
The recent distribution of endemic, disjunct and globally ...

Folia Geobotanica 41: 433–451, 2006
NEW CHROMOSOME NUMBERS FOR PLANT TAXAENDEMIC TO THE BALEARIC ISLANDS
Mercedes Castro1)
& Josep A. Rosselló2*)
1) Facultad de Agronomia, Universidad Central de Venezuela, Apartado 4579, 2101 Maracay, Venezuela
2) Jardí Botànic, Universidad de Valencia, c/Quart 80, E-46008 Valencia, Spain; *) Corresponding author:fax +34 96 386 68 26, e-mail [email protected]
Abstract: Mitotic chromosome numbers are reported from 25 vascular plant taxa, endemic to the BalearicIslands that are poorly known cytogenetically. The chromosome numbers of Anthyllis vulneraria subsp.balearica (2n=12), Cymbalaria fragilis (2n=56), and Polygonum romanum subsp. balearicum (2n=40) weredetermined for the first time. A new chromosome number was found in several populations of Anthyllis hystrix(2n=70) suggesting that this species is decaploid, in contrast to an earlier work reporting a higher ploidy level(2n=12x=84). The new chromosome number 2n=32 was reported in Hypericum hircinum subsp. cambessedesii.It is suggested that the previous count (2n=40) could be explained by the presence of anomalous pentaploid cellsin some tissues, contrasting with the presence of a regular tetraploid complement (2n=32). Cytogeneticobservations suggest that Sibthorpia africana has a diploid chromosome complement of 2n=18, with 0–2accessory chromosomes. Accessory chromosomes are also reported for Phlomis italica, being the first record ofB chromosomes in this genus. Chromosomal instability was found in Galium crespianum and G. friedichiispecies, with three numbers 2n=44, 55 and 66. Two cytotypes differing in ploidy level were documented withinsingle plants. It is suggested that both species share a regular complement of 2n=44 and that the pasthybridization events and formation of regenerating roots from the typical rootstock of G. crespianum and G.friedrichii could be involved in the genesis of chromosome variants through partial endopolyploidy andconcomitant somatic segregation.
Keywords: Accessory chromosomes, Anthyllis, Cymbalaria, Galium, Hypericum, Polygonum
INTRODUCTION
The flora of the Balearic Islands, with about 100 non-apomictic species endemic to thearchipelago, is one of the most diverse of the Mediterranean basin. The origin andevolutionary relationships of this endemic flora has been debated since the phytogeographicapproach of KNOCHE (1922). The identification of the closest relatives of many Balearicendemic taxa is to a high degree uncertain because most of the available knowledge is usingintuitive comparisons based on morphological characters (KNOCHE 1922). Chromosomaldata have served as the basis for a classification of Mediterranean endemic taxa in fourcategories (paleoendemism, apoendemism, patroendemism, and schizoendemism), accordingto their age, taxonomic isolation, and closest relationships (FAVARGER &CONTANDRIOPOULOS 1961). This cytogenetic approach helps extend our knowledge on theendemic flora of the major Mediterranean islands (CARDONA & CONTANDRIOPOULOS 1977,1979). Despite the existence of an earlier cytogenetic review (CONTANDRIOPOULOS &CARDONA 1984), the current stage of karyological knowledge of the Balearic endemic flora isnot satisfactory. Chromosome counts on Balearic endemics are usually based on a very

limited number of observations, and some species lack basic chromosomal data. Recently, westarted a study aimed at evaluating the patterns of karyological evolution in the Balearicendemic flora (CASTRO & ROSSELLÓ 2005), assessing the basic karyological features ofpreviously studied taxa. In this work, we have assessed the mitotic chromosome numbers of25 vascular plant taxa endemic to the Balearic archipelago. With this contribution, nearly halfof the endemic flora of the Balearic archipelago have been revisited.
MATERIALS AND METHODS
Plant material
Living material (either whole plants or cuttings) was collected mostly from naturalpopulations throughout the Balearic Islands. Living plants were transferred and cultivated inpots at the Botanical Garden of Valencia University. From some accessions of Anthyllishystrix, A. vulneraria subsp. balearica, and Lysimachia minoricensis, seeds which wereobtained from the germplasm collection of the Botanical Garden of Sóller (Balearic Islands)were analyzed. Seeds of Launaea cervicornis were provided by M.A. Conesa and M. Mus(University of the Balearic Islands) from individuals collected in the field. Seeds weregerminated on solid agar in Petri dishes in a constant temperature of 20 °C and 12 hours ofwhite light daily. Voucher specimens are preserved at VAL (Table 1).
Chromosome preparation and karyotype analysis
Root tips were pre-treated with 0.002 M 8-hydroxyquinoline solution for 2 h at 4 °C and2 h at room temperature, washed with distilled water, fixed in fresh Carnoy I solution (glacialacetic acid: absolute ethanol; 3:1) overnight and stored in 70% ethanol at 4 °C until use. Forchromosome counts and karyotype determination, the root tips were hydrolyzed for 5–10 minin 1 M HCl at 60 °C, washed and stained in aceto-orcein for 4–6 h. Stained meristems weresquashed in a drop of 45% acetic acid and permanent preparations were made by mounting inCanada balsam. Photomicrographs of well-spread metaphases were taken with an OlympusCamedia C-2000-Z digital camera and processed with Adobe Photoshop 7.0. Chromosomecounts were made from 1–5 individuals per population, by direct observation and from thephotomicrographs of at least five well-spread metaphases per individual. Chromosomemeasurements were made on digital images using the processing image software ImageTool5.0. The idiograms of some taxa showing enough chromosomal resolution were obtainedfrom the chromosome measurements of five well-spread metaphase plates from severalplants. For each metaphase plate, the length of the short (S) and long (L) arms ofchromosomes, as well as the length of satellites, were expressed in relative values (haploidchromosome set = 100%). For centromere position, the nomenclature of LEVAN et al. (1964)was followed.
RESULTS AND DISCUSSION
A summary of the analyzed species and their chromosome numbers presented in this paperis shown in Table 1.
434 M. Castro & J.A. Rosselló

Alliaceae
Allium antoni-bolosii P. PALAU subsp. antoni-bolosiiMallorca, Artà, Talaia Morella (39°46�35� N, 3°21�1� E), 420 m, calcareous crevices, in
open and sunny places, 11.II.2004, M.A. CONESA (VAL 156738). 2n=30 (three individuals).No karyological knowledge was previously available for the Majorcan populations, but the
chromosome number agrees with previous counts from single accessions from Minorca andCabrera islands (GARBARI et al. 1991, ROSSELLÓ et al. 1993). The chromosomes showed awide range of size variation (6–15 µm) and the karyotype was composed of five pairs ofmetacentric, nine submetacentric and one subtelocentric pairs. Chromosome morphology andsize agree with the details given by ROSSELLÓ et al. (1993) for plants from other BalearicIslands, and no interpopulation variation was evident concerning gross karyotypemorphology. No satellite was observed in the individuals studied.
Allium antoni-bolosii P. PALAU subsp. eivissanum (GARBARI et MICELI) N. TORRES etROSSELLÓ
Ibiza, Sant Antoni de Portmany, Es Port des Torrent (38°57�43� N, 1°15�25� E), 25 m,clearing patches with thin soil in low calcareous scrub, 17.XI.2004, N. TORRES & J.A.ROSSELLÓ (VAL 156739). 2n=30 (five individuals).
Ibiza, Sant Josep de sa Talaia, Ses Salines (38°50�47� N, 1°22�30� E), 1 m, basic soil onshaded slopes, 18.XI.2004, N. TORRES & J.A. ROSSELLÓ (VAL 156740). 2n=30 (fiveindividuals).
The two accessions sampled showed the same chromosome number as that reported forplants collected at the type locality (MICELI & GARBARI 1987). The chromosomecomplement is similar in morphology and size to the typical subspecies, suggesting that nomajor chromosomal rearrangements have occurred within the species. Western (subsp.eivissanum; Ibiza and Formentera islands) and eastern (subsp. antoni-bolosii, Mallorca,Minorca, and Cabrera islands) populations are very closely related concerning theirmorphology, leaf anatomy and karyology (ROSSELLÓ et al. 1993). On the basis of the basechromosome numbers reported in the genus (x=7, 8), the Balearic populations are polyploid(MATHEW 1996). A. antoni-bolosii belongs to Allium sect. Scorodon KOCH and is closelyrelated to the A. cupani RAFIN.-A. hirtovaginatum KUNTH complex (MICELI & GARBARI
1987, GARBARI et al. 1979). Interestingly, only chromosome numbers 2n=16 and 2n=14 havebeen reported for the diploid members of the complex, A. cupani and A. hirtovaginatum,respectively (GARBARI et al. 1979). Thus, the hypothesis of an allopolyploid origin ofA. antoni-bolosii from x=7 and x=8 ancestors (now absent from the Balearic Islands) shouldnot be discarded, although other explanations (autotetraploid or allopolyploid origin throughx=8 ancestors and later fusion of a pair of chromosomes) are also plausible. Given thatconcerted evolution of ribosomal multigene families is not the dominating process in Allium(MES et al. 1999), the search for divergent ITS paralogues (or chimeric sequences) inA. antoni-bolosii could be a reliable molecular approach to assess its origin.
Chromosome numbers in Balearic endemics 435

436 M. Castro & J.A. RossellóT
able
1.A
sum
mar
yof
the
anal
yzed
spec
ies
and
acce
ssio
nsan
dth
eirc
hrom
osom
enu
mbe
rs.A
nas
teri
skde
note
sth
atth
eta
xon
isno
tend
emic
toth
eB
alea
ric
Isla
nds.
Tax
onC
hrom
.num
ber
Acc
essi
onV
ouch
er
All
ium
anto
ni-
bolo
sii
P.P
AL
AU
subs
p.anto
ni-
bolo
sii
2n=
30M
allo
rca,
Art
a,T
alai
aM
orel
laV
AL
1567
38A
lliu
manto
ni-
bolo
sii
P.P
AL
AU
subs
p.ei
viss
anum
(GA
RB
AR
Iet
MIC
EL
I)N
.TO
RR
ES
etR
OS
SE
LL
Ó2n
=30
Ibiz
a,S
antA
nton
ide
Por
tman
y,E
sP
ortd
esT
orre
ntV
AL
1567
39Ib
iza,
San
tJos
epde
saT
alai
a,S
esS
alin
esV
AL
1567
40A
nth
ylli
shys
trix
(WIL
LK
.ex
BA
RC
EL
Ó)
CA
RD
ON
A,C
ON
TA
ND
R.
etS
IER
RA
2n=
70M
inor
ca,F
erre
ries
,Bar
ranc
d’A
lgen
dar
VA
L15
6752
Min
orca
,Mer
cada
l,C
apde
Cav
alle
ria
VA
L15
6753
Min
orca
,Fer
reri
es,E
tsA
locs
VA
L15
6754
Min
orca
,Ciu
tade
lla
deM
enor
ca,M
arin
ade
Cur
niol
aV
AL
1567
55A
nth
ylli
svu
lner
ari
aL
.sub
sp.b
ale
ari
ca(C
OS
S.e
xM
AR
ÈS
etV
IGIN
.)O
.BO
LÒ
Set
VIG
O2n
=12
Mal
lorc
a,S
ólle
r,S
erra
d’A
lfàb
iaV
AL
1567
56M
allo
rca,
Sól
ler,
Ser
rade
Son
Tor
rell
aV
AL
1567
57M
allo
rca,
Pui
gM
ajor
,Sa
Com
ade
n’A
rbon
aV
AL
1567
58A
.vu
lner
ari
aL
.sub
sp.g
andoger
i(S
AG
OR
SK
I)W
.BE
CK
ER
*2n
=12
Mal
lorc
a,A
rtà,
Col
ònia
deS
antP
ere
VA
L15
6840
Are
nari
abolo
sii
(CA
NIG
.)L
.SÁ
EZ
etR
OS
SE
LL
Ó2n
=44
Mal
lorc
a,E
scor
ca,P
uig
deM
assa
nell
aV
AL
1567
50A
rist
olo
chia
bia
nori
iS
EN
NE
Net
PA
U2n
=12
Mal
lorc
a,P
olle
nça,
Col
lde
laC
reue
taV
AL
1484
71A
stra
galu
sbale
ari
cus
CH
AT
ER
2n=
16M
allo
rca,
Pol
lens
a,M
orti
tx,P
uig
deS
esP
arad
esV
AL
1567
59C
abre
raV
AL
1567
60B
iscu
tell
aeb
usi
tana
RO
SS
EL
LÓ
,N.T
OR
RE
Set
L.S
ÁE
Z2n
=18
Ibiz
a,S
antM
ique
lde
Bal
ansa
t,E
sP
enya
lde
s’A
guil
aV
AL
1484
72Ib
iza,
San
tMat
eud’
Alb
arca
,Cal
aA
lbar
caV
AL
1567
49C
rocu
sca
mbes
sedes
siJ.
GA
Y2n
=16
Mal
lorc
a,P
olle
nça,
Col
lde
laC
reue
taV
AL
1484
97M
allo
rca,
Esc
orca
,Pui
gM
ajor
VA
L15
6763
Min
orca
,Ciu
tade
lla,
Cal
aM
orel
lV
AL
1567
64M
inor
ca,M
aó,C
apF
avar
itx
VA
L15
6765
Min
orca
,Fer
reri
es,E
lPil
arV
AL
1567
82C
ymbala
ria
fragil
is(J
.J.R
OD
R.)
A.C
HE
VA
L.
2n=
56M
inor
ca,F
erre
ries
,Bar
ranc
d’A
lgen
dar
VA
L15
6778
Euphorb
iafo
ntq
uer
iana
GR
EU
TE
R2n
=20
Mal
lorc
a,E
scor
ca,P
uig
deM
assa
nell
aV
AL
1567
51G
ali
um
cres
pia
num
J.J.
RO
DR
.2n
=55
Mal
lorc
a,E
scor
ca,P
uig
Maj
orV
AL
1484
82M
allo
rca,
Pol
lenç
a,C
olld
eS
aC
reue
taV
AL
1484
832n
=55
,66
Mal
lorc
a,E
scor
ca,P
uig
deM
assa
nell
aV
AL
1567
74M
allo
rca,
Cai
mar
i,S
altd
ela
Bel
laD
ona
VA
L15
6775
2n=
66M
allo
rca,
Pol
lens
a,M
orti
txV
AL
1567
76M
allo
rca,
Sól
ler,
Ser
rad’
Alf
àbia
VA
L15
6777
Gali
um
frie
dri
chii
N.T
OR
RE
S,L
.SÁ
EZ
,MU
Set
RO
SS
EL
LÓ
2n=
44Ib
iza,
San
tJos
epde
saT
alai
a,S
esR
oque
sA
ltes
VA
L15
6728
2n=
66Ib
iza,
San
tJoa
nde
Lab
ritj
a,E
lsC
ingl
esN
egre
sV
AL
1567
27

Chromosome numbers in Balearic endemics 4372n
=55
,66
Ibiz
a,S
antA
nton
ide
Por
tman
y,E
lsA
lls
VA
L15
6729
For
men
tera
,ElP
ilar
,La
Mol
aV
AL
1484
84G
lobula
ria
majo
rice
nsi
sG
AN
D.
2n=
32M
allo
rca,
Esc
orca
,sal
tde
laB
ella
Don
aV
AL
1567
62H
yper
icum
hir
cinum
L.s
ubsp
.cam
bes
sedes
ii(C
OS
S.e
xB
AR
CE
LÓ
)S
AU
VA
GE
2n=
32M
allo
rca,
Esc
orca
,Gor
gB
lau
VA
L15
6725
Launea
cerv
icorn
is(B
OIS
S.)
FO
NT
QU
ER
etR
OT
HM
.2n
=18
Mal
lorc
a,F
elan
itx,
Cal
aS
aN
auV
AL
1567
41M
allo
rca,
Cap
depe
ra,E
sC
arre
gado
rV
AL
1567
42M
allo
rca,
Art
à,C
ala
Mat
zocs
VA
L15
6743
Mal
lorc
a,S
anta
Mar
gali
da,S
onB
auló
VA
L15
6744
Mal
lorc
a,M
anac
or,P
orto
Cri
sto
VA
L15
6745
Min
orca
,Es
Mer
cada
l,T
iran
tV
AL
1567
46M
inor
ca,E
sM
erca
dal,
Bin
imel
.làV
AL
1567
47M
inor
ca,M
aó,B
inid
alí
VA
L15
6748
Lotu
ste
traphyl
lus
L.f
.2n
=14
Mal
lorc
a,E
scor
ca,P
uig
deM
assa
nell
aV
AL
1567
61M
inor
ca,F
erre
ries
,Bar
ranc
d’A
lgen
dar
VA
L15
6800
Min
orca
,Fer
reri
es,C
ala
Gal
dana
VA
L15
6732
Lys
imach
iam
inori
censi
sJ.
J.R
OD
R.
2n=
24B
otan
ical
Gar
den
ofS
ólle
rV
AL
1567
26P
hlo
mis
itali
caL
.2n
=20
Mal
lorc
a,C
alvi
à,S
’Esc
lop
VA
L15
6768
Mal
lorc
a,P
olle
nsa,
Mor
titx
VA
L15
6769
2n=
20+
0-1B
Mal
lorc
a,E
scor
ca,P
uig
deM
assa
nell
aV
AL
1567
66M
allo
rca,
Esc
orca
,Pui
gM
ajor
VA
L15
6767
Min
orca
,Fer
reri
es,S
antA
nton
ide
Rum
aV
AL
1567
70P
oly
gonum
rom
anum
J AC
Q.s
ubsp
.bale
ari
cum
RA
FF
AE
LL
Iet
L.
VIL
LA
R2n
=40
Min
orca
,Ciu
tade
lla,
wes
tfac
eof
Cal
aM
orel
lV
AL
1567
23M
inor
ca,C
iuta
dell
a,ea
stfa
ceof
Cal
aM
orel
lV
AL
1567
71P
rim
ula
aca
uli
s(L
.)L
.sub
sp.b
ale
ari
ca(W
ILL
K.)
GR
EU
TE
Ret
BU
RD
ET
2n=
22M
allo
rca,
Pui
gde
Mas
sane
lla
VA
L15
6772
Mal
lorc
a,E
scor
ca,P
uig
Maj
orV
AL
1567
73R
ubia
bale
ari
ca(W
ILL
K.)
PO
RT
Asu
bsp.
bale
ari
ca2n
=66
Mal
lorc
a,F
orna
lutx
,Bal
itx
d’en
Mig
VA
L14
8493
Mal
lorc
a,C
aim
ari,
Sal
tde
laB
ella
Don
aV
AL
1504
75Sib
thorp
iaafr
icana
L.
2n=
18+
0-1B
Mal
lorc
a,E
scor
ca,P
uig
Maj
orV
AL
1484
97M
allo
rca,
Pol
lenç
a,C
olld
eS
aC
reue
taV
AL
1484
98M
allo
rca,
For
nalu
tx,B
alit
xd’
En
Mig
VA
L14
8499
Min
orca
,Fer
reri
es,n
ear
Es
Pas
des
Rev
ull
VA
L15
0477
Ibiz
a,S
antM
ateu
d’A
lbar
ca,C
ala
Alb
arca
VA
L15
6779
2n=
18+
0-2B
Mal
lorc
a,E
scor
ca,P
uig
deM
assa
nell
aV
AL
1504
76V
iola
jauber
tiana
MA
RÈ
Set
VIG
IN.
2n=
20M
allo
rca,
Pol
lens
a,M
orti
tx,P
uig
deS
esP
arad
esV
AL
1567
81V
iola
odora
taL
.sub
sp.s
tolo
nif
era
(J.J
.RO
DR
.)O
RE
LL
etR
OM
O2n
=20
Min
orca
,Fer
reri
es,B
arra
ncd’
Alg
enda
rV
AL
1567
80

Aristolochiaceae
Aristolochia bianorii SENNEN et PAU
Mallorca, Pollença, Coll de la Creueta (39°56�20� N, 3°16�19� E), 50 m, maritime slopes,15.II.2004, M.A. CONESA, M. MUS & J.A. ROSSELLÓ (VAL 148471). 2n=12 (twoindividuals).
The chromosomes of this species are small (1–1.5 µm) and apparently similar inmorphology; no further karyological details could be observed with the method used. Thesame chromosome count was reported by CARDONA (1976) from a single population fromMallorca.
Asteraceae
Launea cervicornis (BOISS.) FONT QUER et ROTHM.Mallorca, Felanitx, Cala Sa Nau (38°40�8� N, 1°34�28� E), 1 m, coastal slopes, s.dat., M.A.
CONESA (VAL 156741). 2n=18 (two individuals).Mallorca, Capdepera, Es Carregador (39°41�9� N, 3°27�17� E), 1 m, coastal slopes, s.dat.,
M.A. CONESA (VAL 156742). 2n=18 (one individual).Mallorca, Arta, Cala Matzocs (39°45�29� N, 3°23�48� E), 1 m, coastal slopes, s.dat., M.A.
CONESA (VAL 156743). 2n=18 (two individuals).Mallorca, Santa Margalida, Son Bauló (39°45�31� N, 3°9�48� E), 5 m, calcareous cliffs
near the sea, s.dat., M.A. CONESA (VAL 156744). 2n=18 (three individuals).Mallorca, Manacor, Porto Cristo (39°31�59� N, 3°19�32� E), 5 m, coastal slopes, s.dat., M.
MUS (VAL 156745). 2n=18 (three individuals).Minorca, Es Mercadal, Tirant (40°5�13� N, 4°6�9� E), 1 m, coastal slopes, s.dat., M.A.
CONESA (VAL 156746). 2n=18 (three individuals).Minorca, Es Mercadal, Binimel.là (40°5�47� N, 4°2�38� E), 1 m, coastal slopes, s.dat.,
M.A. CONESA (VAL 156747). 2n=18 (three individuals).Minorca, Maó, Binidalí (39°49�29� N, 4°12�12� E), 5 m, coastal slopes, s.dat., M.A.
CONESA (VAL 156748). 2n=18 (four individuals).Chromosomes are 4–7 µm long, predominantly submetacentric and subtelocentric, with
some metacentric pairs. No secondary constrictions were observed in the individuals studied.The same chromosome number was previously reported for two accessions from Minorca(DAHLGREN et al. 1971, CARDONA 1977).
Brassicaceae
Biscutella ebusitana ROSSELLÓ, N. TORRES et L. SÁEZ
Ibiza, Sant Miquel de Balansat, Es Penyal de s’Aguila (39°3�44� N, 1°20�49� E), maritimecliffs, 50 m, 26.V.2002, M.A. CONESA, M. MUS, J.A. ROSSELLÓ & N. TORRES (VAL148472). 2n=18 (two individuals).
Ibiza, Sant Mateu d’Albarca, Cala Albarca (39°3�12� N, 1°21�32� E), 100 m, maritimecliffs, 17.XI.2004, N. TORRES & J.A. ROSSELLÓ (VAL 156749). 2n=18 (one individual).
The karyotype is composed of small chromosomes (1.5–2 µm), showing no visiblecentromeric constrictions. The species was already karyologically analyzed, but only a singleplant was available for study (ROSSELLÓ et al. 1999). Additional plants from the type locality
438 M. Castro & J.A. Rosselló
(Es Penyal de s’Aguila) and from another population showed the same chromosome number,confirming the diploid level for Balearic populations.
Caryophyllaceae
Arenaria bolosii (CANIG.) L. SÁEZ et ROSSELLÓ
Mallorca, Escorca, Puig de Massanella (39°48�13� N, 2°50�53� E), 1250 m, sunny slopes,8.II.2004, M.A. CONESA, M. MUS & J.A. ROSSELLÓ (VAL 156750). 2n=44 (one individual).
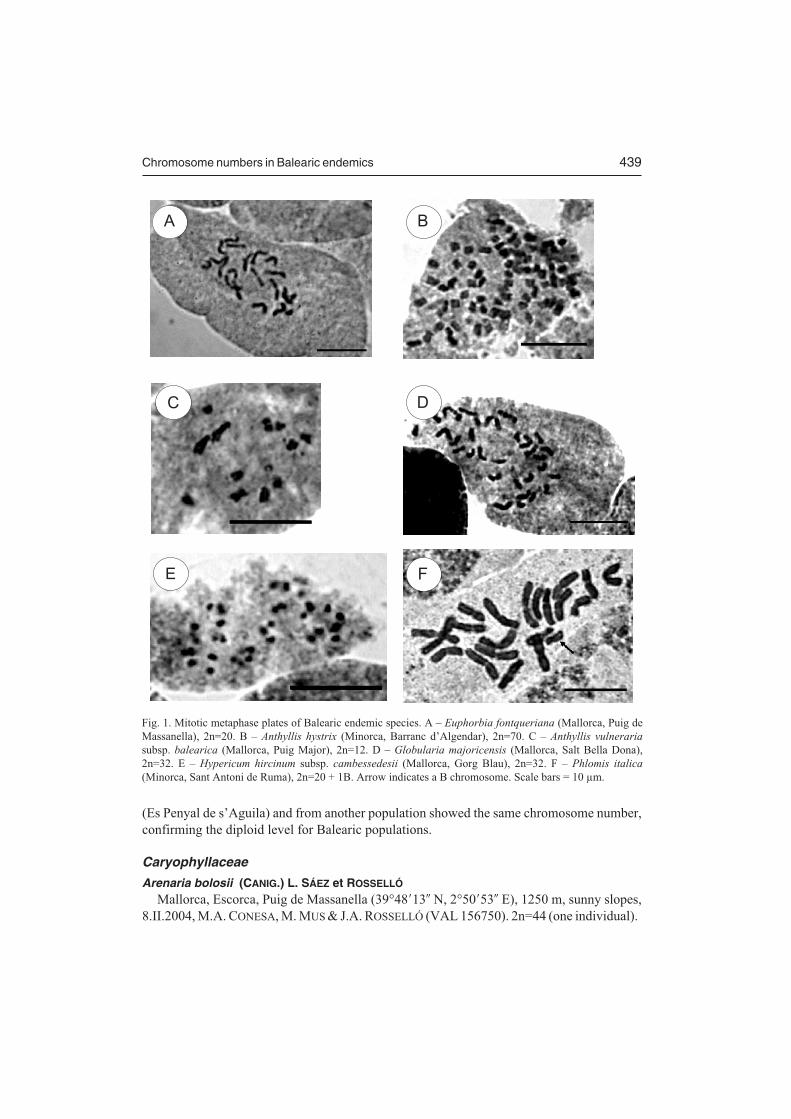
Chromosome numbers in Balearic endemics 439
B
C
E
D
F
A
Fig. 1. Mitotic metaphase plates of Balearic endemic species. A – Euphorbia fontqueriana (Mallorca, Puig deMassanella), 2n=20. B – Anthyllis hystrix (Minorca, Barranc d’Algendar), 2n=70. C – Anthyllis vulneraria
subsp. balearica (Mallorca, Puig Major), 2n=12. D – Globularia majoricensis (Mallorca, Salt Bella Dona),2n=32. E – Hypericum hircinum subsp. cambessedesii (Mallorca, Gorg Blau), 2n=32. F – Phlomis italica
(Minorca, Sant Antoni de Ruma), 2n=20 + 1B. Arrow indicates a B chromosome. Scale bars = 10 µm.
The chromosome complement is composed mainly of small metacentric chromosomes(1–2.5 µm). CARDONA & CONTANDRIOPOULOS (1983) reported the same chromosomenumber for plants growing at the type locality (Puig de Massanella), but the voucher specimencould not be traced and verified. At this site, the related A. grandiflora subsp. glabrescens(WILLK.) G. LÓPEZ et NIETO FEL. also occurs, and intermediate plants of putative hybridorigin can be found. To date, only tetraploids plants of the A. grandiflora L. complex havebeen detected in the Balearic Islands (A. grandiflora subsp. glabrescens, 2n=44; CASTRO &ROSSELLÓ 2005).
440 M. Castro & J.A. Rosselló
A
C
E
B
D
F
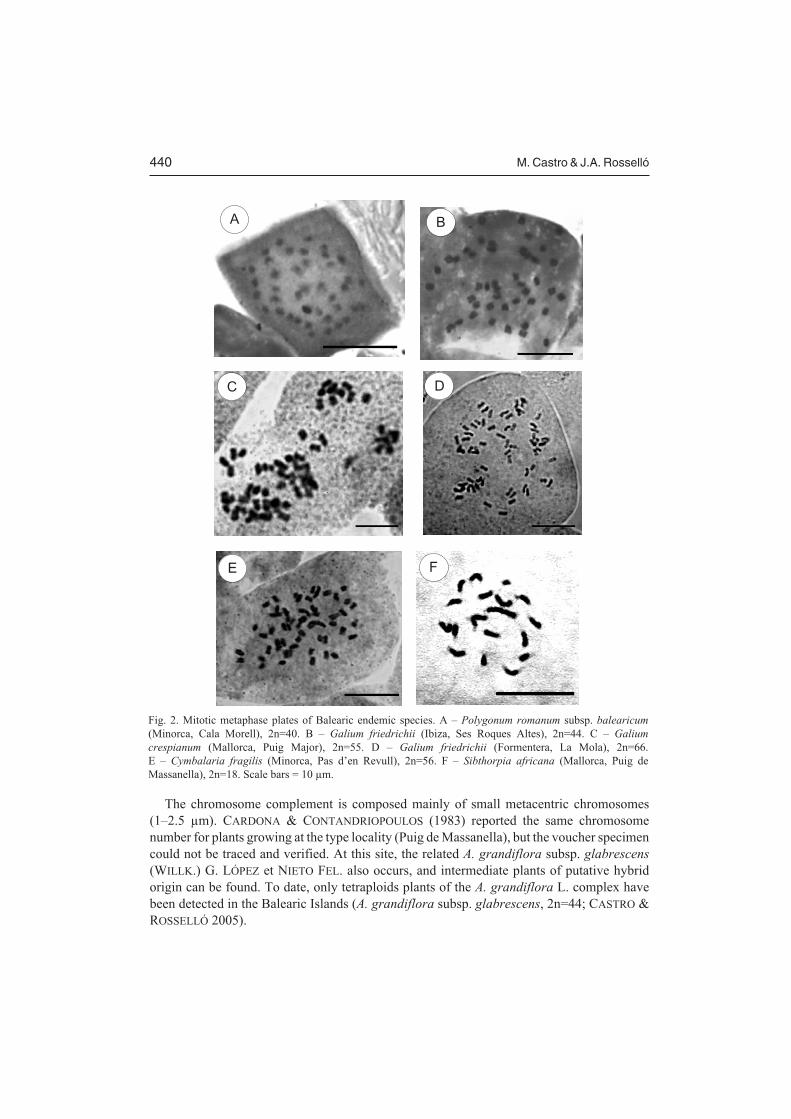
Fig. 2. Mitotic metaphase plates of Balearic endemic species. A – Polygonum romanum subsp. balearicum
(Minorca, Cala Morell), 2n=40. B – Galium friedrichii (Ibiza, Ses Roques Altes), 2n=44. C – Galium
crespianum (Mallorca, Puig Major), 2n=55. D – Galium friedrichii (Formentera, La Mola), 2n=66.E – Cymbalaria fragilis (Minorca, Pas d’en Revull), 2n=56. F – Sibthorpia africana (Mallorca, Puig deMassanella), 2n=18. Scale bars = 10 µm.

Euphorbiaceae
Euphorbia fontqueriana GREUTER
Mallorca, Escorca, Puig de Massanella (39°48�13� N, 2°50�53� E), 1250 m, slopes onsunny places, 8.II.2004, M.A. CONESA, M. MUS & J.A. ROSSELLÓ (VAL 156751). 2n=20(one individual).
Chromosomes are small in size (2–6 µm) showing mainly metacentric and submetacentricpairs (Fig. 1A); only a single pair of subtelocentric chromosomes is present. No secondaryconstrictions were detected in the metaphase plates. Our chromosome count agrees with thereport given by CARDONA (1991) for this species.
Fabaceae
Anthyllis hystrix (WILLK. ex BARCELÓ) CARDONA, CONTANDR. et SIERRA
Minorca, Ferreries, Barranc d’Algendar (39°58�19� N, 3°58�19� E), near Es Pas desRevull, 50 m, 7.II.2004, P. FRAGA & J.A. ROSSELLÓ (VAL 156752). 2n=70 (one individual).
Minorca, Mercadal, Cap de Cavalleria (40°4�41� N, 4°5�26� E), coastal slopes, VIII. 2004,P. FRAGA (VAL 156753). 2n=70 (five individuals).
Minorca, Ferreries, Ets Alocs (40°3�7� N, 3°59�5� E). Plants from the living stock of theBotanical Garden of Sóller (Balearic Islands) (VAL 156754). 2n=70 (three individuals).
Minorca, Ciutadella de Menorca, Marina de Curniola (40°3�10� N, 3°53�27� E), VIII.2004,P. FRAGA (VAL 156755). 2n=70 (five individuals).
Metacentric and submetacentric chromosome pairs are predominant in the complement,but due to their small size (2–3 µm), no other karyological features could be determined(Fig. 1B). The observed 2n=70 number differs from the 2n=84 (n=42) cytotype reported byCARDONA & CONTRANDRIOPOULOS (1983) and later figured by CARDONA et al. (1986) froma single population (Sa Mola de Fornells). These authors reported a normal meiosis in thepollen grains of A. hystrix. However, they pointed out that several bivalents showedconnections (CARDONA et al. 1986: Fig. 6a). A reinterpretation of this figure (using theirfigure 6b on A. hermanniae L. as reference for the visualization of centromeric constrictions)revealed an approximate number of n=35–36, which matches our results obtained on roottissues. Thus, the 2n=84 count for the Balearic plant should be viewed with great caution andshould be verified. On the basis of the 2n=70 counts, A. hystrix is a high polyploid (decaploid,x=7), and contrasts with the diploid level (2n=14) of the closely related A. hermanniae(CARDONA et al. 1986, CUSMA et al. 2002).
Anthyllis vulneraria L. subsp. balearica (COSS. ex MARES et VIGIN.) O. BOLOS et VIGO
Mallorca, Sóller, Serra d’Alfabia (39°43�53� N, 2°42�29� E), 900 m, vertical cliffs,2.XII.2004, M.A. CONESA, P. FRAGA & J.A. ROSSELLÓ (VAL 156756). 2n=12 (oneindividual).
Mallorca, Sóller, Serra de Son Torrella (39°47�8� N, 2°45�59� E), 800 m, calcareous cliffsand slopes, 2.IX.2004, M.A. CONESA, M. MUS & J.A. ROSSELLÓ (VAL 156757). 2n=12 (twoindividuals).
Chromosome numbers in Balearic endemics 441

Mallorca, Puig Major, Sa Coma de n’Arbona (39°47�41� N, 2°46�41� E), vertical cliffs.Seeds from the germplasm collection of the Botanical Garden of Sóller (Balearic Islands)(VAL 156758). 2n=12 (five individuals).
The karyotype of this taxon is composed of small chromosomes (1.5–3.5 µm), with threemetacentric, one submetacentric, and two subtelocentric pairs (Fig. 1C). This is the firstchromosome count of this Majorcan endemic taxon. BENEDÍ (2000) reported 2n=12 for thissubspecies on the basis of the chromosome counts given by DAHLGREN et al. (1971)(C. BENEDÍ, in litt.). These authors, however, clearly stated in their paper that the plantsanalyzed from NE Mallorca and Minorca (where subspecies balearica is absent) belong toA. vulneraria subsp. gandogeri (SAGORSKI) W. BECKER (sub A. vulneraria subsp.praepropera (A. KERN.) BORNM.). Chromosome counts of A. vulneraria subsp. gandogeriwere determined for comparative purposes (Mallorca, Arta, Colonia de Sant Pere (39°43�53�
N, 3°16�6� E), 1 m, rocky soils, 13.X.2004, M.A. CONESA & J.A. ROSSELLÓ VAL 156840)and also showed 2n=12. No significant karyotype differences were found between subspeciesbalearica and gandogeri.
Astragalus balearicus CHATER
Mallorca, Pollensa, Mortitx, Puig de Ses Parades (39°53�6� N, 2°54�23� E), slopes, 400 m,2.XII.2004, M.A. CONESA & J.A. ROSSELLÓ (VAL 156759). 2n=16 (two individuals).
442 M. Castro & J.A. Rosselló
A
C
B
D
Fig. 3. Haploid idiograms of selected Balearic endemic species. A – Lotus tetraphyllus, 2n=14. B – Crocus
cambessedesii, 2n=16. C – Phlomis italica, 2n=20. D – Lysimachia minoricensis, 2n=24. Scale bars = 5 µm.

Cabrera, without locality. Plants from the germplasm collection of the Botanical Garden ofSóller (Balearic Islands) (VAL 156760). 2n=16 (five individuals).
The karyotype of this species is composed of small chromosomes (1.5–3 µm), with fourmetacentric, three submetacentric, and one subtelocentric pairs. The same chromosomenumber was previously reported from single accessions from Mallorca (GUINOCHET &LEFRANC 1972) and Minorca (CARDONA 1977).
Lotus tetraphyllus L.f.Mallorca, Escorca, Puig de Massanella (39°48�13� N, 2°50�53� E), 750 m, slopes on an
evergreen oak forest, 8.II.2004, M.A. CONESA, M. MUS & J.A. ROSSELLÓ (VAL 156761).2n=14 (one individual).
Minorca, Ferreries, Barranc d’Algendar, near Es Pas des Revull (39°58�16� N, 3°57�36�
E), shady slopes 100 m, 7.II.2004, P. FRAGA & J.A. ROSSELLÓ (VAL 156800). 2n=14 (twoindividuals).
Minorca, Ferreries, Cala Galdana towards Cala Macarella (39°56�39� N, 3°56�53� E),100 m, 7.II.2004, P. FRAGA & J.A. ROSSELLÓ (VAL 156732). 2n=14 (two individuals).
The karyotype is composed of medium-sized chromosomes (3–4 µm), showing fivesubmetacentric and two metacentric pairs (Fig. 3A). No secondary constrictions wereobserved. Our results agree with the counts of DAHLGREN et al. (1971) and CARDONA (1973)on two samples from Minorca.
Globulariaceae
Globularia majoricensis GAND.Mallorca, Escorca, salt de la Bella Dona (39°48�14� N, 2°53�41� E), 450 m, calcareous
cliffs in shaded places, 14.II.2004, M.A. CONESA & J.A. ROSSELLÓ (VAL 156762). 2n=32.The karyotype of this species comprises small chromosomes (2–5 µm) mostly metacentric
and submetacentric, with some subtelocentric pairs (Fig. 1D). Our results agree with previouscounts by CONTANDRIOPOULOS (1978) and CARDONA & CONTANDRIOPOULOS (1980)reporting the same chromosome number. The deviating aneuploid number 2n=62 given bySCHWARTZ (1963) for this species has not been confirmed again and should probably bediscarded. In fact, infraspecific cytotypes, either aneuploid or polyploid races, have beenrarely detected in the genus (CONTANDRIOPOULOS 1978). Available evidence suggests thatthis Balearic endemic is tetraploid (x=8). Plants from the Iberian peninsula and southernFrance that are morphologically related to G. majoricensis (e.g. G. vulgaris L., G. valentinaWILLK.) show the tetraploid and (rarely) octoploid levels (VERLAQUE et al. 1995), and nodiploids are known. It is therefore likely that in this complex polyploidy predateddiversification.
Hypericaceae
Hypericum hircinum L. subsp. cambessedesii (COSS. ex BARCELÓ) SAUVAGE
Mallorca, Escorca, Gorg Blau (39°48�46� N, 2°49�29� E), 650 m, calcareous cliffs ina stream, 14.II.2004, M.A. CONESA & J.A. ROSSELLÓ (VAL 156725). 2n=32 (twoindividuals).
Chromosome numbers in Balearic endemics 443

The karyogram of this taxon showed small punctiform chromosomes (1.5–2 µm), withoutvisible centromeric constrictions (Fig. 1E). Surprisingly, a previous count from the samepopulation by REYNAUD (1986) reported 2n=40. According to this author, Hypericum sect.Androsaemum GODR., to which the Balearic taxon belongs, is characterized by x=10. Thisstatement was credited on the basis of her original work on H. grandifolium CHOISY (2n=40),H. hircinum subsp. hircinum (2n=40), and H. �inodorum MILL. (2n=40), as well as onprevious counts (2n=40) given by ROBSON (1981) for the former two taxa. However,contradictory results on species from the section Androsaemum were recently reported byMATZK et al. (2003). These authors indicated original counts for H. androsaemum L.(2n=36), H. foliosum AITON (2n=32), H. grandifolium (2n=32), H. �inodorum (2n=40), andH. hircinum subsp. hircinum (2n=32). Given these constant differences in chromosomenumber within the same taxa of the section Androsaemum made by three independent teams,it is likely premature to invoke technical errors to explain them. However, what is even moresurprising is the report of two different chromosome counts (2n=32 and 2n=40) within thesame population of H. hircinum subsp. cambessedesii, which is composed of less than a fewdozen individuals. REYNAUD (1986) stated that the base chromosome number in the sectionAndrosaemum was x=10. However, this hypothesis can be reinterpreted in a different way inthe light of the 2n=32 counts. If the base chromosome number in the section was x=8 insteadof x=10, the two different numbers within H. hircinum subsp. cambessedesii, 2n=32 and2n=40, might be due to the presence of anomalous pentaploid cells (2n=40) in some tissuesbesides regular tetraploid complement (2n=32). In this respect, it is interesting to note that ourchromosome counts and those from MATZK et al. (2003) were made on root tip cells, whereasREYNAUD (1986) counts were obtained from ovary cells from fixed flowers in the field.Karyological lability could be widespread in the genus since somaclonal variation in respectto chromosome number was reported in tissue culture-derived plants of H. perforatum L.plants (BRUTOVSKÁ et al. 1998).
Iridaceae
Crocus cambessedessi J. GAY
Mallorca, Pollença, Coll de la Creueta (39°56�20� N, 3°16�19� E), 100 m, maritime slopes,18.XI.2003, M.A. CONESA, M. MUS & J.A. ROSSELLÓ (VAL 148497). 2n=16 (twoindividuals).
Mallorca, Escorca, Puig Major (39°47�41� N, 2°46�41� E), 1350 m, slopes and rockcrevices, on shaded exposures, 2.X.2004, M.A. CONESA, M. MUS & J.A. ROSSELLÓ (VAL156763). 2n=16 (three individuals).
Minorca, Ciutadella, west face of Cala Morell (40°3�10� N, 3°52�45� E), 100 m, temporaryponds, 7.II.2004, P. FRAGA & J.A. ROSSELLÓ (VAL 156764). 2n=16 (two individuals).
Minorca, Maó, Cap Favaritx (39°59�43� N, 4°15�54� E), litoral scrub, X.2004, P. FRAGA
(VAL 156765). 2n=16 (two individuals).Minorca, Ferreries, El Pilar (40°3�7� N, 3°58�23� E), litoral scrub, X.2004, P. FRAGA
(VAL 156782). 2n=16 (two individuals).The karyotype is composed of five pairs of metacentric and three pairs of submetacentric
chromosomes (Fig. 3B). Two metacentric pairs are conspicuously longer (10–12 µm) than the
444 M. Castro & J.A. Rosselló

others (3–5 µm). Accessory chromosomes, reported in several species of the genus(BRIGHTON et al. 1973), were not seen. Identical chromosome counts from several Majorcanaccessions of this species were recorded also by BRIGHTON et al. (1973) and NILSSON &LASSEN (1971).
Lamiaceae
Phlomis italica L.Mallorca, Escorca, Puig de Massanella (39°48�13� N, 2°50�53� E), 1250 m, open slopes,
8.II.2004, M.A. CONESA, M. MUS & J.A. ROSSELLÓ (VAL 156766). 2n=20 + 0–1B (twoindividuals).
Mallorca, Escorca, Puig Major (39°47�41� N, 2°46�41� E), 1400 m, open slopes onnorthern exposures, 2.X.2004, M.A. CONESA, M. MUS & J.A. ROSSELLÓ (VAL 156767).2n=20 + 0–1B (five individuals).
Mallorca, Calvia, S’Esclop (39°37�53� N, 2°26�26� E), s.dat., J. GULÍAS (VAL 156768).2n=20 (five individuals).
Mallorca, Pollensa, Mortitx, Puig de ses Parades (39°53�6� N, 2°54�23� E), shady placesnear calcareous cliffs, 3.X.2004, M.A. CONESA & J.A. ROSSELLÓ (VAL 156769). 2n=20(five individuals).
Minorca, Ferreries, Sant Antoni de Ruma (40°0�56� N, 4°1�9� E), 100 m, 6.II.2004,P. FRAGA & J.A. ROSSELLÓ (VAL 156770). 2n=20 + 0–1B (two individuals).
Previously, the number 2n=20 was reported for a single accession from Minorca(CARDONA 1978). The regular chromosome complement is similar in all accessions and it iscomposed of ten metacentric chromosome pairs, ranging in size between 4.5–7 µm (Figs. 1F,3C). Nevertheless, intraindividual variation concerning chromosome numbers was present inmost accessions, and cells showing 20 and 21 chromosomes were usually observed (Fig. 1F).We suggest that B chromosomes are present in Balearic plants. No previous reports pointingout the presence of accessory chromosomes in the genus are known.
Polygonaceae
Polygonum romanum JACQ. subsp. balearicum RAFFAELLI et L. VILLAR
Minorca, Ciutadella, west face of Cala Morell (40°3�10� N, 3°52�45� E), 100 m, temporaryponds, 7.II.2004, P. FRAGA & J.A. ROSSELLÓ (VAL 156723). 2n=40 (three individuals).
Minorca, Ciutadella, east face of Cala Morell (40°3�10� N, 3°53�27� E), 125 m, temporaryponds, 7.II.2004, P. FRAGA & J.A. ROSSELLÓ (VAL 156771). 2n=40 (two individuals).
This is the first chromosome number count of this Majorcan endemic taxon. Nokaryological data concerning other subspecies of P. romanum are currently available. Thekaryotype is composed of small chromosomes (1–1.5 µm), similar in size, without visiblecentromeric constrictions (Fig. 2A). Taxa of Polygonum sect. Polygonum have the basechromosome number x=10, and on this basis it is suggested that P. romanum subsp.balearicum shows a polyploid (tetraploid) level.
Chromosome numbers in Balearic endemics 445

Primulaceae
Lysimachia minoricensis J.J. RODR.Seed collection from the Botanical Garden of Sóller (Mallorca). The species is extinct in
the wild, and the only known population of the species was from Sa Vall (Minorca, Ferreries)(VAL 156726). 2n=24 (five individuals).
Chromosomes ranged between 5–7 µm. The karyotype shows six metacentric and sixsubmetacentric pairs (Fig. 3D). A secondary constriction is visible in the short arms of one ofthe submetacentric chromosome pairs. The same chromosome number was obtained byCARDONA & CONTANDRIOPOULOS (1980), but no satellite chromosomes were observed.
Primula acaulis (L.) L. subsp. balearica (WILLK.) GREUTER et BURDET
Mallorca, Escorca, Puig de Massanella (39°48�13� N, 2°50�53� E), 1250 m, shady creviceson northern exposures, 8.II.2004, M.A. CONESA, M. MUS & J.A. ROSSELLÓ (VAL 156772).2n=22 (two individuals).
Mallorca, Escorca, Puig Major (39°47�41� N, 2°46�41� E), 1400 m, shady crevices onnorthern exposures, 2.X.2004, M.A. CONESA, M. MUS & J.A. ROSSELLÓ (VAL 156773).2n=22 (three individuals).
The chromosomes of this species are small (1–2 µm), similar in size, and without visiblecentromeric constrictions. Our counts agree with the observations of CARDONA &CONTANDRIOPOULOS (1980) on plants from a single accession.
Rubiaceae
Galium crespianum J.J. RODR.Mallorca, Escorca, Puig Major (39°47�41� N, 2°46�41� E), 1400 m, shady slopes and cliffs
on northern exposures, L. SÁEZ & J.A. ROSSELLÓ, 24.V.2003 (VAL 148482). 2n=55 (oneindividual).
Mallorca, Pollença, Coll de Sa Creueta (39°56�20� N, 3°16�19� E), 25 m, maritime slopes,18.XI.2003, M.A. CONESA, M. MUS & J.A. ROSSELLÓ (VAL 148483). 2n=55 (oneindividual).
Mallorca, Escorca, Puig de Massanella (39°48�13� N, 2°50�53� E), 1250 m, calcareouscrevices, 8.II.2004, M.A. CONESA, M. MUS & J.A. ROSSELLÓ (VAL 156774). 2n=55, 66(one individual, mixoploidy).
Mallorca, Caimari, Salt de la Bella Dona (39°48�14� N, 2°53�41� E), 450 m, calcareouscliffs, 5.II.2004, M.A. CONESA & J.A. ROSSELLÓ (VAL 156775). 2n=55, 66 (one individual,mixoploidy).
Mallorca, Pollensa, Mortitx, Puig de ses Parades (39°53�6� N, 2°54�23� E), calcareouscliffs on shady places, 3.X.2004, M.A. CONESA & J.A. ROSSELLÓ (VAL 156776). 2n=66(one individual).
Mallorca, Sóller, Serra d’Alfabia (39°43�53� N, 2°42�29� E), 900 m, vertical cliffs,2.XII.2004, M.A. CONESA, P. FRAGA & J.A. ROSSELLÓ (VAL 156777). 2n=66 (twoindividuals).
See G. friedrichii for the discussion of karyological features of the species.
446 M. Castro & J.A. Rosselló

Galium friedrichii N. TORRES, L. SÁEZ, MUS et ROSSELLÓ
Ibiza, Sant Josep de sa Talaia, Ses Roques Altes (38°54�27� N, 1°14�49� E), IV.2004, N.TORRES (VAL 156728). 2n=44 (one individual).
Ibiza, Sant Joan de Labritja, Els Cingles Negres (39°4�20� N, 1°24�58� E), IV.2004, N.TORRES (VAL 156727). 2n=66 (one individual).
Ibiza, Sant Antoni de Portmany, Els Alls (39°3�44� N, 1°20�49� E), IV.2004, N. TORRES,(VAL 156729). 2n=66 (one individual).
Formentera, El Pilar, La Mola (38°40�8� N, 1°34�28� E), open slopes and maritime cliffs,100 m, 28.VI.2003, J.A. ROSSELLÓ (VAL 148484). 2n=55, 66 (one individual, mixoploidy).
In both Galium species the karyotype is composed of small chromosomes (2–3.5 µm), withmainly metacentric and submetacentric pairs. No other karyological features could bediscerned with the method used. Galium crespianum and G. friedrichii are closely relatedspecies that have been confused in the past. Previously, 2n=44 was reported in both species,based on unpublished data on fixed floral buds by F. Ehrendorfer (TORRES et al. 2001).TORRES et al. (2001) cast some doubts on the accuracy of the 2n=66 record made byCARDONA (1976), since this author stated that the chromosome counts of G. crespianum weremade on mitotic plates from floral buds. However, although the voucher specimen effectivelydid belong to G. crespianum, the plant not only lacked flowers and floral buds but it did notshow any trace of inflorescence development, suggesting that this specimen was not really thefixed specimen used for cytological work. Thus, the possibility was suggested that duringfield work some labeling mistake with other species (e.g. Rubia balearica, 2n=66) growing inthe vicinity occurred. Our work has revealed a complex cytological scenario in both Galiumspecies, including tetraploid (2n=44), pentaploid (2n=55), and hexaploid cytotypes (2n=66),and, in addition, the somatic mosaics that combine two ploidy levels within one individual(mixoploids; Figs. 2B–D). Interestingly, this range of chromosome number variation was notdetected in both species when meiotic studies were made (EHRENDORFER in TORRES et al.2001), and this contrasts with the results reported here, made on mitotic plates from roots ofpotted plants collected in the field. Such variation can be explained by polyploidization andpast hybridization events. The tetraploid level (2n=44) might actually be the original ploidylevel for both species. The hexaploids formed via fertilization of unreduced gametes and thefollowing hybridization between tetraploids and hexaploids could give rise to pentaploidhybrids. Then, the instability of hybrids might lead to somatic segregation in their progenyand to mixoploidy. However, sequences from the ribosomal nuclear ITS region from bothspecies do not show single nucleotide additive polymorphisms (SNAPs) that would supporta hybridization scenario (ROSSELLÓ, unpubl. data). Vegetative reproduction may be the finalcause for such a cytological variation in the two Balearic species. In some plants a particularlevel of ploidy, or polysomaty, is specific to a certain tissue (CLOWES 1961). This has alsobeen confirmed by works in plant tissue culture, where plantlets formed from the callus tissuecan exhibit mutations and chimeras (D’AMATO 1978) or different ploidy levels (HU HAN etal. 1978). Thus, the formation of regenerating roots from the typical rootstock ofG. crespianum and G. friedrichii could be involved in the genesis of chromosome variantsthrough partial endopolyploidy and concomitant somatic segregation. Cytologicalcomparison of transplanted plants with seedlings obtained through seed germination could be
Chromosome numbers in Balearic endemics 447

useful to test the view that both species share a regular complement of 2n=44 and thatchromosomal instability is tissue-specific.
Rubia balearica (WILLK.) PORTA subsp. balearicaMallorca, Fornalutx, Balitx d’en Mig (39°48�13� N, 2°43�52� E), 325 m, cultured olive
fields, on open exposures, 17.XI.2003, M.A. CONESA, M. MUS & J.A. ROSSELLÓ (VAL148493). 2n=66 (one individual).
Mallorca, Caimari, Salt de la Bella Dona (39°48�14� N, 2°53�41� E), 450 m, shady slopesnear the road, 5.II.2004, M.A. CONESA & J.A. ROSSELLÓ (VAL 150475). 2n=66 (oneindividual).
The chromosome complement ranges between 1.5–2 µm, showing metacentric tosubtelocentric pairs. Our results agree with a previous count (CARDONA 1973), based onplants from an unspecified Majorcan population. However, the accessory chromosomesreported by CARDONA (1973) have not been observed in our samples.
Scrophulariaceae
Cymbalaria fragilis (J.J. RODR.) A. CHEVAL.Minorca, Ferreries, Barranc d’Algendar (39°58�16� N, 3°57�36� E), 100 m, near Es Pas des
Revull, vertical cliffs, 6.II.2004, P. FRAGA & J.A. ROSSELLÓ (VAL 156778). 2n=56 (threeindividuals).
The karyotype is composed of small (1.5–3 µm), metacentric and submetacentricchromosomes (Fig. 2E). This is the first chromosome count for this Balearic endemic plant.Cymbalaria aequitriloba (VIV.) A. CHEVAL., C. hepaticifolia (POIR.) WETTST. andC. fragilis are the only species of the genus that show this high polyploid (octoploid) level(SUTTON 1988).
Sibthorpia africana L.Mallorca, Escorca, Puig Major (39°47�41� N, 2°46�41� E), 1400 m, shady slopes and cliffs
on northern exposures, 24.V.2003, L. SÁEZ & J.A. ROSSELLÓ (VAL 148497). 2n=18 + 0–1B(one individual).
Mallorca, Pollença, Coll de Sa Creueta (39°56�20� N, 3°16�19� E), 50 m, maritime slopes,18.XI.2003, M.A. CONESA, M. MUS & J.A. ROSSELLÓ (VAL 148498). 2n=18 + 0–1B (twoindividuals).
Mallorca, Fornalutx, Balitx d’En Mig (39°48�13� N, 2°43�52� E), 325 m, cultured olivefields, on open exposures, 17.XI.2003, M.A. CONESA, M. MUS & J.A. ROSSELLÓ,17.XI.2003 (VAL 148499). 2n=18 + 0–1B (one individual).
Mallorca, Escorca, Puig de Massanella (39°48�13� N, 2°50�53� E), 1250 m, calcareouscrevices on shady exposures, 8.II.2004, M.A. CONESA, M. MUS & J.A. ROSSELLÓ (VAL150476). 2n=18 + 0–2B (one individual).
Minorca, Ferreries, near Es Pas des Revull (39°58�16� N, 3°57�36� E), shady slopes 100 m,7.II.2004, P. FRAGA & J.A. ROSSELLÓ (VAL 150477). 2n=18 + 0–1B (two individuals).
Ibiza, Sant Mateu d’Albarca, Cala Albarca (39°3�12� N, 1°21�32� E), 100 m, slopes nearmaritime cliffs, 17.XI.2004, N. TORRES & J.A. ROSSELLÓ (VAL 156779). 2n=18 + 0–1B(two individuals).
448 M. Castro & J.A. Rosselló

Chromosomes of this Balearic species are uniform in size, 2.5–3.5 µm long, mostlymetacentric and submetacentric. Previously, 2n=20 was reported for S. africana (LENSKI
1966, DAHLGREN et al. 1971, HEDBERG 1975, CARDONA & CONTANDRIOPOULOS 1980).However, we have observed cells showing 18 and 19 chromosomes within a single individualin all accessions (Fig. 2F); even a single individual from Massanella showed 18, 19 and 20chromosomes. This variation is best explained by the presence of a regular complement of2n=18 and a variable number of B chromosomes. The closely related S. europaea L. showsthe same chromosome number (2n=18; HEDBERG 1975, WENTWORTH et al. 1991, MORTON
1993), as that reported here for the Balearic species.
Violaceae
Viola odorata L. subsp. stolonifera (J.J. RODR.) ORELL et ROMO
Minorca, Ferreries, Barranc d’Algendar, near Es Pas des Revull (39°58�16� N,3°57�36� E), 100 m, shady slopes, 7.II.2004, P. FRAGA & J.A. ROSSELLÓ (VAL 156780).2n=20 (three individuals).
The karyotype of this Balearic taxon is composed of small (2–2.5 µm) metacentric andsubmetacentric pairs. Our count agrees with the report by CARDONA (1991) for this taxon, butthe reported accessory chromosomes were not seen.
Viola jaubertiana MARES et VIGIN.Mallorca, Pollensa, Mortitx, Puig de Ses Parades (39°53�6� N, 2°54�23� E), 400 m,
calcareous cliffs, 2.XII.2004, M.A. CONESA & J.A. ROSSELLÓ (VAL 156781). 2n=20 (twoindividuals).
The chromosome sizes of this species range between 1.5–2.5 µm, and the karyotype ismainly composed of metacentric and subtelocentric pairs. No secondary constrictions wereobserved. Our results agree with a previous chromosome count of GUINOCHET & LEFRANC
(1972), but the voucher specimen could not be traced and verified. At the Gorg Blau site, therelated V. alba BESSER also grows there, and intermediate plants of putative hybrid originhave been found (V. �balearica ROSSELLÓ, MAYOL & MUS).
Acknowledgements: We thank our colleagues M.A. Conesa, P. Fraga, M. Mus and N. Torres for their generoushelp with field sampling. Several accessions were obtained from the living and germplasm collections of theBotanical Garden of Sóller (Mallorca) thanks to the facilities given by J.L. Gradaille and M. Vicens. Lastly, thiswork has been greatly improved by the comments of G. Nieto-Feliner and an anonymous referee. This work hasbeen partly supported by funds of the project MMA 034/2002.
REFERENCES
BENEDÍ C. (2000): Anthyllis L. In: AEDO C., CASTROVIEJO S., HERRERO A., ROMERO C., SALGUEIRO F.J. &VELAYOS M. (ed.), Flora Iberica 7/2, Real Jardín Botánico, CSIC, Madrid, pp. 829–863.
BRIGHTON C.A., MATHEW B. & MARCHANT C.J. (1973): Chromosome counts in the genus Crocus (Iridaceae).Kew Bull. 28: 451–464.
BRUTOVSKÁ R., ÈELLÁROVÁ E. & DOLEZ¡EL J. (1998): Cytogenetic variability of in vitro regeneratedHypericum perforatum L. plants and their seed progenies. Pl. Sci. 133: 221–229.
CARDONA M.A. (1973): Contribution a l’étude cytotaxonomique de la flore des Baléares. I. Acta Phytotax.Barcinon. 14: 1–20.
Chromosome numbers in Balearic endemics 449

CARDONA M.A. (1976): Contribución al estudio citotaxonómico de la flora de Baleares. IV. Lagascalia 6:265–274.
CARDONA M.A. (1977): Contribució a l’estudi citotaxònomic de la flora de les Balears III (A contribution to thecytotaxonomic knowledge of the Balearic flora III). Butl. Inst. Catalana Hist. Nat. 41: 83–94.
CARDONA M.A. (1978): Contribució a l’estudi citotaxonomic de les Balears II (A contribution to thecytotaxonomic knowledge of the Balearic flora II). Colloq. Soc. Catalana Biol. 10–11: 51–67.
CARDONA M.A. (1991): [Reports]. In: STACE C.A. (ed.), IOPB Chromosome data 3, Int. Organ. Pl. Biosyst.Newslett. 17: 7–8.
CARDONA M.A. & CONTANDRIOPOULOS J. (1977): L’endemisme dans les flores insulaires méditerranéennes.Mediterranea 2: 49–77.
CARDONA M.A. & CONTANDRIOPOULOS J. (1979): Endemism and evolution in the islands of the WesternMediterranean. In: BRAMWELL D. (ed.), Plants and islands, Academic Press, London, pp. 133–169.
CARDONA M.A. & CONTANDRIOPOULOS J. (1980): Números cromosómicos para la flora espanola 162–182.Lagascalia 9: 272–284.
CARDONA M.A., CONTANDRIOPOULOS J. (1983): [Reports]. In: LÖVE Á. (ed.), IOPB chromosome numberreports LXXIX, Taxon 32: 323–324.
CARDONA M.A., CONTANDRIOPOULOS J. & SIERRA E. (1986): Étude biosystématique d’Anthyllis hystrix deMinorque et d’A. hermanniae de la Méditerranée orientale et centrale. Orsis 2: 5–25.
CASTRO M. & ROSSELLÓ J.A. (2005): Chromosome numbers in plant taxa endemic to the Balearic Islands. Bot.J. Linn. Soc. 148: 219–228.
CLOWES F.A.L. (1961): Apical meristems. Blackwell Scientific Publications, Oxford.CONTANDRIOPOULOS J. (1978): Contribution à l’étude cytogéographique du genre Globularia L. Biol. & Écol.
Médit. 5 : 3–14.CONTANDRIOPOULOS J. & CARDONA M.A. (1984): Caractère original de la flore endémique des Baléares. Bot.
Helv. 94: 101–131.CUSMA T.C., FEOLI L., KOSOVEL V., BACCHETTA G. & PATUI S. (2002): Mediterranean chromosome number
reports. Reports 1312–1314. Fl. Medit. 12: 475–480.DAHLGREN R., KARLSSON T.H. & LASSEN P. (1971): Studies on the flora of the Balearic Islands I.
Chromosome numbers in Balearic angiosperms. Bot. Not. 124: 249–269.D’AMATO F. (1978): Chromosome number variation in cultured cells and regenerated plants. In: THORPE A.
(ed.), Frontiers of plant tissue culture 1978, Proceedings of the 4th International Congress of Plant Cell andTissue Culture, International Association for Plant Tissue Culture, Alberta, pp. 287–295.
FAVARGER C. & CONTANDRIOPOULOS J. (1961): Essai sur l’endémisme. Ber. Schweiz. Bot. Ges. 71: 384–408GARBARI F., CORSI G. & MASINI A. (1991): Anatomical investigations in the Allium cupanii-A. hirtovaginatum
complex. Bot. Chron. 10: 805–808.GARBARI F., GREUTER W. & MICELI P. (1979): The Allium cupanii group: a preliminary taxonomic,
caryological and anatomical study. Webbia 34: 459–480.GUINOCHET M. & LEFRANC M. (1972): [Reports]. In: LÖVE Á. (ed.), IOPB chromosome number reports
XXXVII, Taxon 21: 497.HEDBERG O. (1975): A cytogenetic study of the genus Sibthorpia L. (Scrophulariaceae). Caryologia 28:
251–260.HU HAN H.T.Y., TSENG C.C., OUYANG T.W. & SHING C.K. (1978): Application of anther culture to crop plants.
In: THORPE T.A. (ed.), Frontiers of plant tissue culture, International Association for Plant Tissue Culture,Calgary, pp. 123–130.
KNOCHE H. (1922): Flora balearica. Étude phytogéographique sur les îles Baléares. 3. Roumégous & Déhen,Montpellier.
LENSKI I. (1966): Chromosomenzahlen in der Gattung Sibthorpia (Scrophulariaceae). Naturwissenschaften 53:710–711.
LEVAN A., FREDGA K. & SANDBERG A.A. (1964): Nomenclature for centromeric position on chromosomes.Hereditas 52: 201–220.
MATHEW B. (1996): A review of Allium sect. Allium. Royal Botanic Gardens Kew, Whitstable Litho Ltd, Kent.MATZK F., HAMMER K. & SCHUBERT I. (2003): Coevolution of apomixis and genome size within the genus
Hypericum. Sex Pl. Reprod. 16: 51–58.
450 M. Castro & J.A. Rosselló

MES T.H., FRITSCH R.M., POLLNER S. & BACHMANN K. (1999): Evolution of the chloroplast genome andpolymorphic ITS regions in Allium subg. Melanocrommyum. Genome 42: 237–247.
MICELI P. & GARBARI F. (1987): Allium eivissanum (Alliaceae), a new species from Eivissa (Balearic Isles,Spain). Willdenowia 16: 383–390.
MORTON J.K. (1993): Chromosome numbers and polyploidy in the flora of Cameroon Mountain. Opera Bot.121: 159–172.
NILSSON O. & LASSEN P. (1971): Chromosome numbers of vascular plants from Austria, Mallorca andYugoslavia. Bot. Not. 124: 270–276.
REYNAUD C. (1986): Étude cytotaxonomique des Millepertuis du Bassin méditerranéen et des Îles Canaries.Bull. Soc. Bot. France, Lettres Bot. 133: 167–177.
ROBSON N.K.B. (1981): Studies in the genus Hypericum L. (Guttiferae). 2. Characters of the genus. Bull. Brit.Mus. (Nat. Hist.), Bot. 8: 55–226.
ROSSELLÓ J.A., MUS M., TORRES N., MAYOL M. & IBÁNEZ M.V. (1993): De flora balearica adnotationes(11–13). Candollea 48: 593–600.
ROSSELLÓ J.A., TORRES N. & SÁEZ L. (1999): A new Biscutella (Brassicaceae) species from the westernBalearic islands. Bot. J. Linn. Soc. 129: 155–164.
SCHWARTZ O. (1963): Chromosomenzahlen, Lebensformen und Evolution der Gattung Globularia L. Drudea3: 5–16.
TORRES N., SÁEZ L., MUS M. & ROSSELLÓ J.A. (2001): The taxonomy of Galium crespianum J.J. RODR.(Rubiaceae), a Balearic islands endemic revisited. Bot. J. Linn. Soc. 136: 313–322.
SUTTON D.A. (1988): A revision of the tribe Antirrhineae. Oxford University Press, London.VERLAQUE R., REYNAUD C. & VIGNAL C. (1995): Mediterranean chromosome number reports. Reports
545–551. Fl. Medit. 5: 350–356.WENTWORTH J.E., BAILEY J.P. & GORNALL R.J. (1991): Contributions to a cytological catalogue of the British
and Irish flora, 1. Watsonia 18: 415–417.
Received 13 April 2005, first revision received 7 October 2005, second revision received and accepted 7 March2006
Chromosome numbers in Balearic endemics 451

452
Copyright © 2022 FDOKUMEN




















