Eur J Pharm Biopharm. 2014;87(1):30-39.
269. Li J, Wu H, Hong J, et al. Odorranalectin is a small peptide lectin with potential for drug
delivery and targeting. PLoS One. 2008;3(6):1-10.
270. Liu Q, Shao X, Chen J, et al. In vivo toxicity and immunogenicity of wheat germ agglutinin
conjugated poly(ethylene glycol)-poly(lactic acid) nanoparticles for intranasal delivery to the
brain. Toxicol Appl Pharmacol. 2011;251(1):79-84.
271. Gao X, Chen J, Tao W, et al. UEA I-bearing nanoparticles for brain delivery following
intranasal administration. Int J Pharm. 2007;340(1-2):207-215.
272. Trabulo S, Cardoso AL, Mano M, de Lima MCP. Cell-penetrating peptides-mechanisms of
cellular uptake and generation of delivery systems. Pharmaceuticals. 2010;3(4):961-993.
273. Kanazawa T, Taki H, Tanaka K, Takashima Y, Okada H. Cell-penetrating peptide-modified
block copolymer micelles promote direct brain delivery via intranasal administration. Pharm
Res. 2011;28(9):2130-2139.
274. Kanazawa T, Akiyama F, Kakizaki S, Takashima Y, Seta Y. Delivery of siRNA to the brain
using a combination of nose-to-brain delivery and cell-penetrating peptide-modified nano-
micelles. Biomaterials. 2013;34(36):9220-9226.
275. Taki H, Kanazawa T, Akiyama F, Takashima Y, Okada H. Intranasal delivery of
camptothecin-loaded tat-modified nanomicells for treatment of intracranial brain tumors.
Pharmaceuticals. 2012;5(10):1092-1102.
276. Yan L, Wang H, Jiang Y, et al. Cell-penetrating peptide-modified PLGA nanoparticles for
enhanced nose-to-brain macromolecular delivery. Macromol Res. 2013;21(4):435-441.
277. Kanazawa T, Morisaki K, Suzuki S, Takashima Y. Prolongation of life in rats with malignant

glioma by intranasal siRNA/drug codelivery to the brain with cell-penetrating peptide-
modified micelles. Mol Pharm. 2014;11(5):1471-1478.
278. Torchilin VP. Tat peptide-mediated intracellular delivery of pharmaceutical nanocarriers. Adv
Drug Deliv Rev. 2008;60(4-5):548-558.
279. Kanazawa T, Kaneko M, Niide T, et al. Enhancement of nose-to-brain delivery of
hydrophilic macromolecules with stearate- or polyethylene glycol-modified arginine-rich
peptide. Int J Pharm. 2017;530(1-2):195-200.
280. Xia H, Gao X, Gu G, et al. Low molecular weight protamine-functionalized nanoparticles for
drug delivery to the brain after intranasal administration. Biomaterials. 2011;32(36):9888-
9898.
281. Zhang Q, Jiang X, Jiang W, Lu W, Su L, Shi Z. Preparation of nimodipine-loaded
microemulsion for intranasal delivery and evaluation on the targeting efficiency to the brain.
Int J Pharm. 2004;275(1-2):85-96.
282. Krauland AH, Leitner VM, Grabovac V, Bernkop-Schnürch A. In Vivo Evaluation of a Nasal
Insulin Delivery System Based on Thiolated Chitosan. J Pharm Sci. 2006;95(11):2463-2472.




















