
Nonadrenergic Imidazoline Binding Sites and Amine Oxidase Activities in Fat Cells
19
PDFlib PLOP: PDF Linearization, Optimization, Protection Page inserted by evaluation version www.pdflib.com – [email protected]
-
Upload
independent -
Category
Documents
-
view
3 -
download
0
Transcript of Nonadrenergic Imidazoline Binding Sites and Amine Oxidase Activities in Fat Cells

PDFlib PLOP: PDF Linearization, Optimization, Protection
Page inserted by evaluation versionwww.pdflib.com – [email protected]

Nonadrenergic Imidazoline Binding Sites and Amine Oxidase Activities
in Fat Cells CHRISTIAN CARPENE: LUC MARTI,
ALAN HUDSON,~ AND MAX LAFONTAN Institut National de la Sante‘ et de la Recherche Me‘dicale
(INSERM U 317) Institut Louis Bugnard
Universite‘ Paul SABATIER C. H. U. Rangueil
31054 Toulouse Cedex, France bPsychopharrnacology Unit School of Medical Sciences
University WALK Bristol, BS8 LTD, UK
Numerous organs, tissues, and cell types possess binding sites for imidazoline drugs (for a review see refs. 1-3). These sites have generally been characterized by their binding parameters rather than their biochemical properties or biological activity. At present, sites labeled with high affinity by [3H]para-aminoclonidine4 or other cloni- dine analogs5 are clearly distinct from those labeled by [‘Hlida~oxan.~ The former type, now called the I1-site, is mainly involved in the hypotensive effects of imidazo- lines administered in animals (reviewed in ref. 7 and in this volume). The latter type, called the 12-site, was previously termed the ‘‘nonadrenergic [3H]idazoxan binding site’’ (NAIBS)? the “imidazoline-guanidinium receptive site” (IGRS)? or other acronyms. In early studies, 12-sites were described as pharmacologically8-’0 and
distinct from adrenergic receptors. Recently, imidazoline bindipgs sites were demonstrated to be located essentially in the mitochondrial outer membrane, in the liver,12 but also in the kidney, brain: and adrenal med~1la.l~ The molecular weights (55-70 kD) of the 1,-sites in the kidney,I4 liver,15 and chromaffin cells16 were also reported. However, the previous implications that Iz-sites are involved in the regulation of biological events such as K’ transport,” noradrenaline release: or insulin secretion18 were in apparent disagreement with the mitochondrial location of such sites. Therefore, the search for a biological effect of 1,-site ligands remains open. Recent reports indicate a parallel between 12-sites and another mitochondrial protein, monoamine oxidase (MAO); some inhibitors of MA0 competed in vitro for [3H]idazoxan binding in rat” and rabbit2’ brain membranes. However, the authors
aAddress for correspondence: Dr. Christian Carp&, INSERM U 317, Institut Louis Bugnard, C.H.U. Rangueil, Bit. L3, 31054 Toulouse Ccdex, France.
380

CARPENE et UZ.: IMIDAZOLINE AND AMINE OXIDASE 381
did not argue for a close relation between 12-sites and M A 0 because: (1) the affinity of M A 0 inhibitors (MAOIs) was 10-fold lower for imidazoline binding sites than for MA02'; (2) idazoxan was only a weak competitor of the binding of labeled M A O I S ' ~ ~ ~ ~ ; and (3) in the brain the density of 12-sites was much lower than that of the two MA0 isoenzymes. In accordance with these pharmacological observations, a similar conclusion was drawn from the differences observed in the isoelectric points of MAO-A, MAO-B, and 12-sites purified from kidney.14 Nevertheless, MAO-B and 12-sites were found to be co-regulated in human brain; both exhibited increasing densities with age.21 The increased density of 12-sites was also observed in the brain of depressed suicide victims." Finally, chronic treatment with the MA01 clorgyline downregulates ['Hlidazoxan binding sites in rat brain." When it is noted that MAOIs are largely used for their antidepressive properties, it is also suspected that interactions between 12-sites and MA0 have functional implications. Such interactions between 12-sites and MAO-A or MAO-B could follow from their common location in the outer mitochondria1 membrane, originally described in liver by Tesson and coworkers. l2
All these observations prompted us to determine if the subcellular co-location of MA0 and 12-sites could be extended to a functional relationship and even to a closer homology between the two entities. To obtain such evidence we first tested the capacity of some inhibitors of MA0 to compete for ["Hlidazoxan binding on the imidazoline receptors of white adipose tissue which were previously described in various animal species'0323 including man.24 Secondly, the effects of imidazoline derivatives were compared to those of various MA0 blockers (including MAO-A or MAO-B selective inhibitors) on the M A 0 activity present in adipose tissue. The same approach was used in rat liver, a model well known for its 12-sites'7925 and MA0 activity.26 Finally, we tested if the inhibition of amine oxidases could influence adipocyte responses to lipolytic amines.
The results indicate that among the tested agents, some MA0 inhibitors and imidazoline molecules share displacing capacities of ['Hlidazoxan binding and inhibi- tion of MA0 activity. Some 12-selective imidazoline compounds were evident (Rx 801077, S 15674, and BU 224); they exhibited 100- to 2,000-fold less affinity for az-adrenoceptors than did idazoxan itself. These 12-site ligands inhibited amine oxidation in both liver cells and adipocytes. In addition, ['Hlidazoxan binding was also inhibited in both models by several inhibitors of MA0 which are not imidazoline or guanidinium derivatives: clorgyline, tranylcypromine, and harmaline. Indeed, inhi- bition of MA0 activity represents a novel biological effect of ligands selective for 12-sites and poses the question of the functional interplay or even the homology between Iz-sites and MAO.
CHARACTERIZATION OF NONADRENERGIC IMIDAZOLINE BINDING SITES IN FAT CELLS
['HlIdazoxan total binding on adipocyte membranes can be separated into at least three components, as shown by the competition studies depicted in FIGURE 1. The first component was adrenergic in nature and corresponded to the portion of binding that was displaced by 100 pM of epinephrine. The second portion was recognized by imidazoline derivatives such as UK 14304 and cirazoline but not by epinephrine
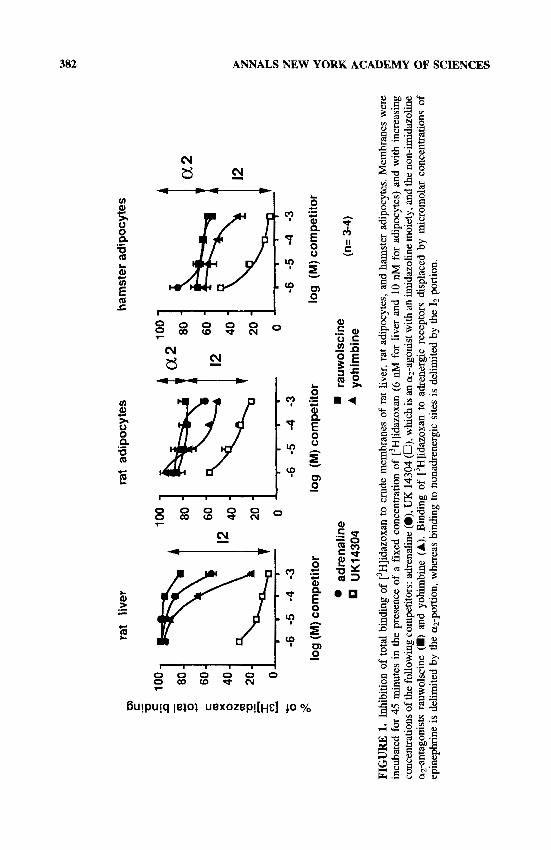
w B
rat
liver
ra
t ad
ipo
cyte
s h
amst
er a
dip
ocy
tes
u)
E
73 .-
..
-
r
-6
-5
-4
-3
-6
-5
-4
-3
-6
-5
-4
-3
0 8
log
(M) c
ompe
titor
lo
g (M
) co
mp
etito
r lo
g (M
) co
mp
etito
r
adre
nal
ine
UK
1430
4 (n
= 3
-4)
rau
wo
lsci
ne
A yo
him
bine
FIGURE 1
. In
hibi
tion
of t
otal
bin
ding
of
[3H
]idaz
oxan
to c
rude
mem
bran
es o
f ra
t liv
er, r
at a
dipo
cyte
s, a
nd h
amst
er a
dipo
cyte
s. M
embr
anes
wer
e in
cuba
ted
for
45 m
inut
es in
the
pre
senc
e of
a f
ixed
con
cent
ratio
n of
[3H
]idaz
oxan
(6
nM f
or li
ver
and
10 n
M f
or a
dipo
cyte
s) a
nd w
ith i
ncre
asin
g co
ncen
tratio
ns of
the
follo
win
g com
petit
ors:
adre
nalin
e (O
), U
K 1
4304
(U), w
hich
is an
cq-
agon
ist w
ith an i
mid
azol
ine m
oiet
y, an
d th
e no
n-im
idaz
olin
e a2
-ant
agon
ists
rauw
olsc
ine
(.) an
d yo
him
bine
(A). B
indi
ng o
f [3
H]id
azox
an to
adr
ener
gic
rece
ptor
s di
spla
ced
by m
icro
mol
ar c
once
ntra
tions
of
epin
ephr
ine i
s de
limite
d by
the
cy2
-por
tion,
whe
reas
bin
ding
to n
onad
rene
rgic
site
s is
del
imite
d by
the
12
porti
on.

CARPENE et a[.: IMIDAZOLINE AND AMINE OXIDASE 383
which, at 1 mM, displaced only a minor part of this binding. The third portion was not competed for by any of the compounds tested. This nonspecific binding, when defined in the presence of 100 pM of cirazoline, averaged 5 and 12% of the total binding at [3H]idazoxan 10 nM in hamster and rat adipocytes, respectively. The same percentages of nonspecific binding were obtained with millimolar concentrations of other imidazoline drugs, such as UK 14304, as illustrated in FIGURE 1. In liver membranes, 5% of ['Hlidazoxan total binding was nonspecific. Moreover, [3H]ida- zoxan mainly labeled nonadrenergic sites in rat liver because 100 pM of epinephrine displaced less than 10% of total binding (FIG. 1). A greater part of the binding was competed for by 1 mM of epinephrine but not by the a2-antagonist rauwolscine. Among the a,,-antagonists tested in competition for ['Hlidazoxan binding, rauwolscine was more selective for a2-adrenergic receptors than for nonadrenergic sites. Another a2-antagonist, yohimbine, recognized part of the nonadrenergic binding sites espe- cially at millimolar concentrations, that is, at doses higher than those needed to block a2-receptors, because the KI value of yohimbine was 3 nM for the a,,-receptors of human a d i p ~ c y t e s . ~ ~ Analysis of the inhibition curves of rauwolscine evidenced a plateau between 1 pM and 1 mM of this agent; the percentage of total Z3H]idazoxan binding resistent to displacement by rauwolscine ranged between 100 and 82% in the liver, 87 and 80% in rat fat cells, and 65 and 55% in hamster fat cells (FIG. 1). This result indicates that the relative proportion of a2-adrenoceptors to imidazoline binding sites was negligible in the rat liver, about one fourth in rat adipocytes, and two thirds in hamster adipocytes. These proportions were confirmed by saturation analyses. For these direct quantifications of nonadrenergic binding sites, increasing concentrations of ['Hlidazoxan were used in the presence of 10 p,M of rauwolscine to preclude binding to a2-adrenoceptors. The a2-adrenoceptors were quantified along with the a2-antagonist [3H]RX 821002. This agent, which is 2-methoxy-idazoxan, did not interfere with the nonadrenergic sites, at least in the nanomolar range, as previously reported in hamster,27 rat,23 and human fat cells.24 In rat fat cells, nonadren- ergic idazoxan binding sites were characterized by an apparent dissociation constant (KD of 15.7 2 2.8 nM and a maximal binding capacity (Bmax) of 290 t 93 fmoV mg protein (n = 5). In hamster adipocyte membranes, the nonadrenergic sites were more numerous: B,,, = 564 2 129 fmoVmg protein (Ko = 9.3 ? 2.2 nM, n = 4). However, fat cell membranes from the hamster also possessed more a2-adrenoceptors than did those from the rat (B,,, of ['H]RX 821002 = 776 2 60 and 39 +- 2 fmoY mg protein, respectively). In rat liver membranes, [3H]idazoxan labeled 450 t 46 fmoYmg protein of nonadrenergic sites (KD = 13.0 * 3.4, n = 14), whereas [3H]RX 821002 labeled only a small amount of binding sites (B,,, = 43 fmoumg protein) of relatively low affinity (KD around 13 nM).
THE FAT CELL: A MODEL FOR DETERMINING THE SELECTIVITY OF LIGANDS FOR NONADRENERGIC
IMIDAZOLINE BINDING SITES
Because white fat cells express both a2-adrenoceptors and nonadrenergic binding sites, it was convenient to verify the distinct pharmacologies of both types of sites on the same cell type. Competition studies of ['Hlidazoxan binding confirmed that

384 ANNALS NEW YORK ACADEMY OF SCIENCES
in rat and hamster fat cells, most of the imidazoline derivatives possessing ( ~ 2 -
adrenergic properties were able to bind to nonadrenergic binding sites (e.g., cirazoline, naphazoline, UK 14304, and idazoxan itself), whereas other imidazolines, such as RX 821002, phentolamine, rilmenidine, and clonidine (not shown), were weak com- petitors. This was in agreement with previous findings in human adipose tissue.24 The low affinity of clonidine allowed nonadrenergic binding sites to be classified as 12-sites according to the recently proposed classification.28q29 The presence of I1-sites on white adipocytes was not tested directly by binding analyses of labeled clonidine derivatives (p-aminoclonidine or p-iodoclonidine) but was excluded from the pattern of competition curves of r3H]clonidine binding. At 100 pM, idazoxan, clonidine, UK 14304, RX 821002, phentolamine, yohimbine, and epinephrine maximally inhibited to the same extent the ['Hlclonidine binding to hamster fat cell membranes; 76 to 88% of the total binding of 8 nM of ligand was inhibited in these conditions. In addition, the maximal binding capacity of ['Hlclonidine was 468 2 76 fmol/mg protein in hamster adipocytes, i.e., slightly less than that reported for [3H]RX 821002. This was in agreement with the respective agonist and antagonist properties of these two radioligands towards a2-adreno~eptors.~~ Accordingly, ['H]clonidine binding to rat adipocytes was at the limits of detection because of the small population of ( ~ 2 -
adrenoceptors in this model (see above and ref. 23). To discriminate new compounds more selective for 12 sites than for a2-adrenocep-
tors, we compared the binding properties of commonly used imidazoline ligands (such as idazoxan or cirazoline) to those of various molecules that can be classified according to their origin: (1) derivatives of idazoxan that were first developed by Reckitt and Colman Laboratories (Hull, England) and now acquired by Pierre Fabre Laboratories (Labige, France), namely, RX 82 1029 or 2-( 1,3-benzodioxanyl)-2-imi- dazoline, and RX 801077 or 2-benzofuran-2-yl-imidazoline; (2) heptyl imidazole derivatives such as S 15 674 (1-methyl 5-n-heptyl imidazole) and S 15430 (2-n- heptylimidazoline), synthesized in the Institut de Recherches Servier (Suresnes, France), in addition to S 15898 (2-(3,3-dicyclopropylpropyl)imidazoline); and (3) BU 224 (2-(4,5-dihydroimidaz-2-yl)-quinoline) and BU 239 (2-(4,5-dihydroimidaz- 2-yl)-quinoxaline) synthesized by Dr. S. Husbands (Bristol, England).
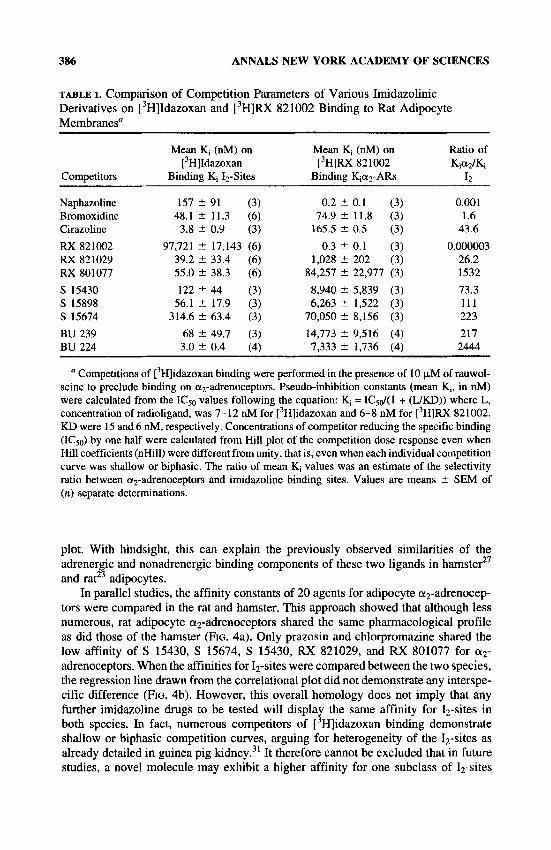
RX 821002 up to 10 p,M did not bind to 12-sites of rat or hamster adipocytes and was the least 12-selective of the tested compounds, justifying its use as an a2- adrenergic ligand (FIG. 2). When also tested for their selectivity, neither RX 821029 nor RX 801077 exhibited a significantly higher affinity than did idazoxan itself (RX 781094) for adipocyte 12-sites, whatever the animal model used (FIG. 2). However, they exhibited much lower affinity in competition for [3H]RX 821002 binding, particu- larly RX 801077, which demonstrated the lowest affinity for a2-adrenoceptors (TABLE 1). The most 12-site-selective agent of the tested compounds was BU 224; it displayed the highest affinity for 12-sites but had an affinity comparable to that of S 15430 or S 15898 with respect to a2-adrenoceptors and was thus endowed with the greatest ratio of KI a2/K1 12. The data in TABLE 1 indicate that RX 801077, S 15674, and BU 224 display the highest selectivity for 12-sites. By contrast, cirazoline, naphazoline, and bromoxidine (UK 14304), which exhibited high affinity for both 12-sites and a2- adrenoceptors, cannot be considered further as highly 12-site-selective.
The use of RX 801077, S 15674, and BU 224 as imidazoline ligands, therefore, is recommended because they offer considerably improved selectivity for 12-sites, at

CARPENE et al.: IMIDAZOLJNE AND AMINE OXIDASE
100.
80 I
60 - 40 - 20.
0-
385
100-
80 - 60 -
40 - 20 - 0-
120 - rat adipocytes ,20 - hamster adipocytes
-10-9 -8 -7 -6 -5 -4 -10 -9 -8 -7 -6 -5 -4 log (M) competitor log (M) competitor
0 idazoxan RX801077 0 RX821002 A RX821029
FIGURE 2. Inhibition of [3H]idazoxan binding by idazoxan derivatives. To prevent the binding to a2-adrenoceptors, binding studies were performed with 8- 1 1 nM [3H]idazoxan in the presence of 10 pM of rauwolscine (rat adipocytes, Zefi panel) or 100 pM of epinephrine (hamster adipocytes, right panel). Data are expressed as a percentage of specific [3H]idazoxan bound after subtraction of the binding not displaced by 100 pM cirazoline. Mean ? SEM of four (hamster) or six (rat) experiments.
least when compared with az-adrenoceptors. However, the need for ligands possessing high affinity and selectivity for nonadrenergic imidazoline binding sites remains crucial in order to label only I-sites, because many investigators still need to mask the a-adrenergic receptors with high concentrations of adrenergic agents mixed with radiolabeled idazoxan or clonidine derivatives. The adipocyte could be a convenient and predictive model to test the selectivity of these ligands. In this regard, the relatively high affinities of Rx 801077 and S 15674 described here on adipocytes were also found for rat liver 12-sites.30
DISTINCT PHARMACOLOGIES OF ADIPOCYTE ( ~ 2 - ADRENERGIC RECEPTORS AND IMIDAZOLINE BINDING
SITES
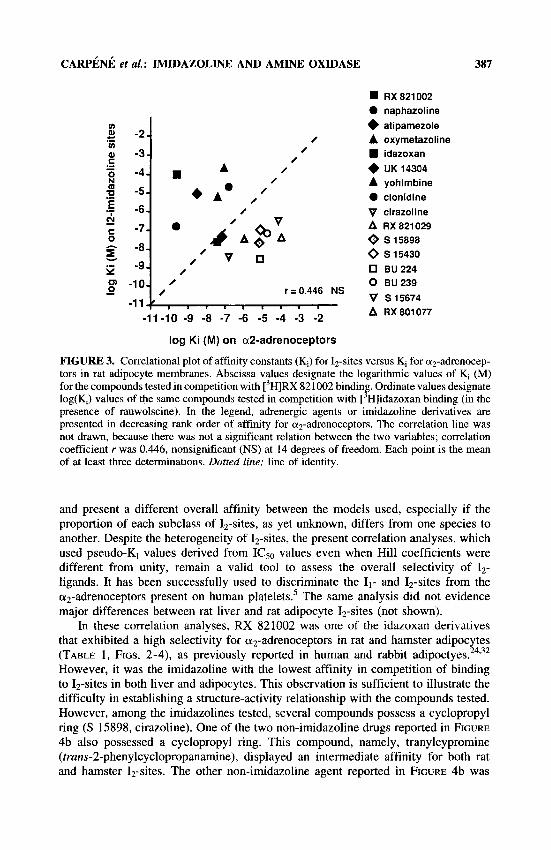
The use of newly synthesized imidazoline derivatives with higher affinity for 12- sites than for az-adrenoceptors could also be used to confirm that these proteins are distinct, as already evidenced by biochemical approaches." As expected, the pharmacological profile of adipocyte 12-sites was not superimposed on that of az- adrenoceptors (FIG. 3). When 16 different drugs including a-agonists, a-antagonists, and imidazoline compounds were analyzed in competition experiments, no correlation was found between their respective affinities for Iz-sites and az-adrenoceptors. Only two compounds, idazoxan and UK 14304, were on the identity line of the correlational
386 ANNALS NEW YORK ACADEMY OF SCIENCES
TABLE 1. Comparison of Competition Parameters of Various Imidazolinic Derivatives on [3H]Idazoxan and r3H]RX 821002 Binding to Rat Adipocyte Membranes"
Competitors
Naphazoline Bromoxidine Cirazoline F S 821002 RX 821029 RX 801077 S 15430 S 15898 S 15674 BU 239 BU 224
Mean Ki (nM) on Mean Ki (nM) on Ratio of [3H]Idazoxan [3H]RX 821002 K&Ki
Binding Ki Iz-Sites Binding Kiaz-ARs 12
157 t 91 (3) 0.2 t 0.1 (3) 0.001 48.1 ? 11.3 (6) 74.9 ? 11.8 (3) I .6 3.8 i 0.9 (3) 165.5 t 0.5 (3) 43.6
97,721 t 17,143 (6) 0.3 t 0.1 (3) 0.000003 39.2 ? 33.4 (6) 1,028 2 202 (3) 26.2 55.0 t 38.3 (6) 84,257 ? 22,977 (3) 1532 122 '' 44 (3) 8,940 2 5,839 (3) 73.3
56.1 ? 17.9 (3) 6,263 2 1,522 (3) 111 314.6 t 63.4 (3) 70,050 t 8,156 (3) 223
68 t 49.7 (3) 14,773 2 9,516 (4) 217 3.0 2 0.4 (4) 7,333 +. 1,736 (4) 2444
Competitions of r3H]idazoxan binding were performed in the presence of 10 p,M of rauwol- scine to preclude binding on uz-adrenoceptors. Pseudo-inhibition constants (mean K,, in nM) were calculated from the ICSo values following the equation: Ki = IC,d( 1 + (LJKD)) where L, concentration of radioligand, was 7-12 nM for [3H]idazoxan and 6-8 nM for r3H]RX 821002. KD were 15 and 6 nM, respectively. Concentrations of competitor reducing the specific binding (ICso) by one half were calculated from Hill plot of the competition dose response even when Hill coefficients (nHill) were different from unity, that is, even when each individual competition curve was shallow or biphasic. The ratio of mean Ki values was an estimate of the selectivity ratio between a,-adrenoceptors and imidazoline binding sites. Values are means ? SEM of (n) separate determinations.
plot. With hindsight, this can explain the previously observed similarities of the adrenergic and nonadrenergic binding components of these two ligands in hamste?' and rat23 adipocytes.
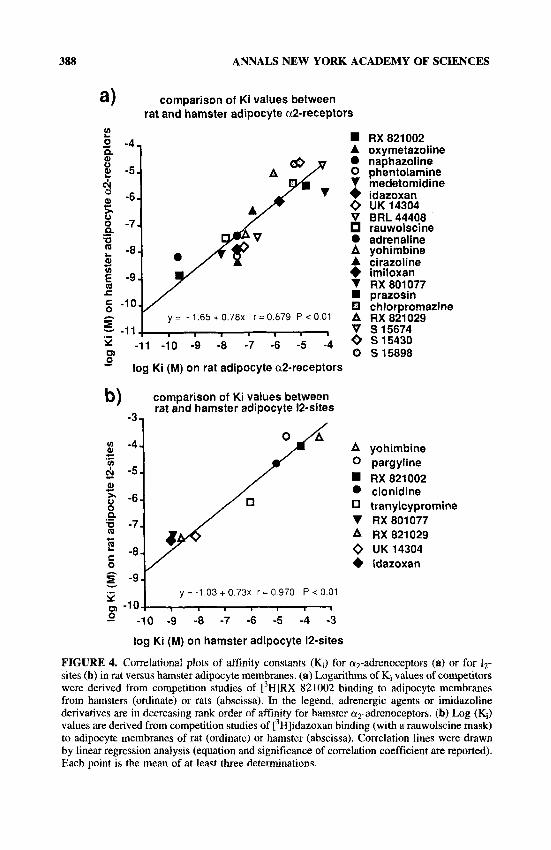
In parallel studies, the affinity constants of 20 agents for adipocyte a2-adrenocep- tors were compared in the rat and hamster. This approach showed that although less numerous, rat adipocyte a2-adrenoceptors shared the same pharmacological profile as did those of the hamster (FIG. 4a). Only prazosin and chlorpromazine shared the low affinity of S 15430, S 15674, S 15430, Rx 821029, and RX 801077 for ( ~ 2 - adrenoceptors. When the affinities for 12-sites were compared between the two species, the regression line drawn from the correlational plot did not demonstrate any interspe- cific difference (FIG. 4b). However, this overall homology does not imply that any further imidazoline drugs to be tested will displa the same affinity for 12-sites in both species. In fact, numerous competitors of [ Hlidazoxan binding demonstrate shallow or biphasic competition curves, arguing for heterogeneity of the 12-sites as already detailed in guinea pig kidney.31 It therefore cannot be excluded that in future studies, a novel molecule may exhibit a higher affinity for one subclass of 12-sites
Y '
CAWENE et ~i.: IMIDAZOLINE AND AMINE OXIDASE
-2.
-3 - -4 - -5 - -6 - -7 - -8 - -9 -
387
-10-
-1 1
0 BU239
v S 15674 rz0.446 NS
/ /
.( , , , , , , , , ,
rn
0
/
/ /
/ /
/ /
/ /
/ /
0
RX 821002 0 naphazoline + atipamezole A oxymetazoline W idazoxan + UK 14304 A yohimbine 0 clonidine V cirazoline A RX 821029 0 S 15898 0 S15430 0 BU224
log Ki (M) on aBadrenoceptors
FIGURE 3. Correlational plot of affinity constants (K,) for Iz-sites versus K, for a2-adrenocep- tors in rat adipocyte membranes. Abscissa values designate the logarithmic values of K, (M) for the compounds tested in competition with [3H]RX 821002 bindin . Ordinate values designate log(K,) values of the same compounds tested in competition with [ Hlidazoxan binding (in the presence of rauwolscine). In the legend, adrenergic agents or imidazoline derivatives are presented in decreasing rank order of affinity for a2-adrenoceptors. The correlation line was not drawn, because there was not a significant relation between the two variables; correlation coefficient r was 0.446, nonsignificant (NS) at 14 degrees of freedom. Each point is the mean of at least three determinations. Dotted line: line of identity.
8 ’
and present a different overall affinity between the models used, especially if the proportion of each subclass of 12-sites, as yet unknown, differs from one species to another. Despite the heterogeneity of 12-sites, the present correlation analyses, which used pseudo-K, values derived from ICs0 values even when Hill coefficients were different from unity, remain a valid tool to assess the overall selectivity of 12- ligands. It has been successfully used to discriminate the 11- and 12-sites from the a2-adrenoceptors present on human platelets5 The same analysis did not evidence major differences between rat liver and rat adipocyte Iz-sites (not shown).
In these correlation analyses, RX 821002 was one of the idazoxan derivatives that exhibited a high selectivity for or2-adrenoceptors in rat and hamster adipocytes (TABLE 1, FIGS. 2-4), as previously reported in human and rabbit a d i p o c t y e ~ . ~ ~ ~ ~ ~ However, it was the imidazoline with the lowest affinity in competition of binding to 12-sites in both liver and adipocytes. This observation is sufficient to illustrate the difficulty in establishing a structure-activity relationship with the compounds tested. However, among the imidazolines tested, several compounds possess a cyclopropyl ring (S 15898, cirazoline). One of the two non-imidazoline drugs reported in FIGURE 4b also possessed a cyclopropyl ring. This compound, namely, tranylcypromine (trans-2-phenylcyclopropanamine), displayed an intermediate affinity for both rat and hamster Iz-sites. The other non-imidazoline agent reported in FIGURE 4b was
388 ANNALS NEW YORK ACADEMY OF SCIENCES
comparison of Ki values between rat and hamster adipocyte a2-receptors
a)
RX 821002 e A oxymetazoline s -4- n 0 naphazoline 0 phentolamine 2 -5. V medetomidine 9 + idazoxan 0 UK14304 a -6- V BRL44408
rauwolscine 0 adrenaline A yohimbine A cirazoline + imitoxan
RX 801077 E, -9. W prarosin
0 = -10. R chlorpromazine
5 -11 V S 15674 Y .;I -;o .; .; .; -6 .j -> 0 ~ 1 5 4 3 0 m 0 S 15898 -
b)
P) 0
e, )I 0 0 -7- P .-
-8. 3 UJ
2
h y = - 1 65 + 0 7 8 ~ r = 0 879 P c 0 01 A RX 821029 .- 0
log Ki (M) on rat adipocyte a2-receptors
comparison of Ki values between rat and hamster adipocyte 12-sites -
A 0
0 0 v A 0 4
yohimbine parg yline RX 821002 clonidine tranylcypromine RX 801077 RX 821029 UK 14304 idazoxan
0 - -10 -9 -8 -7 -6 -5 -4 -3
log Ki (M) on hamster adipocyte 12-sites
FIGURE 4. Correlational plots of affinity constants (Ki) for ap-adrenoceptors (a) or for 1,- sites (b) in rat versus hamster adipocyte membranes. (a) Logarithms of Ki values of competitors were derived from competition studies of [3H]RX 821 002 binding to adipocyte membranes from hamsters (ordinate) or rats (abscissa). In the legend, adrenergic agents or imidazoline derivatives are in decreasing rank order of affinity for hamster a2-adrenoceptors. (b) Log (K;) values are derived from competition studies of ['Hlidazoxan binding (with a rauwolscine mask) to adipocyte membranes of rat (ordinate) or hamster (abscissa). Correlation lines were drawn by linear regression analysis (equation and significance of correlation coefficient are reported). Each point is the mean of at least three determinations.
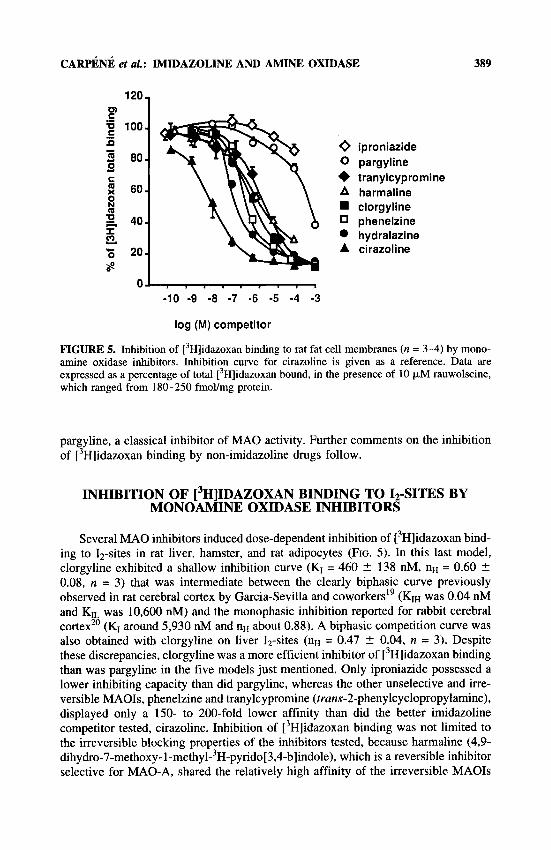
CARPENE et at.: IMIDAZOLINE AND AMINE OXIDASE 389
0 0 + A
0 0 A
iproniaride pargyline tranylcypromine harmaline clorgyline phenelzine hydralazine cirazoline
0- -10 -9 -8 -7 -6 -5 -4 -3
log (M) competitor
FIGURE 5. Inhibition of [3H]idazoxan binding to rat fat cell membranes (n = 3-4) by mono- amine oxidase inhibitors. Inhibition curve for cirazoline is given as a reference. Data are expressed as a percentage of total [3H]idazoxan bound, in the presence of 10 p.M rauwolscine, which ranged from 180-250 fmol/mg protein.
pargyline, a classical inhibitor of MA0 activity. Further comments on the inhibition of [3H]idazoxan binding by non-imidazoline drugs follow.
INHIBITION OF [3H]IDAZOXAN BINDING TO 12-SITES BY MONOAMINE OXIDASE INHIBITORS
Several MA0 inhibitors induced dose-dependent inhibition of [’Hlidazoxan bind- ing to 12-sites in rat liver, hamster, and rat adipocytes (FIG. 5). In this last model, clorgyline exhibited a shallow inhibition curve (KI = 460 2 138 nM, nH = 0.60 2 0.08, n = 3) that was intermediate between the clearly biphasic curve previously observed in rat cerebral cortex by Garcia-Sevilla and coworker^'^ (KIH was 0.04 nM and Kn was 10,600 nM) and the monophasic inhibition reported for rabbit cerebral cortex” (KI around 5,930 nM and nH about 0.88). A biphasic competition curve was also obtained with clorgyline on liver 12-sites (nH = 0.47 2 0.04, n = 3). Despite these discrepancies, clorgyline was a more efficient inhibitor of [ 3H]idazoxan binding than was pargyline in the five models just mentioned. Only iproniazide possessed a lower inhibiting capacity than did pargyline, whereas the other unselective and irre- versible MAOIs, phenelzine and tranylcypromine (truns-2-phenylcyclopropylamine), displayed only a 150- to 200-fold lower affinity than did the better imidazoline competitor tested, cirazoline. Inhibition of [3H]idazoxan binding was not limited to the irreversible blocking properties of the inhibitors tested, because harmaline (4,9- dihydro-7-methoxy- 1 -methyl-3H-pyrido[3,4-b]indole), which is a reversible inhibitor selective for MAO-A, shared the relatively high affinity of the irreversible MAOIs

390 ANNALS NEW YORK ACADEMY OF SCIENCES
(FIG. 5). It must be noted that the most efficient MA01 was hydralazine (KI = 116 ? 46 nM, n = 3) which also possesses antihypertensive properties.
Although these MAOIs do not possess an imidazoline ring in their chemical structure and also are devoid of any imidazole moiety or guanidium group, some of them exhibited a relatively high affinity for ['H]idazoxan binding sites and complete maximal inhibitory capacity comparable to that of some imidazoline derivatives.
Moreover, the MA0 inhibitors competed for [3H]RX 821002 binding on ( ~ 2 - adrenoceptors of rat adipocytes with a noticeably lower affinity than that observed for 12-sites (mean KI > 1 mM for hydralazine). The same observation was made in hamster fat cells.3o To our knowledge, MAOIs are the first molecules that do not belong to the imidazole or guanidium family of derivatives but exhibit affinity for the imidazoline-preferring sites. Because these competitors of ['Hlidazoxan are known to block the MA033 and because MA0 activity and 12-sites are colocalized in the mitochondria1 fraction of various tissues; we compared the effect of some imidazoline derivatives with that of MAOIs on amine oxidation in liver and fat cells.
INHIBITION OF MONOAMINE OXIDASE ACTIVITY BY MAOIs AND 12-SITE LIGANDS
In white adipose tissue, as in liver, amine oxidase activity was measured with crude membrane preparations according to the method of Yu26 using [14C]tyramine as substrate (which is oxidized by both MAO-A and MAO-B). Under the conditions previously described,ln the mean K,,, values for tyramine were 0.33, 0.35, and 0.39 mM for rat adipocytes, hamster adipocytes, and liver cells, respectively. In the three preparations studied, basal oxidation of tyramine was totally abolished in the presence of tranylcypromine (irreversible inhibitor of MAO-AB) (FIG. 6). Pargyline (irrevers- ible inhibitor of MAO-B) and harmaline (reversible inhibitor of MAO-A) were less potent inhibitors of enzyme activity. Their respective dose-dependent inhibition curves were biphasic or shallow, as expected for the presence of both MAO-A and MAO- B acting on the nonspecific substrate ['4C]tyramine.3' In hamster and rat adipocytes, the isoform of MA0 involved in tryamine oxidation remains to be characterized, but the biphasic inhibition curve for pargyline suggests that two MA0 subtypes can coexist in adipose tissue. Furthermore, at least one third of tyramine oxidation by fat cell membranes was resistant to blockade by pargyline or harmaline and was due to an amine oxidase distinct from MAO-A or MAO-B. This enzyme belongs to the semicarbazide-sensitive amine oxidase family (SSAO), as previously described in browd4 and white3' adipocytes. It was not evident in liver cells. Strikingly, in all tissue preparations, various imidazolinic drugs were able to inhibit MA0 activity. Among the drugs tested, the classical imidazoline ligand cirazoline was the most potent (mean ICSo values ranged between 20 and 180 pM), whereas RX 821029 and naphazoline exhibited lower affinity and intrinsic activity (total tyramine oxidation was maximally inhibited by 30-40%, 50-60%, and 60-70% in the presence of 1 mM of these compounds in rat fat cells, hamster fat cells, and liver cells, respectively) (FIG. 6). In addition, S15 674 and RX 801077 totally abolished tyramine oxidation by liver cells in a noncompetitive manner,3o probably by acting on a site distinct from the catalytic one. In addition, the inhibitory action of BU 224 and BU 239 on
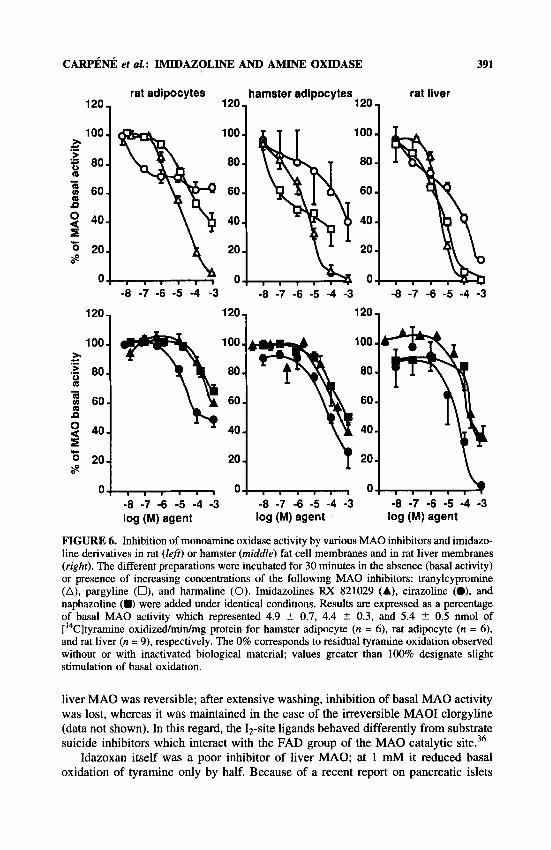
CARPENI~ et UZ.: IMIDAZOLINE AND AMINE OXIDASE
rat adipocytes
1201 100.
'$ 80.
60.
2 40. I
.- 3 > 0 - 4
1201
100.
2 80.
60. n 2 40.
%
0 m
c. .-
- 0
391
-8 -7 -6 -5 -4 -3 -8 -7 -6 -5 -4 -3 -8 -7 -6 -5 -4 -3 120, 120,
-8 -7 -6 -5 -4 -3 log (M) agent
FIGURE 6. Inhibition of monoamine oxidase activity by various M A 0 inhibitors and imidazo- line derivatives in rat (left) or hamster (middle) fat cell membranes and in rat liver membranes (right). The different preparations were incubated for 30 minutes in the absence (basal activity) or presence of increasing concentrations of the following M A 0 inhibitors: tranylcypromine (A), pargyline (O), and harmaline (0). Imidazolines RX 821029 (A), cirazoline (O), and naphazoline (D) were added under identical conditions. Results are expressed as a percentage of basal MA0 activity which represented 4.9 -t 0.7, 4.4 t 0.3, and 5.4 t 0.5 nmol of ['4C]tyramine oxidizedmidmg protein for hamster adipocyte (n = 6), rat adipocyte (n = 6) , and rat liver (n = 9), respectively. The 0% corresponds to residual tyramine oxidation observed without or with inactivated biological material; values greater than 100% designate slight stimulation of basal oxidation.
liver MA0 was reversible; after extensive washing, inhibition of basal MA0 activity was lost, whereas it was maintained in the case of the irreversible MA01 clorgyline (data not shown). In this regard, the Iz-site ligands behaved differently from substrate suicide inhibitors which interact with the FAD group of the M A 0 catalytic site.36
Idazoxan itself was a poor inhibitor of liver MAO; at 1 mM it reduced basal oxidation of tyramine only by half. Because of a recent report on pancreatic islets

392 ANNALS NEW YORK ACADEMY OF SCIENCES
in which agonist and antagonist properties were attributed to several imidazolines according to their effect on insulin secretion,37 we verified if idazoxan could behave as an antagonist of the inhibitory effect of imidazolines on amine oxidase activity. However, complementary studies failed to demonstrate that idazoxan impaired the complete inhibitory action of other imidazolines.30 Therefore, idazoxan cannot be considered a blocker of the imidazoline-induced inhibition of MAO, but rather it is a partial agonist. Clonidine (100 pM) was unable to inhibit more than 20% of MA0 activity, a finding that, in addition to the low affinity of RX 821029 for 1x2- adrenoceptors, implies that the effects observed were independent of a2-adrenergic activation.
When the compounds just mentioned were tested on amine oxidase preparations purified from bovine plasma, tranylcypromine and RX 821029 shared similar 1C50 values (23 and 17 pM) and maximal inhibitory capacity (less than 8% of the basal activity remained in the presence of 1 mM of inhibitor). Naphazoline and cirazoline blocked a large part of the enzyme activity at submillimolar doses. Surprisingly, pargyline and harmaline were ineffective on these preparations.3o
Further studies are required for complete definition of this novel biological effect induced by imidazolines, inhibition of amine oxidase. From the present preliminary studies, it can be concluded that 12-site ligands can inhibit MAO-A and/or MAO-B, because some of them totally counteracted the oxidation of a nonselective substrate, tyramine, in rat liver which is commonly known to possess equal proportions of both isoenzymes.26 Furthermore, kinetic studies demonstrated that the site of action of imidazolines involved in inhibition of oxidase activity seems to be distinct from that of classical “substrate suicide” inhibitors of MAO. Whether the M A 0 enzyme itself, whose two isoforms, A and B, have already been sequenced,” harbors the binding site( s) of imidazolines remains to be elucidated. Such a hypothesis is relevant, because in liver both proteins are located in the mitochondria1 outer membrane,’2*’9 and share a similar molecular weight of around 60 kD.’5*38
INFLUENCE OF THE INHIBITION OF AMINE OXIDATION ON FAT CELL LIPOLYTIC ACTIVITY
Beyond the functional relationship between M A 0 and 12-sites, which is being intensively investigated in the brain,”-2’ it was of interest to define the physiological role of amine oxidation in peripheral organs. Involvement of peripheral MA0 in the degradation of hyperpressive amines was essentially evident during the therapeutic use of some MAOIs as antidepressants. These nonselective inhibitors blocked peripheral MAO-A mainly in the liver and small intestine and prevented the degradation of amines assimilated from foodstuffs. During MA0 blockade, exogenous amines, in- cluding tyramine, were not catabolized, released norepinephrine, and led to excessive, even fatal, hypertension crises, also called the “cheese effect.”36 Whereas liver and intestinal MA0 activities are associated with a protective function against hyperten- sive attacks induced by endogenous or exogenous amines, very little is known about the role of MA0 and SSAO in adipose tissue. Because amines of neuronal or systemic origin are involved in the regulation of lipolysis, we first tested if inhibition of their oxidation could influence their lipolytic capacities.

CARPENE et UI.: IMIDAZOLINE AND AMINE OXIDASE 393
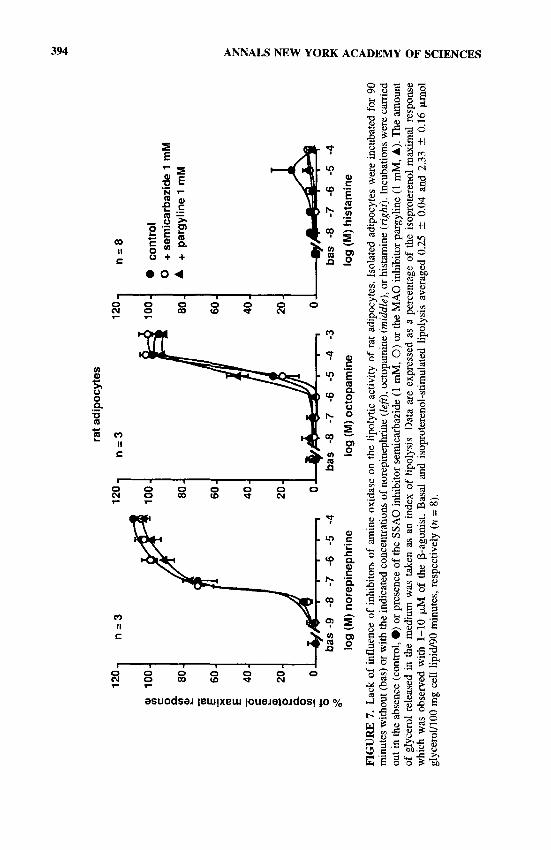
Briefly, blockade of MA0 by 1 mM of pargyline did not influence lipolytic responses to maximal norepinephrine, octopamine (which acts via P3-adrenocep- tors4’), or histamine (which essentially stimulates Hz-receptors41) (FIG. 7). As men- tioned here, another adipocyte enzyme distinct from MA0 is also responsible for the oxidation of amines, SSAO. The SSAO activity was thus blocked by 1 mM of semicarbazide. This failed to modify the lipolytic responses to the three amines studied. This last observation contrasted with the findings of Raimondi et aL4’ who reported an increase in the moderate lipolytic effect of histamine when rat adipocytes were incubated in the presence of 1 mM of ~emicarbazide.~~ Simultaneous blockade of both MA0 and SSAO did not further modify the lipolytic dose-responses reported in FIGURE 7. Catecholamines can also induce antilipolytic responses through activation of a*-adrenoceptors, but no influence of MA0 blockade was evident on a2-adrenergic antilipolysis. Finally, idazoxan itself did not modify basal or stimulated lipolysis. It counteracted the antilipolytic effects of various olz-agonists but had no effect on the antilipolytic responses to adenosine analogs, nicotinic acid, and prostaglandin El (not shown). In this regard, it apparently behaved as a pure az-antagonist, at least when lipolysis was measured, as previously reported on the same cell
Despite the lack of influence of inhibition of amine oxidase activity on the lipolytic response, it is necessary to verify if other aspects of adipocyte metabolism could be influenced by the regulation of amine oxidation. For instance, catecholamines influ- ence not only lipolytic responses but also glucose metabolism and lipogenesis. Another question must therefore be posed: why do fat cells, whose mitochondria1 apparatus is less developed than is that of liver or kidney cells, express a similar number of 12-sites and amine oxidase activity? It can be assumed that adipose cells need to exert a scavenging function by eliminating excess circulating amines in order to preserve their fat stores. This may agree with the finding that orthosympathetic activity is reduced in various models of obesity.
CONCLUSION
Our findings indicate that some imidazoline derivatives and M A 0 inhibitors can inhibit both [?H]idazoxan binding and MA0 activity. Competition of [3H]idazoxan binding by various MA0 inhibitors belonging to the propargylamine, cyclopropylam- ine, or hydrazine families shows that 12-sites not only bind imidazoline or guanidinium derivatives but also exhibit affinity for other families of molecules. Inhibition of MA0 activity represents a novel biological effect of 12-site l ighds which is common to a variety of anatomical locations (adipose tissue and liver). However, the site of action of the imidazolines involved in MA0 inhibition seems distinct from that of “substrate suicide” M A 0 inhibitors. Localization of the 12-sites on the M A 0 itself remains to be demonstrated.
ACKNOWLEDGMENTS
We are grateful to M. Leynaert and P. Collon for their assistance and to A. Cordi (Servier Labs.), D. Nutt (Bristol University), and P. Ladure (P. Fabre Labs.) for providing the imidazoline compounds.
-- ,m
m
._
I
, _,-.
I--”
>”
.--.-..
T t 02 I
l,
au!Jydau!daiou (w) 601 P-
S- 9-
L- 8-
6- Seq
4-
O-
-Y
02
Ot
09
08
00 1
oz 1 E
=U

CARPENE et UZ.: IMIDAZOLINE AND AMINE OXJDASE 395
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
REFERENCES
HIEBLE, J. P. & R. R. RUFFOLO. 1992. Imidazoline receptors: Historical perspective. Fundam. Clin. Pharmacol. 6 (suppl 1): 7-13.
PARINI, A., I. COUPRY, F. TESSON, I. LIMON & C. GARGALIDIS-MOUDANOS. 1992. Imidazo- line-guanidinium receptive site (IGRS): Mitochondria1 distribution and regulation prop- erties. Fundam. Clin. Pharmacol. 6 (suppl 1): 35-40.
REIS, D. J., S. REGUNATHAN, H. WANG, D. L. FEINSTEIN & M. P. MEELEY. 1992. Imidazoline receptors in the nervous system. Fundam. Clin. Pharmacol. 6 (suppl. 1): 23.
ERNSBERGEX, P., M. P. MEELEY, J. J. MA" & D. J. REIS. 1987. Clonidine binds to imidazole binding sites as well as alpha2-adrenoceptors in the ventrolateral medulla. Eur. J. Pharmacol. 134: 1 - 13.
PILETZ, J. E. & K. SLETTEN. 1993. Nonadrenergic imidazoline binding sites on human platelets. J. Pharmacol. Exp. Ther. 267: 1493-1502.
MICHEL, M. C., J. W. REGAN, M. A. GERHARDT, R. R. NEUBIG, P. A. INSEL & H. J. MOTULSKY. 1990. Nonadrenergic [3H]-Idazoxan binding sites are physically distinct from a*-adrenergic receptors. Mol. Pharmacol. 37: 65-68.
BOUSQUET, P., G. BRICCA, M. DONTENWILL, J. FELDMAN, H. GRENEY, A. BELCOURT, J. STUTZMANN & J. TIBRICA. 1992. From the a2-adrenoceptors to the imidazoline preferring receptors. Fundam. Clin. Pharmacol. 6: (suppl. I): 15s-22s.
COWRY, I., R. A. PODEVIN, J. P. DAUSSE & A. PARINI. 1987, Evidence for imidazoline binding sites in basolateral membranes from rabbit kidney. Biochem. Biophys. Res.
HAMILTON, C. A,, J. L. REID & M. A. YAKABU. 1988. [3H]yohimbine and [3H]idazoxan bind to different sites on rabbit forebrain and kidney membranes. Eur. J. Pharmacol.
LANGIN, D. & M. LAFONTAN. 1989. [3H]idazoxan binding at non-alpha2-adrenoceptors in rabbit adipocyte membranes. Eur. J. Pharmacol. 159: 199-203.
PARINI, A., I. COUPRY, R. M. GRAHAM, I. UZIELLI, D. ATLAS & S. LANIER. 1989. Character- ization of an imidazoline/guanidinium receptive site distinct from the az-adrenergic receptor. J. Biol. Chem. 264: I 1874- I 1878.
TESSON, F., C. PRP-BUUS, A. LEMOINE, J. P. PEGORIER & A. PARINI. 1991. Subcellular distribution of imidazoline-guanidinium-receptive sites in human and rabbit liver. Major localization to the mitochondria1 outer membrane. J. Biol. Chem. 266: 155-160.
REGUNATHAN, S., M. P. MEELEY & D. J. REIS. 1993. Expression of non-adrenergic imidazoline sites in chromaffin cells and mitochondrial membranes of bovine adrenal medulla. Biochem. Pharmacol. 45: 1667- 1675.
LIMON, I., I. COUPRY, S. M. LANIER & A. PARINI. 1992. Purification and characterization of mitochondrial imidazoline-guanidinium receptive site from rabbit kidney. J. Biol. Chem. 267: 21645-21649.
LANIER, S. M., B. IVKOVIC, I. SINGH, J. L. NEUMEYER & V. BAKTHAVACHALAM. 1993. Visualization of multiple imidazolinelguanidinium receptive sites. J. Biol. Chem. 268:
WANG, H., S. REGUNATHAN, M. P. MEELEY & D. J. REIS. 1992. Isolation and characteriza- tion of imidazoline receptor protein from bovine adrenal chromaffin cells. Mol. Pharma-
ZONNENSCHEIN, R., S. DIAMANT & D. ATLAS. 1990. Imidazoline receptors in rat liver cells: A novel receptor or a subtype of a*-adrenoceptors? Eur. J. Pharmacol. 190: 203- 215.
CHAN, S. L. F. 1993. Role of a2-adrenoceptors and imidazoline-binding sites in the control of insulin secretion. Clin. Sci. 85: 671-677.
OLMOS, G., A. M. GABILONDO, A. MIRALLES, P. V. ESCRIBA & J. A. GARCIA-SEVILLA. 1993. Chronic treatment with the monoamine oxidase inhibitors clorgyline and pargyline down regulates non-adrenoceptor [3H]idazoxan binding sites in the rat brain. Br. J. Pharmacol. 108: 597-603.
C O ~ . 147: 1055-1060.
146: 345-348.
16047 - 1605 1.
COI. 42: 792-801.

396 ANNALS NEW YORK ACADEMY OF SCIENCES
20.
21.
22.
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
RENOUARD, A,, P. S. WIDDOWSON & A. CORDI. 1993. ['Hlidazoxan binding to rabbit cerebral cortex recognises multiple imidazoline (12-type receptors). Pharmacological characterization and relationship to monoamine oxidase. Br. J. Pharmacol. 109: 625- 631.
SASTRE, M. & J. A. GARCIA-SEVILLA. 1993. Opposite age-dependent changes of alpha2A- adrenoceptors and nonadrenoceptor ['€B]idazoxan binding sites (12-imidazoline sites) in the human brain. Strong correlation of 12 with monoamine oxidase-B sites. J. Neurochem. 61: 881 -889.
MEANA, J. J., F. BARTIJREN, I. MARTIN & J. A. GARCIA-SEVILLA. 1993. Evidence of increased non-adrenoceptor ['HI-idazoxan binding sites in the frontal cortex of de- pressed suicide victims. Biol. Psychiatry 34: 498-501.
CARPENE, C., J. GALITZKY, D. LARROUY, D. LANGW & M. LAFONTAN. 1990. Nonadrenergic sites for imidazolines are not directly involved in the az-adrenergic antilipolytic effect of UK14304 in rat adipocytes. Biochem. Pharmacol. 4 0 437-445.
LANGIN, D., H. PARIS & M. LAFONTAN. 1990. Binding of ['Hlidazoxan and of its methoxy derivative ['HIRX 821002 in human fat cells: ['Hlidazoxan hut not [3H]RX 821002 labels additional non-a2-adrenergic binding sites. Mol. Pharmacol. 37:876- 885.
ATLAS, D. 1991. Clonidine-displacing substance (CDS) and its putative imidazoline recep- tor. New leads for further divergence of alpha2-adrenergic receptor activity. Biochem. Pharmacol. 41: 1541-1549.
Yu, P. H. 1986. Monoamine oxidase. In Neuromethods, Neurotransmitter Enzymes. A. A. Boulton & G. 8. Baker, Eds. Vol. 5: 235-272. Humana Press. Clifton, N.J.
radioligands for the identification of hamster adipocytes alpha2-adrenoceptors. Eur. J. Pharmacol. 171: 145-157.
ERNSBERGER, P. 1992. Heterogcncity of imidazolinc binding sites: Proposed I1 and 12 subtypes. Fund. Clin. Pharmacol. 6: (suppl. 1): 55s.
MICHEL, M. C. & P. ERNSBERGER. 1992. Keeping an eye on the I site: Imidazoline preferring receptors. Trends Pharmacol. Sci. 13: 369-370.
C A R I J ~ N ~ , C., P. COLLON, A. REMAIJRY, A. CORDI, A. HIIDSON, D. NIJTT & M. LAFONTAN. 1995. Inhihition of amine oxidase activity by derivatives that recognize imidazoline Iz sites. J . Pharmacol. Exp. Ther. 272: 681 ~ ~ 6 8 8 .
WIKBERG, J . E. S., S. UHLEN & V. CHAJLANI. 1992. Evidence that drug binding to non- adrencrgic [3H]-idazoxan binding sites (I-receptors) occurs to interacting or intercon- vertible affinity forms of the receptor. Pharmacol. Toxicol. 70: 208-219.
LANGIN, D., H. PARIS, M. DAIJZATS & M. LAPONTAN. 1990. Discrimination between a2- adrenoceptors and ['Hlidazoxan non-adrenergic sites in rabbit fat cells. Eur. J. Pharma-
YOUDIM, M. B. H. & J. P. M. FINBERG. 1991. New directions in monamine oxidase A and B selective inhibitors and substrates. Biochem. Pharmacol. 41: 155- 162.
BARRAND, M. A. & B. A. CALLINGHAM. 1984. Solubilization and some properties of a semi carhazide-sensitive amine oxidase in brown adipose tissue of the rat. Biochem.
RAIMONDI, L., R. PIRISINO, G. IGNESTI, S. CAPECCHI, G. BANCHELLI & F. BUWONI. 1991. Semicarbazide-sensitive amine oxidase activity (SSAO) of rat epididymal white adipose tissue. Biochem. Pharmacol. 41: 467-469.
KYBUR~, E. 1990. New developments in the field of M A 0 inhibitors. Drugs News Perspectives 3: 592-599.
Antagonism of the stimulatory effects of efaroxan and glibenclamide in rat pancreatic islets by the imidazoline, RX 801080. Br. J. Pharmacol. 110: 1017-1022.
BACH, A. W. J., N. C. LAN, D. L. JOHNSON, C. W. ABELL, M. E. BEMBWEK, S. W. KWAN, P. H. SEEBURG & J. C. SHIII. 1988. cDNA cloning of human liver monoamine oxidase
SAULNIER-BLACCIB, E., C. CARPBNE: D. LANGIN & M. LAFONTAN. 1989. Imidazolinic
COI. 188: 261 -272.
J. 222: 467-475.
BROWN, C. A,, S. L. CHAN, M. R. SI'ILLINGS, S. A. SMITH & N. G. MORGAN. 1993.

CAWENE et UZ.: IMIDAZOLINE AND AMINE OXIDASE 397
A and B. Molecular basis of differences in enzymatic properties. Roc. Natl. Acad. Sci. USA 85: 4934-4938.
39. SCHNAITMAN, C., G. ERWIN& J. W. GREENAWALT. 1967. The submitochondriallocalization of monoamine oxidase. J. Cell Biol. 32: 719-735.
40. GALITZKY, J., C. CARPBNI?, M. LAFONTAN & M. BERLAN. 1993. Stimulation spkifique des rkcepteurs P3-adrhergiques du tissu adipeux par l’octopamine. C. R. Acad. Sci. (Paris) 316: 519-523.
41. RAIMONDI, L., G. CONFORTI, G. BANCHELLI, G. IGNESTI, R. PWSINO & F. BUFFONI. 1993. Histamine lipolytic activity and semicarbazide-sensitive amine oxidase (SSAO) of rat white adipose tissue (WAT). Biochem. Pharmacol. 46: 1369- 1376.


















![3H]Cirazoline as a Tool for the Characterization of Imidazoline Sites](https://static.fdokumen.com/doc/165x107/631ef25b7509c0131f0958a9/3hcirazoline-as-a-tool-for-the-characterization-of-imidazoline-sites.jpg)

