
NMR studies of zinc binding in a multi-histidinic peptide fragment
13
PAPER www.rsc.org/dalton | Dalton Transactions NMR studies of zinc binding in a multi-histidinic peptide fragment† Maria Antonietta Zoroddu,* a Serenella Medici, a Massimiliano Peana a and Roberto Anedda b Received 16th July 2009, Accepted 23rd October 2009 First published as an Advance Article on the web 26th November 2009 DOI: 10.1039/b914296g A multi-histidinic peptide and its minimal models have been investigated for Zn(II) binding. We have used NMR spectroscopy to probe the binding of zinc to the three repeats (T 1 R 2 S 3 R 4 S 5 H 6 T 7 S 8 E 9 G 10 ) 3 and to its mono-histidinic minimal models, the 9- and 10-aminoacid fragment. 1 H- 1 H TOCSY, 1 H- 13 C HSQC, 1 H- 1 H NOESY and 1 H- 1 H ROESY multidimensional NMR techniques were performed to understand the details of metal binding sites and the conformational behaviour of the peptides at different pH values and at different ligand to metal molar ratios. Zinc coordination involves imidazole N d of His6 and carboxyl g -O of Glu9 residues; interaction with peptide oxygens of the His6-Thr7 or Thr7-Ser8 bonds in a tetrahedral arrangement with the minimal model peptides, cannot be excluded. Zinc coordination involves, at physiologic pH, all the three imidazole N d donors of His6, His16 and His26 as well as carboxyl g -O of Glu residues in a tetra, penta or octahedral arrangement with the three repeats, the 30-aminoacid fragment. Zinc complexation induces important structural changes with the C-terminal portion of the ligand, constraining it to leave its disordered conformation. Our results give rise to a model of the induced structure of the peptides when bound to zinc. At high pH, amide deprotonation does not take place and hydroxo or high molecular weight polymeric species may be formed. Introduction Recently, histidine rich repeat domains in peptides have drawn considerable attention as models for the study of molecular phenomena associated with metal ion binding in proteins involved in fatal neurodegenerative disorders such as Alzheimer’s (AD) and prion diseases or involved in metal detoxification mechanisms. 1-6 Despite an increasing body of evidence linking metals with neurodegenerative diseases, the precise coordination geometry, the affinity and stoichiometry of binding, as well as what triggers multi-histidinic amyloid-b peptide (Ab) to convert from its soluble form to an amyloidogenic form, are yet to be established. 7 Imbalances in transition metal ions such as copper, zinc or iron are assumed to contribute to Ab deposition. 8 Zinc is an important trace element in human biology. It plays an active role in enzymatic reactions or modulates receptor responses to various neurotransmitters. In particular, Zn(II) ions have been shown to trigger Ab aggregation 9-12 and abnormally high levels of zinc have been found within amyloid deposits in AD patients. 13 Zinc ion levels can be as high as about 1 mM in amyloid plaques. However, the role of zinc is subject to debate 14,15 as Zn(II) ion induces the deposition of Ab but, zinc has also been shown to function as an antioxidant able to protect against oxidative stress. 16,17 Recently, direct spectroscopic evidence showed that Zn(II) ions are bound to Ab via the histidine imidazole rings within senile plaque cores. 18 a Department of Chemistry, University of Sassari, via Vienna 2, 07100, Sassari, Italy. E-mail: [email protected] b Porto Conte Ricerche, Porto Conte, Tramariglio, Alghero, Italy † Electronic supplementary information (ESI) available: Fig. S1: 1 H NMR spectra of the 9aa peptide when free and in the presence of Zn(II). See DOI: 10.1039/b914296g Zinc is also the only metal, other than copper, that induces prion protein (PrP) endocytosis. 19,20 Recent studies on human PrP have indicated that zinc can affect copper binding by either interacting directly with PrP or by indirectly changing the copper binding mode. 21 Zinc, like copper, promotes intermolecular interactions. 22 The presence of His-rich domains in our fragment is similar to those found in prion proteins or Ab peptides, therefore the study of the interaction of the 30-aminoacid peptide with zinc(II) ions can contribute to the understanding of the cru- cial role of poly-imidazole centers in the protein coordination processes. We have previously published data on copper and nickel binding to the two and three histidinic, the 20- and 30-aminoacid fragments (20aa and 30aa), respectively. 23-25 In these studies we have shown that both nickel and copper involve coordination, towards N- terminal region, of main chain amides preceding the His imidazole as well as imidazole nitrogen N d forming, at high pH, a square planar/tetragonal species. One, two or three Cu(II), as well as Ni(II) ions bind to His6, His16 and His26, independently of each other in an intra-repeat binding, at high pH. Multi-histidinic coordination to a single metal ion is involved and macrochelates species in an inter-repeat binding, at low pH are formed. The coordination behaviour towards Zn(II) ions of ends-blocked Ac-RSRSHTSEG-Am (9aa), Ac-TRSRSHTSEG-Am (10aa), and the three repeats Ac-TRSRSHTSEG-TRSRSHTSEG- TRSRSHTSEG-Am (30aa) peptides has been here examined by 1 H- 1 H TOCSY, 1 H- 13 C HSQC, 1 H- 1 H NOESY and 1 H- 1 H ROESY multidimensional NMR techniques in order to understand the details of zinc binding sites and the conformational behaviour of the peptides. NMR experiments were performed in aqueous solution in the pH range 5-8 and at different 0.5 : 1, 1 : 1, 2 : 1 and 3 : 1 metal to ligand molar ratios. 1282 | Dalton Trans., 2010, 39, 1282–1294 This journal is © The Royal Society of Chemistry 2010 Downloaded by Universita' degli Studi di Sassari on 28 December 2010 Published on 26 November 2009 on http://pubs.rsc.org | doi:10.1039/B914296G View Online
Transcript of NMR studies of zinc binding in a multi-histidinic peptide fragment

PAPER www.rsc.org/dalton | Dalton Transactions
NMR studies of zinc binding in a multi-histidinic peptide fragment†
Maria Antonietta Zoroddu,*a Serenella Medici,a Massimiliano Peanaa and Roberto Aneddab
Received 16th July 2009, Accepted 23rd October 2009First published as an Advance Article on the web 26th November 2009DOI: 10.1039/b914296g
A multi-histidinic peptide and its minimal models have been investigated for Zn(II) binding. We haveused NMR spectroscopy to probe the binding of zinc to the three repeats (T1R2S3R4S5H6T7S8E9G10)3
and to its mono-histidinic minimal models, the 9- and 10-aminoacid fragment. 1H-1H TOCSY, 1H-13CHSQC, 1H-1H NOESY and 1H-1H ROESY multidimensional NMR techniques were performed tounderstand the details of metal binding sites and the conformational behaviour of the peptides atdifferent pH values and at different ligand to metal molar ratios. Zinc coordination involves imidazoleNd of His6 and carboxyl g-O of Glu9 residues; interaction with peptide oxygens of the His6-Thr7 orThr7-Ser8 bonds in a tetrahedral arrangement with the minimal model peptides, cannot be excluded.Zinc coordination involves, at physiologic pH, all the three imidazole Nd donors of His6, His16 andHis26 as well as carboxyl g-O of Glu residues in a tetra, penta or octahedral arrangement with the threerepeats, the 30-aminoacid fragment. Zinc complexation induces important structural changes with theC-terminal portion of the ligand, constraining it to leave its disordered conformation. Our results giverise to a model of the induced structure of the peptides when bound to zinc. At high pH, amidedeprotonation does not take place and hydroxo or high molecular weight polymeric species may beformed.
Introduction
Recently, histidine rich repeat domains in peptides have drawnconsiderable attention as models for the study of molecularphenomena associated with metal ion binding in proteins involvedin fatal neurodegenerative disorders such as Alzheimer’s (AD) andprion diseases or involved in metal detoxification mechanisms.1-6
Despite an increasing body of evidence linking metals withneurodegenerative diseases, the precise coordination geometry,the affinity and stoichiometry of binding, as well as what triggersmulti-histidinic amyloid-b peptide (Ab) to convert from its solubleform to an amyloidogenic form, are yet to be established.7
Imbalances in transition metal ions such as copper, zinc oriron are assumed to contribute to Ab deposition.8 Zinc is animportant trace element in human biology. It plays an active rolein enzymatic reactions or modulates receptor responses to variousneurotransmitters. In particular, Zn(II) ions have been shown totrigger Ab aggregation9-12 and abnormally high levels of zinc havebeen found within amyloid deposits in AD patients.13 Zinc ionlevels can be as high as about 1 mM in amyloid plaques. However,the role of zinc is subject to debate14,15 as Zn(II) ion induces thedeposition of Ab but, zinc has also been shown to function as anantioxidant able to protect against oxidative stress.16,17 Recently,direct spectroscopic evidence showed that Zn(II) ions are boundto Ab via the histidine imidazole rings within senile plaquecores.18
aDepartment of Chemistry, University of Sassari, via Vienna 2, 07100,Sassari, Italy. E-mail: [email protected] Conte Ricerche, Porto Conte, Tramariglio, Alghero, Italy† Electronic supplementary information (ESI) available: Fig. S1: 1H NMRspectra of the 9aa peptide when free and in the presence of Zn(II). SeeDOI: 10.1039/b914296g
Zinc is also the only metal, other than copper, that inducesprion protein (PrP) endocytosis.19,20 Recent studies on humanPrP have indicated that zinc can affect copper binding byeither interacting directly with PrP or by indirectly changing thecopper binding mode.21 Zinc, like copper, promotes intermolecularinteractions.22
The presence of His-rich domains in our fragment is similarto those found in prion proteins or Ab peptides, thereforethe study of the interaction of the 30-aminoacid peptide withzinc(II) ions can contribute to the understanding of the cru-cial role of poly-imidazole centers in the protein coordinationprocesses.
We have previously published data on copper and nickel bindingto the two and three histidinic, the 20- and 30-aminoacid fragments(20aa and 30aa), respectively.23-25 In these studies we have shownthat both nickel and copper involve coordination, towards N-terminal region, of main chain amides preceding the His imidazoleas well as imidazole nitrogen Nd forming, at high pH, a squareplanar/tetragonal species. One, two or three Cu(II), as well as Ni(II)ions bind to His6, His16 and His26, independently of each other inan intra-repeat binding, at high pH. Multi-histidinic coordinationto a single metal ion is involved and macrochelates species in aninter-repeat binding, at low pH are formed.
The coordination behaviour towards Zn(II) ions of ends-blockedAc-RSRSHTSEG-Am (9aa), Ac-TRSRSHTSEG-Am (10aa),and the three repeats Ac-TRSRSHTSEG-TRSRSHTSEG-TRSRSHTSEG-Am (30aa) peptides has been here examined by1H-1H TOCSY, 1H-13C HSQC, 1H-1H NOESY and 1H-1H ROESYmultidimensional NMR techniques in order to understand thedetails of zinc binding sites and the conformational behaviourof the peptides. NMR experiments were performed in aqueoussolution in the pH range 5-8 and at different 0.5 : 1, 1 : 1, 2 : 1 and3 : 1 metal to ligand molar ratios.
1282 | Dalton Trans., 2010, 39, 1282–1294 This journal is © The Royal Society of Chemistry 2010
Dow
nloa
ded
by U
nive
rsita
' deg
li St
udi d
i Sas
sari
on
28 D
ecem
ber
2010
Publ
ishe
d on
26
Nov
embe
r 20
09 o
n ht
tp://
pubs
.rsc
.org
| do
i:10.
1039
/B91
4296
GView Online

NMR results allowed us to use molecular modeling approachto determine the three dimensional structure of the Zn-boundpeptide.
Experimental section
Peptide synthesis
Peptides were chemically synthesized using solid phase Fmoc(fluoren-9-ylmethoxycarbonyl) chemistry in an Applied Biosys-tems Synthesizer.26 Peptides were N-terminally acetylated andC-terminally amidated in order to mimic this region of Cap43within the full-length protein. The peptides were removed fromthe resin and deprotected before purification by reverse-phaseHPLC. Fractions were collected and analyzed by MALDI-TOFMS. Fractions containing the peptide of the expected molecularweight were then pooled and lyophilized.
NMR spectroscopy
NMR experiments were performed on a Bruker Avance 600spectrometer equipped with inverse quadruple (QXI) resonanceprobe. Samples used for NMR experiments were 5 mM inconcentration and dissolved in 90/10 (v/v) H2O/D2O solutions.All NMR experiments were performed at 298 K in 5 mm NMRtubes.
2D 1H/13C heteronuclear correlation spectra (HSQC) wereacquired using a phase-sensitive sequence using Echo-Antiecho-TPPI gradient selection with a heteronuclear coupling con-stant JXH = 145 Hz and shaped pulses for all 180 de-gree pulses on f2 channel with decoupling during acquisi-tion, sensitivity improvement and gradients in back-inept wereused.27-29
Relaxation delays of 2 s, 90 degree pulses of about 9.5 ms wereapplied for all experiments. Solvent suppression for 1D, ROESYand TOCSY experiments was achieved using excitation sculptingwith gradients.30 ROESY 2D spectra31 were acquired with spin-lock pulses duration in the range 200-400 ms (optimized for eachpeptide). Long ROESY spin-lock times are believed not to causeimportant indirect effects in low molecular weight oligopeptides.32
ROESY spectra were acquired with 48-64 scans and 512-1024increments, depending on each sample. 1H-1H TOCSY wereperformed using mixing times of 60 ms. The combination ofTOCSY, HSQC and ROESY experiments was used to assignthe spectra of both free and Zn(II)-bound peptides at differentpHs. Assignment of the spectra for both free and Zn(II)-boundpeptides at different pHs was carried out through the combinationof TOCSY, HSQC and ROESY experiments. All NMR data wereprocessed with XWINNMR (Bruker Instruments) software andanalyzed by Sparky 3.1133 and MestRe Nova 5.3.0 (MestrelabResearch S.L.) programs.
Table 1 1H and 13C chemical shifts of free 10aa and Zn(II) bound 10aa at 1 : 1 peptide to metal ratio at pH 7.3
Residue Proton 10aa Free 10aa:Zn(II) 1 : 1 Dd Residue Carbon 10aa Free 10aa:Zn(II) 1 : 1 Dd
ppm ppm ppm ppm ppm ppm
T1 Ha 4.200 4.190 -0.010 T1 Ca 59.591 59.584 -0.007T1 Hb 4.091 4.089 -0.002 T1 Cb 67.148 67.117 -0.031T1 Qg 1.134 1.135 0.001 T1 Cg 18.891 18.849 -0.042R2 Ha 4.244 4.228 -0.016 R2 Ca 53.725 53.583 -0.142R2 Hb1 1.758 1.759 0.001 R2 Cb 28.145 28.139 -0.006R2 Hb2 1.666 1.672 0.006R2 Qg 1.572 1.576 0.004 R2 Cg 24.327 24.355 0.028R2 Qd 3.122 3.117 -0.005 R2 Cd 40.654 40.663 0.009S3 Ha 4.382 4.367 -0.015 S3 Ca 55.682 55.683 0.001S3 Qb 3.790 3.782 -0.008 S3 Cb 61.117 61.105 -0.012R4 Ha 1.793 1.794 0.001 R4 Ca 53.55 53.583 0.033R4 Hb1 1.681 1.681 0.000 R4 Cb 28.145 28.13 -0.015R4 Hb2 1.558 1.552 -0.006R4 Qg 3.125 3.117 -0.008 R4 Cg 24.327 24.355 0.028R4 Qd 1.793 1.794 0.001 R4 Cd 40.654 40.663 0.009S5 Ha 4.370 4.358 -0.012 S5 Ca 55.682 55.683 0.001S5 Qb 3.758 3.749 -0.009 S5 Cb 61.137 61.161 0.024H6 Ha 4.600 4.649 0.049 H6 Ca 54.096 n.d. n.dH6 Hb1 3.072 3.109 0.037 H6 Cb 28.306 27.751 -0.555H6 Hb2 3.037 3.027 -0.010H6 Hd2 6.924 6.945 0.021 H6 Cd2 117.251 n.d. disappearedH6 He1 7.671 7.801 0.130 H6 Ce1 135.931 136.627 0.696T7 Ha 4.306 4.256 -0.050 T7 Ca 59.047 59.408 0.361T7 Hb 4.161 4.171 0.010 T7 Cb 67.167 67.043 -0.124T7 Qg 1.058 1.090 0.032 T7 Cg 18.713 18.859 0.146S8 Ha 4.394 4.372 -0.022 S8 Ca 55.883 55.748 -0.135S8 Qb 3.820 3.811 -0.009 S8 Cb 61.079 61.018 -0.061E9 HN 8.357 8.281 -0.076E9 Ha 4.241 4.245 0.004 E9 Ca 53.857 53.701 -0.156E9 Hb1 2.013 2.043 0.030 E9 Cb 27.581 27.382 -0.199E9 Hb2 1.866 1.882 0.016E9 Qg 2.191 2.239 0.048 E9 Cg 33.626 33.094 -0.532G10 Qa 3.821 3.813 -0.008 G10 Ca 42.331 42.33 -0.001
This journal is © The Royal Society of Chemistry 2010 Dalton Trans., 2010, 39, 1282–1294 | 1283
Dow
nloa
ded
by U
nive
rsita
' deg
li St
udi d
i Sas
sari
on
28 D
ecem
ber
2010
Publ
ishe
d on
26
Nov
embe
r 20
09 o
n ht
tp://
pubs
.rsc
.org
| do
i:10.
1039
/B91
4296
GView Online
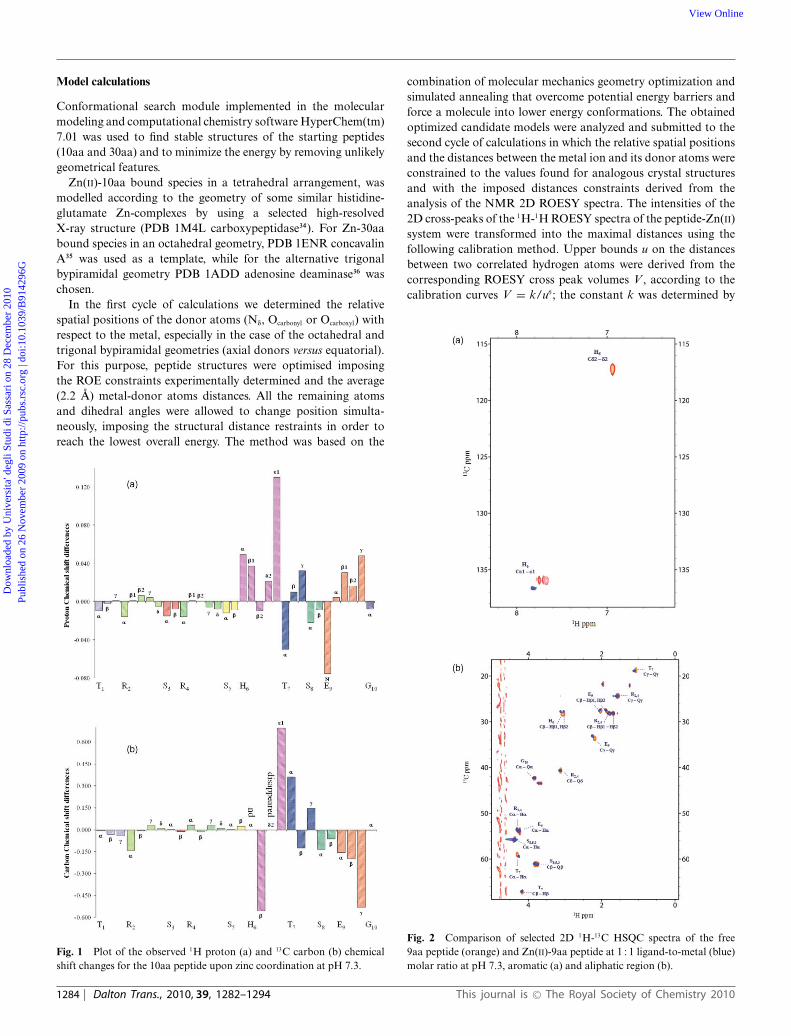
Model calculations
Conformational search module implemented in the molecularmodeling and computational chemistry software HyperChem(tm)7.01 was used to find stable structures of the starting peptides(10aa and 30aa) and to minimize the energy by removing unlikelygeometrical features.
Zn(II)-10aa bound species in a tetrahedral arrangement, wasmodelled according to the geometry of some similar histidine-glutamate Zn-complexes by using a selected high-resolvedX-ray structure (PDB 1M4L carboxypeptidase34). For Zn-30aabound species in an octahedral geometry, PDB 1ENR concavalinA35 was used as a template, while for the alternative trigonalbypiramidal geometry PDB 1ADD adenosine deaminase36 waschosen.
In the first cycle of calculations we determined the relativespatial positions of the donor atoms (Nd, Ocarbonyl or Ocarboxyl) withrespect to the metal, especially in the case of the octahedral andtrigonal bypiramidal geometries (axial donors versus equatorial).For this purpose, peptide structures were optimised imposingthe ROE constraints experimentally determined and the average(2.2 A) metal-donor atoms distances. All the remaining atomsand dihedral angles were allowed to change position simulta-neously, imposing the structural distance restraints in order toreach the lowest overall energy. The method was based on the
Fig. 1 Plot of the observed 1H proton (a) and 13C carbon (b) chemicalshift changes for the 10aa peptide upon zinc coordination at pH 7.3.
combination of molecular mechanics geometry optimization andsimulated annealing that overcome potential energy barriers andforce a molecule into lower energy conformations. The obtainedoptimized candidate models were analyzed and submitted to thesecond cycle of calculations in which the relative spatial positionsand the distances between the metal ion and its donor atoms wereconstrained to the values found for analogous crystal structuresand with the imposed distances constraints derived from theanalysis of the NMR 2D ROESY spectra. The intensities of the2D cross-peaks of the 1H-1H ROESY spectra of the peptide-Zn(II)system were transformed into the maximal distances using thefollowing calibration method. Upper bounds u on the distancesbetween two correlated hydrogen atoms were derived from thecorresponding ROESY cross peak volumes V , according to thecalibration curves V = k/u6; the constant k was determined by
Fig. 2 Comparison of selected 2D 1H-13C HSQC spectra of the free9aa peptide (orange) and Zn(II)-9aa peptide at 1 : 1 ligand-to-metal (blue)molar ratio at pH 7.3, aromatic (a) and aliphatic region (b).
1284 | Dalton Trans., 2010, 39, 1282–1294 This journal is © The Royal Society of Chemistry 2010
Dow
nloa
ded
by U
nive
rsita
' deg
li St
udi d
i Sas
sari
on
28 D
ecem
ber
2010
Publ
ishe
d on
26
Nov
embe
r 20
09 o
n ht
tp://
pubs
.rsc
.org
| do
i:10.
1039
/B91
4296
GView Online
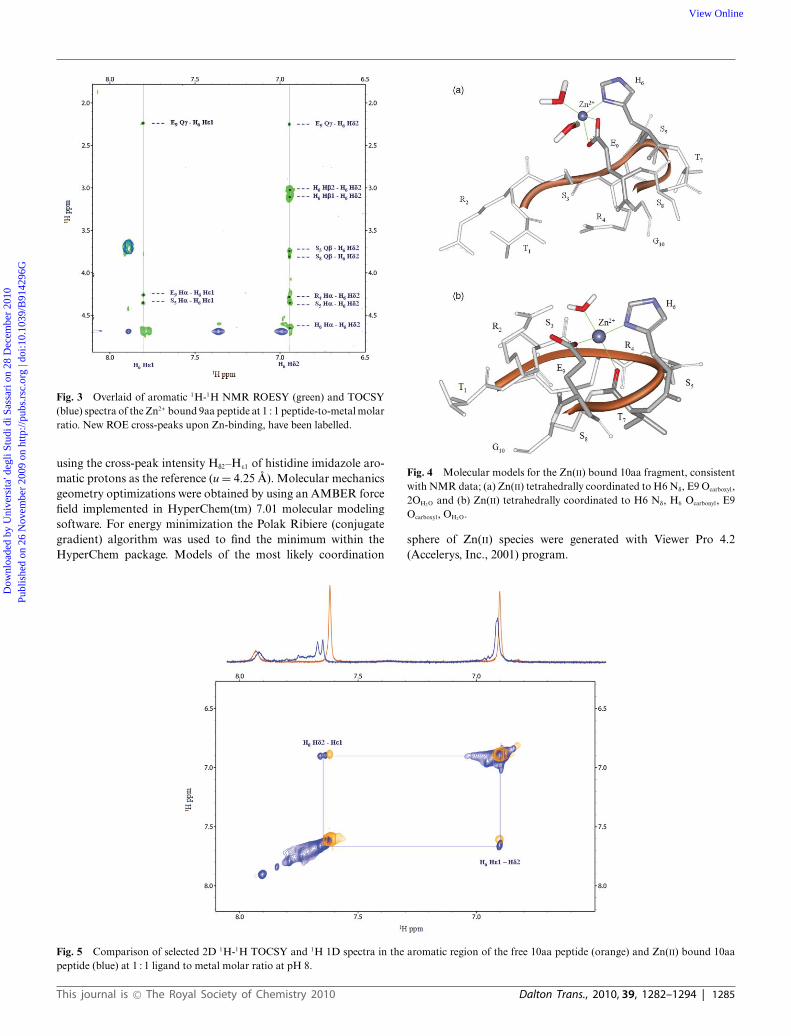
Fig. 3 Overlaid of aromatic 1H-1H NMR ROESY (green) and TOCSY(blue) spectra of the Zn2+ bound 9aa peptide at 1 : 1 peptide-to-metal molarratio. New ROE cross-peaks upon Zn-binding, have been labelled.
using the cross-peak intensity Hd2–He1 of histidine imidazole aro-matic protons as the reference (u = 4.25 A). Molecular mechanicsgeometry optimizations were obtained by using an AMBER forcefield implemented in HyperChem(tm) 7.01 molecular modelingsoftware. For energy minimization the Polak Ribiere (conjugategradient) algorithm was used to find the minimum within theHyperChem package. Models of the most likely coordination
Fig. 4 Molecular models for the Zn(II) bound 10aa fragment, consistentwith NMR data; (a) Zn(II) tetrahedrally coordinated to H6 Nd, E9 Ocarboxyl,2OH2 O and (b) Zn(II) tetrahedrally coordinated to H6 Nd, H6 Ocarbonyl, E9Ocarboxyl, OH2 O.
sphere of Zn(II) species were generated with Viewer Pro 4.2(Accelerys, Inc., 2001) program.
Fig. 5 Comparison of selected 2D 1H-1H TOCSY and 1H 1D spectra in the aromatic region of the free 10aa peptide (orange) and Zn(II) bound 10aapeptide (blue) at 1 : 1 ligand to metal molar ratio at pH 8.
This journal is © The Royal Society of Chemistry 2010 Dalton Trans., 2010, 39, 1282–1294 | 1285
Dow
nloa
ded
by U
nive
rsita
' deg
li St
udi d
i Sas
sari
on
28 D
ecem
ber
2010
Publ
ishe
d on
26
Nov
embe
r 20
09 o
n ht
tp://
pubs
.rsc
.org
| do
i:10.
1039
/B91
4296
GView Online

Results and discussion
The 9aa, the 10aa and the 30aa free peptides have alreadybeen extensively characterized through 1D and 2D NMRspectroscopy.23-25 The combination of mono- and bidimensionalNMR experiments was used to assign the Zn(II)-bound peptidessignals. The NMR studies on the shorter peptides were usefulfor a better understanding of the coordination ability of zinctowards the larger fragment and, in particular, to study thepossible involvement of multiple histidinic sites in the coordinationevents.
In order to characterize zinc binding to the three repeats and toits minimal models, 1H-1H TOCSY, 1H-13C HSQC, 1H-1H NOESYand 1H-1H ROESY multidimensional NMR techniques have beenused.
The behaviour of the peptides toward zinc ions was examinedat different pH values and at different ligand to metal molarratios.
Titration of the ligands in the pH range from 5 to 8, showedthat at pH below 6, when the histidines are mainly protonated, nozinc binding occurs. In fact, complexation starts at pH 6.1 and thecomplexed Zn(II) species are stable up to pH 8.
In Table 1 assignments (at pH 7.3 and at 1 : 1 metal to ligandmolar ratio) and Dd (dbound-dfree) of proton and carbon signals arereported.
Addition of zinc to the peptide caused a number of NMRresonances to be perturbed. A comparison between 0.5 : 1 and1 : 1 Zn(II) to ligand molar ratio spectra at pH 6, shows signalshifts to high or low field, typical for a fast exchange be-tween free and zinc bound peptide, with respect to the NMRtimescale rate (Fig. S1 of electronic supplementary information,ESI†).
In Fig. 1(a) and (b), histograms about all the obtained chemicalshift variations (Dd) for 1H protons and 13C carbons respectively,for the 10aa fragment in the presence of 1 equiv of Zn(II) ion,are reported. From the entity of variations obtained, it is clear
Table 2 1H and 13C chemical shifts of free 30aa peptide and Zn(II) bound at 1 : 1 peptide to metal ratio at pH 6
Residue Proton 30aa Free 30aa:Zn(II) 1 : 1 Dd Residue Carbon 30aa Free 30aa:Zn(II) 1 : 1 Dd
pH6 ppm ppm ppm ppm ppm ppm
T1 HN 8.137 8.140 0.003T1 Ha 4.188 4.194 0.006 T1 Ca 59.607 59.650 0.043T1 Hb 4.096 4.102 0.006 T1 Cb 67.103 67.116 0.013T1 Qg 1.128 1.124 -0.004 T1 Cg 18.916 18.897 -0.019R2 HN 8.280 8.276 -0.004R2 Ha 4.302 4.301 -0.001 R2 Ca 53.530 53.554 0.024R2 Hb1 1.807 1.814 0.007 R2 Cb 28.066 28.029 -0.037R2 Hb2 1.701 1.706 0.005R2 Qg 1.565 1.567 0.002 R2 Cg 24.388 24.404 0.016R2 Qd 3.115 3.117 0.002 R2 Cd 40.665 40.658 -0.007R2 Qe 7.136 7.130 -0.006S3 HN 8.251 8.263 0.012S3 Ha 4.348 4.357 0.009 S3 Ca 55.747 55.779 0.032S3 Qb 3.791 3.785 -0.006 S3 Cb 61.094 61.125 0.031R4 HN 8.330 8.320 -0.010R4 Ha 4.301 4.298 -0.003 R4 Ca 53.751 53.681 -0.070R4 Hb1 1.808 1.813 0.005 R4 Cb 28.066 28.029 -0.037R4 Hb2 1.697 1.705 0.008R4 Qg 1.566 1.567 0.001 R4 Cg 24.388 24.404 0.016R4 Qd 3.116 3.120 0.004 R4 Cd 40.665 40.658 -0.007R4 Qe 7.136 7.132 -0.004S5 HN 8.239 8.205 -0.034S5 Ha 4.344 4.347 0.003 S5 Ca 55.747 55.734 -0.013S5 Qb 3.763 3.762 -0.001 S5 Cb 61.087 61.137 0.050H6 Ha 4.669 4.687 0.018 H6 Ca nd nd ndH6 Hb1 3.167 3.164 -0.003 H6 Cb 27.230 27.154 -0.076H6 Hb2 3.080 3.069 -0.011H6 Hd2 7.082 7.053 -0.029 H6 Cd2 117.358 117.517 0.159H6 He1 8.147 8.046 -0.101 H6 Ce1 134.758 135.869 1.111T7 HN 8.072 8.075 0.003T7 Ha 4.302 4.263 -0.039 T7 Ca 59.213 59.534 0.321T7 Hb 4.163 4.176 0.013 T7 Cb 67.096 67.042 -0.054T7 Qg 1.098 1.119 0.021 T7 Cg 18.825 18.920 0.095S8 HN 8.307 8.288 -0.019S8 Ha 4.401 4.382 -0.019 S8 Ca 55.874 55.885 0.011S8 Qb 3.815 3.812 -0.003 S8 Cb 61.089 61.056 -0.033E9 HN 8.411 8.335 -0.076E9 Ha 4.225 4.239 0.014 E9 Ca 54.334 54.235 -0.099E9 Hb1 2.006 2.030 0.024 E9 Cb 27.561 27.479 -0.082E9 Hb2 1.893 1.904 0.011E9 Qg 2.206 2.241 0.035 E9 Cg 33.562 33.265 -0.297G10 HN 8.351 8.340 -0.011G10 Qa 3.927 3.938 0.011 G10 Ca 42.802 42.822 0.020
1286 | Dalton Trans., 2010, 39, 1282–1294 This journal is © The Royal Society of Chemistry 2010
Dow
nloa
ded
by U
nive
rsita
' deg
li St
udi d
i Sas
sari
on
28 D
ecem
ber
2010
Publ
ishe
d on
26
Nov
embe
r 20
09 o
n ht
tp://
pubs
.rsc
.org
| do
i:10.
1039
/B91
4296
GView Online

that the only protons affected by metal addition are those of theportion from His6 to Glu9 residues, thus indicating the directionof coordination towards C-terminus.
The most affected protons in the aromatic region are those ofimidazole ring of histidine and, in the aliphatic region those ofcarboxyl group of glutamic residue.
The selective spectral changes observed upon Zn additionclearly indicate the involvement of these two residues in metalcoordination.
The large downfield shift of His6 He1 (Dd = 0.130 ppm),compared to the shift of Hd2 (Dd = 0.021 ppm), obtained at pH 7.3,indicates zinc coordination through imidazole Nd. The deshieldingeffect exhibited by He1 protons can be explained by consideringthe inductive effect of imidazolic nitrogens that withdraw electrondensity away from the spinning nuclei.
In addition, the 9aa or the 10aa fragments both display strongZn-induced shifts on Glu9 residue signals in the aliphatic region.In particular, the Hg Glu9 protons are shifted downfield (Dd =0.048 ppm) upon metal binding. This result points to zinc bindingof Glu9 side chain O-carboxylate.
The sharp signals obtained upon zinc addition and the absenceof a decrease in intensity of the shifted signals indicate theformation of a strong zinc binding to both sites.
The almost complete lack of significant broadening or shift ofall the other resonances, together with the observed shifts locatedin the C-terminal portion from His6 to Glu9 residues indicatesthe formation of a macrochelate system, in agreement with thedirection of coordination towards C-terminal.
When peptide binds Zn(II) through imidazole from His6and carboxylic oxygen from Glu9 residues, the chelate ringsize is rather large. Since the stability of a chelate complexgenerally increases as the size of the chelate ring decreases,it is possible that one (or more) of potential ligand atomson the peptide may also stabilize the complex via Zn(II)binding.
Experimental manifestation of this possibility emerged fromthe NMR data, in particular from the strong decrease in intensitytogether with the upfield shift of Thr7 Ha (Dd = -0.050 ppm) andthe strong downfield shift of Thr7 Ca (Dd = 0.361) and, though toa lesser extent, from the upfield shift of Ser8 Ha (Dd = - 0.034 ppm)and Ser8 Ca (Dd = -0.135 ppm). These results indicate an approachof the backbone from Thr7 to Ser8 towards coordinated metalion and favour the view that coordination of the amide carbonyloxygen of the His6-Thr7 and/or Thr7-Ser8 peptide bond could notbe excluded. A possible involvement of additional donor atoms isalso supported by the different Dd found for His Hb1Hb2, Dd Hb1
Table 3 1H and 13C chemical shifts of free 30aa peptide and Zn(II) bound at 1 : 1 peptide to metal ratio at pH 7
Residue Proton 30aa Free 30aa:Zn(II) 1 : 1 Dd Residue Carbon 30aa Free 30aa:Zn(II) 1 : 1 Dd
pH 7 ppm ppm ppm ppm ppm ppm
T1 HN 8.134 8.137 0.003T1 Ha 4.187 4.194 0.007 T1 Ca 59.548 59.638 0.090T1 Hb 4.090 4.099 0.009 T1 Cb 67.158 67.112 -0.046T1 Qg 1.124 1.130 0.006 T1 Cg 18.915 18.989 0.074R2 Ha 4.303 4.297 -0.006 R2 Ca 53.528 53.583 0.055R2 Hb1 1.800 1.806 0.006 R2 Cb 28.111 28.002 -0.109R2 Hb2 1.693 1.707 0.014R2 Qg 1.560 1.562 0.002 R2 Cg 24.370 24.418 0.048R2 Qd 3.108 3.110 0.002 R2 Cd 40.668 40.665 -0.003S3 Ha 4.350 4.362 0.012 S3 Ca 55.745 55.808 0.063S3 Qb 3.787 3.784 -0.003 S3 Cb 61.105 61.105 0.000R4 Ha 4.303 4.294 -0.009 R4 Ca 53.751 53.722 -0.029R4 Hb1 1.800 1.808 0.008 R4 Cb 28.111 28.002 -0.109R4 Hb2 1.693 1.702 0.009R4 Qg 1.560 1.561 0.001 R4 Cg 24.370 24.408 0.038R4 Qd 3.107 3.111 0.004 R4 Cd 40.665 40.665 0.000S5 Ha 4.352 4.350 -0.002 S5 Ca 55.745 55.808 0.063S5 Qb 3.752 3.757 0.005 S5 Cb 61.124 61.161 0.037H6 Ha 4.612 4.657 0.045 H6 Ca n.d. n.d. n.dH6 Hb1 3.075 3.119 0.044 H6 Cb 28.282 27.587 -0.695H6 Hb2 3.022 3.028 0.006H6 Hd2 6.939 6.965 0.026 H6 Cd2 117.249 disappeared disappearedH6 He1 7.765 7.834 0.069 H6 Ce1 135.831 136.917 1.086T7 HN 7.973 8.000 0.027T7 Ha 4.293 4.247 -0.046 T7 Ca 59.055 59.572 0.517T7 Hb 4.153 4.177 0.024 T7 Cb 67.148 67.026 -0.122T7 Qg 1.066 1.108 0.042 T7 Cg 18.740 18.933 0.193S8 Ha 4.395 4.362 -0.033 S8 Ca 55.861 55.808 -0.053S8 Qb 3.814 3.807 -0.007 S8 Cb 61.108 61.018 -0.090E9 HN 8.404 8.273 -0.131E9 Ha 4.219 4.237 0.018 E9 Ca 54.326 54.213 -0.113E9 Hb1 2.004 2.029 0.025 E9 Cb 27.543 27.485 -0.058E9 Hb2 1.888 1.907 0.019E9 Qg 2.195 2.246 0.051 E9 Cg 33.598 33.201 -0.397G10 HN 8.341 8.327 -0.014G10 Qa 3.917 3.937 0.020 G10 Ca 42.814 42.878 0.064
This journal is © The Royal Society of Chemistry 2010 Dalton Trans., 2010, 39, 1282–1294 | 1287
Dow
nloa
ded
by U
nive
rsita
' deg
li St
udi d
i Sas
sari
on
28 D
ecem
ber
2010
Publ
ishe
d on
26
Nov
embe
r 20
09 o
n ht
tp://
pubs
.rsc
.org
| do
i:10.
1039
/B91
4296
GView Online

(0.037 ppm), Dd Hb2 (-0.010 ppm) and the strong shift of His Ha
and Dd Ha (0.049 ppm) upon zinc coordination, as reported inTable 1 and Fig. 1A.
In addition, the effect on Thr7 Hg protons suggests a changeon hydrophobic packing of the Thr7 side-chain upon metalcoordination. Thus, zinc binding via histidine residue results ina structural change that affects the environment of the side chainof Thr7 residue.
These data suggest that complexation results in a relevantrestriction of segmental motion throughout His6-Thr7 bondon the C-terminal portion of the peptide. It could be causedby the Thr7 carbonyl oxygen involvement in the coordinationsphere to give a tetra-coordinated Zn(II) species in a tetrahedralarrangement with the minimal model peptides; water may alsobe involved to complete the coordination sphere around zincion.
Comparison of selected 2D 1H-13C HSQC spectra of the free9aa peptide and Zn(II)-9aa peptide at 1 : 1 ligand-to-metal molarratio at pH 7.3 is reported in Fig. 2. The disappearance in the
aromatic region of Cd2 signal (Fig. 2(a)), which is in an oppositeposition to the site of coordination, could be explained withits approach to a shielding or deshielding area in the boundspecies.
Inter-residual ROESY connectivities among His6 He1, Hd2 andside chain protons of several residues, Glu9 Hg , His6 Ha, Hb, Arg4Ha, Ser5, Ser8 Hb, around the coordination center, have also beendetected (Fig. 3).
All these findings taken together are indicative of the rigidity ofthe peptide portion and of a restriction of the mobility from Arg4to Glu9 residue, upon zinc coordination.
Further metal additions on the 9aa or 10aa fragment yield neg-ligible chemical shift variations indicating the exclusive presenceof a unique 1 : 1 Zn-peptide bound species.
The residues directly involved in the sphere of coordinationshow also large variation in carbon chemical shifts as reported inFig. 1 and Tables 2 and 3.
All the results allowed us to build a model for the Zn tetra-coordinated species.
Fig. 6 (a) Aromatic and (b) aliphatic region of 1H 1D spectra of the free 30aa peptide (orange) and in the presence of 1.0 (blue) equiv of Zn(II) at pH 7.
1288 | Dalton Trans., 2010, 39, 1282–1294 This journal is © The Royal Society of Chemistry 2010
Dow
nloa
ded
by U
nive
rsita
' deg
li St
udi d
i Sas
sari
on
28 D
ecem
ber
2010
Publ
ishe
d on
26
Nov
embe
r 20
09 o
n ht
tp://
pubs
.rsc
.org
| do
i:10.
1039
/B91
4296
GView Online

In Fig. 4 molecular models consistent with NMR data fora tetrahedral arrangement of Zn-{NdOcarboxyl 2OH2} or Zn-{Nd
Ocarboxyl Ocarbonyl OH2} chromophore species, are reported.pH higher than 7.3, resulted in a general broadening of
NMR signals, suggesting changes in zinc coordination. At pH 8,amide deprotonation does not take place and together with thegeneral broadening of all the resonances, shifts and broadeningof imidazolic Hd2 and He1 aromatic signals can be seen (Fig. 5).These changes suggest increase in molecular weight and formationof polymeric species37 or zinc peptide-hydroxo complexes followedby dissociation and possible formation of Zn(OH)4
2- and Zn(OH)2
precipitation which starts to occur above pH 8.The NMR study on shorter peptides was useful for a better
understanding of the coordination behaviour of Zn(II) ionstowards the longer 30-aminoacid fragment and in particular,to study the possible involvement of multiple histidinic sites incoordination events.
The chemical shift changes in each of the three mono-histidinic fragments of the 30aa peptide reflected, in terms of
type and intensity, the values found for the 10aa-Zn(II) systemin a 1 : 1 ligand to metal molar ratio, though some differencescan also be noted. In Fig. 6 an overlaid of aliphatic (a) andaromatic region (b) of 1D spectra for the free 30aa and Zn(II)bound 30aa at 1 : 1 peptide to zinc molar ratio at pH 7, isreported.
In Fig. 7 a comparison of selected 2D 1H-13C HSQC for the free30aa and Zn-30aa peptide at 1 : 1 molar ratio at pH 7, is reported.
It is clear that the longer chain of the 30aa peptide, repre-senting the three decapeptide repeats, displays a unique modeof Zn(II) binding. The most affected protons are those ofaromatic ring of histidines and those of side chains of glutammicresidues.
The three repeats, the 30aa fragment, exhibits in the presence ofzinc, similar effects to those found for the minimal model but alsosome subtle differences.
The pattern of chemical shift variations clearly indicatesZn(II) coordination to the same aminoacid residues and donoratoms as for the minimal models. Like for the 10aa, also
Fig. 7 Comparison of 2D 1H-13C HSQC spectra, aliphatic (a) and (b) and aromatic (c) region of free 30aa peptide (orange) and Zn(II)-30aa peptide at1 : 1 ligand to metal (blue) molar ratio at pH 7.
This journal is © The Royal Society of Chemistry 2010 Dalton Trans., 2010, 39, 1282–1294 | 1289
Dow
nloa
ded
by U
nive
rsita
' deg
li St
udi d
i Sas
sari
on
28 D
ecem
ber
2010
Publ
ishe
d on
26
Nov
embe
r 20
09 o
n ht
tp://
pubs
.rsc
.org
| do
i:10.
1039
/B91
4296
GView Online
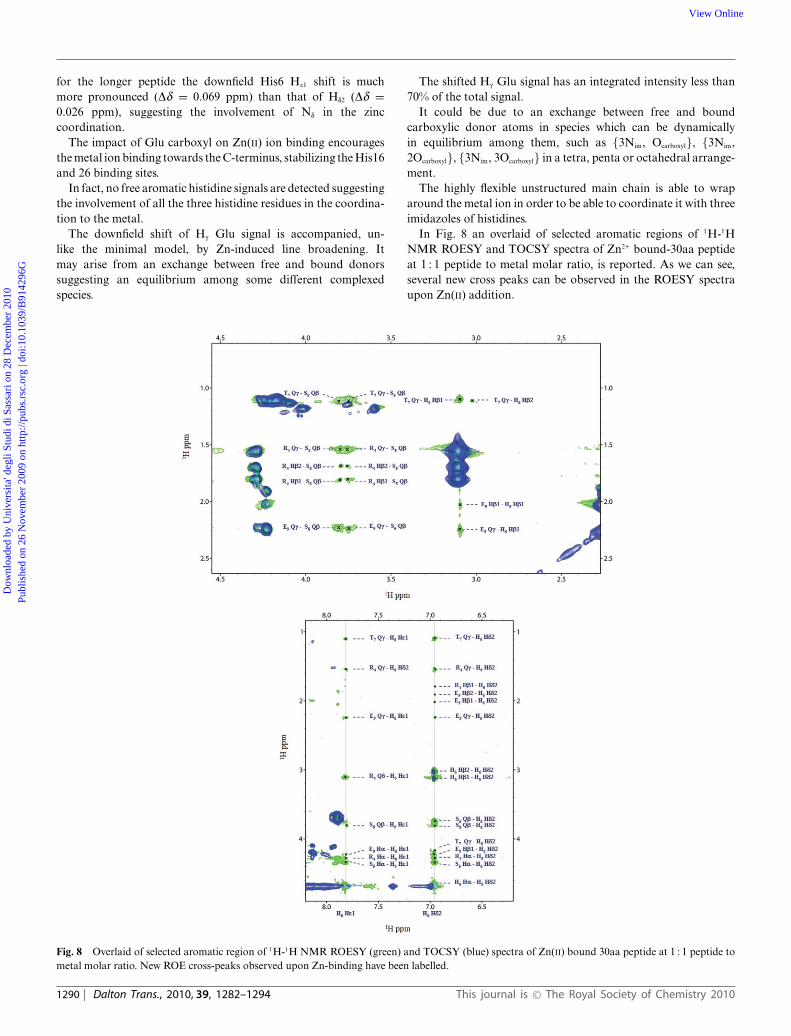
for the longer peptide the downfield His6 He1 shift is muchmore pronounced (Dd = 0.069 ppm) than that of Hd2 (Dd =0.026 ppm), suggesting the involvement of Nd in the zinccoordination.
The impact of Glu carboxyl on Zn(II) ion binding encouragesthe metal ion binding towards the C-terminus, stabilizing the His16and 26 binding sites.
In fact, no free aromatic histidine signals are detected suggestingthe involvement of all the three histidine residues in the coordina-tion to the metal.
The downfield shift of Hg Glu signal is accompanied, un-like the minimal model, by Zn-induced line broadening. Itmay arise from an exchange between free and bound donorssuggesting an equilibrium among some different complexedspecies.
The shifted Hg Glu signal has an integrated intensity less than70% of the total signal.
It could be due to an exchange between free and boundcarboxylic donor atoms in species which can be dynamicallyin equilibrium among them, such as {3Nim, Ocarboxyl}, {3Nim,2Ocarboxyl}, {3Nim, 3Ocarboxyl} in a tetra, penta or octahedral arrange-ment.
The highly flexible unstructured main chain is able to wraparound the metal ion in order to be able to coordinate it with threeimidazoles of histidines.
In Fig. 8 an overlaid of selected aromatic regions of 1H-1HNMR ROESY and TOCSY spectra of Zn2+ bound-30aa peptideat 1 : 1 peptide to metal molar ratio, is reported. As we can see,several new cross peaks can be observed in the ROESY spectraupon Zn(II) addition.
Fig. 8 Overlaid of selected aromatic region of 1H-1H NMR ROESY (green) and TOCSY (blue) spectra of Zn(II) bound 30aa peptide at 1 : 1 peptide tometal molar ratio. New ROE cross-peaks observed upon Zn-binding have been labelled.
1290 | Dalton Trans., 2010, 39, 1282–1294 This journal is © The Royal Society of Chemistry 2010
Dow
nloa
ded
by U
nive
rsita
' deg
li St
udi d
i Sas
sari
on
28 D
ecem
ber
2010
Publ
ishe
d on
26
Nov
embe
r 20
09 o
n ht
tp://
pubs
.rsc
.org
| do
i:10.
1039
/B91
4296
GView Online
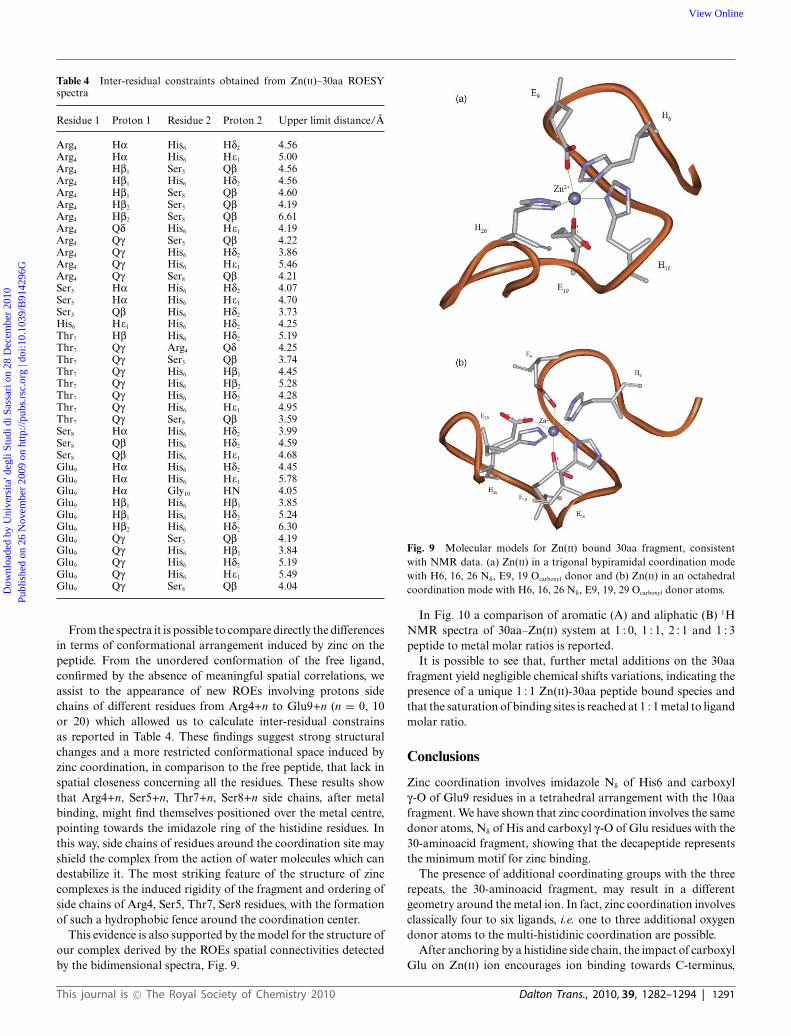
Table 4 Inter-residual constraints obtained from Zn(II)–30aa ROESYspectra
Residue 1 Proton 1 Residue 2 Proton 2 Upper limit distance/A
Arg4 Ha His6 Hd2 4.56Arg4 Ha His6 He1 5.00Arg4 Hb1 Ser5 Qb 4.56Arg4 Hb1 His6 Hd2 4.56Arg4 Hb1 Ser8 Qb 4.60Arg4 Hb2 Ser5 Qb 4.19Arg4 Hb2 Ser8 Qb 6.61Arg4 Qd His6 He1 4.19Arg4 Qg Ser5 Qb 4.22Arg4 Qg His6 Hd2 3.86Arg4 Qg His6 He1 5.46Arg4 Qg Ser8 Qb 4.21Ser5 Ha His6 Hd2 4.07Ser5 Ha His6 He1 4.70Ser5 Qb His6 Hd2 3.73His6 He1 His6 Hd2 4.25Thr7 Hb His6 Hd2 5.19Thr7 Qg Arg4 Qd 4.25Thr7 Qg Ser5 Qb 3.74Thr7 Qg His6 Hb1 4.45Thr7 Qg His6 Hb2 5.28Thr7 Qg His6 Hd2 4.28Thr7 Qg His6 He1 4.95Thr7 Qg Ser8 Qb 3.59Ser8 Ha His6 Hd2 3.99Ser8 Qb His6 Hd2 4.59Ser8 Qb His6 He1 4.68Glu9 Ha His6 Hd2 4.45Glu9 Ha His6 He1 5.78Glu9 Ha Gly10 HN 4.05Glu9 Hb1 His6 Hb1 3.85Glu9 Hb1 His6 Hd2 5.24Glu9 Hb2 His6 Hd2 6.30Glu9 Qg Ser5 Qb 4.19Glu9 Qg His6 Hb1 3.84Glu9 Qg His6 Hd2 5.19Glu9 Qg His6 He1 5.49Glu9 Qg Ser8 Qb 4.04
From the spectra it is possible to compare directly the differencesin terms of conformational arrangement induced by zinc on thepeptide. From the unordered conformation of the free ligand,confirmed by the absence of meaningful spatial correlations, weassist to the appearance of new ROEs involving protons sidechains of different residues from Arg4+n to Glu9+n (n = 0, 10or 20) which allowed us to calculate inter-residual constrainsas reported in Table 4. These findings suggest strong structuralchanges and a more restricted conformational space induced byzinc coordination, in comparison to the free peptide, that lack inspatial closeness concerning all the residues. These results showthat Arg4+n, Ser5+n, Thr7+n, Ser8+n side chains, after metalbinding, might find themselves positioned over the metal centre,pointing towards the imidazole ring of the histidine residues. Inthis way, side chains of residues around the coordination site mayshield the complex from the action of water molecules which candestabilize it. The most striking feature of the structure of zinccomplexes is the induced rigidity of the fragment and ordering ofside chains of Arg4, Ser5, Thr7, Ser8 residues, with the formationof such a hydrophobic fence around the coordination center.
This evidence is also supported by the model for the structure ofour complex derived by the ROEs spatial connectivities detectedby the bidimensional spectra, Fig. 9.
Fig. 9 Molecular models for Zn(II) bound 30aa fragment, consistentwith NMR data. (a) Zn(II) in a trigonal bypiramidal coordination modewith H6, 16, 26 Nd, E9, 19 Ocarboxyl donor and (b) Zn(II) in an octahedralcoordination mode with H6, 16, 26 Nd, E9, 19, 29 Ocarboxyl donor atoms.
In Fig. 10 a comparison of aromatic (A) and aliphatic (B) 1HNMR spectra of 30aa–Zn(II) system at 1 : 0, 1 : 1, 2 : 1 and 1 : 3peptide to metal molar ratios is reported.
It is possible to see that, further metal additions on the 30aafragment yield negligible chemical shifts variations, indicating thepresence of a unique 1 : 1 Zn(II)-30aa peptide bound species andthat the saturation of binding sites is reached at 1 : 1 metal to ligandmolar ratio.
Conclusions
Zinc coordination involves imidazole Nd of His6 and carboxylg-O of Glu9 residues in a tetrahedral arrangement with the 10aafragment. We have shown that zinc coordination involves the samedonor atoms, Nd of His and carboxyl g-O of Glu residues with the30-aminoacid fragment, showing that the decapeptide representsthe minimum motif for zinc binding.
The presence of additional coordinating groups with the threerepeats, the 30-aminoacid fragment, may result in a differentgeometry around the metal ion. In fact, zinc coordination involvesclassically four to six ligands, i.e. one to three additional oxygendonor atoms to the multi-histidinic coordination are possible.
After anchoring by a histidine side chain, the impact of carboxylGlu on Zn(II) ion encourages ion binding towards C-terminus,
This journal is © The Royal Society of Chemistry 2010 Dalton Trans., 2010, 39, 1282–1294 | 1291
Dow
nloa
ded
by U
nive
rsita
' deg
li St
udi d
i Sas
sari
on
28 D
ecem
ber
2010
Publ
ishe
d on
26
Nov
embe
r 20
09 o
n ht
tp://
pubs
.rsc
.org
| do
i:10.
1039
/B91
4296
GView Online

Fig. 10 Comparison of (a) aromatic and (b) aliphatic region of 1H NMR spectra of Zn(II)-30aa peptide system in 1 : 0, 1 : 1, 1 : 2 and 1 : 3 peptide tometal molar ratio at pH 7.
stabilizing the His16 and then the third consecutive His26 bindingsite from the peptide sequence. Nevertheless, the selective observedreduction on intensity and line broadening of carboxyl Hg Glusignal seems to point towards a less precise binding mode ofmetal which prevails for the 30aa peptide, compared to the shorterfragment.
In fact, the shift and broadening of Hg Glu signal can beinterpreted as due to a slow, intermediate chemical exchange rateregime with respect to the NMR timescale, between Zn-bound to
all the donor atoms and possibly, free carboxyl oxygens, to givedifferent species in a dynamic equilibrium among them, whichmight involve N3O, N3O2 and eventually, N3O3 chromophores ina tetra, penta or an octahedral arrangement.
Thus an equilibrium among some different coordinated speciescan be assumed. Actually, as Zn(II) ion can easily become penta co-ordinated, besides the three histidines, two intermediate carboxylgroups can be simultaneously involved in the zinc coordination inorder to decrease the size of the chelate rings.
1292 | Dalton Trans., 2010, 39, 1282–1294 This journal is © The Royal Society of Chemistry 2010
Dow
nloa
ded
by U
nive
rsita
' deg
li St
udi d
i Sas
sari
on
28 D
ecem
ber
2010
Publ
ishe
d on
26
Nov
embe
r 20
09 o
n ht
tp://
pubs
.rsc
.org
| do
i:10.
1039
/B91
4296
GView Online

We conclude that the coordination site of Zn-30aa fragment ismost likely made of 5 to 6 donor atoms including all the threehistidines, His6, 16, 26 and two or eventually three carboxylic sidechains of Glu9, 19 and 29, which can complete the donor set in anapical position, by forming macrochelate systems.
When Zn binds the C-terminal portion of each fragment, itfolds around the metal, inducing strong conformational changes,constraining it to leave the disordered conformation. The metalinduced “structuring” may have implication in the biologicalfunctions of the protein fragments. It strongly perturbs its normalconformation and could promote its pathological aggregation.
In order to focus on the structural changes caused by the metalion, we build a model of the induced structure of the peptide whenbound to zinc. A trigonal bypiramidal as well as an octahedralgeometry around the metal ion seem to fit experimental constrainsobtained from the ROESY NMR results.
Like Cu(II), Zn(II) ion shows to have strong affinity for histidineimidazole rings. The presence of three imidazoles within thepeptide sequence allows zinc ion to form poly-imidazole bindingmode which dominates in the physiological pH range. Coppershares with zinc the multi-histidinic binding mode but, unlike zinc,Cu(II) binding prevails at mild acidic conditions, as we previouslyreported.38
The multi-imidazole coordination mode may be critical for thebiological implications.
It is possible that Zn(II) ions will compete for the same histidineligands as Cu(II). Thus, taking into account that the levels of zincand copper are elevated in amyloid deposits, 1mM and 400 mM,respectively, zinc may play a neuroprotective role since Ab maycapture redox inactive zinc ions, thereby preventing Ab fromparticipating in redox cycling with other metals, in particular,Cu(II) ions. Therefore, zinc could have a competitive role asredox silencing Cu(II) ions or, as promoting peptide aggregates,and these ligands may act as seeding factors in the formationof amyloid plaques. Under physiological conditions, Ab has apropensity to bind zinc, whereas under mild acidic conditions, asin physiologic acidosis following an inflammatory process, copperis preferentially bound.18,39,40
This difference between copper and zinc affinity at differentpH values, has been suggested to be a pathogenic mechanismof Ab and is also consistent with the diverse role of the twometals in affecting the interaction of prion peptides with modelmembranes.41
Thus, we can conclude that such a common multi-histidinicbinding mode could consequently suggest a common biologicalrole operating in histidine rich domains proteins.
We believe that our study could give an additional clue tounderstanding the role of zinc binding in crucial multi-histidinicprotein, such as prion protein or amyloid precursor proteininvolved in neurodegenerative disorders.
Acknowledgements
This work was supported by the Regione Autonoma Sardegna“Master and Back” programme and the Fondazione Bancodi Sardegna, Sassari, Sardegna, Italy. CERM (University ofFlorence) and Prof. Ivano Bertini are gratefully acknowledgedfor the use of spectroscopic facilities. Porto Conte Ricerche,
Tramariglio, Alghero is gratefully acknowledged for the use ofNMR facilities.
References
1 A. Sigel, H. Sigel and R. K. O. Sigel, (Eds), (Neurodegenerative Diseasesand Metal Ions in Life Sciences, Wiley, Chichester, 2006, Vol. 1.
2 C. D. Syme, R. C. Nadal, S. E. J. Rigby and J. H. Viles, J. Biol. Chem.,2004, 279, 18169–18177.
3 G. S. Jackson, I. Murray, L. L. P. Hosszu, N. Gibbs, J. P. Waltho,A. R. Clarke and J. Collinge, Proc. Natl. Acad. Sci. U. S. A., 2001, 98,8531–8535.
4 G. L. Millhauser, Annu. Rev. Phys. Chem., 2007, 58, 299–320.5 E. Gaggelli, H. Kozlowski, D. Valensin and G. Valensin, Chem. Rev.,
2006, 106, 1995–2044.6 R. P. Bonomo, G. Impellizzeri, G. Pappalardo, E. Rizzarelli and G.
Tabbı, Chem.–Eur. J., 2000, 6, 4195–4202.7 P. Faller and C. Hureau, Dalton Trans., 2009, 1080–1094.8 A. I. Bush, Trends Neurosci., 2003, 26, 207–214.9 P. W. Mantyh, J. R. Ghilardi, S. Rogers, E. DeMasters, C. J. Allen, E. R.
Stimson and J. E. Maggio, J. Neurochem., 1993, 61, 1171–1174.10 A. I. Bush, W. H. Pettingell, G. Multhaup, M. de Paradis, J. P. Vonsattel,
J. F. Gusella, K. Beyreuther, C. I. Masters and R. E. Tanzi, Science,1994, 265, 1464–1467.
11 S. Zirah, S. A. Kozin, A. K. Mazur, A. Blond, M. Cheminant, I. Segalas-Milazzo, P. Debey and S. Rebuffat, J. Biol. Chem., 2005, 281, 2151–2161.
12 W. P. Esler, E. R. Stimson, J. M. Jennings, J. R. Ghilardi, P. W. Mantyhand J. E. Maggio, J. Neurochem., 1996, 66, 723–732.
13 S. W. Suh, K. B. Jensen, M. S. Jensen, D. S. Silva, P. J. Kesslak, G.Danscher and C. J. Frederickson, Brain Res., 2000, 852, 274–278.
14 M. P. Cuajungco and K. Y. Faget, Brain Res. Rev., 2003, 41, 44–56.
15 M. P. Cuajungco, C. J. Frederickson and A. I. Bush, SubcellularBiochemistry, 2005, 38, 235–254.
16 C. C. Curtain, F. Ali, I. Volitakis, R. A. Cherny, R. S. Norton, K.Beyreuther, C. J. Barrow, C. L. Masters, A. I. Bush and K. J. Barnham,J. Biol. Chem., 2001, 276, 20466–20473.
17 M. Valko, H. Morris and M. T. Cronin, Curr. Med. Chem., 2005, 12,1161–1208.
18 T. Miura, K. Suzuki, N. Kohata and H. Takeuchi, Biochemistry, 2000,39, 7024–7031.
19 P. C. Pauly and D. A. Harris, J. Biol. Chem., 1998, 273, 33107–33110.
20 W. S. Perera and N. M. Hooper, Curr. Biol., 2001, 11, 519–523.21 A. G. Kenvard, L. J. Bartolotti and C. S. Burns, Biochemistry, 2007, 46,
4261–4271.22 E. D. Walter, D. J. Stevens, M. P. Visconte and G. L. Millhauser, J. Am.
Chem. Soc., 2007, 129, 15440–15441.23 M. A. Zoroddu, M. Peana, T. Kowalik-Jankowska, H. Kozlowski and
M. Costa, J. Inorg. Biochem., 2004, 98, 931–939.24 M. A. Zoroddu, T. Kowalik-Jankowska, M. Peana, S. Medici and H.
Kozlowski, Dalton Trans., 2008, 6127–6134.25 M. A. Zoroddu, M. Peana and S. Medici, Dalton Trans., 2009, 5523–
5534.26 J. Meienhofer, M. Waki, E. P. Heimer, T. J. Lambros, R. C. Makofske
and C. D. Chang, Int. J. Pept. Protein Res., 1979, 13, 35–42.27 A. G. Palmer III, J. Cavanagh, P. E. Wright and M. Rance, J. Magn.
Reson., 1991, 93, 151–170.28 L. E. Kay, P. Keifer and T. Saarinen, J. Am. Chem. Soc., 1992, 114,
10663–10665.29 J. Schleucher, M. Schwendinger, M. Sattler, P. Schmidt, O. Schedletzky,
S. J. Glaser, O. W. Sorensen and C. Griesinger, J. Biomol. NMR, 1994,4.
30 T. L. Hwang and A. J. Shaka, J. Magn. Reson., Ser. A, 1995, 112,275–279.
31 A. Bax and D. G. Davis, J. Magn. Reson., 1985, 63, 207–213.32 W. Bocian, P. Borowicz, J. Mikołajczyk, J. Sitkowski, A. Tarnowska,
E. Bednarek, T. Głabski, B. Tejchman-Małecka, M. Bogiel and L.Kozerski, Biopolymers, 2008, 89, 820–830.
33 T. D. Goddard and D. G. Kneller, SPARKY 3, University of California,San Francisco.
34 A. Kilshtain-Vardi, M. Glick, H. M. Greenblatt, A. Goldblum and G.Shoham, Acta Crystallogr., 2003, 59, 323–333.
This journal is © The Royal Society of Chemistry 2010 Dalton Trans., 2010, 39, 1282–1294 | 1293
Dow
nloa
ded
by U
nive
rsita
' deg
li St
udi d
i Sas
sari
on
28 D
ecem
ber
2010
Publ
ishe
d on
26
Nov
embe
r 20
09 o
n ht
tp://
pubs
.rsc
.org
| do
i:10.
1039
/B91
4296
GView Online

35 J. Bouckaert, F. Poortmans, L. Wyns and R. Loris, J. Biol. Chem., 1996,271, 16144–16150.
36 D. K. Wilson and F. A. Quicho, Biochemistry, 1993, 32, 1689–1694.
37 C. D. Syme and J. H. Viles, Biochim. Biophys. Acta, Proteins Proteomics,2006, 1764, 246–256.
38 M. A. Zoroddu, S. Medici and M. Peana, J. Inorg,. Biochem,DOI: 10.1016/j.jinorgbio.2009.06.008.
39 Y. Yoshiike, K. Tanemura, O. Murayama, T. Agagi, M. Murayama, S.Sato, X. Sun, N. Tanaka and A. Takashima, J. Biol. Chem., 2001, 276,32293–32299.
40 C. S. Atwood, R. D. Moir, M. A. Hartshorn, R. E. Tanzi and A. I.Bush, J. Biol. Chem., 1998, 273, 12817–12826.
41 L. De Gioia, C. Selvaggini, E. Ghibaudi, L. Diomede, O. Bugiani,G. Forloni, F. Tagliavini and M. Salmona, J. Biol. Chem., 1994, 269,7859–7862.
1294 | Dalton Trans., 2010, 39, 1282–1294 This journal is © The Royal Society of Chemistry 2010
Dow
nloa
ded
by U
nive
rsita
' deg
li St
udi d
i Sas
sari
on
28 D
ecem
ber
2010
Publ
ishe
d on
26
Nov
embe
r 20
09 o
n ht
tp://
pubs
.rsc
.org
| do
i:10.
1039
/B91
4296
GView Online




















