
New Achievement in Panax vietnamensis Research
15
43 Abstract Panax vietnamensis Ha et Grushv., an endemic Panax genus of Vietnam, is a well known Vietnamese ginseng (Ngoc Linh ginseng) rich in pharmaceutical compounds, most importantly saponin. Its cultivation takes a long time, generally 5–7 years, and needs extensive efforts to quality control in the face of environmen- tal stresses including soil, shade, climate, pathogens and pests. In vitro techniques have been widely explored for rapid and efficient production of ginseng biomass and ginsenosides. The establishment of cell and adventitious root cultures of P. vietnamensis opens the way to commercial applications. Various physiological and biochemical parameters affecting the biomass production and ginsenoside accu- mulation have been investigated. These parameters are effect of various phytohor- mones, sucrose and activated charcoal (AC) on shoot regeneration and proliferation from callus, and adventitious and secondary root formation. The saponin analysis of calli and roots showed the presence of ginsenoside-Rg 1 , majonoside-R 2 , and ginsenoside-Rb 1 . These results indicated that P. vietnamensis biomass has a great potential to produce saponin as a new source for the pharmaceutical and cosmetic industry. Abbreviations AC Activated charcoal MS Murashige and Skoog NAA α-napthaleneacetic acid 2, 4-D 2, 4-dichlorophenoxyacetic acid TDZ Thidiazuron IBA Indole-3-butyric acid Chapter 3 New Achievement in Panax vietnamensis Research Duong Tan Nhut, Nguyen Thanh Hai, Nguyen Phuc Huy, Hoang Xuan Chien and Nguyen Ba Nam S. M. Jain and S. Dutta Gupta (eds.), Biotechnology of Neglected and Underutilized Crops, DOI: 10.1007/978-94-007-5500-0_3, © Springer Science+Business Media Dordrecht 2013 D. T. Nhut (*) · N. T. Hai · N. P. Huy · H. X. Chien · N. B. Nam Tay Nguyen Institute of Biology, 116 Xo Viet Nghe Tinh, Dalat, Lam Dong, Vietnam e-mail: [email protected]
-
Upload
independent -
Category
Documents
-
view
6 -
download
0
Transcript of New Achievement in Panax vietnamensis Research

43
Abstract Panax vietnamensis Ha et Grushv., an endemic Panax genus of Vietnam, is a well known Vietnamese ginseng (Ngoc Linh ginseng) rich in pharmaceutical compounds, most importantly saponin. Its cultivation takes a long time, generally 5–7 years, and needs extensive efforts to quality control in the face of environmen-tal stresses including soil, shade, climate, pathogens and pests. In vitro techniques have been widely explored for rapid and efficient production of ginseng biomass and ginsenosides. The establishment of cell and adventitious root cultures of P. vietnamensis opens the way to commercial applications. Various physiological and biochemical parameters affecting the biomass production and ginsenoside accu-mulation have been investigated. These parameters are effect of various phytohor-mones, sucrose and activated charcoal (AC) on shoot regeneration and proliferation from callus, and adventitious and secondary root formation. The saponin analysis of calli and roots showed the presence of ginsenoside-Rg1, majonoside-R2, and ginsenoside-Rb1. These results indicated that P. vietnamensis biomass has a great potential to produce saponin as a new source for the pharmaceutical and cosmetic industry.
Abbreviations
AC Activated charcoalMS Murashige and SkoogNAA α-napthaleneacetic acid2, 4-D 2, 4-dichlorophenoxyacetic acidTDZ ThidiazuronIBA Indole-3-butyric acid
Chapter 3New Achievement in Panax vietnamensis Research
Duong Tan Nhut, Nguyen Thanh Hai, Nguyen Phuc Huy, Hoang Xuan Chien and Nguyen Ba Nam
S. M. Jain and S. Dutta Gupta (eds.), Biotechnology of Neglected and Underutilized Crops, DOI: 10.1007/978-94-007-5500-0_3, © Springer Science+Business Media Dordrecht 2013
D. T. Nhut (*) · N. T. Hai · N. P. Huy · H. X. Chien · N. B. Nam Tay Nguyen Institute of Biology, 116 Xo Viet Nghe Tinh, Dalat, Lam Dong, Vietname-mail: [email protected]

44 D. T. Nhut et al.
3.1 Introduction
The word “ginseng” derives from the Chinese term, based on the roots of sev-eral distinct species of plants, mainly Korean or Asian ginseng (Panax ginseng), Siberian ginseng (Eleutherococcus senticosus), and American ginseng (Panax quinquefolius). All these species belong to the Araliaceae family. They are well known for various effects on human health. Recently, researchers in Asia and the West have been paying great attention to ginseng research. So far, more than 200 ginsenosides and non-saponin constituents have been isolated (Zang 2006). Ginsenosides show significant biological activities in physiological and pathologi-cal aspects. The demand for ginseng has dramatically increased worldwide, and has become very expensive due to inability to increase production. Moreover, tra-ditional methods take longer time, 5–7 years, and challenge the production cycles. Plant biotechnology is an efficient alternative approach for the mass production of ginseng and its active components. Several commercial products including health foods, drinks and cosmetics are produced from powder and extracts of ginseng cell cultures. The ginseng culture has continued to attract considerable research and development efforts in the recent years and scientists seek to understand and opti-mize the culture conditions. In Vietnam, P. vietnamensis Ha et Grushv. was found on highland of Central Vietnam in 1973, and was regarded as a precious medici-nal plant of local residents. P. vietnamensis is routinely used by the Sedang ethnic group, unknown to others, and use as a life-saving plant drug to treat many serious diseases, and to enhance body stamina during long journeys in high mountains. Nowadays P. vietnamensis supply is very limited because it grows slowly mainly in Ngoc Linh mountain area. Due to excessively harvesting, the ginseng is among 250 endangered species at high risk of extinction (Vietnam’s Red Data book). This ginseng species is used specifically for certain physical actions: anti-stress, anti-depression, in vitro and in vivo antioxidation, etc. Saponin triterpenoic compounds are the main effective group of compounds in the species. Chemical studies of the constituents of the plant have identified 23 saponins including 14 new com-pounds. Some of them are common to P. ginseng, such as protopanaxadiol and protopanaxatriol saponins, however, high concentration in this species. In addition, high concentration of ocotillol saponins, i.e., majonoside-R2 (5.3 % of the dried rhizome) was determined. Moreover, P. vietnamensis possessed the highest dam-maran-frame saponin (12–15 %) and saponin content among Panax genus. With these special features, this ginseng is one of the most valuable species in Vietnam and worldwide (Dong et al. 2007).
The initial work on propagation and culture medium improvement of this ginseng species had limited success Dung et al. (1995). Nhut et al. (2009a, 2009b) studied secondary root formation of P. vietnamensis, and the best results were obtained on SH medium supplemented with 3.0 mg/l 2,4-D. Jacques et al. (2007) investigated optimum conditions to increase ginseng biomass in bioreactor. Recently, Duong et al. (2008) analyzed to quantify ginsenoside-Rg1, -Rg2, -Rd in cell extract from gin-seng biomass by HPLC. Apparently, biomass collection and saponin analysis are

453 New Achievement in Panax vietnamensis Research
essential to validate in vitro growth. In the current research, the work is being carried out on the effects of medium compositions, culture conditions, explant size for in vitro multiplication of P. vietnamensis from callus induction to shoot and root regen-eration and quantification of saponin from biomass collected from in vitro cultures.
3.2 Micropropagation and Morphogenetic Programs
Since the traditional propagation of P. vietnamensis through seed and stem cut-tings is recalcitrant, in vitro techniques especially micropropagation would be an ideal approach for mass production; ginseng large-scale cell suspension culture for the production of pharmaceutical active compounds and fresh raw materials. Leaves and petioles explants of P. vietnamensis, grown at Tay Nguyen Institute of Biology, used to induce calli; washed with 5 % sodium hypochlorite solu-tion (Nhut et al. 2010), and followed by 2 h continuous washing with water. Explants were treated with 70 % alcohol while shaking for 30 s, and continuously rinsed 4–5 times with sterile distilled water; treated with 0.1 % HgCl2 contain-ing a few drops of Tweens-20, for 5 min, and washed in distilled water to remove HgCl2. The sterilized leaves were chopped in small pieces with a scalpel, size 1.0 × 10 cm. The petioles were vertically cleft, and cut in 1.0 cm long pieces.
The culture media used were MS basal medium (Murashige and Skoog 1962), modified MS ½ (originated essential minerals and half of microminerals) and modified MS ½ (half essential and microminerals) media supplemented with 30 g/l sucrose, 8.0 g/l agar and pH 5.7. The suitable culture conditions for callus induc-tion and development, shoot regeneration and proliferation are: 25 ± 2 °C, 16 h photoperiod, light intensity of 40 μ mol.s−1.m−2 fluorescent light, and relative humidity 75–80 %. Root regeneration and proliferation carried out in the darkness.
3.2.1 Callus Induction
It is important to standardize the protocol for ginseng shoot regeneration through callus. The regenerated plants are genetically identical to the original material, pathogen free and produce a large number of plantlets in a very short duration. However, the response of explants for callus induction and regeneration differ in different media. Therefore, it is required to establish an effective protocol for rapid plant regeneration from leaf and petiole explants.
3.2.1.1 Effect of Auxin Types and Concentration on Callus Induction from Leaf and Petiole
Research on Panax genus showed that callus induction stage usually required a combination of cytokinins and auxins. When Korean ginseng seeds are cultured,

46 D. T. Nhut et al.
MS induction medium supplemented with 1 mg/l 2,4-D and 0.01 mg/l kinetin was used (Arya et al. 1993) while leaf and other explants were cultured MS medium containing 1 mg/l 2,4-D and 0.1 mg/l kinetin (Lim et al. 1997). Nhut et al. (2011) used 0.2 mg/l TDZ in combination with 1.0 mg/l 2,4-D to investigate the effects on callus induction from P. vietnamensis leaf explants.
The effect of 2,4-D on callus induction from leaf and petiole explants after 8 weeks of culture is shown in Fig. 3.1. Among the 3 auxins (2,4-D, IBA, NAA) tested in the induction medium, 2,4-D exhibited the ability to stimulate cal-lus formation from leaf and petiole. On the induction culture medium containing 1.0 mg/l 2,4-D, cultured explants produced highest callus formation, 90 % from leaf and 100 % from petiole explants with a high number of rigid structure and bright yellow calli. 3.0 mg/l 2,4-D induced crystal callus formation on the same explants (Nhut et al. 2009a, 2009b). Radhakrishnan et al. (2001) suggested that the cells can only utilize limited amount of auxin and over-use of auxins at any level inhibits cell growth. Therefore, above 3.0 mg/l 2,4-D is not suitable for callus induction from P. vietnamensis leaves.
3.2.1.2 Effect of Lighting Condition on Callus Induction from Leaf and Petiole
Normally, induced callus cultures are kept under light or in the darkness depending on the type of explants, however, leaf-derived callus is maintained in the darkness. In some cases, culture explants produce better calli under the light. The previ-ous reports indicated that the number and quality of calli, grown in the darkness, are lower than in the light due to crystalline formation especially in the medium supplemented with 3.0 mg/l 2,4-D (Fig. 3.2a, b). These observations validate the results on Korean ginseng (Lim et al. 1997). Therefore, the lighting period 16 h/day is ideal to stimulate callus formation from P. vietnamensis leaves.
Fig. 3.1 Effect of 2,4-D on callus induction from leaf and petiole

473 New Achievement in Panax vietnamensis Research
3.2.1.3 Effect of Auxin Types and Concentration on Callus Multiplication
Our results showed that after multiplication stage, calli cultured on the medium supplemented with 0.5 mg/l IBA and the medium containing 1.0 mg/l 2,4-D had the highest dry weight and highest increase in callus dry weight, respectively (Fig. 3.2c, d).
Medina et al. (1998) demonstrated that carbohydrate is responsible for most of callus dry weight. The main carbon source in the culture medium is sucrose, and sugar utilization by callus depends on the medium and explant source - cotyledon, petiole, hypocotyl and leaf. The utilization of both auxin and cytokinin may help calli improving sugar and other nutrients absorption from the culture medium, and results in callus development and biomass dry weight increase. According to our results, IBA is more effectively used together with TDZ, rather with NAA and 2,4-D. Dry weight ratio of calli in 0.5 mg/l IBA-containing medium is highest among 3 treatments using 3 auxins. Although dry weight of calli in IBA-containing
Fig. 3.2 Callus formation and multiplication of P. vietnamensis. a Callus formation from leaves on medium containing 3.0 mg/l 2,4-D under 16 h photoperiod. b Callus formation from leaves on medium containing 3.0 mg/l 2,4-D in the darkness. c Callus multiplication on medium containing 1.0 mg/l 2,4-D under 16 h photoperiod. d Callus multiplication on medium containing 1.0 mg/l 2,4-D in the darkness

48 D. T. Nhut et al.
medium is highest, 2,4-D increased the highest dry weight ratio and relative high callus fresh weight. Alternatively, calli had the best conditions for growth and were highly regenerative in 2,4-D-containing medium.
3.2.1.4 Effect of Explant Size on Callus Development
Explant size is one of the most critical factors in in vitro multiplication. The initial difference in explant size can lead to a significant difference in cell density during the multiplication process, which can lead to many other changes in the culture medium (Akalezi et al. 1999). When investigating the effects of initial callus size on callus development, we learned that the smallest size (0.5 × 0.5 cm) gave the highest increase in fresh and dry weight. Some products released during the callus growth in the culture medium, which have toxic feedback to calli growth. Garcia and John (1983) observed that tobacco calli grown in the presence of 0.5–25 mg 2,4-D, higher internal ethane and ethylene were produced in the medium, which later decelerated callus multiplication. Also at same 2,4-D concentration, bigger callus size could produce more ethylene and ethane would be toxic to further cal-lus development of P. vietnamensis. Therefore, small callus size is ideal to pro-duce less ethylene and ethane and has higher rate of development.
3.2.2 Shoot Formation
3.2.2.1 Shoot Regeneration from Callus
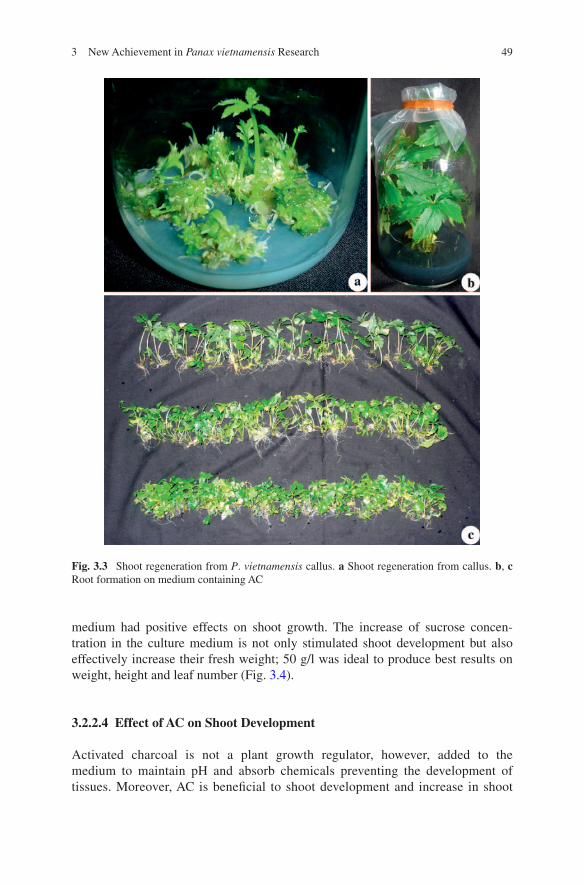
The auxin and cytokinin ratio plays an important role in shoot regeneration. Cytokinins usually promote shoot formation, which can be stimulated with a low concentration of auxins. The studies on the shoot regeneration ability in P. vietna-mensis callus with the addition of BA and NAA in the culture medium indicated that 1.0 mg/l BA and 1.0 mg/l NAA gave the highest shoot number (6.3 shoots/explant) with an average weight 0.185 g (Fig. 3.3a).
3.2.2.2 Effect of BA on Shoot Development
With the addition of 1.0 mg/l BA and 0.5 mg/l NAA in the culture medium, the best response was on shoot regeneration from P. vietnamensis callus, 0.87 g fresh weight of new shoots and height 6.16 cm.
3.2.2.3 Effect of Sucrose Concentration on Shoot Development
Normally, shoot regeneration requires 30–120 g/l sucrose in the culture medium. P. vietnamensis shoot development showed that adding sucrose into culture
493 New Achievement in Panax vietnamensis Research
medium had positive effects on shoot growth. The increase of sucrose concen-tration in the culture medium is not only stimulated shoot development but also effectively increase their fresh weight; 50 g/l was ideal to produce best results on weight, height and leaf number (Fig. 3.4).
3.2.2.4 Effect of AC on Shoot Development
Activated charcoal is not a plant growth regulator, however, added to the medium to maintain pH and absorb chemicals preventing the development of tissues. Moreover, AC is beneficial to shoot development and increase in shoot
Fig. 3.3 Shoot regeneration from P. vietnamensis callus. a Shoot regeneration from callus. b, c Root formation on medium containing AC

50 D. T. Nhut et al.
fresh weight. Our results indicated that an increase in AC concentration could lead to a considerable change in either shoot weight or height, but not the num-ber of leaves. The highest shoot weight was 1.01 g/shoot with the addition of 2.0 g/l AC in the culture medium, an increase by 1.9 fold with control (Fig. 3.3b, c). Thus, 2.0 g/l AC is the optimal concentration for P. vietnamensis shoot development.
The calli of P. vietnamensis was successfully induced from rhizome explants on MS medium containing 2,4-D and TDZ under 16 h photoperiod. The highest callus biomass yield was obtained on MS medium supplemented with 1.0 mg/l 2,4-D and 0.2 mg/l TDZ. The highest number of regenerated shoots from callus was in MS medium amended with 1.0 mg/l BA, 1.0 mg/l NAA and 50 g/l sucrose. In plantlet formation phase, 1/2 MS medium containing 1.0 mg/l BA, 0.5 mg/l NAA. 50 g/l sucrose and 2.0 g/l AC. After 6 months, under the lighting condi-tions 70 % red LEDs and 30 % blue LEDs is the most suitable medium and condi-tion for plantlet formation. After 6 months growth in the nursery, in vitro derived plantlet growth rate was faster than seed-derived plant growth rate and the survival rate was 87 %.
3.3 Adventitious Root Formation
In general, ginseng adventitious roots are an excellent starting material for the production of bioactive secondary metabolites without forging genes. However, adventitious root regeneration requires an understanding of basic research on plant physiology and biochemistry. So far, few reports are available on P. vietnamensis adventitious root and secondary root cultures. The more knowledge of the metab-olism and mineral nutrient uptake in root cultures, the more strategies for large-scale production need to be accomplished. Therefore, several studies were carried out to investigate to optimize environmental factors, which have crucial role in growth and development of ginseng explant in vitro.
Fig. 3.4 Effect of sucrose concentration on shoot regeneration from callus

513 New Achievement in Panax vietnamensis Research
3.3.1 Adventitious Root Formation from Callus
While investigating the effects of three auxins (IAA, IBA, NAA) we learned that IAA was not suitable for P. vietnamensis root formation. NAA and IBA could well stimulate rooting process. The two best results were obtained with the addition of 3.0 mg/l NAA and 5.0 mg/l IBA which gave 100 % root formation, 8.7 roots/explant, fresh weight root to explant ratio by an average of 21.88 %, and a longest root average of 13 mm; and 100 % root formation, 4.8 roots/explant, fresh weight root to explant ratio by an average of 15.81 %, and a longest root average of 18 mm, respectively (Fig. 3.5a). These phenomena could be explained by higher activity of synthetic auxins (IBA, NAA, 2,4-D) than the natural one (IAA). IAA was not able to stimulate root formation due to its low biological activity and its sensitivity to enzyme activity. George and Sherington (1984) observed that IAA, IBA and NAA used for root formation, among them, IBA was most effective. Moreover, auxin not only stimulated root development but also increased explant fresh weight; NAA and IBA were more effective than IAA (Kull and Arditti 2002).
3.3.2 Adventitious Root Multiplication
In order to choose the most suitable auxin for adventitious root regeneration and multiplication in P. vietnamensis, we continued to multiply adventitious roots using two auxin types, IBA and NAA. Our experiments indicated that NAA was suitable for adventitious root multiplication of P. vietnamensis. NAA at concen-tration 5.0 mg/l was optimal for root multiplication with the highest root forma-tion rate (60%), the highest secondary root formation (9 roots/explant) and the highest weight increase (average fresh weight: 390 ± 20 mg, increase 3.5 folds
Fig. 3.5 Effects of auxins on root formation and multiplication from P. vietnamensis callus. a Adventitious root formation from callus. b Root multiplication on medium supplemented with NAA

52 D. T. Nhut et al.
over the original one). In addition, five among six treatments supplemented with NAA showed root formation while IBA showed four among six treat-ments. As a result, 3.0 mg/l NAA is most suitable for root formation from cal-lus and 5.0 mg/l NAA is most suitable for adventitious root multiplication at P. vietnamensis.
3.3.3 Saponin of In Vitro Cultured P. vietnamensis Biomass
TLC analysis was used to determine Rf values of the compounds by their positions and colors showing on the plate. The results indicated that MR2 G-Rg1, and G-Rb1 were present in calli when compared with explant color and position on the plate with the standard compounds. Especially, the color chart from root weight showed the presence of three standard ginsenosides.
Ginsenoside types of callus and in vitro cultured biomass of ginseng genus depended on explant sources and supplemented auxin (Bonfill et al. 2002; Furuya et al. 1986). The ratio of group Rb/Rg in 2 year-old Korean ginseng rootstalk was 0.49 in the presence of 2,4-D after 5 weeks of culturing (Bonfill et al. 2002). William (2000) reported Rb group amount in 2 year-old Korean ginseng rootstalk was lower than Rg (0.6 vs. 1.0 %), indicating that saponin accumulation in Korean Ginseng callus is similar with natural explants. When trying to produce saponin from adventitious roots of Korean ginseng, Langhansova et al. (2005) found that total weight of ginsenosides of cultured roots in a bioreactor was about 14.48 mg/g biomass, while natural ginseng root contained 33.12 mg/g. In the total biomass, 5.02 mg/g G-Rb and 9.46 mg/g G-Rg compounds are present when compared with 15.06 mg/g G-Rb and 18.06 mg/g G-Rg in roots of Korean ginseng (Langhansova et al. 2005). These results showed that even in in vitro conditions, collected bio-mass is able to synthesize compounds, which are similar to the original explants.
Although there is no evidence of ginsenoside Rb and Rg group presence, our results showed that G-Rg1, a representative of 20(S)-protopanaxatriol group in callus, is present in very low amount in the original P. vietnamensis leaves. Analyzing saponin components in P. vietnamensis leaves showed 20(S)-protopanaxadiol derivatives, but not G-Rg1, hold a high ratio among saponin in stems and leaves (Dong et al. 2007); shows inability of G-Rb1 detection in leaf and shoot-derived callus. Moreover, majonoside-R2 was present in callus, absent in saponin components from leaves. Therefore, auxin and cytokinin had some influence on the multiplication process, and callus cells can synthesize majono-side-R2 themselves. This saponin is critical for medical uses of this ginseng.
Metabolite extracts from P. vietnamensis tissue grown in nature (1), and bio-mass of the petiole lTCL-derived calli (2) were run on TLC plate along with authentic standards of MR2, G-Rb1 and G-Rg1 (Table 3.1, Fig. 3.6). The results showed that biomass of calli included MR2, G-Rb1 and G-Rg1. Furthermore, metabolite extracted from biomass of calli also had the other bands correspond-ing to those present in extract from plant grown in nature, suggesting that calli had

533 New Achievement in Panax vietnamensis Research
similar chemical profile with those in natural environment. Saponins from in vitro P. vietnamensis calli were also analyzed using high-performance liquid chroma-tography with photodiode array detector at 190 nm (for MR2), and 203 nm (for G-Rb1, and G-Rg1) (Figs. 3.7, 3.8). With authentic saponin standards, HPLC anal-ysis revealed that all three important saponins of P. vietnamensis were present in the petiole lTCL-derived calli at high abundance. They included MR2 (0.424 %), G-Rg1 (0.061 %), and G-Rb1 (0.087 %).
In P. vietnamensis adventitious roots, all three main saponin groups were present: G-Rb1 representative of 20(S)-protopanaxadiol group, G-Rb1 representa-tive of 20(S)-protopanaxatriol group, and MR2 representative of ocotillol group. Although these compounds were not quantified yet, according to the strength of visualized colors, we could infer that MR2 has the highest amount, followed with G-Rg1 and finally with G-Rb1. These results are consistent with saponin
Table 3.1 Result of the saponin content in petiole lTCL-derived callus sample of P. vietnamen-sis
Weight of sample (mg) Standard Peak area
Content of saponin in sample
(μg) (%)
565.3 G-Rg1 924368 ± 23 0.320324 ± 0.00001 0.061MR2 483832 ± 28 2.213238 ± 0.00013 0.424G-Rb1 378858 ± 25 0.454827 ± 0.00003 0.087
Fig. 3.6 Fractions eluted from petiole lTCL-derived callus sample and reference sample. 1 Ref-erence sample. 2 Biomass of calli. 3 MR2. 4 G-Rb1. 5 G-Rg1

54 D. T. Nhut et al.
components presenting in natural P. vietnamensis roots, with 50 % MR2 in total saponin in root and rootstalk.
Besides chemical bands, which had the same positions with standard ones, there were different bands with other colors (green, or yellow, etc.) on the TLC plate. These alien bands indicated that auxin and cytokinin had stimulated the syn-thesis of non-saponin compounds in callus development. Types and compositions of these compounds are yet to be identified. In addition, there were some other bands, which almost had the same positions with two standard samples, Korean
Fig. 3.7 HPLC analysis of in vitro P. vietnamensis callus with PDA detection at UV wavelength 190 nm
Fig. 3.8 HPLC analysis of in vitro P. vietnamensis callus with PDA detection at UV wavelength 203 nm

553 New Achievement in Panax vietnamensis Research
ginseng and P. vietnamensis. There may be some other saponins, besides the standard ones, with low concentration, in ginseng callus cultures.
These results suggested that this lTCL morphogenesis protocol is feasible for in vitro culture of P. vietnamensis. There is no difference in saponin profile obtained from calli compared to field- grown plants.
3.4 Conclusion and Prospects
In recent years, P. vietnamensis has been extensively studied on pharmacologi-cal aspects, micropropagation and biomass production. The results suggested that P. vietnamensis regeneration and adventitious root protocols should strictly fol-low the steps regarding many biological and physical factors in the culture con-dition. P. vietnamensis callus was successfully induced from leaves and petioles cultured in MS medium supplemented with 1.0 mg/l 2,4-D, 0.2 mg/l TDZ under 16 h photoperiod. In callus multiplication stage, MS medium containing 1.0 mg/l 2,4-D, 0.3 mg/l TDZ was suitable for callus induction. Callus with small size (~0.5 × 0.5 cm) showed high growth rate. Callus regeneration was optimal on MS medium amended with 1.0 mg/l BA and 1.0 mg/l NAA. Subsequently, ½ MS
Fig. 3.9 Regeneration protocol of P. vietnamensis. a Callus induction. b Shoot and plantlet for-mation from callus. c Plantlet acclimatization. d Six-month-ginseng plant derived from in vitro plantlets

56 D. T. Nhut et al.
medium containing 1.0 mg/l BA, 0.5 mg/l NAA, 50 g/l sucrose and 2.0 g/l AC was most suitable for shoot development. In case of callus-derived root formation, the callus culture on MS ½ medium added with 3.0 mg/l NAA produced the highest root formation rate, root number and root fresh weight. MS ½ medium containing 5.0 mg/l NAA stimulated root multiplication and gave the highest secondary root formation. Saponin analysis showed that G-Rg1, MR2 and G-Rb1 present in callus and root biomass.
Recently Nhut et al. (2011) demonstrated high efficiency of plant regenera-tion with applied biotechnology in P. vietnamensis (Fig. 3.9). Biomass production through adventitious root is applicable to commercial propagation of P. vietnamensis. However, many problems still exist to scale-up and to improve saponin quantity in bio-mass. The main problems are: difficulties in controlling biochemical pathway, prepara-tion of explant and determination of the optimum culture conditions. They all depend on explant type, and different factors affected in culture process.
The efficient production of P. vietnamensis adventitious roots and bio-mass has great future prospect to enhance productive of pharmacy and cosmetic ginsenosides.
Acknowledgments The authors would like to thank the National Foundation for Science and Technology Development (Ministry of Science and Technology, Vietnam), the Committee for Local Science and Technology (Ministry of Science and Technology, Vietnam) and the Department of Application and Development of Technology (Vietnam Academy of Science and Technology) for the financial support.
References
Akalezi CO, Liu S, Li QS, Yu JT, Zhong JJ (1999) Combined effects of initial sucrose concentra-tion and inoculum size on cell growth and ginseng saponin production by suspension cultures of Panax ginseng. Process Biochem 34:639–642
Arya S, Arya IDI, Eriksson T (1993) Rapid multiplication of adventitious somatic embryos of Panax ginseng. Plant Cell Tiss Org Cul 34:157–162
Bonfill M, Rosa MC, Javier P, Teresa PM, Morales C (2002) Influence of auxins on organogen-esis and ginsenoside production in Panax ginseng calluses. Plant Cell Tiss Org Cul 68:73–78
Dong NT, Luan TC, Huong NTT (2007) Ngoc Linh Ginseng and some medicinal plants belong to Ginseng family. Science and Technology Publishing House, Vietnam
Dung NN (1995) Propagation of Panax vietnamensis by biological technology. Agricultural Publishing Houses, Vietnam
Duong VB, Dao Don DV, Long NV, Luong HV, Lau TV (2008) Study on quantitative analysis of some main ginsenosides of Ngoc Linh Ginseng by HPLC method. Med J 390:41–43
Furuya T, Yoshikawa T, Ushiyama K, Oda H (1986) Formation of plantlets from callus cultures of ginseng (Panax ginseng). Experientia 42:193–194
Garcia FG, John WE (1983) Ethylene and ethane production in 2,4-D treated and salt treated tobacco tissue cultures. Ann Bot 51:287–295
George EF, Sheringtoln PD (1984) Plant propagation by tissue culture. Exegetics Ltd., EversleyJacques P, Kevers C, Gaspar T, Dommes J, Thonart P (2007) Condition Panax vietnamensis cell
mass production in bioreactors. Acta Bot Gallica 154(1):21–26Kull T, Arditti J (2002) Orchid biology reviews and perspective, vol 8. Kluwer Academic
Publishers, The Netherland, pp 443–487

573 New Achievement in Panax vietnamensis Research
Langhansová L, Maršík P, Vanek T (2005) Production of saponins from Panax ginseng suspen-sion and adventitious root cultures. Biol Plantarium 49(3):463–465
Lim HT, Lee HS (1997) Regeneration of Panax ginseng C. A. Meyer by organogenesis DNA analysis of regenerants. Plant Cell Tiss Org Cul 49:179–187
Medina M, Villalobos N, Cruz PJDL, Dorado A, Guerra H (1998) Effect of culture medium and light conditions on the morphological characteristics and carbohydrate contents of Medicago strasseri calli. Acta Physiol Plant 20(4):383–392
Murashige T, Skoog F (1962) A revised medium for rapid growth and bioassays with tobacco tis-sue cultures. Physiol Plant 15:473–497
Nhut DT, Luan VQ, Binh NV, Phong PT, Huy BN, Ha DTN, Tam PQ, Nam NB, Hien VT, Vinh BT, Hang LTM, Ngoc DTM, Thao LB, Luan TC (2009a) The effects of some factors on in vitro biomass production of vietnamese ginseng (Panax vietnamensis ha et grushv.) and pre-liminary analysis of saponin content. J Biotechnol Viet 7(3):365–370
Nhut DT, Tam PQ, Luan VQ, Binh NV, Ha DTN, Chien HX, Nhan NCT Nam NB, Truc NB, Thuy LNM, Nguyen PLH, Hien VT, Hang LTM, Hang NTT, Hai NT (2009b) Adventitious and secondary root formation of Panax vietnamensis cultured in vitro. In: Proceedings of biotechnology conference of southern provinces of Vietnam. Science and Technology Publishers, pp. 252–258
Nhut DT, Hang LTM, Vinh BT, Tam PQ, Nam NB, Nhan NCT, Chien HX, Thuy LNMT, Hien VT, Binh NV, Luan VQ, Luan TC, Duc DT (2010) Identification of saponin content and resi-due of some phytohormones in callus, shoot and root of Panax vietnamensis cultured in vitro. J Biotechnol Viet 8(2):189–202
Nhut DT, Huy NP, Luan VQ, Binh NV, Nam NB, Thuy LNM, Ha DTN, Chien HX, Huong TT, Cuong HV, Cuong LK, Hien VT (2011) Shoot regeneration and micropropagation of Panax vietnamensis Ha et Grushv. from ex vitro leaf-derived callus. Afr J Biotechnol 10(84):19499–19504
Radhakrishnan T, Murthy TGK, Chandran K, Banyopadhyay A (2001) Somatic embryogenesis in Arachis hypogaea revisited. Aust J Bot 49:753–759
William EC (2000) Ginseng. Harwood Academic Publishers, The Gordon and Breach Publishing Group
Zhang JT (2006) The chemistry, metabolism and biological activities of ginseng. Chemical Industry Press, Beijing, pp 3–17




















