New Approaches in the Study of Stimulus-Secretion Coupling in Anterior Pituitary Cells
18
New Approaches in the Study of Stimulus-Secretion Coupling in Anterior Pituitary Cells PIERRE-MARIE LLEDO,“.’ PHILIPPE VERNIER,“ WILLIAM T. MASON,b AND ROBERT ZOREC‘ aInstitut Arfred Fessard Centre National de la Recherche Scientij?que 91198 Gf-sur- Yvette Cgdex, France JEAN-DIDIER VINCENT,” bDepartment of Neurobiology Agriculture Food Research Council Babraham Cambridge CB2 4AT, United Kingdom ‘Laboratory of Neuroendocrinology Institute of Pathophysiology University of LjubIjana School of Medicine 61105 Ljubljana, Slovenia INTRODUCTION The eukaryotic cell is divided into a number of compartments bounded by mem- branes, including the nuclear envelope, endoplasmic reticulum, Golgi apparatus, lysosomes, various classes of endosomes, and a number of still poorly defined organ- elles. Membranes compartmentalize, but also isolate cells from their immediate envi- ronment. The process of exocytosis is an important means to overcome such isolation because secretory products may be released into the external milieu through both constitutive and regulated secretion. It is now well known that the flow of material through successive organelles of the secretory and endocytotic pathways is mediated by transport vesicles. These lipid vesicles are formed by budding of “donor” compartments and are precisely targeted to the membrane of the cognate “acceptor” compartment, with which they fuse. Clearly, membrane fusion must be a specific and carefully controlled process; other- wise, the vesicle “cargo” will be delivered to an incorrect acceptor compartment. Such a situation would destroy the highly differentiated, compartmentalized structure 301
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of New Approaches in the Study of Stimulus-Secretion Coupling in Anterior Pituitary Cells

New Approaches in the Study of Stimulus-Secretion Coupling in
Anterior Pituitary Cells PIERRE-MARIE LLEDO,“.’
PHILIPPE VERNIER,“
WILLIAM T. MASON,b AND ROBERT ZOREC‘
aInstitut Arfred Fessard Centre National de la Recherche Scientij?que
91198 Gf-sur- Yvette Cgdex, France
JEAN-DIDIER VINCENT,”
bDepartment of Neurobiology Agriculture Food Research Council
Babraham Cambridge CB2 4AT, United Kingdom
‘Laboratory of Neuroendocrinology Institute of Pathophysiology
University of LjubIjana School of Medicine 61105 Ljubljana, Slovenia
INTRODUCTION
The eukaryotic cell is divided into a number of compartments bounded by mem- branes, including the nuclear envelope, endoplasmic reticulum, Golgi apparatus, lysosomes, various classes of endosomes, and a number of still poorly defined organ- elles. Membranes compartmentalize, but also isolate cells from their immediate envi- ronment. The process of exocytosis is an important means to overcome such isolation because secretory products may be released into the external milieu through both constitutive and regulated secretion.
It is now well known that the flow of material through successive organelles of the secretory and endocytotic pathways is mediated by transport vesicles. These lipid vesicles are formed by budding of “donor” compartments and are precisely targeted to the membrane of the cognate “acceptor” compartment, with which they fuse. Clearly, membrane fusion must be a specific and carefully controlled process; other- wise, the vesicle “cargo” will be delivered to an incorrect acceptor compartment. Such a situation would destroy the highly differentiated, compartmentalized structure
301

302 ANNALS NEW YORK ACADEMY OF SCIENCES
of the eukaryotic cell and would disrupt the ordered, vectorial, sequential processing and trafficking of newly synthesized products. The means by which eukaryotic cells achieve such a degree of fidelity has been recently the topic of intense investigations that have highlighted the role of low-molecular-weight GTP-binding proteins.
The patch-clamp technique offers a number of tools to study the signal pathways involved. First, it allows the recording of whole-cell currents that are involved in the regulation of [Caz+Ii. Second, in favorable cases, it provides an assay for the secretory process itself through membrane capacitance measurements by providing direct measurement of surface area. The technique has such high-quality resolution that fusion events of even single vesicles can be recorded. Third, it is readily combined with the measurement of [Ca2+Ii by the calcium indicator dyes (i.e., fura-2, indo-1, fluo-3). Finally, it allows introduction of solutions of known composition into the cell interior (because there is rapid diffusional exchange between a patch pipette milieu and a small cell compartment) in the tight-seal whole-cell recording configuration.'
In this review, we describe some of these tools that have helped us to identify some molecular steps involved in the study of regulated secretion in anterior pituitary cells.
ANTERIOR PITUITARY CELLS: A MODEL FOR THE STUDY OF REGULATED SECRETORY EVENTS
Endocrine tissue share many properties with their neuronal counterpart. Secretory products from endocrine cells, including small molecules and expressed proteins, are (as in neurons) sequestered in two different cytosolic organelles: small vesicles and secretory granules (for a review, see reference 2). Vesicles (or synaptic vesicles in neurons) are small and very homogeneous in diameter (about 50 nm) and they may contain chemical neurotransmitters such as acetylcholine, GABA, glutamate, glycine, and the biogenic amines, but they do not contain soluble protein. Secretory granules (or large, dense vesicles in neurons) are larger (70-400 nm) and have a characteristic dense core in electron micrographs due to their content of soluble protein. Both classes of organelles release their contents by exocytosis, but the way by which they acquire these contents is different (see reference 2 for a review). Although the current view is that these two populations of vesicles are distinct and independent from one another, the possibility that the smaller vesicles are derived from the larger ones has not been ruled out definitively.
Even if there is still some debate, it has been generally accepted that vesicles are involved in the release of both neurotransmitters and hormonal products. In the pioneering work of D o ~ g l a s , ~ it was already suggested that hormonal release is an exocytotic process. When assaying the materials released from chromaffin cells with biogenic amines (norepinephrine and epinephrine), Douglas also found ATP, chro- mogranins, and dopamine P-hydroxylase. Moreover, these constituents were present in the same molar ratios within the secretory vesicles isolated from the medulla by centrifugation.
The adenohypophysis is another endocrine tissue composed of different cells se- creting a variety of hormones. These are stored within the cells in dense core granules

LLEDO et al.: ANTERIOR PITUITARY CELLS 303
and are released by appropriate stimuli. In addition to dense core granules, anterior pituitary cells contain small translucent vesicles, which are very simiIar to synaptic vesicles not only in the morphological appearance, but also with regard to their protein constit~ents.~.' For example, chromogranins and secretogranins found in secretory granules and small vesicles of endocrine cells have also been detected in their neuronal counterparts (for a review, see reference 6). It is further noteworthy that membrane proteins from synaptic vesicles such as synaptotagmin, a calcium-sensor protein (quoted also p65), have also been found associated with secretory granule membranes. All these findings indicate that neurons and endocrine cells are very similar with respect to the composition of their characteristic small vesicles and large secretory granules, besides having a variety of molecular, biochemical, and functional similarities.' Such similarities in the molecular machinery for secretion suggest that processes responsible for secretion of neurotransmitters and hormones are likely to have evolved from the trafficking machinery that controls secretion in more simple cells such as in yeast.8
Membrane Capacitance Reveals Exocytotic and Endocytotic Activities
Incorporation of membrane vesicles into the plasma membrane is a consequence of the process of exocytosis. As a result, the total cell surface may increase concomi- tantly with the secretory activity, depending on the number of fusing vesicles and on the respective rate of both exocytotic membrane fusion and endocytotic membrane retrieval. Therefore, it is possible to estimate the secretory activity of a single cell by quantifying the cell membrane capacitance, which is proportional to the membrane surface area.
As early as 1776, long before the nature of electricity was understood, Cavendish wondered whether the thin membranes of the cells in the electric organ of Torpedo (the electric ray) might not function like the glass walls of Leiden jars, which were used in that time to store electricity. Today, because cell membranes are able to maintain a difference of electrical potential, we know that they exhibit the property of electrical capacitance. Depending on the desired resolution, various methods can be used to assess membrane capacitance: (i) the pseudorandom binary sequence technique uses the transfer function of a given stimulation to obtain membrane capacitance;' (ii) the time-domain technique determines capacitance from the current relaxation in response to a voltage step;" and (iii) the two-phase lock-in amplifier technique derives passive membrane parameters by delivering a sinusoidal voltage command and measuring the resulting current response at two orthogonal phases." Only the last of these methods allows capacitance to be monitored in real time while providing the highest resolution.
Measurements of cell membrane capacitance (C,) have been applied to the investi- gation of the influence of [CaZ+Ii on exocytosis of cultured lactotrophs.I2 Intracellular [Ca2+] was modified by either dialyzing the cytosol with different calcium-containing pipette solutions or changing the membrane potential. High calcium resulted in a large C,,, increase, demonstrating the role for [Ca2+Ii in stimulus-secretion coupling as previously suggested in pituitary cells.13 Tonic calcium influx through voltage-gated

304 ANNALS NEW YORK ACADEMY OF SCIENCES
calcium channels, seen to occur at resting membrane potential, was found to play a major role in secretory activity monitored by C,.
More recently, a number of other signaling pathways have been suggested to modulate hormonal release from pituitary cells. In gonadotropes, breakdown of polyphosphoinositides induces calcium oscillations that trigger rhythmic exocyt~sis'~ (see also Hille et al. in this volume). In lactotrophs, cAMP was found to increase the magnitude and rate of calcium-induced exocytosis. In contrast, cAMP had no detectable effect on C,,, when intracellular calcium was It can therefore be concluded that cAMP facilitates calcium-induced secretion by acting directly on the secretory apparatus of anterior pituitary cells. The role of GTP-binding proteins on calcium-induced exocytosis has also been investigated in lactotrophs using nonhydro- lyzable GTP analogues (GTP-y-S and GMP-PNP) to irreversibly activate all GTPases. In this way, two distinct effects of G-protein activation could be distin- guished: the maximum C,,, increase due to intracellular calcium injection was dimin- ished, whereas the rate of C, increase (AC,,,/At) was facilitated, revealing a converse stimulatory role of G proteins in the translocation of secretory granules to the fusion sites. l6
PACKAGING AND STORAGE IN DIFFERENT SECRETORY ORGANELLES
There is abundant evidence that small-molecule transmitters are located in vesicles (for recent reviews, see references 2 and 17). Vesicular stores constitute a large reserve of transmitter that is protected from intracellular catabolism. These vesicles may contain small-molecule transmitters or neuroactive peptides. Because these latter are synthesized as secretory products, it can be assumed that essentially all of the peptides within an endocrine cell or a neuron are packaged within vesicles. Unlike small- molecule transmitters, none of these peptides are synthesized in the cytosol and no mechanism for regulating their cytoplasmic concentration need exist. The absence of specific enzymes for controIling the intracellular store of these messengers is an important feature.
The way by which secretory vesicles acquire their contents is different between vesicles and granules. Packaging into synaptic vesicles is performed from cytoplasmic pools; hence, synaptic vesicle membranes contain (i) electrogenic proton pumps mak- ing the inside positive with respect to the outside'" and (ii) chloride channels that dissipate the membrane potential and allow the buildup of an acidic environment inside the vesicle^.'^ The electrochemical gradient generated by the proton pump drives the accumulation of transmitters via several classes of neurotransmitter transporters. Therefore, after releasing their contents, the secretory vesicles reform and fill with transmitter from the cytoplasmic pools.
On the other hand, because secretory granules contain protein, they cannot refill in the cytoplasm after undergoing exocytosis. Hence, packaging and storage into secretory granules occurs in the Golgi complex rather than in the cytoplasm.

LLEDO et al. : ANTERIOR PITUITARY CELLS 305
THE ROLE OF CALCIUM IN EXOCYTOSIS
The concentration of free intracellular calcium ([Ca2+]i) has long been proposed to control a number of cellular events, such as membrane permeability, intracellular calcium release, and activation of enzymes or genes. In line with such a central role, calcium has also been recognized as the most important regulator of secretory processes. In his concept of “stimulus-secretion coupling”, Douglas3 proposed that releasable secretory granules docked at the plasma membrane fuse when calcium enters during membrane depolarization through voltage-gated calcium channels. Ex- citable tissues, such as neurons and endocrine cells, have several systems that regulate [Ca’+], levels. Calcium influx into the cytoplasm is largely mediated by voltage- sensitive and ligand-gated plasma membrane channelszo and by release from internal organelle compartments.21 On the other hand, mechanisms of reducing [Ca*+], levels include plasma membrane extrusion systems, such as the Ca2+-ATPase and the Na+- Ca2+ exchanger, organelle sequestration, and Caz+-binding proteins. All of these processes control both the resting [Ca2+]i levels and the regulation of [Ca2+Ii transients occurring as a result of electrical activity or other environmental chemical signals such as neurotransmitters. We shall first describe the calcium influx through voltage- sensitive calcium channels before proceeding to analyze some of the intracellular mechanisms involved in [Ca2+], homeostasis. Finally, some of the molecular steps that link intracellular calcium and exocytosis will be briefly described.
Calcium Influx through Voltage-gated Channels
To date, at least four major types of voltage-dependent calcium channels have been described in excitable cells (called T-, L-, N-, and P-channels) (for reviews, see references 20 and 22). These channels are distinguishable from one another by their voltage-dependence, time course of inactivation, and sensitivity to various antago- nists.
Because dopamine acts primarily as an inhibitory factor on prolactin release from lactotrophs, we have investigated its effects on voltage-gated calcium channel^.'^ We found that dopamine D2 receptor stimulation induced a reduction of both T- and L-type channel activity through two distinct mechanisms. Whereas dopamine de- presses the L-type current through a shift in the voltage-dependence of the activation curve towards depolarized values, without a change in current kinetics, it reduces the T-type current by increasing its tendency to inactivate. It should be added here that the mechanisms of action of acute dopamine application on ionic channel activity are quite similar to the well-established effects of a number of inhibitory transmitters (for a review, see reference 24). However, much less is known about modulation of ionic channels by chronic treatment with transmitters, which in some cases could mimic a physiological state. As an illustration of this long-term modulation, we have studied the effects of chronic stimulation of D1 receptors on ionic channel density in rat lactotroph cells, maintained in primary culture. In dopamine-free culture me- dium, the density of two different voltage-activated potassium currents remains con- stant with the duration of culture, whereas the densities of both types of calcium channels (L- and T-type) increase markedly. Because chronic dopamine D2 receptor

306 ANNALS NEW YORK ACADEMY OF SCIENCES
stimulation prevents this effect, it was concluded that the calcium current development reported in vitro could result from a loss of dopaminergic inhibition. These dopamine- induced effects on calcium channel density were reported to be CAMP-independent and it is probable that a reduction of protein kinase C activity mediates, at least in part, this chronic effect. Therefore, dopamine inhibition of calcium current involves both short- and long-lasting effects via the dopamine D2 receptor in lactotrophs. These findings may represent an aspect of the mechanism by which dopamine application tonically inhibits prolactin secretion.
Homeostasis of the Intracellular Free Calcium Concentration
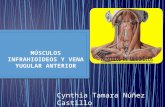
Among intracellular processes that regulate [Ca2+Ii, calcium-binding proteins play a central role because of their high and fast buffering capacity, their relatively high mobility within the cytoplasm, and their high affinity for calcium. In this respect, parvalbumin and the M, 28,000 protein ~ a 1 b i n d i n - D ~ ~ are believed to play an im- portant role in controlling [Ca2 +]i.25 We have used calbindin-transfected GH3 cell lines to examine the effect of calbindin on ionic channel activities and [Ca2+]i homeostasis. Because ionic channel activities are very susceptible to alterations during the whole- cell recording technique, the patch was perforated with the pore-forming antibiotic nystatin26 in these experiments, to enable recording of ionic currents. This allowed us to measure ionic currents and [Ca2+Ii levels simultaneously, without outward diffusion of cytoplasmic constituents and without introducing alien calcium buffers into the cell (FIGURE 1). Thus, we were able to measure calcium entry through voltage- sensitive calcium channels and to assess simultaneously the [Ca2+], level. Interestingly, only about 1/100th of the calcium influx through the plasma membrane of anterior pituitary cells was seen as free calcium at the peak of the [Ca2+Ii transient, revealing that most of the calcium ( 2 99%) that enters through calcium channels binds to endogenous calcium buffers.
This percentage is in reasonable agreement with the value used by Smith and ZuckerZ8 to fit Arsenazo I11 signals in Aplysia neurons. We have also examined the direct effects of introducing purified calbindin and other synthetic products known to be calcium buffers by recording responses in the whole-cell configuration. Our findings from these different experimental approaches indicate that the presence of calbindin affects both calcium influx through voltage-dependent calcium channels and [Ca2+Ii homeostasis. Therefore, this protein exhibits the characteristics of both a “buffer” protein, by binding internal calcium, and a “trigger” protein, by affecting calcium-regulating proteins such as calcium channels or the plasma membrane Ca2+/ Mg*+-ATPase. Through such a dual effect, calcium-binding proteins may play an important role in regulating exocytosis and could act as molecular switches whose “on” and “off’ states are triggered by binding and release of calcium. Conserved structure and mechanism in myriad versions of the switch (from bacterial to verte- brates) suggest that they all derive from a single primordial protein, repeatedly modi- fied in the course of evolution to perform a large variety of functions.
As reported above, the role of calcium release from internal pools has recently been demonstrated to regulate exocytosis in anterior pituitary cells. Tse and colleagues have shown that gonadotropin-releasing hormone induced the rhythmic release of
LLEDO et nl. : ANTERIOR PITUITARY CELLS 301
1 OOpA _yI_ ICa
1 OOms
I --"
Y
1 sec [Cali
FIGURE 1. Calcium current recordings and [Ca2+], change in depolarized cells. A GH3 cell was loaded with indo-1-AM. From a holding potential of -80 mV, the cell was depolarized to 0 mV (medium trace) for 200 ms using the perforated patch method.26 Calcium currents (upper trace) were corrected for leak current by adding four times the current activated during 20-mV hyperpolarizing pulses. External calcium concentration was 2.5 mM. The respective amount of charge flowing into the cell was 1.45 x lo-" C for a cell volume of 1220 pm3. [Ca2+], recordings (lower trace) were evaluated using photometric measurement^.^'
calcium from an inositol 1,4,Striphosphate (IP+sensitive store in pituitary gonado- tropesI4 (see also Hille et al. in this volume). Exocytosis measured in single gonado- tropes revealed that each elevation of [Ca2+]i induced a burst of exocytosis. From this finding, it was concluded that agonist-induced oscillations of [Ca2+]i in secretory cells may be a mechanism t o optimize the secretory output while avoiding the toxic effects of sustained elevation of [Ca2+]i.
The Exocytotic Machinery Is Regulated by Calcium
The role of the intracellular calcium concentration in secretion from endocrine cells has been studied in three ways: (i) the effect of elevated [Ca2+]i on the secretion

308 ANNALS NEW YORK ACADEMY OF SCIENCES
rate has been measured in permeabilized cell^;^^^^^ (ii) the average [Ca2+]i has been assessed during secretion triggered by depolarization or release of calcium from internal stores; and (iii) the secretion stimulated by perfusion of cells with solutions of different calcium concentrations through a whole-cell patch pipette has been moni- tored. These studies have shown that there is a threshold level of peak [Ca”], for hormone release to occur. More interestingly, the release of hormones and neurotrans- mitters has been found to be linearly related to the time integral of [Ca2+], elevation above this threshold.”
With dialysis and perfusion of calcium buffers, the rate of secretion achieved for a given [Ca2+]i is substantially less pronounced than that observed in response to a brief depolarization. This could be taken to indicate that depolarizing pulses open calcium channels such that [CaZ+li is locally elevated near release sites to levels sub- stantially higher than the average [Ca2+Ii recorded with photometric techniques. Consistent with such an hypothesis, even at high cytoplasmic [Ca2+]i (300-1000 nM), secretion rates during dialysis always remained clearly lower than those obtained during voltage pulses. Several other means of rapidly elevating [Ca’+]i also yield higher secretion rates. Conversely, the total amount of secretion is always much larger with calcium dialysis than with rapid calcium elevation using caged calcium ions or depolarization-induced calcium entry. These observations may be related to the existence of multiple pools of hormone, that is, a relatively small, rapidly releas- able pool and a larger reserve pool. In this respect, both movement of vesicles between pools and secretion have recently been shown to be sensitive to the intracellular calcium l e ~ e l . ~ ~ . ’ ~
THE ROLE OF SMALL GTP-BINDING PROTEINS IN EXOCYTOSIS
The superfamily of Ras-related, small GTP-binding proteins currently comprises over 50 members, which have been found to regulate a large spectrum of elementary cellular processes. Among them, the Rab gene subfamily members have in common not only their structural features, but also their ability to regulate intracellular vesicle traffic and sorting at the plasma membrane.34*35
By analogy with other guanine nucleotide-binding proteins, the Rab proteins are considered inactive in their GDP-bound form. This form is stabilized by guanine nucleotide dissociation inhibitors (GDI). Upon stimulation by a guanine nucleotide releasing protein (GNRP), the GDP is exchanged for GTP and the Rab proteins switch to their active, GTP-bound form. This state is quasi-irreversible until the Rab proteins hydrolyze the bound GTP to GDP; this effect is stimulated by a GTPase- activating protein (GAP). Thus, the proteins become inactivated, allowing the process to be repeated. Hence, as with other G proteins, Rab proteins behave as molecular switches cycling from an active (GTP-bound) to an inactive (GDP-bound) conforma- t i ~ n . ~ ~ . ~ ’ Because the general role of G proteins is to control the specificity and the temporal coherence of intermolecular recognition processes, it is very likely that Rab proteins act as regulators of vesicular targeting from an upstream to a downstream compartment .38

LLEDO et al.: ANTERIOR PITUITARY CELLS 309
Distribution and Function of Rab Proteins
Genetic mutants of yeast deficient for the function of two proteins belonging to the Rab family, SEC4 and YPTI, have provided the strongest evidence that Rab proteins may control the vectorial traffic of intracellular v e ~ i c l e s . ~ ~ ~ ~ The primary structure of the yeast SEC4 and YPTl gene products revealed that they belong to the Ras-like (or “small”, low molecular weight) G proteins. Immunolabeling studies show that SEC4 is present on the membrane vesicle^.^^^^ A search for the mammalian counterparts of SEC4 has led to the identification of a large number of Ras-related proteins termed Rab proteins (for “Ras-like proteins from rat brain”).
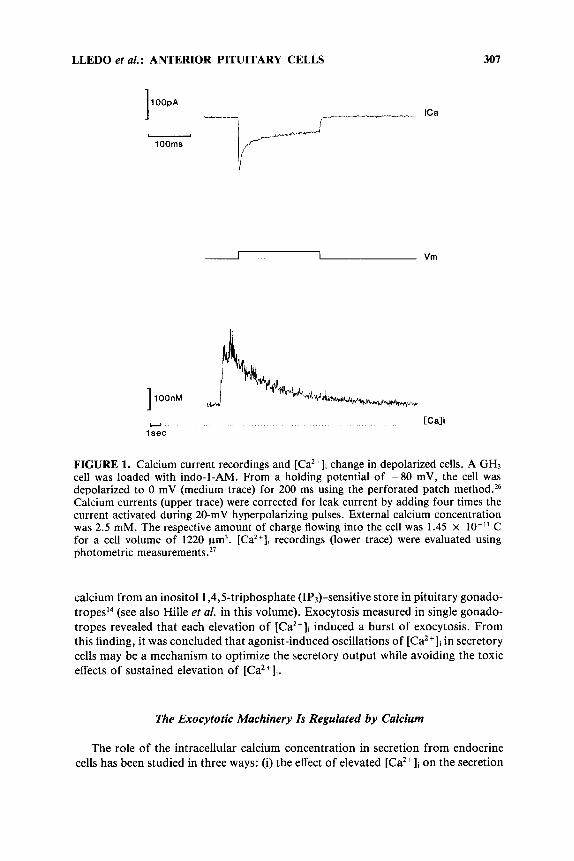
It is now clear that each Rab family member has distinct subcellular locations within cells and that most of the organelles involved in exocytosis or endocytosis possess at least one distinct member .5,41 Depending essentially upon the sequence of their C-terminus, each member of the Rab protein family seems to be associated with a particular vesicular compartment (see Pfeffer35 for a review). In addition, a posttranslational isoprenylation of cysteine residues at the C-terminus of the Rab proteins is required for their functional association with the vesicular membrane.42,43 Although most of the Rab family members are uniformly distributed in mammalian cells, they could also be involved in more differentiated secretory or compartmental- ized processes specific to neurons, endocrine cells, or epithelial cells. In this respect, it has been reported that Rab3A is specifically expressed in where it could control the recruitment of synaptic vesicles and their fusion with the plasma membrane during exo~ytos is .~~ .~* Other members of the Rab3 subfamily, termed Rab3B and Rab3C,49-s2 are thought to be associated with the membrane of secretory vesicles or granules, although this has not yet been demonstrated. In this respect, we have found that Rab3B is the major form found in rat anterior and intermediate pituitary gland, in contrast to Rab3A, which is highlyexpressed in selected brain areas like hippocampus, cerebellum, or thalamic n u ~ l e i ~ ~ - ~ ~ (FIGURE 2A). Some overlap expression for Rab3A and Rab3B mRNA also exists in the olfactory tubercle or striatum. More recently, Baldini and colleagues reported the cloning of Rab3D from mouse, which is proposed to control insulin-induced exocytosis in a d i p ~ c y t e s . ~ ~ This latter protein is homologous to the rat Rabl6 protein that is mainly expressed in lung, but in also many other peripheral and neuronal t iss~es.’~ Whether the different Rab3 proteins are functionally identical proteins expressed in different cell types or proteins associated with different kinds of secretory vesicles awaits further investiga- tion.
Assignment of SpeciJic Small G Proteins Involved in Exocytosis
The use of antibodies is an interesting approach to characterize the function of a specific protein (for a review, see reference 58) . However, there are inherent prob- lems in the use of antibodies, including the difficulty of introducing them into the cell, the low affinity of many antibodies directed against peptides, and uncertainty as to their specificity for preventing molecular interactions because of their large size. A more accurate method is to selectively impair the synthesis of specific proteins by introducing DNA oligonucleotides corresponding to the antisense orientation of
310 ANNALS NEW YORK ACADEMY OF SCIENCES
FIG
UR
E 2
. (A
) Nor
ther
n bl
ot a
naly
sis o
f 10
pg
of to
tal R
NA
ext
ract
ed f
rom
var
ious
tiss
ues
and
succ
essi
vely
hyb
ridiz
ed w
ith R
ab3B
and
R
ab3A
"P-la
bele
d cD
NA
pro
bes -
top:
hyb
ridiz
atio
n w
ith th
e ra
t Rab
3A p
robe
(aut
orad
iogr
am e
xpos
ed fo
r 8 ho
urs)
; bot
tom
: hyb
ridiz
atio
n w
ith th
e ra
t Rab
3B p
robe
(aut
orad
iogr
am e
xpos
ed fo
r 4 d
ays)
. (B
) Seq
uenc
es o
f the
kno
wn
Rab
3 pr
otei
ns a
nd th
e po
sitio
n of
the
antis
ense
an
d se
nse
olig
onuc
leot
ides
we
have
use
d. A
ntis
ense
olig
onuc
leot
ides
(ar
row
to
the
left)
: N
63 a
nd P
T3
over
lap
the
tran
slat
ion
initi
atio
n co
don
of R
ab3B
(bol
d lin
e) a
nd R
ab3A
(do
tted
line)
, res
pect
ivel
y. S
ense
olig
onuc
leot
ides
(ar
row
to th
e rig
ht):
N62
and
PT5
cor
resp
ond
to th
e nu
cleo
tides
188
to 2
08, r
elat
ive
to th
e fir
st A
TG
in th
e co
ding
reg
ion
of R
ab3B
(bol
d lin
e) a
nd R
ab3A
(do
tted
line)
, res
pect
ivel
y.
An
addi
tiona
l ol
igon
ucle
otid
e, N
61, c
ompl
emen
tary
to a
reg
ion
of t
he 3
'-unt
rans
late
d pa
rt o
f R
ab3B
mR
NA
, w
as a
lso
used
.

LLEDO et al.: ANTERIOR PITUITARY CELLS 31 1
the respective mRNAs. These antisense probes specifically block the translation of the corresponding message into protein. This strategy has been effective in reducing the concentration of a number of proteins or in preventing their induction, provided that the turnover rate of the protein to be blocked is sufficiently fast with regard to the limited lifetime of the antisense oligonucleotides them~elves .~~ Addition of specific antisense oligonucleotides to the cell medium has been used to cause depletion of G proteins in fibroblasts.m Microinjection, however, is a more direct and reliable route for the introduction of antisense oligonucleotides into cells and the efficacy of this method has been demonstrated in recent experiments. Kleuss et aL61,62 successfully blocked the expression of specific G proteins in cells of the anterior pituitary GH3 cell line by microinjection with antisense oligonucleotides. However, the introduction of the oligonucleotides into the cells by nuclear microinjection promoted the loss of a large number of injected cells. To prevent this drawback, we have developed a new antisense approach that allows the loading of DNA probes by diffusion from the patch pipette into the c y t o p l a ~ m . ~ ~ ~ ~ The technique consists of a sequential patch- clamp procedure that allows recordings from the same injected cell at several-day intervals. During the first whole-cell recording, the cells are loaded with the antisense- DNA and then, after various periods of time, the same cells are recorded again to determine the effects of protein absence on a specific cellular response.
We designed desoxyribonucleotides in an antisense orientation to Rab3A and Rab3B mRNAs to suppress the expression of these molecules. The antisense oligonu- cleotides PT3 and N63, overlapping the translation initiation site of Rab3A and Rab3B mRNAs, respectively (FIGURE 2B), inhibited the in vitro translation of the RNAs. In contrast, other oligonucleotides -either sense or antisense- were ineffi- cient. Only antisense for Rab3B, but not antisense for Rab3A, was efficient in reducing Rab3-like immunoreactivity on Western blots of the in vitro translation products of pituitary polyA-RNA.
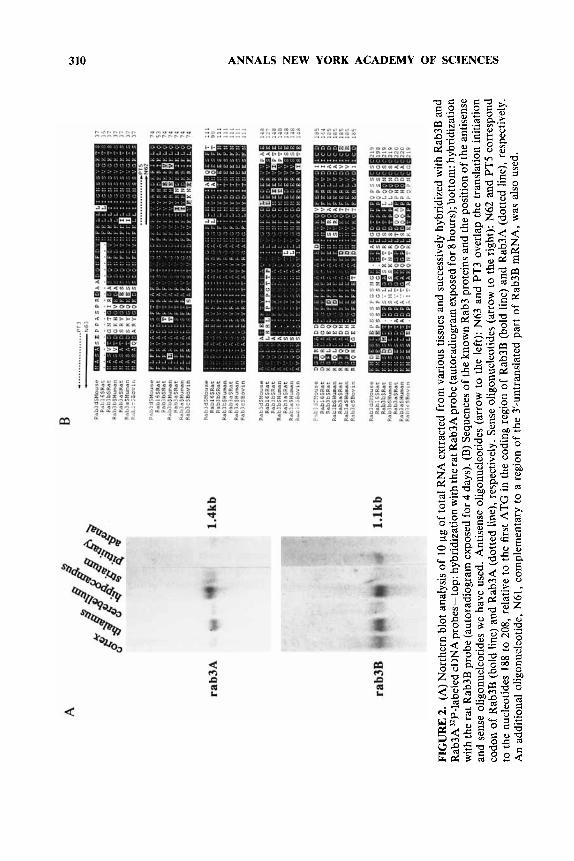
To check the effects of “knocking out” the expression of a target protein by our sequential patch-clamp procedure, we thus visualized cells by immunofluorescence microscopy after loading with sense or antisense probes. In control cells, immuno- staining for Rab3 was observed homogeneously over the cytoplasm (FIGURE 3). By contrast, the expression of immunoreactive Rab protein was strongly suppressed in cells stained 72 h after loading with antisense directed against Rab3B (FIGURE 3c). Interestingly, cells injected with the Rab3A antisense showed a fluorescence similar to nonloaded cells at all times after loading (FIGURE 3a). This finding demonstrates the specificity of our antisense probe in suppressing the Rab3B expression, the pre- dominant form of Rab3, present in anterior pituitary cells.
To investigate the function of Rab3B protein in the calcium-induced exocytosis, we made electrophysiological observations several days after loading the cell with specific oligonucleotides. To monitor changes in membrane capacitance (C,), whole- cell recordings with pipette solutions of various calcium concentrations were used. In control experiments, the increase in C, related to exocytosis is induced by dialysis of the cytosol with 1 pM calcium, as shown p r e v i o ~ s l y . ’ ~ ~ ~ ~ ~ ~ ~ The secretory response (AC,) measured as a change in C,,, relative to resting C,,, at 7.5 min after the start of the recording was increased by about 40% (FIGURE 4; upper trace). In contrast, cells preloaded with antisense oligonucleotide directed against Rab3B showed a reduced
312 ANNALS NEW YORK ACADEMY OF SCIENCES
FIGURE 3. Immunocytochemical detection of Rab3 proteins in cultured rat anterior pituitary cells. Cells were photographed sequentially to visualize marked cells with about 50,000 molecules of injected antisense oligonucleotides for Rab3 and FITC-IgG labeling of Rab3. (a) Phase-contrast image showing a cell 72 hours after the whole-cell recording and loading with oligonucleotide PT3 (arrow). (6) FITC-fluorescence image of the same cells as in part a showing similar staining among the loaded cell (arrow) and nonloaded cells. (c & d) FITC-fluorescence images of cells injected with oligonucleotide for Rab3B (arrows) and observed 72 hours or 110 hours afterwards, respectively. The immunofluo- rescence intensity was reduced only in the cell loaded for a duration of 72 hours with antisense (c).
secretory response 48 hours after loading, with ACm of about 15% (FIGURE 4; lower trace).
This inhibition was time-dependent because a diminution of secretory responses (i) was never observed 6 h after antisense loading, (ii) developed only partially at 24 h , and (iii) was maintained up to 72 h (FIGURE 5 ) .
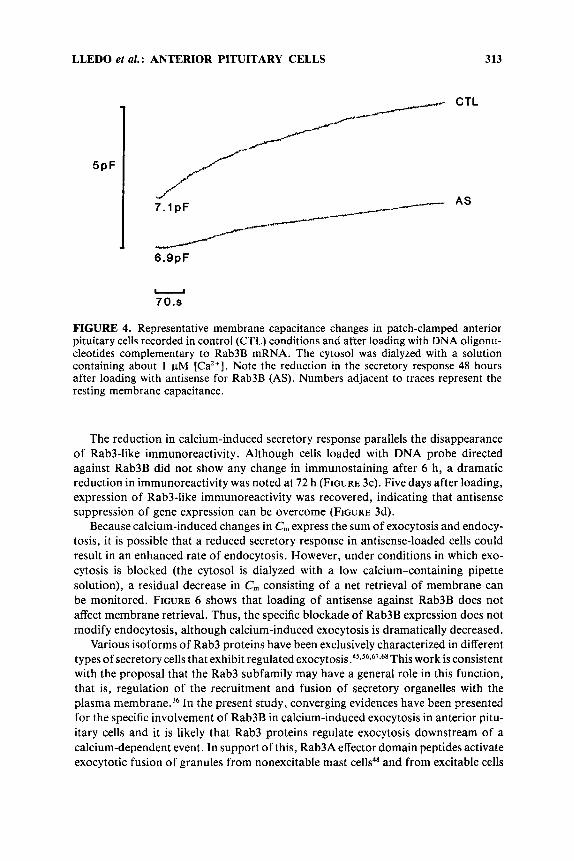
LLEDO et al. : ANTERIOR PITUITARY CELLS 313
.,
5PF
A 6.9pF
CTL
AS
. 70.9
FIGURE 4. Representative membrane capacitance changes in patch-clamped anterior pituitary cells recorded in control (CTL) conditions and after loading with DNA oligonu- cleotides complementary to Rab3B mRNA. The cytosol was dialyzed with a solution containing about 1 pM [Ca’+]. Note the reduction in the secretory response 48 hours after loading with antisense for Rab3B (AS). Numbers adjacent to traces represent the resting membrane capacitance.
The reduction in calcium-induced secretory response parallels the disappearance of Rab3-like immunoreactivity. Although cells loaded with DNA probe directed against Rab3B did not show any change in immunostaining after 6 h, a dramatic reduction in immunoreactivity was noted at 72 h (FIGURE 3c). Five days after loading, expression of Rab3-like immunoreactivity was recovered, indicating that antisense suppression of gene expression can be overcome (FIGURE 3d).
Because calcium-induced changes in C,,, express the sum of exocytosis and endocy- tosis, it is possible that a reduced secretory response in antisense-loaded cells could result in an enhanced rate of endocytosis. However, under conditions in which exo- cytosis is blocked (the cytosol is dialyzed with a low calcium-containing pipette solution), a residual decrease in C,,, consisting of a net retrieval of membrane can be monitored. FIGURE 6 shows that loading of antisense against Rab3B does not affect membrane retrieval. Thus, the specific blockade of Rab3B expression does not modify endocytosis, although calcium-induced exocytosis is dramatically decreased.
Various isoforms of Rab3 proteins have been exclusively characterized in different types of secretory cells that exhibit regulated exocyt~sis.~~,~~,~~.~~This work is consistent with the proposal that the Rab3 subfamily may have a general role in this function, that is, regulation of the recruitment and fusion of secretory organelles with the plasma membrane.36 In the present study, converging evidences have been presented for the specific involvement of Rab3B in calcium-induced exocytosis in anterior pitu- itary cells and it is likely that Rab3 proteins regulate exocytosis downstream of a calcium-dependent event. In support of this, Rab3A effector domain peptides activate exocytotic fusion of granules from nonexcitable mast cells48 and from excitable cells
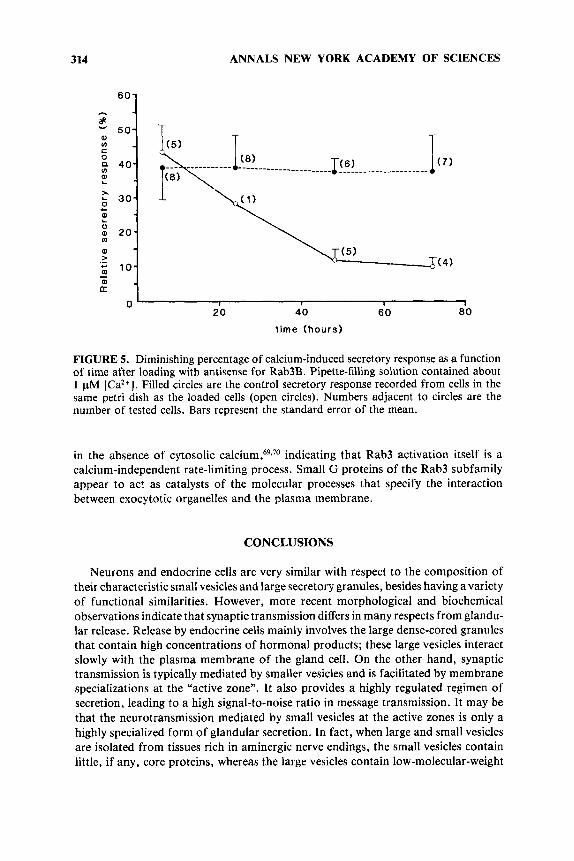
314
6 0 -
h ae - 5 0 - a, ln C 0 a 40-
?!
?! : 20-
;;; 10-
% ; 30- - ln
a, > .- - m u
ANNALS NEW YORK ACADEMY OF SCIENCES
0 ’ I I I 1 20 40 6 0 80
t ime (hours)
FIGURE 5. Diminishing percentage of calcium-induced secretory response as a function of time after loading with antisense for Rab3B. Pipette-filling solution contained about 1 FM [Ca2+]. Filled circles are the control secretory response recorded from cells in the same petri dish as the loaded cells (open circles). Numbers adjacent to circles are the number of tested cells. Bars represent the standard error of the mean.
in the absence of cytosolic cal~ium,6~*’~ indicating that Rab3 activation itself is a calcium-independent rate-limiting process. Small G proteins of the Rab3 subfamily appear to act as catalysts of the molecular processes that specify the interaction between exocytotic organelles and the plasma membrane.
CONCLUSIONS
Neurons and endocrine cells are very similar with respect to the composition of their characteristic small vesicles and large secretory granules, besides having avariety of functional similarities. However, more recent morphological and biochemical observations indicate that synaptic transmission differs in many respects from glandu- lar release. Release by endocrine cells mainly involves the large dense-cored granules that contain high concentrations of hormonal products; these large vesicles interact slowly with the plasma membrane of the gland cell. On the other hand, synaptic transmission is typically mediated by smaller vesicles and is facilitated by membrane specializations at the “active zone”. It also provides a highly regulated regimen of secretion, leading to a high signal-to-noise ratio in message transmission. It may be that the neurotransmission mediated by small vesicles at the active zones is only a highly specialized form of glandular secretion. In fact, when large and small vesicles are isolated from tissues rich in aminergic nerve endings, the small vesicles contain little, if any, core proteins, whereas the large vesicles contain low-molecular-weight

LLEDO et al.: ANTERIOR PITUITARY CELLS 315
8.9 pF
CTL
8.5 pF
FIGURE 6. Representative membrane capacitance changes in patch-clamped anterior pituitary cells recorded in control conditions and after loading with DNA oligonucleotides complementary to Rab3B mRNA. The cytosol was dialyzed with a solution containing about lo-* M [Ca*+]. The endocytotic response recorded 48 hours after loading with antisense for Rab3B (AS) is not significantly different from the control record (CTL). Numbers adjacent to traces represent the resting membrane capacitance.
amine transmitters, core proteins, and peptides. Because all the neuropeptides are found primarily, or even perhaps exclusively, in larger vesicles of neurons, nerve terminals would seem t o maintain the more “primitive” mechanism of release used by gland cells.
Membrane trafficking along the exocytotic and endocytotic pathways is mediated by a number of vesicular intermediates. This vesicular transport is vectorial because each step is mediated by specific targeting, docking, and fusion events. Among these processes, calcium- and GTP-regulated exocytosis lead to the final stage of the secre- tory pathway. The combination of yeast genetics, biochemistry of synaptic proteins, and in vitro reconstitution of vesicular transport has provided convincing evidence for a common mechanism that involves elements conserved through evolution such as calcium-sensor proteins and GTP-binding proteins.
REFERENCES
1. MARTY, A. & E. NEHER. 1983. Tight seal whole-cell recording. In Single-Channel Recording.
2. KELLY, R. B. 1993. Storage and release of neurotransmitters. CelVNeuron (Suppl.) 72: 43-53. 3. DOUGLAS, W. W. 1968. Stimulus-secretion coupling: the concept and clues from chromaffin and
other cells. Br. J. Pharmacol. 34: 451-474. 4. JAHN, R. & P. DE CAMILLI. 1991. Membrane proteins of synaptic vesicles: markers for neurons
and neuroendocrine cells; tools for the study of neurosecretion. In Markers for Neurons and Neuroendocrine Cells: Molecular and Cell Biology, Diagnostic Applications. First edition. M. Gratzl & D. K. Langley, Eds.: 25-92. Verlag-Chemie. Weinheim.
B. Sakman & E. Neher, Eds.: 107-121. Plenum. New York.

316 ANNALS NEW YORK ACADEMY OF SCIENCES
5 .
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21. 22.
23.
24.
25.
26.
27.
28.
29.
30.
SUDHOF, T. C. & R. JAHN. 1991. Proteins of synaptic vesicles involved in exocytosis and mem- brane recycling. Neuron 6: 665-677.
SOMOGYI, P., A. J. HODGSON, R. W. DEPOTTER, R. FISCHER-COLBRIE, M. SCHOBER. H. WINKER &I. W. CHUBB. 1984. Chromogranin immunoreactivity in the central nervous system: immuno- chemical characterization, distribution, and relationship to catecholamine and enkephalin pathways. Brain Res. Rev. 8: 193-230.
GRATZL, M. & D. K. LANGLEY, Eds. 1991. Markers for Neurons and Neuroendocrine Cells: Molecular and Cell Biology, Diagnostic Applications. First edition. Verlag-Chemie. Wein- heim.
BENNETT, M. K. & R. H. SCHELLER. 1993. The molecular machinery for secretion is conserved from yeast to neurons. Proc. Natl. Acad. Sci. U.S.A. 90: 2559-2563.
FERNANDEZ, J. M., F. BEZANILLA& R . E. TAYLOR. 1982. Distribution and kinetics of membrane dielectric polarization. J. Gen. Physiol. 79: 41-67.
LINDAU, M. & E. NEHER. 1988. Patch-clamp techniques for time-resolved capacitance measure- ments in single cells. Pfliigers Arch. Gen. Physiol. 411: 137-146.
NEHER, E. & A. MARTY. 1982. Discrete changes of cell membrane capacitance observed under conditions of enhanced secretion in bovine adrenal chromaffin cells. Proc. Natl. Acad. Sci. U.S.A. 79: 6712-6716.
ZOREC, R., F. HENIGMAN, W. T. MASON & M. KORDAS. 1991. Electrophysiological study of hormone secretion by single adenohypophyseal cells. Methods Neurosci. 4: 194-210.
TARASKEVICH, P. S. & W. W. DOUGLAS. 1984. Electrical activity in adenohypophyseal cells and effects of hypophyseotropic substances. Fed. Proc. Fed. Am. SOC. Exp. Biol. 43: 2373-2378.
TSE, A., F. W. TSE, W. ALMERS & B. HILLE. 1993. Rhythmic exocytosis stimulated by GnRH- induced calcium oscillations in rat gonadotropes. Science 260: 82-84.
SIKDAR, S. K., R. ZOREC & W. T. MASON. 1990. CAMP directly facilitates calcium-induced exocytosis in bovine lactotrophs. FEBS Lett. 273: 150-154.
SIKDAR, S. K., R. ZOREC, D. BROWN & W. T. MASON. 1989. Dual effects of G-protein activation on calcium-dependent exocytosis in bovine lactotrophs. FEBS Lett. 253: 88-92.
STEVENS, C. F. 1993. Quanta1 release of neurotransmitter and long-term potentiation. Cell/ Neuron (Suppl.) 72: 55-64.
FLOOR, E., P. S. LEVENTHAL & S. F. SCHAEFFER. 1990. Partial purification and characterization of the vacuolar H+-ATPase of mammalian synaptic vesicles. J. Neurochem. 55: 1663-1670.
TABB, J. S. , P. E. KOSH, R. VAN DYKE & T. UEDA. 1992. Glutamate transport into synaptic vesicles: roles of membrane potential, pH gradient, and intravesicular pH. J. Biol. Chem. 267: 1541 2-15418.
BEAN, B. P. 1989. Classes of calcium channels in vertebrate cells. Annu. Rev. Physiol. 51: 367- 384.
CARAFOLI, E. 1987. Intracellular calcium homeostasis. Annu. Rev. Biochem. 5 6 395-433. SNUTCH, P. & P. B. REINER. 1992. Calcium channels: diversity of form and function. Curr.
Opin. Neurobiol. 2: 247-253. LLEDO, P-M., V. HOMBURGER, J. BOCKAERT & J-D. VINCENT. 1992. Differential G protein-
mediated coupling of D1 dopamine receptors to K' and Ca2+ currents in rat anterior pituitary cells. Neuron 8: 455-463.
BEECH, D. J., L. BERNHEIM & B. HILLE. 1992. Pertussis toxin and voltage dependence distinguish multiple pathways modulating calcium channels of rat sympathetic neurons. Neuron 8: 97- 106.
CHRISTAKOS, S., C. GABRIELIDES & W. B. RHOTEN. 1989. Vitamin D-dependent calcium binding proteins: chemistry, distribution, functional considerations, and molecular biology. Endocr. Rev. 10: 3-26.
HORN, R. &A. MARTY. 1988. Muscarinic activation of ionic currents measured by a new whole- cell recording method. J. Gen. Physiol. 92: 145-159.
LLEDO, P-M., B. SOMASUNDARAM, J. MORTON, P. EMSON & W. T. MASON. 1992. Stable transfec- tion of ~ a 1 b i n d i n - D ~ ~ ~ into the GH, cell line alters Ca2+ currents and intracellular Ca2+ homeo- stasis. Neuron 9 943-954.
SMITH, S. J. & R. S. ZUCKER. 1980. Aequorin response facilitation and intracellular calcium accumulation in molluscan neurones. J. Physiol. 300: 167-196.
BAKER, P. F. & D. E. KNIGHT. 1978. Calcium-dependent exocytosis in bovine adrenal medullary cells with leaky plasma membranes. Nature (London) 276: 620-622.
GOMPERTS, B. D. & J. M. FERNANDEZ. 1985. Technique for membrane permeabilization. Trends Biochem. Sci. 10: 414-417.

LLEDO et al.: ANTERlOR PITUITARY CELLS 317
31.
32.
33.
34.
35.
36.
37. 38. 39.
40.
41.
42.
43.
44.
45.
46.
47.
48.
49,
50.
51.
52.
53.
54.
5 5 .
56.
PENG, Y. Y. & R. S. ZUCKER. 1993. Release of LHRH is linearly related to the time integral of presynaptic calcium elevation above a threshold level in bullfrog sympathetic ganglia. Neuron 10: 465-473.
NEHER, E. & R. ZUCKER. 1993. Multiple calcium-dependent processes related to secretion in bovine chromaffin cells. Neuron 10: 21-30.
THOMAS, P., J . G. WONG & W. ALMERS. 1993. Millisecond studies of secretion in single rat pituitary cells stimulated by flash photolysis of caged calcium. EMBO J. 12: 303-306.
VALENCIA, A., P. CHARDIN, A. WITTINGHOFER & C. SANDER. 1991. The ras protein family: evolutionary tree and role of conserved amino acids. Biochemistry 30: 4637-4648.
PFEFFER, S. R. 1992. GTP-binding proteins in intracellular transport. Trends Cell Biol. 2: 41- 46.
BOURNE, H. R., D. A. SANDERS & F. MCCORMICK. 1990. The GTPase superfamily: a conserved switch for diverse cell functions. Nature (London) 348: 125-132.
GRAND, R. J. & D. OWEN. 1991. The biochemistry of rus p21. Biochem. J. 2 7 9 609-631. BOURNE, H. R. 1988. Do GTPases direct membrane traffic in secretion? Cell 53: 669-671. SALMINEN, A. & P. J. NOVICK. 1987. A ras-like protein is required for a post-Golgi event in
yeast secretion. Cell 47: 527-538. GOUD, B., A. SALMINEN, N. C. WALWORTH & P. J. NOVICK. 1988. A GTP-binding protein
required for secretion rapidly associates with secretory vesicles and the plasma membrane in yeast. Cell 53: 753-768.
CHAVRIER, P., J. P. GORVEL, E. STELZER, K. SIMONS, J . GRUENBERG&M. ZERIAL. 1991. Hyperva- riable C-terminal domain of rub proteins acts as a targeting signal. Nature (London) 353:
MUSHA, T., M. KAWATA & Y. TAKAI. 1992. The geranylgeranyl moiety, but not the methyl moiety of the smg-25A/rub3A protein is essential for the interaction with membrane and its inhibitory GDP/GTP exchange protein. J. Biol. Chem. 267: 9821-9825.
KHOSRAVI-FAR, R., G. J . CLARK, K. ABE, A. D. Cox, T. MCLAIN, R. J . LUTZ, M. SINENSKI & C. J . DER. 1992. Ras (CXXX) and rub (CC/CXC) prenylation signal sequences are unique and functionally distinct. J. Biol. Chem. 267: 24363-24368.
MIZOGUCHI, A., S. KIM, T. UEDA, A. KIKUCHI, H. YORIFUII, N. HIROKAWA & Y. TAKAI. 1990. Localization and subcellular distribution of smg p25A, a ras p21-like GTP-binding protein, in rat brain. J. Biol. Chem. 265: 11872-11879.
DARCHEN, F., A. ZAHRAOUI, F. HAMMEL, M-P. MONTEILS, A. TAVITIAN & D. SCHERMAN. 1990. Association of the GTP-binding protein Rub3A with bovine adrenal chromaffin granules. Proc. Natl. Acad. Sci. U.S.A. 87: 5692-5696.
MATTEOLI, M., K. TAKEI, R. CAMERON, P. HURLBUT, P. A. JOHNSTON, T. C. S~~DHOF, R. JAHN & P. DE CAMILLI. 1991. Association of Rub3A with synaptic vesicles at late stages of the secretory pathway. J . Cell Biol. 3: 625-633.
FISCHER VON MOLLARD, G., T. C. S~~DHOF & R. JAHN. 1991. A small GTP-binding protein dissociates from synaptic vesicles during exocytosis. Nature (London) 349: 79-81.
OBERHAUSER, A. F., J. R. MONCK, W. E. BALCH & J. M. FERNANDEZ. 1992. Exocytotic fusion is activated by Rub3A peptides. Nature (London) 360: 270-273.
TOUCHOT, N., P. CHARDIN & A. TAVITIAN. 1987. Four additional members of the rus gene superfamily isolated by an oligonucleotide strategy: molecular cloning of YPT-related cDNAs from a rat brain library. Proc. Natl. Acad. Sci. U S A . 84: 8210-8214.
MATSUI, Y., A. KIKUCHI, J. KONDO, T. HISHIDA, Y. TERAMSHI & Y. TAKAI. 1988. Nucleotide and deduced amino acid sequences of a GTP-binding protein family with molecular weights of 25,000 from bovine brain. J . Biol. Chem. 263: 11071-11074.
ZAHRAOUI, A., N. TOUCHOT, P. CHARDIN &A. TAVITIAN. 1988. Complete coding sequences of the ras related rub3 and 4 cDNAs. Nucleic Acids Res. 16: 1204.
VERNIER, P., F. DARCHEN & J. MALLET. 1993. Tissular expression and posttranslational modifi- cation distinguish rub3A from rub3B in the rat. Manuscript in preparation.
AYALA, J., B. OLOFSSON, A. TAVITIAN & A. PROCHIANTZ. 1989. Developmental and regional regulation of rub3, a new brain specific rus-like gene. J. Neurosci. Res. 22: 241-246.
MOYA, K. L., B. TAVITIAN, A. ZAHRAOUI & A. TAVITIAN. 1992. Localization of the ras-like rub3A protein in the adult rat brain. Brain Res. 590: 118-127.
STETTLER, O., F. NOTHIAS, B. TAVITIAN & P. VERNIER. 1993. Differential expression of rub3 genes in the rat brain. Manuscript in preparation.
BALDINI, G., T. HOHL, H. Y. LIN&H. F. LODISH. 1992. Cloningof aRub3 isotypepredominantly expressed in adipocytes. Proc. Natl. Acad. Sci. U.S.A. 8 9 5049-5052.
769-772.

318 ANNALS NEW YORK ACADEMY OF SCIENCES
57.
58.
59.
ELFERINK, L. A,, K. ANZAI & R. H. SHELLER. 1992. Rub 15, a novel low molecular weight GTP-binding protein specifically expressed in the rat brain. J. Biol. Chem. 267: 5768-5775.
MILLIGAN, G. 1988. Techniques used in the identification and analysis of function of pertussis toxin-sensitive guanine binding proteins. Biochem. J . 255: 204-21 1.
HEIKKILA, R., G. SCHWAB, E. WICKSTROM, S. L. LOKE, D. H. PLUZNIK, R. WATT & L. M. NECKERS. 1987. A c-mycantisense oligonucleotide inhibits entry into S phase, but not progress from Go to G,. Nature (London) 328: 445-449.
WANG, H-Y., D. C. WATKINS & C. C. MALBON. 1992. Antisense oligonucleotides to G, protein a-subunit sequence accelerate differentiation of fibroblast to adipocytes. Nature (London)
KLEUSS, C., J. HESCHELER, C. EWEL, W. ROSENTHAL, G. SCHULTZ & B. WITTIG. Assignment of G-protein subtypes to specific receptors inducing inhibition of calcium currents. Nature (London) 353: 43-48.
62. KLEUSS, C., H. SCHERUBL, J . HESCHELER, G. SCHULTZ & B. WITTIG. 1992. Different P-subunits determine G-protein interaction with transmembrane receptors. Nature (London) 358: 424- 426.
BAERTSCHI, A. J., Y. AUDIGIER, P-M. LLEDO, J. M. ISRAEL, J. BOCKAERT & J-D. VINCENT. 1992. Dialysis of lactotropes with antisense oligonucleotides assigns G-protein subtypes to their channel effectors. Mol. Endocrinol. 6: 2257-2265.
LLEDO, P-M., P. VERNIER, J-D. VINCENT, W. T. MASON & R. ZOREC. 1993. Inhibition of rub3B expression attenuates calcium-dependent exocytosis in rat anterior pituitary cells. Nature (Lon- don) 364: 540-544.
65. THOMAS, P., A. SURPRENANT & W. ALMERS. 1990. Cytosolic Ca2+, exocytosis, and endocytosis in single melanotrophs of the rat pituitary. Neuron 5: 723-733.
66. ZOREC, R., S. K. SIKDAR& W. T. MASON. 1991. Increased cytosolic calcium stimulatesexocytosis in bovine lactotrophs. J. Gen. Physiol. 97: 473-497.
67. FISCHER VON MOLLARD, G., G. A. MIGNERY, M. BAUMERT, M. S. PERIN, T. J . HANSON, P. M. BURGER, R. JAHN & T. C. SUDHOF. 1990. Rub3 is a small GTP-binding protein exclusively localized to synaptic vesicles. Proc. Natl. Acad. Sci. U.S.A. 87: 1988-1992.
REGAZZI, R., A. KIKUCHI, Y. TAKAI & C. B. WOLLHEIM. 1992. The small GTP binding proteins in the cytosol of insulin-secreting cells are complexed to GDP-dissociation inhibitor proteins. J. Biol. Chem. 267: 17512-17519.
PADFIELD, P. J., W. E. BALCH & J. D. JAMIESON. 1992. A synthetic peptide of the rub3a effector domain stimulates amylase release from permeabilized pancreatic acini. Proc. Natl. Acad. Sci. U.S.A. 89: 1656-1660.
EDWARDSON, J. M., C. M. MACLEAN & G. J . LAW. 1993. Synthetic peptides of the rob3 effector domain stimulate a membrane fusion event in regulated exocytosis. FEBS Lett. 320: 52-56.
60.
358: 334-337. 61.
63.
64.
68.
69.
70.