
Developing a Digital Artifact for the Sustainable Presentation ...

lable at ScienceDirect
Journal of Human Evolution 57 (2009) 688–696
Contents lists avai
Journal of Human Evolution
journal homepage: www.elsevier .com/locate/ jhevol
Newly discovered fossil- and artifact-bearing deposits, uranium-series ages,and Plio-Pleistocene hominids at Swartkrans Cave, South Africa
Morris B. Sutton a,b, Travis Rayne Pickering a,c,*, Robyn Pickering d, C.K. Brain e, Ronald J. Clarke a,Jason L. Heaton f, Kathleen Kuman a,b
a Institute for Human Evolution, University of the Witwatersrand, WITS 2050, Johannesburg, South Africab School of Geography, Archaeology and Environmental Studies, University of the Witwatersrand, WITS 2050, Johannesburg, South Africac Department of Anthropology, University of Wisconsin, Madison, Wisconsin 53706, USAd Institute for Geological Sciences, University of Bern, CH-3012, Bern, Switzerlande Transvaal Museum, Pretoria, South Africaf Department of Biology, Birmingham-Southern College, Birmingham, Alabama 35254, USA
a r t i c l e i n f o
Article history:Received 30 December 2008Accepted 18 May 2009
Keywords:Site formationU-Th disequilibrium datingParanthropus (Australopithecus) robustusMiddle Stone Age archaeology
* Corresponding author.E-mail address: [email protected] (T.R. Pickerin
1 Unless monophyly of the robust australopithecinconvincingly, TRP and JLH object to the use of Paranprefer the use of Australopithecus robustus. Howeveinclination of many specialists, including RJC, we emplthroughout the rest of this paper.
0047-2484/$ – see front matter � 2009 Elsevier Ltd.doi:10.1016/j.jhevol.2009.05.014
a b s t r a c t
We report on new research at Swartkrans Cave, South Africa, that provides evidence of two previouslyunrealized artifact- and fossil-bearing deposits. These deposits underlie a speleothem dated by theuranium-thorium disequilibrium technique to 110,000� 1,980 years old, the first tightly constrained,geochronological date available for the site. Recovered fauna from the two underlying depos-itsdincluding, prominently, the dental remains of Paranthropus (Australopithecus) robustus from theuppermost layer (Talus Cone Deposit)dindicate a significantly older, late Pliocene or early Pleistoceneage for these units. The lowest unit (LB East Extension) is inferred to be an eastward extension ofthe well-known Lower Bank of Member 1, the earliest surviving infill represented at the site. Thedate acquired from the speleothem also sets the maximum age of a rich Middle Stone Age lithicassemblage.
� 2009 Elsevier Ltd. All rights reserved.
Introduction
Swartkrans Cave is an important paleoanthropological sitelocated w40 km northwest of Johannesburg, in Gauteng Province,South Africa (Fig.1). Robert Broom and John Robinson’s (e.g., Broom,1949; Broom and Robinson, 1949, 1950, 1952) early research atSwartkrans (1948–1949 and 1951–1953) provided large samples ofthe Plio-Pleistocene hominid Paranthropus (Australopithecus)robustus1 and demonstrated that it was synchronous and sympatricwith Homo ergaster, a species that is probably a direct ancestor ofmodern humans. C.K. Brain’s (e.g., 1970, 1981, 1993a; Brain et al.,1988) subsequent work at the site (1965–1986) resulted in therecovery of large fossil and archaeological samples that, becausethey were collected with acute regard for stratigraphy andtaphonomy, provide essential context for those extinct hominids.
g).es can be established more
thropus robustus and insteadr, to conform to the currentoy the designation P. robustus
All rights reserved.
Importantly, Brain analyzed these samples within a robust actual-istic framework, allowing him to draw novel inferences about earlyhominid behavior at Swartkrans between c.1.8 and 1.0 Ma. Includedamong Brain’s most important results were: (1) the recognition thatmany of the hominids were collected in the cave as the prey of largecarnivores (Brain, 1981, 1993b); (2) the identification of bone toolsused by hominids for digging activities (Brain and Shipman, 1993);and (3) the identification of burned bones from Member 3, indi-cating some of the earliest known control of fire by hominids at c.1.0 Ma (Brain and Sillen, 1988). Brain also recovered archaeologicalsamples that include the second largest collection of DevelopedOldowan/Early Acheuelan stone tools in southern Africa (Clark,1993; Field,1999) and the largest and most behaviorally informativecollections of large mammal remains butchered by Earlier Stone Age(ESA) hominids in the subregion (Brain, 1993b; T.R. Pickering et al.,2004, 2005, 2007, 2008).
Cave form and dating
The stratigraphy of Swartkrans is extremely complex, havingformed through a number of cycles of deposition and erosion.Recognizing that erosion has obscured or removed all trace of many

Fig. 1. The global (a), international (b), and local (c) positions of Swartkrans Cave. The cave is part of a UNESCO World Heritage Site, called the ‘‘Cradle of Humankind’’ (‘‘COH’’), asoutlined in (c). Major ‘‘COH’’ paleontological sites, including Swartkrans, are indicated. Important South African paleoanthropological sites outside the ‘‘COH’’ are indicated anddistinguished from cities by boldface and non-italicization on the political map (b).
M.B. Sutton et al. / Journal of Human Evolution 57 (2009) 688–696 689
of these cycles, Brain (1993c) identified six remaining depositionalunits at Swartkrans (Fig. 2). Oldest to youngest, these units are theLower Bank (LB) of Member 1, the Hanging Remnant (HR) ofMember 1, Member 2, Member 3, Member 4, and Member 5. Balteret al. (2008) used uranium-lead (U-Pb) dating of bovid toothenamel to produce ages of 1.83�1.38 Ma for Member 1,1.36� 0.29 Ma for Member 2, and 0.83� 0.21 Ma for Member 3.Electron spin resonance ages on hominid and bovid tooth enamelfrom the HR are 2.11�0.21 Ma and 1.63� 0.16 Ma (Blackwell, 1994;Curnoe et al., 2001). However, enamel is an open isotopic system,with the potential for diagenetic gain and loss of U and Pb;considerable error ranges, up to 60% in the case of the U-Pb age forMember 1, are therefore associated with the age calculations.
Thus, the currently best accepted dating of Members 1–3 relieson biostratigraphy, which, on balance, places each unit between c.1.8–1.0 Ma. Parsing of the bovid and equid data leads some (e.g.,Vrba, 1985; Churcher and Watson, 1993) to assign dates morespecifically, with Member 1 at 1.7 Ma, Member 2 at 1.5 Ma, andMember 3 at 1.0 Ma. Brain (1993c) provides comprehensive justi-fication for the assigned chronological order of successive infillingsat Swartkrans and corroborating evidence of their discrete natures.
Members 1, 2, 3, and 5 occupy the main, excavated area ofSwartkrans (Fig. 2). During 1980–1981, Brain dug an east-westarchaeological test trench (w7 m�w3 m�w1 m) within anexisting lime miners’ trench and through the surface of theMember 4 deposit, which lies to the east of the main area of the site.Brain’s goals with this test trenching were to identify the east wallof the cave and to determine the relationship of Member 4 with therest of the cave system. Member 4 is a colluvial deposit thatoccupies an area of w62 m2 on the cave’s surface. More than 1500stone artifacts, many of them diagnostically Middle Stone Age(MSA), but no fossils, were recovered from the Member 4 testtrench dug by Brain. There is no evidence of diagenetic bonedissolution (see Karkanas et al., 2000) to explain the lack of fossils,and their deficiency may indicate the absence of a cave roof over-hanging the area now occupied by Member 4 during the periodwhen MSA sediments and artifacts were deposited. Bone preser-vation within dolomitic limestone caves is due largely to itsencasement in calcified conglomerates (i.e., sedimentary breccia),and bone breccia formation in a cave is typically dependent on anoverhanging roof from which lime-bearing water drips onto the
floor and calcifies sediments and its included bones and stone clasts(Brain, 1958, 1981; Latham, 1999). Alternatively, bones may havenever been deposited in the area during the MSA. Based on ouranalysis of the stone artifact assemblage, and particularly itsabundance of formal tools, we infer that hominids both manufac-tured and used tools in proximity to the Member 4 area (Suttonet al., unpublished data). It is unlikely that the cave was actuallyused by hominids; instead, anthropogenic activity probablyoccurred outside on the hillside near the cave entrance. Slopewashand gravitation then transported the sediment and its includedartifacts down the hillside where it was contained by dolomiteblocks. The Member 4 colluvium extends down into the cave as theouter layer of a talus cone. The cone is at least 12 m in depth, witha 40� slope and a base of 19 m across its north face. The west side ofthis talus cone abuts the lower extent of the LB of Member 1 (Fig. 2).
Swartkrans Paleoanthropological Research Project (SPRP)
In 2005, we initiated the SPRP, a new program of research atSwartkrans coordinated by CKB and directed by TRP. The SPRP iscurrently focusing excavations on the surface of Member 4. Fortu-itously, we are also able to excavate the north side of the large taluscone on which the Member 4 MSA deposit rests, w10 m below oursurface excavation. The underground excavation is accessedthrough a natural aven that was subsequently expanded by theactivities of lime miners operating at Swartkrans in the mid-20thcentury. A lime miners’ cut into the north side of the talus conerevealed a decalcified deposit of cave sediment. This cuttingextended into the cone w2 m and effectively removed a wedge-shaped section of deposit 50–110 cm deep from the talus cone wall.Our underground excavation began at the miners’ cutting in 2006and has since expanded. Here we report on our initial findings.
Methods
Surface excavation
An EDM laser theodolite was used to establish a square metergrid over the floor of Brain’s original 1980–1981 test trench on thesurface of Member 4 (Fig. 2). The Member 4 grid is now part of Brain’soverall grid system, with the square coordinates used in the current

Fig. 2. Oblique photographic view of the surface of Swartkrans Cave (a), taken from the southwestern edge of the site, as indicated by the starred black arrow on the plan view (b).Note the spatial relationships of the Hanging Remnant and Lower Bank of Member 1 and the Member 4 area, and especially the proximity of the latter two units. The plan viewillustrates several of the remaining surficial deposits of the Swartkrans Formation (abbreviations: HR¼Member 1 Hanging Remnant; LB¼Member 1 Lower Bank; M3¼Member 3;M4¼Member 4), including overlap of discrete stratigraphic units (b). Member 2 is nearly entirely removed by previous excavation and is thus not pictured to provide clearer viewof the distribution of the remaining deposits; Member 5, an 11,000 year old deposit, is also removed in plan view to provide clearer indication of the Member 1 deposits in thenorth-central portion of the site. Detail of the Member 4 area, with our excavation grid superimposed, is shown in (c). The Member 4 area includes the Middle Stone Age (MSA)surface excavation (shaded) and the underlying Talus Cone Deposit and Lower Bank East Extension deposit, exposed in our excavations to the north (open circles) of the surfaceexcavation. Member 4 area stratigraphy is shown schematically in profile in (d): the Lower Bank East Extension is a largely decalcified yellowish-red (5YR 4/6), sandy silt, with onlyoccasional very small to small clasts of subangular dolomite that contains fossils and has areas of remaining calcification; the Talus Cone Deposit is >12 m thick and is composed ofdark red (2.5YR 3/6) decalcified silty sand, with small to medium sized (10–25 cm) clasts of angular and sub-angular dolomite, contains an abundance of fossils, but no MSA stoneartifacts deeper than its outer 20 cm; the SWK 4 flowstone is 2–3 cm thick, with a mean U-Th age of 110,000�1,980 years old; the MSA tool-bearing deposit is 110–120 cm in depthand composed of dark reddish brown (5YR 3/3), loose, non-calcified silty sand, with abundant small to large clasts of angular dolomite that are heavily stained with manganese.
M.B. Sutton et al. / Journal of Human Evolution 57 (2009) 688–696690
excavation corresponding to each square’s position within the cavesystem. Each square was excavated by quadrant (50 cm� 50 cm) at5 cm and 10 cm depths. Six squares, 2N 21E, 2N 22E, 3N 21E, 3N 22E,4N 21E, and 4N 22E, were excavated. The first four squares occurwithin Brain’s 1980–1981 trench. Squares 4N 21E and 4N 22E wereopened to provide a sample from the original surface of the depositsince their north halves are beyond the north wall of the trench. Allartifacts�20 mm in length were recorded by position with the EDMbefore removal. Each quadrant level was excavated separately andthen sieved using a 5 mm mesh over a 2 mm mesh screen. Thematerial from the 5 mm mesh was then wet sieved. The 2–5 mmfraction material was sorted to ensure the collection of small bonefragments, microfaunal remains, and stone artifact chips.
Underground excavation
We excavated four square meters (9N 18E, 9N 19E, 10N 18E, and10N 19E) to a depth of 175 cm underground, on the northside of theMember 4 talus, using the same methods as the surface excavation.
In addition, so that deposit formation processes might be bestinferred, we conducted preliminary fabric analyses on sedimentaryclasts included in the underground strata. Fabric analysis plots theangular values of direction for stones in a stratum in order toproduce hypotheses of the deposit’s origins and dynamics of itsaccumulation. Fabric analysis data collection followed the methodsof Bertran and Texier (1995) and Lenoble and Bertran (2004): usinga Brunton� GEO Transit instrument inclination and orientationmeasurements were taken on all stones >2 cm in length and withan elongation ratio (length/width; Drake, 1974) of >1.6. Fabricanalysis data were plotted with Stereonet software (developed byRichard W. Allmendinger).
Dating
A sample of the flowstone (SWK 4) underlying the Member 4MSA unit was extracted for U-Th dating with hammers and chiselsfrom the northside of the underground talus cone, beneath theMember 4 surface. Blocks of the flowstone sample were subsampled

M.B. Sutton et al. / Journal of Human Evolution 57 (2009) 688–696 691
(SWK 4-1, 4-4, and 4-5) mechanically using a hand-held diamond-wheel saw. The subsamples were then cleaned ultrasonically indistilled H2O. Analytical procedures for the separation and concen-tration of U and Th, as described by Ivanovich and Harmon (1992),were undertaken in the clean laboratory environment (PicoTrace�)of the Institute for Geological Sciences, University of Bern,Switzerland. Uranium and thorium isotopes were measured sepa-rately on a Nu Instruments� multicollector ICP-Mass spectrometerfollowing the protocol described by Fleitmann et al. (2007).
Hominid fossil analysis
We provide preliminary descriptions of hominid dental remainsrecovered from our underground excavations in the Member4 area. Each specimen was examined using a low power binocularmicroscope and, when possible, measurements were taken witha taper-ended, dial vernier caliper. Enamel thickness was measuredalong natural fracture surfaces for two permanent postcanine teethusing methods described by White et al. (2006). Because one ofthese specimens is not assignable to a more specific category oftooth, we cannot distinguish at which cusps its fracture surfaceoccurs, but we were able to ensure that measurements were alonga radial section, ‘‘i.e., on sections that were considered to approx-imately exhibit local minimal distances from the enamel-dentinejunction (EDJ) to the outer enamel surface’’ (White et al., 2006:Supplementary Discussion 2). For the fragment that we were ableto categorize as a right upper first molar, we took measurementsalong radial sections at the metacone and paracone and at themesial and distal crown face. Other analytical techniques might, insome cases, come closer to capturing enamel thickness acrossa tooth’s entire crown (see review in Teaford, 2006), but becausethe new Swartkrans specimens are already fragmentary we areunwilling to sacrifice any more of each to provide more detailedmeasurements.
Results
Surface excavation
The excavation on the floor of Brain’s 1980-1981 test trench(squares 2N 21E, 2N 22E, 3N 21E, 3N 22E) immediately yieldedstone artifacts. Approximately 40 artifacts per 5 cm level wererecovered in the upper levels (40 cm) of square 2N 21E. However,beyond these depths only a few artifact flake fragments per levelwere recovered, and eventually at w60 cm below the floor ofBrain’s original test trench the deposit became archaeologicallysterile. A similar pattern existed in the upper levels of 2N 22E, 3N21E, and 3N 22E, and it was decided to halt excavations of thesesquares and continue down in 2N 21E to determine the naturalcontour of the deposits. Artifact density parallels a change in thematrix. The floor and walls of the test trench are composed of a darkreddish, slightly organic, silty sand. Then, at w40 cm, a moreconsolidated, reddish silty sand is exposed. This relatively consol-idated layer continues to a depth of 130 cm, at which point exca-vation on 2N 21E was discontinued. Squares 4N 21E and 4N 22E,which began at the original surface above Brain’s trench, alsoyielded dense artifact levels until reaching the matrix change orunderlying deposit. It is evident the MSA artifacts exist in the upper110 cm (from original untrenched surface) layer, which overliesa larger Talus Cone Deposit (TCD). Thus far w4000 artifacts havebeen recovered from the surface excavations in the top 110 cm ofthe colluvial deposits.
A detailed report on the MSA artifacts is being prepared (Suttonet al., unpublished data). However, a general summary here providesimportant insights into the nature of the assemblage (Fig. 3). The
entire assemblage is composed of three primary raw materials:quartzite, quartz, and diabase. Of the cores and cortical flakes themajority reflects a river cobble origin for the lithics and indicatesfurther that the nearby Bloubank River gravels were the primarysource of raw materials. A full size range of flakes and blades isrepresented in the assemblage. The broad range of flake size and thesubstantial percentage (>55%) of small (<20 mm in maximumdimension) flaking debris indicate a high level of ‘‘site capture’’ andthe near primary context of the assemblage (e.g., Schick, 1987).Corroborating this inference, most of the artifacts are in ‘‘fresh’’condition; only 19% have weathered surfaces and that weatheredmaterial mostly consists of diabase pieces. Less than 5% of the�2 mm flake debris subassemblage exhibits edge damage. Collec-tively, these observations suggest rapid burial and minimal move-ment of artifacts within the colluvium (e.g., little or no trampling)after their deposition.
Underground excavation
A small number of stone artifacts with clear MSA attributes wasrecovered in the upper/outer 10–15 cm of the Member 4 talus conein squares 9N 18E and 9N 19E. This was also the case for squares10N 18E and 10N 19E, but in the latter square we also uncovereda 2–3 cm thick flowstone at w18 cm in depth (Figs. 2d, 4). Werecovered fossilized bone, including hominid specimens (seebelow), but only a few nondiagnostic artifact flake fragments belowthe speleothem in the TCD. We infer that the flowstone isa surviving portion of an extensive calcite capstone formed over thenorth portion of the talus cone, clearly separating the overlyingMSA levels from two distinct underlying fossiliferous strata. Otherremnants of this capping flowstone were revealed in the exposedportions of the talus cone wall in squares 9N 18E and 9N 19E.However, because this section is cut deeper in the talus cone andthe dolomite cave roof angles sharply above it, the flowstone wasbroken and incorporated into the talus deposit in this area.
Excavation continued in all four square meters to a depth of 2 m.At w110 cm we reached a clear unconformity between the TCD andan underlying stratified deposit (Figs. 2d and 4). This lowest depositcontains fossils, but no stone artifacts have been recovered. Thedeposit is also distinct sedimentologically from the overlying TCDand instead appears identical to the LB of Member 1 in its matrixcolor and consistency. We infer this deposit is actually an eastwardextension of the LB and designate it the LB East Extension, asdistinguished from the overlying TCD.
Fabric patterns of the stone embedded in the underground sedi-ments support the inference of two distinct deposits below the datedflowstone (Fig. 4). We note, however, that fabric pattern results shouldbe regarded as preliminary since Lenoble and Bertran (2004)recommend a sample size of 40–50 measured pieces to ensurea stable Vector Magnitude. Because the Swartkrans dolomiteweathers predominantly into cube-shaped blocks, elongated pieces>2 cm in our underground excavations are rare and only 10 pieces perdeposit were suitable for measurement. Thus, even though our fabricdata were collected from several excavated levels, our overall samplesstill fall short of the recommended sample size. With that in mind, ourresults do show clear patterns. Pieces in the LB East Extension wereoriented east-northeast, reflecting a sediment flow direction from theLB, meeting the predictions of a hypothesis that those two ‘‘deposits’’are actually one homogenous and contiguous infill. The LB EastExtension stones are relatively flatly inclined (plunge range¼ 0�–20�;mean¼ 4�). In contrast, the TCD pieces are oriented north, with theirdips (plunge range¼ 20�–40�; mean¼ 30.5�) clustered around theoverall slope angle of the talus cone (¼40�). This is what is predictedfor stones filtering down the outside edge of the talus cone from theoverlying, forming MSA deposit.

Fig. 3. Diagnostic Middle Stone Age stone tools from new excavations of Swartkrans Member 4: quartzite Levallois core (a); quartzite preferential flake core (b); quartzite bladecores (c, d); diabase denticulated blades (e, f); quartz convex end scrapers (g, h); quartzite denticulated flake (i); and quartzite denticulated blade (j).
M.B. Sutton et al. / Journal of Human Evolution 57 (2009) 688–696692
Analyses of the paleoanthropological materials from bothunderground deposits are ongoing, but we provide a preliminaryassessment of their condition here. As with other decalcified unitsat Swartkrans, the faunas from the TCD and LB East Extension arehighly comminuted. Thus far, we have identified only the recoveredhominid specimens to the species level; the presence of P. robustusfossils (described below) places the TCD as significantly older thanthe MSA level and the flowstone (see below) that both occur aboveit. Based on superposition, we currently infer that the LB EastExtension must be still older than the overlying MSA, capstone, andTCD units. The families Cercopithecidae, Canidae, Viverridae,Felidae, Bovidae, and Procaviidae are represented in both of thenewly recognized, underground deposits, taxa typical of other Plio-Pleistocene assemblages from Swartkrans (microfaunal remains areabundant but have not yet been identified more specifically).
Dating
Overall, U concentrations in subsamples SWK 4-1, 4-4, and 4-5 areconsistently w60 ppb, which is in the same range as other U-Th dated
flowstones from this region (R. Pickering et al., 2007). 234U/238U ratiosare all above equilibrium, as to be expected for samples of this age.230Th/232Th ratios are well above 30, indicating that detritalcontamination is not a concern with the samples (Schwarcz, 1986).Table 1 summarizes U-Th disequilibrium dating results on the SWK4 subsamples, with a mean age of 110,330�1,980 years old for theflowstone layer. Because the flowstone from which the dated SWK4 subsamples derived is a discrete layer of rigid, well preserved,nonporous calcite, the 110,330�1,980 years old date thus providesthe maximum age of the Member 4 MSA artifacts.
Hominid fossils
There are three hominid dental fossils, all of which derive fromthe TCD level under the north face flowstone and above the LB EastExtension (Table 2). All three specimens display taxonomicallydiagnostic morphology that permits attribution to the speciesP. robustus (Fig. 5).
Specimen SWT/TC-2 is the lingual two-thirds of the crown ofa small right upper first molar. It is broken mesiodistally through
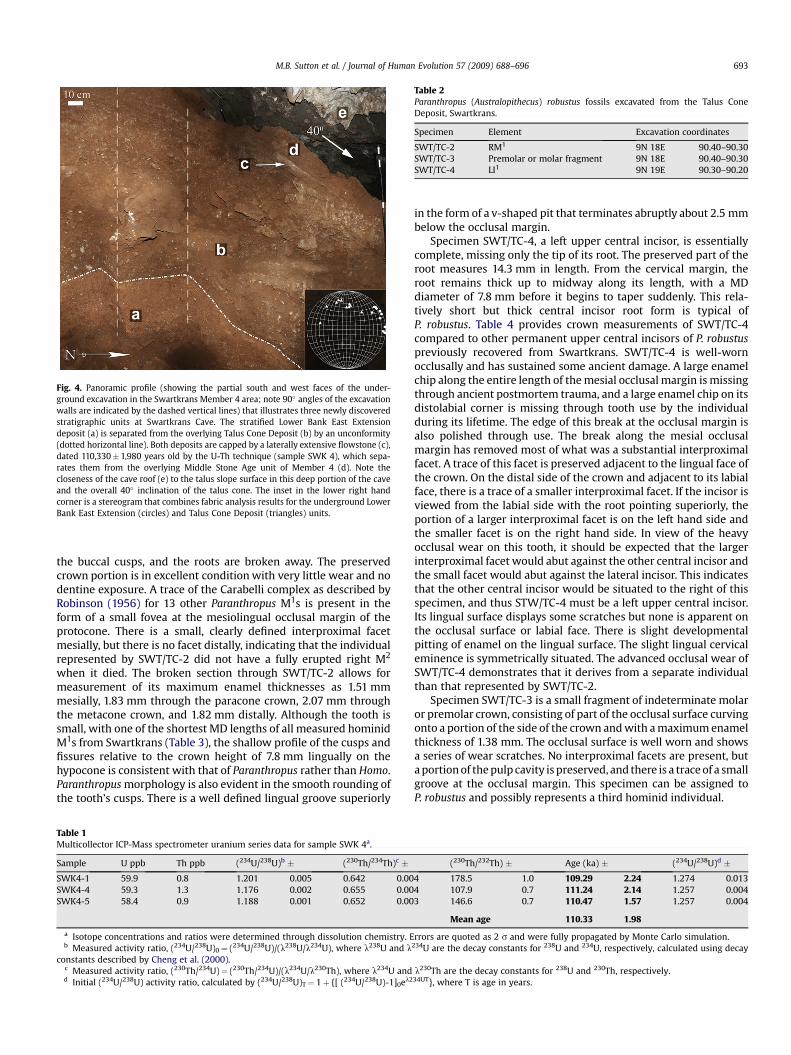
Fig. 4. Panoramic profile (showing the partial south and west faces of the under-ground excavation in the Swartkrans Member 4 area; note 90� angles of the excavationwalls are indicated by the dashed vertical lines) that illustrates three newly discoveredstratigraphic units at Swartkrans Cave. The stratified Lower Bank East Extensiondeposit (a) is separated from the overlying Talus Cone Deposit (b) by an unconformity(dotted horizontal line). Both deposits are capped by a laterally extensive flowstone (c),dated 110,330� 1,980 years old by the U-Th technique (sample SWK 4), which sepa-rates them from the overlying Middle Stone Age unit of Member 4 (d). Note thecloseness of the cave roof (e) to the talus slope surface in this deep portion of the caveand the overall 40� inclination of the talus cone. The inset in the lower right handcorner is a stereogram that combines fabric analysis results for the underground LowerBank East Extension (circles) and Talus Cone Deposit (triangles) units.
Table 2Paranthropus (Australopithecus) robustus fossils excavated from the Talus ConeDeposit, Swartkrans.
Specimen Element Excavation coordinates
SWT/TC-2 RM1 9N 18E 90.40–90.30SWT/TC-3 Premolar or molar fragment 9N 18E 90.40–90.30SWT/TC-4 LI1 9N 19E 90.30–90.20
M.B. Sutton et al. / Journal of Human Evolution 57 (2009) 688–696 693
the buccal cusps, and the roots are broken away. The preservedcrown portion is in excellent condition with very little wear and nodentine exposure. A trace of the Carabelli complex as described byRobinson (1956) for 13 other Paranthropus M1s is present in theform of a small fovea at the mesiolingual occlusal margin of theprotocone. There is a small, clearly defined interproximal facetmesially, but there is no facet distally, indicating that the individualrepresented by SWT/TC-2 did not have a fully erupted right M2
when it died. The broken section through SWT/TC-2 allows formeasurement of its maximum enamel thicknesses as 1.51 mmmesially, 1.83 mm through the paracone crown, 2.07 mm throughthe metacone crown, and 1.82 mm distally. Although the tooth issmall, with one of the shortest MD lengths of all measured hominidM1s from Swartkrans (Table 3), the shallow profile of the cusps andfissures relative to the crown height of 7.8 mm lingually on thehypocone is consistent with that of Paranthropus rather than Homo.Paranthropus morphology is also evident in the smooth rounding ofthe tooth’s cusps. There is a well defined lingual groove superiorly
Table 1Multicollector ICP-Mass spectrometer uranium series data for sample SWK 4a.
Sample U ppb Th ppb (234U/238U)b � (230Th/234Th)c �
SWK4-1 59.9 0.8 1.201 0.005 0.642 0.00SWK4-4 59.3 1.3 1.176 0.002 0.655 0.00SWK4-5 58.4 0.9 1.188 0.001 0.652 0.00
a Isotope concentrations and ratios were determined through dissolution chemistry. Eb Measured activity ratio, (234U/238U)0¼ (234U/238U)/(l238U/l234U), where l238U and l
constants described by Cheng et al. (2000).c Measured activity ratio, (230Th/234U)¼ (230Th/234U)/(l234U/l230Th), where l234U andd Initial (234U/238U) activity ratio, calculated by (234U/238U)T¼ 1þ {[ (234U/238U)-1]0el2
in the form of a v-shaped pit that terminates abruptly about 2.5 mmbelow the occlusal margin.
Specimen SWT/TC-4, a left upper central incisor, is essentiallycomplete, missing only the tip of its root. The preserved part of theroot measures 14.3 mm in length. From the cervical margin, theroot remains thick up to midway along its length, with a MDdiameter of 7.8 mm before it begins to taper suddenly. This rela-tively short but thick central incisor root form is typical ofP. robustus. Table 4 provides crown measurements of SWT/TC-4compared to other permanent upper central incisors of P. robustuspreviously recovered from Swartkrans. SWT/TC-4 is well-wornocclusally and has sustained some ancient damage. A large enamelchip along the entire length of the mesial occlusal margin is missingthrough ancient postmortem trauma, and a large enamel chip on itsdistolabial corner is missing through tooth use by the individualduring its lifetime. The edge of this break at the occlusal margin isalso polished through use. The break along the mesial occlusalmargin has removed most of what was a substantial interproximalfacet. A trace of this facet is preserved adjacent to the lingual face ofthe crown. On the distal side of the crown and adjacent to its labialface, there is a trace of a smaller interproximal facet. If the incisor isviewed from the labial side with the root pointing superiorly, theportion of a larger interproximal facet is on the left hand side andthe smaller facet is on the right hand side. In view of the heavyocclusal wear on this tooth, it should be expected that the largerinterproximal facet would abut against the other central incisor andthe small facet would abut against the lateral incisor. This indicatesthat the other central incisor would be situated to the right of thisspecimen, and thus STW/TC-4 must be a left upper central incisor.Its lingual surface displays some scratches but none is apparent onthe occlusal surface or labial face. There is slight developmentalpitting of enamel on the lingual surface. The slight lingual cervicaleminence is symmetrically situated. The advanced occlusal wear ofSWT/TC-4 demonstrates that it derives from a separate individualthan that represented by SWT/TC-2.
Specimen SWT/TC-3 is a small fragment of indeterminate molaror premolar crown, consisting of part of the occlusal surface curvingonto a portion of the side of the crown and with a maximum enamelthickness of 1.38 mm. The occlusal surface is well worn and showsa series of wear scratches. No interproximal facets are present, buta portion of the pulp cavity is preserved, and there is a trace of a smallgroove at the occlusal margin. This specimen can be assigned toP. robustus and possibly represents a third hominid individual.
(230Th/232Th) � Age (ka) � (234U/238U)d �
4 178.5 1.0 109.29 2.24 1.274 0.0134 107.9 0.7 111.24 2.14 1.257 0.0043 146.6 0.7 110.47 1.57 1.257 0.004
Mean age 110.33 1.98
rrors are quoted as 2 s and were fully propagated by Monte Carlo simulation.234U are the decay constants for 238U and 234U, respectively, calculated using decay
l230Th are the decay constants for 238U and 230Th, respectively.34UT}, where T is age in years.
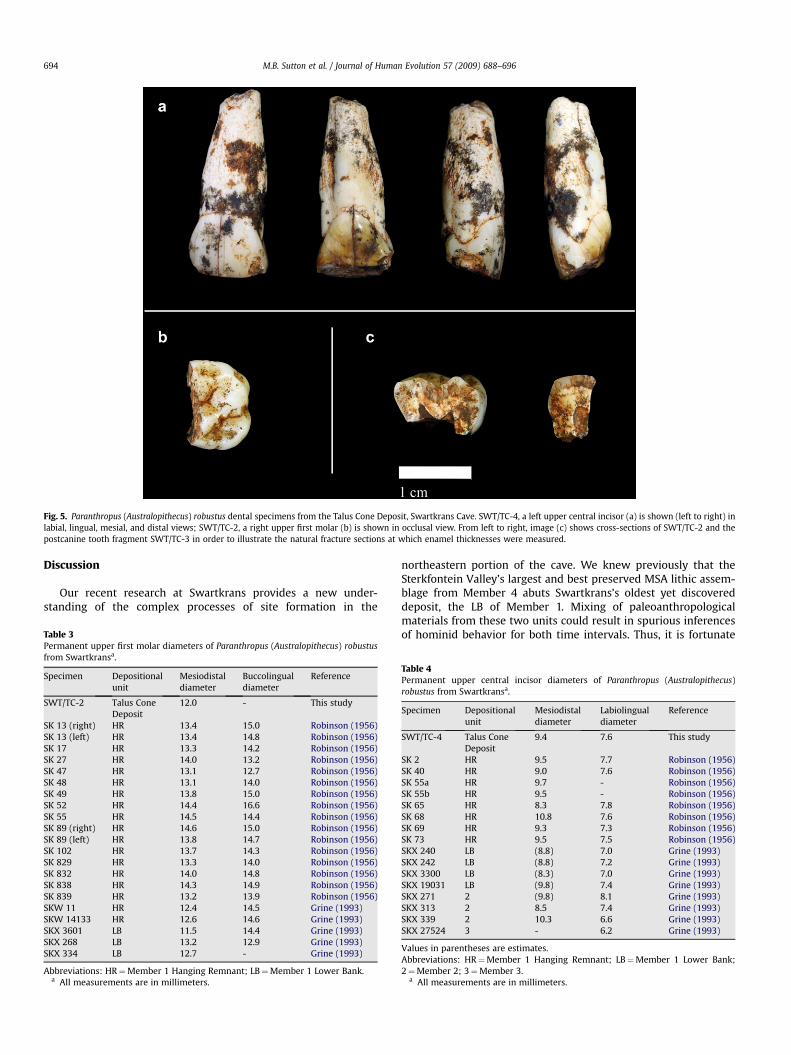
Fig. 5. Paranthropus (Australopithecus) robustus dental specimens from the Talus Cone Deposit, Swartkrans Cave. SWT/TC-4, a left upper central incisor (a) is shown (left to right) inlabial, lingual, mesial, and distal views; SWT/TC-2, a right upper first molar (b) is shown in occlusal view. From left to right, image (c) shows cross-sections of SWT/TC-2 and thepostcanine tooth fragment SWT/TC-3 in order to illustrate the natural fracture sections at which enamel thicknesses were measured.
M.B. Sutton et al. / Journal of Human Evolution 57 (2009) 688–696694
Discussion
Our recent research at Swartkrans provides a new under-standing of the complex processes of site formation in the
Table 3Permanent upper first molar diameters of Paranthropus (Australopithecus) robustusfrom Swartkransa.
Specimen Depositionalunit
Mesiodistaldiameter
Buccolingualdiameter
Reference
SWT/TC-2 Talus ConeDeposit
12.0 - This study
SK 13 (right) HR 13.4 15.0 Robinson (1956)SK 13 (left) HR 13.4 14.8 Robinson (1956)SK 17 HR 13.3 14.2 Robinson (1956)SK 27 HR 14.0 13.2 Robinson (1956)SK 47 HR 13.1 12.7 Robinson (1956)SK 48 HR 13.1 14.0 Robinson (1956)SK 49 HR 13.8 15.0 Robinson (1956)SK 52 HR 14.4 16.6 Robinson (1956)SK 55 HR 14.5 14.4 Robinson (1956)SK 89 (right) HR 14.6 15.0 Robinson (1956)SK 89 (left) HR 13.8 14.7 Robinson (1956)SK 102 HR 13.7 14.3 Robinson (1956)SK 829 HR 13.3 14.0 Robinson (1956)SK 832 HR 14.0 14.8 Robinson (1956)SK 838 HR 14.3 14.9 Robinson (1956)SK 839 HR 13.2 13.9 Robinson (1956)SKW 11 HR 12.4 14.5 Grine (1993)SKW 14133 HR 12.6 14.6 Grine (1993)SKX 3601 LB 11.5 14.4 Grine (1993)SKX 268 LB 13.2 12.9 Grine (1993)SKX 334 LB 12.7 - Grine (1993)
Abbreviations: HR¼Member 1 Hanging Remnant; LB¼Member 1 Lower Bank.a All measurements are in millimeters.
northeastern portion of the cave. We knew previously that theSterkfontein Valley’s largest and best preserved MSA lithic assem-blage from Member 4 abuts Swartkrans’s oldest yet discovereddeposit, the LB of Member 1. Mixing of paleoanthropologicalmaterials from these two units could result in spurious inferencesof hominid behavior for both time intervals. Thus, it is fortunate
Table 4Permanent upper central incisor diameters of Paranthropus (Australopithecus)robustus from Swartkransa.
Specimen Depositionalunit
Mesiodistaldiameter
Labiolingualdiameter
Reference
SWT/TC-4 Talus ConeDeposit
9.4 7.6 This study
SK 2 HR 9.5 7.7 Robinson (1956)SK 40 HR 9.0 7.6 Robinson (1956)SK 55a HR 9.7 - Robinson (1956)SK 55b HR 9.5 - Robinson (1956)SK 65 HR 8.3 7.8 Robinson (1956)SK 68 HR 10.8 7.6 Robinson (1956)SK 69 HR 9.3 7.3 Robinson (1956)SK 73 HR 9.5 7.5 Robinson (1956)SKX 240 LB (8.8) 7.0 Grine (1993)SKX 242 LB (8.8) 7.2 Grine (1993)SKX 3300 LB (8.3) 7.0 Grine (1993)SKX 19031 LB (9.8) 7.4 Grine (1993)SKX 271 2 (9.8) 8.1 Grine (1993)SKX 313 2 8.5 7.4 Grine (1993)SKX 339 2 10.3 6.6 Grine (1993)SKX 27524 3 - 6.2 Grine (1993)
Values in parentheses are estimates.Abbreviations: HR¼Member 1 Hanging Remnant; LB¼Member 1 Lower Bank;2¼Member 2; 3¼Member 3.
a All measurements are in millimeters.

M.B. Sutton et al. / Journal of Human Evolution 57 (2009) 688–696 695
that we are able to distinguish four clear stratigraphic divisions inour underground excavations. We summarize following their orderof discovery (youngest to oldest), rather than by conventionaldescription of stratigraphic units, from oldest to youngest:
(1) The uppermost layer is the MSA tool-bearing deposit, 110–120 cm in depth and composed of dark reddish-brown (5YR3/3), loose, noncalcified silty sand, with abundant small to largeclasts of angular dolomite that are heavily stained withmanganese. Unlike the MSA level at nearby Sterkfontein, wheresome diagnostic artifacts co-occur with reworked ESA mate-rials (Reynolds et al., 2007), the Swartkrans MSA artifactassemblage does not show any incorporation of older,reworked artifacts, and its large size renders it more informa-tive of MSA hominid behavior and technology. A probablyyounger (88,700�1,600–62,900�1,300 years old) MSA arti-fact assemblage from Plover’s Lake, only w6 km northeast ofSwartkrans, has not yet been published in detail (de Ruiteret al., 2008).
(2) The MSA stratum overlies a now-fragmented speleothem, witha mean U-Th age of 110,000�1,980 years old, which sets themaximum age for the MSA and serves as a capstone for twounderlying strata that necessarily must be older than c. 110,000years old.
(3) Immediately beneath the dated flowstone is the large TCD,>12 m thick. The TCD, a dark red (2.5YR 3/6) decalcified siltysand, with small to medium sized (10–25 cm) clasts of angularand subangular dolomite, contains an abundance of fossils butno MSA stone artifacts deeper than its outer 20 cm. The pres-ence of P. robustus fossils in the TCD corroborates the inferencebased on superposition that it is of greater (late Pliocene orearly Pleistocene) age than the overlying flowstone and MSAunits.
(4) The lowest sedimentary unit, a yellowish-red (5YR 4/6), sandysilt, with only occasional very small to small clasts of sub-angular dolomite, that contains fossils is termed the LB EastExtension. The LB East Extension is largely decalcified, but ithas areas of remaining calcification. The matrix of the LB EastExtension is consistent with that of the LB of Member 1 fromBrain’s original excavation area on the cave’s surface (Fig. 2),and we infer that the former is an eastward extension of thelatter.
We combine our new observations with Brain’s (1993c)previous interpretation of the formation of the Swartkrans Cavedeposits in the following, revised reconstruction, which focuses onthe deposits of Swartkrans northeast:
(1) Sometime probably in excess of 1.7 Ma, joints in the roof of anunderground cavern that was to become Swartkrans Caveopened between the ground surface and the cavern. This shaftbegan to admit surface-derived soil materials, which formeda talus slope below the shaft and extended toward the north-west corner of the cave; this deposit is now referred to as the LBof Member 1 (Brain, 1993c). We now infer that the infilling alsocontinued laterally toward the east for w50 m until it reachedthe east wall of the cave. The passage through which thiseastward extension of the LB traveled was very narrow due tothe extreme thickness of the cave’s roof near its north wall; thisnarrow channel between the roof and floor of the cave leveledthe LB East Extension deposit, as reflected in fabric orientation(ENE) in that portion of the cave. Eventually, but still in theearly Pleistocene (c. 1.8–1.0 Ma) based on faunal dates for thecontiguous surface LB of Member 1, the eastern part of the cavebecame choked with LB East Extension sediments.
(2) More recently in the Pleistocene, the continued dissolution ofdolomite by groundwater resulted in a partial collapse ofa section of cave roof in the Member 4 area. The resulting avenallowed the infilling of the Member 4 area TCD, as evidenced bythe large roof spall in the contact zone of the LB East Extensionand the overlying TCD and by the presence of surface dolomiterubble in the lower portions of the TCD. Fabric orientationreflects the steep northward advance of the talus formation.The talus cone eventually filled the opening, once againchoking off the area from further infill.
(3) A flowstone layer was deposited over the TCD in the latePleistocene, c. 110,000 years ago, indicating that the cave waspartially closed during this time and that its roof was relativelyintact.
(4) A MSA deposit covered the top of the talus cone in the Member4 area, with some material filtering down the north surface ofthe cone. The density of stone tools and absence of bonesuggest deposition of MSA materials after the dissolution andcollapse of the cave roof, or, alternatively, that the materialswere deposited just beyond the protection of an existing roof.Preliminary analysis of the MSA stone artifacts shows thata majority of the assemblage is in fresh condition with onlyminimal edge damage, suggesting little colluvial movement ofthe artifacts.
Conclusions
In 1993, C.K. Brain summed up his interpretation of siteformation at Swartkrans with a prescient statement: ‘‘The processof discovery is by no means over and I am convinced that as furtherwork is done there in the future, new surprises will surface, withthe result that revisions of the interpretation presented here will beneeded’’ (Brain, 1993c: 23). This paper describes the initial stage ofthe reinterpretation of Swartkrans, substantiating Brain’s earlierview of depositional and erosional cycles at the site and addingfurther detail to the history of infilling in the important north-eastern portion of the site.
Beyond reinterpretations of site formation, our augmentation ofthe MSA archaeological assemblage from Member 4 will provideimportant inferences relevant to the emergence of modern humanbehavior during the middle Pleistocene in southern Africa.Although there is a wealth of MSA sites in South Africa, the lithicsample from Swartkrans Member 4 is the largest and leastdisturbed from the Sterkfontein Valley. Further, the recovery ofadditional P. robustus fossils from the newly recognized TCD ofSwartkrans confirms the cave’s importance to understanding thatspecies’ evolutionary history and taxonomic status.
Acknowledgments
The Swartkrans Paleoanthropology Research Project (SPRP) issupported by grants to T.R. Pickering from the National ScienceFoundation (USA), the L.S.B. Leakey Foundation, and the Palae-ontology Scientific Trust (PAST), to Sutton from the NationalResearch Foundation (South Africa) and PAST, and to Kuman fromPAST. R. Pickering was supported by the Swiss National ScienceFoundation. We thank the following individuals, institutions, andcompanies (in alphabetical order) for their assistance with ourproject: African Explosives; the Brain family; Laurent Bruxelles;Anna Craven-Sutton; John Cruise; Manuel Domınguez-Rodrigo;Amy Egeland; Charles Egeland; Andrew Phaswana; the T.R. Pick-ering family; Stephany Potze; Dusty van Rooyen; the School ofGeography, Archaeology and Environmental Studies, University ofthe Witwatersrand; Francis Thackeray; the Transvaal Museum;

M.B. Sutton et al. / Journal of Human Evolution 57 (2009) 688–696696
Sarah Zwodeski. Thanks to Susan Anton, the Associate Editor andreviewers, and Darryl de Ruiter for comments that improved themanuscript. This is paper No. 2 in the SPRP publication series.
References
Balter, V., Blichet-Toft, J., Braga, J., Telouk, P., Thackeray, J.F., Albarede, F., 2008. U-Pbdating of fossil enamel from the Swartkrans Pleistocene hominid site, SouthAfrica. Earth Planet. Sci. Lett. 267, 236–246.
Bertran, P., Texier, J.P., 1995. Fabric analysis: application to Paleolithic studies.J. Archaeol. Sci. 22, 521–535.
Blackwell, B.A., 1994. Problems associated with reworked teeth and electron spinresonance (ESR) dating. Quatern. Sci. Rev. 13, 651–660.
Brain, C.K., 1958. The Transvaal Ape-Man-Bearing Cave Deposits. TransvaalMuseum, Pretoria.
Brain, C.K., 1970. New finds at the Swarkrans australopithecine site. Nature 225,1112–1119.
Brain, C.K., 1981. The Hunters or the Hunted? An Introduction to African CaveTaphonomy. University of Chicago Press, Chicago.
Brain, C.K. (Ed.), 1993a. Swartkrans: A Cave’s Chronicle of Early Man. TransvaalMuseum, Pretoria.
Brain, C.K., 1993b. A taphonomic overview of the Swartkrans fossil assemblages. In:Brain, C.K. (Ed.), Swartkrans: A Cave’s Chronicle of Early Man. TransvaalMuseum, Pretoria, pp. 257–264.
Brain, C.K., 1993c. Structure and stratigraphy of the Swartkrans Cave in the light ofthe new excavations. In: Brain, C.K. (Ed.), Swartkrans: A Cave’s Chronicle ofEarly Man. Transvaal Museum, Pretoria, pp. 23–33.
Brain, C.K., Shipman, P., 1993. The Swartkrans bone tools. In: Brain, C.K. (Ed.), Swartk-rans: A Cave’s Chronicle of Early Man. Transvaal Museum, Pretoria, pp. 195–215.
Brain, C.K., Sillen, A., 1988. Evidence from the Swartkrans Cave for the earliest use offire. Nature 336, 464–466.
Brain, C.K., Churcher, C.S., Clark, J.D., Grine, F.E., Shipman, P., Susman, R.L., Turner, A.,Watson, V., 1988. New evidence of early hominids, their culture and environ-ment from the Swartkrans Cave, South Africa. S. Afr. J. Sci. 84, 828–835.
Broom, R., 1949. Another new type of fossil ape-man (Paranthropus crassidens).Nature 163, 57.
Broom, R., Robinson, J.T., 1949. A new type of fossil man. Nature 164, 322.Broom, R., Robinson, J.T., 1950. Man contemporaneous with the Swartkrans ape-
man. Am. J. Phys. Anthropol 8, 151–156.Broom, R., Robinson, J.T., 1952. Swartkrans Ape-Man Paranthropus crassidens.
Transvaal Museum, Pretoria.Cheng, H., Edwards, R.L., Hoff, J., Gallup, C.D., Richards, D.A., Asmerom, Y., 2000. The
half-lives of uranium-234 and thorium-230. Chem. Geol. 169, 17–33.Churcher, C.S., Watson, V., 1993. Additional fossil Equidae from Swartkrans. In:
Brain, C.K. (Ed.), Swartkrans: A Cave’s Chronicle of Early Man. TransvaalMuseum, pp. 137–150.
Clark, J.D., 1993. Stone artifact assemblages from members 1–3, Swartkrans Cave.In: Brain, C.K. (Ed.), Swartkrans: A Cave’s Chronicle of Early Man. TransvaalMuseum, Pretoria, pp. 167–194.
Curnoe, D., Grun, R., Taylor, L., Thackeray, F., 2001. Direct ESR dating of a Pliocenehominin from Swartkrans. J. Hum. Evol. 40, 379–391.
Drake, P., 1974. Till fabric control by clast shape. Geol. Soc. Am. Bull. 85, 247–250.Field, A.S., 1999. An Analytic and Comparative Study of the Earlier Stone Age
Archaeology of the Sterkfontein Valley. M.Sc. thesis, University of the Witwa-tersrand, Johannesburg.
Fleitmann, D., Burns, S.J., Mangini, A., Mudelsee, M., Kramers, J.D., Villa, I., Neff, U.,Al-Subbary, A.A., Buettner, A., Hippler, D., Matter, A., 2007. Holocene ITCZ and
Indian monsoon dynamics recorded in stalagmites from Oman and Yemen(Socotra). Quatern. Sci. Rev. 26, 170–188.
Grine, F.E., 1993. Description and preliminary analysis of new hominid craniodentalfossils from the Swartkrans formation. In: Brain, C.K. (Ed.), Swartkrans: A Cave’sChronicle of Early Man. Transvaal Museum, Pretoria, pp. 167–194.
Ivanovich, M., Harmon, R.S. (Eds.), 1992. Uranium Series Disequilibrium: Applica-tions to Earth, Marine and Environmental Sciences. Claredon, Oxford.
Karkanas, P., Bar-Yosef, O., Goldberg, P., Weiner, S., 2000. Diagenesis in prehistoriccaves: the use of minerals that form in situ to assess the completeness of thearchaeological record. J. Archaeol. Sci. 27, 915–929.
Latham, A.G., 1999. Cave breccias and archaeological sites. Capra 1.Lenoble, A., Bertran, P., 2004. Fabric of Paleolithic levels: methods and implications
for site formation processes. J. Archaeol. Sci. 31, 457–459.Pickering, R., Hancox, P.J., Lee-Thorp, J.A., Grun, R., Mortimer, G.E., McCulloch, M.,
Berger, L.R., 2007. Stratigraphy, U-series chronology and palaeoenvironments atGladysvale Cave: insights into the climatic control of South African homininbearing caves. J. Hum. Evol. 53, 602–619.
Pickering, T.R., Domınguez-Rodrigo, M., Egeland, C.P., Brain, C.K., 2004. New dataand ideas on the foraging behaviour of Early Stone Age hominids at SwartkransCave, South Africa. S. Afr. J. Sci. 100, 215–219.
Pickering, T.R., Domınguez-Rodrigo, M., Egeland, C.P., Brain, C.K., 2005. The contri-bution of limb bone fracture patterns to reconstructing early hominid behaviorat Swartkrans Cave (South Africa): archaeological application of a new analyt-ical method. Int. J. Osteoarchaeol. 15, 247–260.
Pickering, T.R., Domınguez-Rodrigo, M., Egeland, C.P., Brain, C.K., 2007. Carcassforaging by early hominids at Swartkrans Cave (South Africa): a new investiga-tion of the zooarchaeology and taphonomy of Member 3. In: Pickering, T.R.,Schick, K., Toth, N. (Eds.), Breathing Life into Fossils: Taphonomic Studies in Honorof C.K. (Bob) Brain. Stone Age Institute Press, Bloomington (IN), pp. 233–253.
Pickering, T.R., Egeland, C.P., Domınguez-Rodrigo, M., Brain, C.K., Schnell, A.G., 2008.Testing the ‘‘shift in the balance of power’’ hypothesis at Swartkrans, SouthAfrica: hominid cave use and subsistence behavior in the early Pleistocene. J.Anthropol. Archaeol. 27, 30–45.
Reynolds, S.C., Clarke, R.J., Kuman, K., 2007. The view from Lincoln Cave: mid- to latePleistocene fossil deposits from the Sterkfontein hominid site, South Africa. J.Hum. Evol. 53, 260–271.
Robinson, J.T., 1956. The Dentition of the Australopithecinae. Transvaal Museum,Pretoria.
de Ruiter, D., Brophy, J.K., Lewis, P.J., Churchill, S.E., Berger, L.R., 2008. Faunalassemblage composition and paleoenvironments of Plovers Lake, a MiddleStone Age locality in Gauteng Province, South Africa. J. Hum. Evol. 55, 1102–1117.
Schick, K.D., 1987. Stone Age Sites in the Making: Experiments in the Formation andTransformation of Archaeological Occurrences. British Archaeological Reports.International Series 319, Oxford.
Schwarcz, H.P., 1986. Geochronology and isotopic geochemistry of speleothems. In:Fritz, P., Fontes, J. (Eds.), Handbook of Environmental Isotope Geochemistry, vol.2. Elsevier, Amsterdam, pp. 271–303.
Teaford, M.F., 2006. What do we know and not know about diet and enamelstructure? In: Ungar, P. (Ed.), Early Hominin Diets: The Known, the Unknown,and the Unknowable. Oxford University, Oxford, pp. 56–76.
Vrba, E.S., 1985. Early hominids in southern Africa: updated observations onchronological and ecological background. In: Tobias, P.V. (Ed.), HominidEvolution: Past, Present and Future. Alan R. Liss, New York, pp. 195–200.
White, T.D., WoldeGabriel, G., Asfaw, B., Ambrose, S., Beyene, Y., Bernor, R.L.,Boisserie, J.-R., Currie, B., Gilbert, H., Haile-Selassie, Y., Hart, W.K., Hlusko, L.J.,Howell, F.C., Kono, R.T., Lehmann, T., Louchart, A., Lovejoy, C.O., Renne, P.R.,Saegusa, H., Vrba, E.S., Wesselman, H., Suwa, G., 2006. Asa Issie, Aramis and theOrigin of Australopithecus. Nature 440, 883–889.
Copyright © 2022 FDOKUMEN




















