![t] x| f} + of] hgf -cf=j](https://static.fdokumen.com/doc/165x107/631922cab41f9c8c6e099861/t-x-f-of-hgf-cfj.jpg)
New records of toxic algae Heterosigma cf. akashiwo and Gymnodinium cf. mikimotoi in the...
13
This article was downloaded by: [Dokuz Eylul University ] On: 05 May 2014, At: 01:28 Publisher: Taylor & Francis Informa Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK Israel Journal of Plant Sciences Publication details, including instructions for authors and subscription information: http://www.tandfonline.com/loi/tips20 New records of toxic algae Heterosigma cf. akashiwo and Gymnodinium cf. mikimotoi in the hypereutrophic Izmir Bay (Aegean Sea): Coupling between organisms and water quality parameters NIHAYET BIZSEL a & K. CAN BIZSEL a a Institute of Marine Science and Technology, Dokuz Eylul University, 1884-8 Sokak 10, Izmir, Turkey Published online: 14 Mar 2013. To cite this article: NIHAYET BIZSEL & K. CAN BIZSEL (2002) New records of toxic algae Heterosigma cf. akashiwo and Gymnodinium cf. mikimotoi in the hypereutrophic Izmir Bay (Aegean Sea): Coupling between organisms and water quality parameters, Israel Journal of Plant Sciences, 50:1, 33-44 To link to this article: http://dx.doi.org/10.1560/046H-XV7W-7BQQ-TCTX PLEASE SCROLL DOWN FOR ARTICLE Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) contained in the publications on our platform. However, Taylor & Francis, our agents, and our licensors make no representations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose of the Content. Any opinions and views expressed in this publication are the opinions and views of the authors, and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be relied upon and should be independently verified with primary sources of information. Taylor and Francis shall not be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and other liabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to or arising out of the use of the Content. This article may be used for research, teaching, and private study purposes. Any substantial or systematic reproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in any form to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http:// www.tandfonline.com/page/terms-and-conditions
Transcript of New records of toxic algae Heterosigma cf. akashiwo and Gymnodinium cf. mikimotoi in the...

This article was downloaded by: [Dokuz Eylul University ]On: 05 May 2014, At: 01:28Publisher: Taylor & FrancisInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: MortimerHouse, 37-41 Mortimer Street, London W1T 3JH, UK
Israel Journal of Plant SciencesPublication details, including instructions for authors and subscription information:http://www.tandfonline.com/loi/tips20
New records of toxic algae Heterosigma cf.akashiwo and Gymnodinium cf. mikimotoi in thehypereutrophic Izmir Bay (Aegean Sea): Couplingbetween organisms and water quality parametersNIHAYET BIZSEL a & K. CAN BIZSEL aa Institute of Marine Science and Technology, Dokuz Eylul University, 1884-8 Sokak 10,Izmir, TurkeyPublished online: 14 Mar 2013.
To cite this article: NIHAYET BIZSEL & K. CAN BIZSEL (2002) New records of toxic algae Heterosigma cf. akashiwo andGymnodinium cf. mikimotoi in the hypereutrophic Izmir Bay (Aegean Sea): Coupling between organisms and water qualityparameters, Israel Journal of Plant Sciences, 50:1, 33-44
To link to this article: http://dx.doi.org/10.1560/046H-XV7W-7BQQ-TCTX
PLEASE SCROLL DOWN FOR ARTICLE
Taylor & Francis makes every effort to ensure the accuracy of all the information (the “Content”) containedin the publications on our platform. However, Taylor & Francis, our agents, and our licensors make norepresentations or warranties whatsoever as to the accuracy, completeness, or suitability for any purpose ofthe Content. Any opinions and views expressed in this publication are the opinions and views of the authors,and are not the views of or endorsed by Taylor & Francis. The accuracy of the Content should not be reliedupon and should be independently verified with primary sources of information. Taylor and Francis shallnot be liable for any losses, actions, claims, proceedings, demands, costs, expenses, damages, and otherliabilities whatsoever or howsoever caused arising directly or indirectly in connection with, in relation to orarising out of the use of the Content.
This article may be used for research, teaching, and private study purposes. Any substantial or systematicreproduction, redistribution, reselling, loan, sub-licensing, systematic supply, or distribution in anyform to anyone is expressly forbidden. Terms & Conditions of access and use can be found at http://www.tandfonline.com/page/terms-and-conditions

© 2002 Laser Pages Publishing Ltd., Jerusalem
Israel Journal of Plant Sciences Vol. 50 2002 pp. 33–44
*Author to whom correspondence should be addressed. E-mail:[email protected]
New records of toxic algae Heterosigma cf. akashiwo and Gymnodiniumcf. mikimotoi in the hypereutrophic Izmir Bay (Aegean Sea):Coupling between organisms and water quality parameters
NIHAYET BIZSEL* AND K. CAN BIZSEL
Institute of Marine Science and Technology, Dokuz Eylul University, 1884-8 Sokak 10, Izmir, Turkey
(Received 3 April 2001 and in revised form 19 August 2001)
ABSTRACT
The red tide flagellates, Heterosigma cf. akashiwo Hada (Raphidophyceae) andGymnodinium cf. mikimotoi Miyake et Kominami (Dinophyceae), which have well-known ichthyotoxic effects, have been recorded for the first time in Izmir Bay,Turkey. They were particularly abundant in the hypereutrophic Inner Bay and theeutrophic Middle Bay in September and November 1998. In the Inner Bay, themaximum cell density of H. akashiwo was 222,300 cells/l and that of G. mikimotoiwas 15,900 cells/l on 2 September 1998. Water quality and meteorological condi-tions measured and/or recorded during this period are discussed in relation tophytoplankton proliferations. The reported environmental conditions that enhancethe growth of these two species (i.e., phosphate, nitrogen, and iron enrichments;temperature; and salinity) have been reconfirmed. Since there has been a monitoringprogram in Izmir Bay for more than 20 years, and both species are cyst-formingorganisms with rapid reproduction and mortality properties, it is likely that they haveoccurred much earlier than when this study was carried out. The cyst-forming abilityof these species suggests that their introduction into the bay might be via ballastwaters of long-distance cargo ships. Finally, the sediment-dredging activity beingcarried out at irregular intervals in the bay could be considered a potential mechani-cal factor that favors the growth and reproduction of these species.
INTRODUCTION
Among 5000 species of extant marine phytoplankton(Sournia et al., 1991), some 300 species can at timesoccur in such high numbers that they discolor the sur-face of the sea, resulting in so-called “red tides”, whileonly 40 or so species have the capacity to producepotent toxins that can find their way through fish andshellfish to humans (UNESCO, 1995). “Culturaleutrophication” from domestic, industrial, and agricul-tural wastes can stimulate harmful algal blooms. Forexample, the number of red tides (mainly Gymnodiniummikimotoi, Gonyaulax polygramma, Noctiluca scint-illans, and Prorocentrum minimum) in Hong KongHarbor in the period 1976 to 1986 shows a strikingrelationship with the 6-fold increase in human popula-
tion and the concurrent 2.5-fold increase in nutrientloading, contributed mainly by untreated domestic andindustrial wastes (UNESCO, 1995).
Toxins produced by certain phytoplankton, particu-larly dinoflagellates, are among the most potent poisonsknown to man. Unfortunately, there is no clear-cut cor-relation between algal concentrations and their poten-tially harmful effects. Dinoflagellate species such asDinophysis and Alexandrium can contaminate shellfishwith toxins, even at low concentrations (UNESCO,1995). Gymnodinium mikimotoi and Heterosigmaakashiwo are known as fish killers (Hoe Chang et al.,1993). When concentrations of Heterosigma exceed
Dow
nloa
ded
by [
Dok
uz E
ylul
Uni
vers
ity ]
at 0
1:28
05
May
201
4

Israel Journal of Plant Sciences 50 2002
34
several million per liter in the Strait of Georgia andadjacent waters, some fish species die. The largest fishspecies are more sensitive than smaller ones and somefish species are more sensitive than other species (HoeChang et al., 1993). Massive fish kills in Port PhillipBay, Melbourne in the early 1950s were associated withbrown water discolorations caused by a similarGymnodinium species. This species has been implicatedin massive kills of benthic invertebrates (lugworms, seaurchins, etc.) and of both wild and farmed fishes. Theprecise mechanisms causing fish death are still poorlyunderstood; both purely physical mechanisms such asincreased sea water viscosity due to secretion of algalmucilages, and chemical mechanisms such as the pro-duction of hemolytic substances, have been proposed(Hallegraeff, 1991).
In several Japanese studies, it was also found thatHeterosigma responded to increased micronutrients, par-ticularly iron (Taylor and Haigh, 1993). Heterosigmaakashiwo blooms require metals, such as iron and man-ganese, in addition to nitrogen, phosphorus, and vitaminB
12 (Honjo, 1993). Both nitrogen and phosphorus can
affect toxin content in all toxic marine dinoflagellates.The exact relationship is not clearly understood, how-
ever, but field evidence may afford some insights.Smayda (1989, cited in Tomas and Baden, 1993) notedthe changing nitrogen and phosphorus concentrationsover time in areas where toxic blooms have been morefrequently noted. Not only the absolute amounts ofthose nutrients, but also changing ratios of N:P, N:Si,and P:Si over time can favor increase in biomass as wellas influence species composition and succession. In ad-dition to these observations, phosphate-limiting condi-tions are generally noted (Tomas and Baden, 1993).
These two toxic organisms are reported in Izmir Bayfor the first time. The prevailing environmental condi-tions in the bay are also discussed.
STUDY AREA
Izmir Bay is one of the most polluted estuaries in theMediterranean Sea (Bizsel and Uslu, 2000). It can bedivided into three parts; Inner, Middle, and Outer IzmirBay (Fig. 1). In addition to untreated wastewaters origi-nating from the increasing population (exceeding 3million) and substantial industrial development, inten-sive harbor activities in the bay and agriculture in thesurrounding areas have added considerable pollution,
Fig. 1. Locations of sampling stations in Ιzmir Bay.
Dow
nloa
ded
by [
Dok
uz E
ylul
Uni
vers
ity ]
at 0
1:28
05
May
201
4

Bizsel and Bizsel / New records of toxic algae Heterosigma akashiwo and Gymnodinium mikimotoi in Izmir Bay
35
particularly to the Inner Izmir Bay. Therefore, signifi-cant phosphorus, nitrogen, and iron enrichments havebeen observed in the water of Inner Izmir Bay (Bizseland Uslu, 2000).
The average salinity of the surface layer in the bayvaries seasonally, between 37.02 psu (in January) and39.16 (in October). The average temperature of the sur-face has its minimum, 11.2 ºC, in January and its maxi-mum, 26.6 ºC, in July (Bizsel, 1996). Phytoplanktonbloom every spring, including harmful red tides that arecurrently spreading and increasing along the coast ofIzmir Bay. The first record of red tide in Izmir Bay andof fish death was recorded in the middle 1950s(Numann, 1955; Acara and Nalbandoglu, 1960). The fre-quency of red tides has increased for the last two decades,and is closely related to the progressively increasingeutrophication with a similar mechanism to that explainedby Honjo (1993) for Japanese waters. In Izmir Bay,Noctiluca scintillans has formed red tides very frequently,but also Prorocentrum micans, P. triestinum, Ceratiumsp., Nitzschia sp., Pseudo-nitzschia pungens group, Thal-assiosira sp., Eutreptiella sp., and Mesodinium rubrumare potential causative organisms (Koray et al., 1992).
MATERIALS AND METHODS
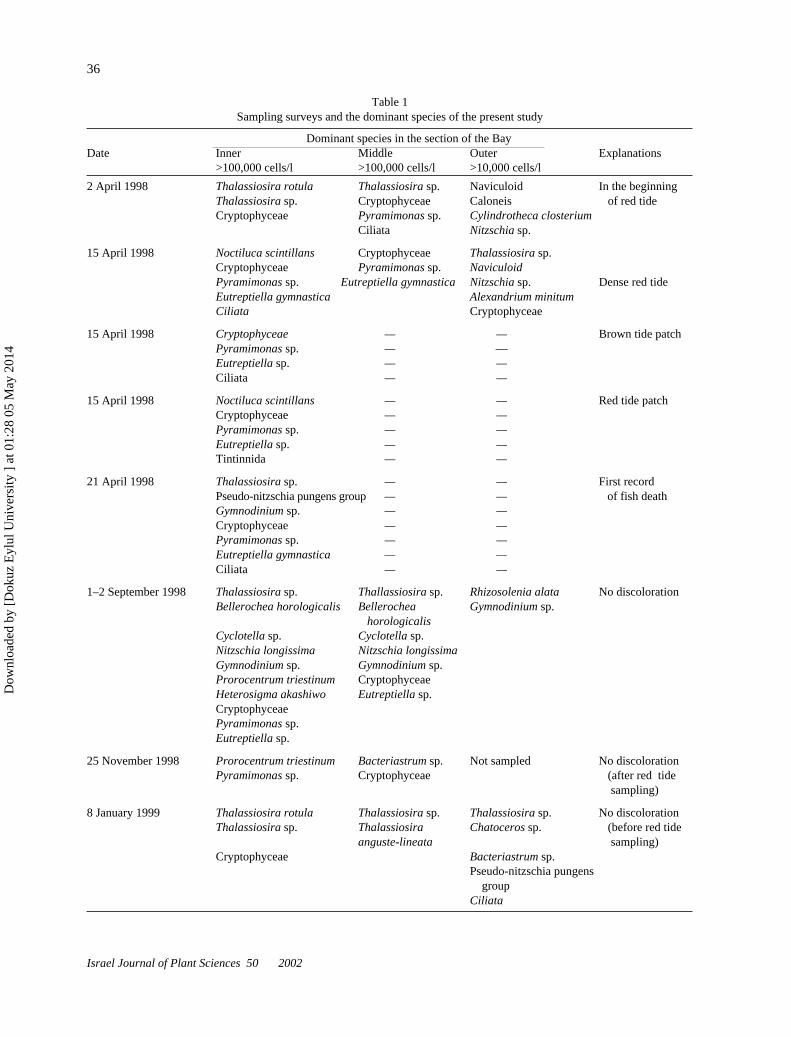
The samples were collected from ten stations (Fig. 1)selected by considering the hydrodynamic features of thebay (DEU-DBTE, 1999). The surveys were carried outin four phases. The first one was at the beginning ofApril (2.iv.1998) and during the red tide (15.iv.1998).The second one was during the early autumn bloom(2.ix.1998), while the third and fourth were in the lateautumn (25.xi.1998) and midwinter (8.i.1999). Thethird and fourth phases were carried out to represent theconditions prior to the beginning of the spring redtides. The survey periods and the dominant phyto-plankton species are shown in Table 1. Phytoplank-ton samples taken from surface water (0.3–0.5 m)were preserved with Lugol solution. Afterward, thesamples were transferred to sedimentation columns.When the suspended materials had sunk to the bot-tom of the column, the excess water above thesedimented material was removed carefully, concentrat-ing the volume of samples to 10 ml. Several drops fromeach sample were counted by using the single-droptechnique (Semina, 1978). Each sample was counted atleast five times.
Different sources were used to identify the phyto-plankton (Sournia, 1968; Drebes, 1974; Balech, 1988;Hallegraeff, 1991; Balech, 1995; Nezan, 1996;IFREMER, 1997; Paulmier, 1997; Tomas, 1997). Themicrophotographs of Heterosigma akashiwo and
Gymnodinium mikimotoi were taken with an Olympusinverted microscope in the Institut Français de recher-che pour l’exploitation de la mer (IFREMER) labora-tory Concarneau, France.
Oceanographic data were collected using a Sea BirdCTD probe which is an automatic device for real-timemeasurements of conductivity, temperature, and depth.The same device makes it possible to obtain density andsalinity via calculations based on the parameters mea-sured. pH was measured using a Hanna pH meter. Dis-solved oxygen (DO) was determined by conventionalWinkler titration. The samples for chlorophyll-a (Chl-a)measurement were concentrated on GF/F filters andextracted with 90% acetone solution. The fluorescenceintensity of clear extracts was then measured (Stricklandand Parsons, 1972) using a Turner Design fluorometer.The analyses of macronutrient samples were based onthe methods of Koroleff (1983). The samples taken fortotal dissolved phosphorus were filtered through aprebaked (450 ºC for 2 h) GF/F filter (Solorzano andSharp, 1980). The analysis method of Chelex-labilesoluble iron form as a micronutrient was given inTUBITAK (2001). Humic substances were measuredby using a PTI QM1 fluorescence spectrophotometer(Wedborg et al., 1994). For particulate organic C (POC)and particulate organic N (PON), the sample waterswere filtered first through a plankton net with 200-µmmesh and then through about 0.7-µm precombusted GF/Ffilters with volumes ranging between 150–1000 ml, de-pending on the clogging rate of the filter. Following acidtreatment, POC and PON were measured with a CarloErba Ea 1108 model CHN analyzer. All parameterstogether with the abundance (cell/L) values of the twonew recorded species were analyzed in order to deter-mine any significant correlation. For correlation analy-sis, Pearson product moment correlation was used.
RESULTS AND DISCUSSION
The two species, Heterosigma akashiwo and Gymno-dinium mikimotoi have been recorded in Izmir Bay forthe first time. Nevertheless, these species have started tobe reported frequently since 1990s from different geo-graphical locations around the world, particularly fromtemperate and subtropical embayments in Japan, Korea,Singapore, New Zealand, England, Belgium, eastern andwestern areas of North America, Bermuda, and Chile(Honjo, 1991, 1993; Koike et al., 1993; Konovalova,1993; Taylor and Haigh, 1993; Yoshinaga et al., 1995;Tangen, 1997; Kimura et al., 1999).
There are four possible explanations for the augmen-tation in frequency and abundance of algal bloomsaround the world, particularly for toxic species:
Dow
nloa
ded
by [
Dok
uz E
ylul
Uni
vers
ity ]
at 0
1:28
05
May
201
4
Israel Journal of Plant Sciences 50 2002
36
Table 1Sampling surveys and the dominant species of the present study
Dominant species in the section of the BayDate Inner Middle Outer Explanations
>100,000 cells/l >100,000 cells/l >10,000 cells/l
2 April 1998 Thalassiosira rotula Thalassiosira sp. Naviculoid In the beginningThalassiosira sp. Cryptophyceae Caloneis of red tideCryptophyceae Pyramimonas sp. Cylindrotheca closterium
Ciliata Nitzschia sp.
15 April 1998 Noctiluca scintillans Cryptophyceae Thalassiosira sp.Cryptophyceae Pyramimonas sp. NaviculoidPyramimonas sp. Eutreptiella gymnastica Nitzschia sp. Dense red tideEutreptiella gymnastica Alexandrium minitumCiliata Cryptophyceae
15 April 1998 Cryptophyceae — — Brown tide patchPyramimonas sp. — —Eutreptiella sp. — —Ciliata — —
15 April 1998 Noctiluca scintillans — — Red tide patchCryptophyceae — —Pyramimonas sp. — —Eutreptiella sp. — —Tintinnida — —
21 April 1998 Thalassiosira sp. — — First recordPseudo-nitzschia pungens group — — of fish deathGymnodinium sp. — —Cryptophyceae — —Pyramimonas sp. — —Eutreptiella gymnastica — —Ciliata — —
1–2 September 1998 Thalassiosira sp. Thallassiosira sp. Rhizosolenia alata No discolorationBellerochea horologicalis Bellerochea Gymnodinium sp.
horologicalisCyclotella sp. Cyclotella sp.Nitzschia longissima Nitzschia longissimaGymnodinium sp. Gymnodinium sp.Prorocentrum triestinum CryptophyceaeHeterosigma akashiwo Eutreptiella sp.CryptophyceaePyramimonas sp.Eutreptiella sp.
25 November 1998 Prorocentrum triestinum Bacteriastrum sp. Not sampled No discolorationPyramimonas sp. Cryptophyceae (after red tide
sampling)
8 January 1999 Thalassiosira rotula Thalassiosira sp. Thalassiosira sp. No discolorationThalassiosira sp. Thalassiosira Chatoceros sp. (before red tide
anguste-lineata sampling)Cryptophyceae Bacteriastrum sp.
Pseudo-nitzschia pungens groupCiliata
Dow
nloa
ded
by [
Dok
uz E
ylul
Uni
vers
ity ]
at 0
1:28
05
May
201
4
Bizsel and Bizsel / New records of toxic algae Heterosigma akashiwo and Gymnodinium mikimotoi in Izmir Bay
37
1. increased scientific awareness of toxic species;2. increased utilization of coastal waters for aquaculture;3. stimulation of plankton blooms by cultural eutrophi-
cation and/or unusual climatological conditions;4. transport of dinoflagellate resting cysts, either in
ships’ ballast water or associated with translocationof shellfish stocks from one area to another.
Species as new records
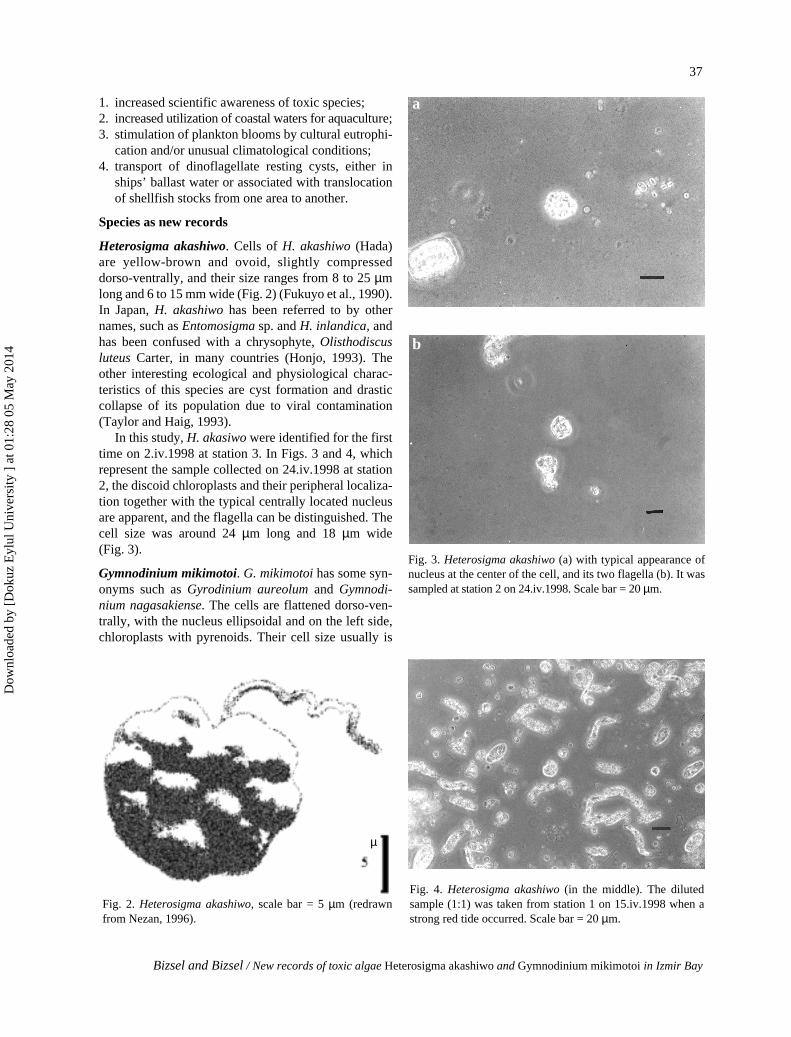
Heterosigma akashiwo. Cells of H. akashiwo (Hada)are yellow-brown and ovoid, slightly compresseddorso-ventrally, and their size ranges from 8 to 25 µmlong and 6 to 15 mm wide (Fig. 2) (Fukuyo et al., 1990).In Japan, H. akashiwo has been referred to by othernames, such as Entomosigma sp. and H. inlandica, andhas been confused with a chrysophyte, Olisthodiscusluteus Carter, in many countries (Honjo, 1993). Theother interesting ecological and physiological charac-teristics of this species are cyst formation and drasticcollapse of its population due to viral contamination(Taylor and Haig, 1993).
In this study, H. akasiwo were identified for the firsttime on 2.iv.1998 at station 3. In Figs. 3 and 4, whichrepresent the sample collected on 24.iv.1998 at station2, the discoid chloroplasts and their peripheral localiza-tion together with the typical centrally located nucleusare apparent, and the flagella can be distinguished. Thecell size was around 24 µm long and 18 µm wide(Fig. 3).
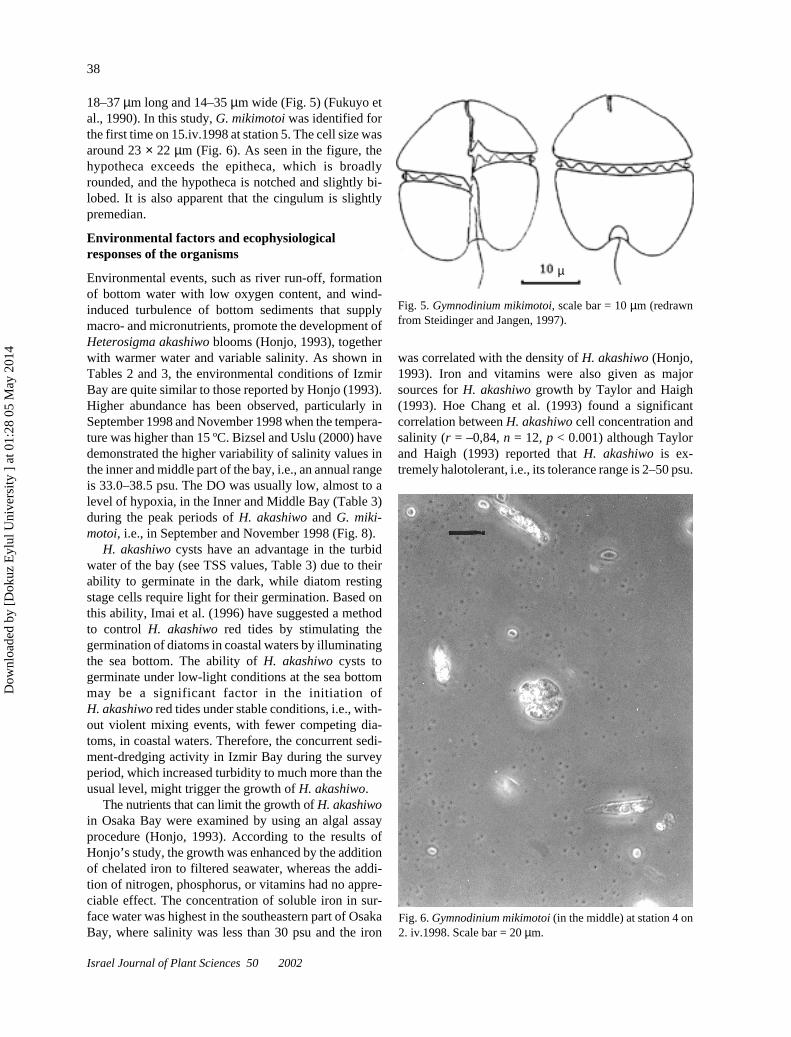
Gymnodinium mikimotoi. G. mikimotoi has some syn-onyms such as Gyrodinium aureolum and Gymnodi-nium nagasakiense. The cells are flattened dorso-ven-trally, with the nucleus ellipsoidal and on the left side,chloroplasts with pyrenoids. Their cell size usually is
Fig. 3. Heterosigma akashiwo (a) with typical appearance ofnucleus at the center of the cell, and its two flagella (b). It wassampled at station 2 on 24.iv.1998. Scale bar = 20 µm.
Fig. 2. Heterosigma akashiwo, scale bar = 5 µm (redrawnfrom Nezan, 1996).
Fig. 4. Heterosigma akashiwo (in the middle). The dilutedsample (1:1) was taken from station 1 on 15.iv.1998 when astrong red tide occurred. Scale bar = 20 µm.
a
b
µ
Dow
nloa
ded
by [
Dok
uz E
ylul
Uni
vers
ity ]
at 0
1:28
05
May
201
4
Israel Journal of Plant Sciences 50 2002
38
18–37 µm long and 14–35 µm wide (Fig. 5) (Fukuyo etal., 1990). In this study, G. mikimotoi was identified forthe first time on 15.iv.1998 at station 5. The cell size wasaround 23 × 22 µm (Fig. 6). As seen in the figure, thehypotheca exceeds the epitheca, which is broadlyrounded, and the hypotheca is notched and slightly bi-lobed. It is also apparent that the cingulum is slightlypremedian.
Environmental factors and ecophysiologicalresponses of the organisms
Environmental events, such as river run-off, formationof bottom water with low oxygen content, and wind-induced turbulence of bottom sediments that supplymacro- and micronutrients, promote the development ofHeterosigma akashiwo blooms (Honjo, 1993), togetherwith warmer water and variable salinity. As shown inTables 2 and 3, the environmental conditions of IzmirBay are quite similar to those reported by Honjo (1993).Higher abundance has been observed, particularly inSeptember 1998 and November 1998 when the tempera-ture was higher than 15 ºC. Bizsel and Uslu (2000) havedemonstrated the higher variability of salinity values inthe inner and middle part of the bay, i.e., an annual rangeis 33.0–38.5 psu. The DO was usually low, almost to alevel of hypoxia, in the Inner and Middle Bay (Table 3)during the peak periods of H. akashiwo and G. miki-motoi, i.e., in September and November 1998 (Fig. 8).
H. akashiwo cysts have an advantage in the turbidwater of the bay (see TSS values, Table 3) due to theirability to germinate in the dark, while diatom restingstage cells require light for their germination. Based onthis ability, Imai et al. (1996) have suggested a methodto control H. akashiwo red tides by stimulating thegermination of diatoms in coastal waters by illuminatingthe sea bottom. The ability of H. akashiwo cysts togerminate under low-light conditions at the sea bottommay be a significant factor in the initiation ofH. akashiwo red tides under stable conditions, i.e., with-out violent mixing events, with fewer competing dia-toms, in coastal waters. Therefore, the concurrent sedi-ment-dredging activity in Izmir Bay during the surveyperiod, which increased turbidity to much more than theusual level, might trigger the growth of H. akashiwo.
The nutrients that can limit the growth of H. akashiwoin Osaka Bay were examined by using an algal assayprocedure (Honjo, 1993). According to the results ofHonjo’s study, the growth was enhanced by the additionof chelated iron to filtered seawater, whereas the addi-tion of nitrogen, phosphorus, or vitamins had no appre-ciable effect. The concentration of soluble iron in sur-face water was highest in the southeastern part of OsakaBay, where salinity was less than 30 psu and the iron
Fig. 6. Gymnodinium mikimotoi (in the middle) at station 4 on2. iv.1998. Scale bar = 20 µm.
was correlated with the density of H. akashiwo (Honjo,1993). Iron and vitamins were also given as majorsources for H. akashiwo growth by Taylor and Haigh(1993). Hoe Chang et al. (1993) found a significantcorrelation between H. akashiwo cell concentration andsalinity (r = –0,84, n = 12, p < 0.001) although Taylorand Haigh (1993) reported that H. akashiwo is ex-tremely halotolerant, i.e., its tolerance range is 2–50 psu.
µ
Fig. 5. Gymnodinium mikimotoi, scale bar = 10 µm (redrawnfrom Steidinger and Jangen, 1997).
Dow
nloa
ded
by [
Dok
uz E
ylul
Uni
vers
ity ]
at 0
1:28
05
May
201
4
Bizsel and Bizsel / New records of toxic algae Heterosigma akashiwo and Gymnodinium mikimotoi in Izmir Bay
39
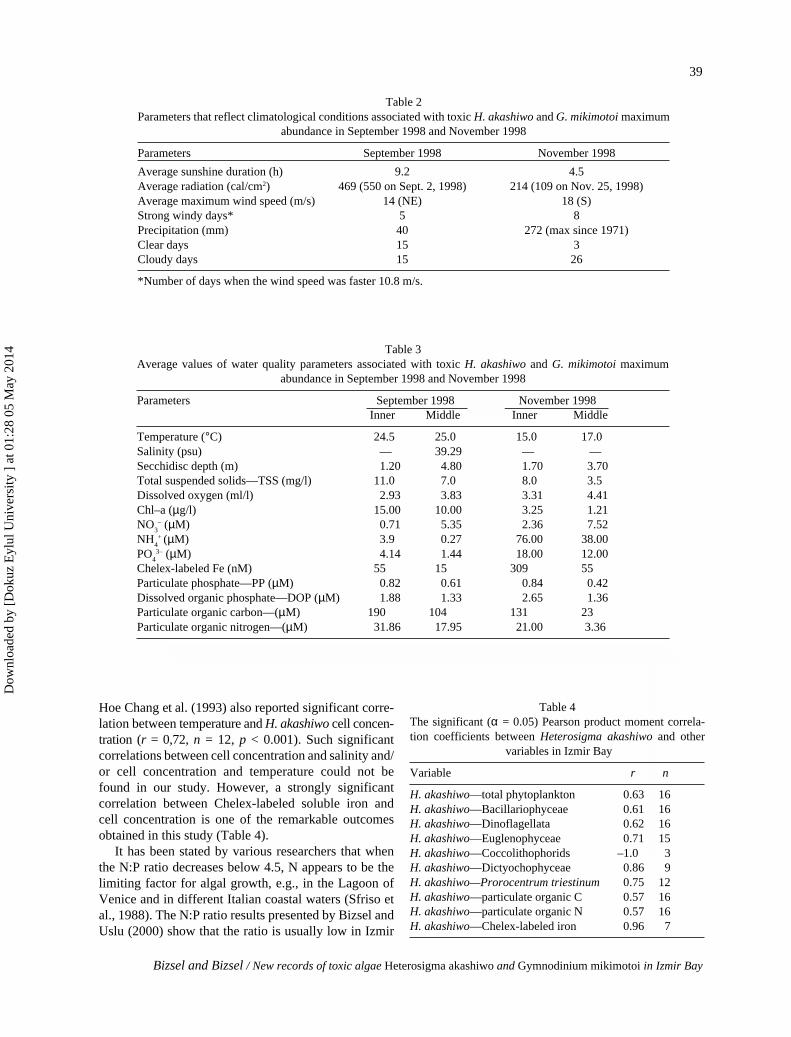
Hoe Chang et al. (1993) also reported significant corre-lation between temperature and H. akashiwo cell concen-tration (r = 0,72, n = 12, p < 0.001). Such significantcorrelations between cell concentration and salinity and/or cell concentration and temperature could not befound in our study. However, a strongly significantcorrelation between Chelex-labeled soluble iron andcell concentration is one of the remarkable outcomesobtained in this study (Table 4).
It has been stated by various researchers that whenthe N:P ratio decreases below 4.5, N appears to be thelimiting factor for algal growth, e.g., in the Lagoon ofVenice and in different Italian coastal waters (Sfriso etal., 1988). The N:P ratio results presented by Bizsel andUslu (2000) show that the ratio is usually low in Izmir
Table 2Parameters that reflect climatological conditions associated with toxic H. akashiwo and G. mikimotoi maximum
abundance in September 1998 and November 1998
Parameters September 1998 November 1998
Average sunshine duration (h) 9.2 4.5Average radiation (cal/cm2) 469 (550 on Sept. 2, 1998) 214 (109 on Nov. 25, 1998)Average maximum wind speed (m/s) 14 (NE) 18 (S)Strong windy days* 5 8Precipitation (mm) 40 272 (max since 1971)Clear days 15 3Cloudy days 15 26
*Number of days when the wind speed was faster 10.8 m/s.
Table 4The significant (α = 0.05) Pearson product moment correla-tion coefficients between Heterosigma akashiwo and other
variables in Izmir Bay
Variable r n
H. akashiwo—total phytoplankton 0.63 16H. akashiwo—Bacillariophyceae 0.61 16H. akashiwo—Dinoflagellata 0.62 16H. akashiwo—Euglenophyceae 0.71 15H. akashiwo—Coccolithophorids –1.0 3H. akashiwo—Dictyochophyceae 0.86 9H. akashiwo—Prorocentrum triestinum 0.75 12H. akashiwo—particulate organic C 0.57 16H. akashiwo—particulate organic N 0.57 16H. akashiwo—Chelex-labeled iron 0.96 7
Table 3Average values of water quality parameters associated with toxic H. akashiwo and G. mikimotoi maximum
abundance in September 1998 and November 1998
Parameters September 1998 November 1998Inner Middle Inner Middle
Temperature (°C) 24.5 25.0 15.0 17.0Salinity (psu) — 39.29 — —Secchidisc depth (m) 1.20 4.80 1.70 3.70Total suspended solids—TSS (mg/l) 11.0 7.0 8.0 3.5Dissolved oxygen (ml/l) 2.93 3.83 3.31 4.41Chl–a (µg/l) 15.00 10.00 3.25 1.21NO
3– (µM) 0.71 5.35 2.36 7.52
NH4+
(µM) 3.9 0.27 76.00 38.00
PO4
3– (µM) 4.14 1.44 18.00 12.00Chelex-labeled Fe (nM) 55 15 309 55Particulate phosphate—PP (µM) 0.82 0.61 0.84 0.42Dissolved organic phosphate—DOP (µM) 1.88 1.33 2.65 1.36Particulate organic carbon—(µM) 190 104 131 23Particulate organic nitrogen—(µM) 31.86 17.95 21.00 3.36
Dow
nloa
ded
by [
Dok
uz E
ylul
Uni
vers
ity ]
at 0
1:28
05
May
201
4
Israel Journal of Plant Sciences 50 2002
40
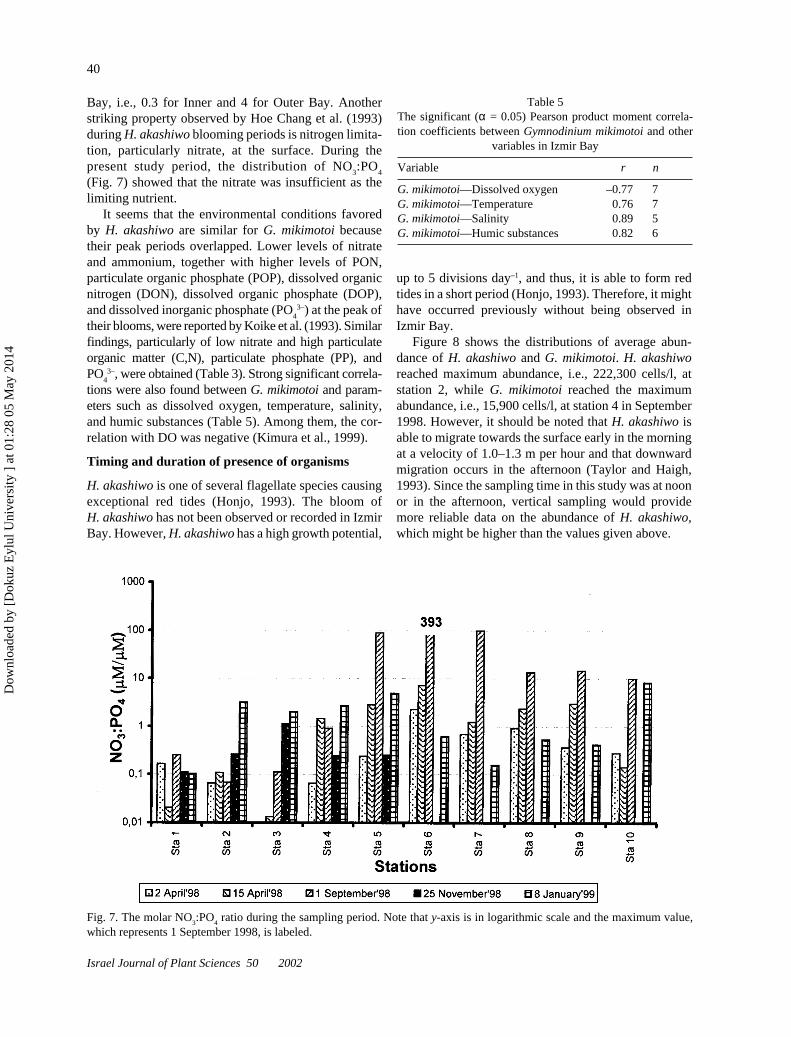
Bay, i.e., 0.3 for Inner and 4 for Outer Bay. Anotherstriking property observed by Hoe Chang et al. (1993)during H. akashiwo blooming periods is nitrogen limita-tion, particularly nitrate, at the surface. During thepresent study period, the distribution of NO
3:PO
4
(Fig. 7) showed that the nitrate was insufficient as thelimiting nutrient.
It seems that the environmental conditions favoredby H. akashiwo are similar for G. mikimotoi becausetheir peak periods overlapped. Lower levels of nitrateand ammonium, together with higher levels of PON,particulate organic phosphate (POP), dissolved organicnitrogen (DON), dissolved organic phosphate (DOP),and dissolved inorganic phosphate (PO
43–) at the peak of
their blooms, were reported by Koike et al. (1993). Similarfindings, particularly of low nitrate and high particulateorganic matter (C,N), particulate phosphate (PP), andPO
43–, were obtained (Table 3). Strong significant correla-
tions were also found between G. mikimotoi and param-eters such as dissolved oxygen, temperature, salinity,and humic substances (Table 5). Among them, the cor-relation with DO was negative (Kimura et al., 1999).
Timing and duration of presence of organisms
H. akashiwo is one of several flagellate species causingexceptional red tides (Honjo, 1993). The bloom ofH. akashiwo has not been observed or recorded in IzmirBay. However, H. akashiwo has a high growth potential,
up to 5 divisions day–1, and thus, it is able to form redtides in a short period (Honjo, 1993). Therefore, it mighthave occurred previously without being observed inIzmir Bay.
Figure 8 shows the distributions of average abun-dance of H. akashiwo and G. mikimotoi. H. akashiworeached maximum abundance, i.e., 222,300 cells/l, atstation 2, while G. mikimotoi reached the maximumabundance, i.e., 15,900 cells/l, at station 4 in September1998. However, it should be noted that H. akashiwo isable to migrate towards the surface early in the morningat a velocity of 1.0–1.3 m per hour and that downwardmigration occurs in the afternoon (Taylor and Haigh,1993). Since the sampling time in this study was at noonor in the afternoon, vertical sampling would providemore reliable data on the abundance of H. akashiwo,which might be higher than the values given above.
Table 5The significant (α = 0.05) Pearson product moment correla-tion coefficients between Gymnodinium mikimotoi and other
variables in Izmir Bay
Variable r n
G. mikimotoi—Dissolved oxygen –0.77 7G. mikimotoi—Temperature 0.76 7G. mikimotoi—Salinity 0.89 5G. mikimotoi—Humic substances 0.82 6
Fig. 7. The molar NO3:PO
4 ratio during the sampling period. Note that y-axis is in logarithmic scale and the maximum value,
which represents 1 September 1998, is labeled.
Dow
nloa
ded
by [
Dok
uz E
ylul
Uni
vers
ity ]
at 0
1:28
05
May
201
4

Bizsel and Bizsel / New records of toxic algae Heterosigma akashiwo and Gymnodinium mikimotoi in Izmir Bay
41
Table 6The profile of relatively abundant species in the phytoplanktonpopulation in September 2000 at Sta 2. Abundance values
are given as the mean values with their standard deviation
Species Abundance(×103 cells/l)
Eutreptiella sp. (Euglenaphyceae) 4.000 ± 600Nitzschia longissima (Diatom) 630 ± 350Cyclotella sp. (Diatom) 240 ± 175Thalassiosira sp. (Diatom) 190 ± 50Bellerochea horologicalis (Diatom) 125 ± 17Gymnodinium sp. (Dinoflagellata) 200 ± 100Prorocentrum triestinum (Dinoflagellata) 125 ± 60Pyramimonas sp. (Prasinophyceae ) 150 ± 80Cryptophyceae 130 ± 100Unidentified species 350 ± 135
Associated species
It is a well-known ecological observation that somespecies always co-exist with certain species and notwith others (Honjo, 1993). Honjo (1993) explained thatH. akashiwo, by excreting allelopathic substances, isable to suppress the growth of Skeletonema costatumwhile it enhances the growth of Prorocentrum triest-inum. In the present study, all the species which arerelatively abundant when higher quantities of H. akashiwoand G. mikimotoi were observed at station 2 in September1998 (Fig. 8) are listed in Table 6. Skeletonemacostatum was not recorded whereas Prorocentrumtriestinum was present with 125,000 cells/l. Thus, asignificant positive correlation was found between num-bers of H. akashiwo and P. triestinum (Table 4). How-ever, it should be noted that the actual dominant species
Fig. 8. Distribution of average abundances of Heterosigma akashiwo and Gymnodinium mikimotoi during the study.
Dow
nloa
ded
by [
Dok
uz E
ylul
Uni
vers
ity ]
at 0
1:28
05
May
201
4

Israel Journal of Plant Sciences 50 2002
42
was Eutreptiella sp. (Euglenophyceae) during the sameperiod and at the same station. Nevertheless, the previ-ous records on Skeletonema costatum in the bay (Bizseland Uslu, 2000; Gunciner, 2000; Bizsel et al., 2001)have shown that this species can usually be found inrelative abundance throughout the year, and sometimescan dominate in the phytoplankton, i.e., with 56 × 106
cells/l, comprising 93% all phytoplankton in 1994 atstation 1 (Bizsel and Uslu, 2000) and reported as thesecond dominant species during spring bloom in 1996by Gunciner (2000). In these previous records, there areno records on either H. akashiwo or G. mikimotoi. Con-sequently, Honjo’s (1993) explanation mentioned abovecan be relevant for the significant correlation observedbetween H. akashiwo and P. triestinum. It should alsobe noted that there are 6 toxic species other thanH. akashiwo and G. mikimotoi that are listed in Table 7.
CONCLUSION
1. It is likely that both species, H. akashiwo andG. mikimotoi, have been distributed in Izmir Bay forsome time.
2. As a field observation, the previous conclusionsreached by mainly Japanese scientists have been sup-ported. The environmental conditions which directlyfavor the growth of the two species are enhanced P,N, Fe loadings, water temperature, salinity, and thesediment dredging as a site-specific crucial factor forIzmir Bay.
3. Among the nutrients, iron is particularly importantfor H. akashiwo. An increased supply of chelex-labile iron from increased river runoff and dredgingactivity in the Inner Bay (Table 4) is likely to be aprevailing factor in the presence of H. akashiwo.
4. Positive responses of H. akashiwo and G. mikimotoi,at marginal values of N:P ratio, pH, DO, andtemperature, indicate their advantages in competing
with other species, as a sign of their opportunistcharacter.
5. A H. akashiwo bloom is a potential reason for sup-pressing S. costatum growth. Therefore, the mecha-nisms related to the marginal conditions mentionedabove deserve to be studied carefully in order tounderstand species-specific interactions between nu-trients and phytoplankton with special emphasize oniron.
6. Several studies that were conducted to clarify cystpopulation dynamics in order to predict the potentialof harmful algal bloom outbreaks, reached a com-mon conclusion that the distribution and abundanceof flagellate cysts in sediments are likely to be veryimportant in initiating harmful algal blooms. Al-though Izmir Bay has been monitored yearly formore than 20 years, there are so far no data on thecomposition and distribution of the dinoflagellatecysts in the sediment and water of the bay. Therefore,it is important to establish a program of continuingmonitoring surveys for investigating the cysts in or-der to be able to improve our knowledge on harmfulalgal blooms, and thereby, to prevent their toxiceffects in humans, as well as the mass mortalities inaquatic living resources.
ACKNOWLEDGMENTS
The Turkish Scientific and Technical Research Council(TUBITAK-YDABCAG-618G) supported this study.The authors would like to thank the Institute of MarineSciences and Technology of Dokuz Eylul University inIzmir for the generous support given for laboratoryfacilities, and the Provincial Directorate of Agricultureand Rural Affairs for the research vessel Yunus. We aregrateful to Elizabeth Nezan for guiding us in identifica-tions of phytoplankton species and to Patrick Lassus forhis helpful comments from IFREMER (France).
Table 7Informative data on the toxic and potentially toxic species: The abundance values in terms of maxima, means, and standard
deviations; sampling dates and sites; special remarks, if any
Species Sampling date Abundance Max Remarks[Mean ± SD](×103 cells/l)
Pseudo-nitzschia sp. 21.iv.1998 410 [95 ± 147] First day of fish mortality, at Sta 2Alexandrium minutum 21.iv.1998 410 [68 ± 141] First day of fish mortality, at Sta 2Dinophysis rotundata 24.iv.1998 9 [1.3 ± 3.4] Fish mortality continues at Sta 2Prorocentrum minimum 24.iv.1998 104 [26 ± 39] Fish mortality continues at Sta 2Dinophysis caudata 2.ix.1998 28 [10 ± 13] at Sta 3Gonyaulax grindleyi 2.ix.1998 3 [0.8 ± 0.3] at Sta 9
Dow
nloa
ded
by [
Dok
uz E
ylul
Uni
vers
ity ]
at 0
1:28
05
May
201
4

Bizsel and Bizsel / New records of toxic algae Heterosigma akashiwo and Gymnodinium mikimotoi in Izmir Bay
43
REFERENCES
Acara, A., Nalbandoglu, U. 1960. Preliminary report on thered tide outbreak in the Gulf of Izmir. Rapp. P.-V. Reun.Comm. Int. Exploit. Sci. Mer. Mediterr. 15: 33–38.
Balech, E. 1988. Los Dinoflagelados del AtlanticoSudoccidental. Publicaciones especiales del InstituttoEspanol de Oceanografia. No. 1. Madrid, Spain, 310 pp.
Balech, E. 1995. The genus Alexandrium Halim (Dino-flagellata). Sherkin Island Co. Cork, Sherkin Island, Ire-land, 115 pp.
Bizsel, N. 1996. Biogeochemical distribution of phosphorusfractions in Izmir Bay. Ph.D. thesis, Dokuz Eylal Univ.,Izmir, 188 pp.
Bizsel, N., Uslu, O. 2000. Phosphate, nitrogen and iron enrich-ments in the polluted Izmir Bay, Aegean Sea. Mar.Environ. Res. 49: 101–122.
Bizsel, N., Ozturk, M., Biszel, K.C., Benli, H.A. 2000. Ironforms and its role on the phytoplankton ecology in IzmirBay. TUBITAK Report, Ankara, 42 pp. (in Turkish).
Bizsel, N., Benli, H.A., Bizsel, K.C., Metin, G. 2001. A synop-tic study on phosphate and phytoplankton relationship inthe hypereutrophicated Izmir Bay (Aegean Sea). Turk. J.Eng. Environ. Sci. 25: 89–99.
DEU-DBTE, 1999. Izmir Korfezi 1994–1998. DenizArastirmalari Raporu, Izmir, 224 pp. (in Turkish).
Drebes, G. 1974. Marines phytoplankton. Georg ThiemeVerlag, Stuttgart, 186 pp.
Fukuyo, Y., Takano, H., Chihara, M., Matsuoka, K. 1990. Redtide organisms in Japan—an illustrated taxonomic guide.Uchida Rokakuho, Tokyo, 430 pp.
Gunciner, U. 2000. Red tide Doneminde Izmir Ic Korfezifitoplanktonunun Temporal Degisimleri. DEU DenizBilimleri ve Teknolojisi Enstitusu, Izmir, 138 pp.(in Turkish).
Hallegraeff, G.M. 1991. Aquaculturists’ guide to harmfulAustralian microalgae. CSIRO, Tasmania, 112 pp.
Hoe Chang, F., Pridmore, R., Boustead, N. 1993. Occurrenceand distribution of Heterosigma akashiwo (Raphid-ophyceae) in a 1989 bloom in Big Glory Bay, NewZealand. In: Smayda, T.J., Shimizu, Y., eds. Toxic phy-toplankton blooms in the sea. Fifth International Confer-ence on Toxic Marine Phytoplankton, Newport, RhodeIsland, USA, 28 October 1991. Newport, Rhode Island,USA, Elsevier, Amsterdam, Vol. 3, pp. 675–680.
Honjo, T. 1991. A relationship between winter water tempera-ture and timing of summer Gymnodinium nagasakiense redtides in Gokasyo Bay. Nippon Suisan Gakkaishi 57: 1679–1682.
Honjo, T. 1993. Overview on bloom dynamics and physi-ological ecology of Heterosigma akashiwo. In: Smayda,T.J., Shimizu, Y., eds. Toxic phytoplankton blooms in thesea. Fifth International Conference on Toxic Marine Phy-toplankton, Newport, Rhode Island, USA, 28 October1991. Elsevier, Amsterdam, Vol. 3, pp. 33–41.
Imai, I., Itakura, S., Yamaguchi, M., Honjo, T. 1996. Harmfuland toxic algal bloom. In: Yasumoto, T., Oshima, Y.,Fukuyo, Y. eds. Intergovernmental Oceanographic Com-
mission, UNESCO, Paris.IFREMER. 1997. Phytoquiz—1: Marine flagellates. A self-
training system for phytoplankton identification.IFREMER Service des Editions Scientifiques et Tech-niques. Web Page: http://www.ifremer.fr/delao/surveil-lance/phytoquiz.
Kimura, B., Kamizono, M., Takuya, E., Koizumi, Y.,Murakami, M., Honjo, T. 1999. Population development ofthe red tide dinoflagellate Gymnodinium mikimotoi in in-shore waters of Japan. Plankton Biol. Ecol. 46(1): 37–47.
Koike, Y., Nakaguchi, Y., Hiraki, K., Takeuchi, T., Kokubo,T., Ishimaru, T. 1993. Species and concentrations of sele-nium and nutrients in Tanabe Bay during Red Tide due toGymnodinium nagasakiense. J. Oceanogr. Vol. 49, pp.641–656.
Konovalova, G.V. 1993. Toxic and potentially toxic di-noflagellates from the Far East coastal waters of Russia. In:Smayda, T.J., Shimizu, Y., eds. Toxic phytoplanktonblooms in the sea. Fifth International Conference on ToxicMarine Phytoplankton, Newport, Rhode Island, USA, 28October 1991. Elsevier, Amsterdam, Vol. 3, pp. 275–279.
Koray, T., Buyukisik, B., Parlak, H., Gokpinar, S. 1992. IzmirKorfezi’nde deniz suyu kalitesini etkileyen tek hucreliorganizmalar: Red tide ve diger asırı ureme olayları. Doga,Turk. J. Biol. 16: 135–157 (in Turkish).
Koroleff, F. 1983. Determination of nutrients. In: Grassoff, K.,Ehrhardt, M., Kremling, K., eds. Methods of seawateranalysis. 2nd ed. Verlag Chemie, Berlin, pp. 134–136.
Nezan, E. 1996. Surveillance du phytoplacton marin: manuelillustré adapté à la formation des analystes. IFREMER,Concarneau, France.
Numann, W. 1955. Izmir Körfezi’nde “Balık kırılması”hadisesi. Hidrob. Mec., Ser. A 3: 90–93 (in Turkish).
Paulmier, P.G. 1997. Direction des ressources vivantes.Institut Français de recherche pour l’exploitation de la mer(IFREMER), Concarneau, France.
Semina, H.J. 1978. The size of cells. In: Sournia, A., ed. Phyto-plankton manual. UNESCO Monographs on Oceano-graphic Methodology 6. UNESCO, Paris, pp. 233–237.
Sfriso, A., Pavoni, B., Marcomini, A., Orio, A.A. 1988. An-nual variations of nutrients in the lagoon of Venice. Mar.Pollut. Bull. 19: 54–60.
Solorzano, L., Sharp, J.H. 1980. Determination of total dis-solved phosphorus and particulate phosphorus in naturalwaters. Limnol. Oceanogr. 25: 754–758.
Sournia, A. 1968. Diatomees planctonique du canal deMozambique et de I’lle Maurice. ORSTOM, Paris.
Sournia, A., Chretiennot-Dinet, M.J., Richard, M. 1991. Ma-rine phytoplankton: how many in the world ocean? J.Plankton Res. 13: 1093–1099.
Steidinger, K.A., Jangen, K. 1997. Dinoflagellates. In: Tomas,C.R., ed. Identifying marine phytoplankton. AcademicPress, San Diego, pp. 387–584.
Strickland, I.D.H., Parsons, T.R. 1972. A practical handbookof sea water analysis. Fish. Res. Board Can., p. 167.
Tangen, K. 1997. Blooms of Gyrodinium aureolum(Dinophyceae) in North European waters, accompanied bymortality in marine organisms. Sarsia 63: 123–133.
Dow
nloa
ded
by [
Dok
uz E
ylul
Uni
vers
ity ]
at 0
1:28
05
May
201
4

Israel Journal of Plant Sciences 50 2002
44
Taylor, F.J.R., Haigh, R. 1993. The ecology of fish killingblooms of the chloromonad flagellate Heterosigma in theStrait of Georgia and adjacent waters. In: Smayda, T.J.,Shimizu, Y., eds. Toxic phytoplankton blooms in the sea.Fifth International Conference on Toxic Marine Phy-toplankton, Newport, Rhode Island, USA, 28 October1991. Elsevier, Amsterdam, Vol. 3, pp. 705–710.
Tomas, C.R., Baden, D.G. 1993. The influence of phosphorussource on the growth and cellular toxin content of thebenthic Dinoflagellate Prorocentrum lima. In: Smayda,T.J., Shimizu, Y., eds. Toxic phytoplankton blooms in thesea. Fifth International Conference on Toxic Marine Phy-toplankton, Newport, Rhode Island, USA, 28 October1991. Elsevier, Amsterdam, Vol. 3, pp. 565–570.
Tomas, C.R. 1997. Identifying marine phytoplankton. Aca-
demic Press, San Diego, 858 pp.UNESCO. 1995. Manual on harmful marine microalgea. In:
Hallegraeff, G.M., Anderson, D.M., Cembella, A.D., eds.Intergovernmental Oceanographic Commission (IOC)Manuals and Guides No. 33. UNESCO, Paris.
Wedborg, M., Skoog, A., Fogelgvist, E. 1994. Organic carbonand humic substances in the Baltic Sea, The Kattegatt andthe Skagerrak. In: Senesi, N., Miano, T.M., eds. Humicsubstances in the global environment and implications onhuman health. Elsevier, Amsterdam, pp. 917–924.
Yoshinaga, I., Kawai, T., Ishida, Y. 1995. Lysis ofGymnodinium nagasakiense by marine bacteria. In:Lassus, P., Arzul, G., Erard-Le Denn, E., Gentien, P.,Marcaillou-Le Baut, C., eds. Harmful marine algal blooms.Lavoisier, Paris, pp. 687–692.
Dow
nloa
ded
by [
Dok
uz E
ylul
Uni
vers
ity ]
at 0
1:28
05
May
201
4

![AcVk Cf]V Z_ >RYRcRdYecR aRceZVd cV]ZVgVU](https://static.fdokumen.com/doc/165x107/6328451de491bcb36c0baf97/acvk-cfv-z-ryrcrdyecr-arcezvd-cvzvgvu.jpg)
![cf}}Bf]]lus ;DklQ a ''n]]l^g](https://static.fdokumen.com/doc/165x107/631a95471a1adcf65a0f21b9/cfbflus-dklq-a-nlg.jpg)




![#@cf}+jflif{s k|ltj]bg @)&%–)&^](https://static.fdokumen.com/doc/165x107/6331ac16ba79697da50fe7f4/cfjflifs-kltjbg-.jpg)

![tgx'+lhNnfsf]cf=j= 2073/074 sf]nflu :jLs[t](https://static.fdokumen.com/doc/165x107/6323d4d1117b4414ec0c7fc9/tgxlhnnfsfcfj-2073074-sfnflu-jlst.jpg)










