
Neuroendocrine–immune (NEI) circuitry from neuron–glial interactions to function: Focus on...
18
Introduction Substantial evidence from several experimental and clinical studies strongly indicates that bidirectional communication between the neuroendocrine and immune systems represents one cardinal feature of a living organism, both in healthy and diseased states. Indeed, since the early studies of Pierpaoli, Sorkin and Besedovsky, 1–4 evidence accumulated in the last decades has clearly documented the vital importance of interacting neuroendocrine–immune (NEI) networks in the regulation of physiological homeostatic mechanisms, in particular with regard to reproductive functions. 5–16 The concept of integrated bidirectionally regulated NEI axes is underscored during preperinatal life, when exposure to abnormal hormonal levels or to endotoxin results in major alterations in the programming of both neuroendocrine and immunological regulatory mechanisms. 4,13,14,17–23 Indeed, a number of experimental and epidemiological studies clearly suggest that early events occurring in fetal life powerfully modulate an individual’s predisposition to different patho- logic states in the adult life. 21–23 Two major endocrine axes dramatically impinge on neuro- immunomodulation: the hypothalamic–pituitary–gonadal (HPG) and the hypothalamic–pituitary–adrenocortical (HPA) axes. The brain–pituitary–reproductive axis and the brain– thymus–lymphoid axis have been shown to communicate via an array of internal mechanisms of communication that use similar signals (neurotransmitters, peptides, growth factors, hormones) acting on similar recognition targets (the recep- tors). 10,11,14,15,24 Moreover, such communication networks form the basis for the control of each step and every level of reproductive physiology. 24 One principal conveying signal is luteinizing hormone-releasing hormone (LHRH), the key reproductive hormone coordinating the major features of mammalian reproduction (Fig. 1). The hypothalamic deca- peptide can modulate immune functions directly, through Immunology and Cell Biology (2001) 79, 400–417 Special Feature Neuroendocrine–immune (NEI) circuitry from neuron–glial interactions to function: Focus on gender and HPA–HPG interactions on early programming of the NEI system MC MORALE, 1 F GALLO, 1 C TIROLO, 1 N TESTA, 1 S CANIGLIA, 1 N MARLETTA, 2 V SPINA-PURRELLO, 2 R AVOLA, 2 F CAUCCI, 3 P TOMASI, 3 G DELITALA, 3 N BARDEN 4 and B MARCHETTI 1,5 Departments of 5 Pharmacology and 3 Endocrinology, Medical School, University of Sassari, Sassari, 1 Department of Neuropharmacology, OASI Institute (IRCCS) for Research and Care on Mental Retardation and Brain Ageing, Troina, 2 Chemical Science, University of Catania, Catania, Italy and 4 Neuroscience and Department of Anatomy and Physiology, Laval University, Quebec, Canada Summary Bidirectional communication between the neuroendocrine and immune systems during ontogeny plays a pivotal role in programming the development of neuroendocrine and immune responses in adult life. Signals generated by the hypothalamic–pituitary–gonadal axis (i.e. luteinizing hormone-releasing hormone, LHRH, and sex steroids), and by the hypothalamic–pituitary–adrenocortical axis (glucocorticoids (GC)), are major players coordinating the development of immune system function. Conversely, products generated by immune system acti- vation exert a powerful and long-lasting regulation on neuroendocrine axes activity. The neuroendocrine–immune system is very sensitive to preperinatal experiences, including hormonal manipulations and immune challenges, which may influence the future predisposition to several disease entities. We review our work on the ongoing mutual regulation of neuroendocrine and immune cell activities, both at a cellular and molecular level. In the central nervous system, one chief compartment is represented by the astroglial cell and its mediators. Hence, neuron–glial signalling cascades dictate major changes in response to hormonal manipulations and pro-inflammatory triggers. The interplay between LHRH, sex steroids, GC and pro-inflammatory mediators in some physiological and patho- logical states, together with the potential clinical implications of these findings, are summarized. The overall study highlights the plasticity of this intersystem cross-talk for pharmacological targeting with drugs acting at the neuroendocrine–immune interface. Key words: corticotropin releasing factor, gender, hypothalamic–pituitary–adrenocortical (HPA) axis, hypothalamic– pituitary–gonadal (HPG) axis, luteinizing hormone-releasing hormone (LHRH), neuron–glial interactions, stress. Correspondence: Prof. B Marchetti, Professor of Pharmacology, Medical School, University of Sassari and Laboratory Head, Neuropharmacology, OASI Institute for Research and Care on Mental Retardation and Brain Ageing, Via Conte Ruggero 73, 94018 Troina (EN), Italy. Email: [email protected] Received 17 May 2001; accepted 17 May 2001.
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Neuroendocrine–immune (NEI) circuitry from neuron–glial interactions to function: Focus on...

Introduction
Substantial evidence from several experimental and clinicalstudies strongly indicates that bidirectional communicationbetween the neuroendocrine and immune systems representsone cardinal feature of a living organism, both in healthy anddiseased states. Indeed, since the early studies of Pierpaoli,Sorkin and Besedovsky,1–4 evidence accumulated in the lastdecades has clearly documented the vital importance of interacting neuroendocrine–immune (NEI) networks in theregulation of physiological homeostatic mechanisms, in particular with regard to reproductive functions.5–16 Theconcept of integrated bidirectionally regulated NEI axes isunderscored during preperinatal life, when exposure toabnormal hormonal levels or to endotoxin results in major
alterations in the programming of both neuroendocrine andimmunological regulatory mechanisms.4,13,14,17–23 Indeed, anumber of experimental and epidemiological studies clearlysuggest that early events occurring in fetal life powerfullymodulate an individual’s predisposition to different patho-logic states in the adult life.21–23
Two major endocrine axes dramatically impinge on neuro-immunomodulation: the hypothalamic–pituitary–gonadal(HPG) and the hypothalamic–pituitary–adrenocortical (HPA)axes. The brain–pituitary–reproductive axis and the brain–thymus–lymphoid axis have been shown to communicate viaan array of internal mechanisms of communication that usesimilar signals (neurotransmitters, peptides, growth factors,hormones) acting on similar recognition targets (the recep-tors).10,11,14,15,24 Moreover, such communication networksform the basis for the control of each step and every level ofreproductive physiology.24 One principal conveying signal isluteinizing hormone-releasing hormone (LHRH), the keyreproductive hormone coordinating the major features ofmammalian reproduction (Fig. 1). The hypothalamic deca-peptide can modulate immune functions directly, through
Immunology and Cell Biology (2001) 79, 400–417
Special Feature
Neuroendocrine–immune (NEI) circuitry from neuron–glialinteractions to function: Focus on gender and HPA–HPGinteractions on early programming of the NEI system
MC MORALE, 1 F GALLO, 1 C TIROLO, 1 N TESTA, 1 S CANIGLIA, 1 N MARLETTA, 2
V SPINA-PURRELLO, 2 R AVOLA, 2 F CAUCCI, 3 P TOMASI, 3 G DELITALA, 3
N BARDEN 4 and B MARCHETTI 1 , 5
Departments of 5Pharmacology and 3Endocrinology, Medical School, University of Sassari, Sassari, 1Departmentof Neuropharmacology, OASI Institute (IRCCS) for Research and Care on Mental Retardation and Brain Ageing,Troina, 2Chemical Science, University of Catania, Catania, Italy and 4Neuroscience and Department of Anatomyand Physiology, Laval University, Quebec, Canada
Summary Bidirectional communication between the neuroendocrine and immune systems during ontogeny playsa pivotal role in programming the development of neuroendocrine and immune responses in adult life. Signals generated by the hypothalamic–pituitary–gonadal axis (i.e. luteinizing hormone-releasing hormone, LHRH, andsex steroids), and by the hypothalamic–pituitary–adrenocortical axis (glucocorticoids (GC)), are major playerscoordinating the development of immune system function. Conversely, products generated by immune system acti-vation exert a powerful and long-lasting regulation on neuroendocrine axes activity. The neuroendocrine–immunesystem is very sensitive to preperinatal experiences, including hormonal manipulations and immune challenges,which may influence the future predisposition to several disease entities. We review our work on the ongoingmutual regulation of neuroendocrine and immune cell activities, both at a cellular and molecular level. In the centralnervous system, one chief compartment is represented by the astroglial cell and its mediators. Hence, neuron–glialsignalling cascades dictate major changes in response to hormonal manipulations and pro-inflammatory triggers.The interplay between LHRH, sex steroids, GC and pro-inflammatory mediators in some physiological and patho-logical states, together with the potential clinical implications of these findings, are summarized. The overall studyhighlights the plasticity of this intersystem cross-talk for pharmacological targeting with drugs acting at the neuroendocrine–immune interface.
Key words: corticotropin releasing factor, gender, hypothalamic–pituitary–adrenocortical (HPA) axis, hypothalamic–pituitary–gonadal (HPG) axis, luteinizing hormone-releasing hormone (LHRH), neuron–glial interactions, stress.
Correspondence: Prof. B Marchetti, Professor of Pharmacology,Medical School, University of Sassari and Laboratory Head, Neuropharmacology, OASI Institute for Research and Care onMental Retardation and Brain Ageing, Via Conte Ruggero 73, 94018Troina (EN), Italy. Email: [email protected]
Received 17 May 2001; accepted 17 May 2001.

local paracrine–autocrine effects in immune cells, and indi-rectly through activation of the HPG axis. Thus, neonatalmanipulations of HPG axis functionality during the firstweek of life result in long-lasting influences in the develop-ment of T- and B-cell dependent functions in adult animals,whereas immune system activation during neonatal life haslong-lasting consequences for reproductive physiology.25
Based primarily on the pioneering work of H Selye,26,27 inter-actions between stress and immune responses have beenclearly demonstrated to have a major impact in the coordina-tion of general homeostatic mechanisms.26–31 Indeed, the HPAaxis is subjected to programming by early life events, andearly life exposure to endotoxin can alter HPA function andpredisposition to inflammation.17,18,20,27 In this context, bidirectional communication between the neuroendocrine andimmune systems represents a critical level modulating thesusceptibility to certain inflammatory, autoimmune and
degenerative disorders of the central nervous system(CNS).31–38 Signals generated by the HPG axis, such as oestro-gens, also remarkably interact at different levels of the stressaxis. Thus, a complex interplay between genotype, the circu-lating gonadal and adrenal hormones, the intrinsic capabilityto respond to stressful stimuli, coupled with a specificimmunological setting, may contribute to the modulation ofan individual’s predisposition to several disease entities.
In both medicine and, especially, psychiatry, the gender-differentiated predispositions to a number of illnessses arehistorically exemplified by the difference in the prevalence ofdepression in women, the vulnerability associated with oralcontraceptives, abortion, the premenstrual period, puerpuerium,and menopause.39 By acting on the neuroendocrine andimmune systems during the stress response, life events maygreatly affect homeostasis and favour the appearance ofdisease. One of the best examples of this psychoimmune–
Neuroendocrine–immune circuitry 401
Figure 1 Schematic representa-tion of the possible interactionsbetween the hypothalamus–hypophyseal–gonadal axis and thethymus, with luteinizing hormone-releasing hormone (LHRH)serving as a major channel ofcommunication. HypothalamicLHRH governs the release of thepituitary gonadotropins luteiniz-ing hormone (LH) and follicle-stimulating hormone (FSH),responsible for gonadal produc-tion of the sex steroids. Thegonadal hormones in turn, feedback information to the thymusand hypothalamus. At the thymuslevel, sex steroids act on specificreceptors present on the reticulo-epithelial matrix, and induce bothup and downregulation of targetgenes involved in the control of T-cell responses. In contrast, thesex steroid background alters theproduction of thymic peptides(thymosins) and neuropeptides,such as LHRH, with autocrine/paracrine regulatory influencewithin the thymic microenviron-ment. The direct neural pathwaysinnervating immune and endocrineorgans together with the modula-tory influence of glucocorticoidsand catecholamines are also indicated.

neuroendocrine circuit was revealed by Galen, in approxi-mately 200 AD, when he observed increased susceptibility tobreast cancer in melancholic women, compared to sanguinewomen. If women are currently thought of as having a higherrisk than men for developing certain psychiatric, and mostautoimmune, disorders, they also seem to be more resistant tothe development of a number of neurological diseases,including Alzheimer’s disease.40–44 Much experimental liter-ature has established that the gonadal hormones are respon-sible not only for the sexual differentiation of neural circuitry,which mediates a variety of reproductive behaviours andphysiological mechanisms, but also for the generation of sexually driven immunological dimorphisms, as well as thesex-linked differential response of the HPA axis. Interest-ingly, the female hormones, oestrogens, have receivedincreased attention and consideration thanks to the apprecia-tion of their ‘beneficial/buffering’ (in a word neuroprotec-tive) mechanisms.40–45 The precise underlying biochemicaland molecular mechanism(s) are far from being completelydisclosed. Besides others, the crucial interaction of this classof hormones at the neuroendocrine–immune interface can beeasily anticipated. A key player, the brain astrocytic compart-ment, must be added to this scenario. Indeed, astrocytes areelements of the CNS that share with the neuronal, endocrineand immune cells similar recognition and transduction capabilities, and that represent key entities during the development of the CNS in the adult brain as well as duringageing and ageing-associated pathologies, such as neuro-degenerative disorders.46–49 Thus, it is not surprising that theHPG (and specifically LHRH and oestrogens) and HPA (inparticular glucocorticoid (GC)) axes, powerfully and dynam-ically interact with the astroglial compartment, realizing afurther level of NEI integration.46,47
In this paper we will review our work on neuroendocrineimmunomodulation with a major focus on LHRH, sexsteroids and interactions between the HPG and HPA axes inimmune system regulation. First, the bidirectional communi-cation between LHRH neurons and glial cells and the influ-ence of oestrogens and glucocorticoids in modulating theactivity of these key CNS elements, will be reviewed, with amajor focus on hormonally induced neuron–glial plasticityand the response of the glial compartment to inflammation.Second, the role of LHRH as the main coordinator of theHPG–thymic axis, in different physiological/pathologicalstates is summarized, with a particular emphasis on the mol-ecular mechanisms involved in HPG–HPA interactions inmediating sex differences in immunological responses.Finally, the impact of a dysfunctional HPA system in a trans-genic mouse model expressing a glucocorticoid receptor(GR) antisense RNA, for the programming of NEI functionswill be briefly summarized.
Astroglial cell compartment as a first level ofregulation: Glia is a source and a target ofneuroendocrine and immunomodulators
An extraordinary body of evidence has now been assembled bydifferent investigators from all fields of neuroscience, sup-porting a key role for the glial cell in neuronal physiopathology.Indeed, nearly a century and a half since the development ofknowledge about neuron–glia interactions,50–52 the possibility
of signals passing from neurons to glial cells, and thus toother neurons opens up many scenarios for intercellular/intra-cellular cross-talk within single cells of the CNS (Fig. 2). Thefunctioning of the nervous system depends on a continousand sophisticated interrelationship between neuronal and glialcells. There are two broad subgroups of glial cells: the macro-glia, which consist of astrocytes, oligodendrocytes and epen-dymal cells, and the microglia. In recent years, an array ofneurotransmitters, receptors, ion channels, adhesion mole-cules and trophic factors have been revealed to be associatedwith glial cells.52 An insight into the impact of hormones generated by the HPG and HPA axes in neuron–astroglialsignalling is presented.
Luteinizing hormone-releasing hormone, sex steroidsand glucocorticoids as mediators of neuron–gliaplasticity: Implications for neuroendocrine–immunecommunications
Luteinizing hormone-releasing hormone, a decapeptide manufactured by highly specialized neuroendocrine cells, isthe key regulator of the hypothalamic–hypophyseal–gonadalaxis and is essential for reproductive competence53,54 (Fig. 1).There is a unique functional interplay between the LHRHneuronal system and the immune ‘world’, reflected at theCNS level, by a bidirectional interaction between the differ-entiating LHRH neuron and the maturing astroglial cell. Glialcells play active roles from embryonic development to adult-hood. In particular, astrocytes participate in several aspects ofneuronal growth and differentiation both by providingcell–cell contacts, and by secreting neuronal growth factors.52
In turn neurons influence the cellular behaviour of astro-cytes by secreting substances in the microenvironment.52
Increasing evidence clearly supports the notion that
MC Morale et al.402
Figure 2 Relationship between astrocytes and other brain elements. The schematic drawing illustrates several possible contacts between the astrocyte and (1) a synaptic cleft; (2) otherastrocyte networks; (3) capillary/blood vessels; (4) neuronal cellbodies; (5) node of Ranvier.
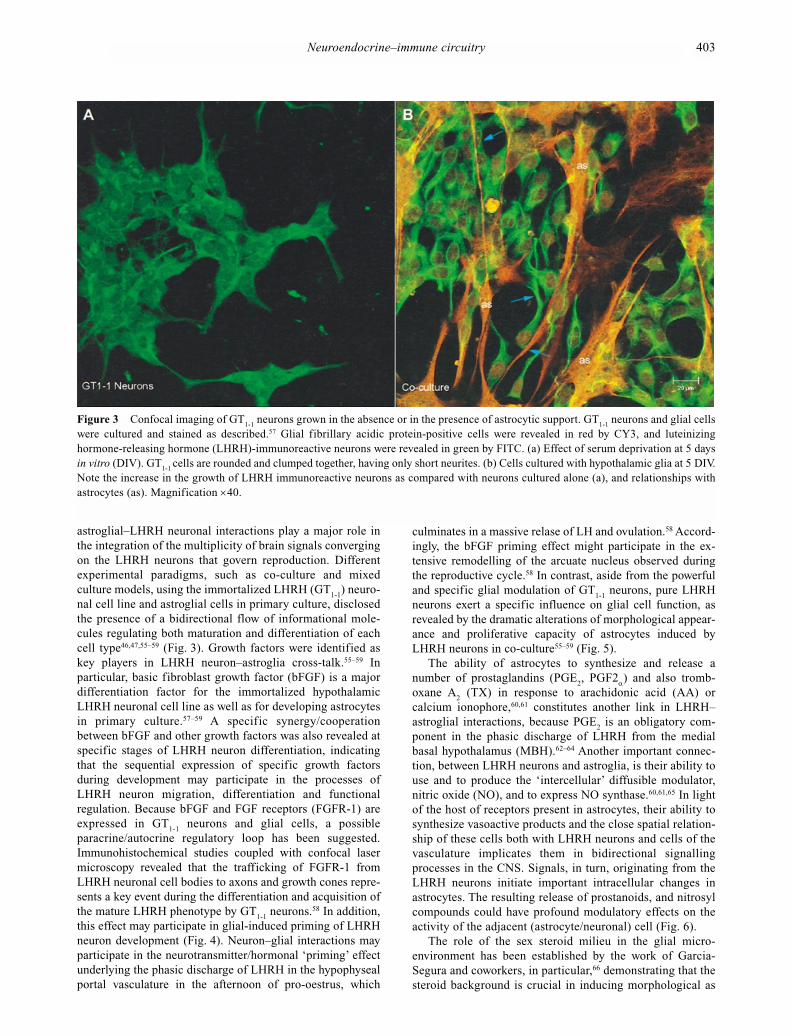
astroglial–LHRH neuronal interactions play a major role inthe integration of the multiplicity of brain signals convergingon the LHRH neurons that govern reproduction. Differentexperimental paradigms, such as co-culture and mixedculture models, using the immortalized LHRH (GT
1-1) neuro-
nal cell line and astroglial cells in primary culture, disclosedthe presence of a bidirectional flow of informational mole-cules regulating both maturation and differentiation of eachcell type46,47,55–59 (Fig. 3). Growth factors were identified as key players in LHRH neuron–astroglia cross-talk.55–59 In particular, basic fibroblast growth factor (bFGF) is a majordifferentiation factor for the immortalized hypothalamicLHRH neuronal cell line as well as for developing astrocytesin primary culture.57–59 A specific synergy/cooperationbetween bFGF and other growth factors was also revealed atspecific stages of LHRH neuron differentiation, indicatingthat the sequential expression of specific growth factorsduring development may participate in the processes ofLHRH neuron migration, differentiation and functional regulation. Because bFGF and FGF receptors (FGFR-1) areexpressed in GT
1-1neurons and glial cells, a possible
paracrine/autocrine regulatory loop has been suggested.Immunohistochemical studies coupled with confocal lasermicroscopy revealed that the trafficking of FGFR-1 fromLHRH neuronal cell bodies to axons and growth cones repre-sents a key event during the differentiation and acquisition ofthe mature LHRH phenotype by GT
1-1neurons.58 In addition,
this effect may participate in glial-induced priming of LHRHneuron development (Fig. 4). Neuron–glial interactions mayparticipate in the neurotransmitter/hormonal ‘priming’ effectunderlying the phasic discharge of LHRH in the hypophysealportal vasculature in the afternoon of pro-oestrus, which
culminates in a massive relase of LH and ovulation.58 Accord-ingly, the bFGF priming effect might participate in the ex-tensive remodelling of the arcuate nucleus observed duringthe reproductive cycle.58 In contrast, aside from the powerfuland specific glial modulation of GT
1-1neurons, pure LHRH
neurons exert a specific influence on glial cell function, asrevealed by the dramatic alterations of morphological appear-ance and proliferative capacity of astrocytes induced byLHRH neurons in co-culture55–59 (Fig. 5).
The ability of astrocytes to synthesize and release anumber of prostaglandins (PGE
2, PGF2α) and also tromb-
oxane A2
(TX) in response to arachidonic acid (AA) orcalcium ionophore,60,61 constitutes another link in LHRH–astroglial interactions, because PGE
2is an obligatory com-
ponent in the phasic discharge of LHRH from the medialbasal hypothalamus (MBH).62–64 Another important connec-tion, between LHRH neurons and astroglia, is their ability touse and to produce the ‘intercellular’ diffusible modulator,nitric oxide (NO), and to express NO synthase.60,61,65 In lightof the host of receptors present in astrocytes, their ability tosynthesize vasoactive products and the close spatial relation-ship of these cells both with LHRH neurons and cells of thevasculature implicates them in bidirectional signallingprocesses in the CNS. Signals, in turn, originating from theLHRH neurons initiate important intracellular changes inastrocytes. The resulting release of prostanoids, and nitrosylcompounds could have profound modulatory effects on theactivity of the adjacent (astrocyte/neuronal) cell (Fig. 6).
The role of the sex steroid milieu in the glial micro-environment has been established by the work of Garcia-Segura and coworkers, in particular,66 demonstrating that thesteroid background is crucial in inducing morphological as
Neuroendocrine–immune circuitry 403
Figure 3 Confocal imaging of GT1-1
neurons grown in the absence or in the presence of astrocytic support. GT1-1
neurons and glial cellswere cultured and stained as described.57 Glial fibrillary acidic protein-positive cells were revealed in red by CY3, and luteinizinghormone-releasing hormone (LHRH)-immunoreactive neurons were revealed in green by FITC. (a) Effect of serum deprivation at 5 daysin vitro (DIV). GT
1-1 cells are rounded and clumped together, having only short neurites. (b) Cells cultured with hypothalamic glia at 5 DIV.
Note the increase in the growth of LHRH immunoreactive neurons as compared with neurons cultured alone (a), and relationships withastrocytes (as). Magnification ×40.

well as functional changes in the astroglial cell compartment.In response to oestrogens, astrocytes appear to participate inthe remodelling of synaptic contacts on hypothalamic neuronsthat control the release of pituitary secretions in rodents andprimates.66 This work is substantiated by findings indicatingthat the morphology, immunoreactivity, enzymatic activity,and gene expression of astroglia are sexually dimorphic inseveral brain areas and/or are modified by different in vivo/invitro experimental manipulations. Glial cells have been shownto harbour receptors for oestradiol and progesterone,67 andoestradiol is able to induce the appearance of progesteronereceptors.68 In particular, oligodendrocytes, are known to becapable of synthesizing steroids such as pregnenolone andprogesterone, and evidence has been presented for the
presence of receptors for these hormones on cultured cells.67
Astrocytes were found to possess very few progesteronereceptors (PR); those found were confined to cells derivedfrom female animals. In contrast, oligodendrocytes preparedfrom both male and female animals possessed PR and,although more abundant in cultures from females, receptors incells from both sexes were increased by exposure to oestro-gens. Because the effect of oestrogen on astroglial cells canvary according to the specific CNS region, the sex of theanimal and the stage of glia/neuron maturation and the presence of growth factors, the resulting response of the neuronal compartment is dictated by the specific changes ofthe microenvironment in a specific physiological/pathologicalcondition.68
MC Morale et al.404
Figure 4 Effect of basic fibroblast growth factor (bFGF) priming on distribution of fibroblast growth factor receptor-1 (FGFR-1) (a,c)and neurofilament fluorescence in GT
1-1(b,d) neurons following growth factor treatment. Cells were double-labelled and revealed as
described.58 Fibroblast growth factor receptor-1 immunoreactivity was visualized at 3 days in vitro (DIV) by confocal laser microscopy inGT
1-1neuronal cultures after bFGF priming (3.5 ng/mL) alone (a,b) and 24 h after application of insulin-like growth factor I
(IGF-I) (c,d). At 3 DIV 12 h after bFGF priming, FGFR-1 immunofluorescence is evident though the soma, neurites and growth cones ofNF-immunolabeled cells (a). Note ‘hot spots’ of FGFR-1 at the growth cone endings (a). When IGF-I is applied 12 h after bFGF priming,FGFR-1 immunofluorescent reaction is increased throught the soma, neurites and growth cones. Note the neuronal growth cones and extensive neurite extension after bFGF + IGF-I treatment (white and blue arrows). Magnification ×20. Scale bar 10 µm.

There is abundant evidence that cultured glia possess corticosteroid receptors.69–71 Adrenal steroids activate twoclasses of intracellular receptors, the mineralcorticoid (MR)or type I receptor, and the GR or type II receptor.69 Ligandbinding studies have demonstrated the presence of a singlepopulation of GR in both astrocytes and oligodendrocytes.69,70
Glucocorticoids are known to modulate the expression of avariety of glial proteins, including glial fibrillary acidicprotein (GFAP), glutamine synthetase (GS), myelin basicprotein (MBP), and glycerol phosphate dehydrogenase.69,70
Using an in vitro model of developing neonatal rat glial cells,we studied developmental expression of GR as a function of
Neuroendocrine–immune circuitry 405
Figure 5 Confocal imaging of astroglial cells grown in the absence or in the presence of GT1-1
neurons. Astroglial cells cultured (a) aloneand (b) in the presence of luteinizing hormone-releasing hormone (LHRH) neurons treated as described57–59 and immunostained for glialfibrillary acidic protein (GFAP), revealed by FITC (in green). (a) Astrocytes at 14 days in vitro (DIV). Glial fibrillary acidic protein-positive cells are flat and poligonal. (b) Glial cells co-cultured with GT
1-1 neurons. Note the characteristic process bearing morphology of
GFAP-immunoreactive cells (blue arrows). Magnification ×40.
Figure 6 Dynamic interactionbetween the astroglial cell com-partment, the endothelial and theluteinizing hormone-releasinghormone (LHRH) neurons. Onselective stimulation astrocytesmay release products able to alterthe vascular endothelium. Theexpression of receptors on astro-cytes, their ability to synthesizevasoactive products, and the closespatial relationships of these cellsboth with neurons and cells of thevasculature implicate astroglialcells in bi-directional signallingprocesses in the central nervoussystem. (PG), prostaglandin; (PAF),platelet activating factor; (TXA
2),
tromboxane A2; (NO), nitric oxide;
(ADRF (NO)), astrocyte-derived(vaso)-relaxing factor (nitricoxide). (ET-1), endothelin-1; (LT),leukotrienes.

MC Morale et al.406
time in culture and found low levels of GR mRNA expressedat 8 days in vitro (DIV) with a progressive increase between12 and 20 DIV, reaching a plateau thereafter, with the mRNAremaining elevated up to 50 DIV.71 Astroglia respond to LPSand dexamethazone with a strong morphological effect.Astrocytes assume a stellate shape and extend processes(Fig. 7). Activation of the astrocyte compartment by LPSboth in vitro and in vivo results in massive production of NO,whereas GC are potent suppressors of inducible NO synthase(iNOS) activity, thus resulting in decreased levels of thisimmunomodulator.38 In the intact brain, glial cells have beenshown to respond to GC, and GFAP mRNA levels are undertranscriptional control by adrenal hormones after acute orchronic treatments both in vivo and in vitro.72–74 Thus, asophisticated circuitry involving the stress hormones and theresponse of the astroglial cell compartment underscores theageing process, both in healthy and diseased states.72
From the presented information, the dynamics of thecross-talk between neurons and astrocytes appears to be verycomplex. It should be noted that neurons and glial cells arelikely to be exposed to a number of different extracellular signalling molecules that may vary from moment to moment,and as a function of the particular physiological status (e.g.sex, age, stressful situation). Such sophisticated regulatorynetwork is likely to orchestrate the final CNS response in a number of physiopathological conditions, and thus play amajor role in NEI communications. The multiple interactionsbetween endogenous or exogenous GC, growth factors, thesex steroid background and inflammatory mediators pro-duced by each glial cell compartment set into motion duringbrain inflammation, may thus have important consequencesfor neural cell regulation.
Luteinizing hormone-releasing hormone orchestratesthe hypothalamic–pituitary–gonadal–thymic axis
The exquisite interplay between the LHRH neuronal systemand immune homeostasis is well reflected by the strong interaction linking reproductive and immune functions. Thephysiological development of an operative HPG axis neces-sitates the presence of an intact immune system and normalimmune function, because immunosuppressed or incompe-tent animals show numerous reproductive disorders.75–81
Moreover, inflammatory and infectious diseases often co-incide with changes in reproductive functions including adecline in fertility, an increased incidence of spontaneousabortion, and full-term birth of abnormal progeny. In con-trast, the reproductive immune interactions are of majorimportance during carcinogen-induced mammary tumours,as revealed by the powerful impact at the level of the thymusgland and the cell-mediated and humoral responses duringmammary carcinogenesis.80 Of major importance, the con-comitant treatment with thymic hormones (thymosin beta 4)and a time-specific schedule, results in significant normal-ization of a number of neuroendocrine parameters andinduces a significant regression of tumour growth.80 Indeed,the primary communication between the immune and thereproductive systems is known to involve the thymus and itspeptide secretion, it seems important to place the LHRHeffects within the context of an hypothalamic–pituitary–gonadal–thymic axis (HPGT). Communication between the
Figure 7 Confocal imaging of astroglial cells grown in theabsence or the presence of LPS or dexamethazone (DEX).Astroglial cells cultured for 14 days in vitro (DIV) were sub-jected to serum deprivation for 24 h, treated subsequently withserum albumin (BSA, A), or LPS (100 ng/mL) or dexametha-zone (DEX, 10–10 mol/L) for 24 h, and immunostained for glialfibrillary acidic protein (GFAP) (revealed by FITC, in green) andBrdU (revealed by Cy3, in red).57 (a) Astrocytes at 14 DIV. Glial fibrillary acidic protein-positive cells are flat and poligonal.Only one GFAP-positive cell incorporates BrdU (white arrow).Glial cells cultured in the presence of (b) LPS or (c) DEX. NoteBrdU incorporation within the nuclei of GFAP-positive cells.Also note the acquisition of the process bearing morphology of GFAP-immunoreactive cells (blue arrows). Magnification×40.

gonadal axis and the lymphoid organs has been proposed forover a century,81 and the studies of Grossman and cowork-ers76–78 have even emphasized the existence of a reciprocalrelationship between the HPG and the brain–thymus–lymphoid axis. A schematic representation of the possibleinteractions between the HPG axis and the thymus, withLHRH serving as a primary channel for communication isgiven in Fig. 1. As observed, a bidirectional network carryinginformation to both the immune and the neuroendocrinereproductive systems, via LHRH, is depicted. Direct aminer-gic and peptidergic innervations of both the gonads and theimmune organs are also illustrated. While hypophyseal andgonadal hormones feedback information to the thymic cell,providing a modulatory system and regulating thymic cellmaturation and thymic peptide production, the thymus and itspeptide secretion can modulate gonadotropin secretion via adirect action at the hypothalamic LHRH neuronal level.
Luteinizing hormone-releasing hormone and LHRHreceptors in immune organs: Presence of aparacrine–autocrine regulatory loop
Specific LHRH binding sites are present in mouse blood lymphocytes,82 in rat mast cells,83 thymocyte and splenocyte84
cultures, as well as in cultured porcine lymphocytes.85 Inaddition, LHRH mRNA and/or LHRH-like molecules havebeen identified in rat thymus, in thymocyte and splenocytecultures and in human peripheral T lymphocytes.86–88 More-over, by synthesizing and sequencing the rat thymus, LHRHthymocyte and hypothalamic LHRH are shown to be identi-cal, with the sequence data obtained 5′ and 3′ to the openreading frame being almost identical to hypothalamic LHRHmRNA.86 Not only is LHRH mRNA expressed in the primaryimmune organ, but it also appears to be developmentally regulated.86 Using the rat immature T-cell line Nb2, Wilsonet al.89 have reported that the LHRH gene is regulated by pro-lactin (PRL), at various times during the cell cycle. Moreover,an alternatively spliced LHRH mRNA exists in Nb2 cells andmay produce a new truncated gonadotropin-associatedpeptide (GAP).89 Finally, the SH gene found on the oppositestrand of the LHRH gene, is expressed in lymphocytes at thesame time and in the same manner as the LHRH gene.89
Messenger RNA transcripts coding for the LHRH receptor(LHRH-R) were demonstrated to be present in immuno-logically com-petent cells. Using reverse transcription–polymerase chain reaction (RT–PCR) and RNA from Nb2cells the LHRH-R PCR fragment was sequenced and clonedand the DNA sequence shown to be identical to that pre-viously described. In analogy to what was observed at theanterior pituitary level for the LHRH-R protein,90 in Nb2cells the LHRH receptor mRNA was shown to drasticallydecrease immediately after PRL administration,89 suggestingpossible paracrine regulation between the two hormoneswithin immune cells.
Luteinizing hormone-releasing hormone coordinatesthe hormonal–immune network during pregnancy andthe oestrous cycle
The powerful action of LHRH in setting the hormonal homeo-stasis to high circulating levels of sex steroid hormones
during pregnancy as well as the dynamic alterations in theconcentration of prolactin, cortisol and other hormonalfactors may clearly modify immune responsiveness. Theincrease in sex steroids that takes place in all mammalianspecies during pregnancy may potentially assist in preventingthe maternal–fetal rejection response.15 If this rejectionresponse is not depressed, termination of pregnancy may takeplace before completion of term. However, in most pregnantwomen, the cell-mediated immune response is markedlydepressed and pregnancy is maintained.15,44 This can alsoaccount for the observation that human skin homograftssurvive longer on pregnant hosts than on non-pregnant hosts.A number of studies clearly indicate that progesterone par-ticipates in the regulation of the immune response duringpregnancy. As to the mechanisms of action of sex steroids inmodulating immune responsiveness, the presence of recep-tors for oestrogens and testosterone on the reticulo-epithelialmatrix of the thymus and peripheral lymphocytes44,78 arguesin favour of a direct effect of these steroids at the level ofimmune organs and cells.
Specifically, intrathymic LHRH system activity variesaccording to the phases of the oestrous cycle, but moreimportantly a sexual dimorphic pattern of LHRH synthesisaccompanies the sex-dimorphic immune response duringontogeny and cyclicity.15,44 Within the thymus, LHRH mRNAconcentration exhibits clear sex-dependent fluctuation duringthe rat oestrous cycle, which is accompanied by a change inthymocyte sensitivity to LHRH with a maximal response tothe natural decapeptide in pro-oestrus. It is likely that thefluctuation of LHRH mRNA observed in the thymus mayreflect the interplay of a number of endocrine influences onthymocytes. The fact that an LHRH-antisense is able tocounteract the mitogen-induced lymphocyte proliferationclearly underlines a physiological paracrine/autocrine role ofintralymphocytic LHRH in the control of lymphocyte respon-siveness.43,44 The dramatic decrease in both rodent thymocyteand human peripheral blood lymphocyte response to T-depen-dent mitogens after ovulation coupled with the diminishedlymphocyte response to LHRH in that particular reproductivephase would indicate a possible participation of LHRH inthat phenomenon and that sex steroids are capable of modu-lating that effect. Such a mechanism is then likely to partici-pate in the reduction of immune responsiveness observedduring pregnancy.15,43,44 It seems tempting to speculate thatsex steroids altering the expression of target genes at theimmune system level may modify the responsiveness of theimmune cell to different hormones and neurotransmitters.Indeed, the ability of oestrogens to directly alter LHRH geneexpression has been demonstrated in the placenta.91
Sympathetic nervous system activity through ββ-adrenergic receptor gene expression is involved inimmunological sex-dimorphism
Using in vitro autoradiography, we demonstrated the presenceof β
2-adrenergic receptors (β
2AR) in the rat thymus.92,93 In the
thymus, norepinephrine (NE) could, therefore, act via theβ
2AR both as a paracrine hormone available to receptors on
thymic cells, and as a localized transmitter in nerve terminalsthat directly contact cortical thymocytes, mast cells andeosinophils to modulate immune functions. The thymic
Neuroendocrine–immune circuitry 407

β2AR are preferentially found in the medullar compartment
of the gland and show a clear sexual dimorphism in receptororganization during sexual maturation.93 The expression of aβ
2AR in the rat thymus was further confirmed by the pres-
ence in the thymic tissue of a mRNA species of 2.3 kb, whichspecifically hybridized with a cDNA encoding the full codingsequence of the human β
2AR.94,95 Interestingly, the β
2AR-
stimulated adenylyl cyclase is markedly modulated by the sexsteroid hormone background.95 Indeed, the β
2AR observed in
thymic tissue preparations is functionally coupled to theadenylyl cyclase system, with a sensitivity characteristic of aβ
2subtype receptor. Both a high affinity state of the receptor
for isoproterenol and an isoproterenol-stimulated adenylylcyclase activity could be detected in the thymus membranepreparations, with the guanine nucleotide converting all ofthe high-affinity state receptors into low affinity state recep-tors.95 Furthermore, an almost 70% loss of the enzyme inresponse to GTP followed castration, suggesting that ovariansteroids may exert potent effects on the β
2AR signalling
pathway.95 Consistent with previous results obtained in othertissues, parallel changes in both β
2AR density and β
2AR
mRNA levels followed the hormonal changes associated withthe rat oestrous cycle, pregnancy or castration (Fig. 8). Suchquantitative changes suggest a subtype-specific hormonalregulation of the β
2AR population.95 The results on the sex
steroid-dependent modulation of β2AR-stimulated cAMPaccumulation and proliferation in rat thymus are consistentwith other data obtained in the mammary gland, where theGTP-stimulated adenylate cyclase showed the highest activ-ity during pregnancy.96 Although the mechanism throughwhich β-adrenergic stimulation alters thymocyte proliferationis unknown, one attractive possibility involves interaction atthe level of second messengers. In fact, in T cells, one earlytransmembrane signal in response to stimulation of the T-cellreceptor is the activation of phospholipase C, causing therelease of inositol phosphates and calcium mobilization frominternal stores and translocation of protein kinase C (PKC).These signals are associated with IL-2 receptor α-chainexpression and production of cytokines, leading to cell pro-liferation.97 Moreover, the membrane translocation and acti-vation of Ca2+-phospholipid-dependent PKC is accompaniedby a dramatic decrease in intracellular cAMP. Conversely,stimulation of adenylyl cyclase is known to provideinhibitory signals for T-cell proliferation. In mature T-cellsIL-2 is inducible via agonists of the phospho-inositidepathway, resulting in a series of cascading interactions inducing cell proliferation.94,95,97
Hypothalamic–pituitary–gonadal–hypothalamic–pituitary–adrenocortical interactionsand immunological sex-dimorphism
Glucocorticoid hormones are crucial hormones in the controlof immunity, being among the most potent anti-inflamma-tory, anti-allergic and immunosuppressive agents known, andthey act in a very complex way, at various steps of theimmune response.98–100 The effects of corticosteroids on cellsof the immune system, as in other corticosteroid responsivecells, are mediated through both soluble and nuclear GR.98–100
Oestrogens and glucocorticoids have been shown to influenceGR mRNA concentrations in the rat brain and pituitary.
Similar control mechanisms in the thymus could have impor-tant implications on the regulatory influence of glucocorti-coids on immune functions. One of the major immunecompartments where glucocorticoids exert their effects isrepresented by the thymus gland. Since the observation thatglucocorticoids induce atrophy of the cortical area of thethymus there have been many studies on the morphologicalchanges and cellular events that lead to loss of cell viability.The dramatic alterations of thymus architecture and functionaccompanying sexual maturation, clearly indicate a possiblerole for sex steroids in the processes of thymic-programmedcell death.101 To clarify this issue we studied changes in GRgene expression under physiological as well as pharmacolog-ical conditions accompanied with marked variations in thesex steroid hormonal milieu, and correlated such changeswith alterations in immune responsiveness. Our results haveclearly indicated that thymic GR mRNA concentration is
MC Morale et al.408
Figure 8 Hormonal regulation of β2-adrenergic receptor (β
2AR)
concentration and mRNA content. Poly(A)+ RNA samples fromthe different experimental groups were fractionated by electro-phoresis on a 1.2% agarose–7% formaldehyde gel, blotted ontonylon filters and probed with 32P-labelled β-adrenergic receptorcDNA (2.5 × 106 dpm/mL) or actin cDNA (2.0 × 106 dpm/mL).95
The relative abundance of the β2AR mRNA are expressed as a
percentage of their values (β2AR mRNA/β-actin mRNA ratios)
relative to di-oestrous 1 (D1). Control is normalized to 100%.Results are the mean ± SEM of 4–6 independent determinations.Representative experiment showing the strong stimulation ofβ
2AR at pro-oestrous (P) compared to di-oestrous phases of the
cycle. Also note the sharp increase in mRNA concentration in sexsteroid treated castrated (OVX) and pregnant (PREG) rats com-pared to oestrus (E) or OVX animals.95

under the control of gonadal and adrenal hormones.102 Weobserved an upregulation of GR mRNA content during theluteal (i.e. oestrous) phase, and a down-modulation of GRtranscript on pro-oestrus, corresponding to the phase ofmaximal oestrogenic stimulation.102 The hormonal sensitivityof type II GR in the thymus is further substantiated by thesharp decrease in GR transcript during pregnancy, while thehormonal milieu of lactation leads to a normalization of GRmRNA concentration to levels measured in di-oestrous rats.It is therefore possible that the hormonal milieu of lactationstimulates GR transcripts within the thymus, and that theoestradiol to progesterone ratio may be critical in the regula-tion of type II GR mRNA.102 Further support for a sexsteroidal modulation of thymic GR gene expression wasgiven by the sharp increase of GR transcript levels after
castration, and the specific reversal of the castration-inducedrise measured after a concomitant treatment with oestrogens(Fig. 8). In addition, the ability of corticosterone in vitro toinfluence a cell-mediated immune response in the thymus(i.e. the blastogenic transformation of thymocytes) seems todepend on the sex steroid hormone milieu.102 Thus, the hor-monal conditions accompanied by higher GR transcript levelsin the thymus (i.e. OVX, oestrus) are characterized by a highdegree of corticosterone inhibition of T-cell proliferation. Incontrast, the endogenous adrenal tone is able to influence thymocyte proliferation, and treatment with corticosterone ofthymocyte cell cultures from adrenalectomized rats produceda sharp inhibition.102 Of interest, the postnatal developmentof GR expression within the thymus is sexually dimorphic.13
The ability of corticosterone to directly inhibit both GR
Neuroendocrine–immune circuitry 409
Figure 9 Receptors coupled to intracellular signalling systems in T lymphocytes. Intracellular pathways elicited by signalling throughthe T-cell CD3 receptor complex (TCR-CD3); the β
2-adrenergic receptor (β
2AR), the luteinizing hormone-releasing hormone (LHRH)
receptor, and steroid receptors modulating transcription of target receptor genes. The stimulatory pathway (mobilizing membranepolyphosphoinositides) and converging to T-cell proliferation may be activated by either the operation of the LHRH-R or the TCR-CD3complex. Cross-talk between second messenger systems (protein kinase C (PKC) and protein kinase A (PKA) adjusts/downregulates thestimulatory signals. Oestrogens modulate the amplitude/degree of T-cell activation by acting on the LHRH, βAR and type II glucocorti-coid receptor (GR) gene expression, via hormone responsive elements (HRE), resulting in an amplification of inhibitory/stimulatory path-ways. An example of the effect of oestrogens on β
2AR gene expression is illustrated together with interaction of cAMP responsive
elements (CREB) modulating the β2AR cycle. (DAG), diacylglycerol; (G), G protein; (G
S), guanine nucleotide stimulatory; (HSP90), heat
shock protein 90; (PIP2), phosphatidylinositol biphosphate.

transcript levels as well as a cell-mediated immune responsewithin the thymus, and the modulation of such inhibitoryeffects by the sex steroid hormone milieu, may offer anexplanation and a molecular mechanism whereby stress maybe deleterious for reproduction, also via immunomodulation.In contrast, hormonally mediated alterations in immunitymight also have a pathological implication in sexually relatedimmune diseases. For example, in mice and humans, lupuserythematosus is more prevalent in females and oestrogenaccelerates the disease process, while menstruation is knownto exacerbate idiopathic thrombocytopenia purpura. Sex steroidhormone milieu might also have a role in controlling thestress response through immunomodulation. Oestrogens havebeen shown to inhibit IL-6 production in IL-1-stimulatedcells.103,104 Because IL-6 in turn can enhance cortisol levelsby activating the HPA axis,105 it would then follow that the oestrogenic status might influence the glucocorticoid–lymphokine interactions. It seems, therefore, highly possiblethat the degree of susceptibility to, and severity of, inflam-matory diseases in response to a given pro-inflammatorytrigger, may depend on the influence of sex steroids on GRactivity. However, it is possible that the sexual dimorphism of immune system activity is related to differential regulationof the HPA axis activity by oestrogens because sexual dimor-phism has also been described in HPA axis regulation.106–108
A number of studies conducted in the last few decades haveclearly demonstrated that the neuroendocrine response tostress displays profound sex-specific differences, the mani-festation of which largely depends on the presence ofgonadal steroids. Distinct sex differences in the genes encoding corticotropin-releasing hormone (CRH) in the hypo-thalamus and GR in the hippocampus have been identified.Several previous studies have suggested an influence on thegonadal steroid milieu on CRH and and corticosteroid recep-tors. Studies of Patchev and Almeida45 demonstrate thatgonadal setroids exert facilitating effects on the GC-mediatedtranscriptional regulation of CRH and GR genes in the ratbrain. Interestingly, the absence of a GC feedback mechanisminduces a persistent increment of adrenocorticotropichormone (ACTH) secretion, thus overriding the sexual dimorphism in plasma ACTH concentration.109
All together the information presented suggest that oestro-gens may induce, on the one hand, an increase in β
2AR and
LHRH activities in the thymus, resulting in an increased sensitivity of the thymic cell to locally released and/or circulating catecholamines, as well as to endogenousLHRH, not excluding possible intersystems paracrine (i.e. catecholaminergic-LHRH-ergic)/autocrine/intercrinecross-talk (Fig. 9). In contrast, a sharp decrease of both GRtranscripts and thymocyte response to corticosterone followsE
2treatment/exposure. Such oestrogen-induced up or down-
regulation of the β2AR, LHRH and GR mRNA levels in the
thymus may then result in sophisticated control of lympho-cyte sensitivity to endogenous hormones and constitute amolecular mechanism of NEI integration (see Fig. 9).
Luteinizing hormone-releasing hormone and LHRH-agonists as immunological response modifiers
Considerable evidence has been provided that LHRH and its agonists and antagonist analogues are able to markedly
influence immune functions. In adult rats, LHRH and itsanalogues have been shown to increase the absolute and relative thymic weight,110–113 and to directly influencethymus morphology and thymocyte proliferative activity ofhypophysectomized male rats.84,114 More importantly, LHRHagonist (LHRH-A) treatment of ageing rats results in a significant increase in thymus weight, and restoration of the morphological appearance of the gland, and to reversethe ageing-induced impairment of thymocyte proliferativecapacity in response to policlonal T-dependent mito-gens.84,110,112 The biochemical mechanism of action of LHRHafter the binding of the neuropeptide to its receptor presentin the immune cell, appears to be the mobilization ofpolyphosphoinositide hydrolysis, translocation of PKC andupregulation of IL-2 receptor expression.97 The physiologi-cal significance of the intra LHRH system within the rodentthymus has also been investigated.25,116,117 Treatment ofneonatal rats with a very potent LHRH-antagonist resultedin irreversible alterations in a series of immune parametersincluding thymus morphology, cell-mediated and humoralimmune responses.25,116 Human peripheral T-cell subsetsproduce LHRH and its production is increased in T cellsactivated with phytohaemagglutinin (PHA).88 This findingcoupled with the expression of LHRH mRNA in human lymphocytes further supports the idea of LHRH as an important immunomodulator. Luteinizing hormone-releasing hormone-A modulates the expression of murinelupus in a gonadal steroid-independent fashion, because castrated (SWR X NZB) F1 mice treated with LHRH antagonist displayed decreased total IgG and anti-DNA anti-body concentrations, delayed renal disease, and significantlyprolonged survival.118,119 Luteinizing hormone-releasinghormone agonist might act on the immune system directly,by a direct effect on B- or T-lymphocytes. Alternatively, theeffects might be indirect, through a reduction in gonado-tropins or alterations in cytokine production by immunecells.
Thus, the fact that LHRH and its potent analogues are ableto modulate immune system function either independentlythrough effects on sex hormone production, or indirectlythrough modulation of the HPG, is of special interest, as theysupport the possibility that hormones other than gonadalsteroids might contribute to the well-known sex differences in the expression of autoimmune diseases. Studies currentlyin progress in physiological states and in a number of humanendocrinopathies underscore the presented mutual regulationbetween the neuroendocrine and immune systems, and, inparticular, the interaction between the HPG and HPA axis in neuroendocrine–immune homeostasis.
The overall advances in reproductive immunology, summarized following the achievements in LHRH-immunefield, clearly highlight a key role of the decapeptide as aunique channel for bidirectional communication within the neuroimmune–reproductive axis. Thus, the teleologicalassumption that the chief hormone governing the fertilitycapacity of a living organism ‘should’ directly signal themaster gland of the immune system (i.e. the thymus)84 isindeed corroborated by biochemical, molecular, as well asclinical evidence, all supporting the concept that this neuropeptide, may indeed behave as an immunologicalresponse modifier.
MC Morale et al.410

The stress axis, gender and autoimmunity
Under stressful situations, the HPA axis is activated. Thisactivation is driven by CRH produced by parvocellularneurons in the paraventricular nucleus (PVN) of the hypo-thalamus, and results in the stimulation of GC from theadrenal cortex31 (Fig. 10). Glucocorticoids, by binding tocognate receptors, exert a negative feedback action at thepituitary, hypothalamic and extrahypothalamic levels. Acti-vation of the immune system in various infectious diseasestates or following administration of bacterial endotoxin,such as LPS, is accompanied by a marked and long-lastingelevation in the circulating levels of GC.31 This elevation isbelieved to depend on the production and release of cytokinesby activated cells of the immune system. The cytokine-induced activation of the HPA axis occurs primarily by induc-ing the secretion of CRH. The products released from theactivated immune cells can not only affect the activity of theHPA axis, but can also trigger a sympathetic reflex mecha-nism contributing to the final dynamic regulation of theimmune response.94 As a consequence of the coordinatedinteraction between the HPA axis and the immune system,GC secretion from the adrenal glands is increased during thecourse of an immune response, and this mechanism limits themagnitude of the inflammatory reaction to an immunogenicstimulus.
As reviewed in the previous sections, there are complexinteractions between the stress axis, the autonomous (sympa-thetic) nervous system, LHRH, and the sex steroid hormonemilieu in immune system responses to different triggers.
Interruption of the interaction between the neuroendocrine(in particular the HPA axis) and the immune systems is thought to contribute to the pathogenesis of different autoimmune diseases, including tyroiditis, arthritis andexperimental autoimmune encephalomyelitis (EAE).31–38,120
Experimental autoimmune encephalomyelitis, the principalanimal model for multiple sclerosis (MS), and MS have allthe hallmarks of an inflammatory disease of the CNS.121
Here, the regulatory mechanisms that guard against auto-immunity are bypassed, resulting in CNS inflammation.121 InMS, the immune system attacks self-molecules within whitematter and spinal cord, resulting in a progressive decline ofmotor and sensory functions and permanent disability.121
A number of studies have reported that susceptibility toEAE may be influenced by variation in the production ofendogenous GC. In fact, the adrenocortical response iscentral to recovery from EAE in the Lewis (Lew) rat, asreflected by the increased severity of the disease in adrenal-ectomized animals.31–38,122 In contrast, gender has a powerfulimpact on autoimmune diseases, and differences in EAEinduction and MS have been clearly documented.123 Womenhave a more active immune response and a concomitantlyhigher incidence of autoimmune diseases when compared tomen. Oestrogens can depress T-cell-dependent immune func-tions, but enhance B-cell immune responses, thereby aggra-vating B-cell-dependent diseases. The ability of GC toinhibit immune system overreactions and the modulation ofsuch inhibitory effects by the sex steroid hormone milieu,may be explained by mechanisms comparable to the onesreviewed in previous sections.
Neuroendocrine–immune circuitry 411
Figure 10 Alterations of glucocorticoid receptor (GR) mRNA transcript levels in female thymus after castration (OVX) and sex steroidreplacement. Total RNA was extracted from the thymus of Sprague-Dawley female rats at di-oestrous 1 (D1), 3 weeks after castration(OVX) in the absence or presence of sex steroid treatments, and hybridized on northern blots.102 Results are the mean ± SEM of individual determinations in each thymic preparation (6 animals/group), expressed as percentages of GR mRNA/β-actin mRNA ratios relative to control values normalized to 100%. ANOVA shows a significant increase of GR transcript levels after castration, whereas treatment with oestrogens (E
2) counteracted OVX-induced GR mRNA rise. This effect was specific for oestrogen, because progesterone
(P) failed to induce a decrease in the stimulated GR transcripts levels. * P < 0.01 versus OVX; **P < 0.01 versus all experimental groups(by Dunkan-Kramer test).102

Glucocorticoid receptors, pro-inflammatory mediatorsand autoimmunity
Two types of receptors, the type I (mineral corticoid, MR),and type II (glucocorticoid, GR) receptors mediate theeffects of GC, although the GR are primarily involved in thefeedback regulation of the HPA axis.98–100 In general, MRbinding is at a maximum in the nervous tissue, namely in the hippocampus, whereas GR reach their highest levels inlymphoid organs, particularly in the thymus where GR gene expression is markedly elevated.102 Accordingly, T lymphocytes are targets of GC that mediate many of theirproperties including tissue-selective homing, activation andexpansion, as well as their programmed cell death and antigen-specific selection. After binding to their cytoplasmatic GR,translocation of a GR complex to the nucleus and binding toGC-responsive elements (GRE), a variety of genes encodingfor inflammatory mediators are known to be modulated byGC.124,125 Generally, GC inhibit the transcription of severalcytokines that are involved in inflammation (IL-1, TNF-α,GM-CSF, IL-3, IL-4, IL-5, IL-6 and IL-8).124,125 Moreover,the production of lipid mediators, such as prostaglandins, isinhibited and the expression of adhesion molecules (inter-cellular adhesion molecule-1 (ICAM-1) and E-selectin) isreduced. Glucocorticoids have also been found to inhibitother transcription factors, such as nuclear factor-kappaB(NF-κB) and activator protein-1 (AP-1).125 Glucocorticoidsare known to modulate the generation of T helper lympho-cytes from Th-1 cells, preferentially producing IL-2, IFN-γ,mediating cellular immune reactions such as EAE to the Th-2 type cells, preferentially secreting IL-4, IL-10 and IL-13, mediating humoral immune responses, including anti-body-mediated autoimmune responses.31 The exact mech-anisms of GC action in autoimmune diseases such as EAEand MS are unknown, but they may interfere at multiplelevels of the cascading reactions activated during auto-immune disease (see Fig. 12).
Transgenic mice with a dysfunctional GR: Impact ondevelopmental alterations of immune system function
The transgenic (Tg) mouse used as a model for impaired GRfunction was produced by the insertion of a transgeneexpressing constitutive antisense RNA against GR.126 TheseTg mice show reduced GR mRNA in the brain, pituitary,thymus and spleen, reduced brain thymic and splenic GRbinding, and reduced HPA axis sensitivity to GC.10,11,126–129
As a consequence of the impaired GR function, the regulationof the HPA axis in these animals is disturbed.127–130 Trans-genic mice display exagerated responses to stress and toexogenously administrated CRH, whereas the responses incorticosterone are normal because of the hyposensitivity ofthe adrenal gland to ACTH.126–129 Hence, these Tg mice showreduced GR capacity, which is not compensated for by ele-vations in circulating GC levels. At the immune system level,these Tg mice showed alterations in both trafficking andresponsiveness of T lymphocytes.12,13 Interestingly enough,the sex difference accompanying the postnatal developmentof GR gene expression within the thymus gland is abrogatedin GR-deficient mice.13 Moreover, the postnatal developmentof cell-mediated immune response was markedly affected,
with a suppression of the typical sex-dependent difference in immune cell reactivity and higher proliferative responsesto different mitogens in both male and female Tg mice.13 Inaddition, a partial blockade of T-cell differentiation and profound alterations of the thymic microenvironment, whichpersisted into adulthood were observed.130 All together, these
MC Morale et al.412
Figure 11 The hypothalamic–pituitary–adrenocortical axis(HPA) within the neuroendocrine–immune network. The mainregulation of stress-related activity of the HPA axis occurs at thelevel of the hypothalamus (i.e. the parvocellular components ofthe paraventricular nucleus of the hypothalamus).31 Corticotropin-releasing hormone (CRH) and arginine vasopressin are secretedinto the hypophyseal portal circulation in a pulsatile fashion andact synergistically to stimulate adrenocorticotropic hormone(ACTH) by the corticotropic cells, to release the end hormone ofthe stress axis, corticosterone, or cortisol. Aside from the relaseof ACTH and β-endorphin from the pituitary, during the stressresponse, catecholamines are also released from the adrenalglands. During the course of an immune response, the increasedlevels of glucocorticoids act at different (thymus, spleen, lym-phoid cells) levels of the immune axis to limit the magnitude ofthe inflammatory reaction, whereas products from activatedimmune cells feedback information at the hypothalamic–pituitarylevel modulating the release of CRH and ACTH.31

data suggest a key role for GC in coordinating the physio-logical dialogue between the neuroendocrine and the immunesystems. As recalled, neonatal exposure to either syntheticGC or to stress situations has long-term effects on the respon-sivity of the HPA axis. Given the involvement of GC in thecontrol of cellular and humoral immunities, all conditons thataffect the ontogeny of the HPA axis may have importantimplications for individual susceptibility to the developmentof a number of diseases.38 Thus, these GR-deficient mice represent useful models in elucidating the role of the HPAaxis and the interaction with the HPG system in program-ming neuroendocrine and immune reactions in later life.
Conclusions
In the present article we have summarized some aspects ofNEI interactions and discussed some potential clinical impli-cations of this network. The functional interplay between theLHRH system and immune mediators can be studied at theCNS level, where a bidirectional functional interactionbetween the differentiating LHRH neuron and the maturingastroglial cell has been demonstrated, and ongoing inter-actions between sex steroids, growth factors and glial signalssubserve the cyclic variations of the HPG activities responsi-ble for physiological regulation of oestrous/menstrual cycle.Hormonally induced changes in glial plasticity and neuron–glial interactions may dictate some major changes of CNSoutput, and thus actively participate in the different sexdimorphic responses, underlying neuroimmunomodulation.Moreover, the astroglial level represents a major target for stress hormones and inflammatory mediators, thus co-ordinating the CNS response to inflammation, ageing and
neurodegeneration. A specific emphasis was given to oneprincipal system, that is, the LHRH receptor-signallingsystem. All together the presented information supports theconcept that LHRH participates both at central and peripherallevels in the interaction between the neuroendocrine andimmune systems. The reciprocity of the NEI signallingsystems is further supported by the ability of sex steroids tomodulate thymus-dependent immune function via directeffects on specific target genes involved in the developmentof sex dimorphism and sex-dimorphic immune responses,including the downregulation of the immune responseobserved during pregnancy. The widespread therapeuticapplication of LHRH and its potent agonistic and antagonis-tic analogues in a large number of pathologies, such as pae-diatric, gynaecological, urologic and oncologic medicine,underlines the potential clinical implications of the describedexperimental findings. This and other studies may open anew chapter in the neuroendocrinology of reproduction onthe potential immune capabilities of hormones and antihor-mones that are widely used in a variety of endocrinopathies.
The central role of GC in the interactive communicationbetween the neuroendocrine and immune systems is under-scored by the vital role of GC in autoimmunity. Interactionbetween gender and the HPA axis is well exemplified by thesex-related differences in the development of an autoimmunedisease such as EAE. Such interactions may have potentialphysiopathological consequences and shape an individual’scapability to become more or less vulnerable to a number ofdisease entities. Transgenic mice with a dysfunctional GRfrom early embryonic life are useful probes to study suchinteractions at a biochemical and molecular level. The char-acterization of the events and their underlying mechanismsduring disease states, involving the neuroendocrine–immuneunit, will help to understand the development of the diseaseand to generate new ideas for pharmacological targeting withdrugs acting at the NEI interface.
Acknowledgements
This research has been supported by grants to B Marchettifrom the Italian National Research Center (CNR), the ItalianMinistry of Scientific and Technological Research(MURST), the Italian Ministry of Health, and OASI (IRCCS)the Scientific Institution for Research and Care on MentalRetardation and Brain Ageing, Italy.
References
1 Pierpaoli W, Sorkin E. Alteration of adrenal cortex and thyroidin mice with congenital absence of the thymus. Nat. New Biol.1972; 238: 282–5.
2 Pierpaoli W, Sorkin E. Hormones, thymus and lymphocyte function. Experentia 1972; 28: 1385–9.
3 Pierpaoli W, Besedovsky HO. Interdependece of the thymus inprogramming neuroendocrine functions. Clin. Exp. Immunol.1975; 20: 323–9.
4 Pierpaoli W, Koop HG, Muller J, Keller M. Interdependencebetween neuroendocrine programming and the generation ofimmune recognition in ontogeny. Cell. Immunol. 1977; 29:16–26.
5 Marchetti B. Involvment of the thymus in reproduction. Prog.Neuroendocrinol. Immunol. 1989; 2: 64–9.
Neuroendocrine–immune circuitry 413
Figure 12 Schematic presentation of the key role played byactivated macrophages and microglial cells to the inflammatoryprocess within the central nervous system (CNS) during experi-mental autoimmune encephalomyelitis (EAE) and the interactionwith glucocorticoids (GC). After activation macrophages andmicroglial cells produce a variety of cytokines and inflammatorymediators that lead to progression of the immune response andlocal tissue damage. Glucocorticoids via their receptors (GR) canmodulate each step of the inflammatory process.38 (ROS), reac-tive oxygen species; (NO), nitric oxide.

6 Blalock JE. Chemical Immunology. Basel: Karger, 1992.7 Blalock JE. Shared ligands and receptors as a molecular mech-
anism for communication between the immune and neuro-endocrine systems. Ann. NY Acad. Sci. 1994; 741: 292–8.
8 McCann SM, Milenkovic L, Gonzalez MC, Lyson K, Karanth S,Rettori V. Endocrine aspects of neuroimmunomodulation:Methods and overview. In: de Souza EB (ed.). Neurobiology ofCytokines, Part a, Vol. 16. Methods in Neurosciences. SanDiego: Academic Press, 1993; 187–210.
9 Pierpaoli W. The pineal gland as ontogenetic scanner of reproduction, immunity, and aging. The aging clock. Ann. NYAcad. Sci. 1994; 741: 46–9.
10 Marchetti B, Morale MC, Guarcello V et al. Crosstalk commu-nication in the neuroendocrine-reproductive axis: age-dependentalterations in the common communication networks. Ann. NYAcad. Sci. 1990; 594: 309–25.
11 Marchetti B, Morale MC, Guarcello V, Cutuli N, Gallo F,Scapagnini U. The neuroendocrine-immune connections in thecontrol of reproductive functions. In: Adashi EY, Mancuso S(eds). Major Advances in Human Female Reproduction. Milan:Serono Symposia Publications from Raven Press, 1990; 279–89.
12 Marchetti B, Peiffer A, Morale MC, Batticane N, Gallo F,Barden N. Transgenic animals with impaired type II glucocorti-coid receptor expression: a model to study aging of the neuro-endocrine immune system. Ann. NY. Acad. Sci. 1994; 719:308–27.
13 Morale MC, Batticane N, Gallo F, Barden N, Marchetti B. Disruption of hypothalamic-pituitary-adrenocortical system intransgenic mice expresing type glucocorticoid receptor anti-sense ribonucleic acid permanently impairs T-cell functions:effects on T-cell trafficking and T-cell responsiveness duringpost-natal maturation. Endocrinology 1995; 136: 3949–60.
14 Marchetti B, Gallo F, Farinella Z, Morale MC. Neuro-endocrineimmunology (NEI) at the turn of the century: towardsa molecular understanding of basic mechanisms and implicationsfor reproductive physiopathology. Endocrine 1995; 3: 845–61.
15 Marchetti B, Gallo F, Farinella Z, Morale MC. Unique neuro-endocrine-immune (NEI) interactions during pregnancy. In:Kendal M, Marsh J (eds). The Physiology of Immunity. London:CRC Press, 1996; 297–328.
16 Pierpaoli W, Bulian D, Dall’Ara A et al. Circadian melatoninand young-to-old pineal grafting postpone aging and maintainjuvenile conditions of reproductive functions in mice and rats.Exp. Gerontol. 1997; 32: 587–602.
17 Shanks N, Larocque S, Meaney M. Neonatal endotoxin expo-sure alters the development of the hypothalamic-pituitaryadrenal axis: early illness and later responsivity to stress. J. Neu-rosci. 1995; 15: 376–84.
18 Meaney MJ, Diorio J, Francis D et al. Early environmental reg-ulation of forebrain glucocorticoid receptor gene expression:Implications for the adrenocortical response to stress. Dev. Neu-rosci. 1996; 18: 49–72.
19 Bakker JM, Kendall MD. Prenatal stress and immunocompe-tence in later life. Dev. Brain Dysfunct. 1997; 10: 445–61.
20 Catalani A. Neonatal exposure to glucocorticoids: long termendocrine and behavioral effects. Dev. Brain Dysfunct. 1997; 10:393–404.
21 Marchetti B, Gallo F, Tirolo C et al. Developmental con-sequences of hypothalamic-pituitary-adrenocortical system disruption: Impact on thymus gland maturation and the susceptibility to develop neuroimmune diseases. Dev. BrainDysfunct. 1997; 10: 503–27.
22 Barker DJ. In utero programming of chronic disease. Clin. Sci.1998; 95: 115–28.
23 Ahbon E, Gogvadze V, Chen M, Celsi G, Ceccatelli S. Prenatalexposure to high levels of glucocorticoids increase the susceptibility of cerebellar granule cells to oxidative stress-induced cell death. Proc. Natl Acad. Sci. USA 2000; 97: 14726–30.
24 Marchetti B, Gallo F, Romeo C, Farinella Z, Morale MC. Theluteinizing hormone-releasing hormone (LHRH) receptors inthe neuroendocrine immune network: biochemical bases andimplications for reproductive physiopathology. Ann. NY Acad.Sci. 1996; 784: 209–36.
25 Morale MC, Batticane N, Bartoloni G et al. Blockade of centraland peripheral luteinizing hormone-releasing hormone (LHRH)receptors in neonatal rats with a potent LHRH-antagonistinhibits the morphofunctional development of the thymus andmaturation of cell-mediated and humoral immune responses.Endocrinology 1991; 128: 1073–85.
26 Selye H. The Stress of Life. New York: McGraw-Hill, 1976.27 Levine S. Developmental correlates of stress system dys-
function. Dev. Brain Dysfunct. 1997; 10: 347–9.28 Pierpaoli W, Lesnikov VA. Pineal control of stress, distress and
aging: the melatonin evidence. Dev. Brain Dysfunct. 1997; 10:528–37.
29 Spector NH. The great Hans Selye and the great stress muddle.Dev. Brain Dysfunct. 1997; 10: 538–7.
30 Regelson W, Kalimi M. Hormesis, allostasis, and Selye’s generalalarm reaction: cardiomiopathy, calciphylaxis and catatoxic(protectiven steroids) related to current interest to heat shockprotein, antioxidant, and dehydroepiandrosterone. Dev. BrainDysfunct. 1997; 10: 359–370.
31 Crousos GP. The stress response and immune function: Clinicalimplications. Ann. NY Acad. Sci. 2000; 917: 38–67.
32 Sternberg EM, Hill JM, Chrousos JP et al. Inflammatory mediator-induced hypothalamic-pituitary-adrenal axis activa-tion is defective in streptococcal cell wall arthritis-susceptibleLewis rats. Proc. Natl Acad. Sci. USA 1989; 86: 2374–8.
33 Sternberg EM, Wilder RI, Gold PW, Chrousos GP. A defect inthe central component of the immune system-hypothalamic-pituitary adrenal axis feedback loop is associated with suscepti-bility to experimental arthrirtis and other inflammatory diseases.Ann. NY Acad. Sci. 1990; 594: 289–92.
34 Wick GM, Schwarz S, Kroemer G. Immunoendocrine commu-nication via the hypothalamo-pituitary-adrenal axis in auto-immune diseases. Endocrine Rev. 1993; 14: 539–63.
35 Cizza G, Sternberg EM. The role of the hypothalamic-pituitary-adrenal axis in susceptibility to autoimmune/inflammatory diseases. Immunomethods 1994; 5: 73–8.
36 Mason D. Genetic variation in the stress response: susceptibil-ity to experimental and implications for human inflammatorydisease. Immunol. Today 1991; 12: 57–60.
37 Chrousos GP. The hypothalamic-pituitary-adrenal axis andimmune-mediated inflammation. N. Engl. J. Med. 1995; 332:1351–62.
38 Marchetti B, Morale MC, Tirolo C, Testa N, Amor S, DijkstraCD, Barden N. Stress, the immune system and vulnerability todegenerative disorders of the central nervous system in trans-genic mice expressing a glucocorticoid receptor (GR) antisenseRNA. Brain Res. Rev. (in press).
39 Parry BL. Mood disorders linked to the reproductive cycle inwomen. In: Bloom FE, Kupfer DJ (eds). Psychopharmacology.The Fourth Generation of Progress New York: Raven Press,1995; 1029–42.
40 Behl C, Skutella T, Lezzoulac F et al. Neuroprotection againstoxidative stress by estrogens: structure-activity relationship.Mol. Pharmacol. 1997; 110: 535–41.
MC Morale et al.414

41 Schneider LS, Finch CE. Can estrogens prevent neurodegenera-tion? Drugs Aging 1997; 11: 87–95.
42 Dubal DB, Kashon ML, Pettigrew LC et al. Estradiol protectsagainst ischemic injury. J. Cereb. Blood Flow Metab. 1998; 18:1253–8.
43 Marchetti B, Morale MC, Gallo F et al. The hypothalamic-pituitary-gonadal axis and the immune system. In: Ader R,Felten DL, Cohen N (eds). Psychoneuroimmunology. 3rd edn,Vol. 1. New York: Academic Press, 2001; 363–87.
44 Marchetti B, Gallo F, Farinella Z et al. Gender, neuroen-docrine–immune interactions and neuron-glial plasticity: Roleof luteinizing hormone releasing hormone (LHRH). Neuroim-munomodulation: Perspective at the New Millenium. Ann. NYAcad. Sci. 2000; 917: 678–709.
45 Patchev VK, Almeida OFX. Gonadal steroids exert facilitating‘buffering effects’ on glucocorticoid-mediated transcriptionalregulation of corticotropin-releasing hormone and corticosteroidreceptor genes in rat brain. J. Neurosci. 1996; 16: 7077–89.
46 Marchetti B. The LHRH-astroglial network of signals as a modelto study neuroimmune interaction: assessment of messengersystems and transduction mechanisms at cellular and molecularlevels. Neuroimmunomodulation 1996; 3: 1–27.
47 Marchetti B. Cross-talk signals in the CNS. Role of neuro-trophic and hormonal factors, adhesion molecules and inter-cellular signaling agents in luteinizing hormone-releasinghormone (LHRH)-astroglia interactive network. Trends Biosci.1997; 2: 1–32.
48 Benveniste EN. Cytokine production. In: Kattenmann H,Ransom BR (eds). Neuroglia. Oxford: Oxford University Press,1995; 700–16.
49 Giulian D. Microglia, cytokines, and cytotoxins: modulators of cellular responses after injury to the central nervous system.J. Immunol. Immunopharmacol. 1990; 10: 15–21.
50 Ramon Y, Cajal S. Sobre un nuevo proceder de impregnacion dela neuroglia y sus resultados en los centros nerviosos del hombrey animales. Trab. Lab. Invest. Biol. Univ. Madrid 1913; 11:219–37.
51 Horstmann E. Die faserglial des selachiergehirns. Z. Zellforsh.1954; 39: 588–617.
52 Kettenmann H, Ranson BR. Neuroglia. Oxford: Oxford Univer-sity Press, 1995.
53 Knobil E, Hotchkiss J. The menstrual cycle and its neuro-endocrine control. In: Knobil E, Neill J (eds). The Physiology ofReproduction. New York: Raven Press, 1988.
54 Marchetti B, Gallo F, Tirolo C, Testa N, Farinella Z, Morale MC.Luteinizing hormone-releasing hormone (LHRH) is a primarysignaling molecule in the neuroimmune network of signals. Ann.NY Acad. Sci. 1998; 840: 205–48.
55 Gallo F, Morale MC, Avola R, Marchetti B. Cross-talk betweenluteinizing hormone-releasing hormone (LHRH) neurons andastroglial cells: developing glia release factors that accelerateneuronal differentiation and stimulate LHRH release from theGT1 cell line and LHRH neurons stimulate astroglia prolifera-tion. Endocrine J. 1993; 3: 863–74.
56 Gallo F, Morale MC, Farinella Z, Avola R, Marchetti B. Growthfactors released from astroglial cells in primary culture partici-pate in the crosstalk between luteinizing hormone-releasinghormone (LHRH) neurons and astrocytes: Effects on LHRHneuronal proliferation and secretion. Ann. NY Acad. Sci. 1996;784: 513–16.
57 Gallo F, Morale MC, Purrello V et al. Basic fibroblast growthfactor (bFGF) acts on both neurons and glia to mediate theneurotrophic effects of astrocytes on LHRH neurons in culture.Synapse 2000; 36: 233–53.
58 Gallo F, Morale MC, Tirolo C et al. Basic fibroblast growthfactor (bFGF) priming increases the response of LHRH neuronsto neurotrophic factors. J. Neuroendocrinol. 2000; 12: 941–59.
59 Avola R, Spina-Purrello V, Gallo F et al. Immortalized hypo-thalamic LHRH neurons induce a functional shift in astrogliaresponsiveness to growth factors. Int. J. Dev. Neurosci. 2000; 18:743–63.
60 Pearce B, Wilkin GP. Eicosanoids, purine, and hormone recep-tors. In: Kattenmann H, Ransom BR (eds). Neuroglia. Oxford:Oxford University Press, 1995; 377–86.
61 Murphy S, Bruner G, Simmons ML. The role of polyphos-phoinositides in agonist-evoked release of vasoactive factorsfrom astrocytes. In: Yu ACH, Hertz L, Norenberg MD, SykováE, Waxman S (eds). Neuronal–Astrocytic Interactions. Amster-dam: Elsevier Science Publishers, 1992; 153–62.
62 Negro-Vilar A, Conte D, Valenca M. Transmembrane signalsmediating neural peptide secretion: role of protein kinase C acti-vators and arachidonic acid metabolites in luteinizing hormone-releasing hormone. Endocrinology 1986; 119: 2796–802.
63 Ojeda SR, Urbanski HF, Katz KH, Costa ME, Conn PM. Acti-vation of two different but complementary biochemical pathwaysstimulates release of hypothalamic luteinizing hormone-releasinghormone. Proc. Natl Acad. Sci. USA 1986; 83: 4932–6.
64 Rettori V, Gimeno MF, Karara A, Gonzalez MC, McCann SM.Interleukin-1α inhibits prostaglandin E
2release to suppress
pulsatile release of luteinizing hormone but not follicle-stimulating hormone. Proc. Natl Acad. Sci. USA 1991; 88:2763–5.
65 Rettori V, Belova A, Kamat K, Lyson K, McCann SM. Blockadeby interleukin-1α of the nitricoxidergic control of luteinizinghormone-releasing hormone in vivo and in vitro. Neuro-immunomodulation 1994; 1: 86–91.
66 Naftolin F, Mor G, Horvath TL et al. Synaptic remodeling in thearcuate nucleus during the estrous cycle is induced by estrogenand precedes the preovulatory gonadotropin surge. Endocrinol-ogy 1996; 137: 5576–80.
67 Yung-Testas I, Renoir JM, Brugnard GL, Greene EE. Demon-stration of steroid hormones receptor and steroid action inprimary cultures of rat glial cells. J. Steroid Biochem. Mol. Biol.1992; 41: 3–8.
68 Marletta N, Licciardello D, Cormaci GF et al. Effect of 17-betaestradiol and epidermal growth factor on DNA and RNA labeling in astroglial cells during development, maturation anddifferentiation in culture. Mech. Ageing Dev. 2001; 122:1059–72.
69 Chou Y, Luttge WG, Sumnersa G. Expression of mineral-corticoid type I and glucocorticoid type II receptors in astro-cytes and glia as a function of time in culture. Dev. Brain Res.1991; 61: 55–61.
70 Cheng JD, De Vellis J. Oligodendrocytes as glucocorticoidstarget cells: functional analysis of the glycerol phosphate dehydrogenase gene. J. Neurosci. Res. 2000; 59: 436–45.
71 Avola R, Spina-Purrello V, Morale MC et al. Insulin-like growthfactor effects on ADP-rybosylation processes and interactionswith glucocorticoids during maturation and differentiation ofastroglial cells in primary culture. In: Muller E (ed.). Igfs in theNervous System. Berlin: Springer-Verlag, 1998; 127–34.
72 Finch CE. Stress mechanisms and the age-related increase inGFAP transcription in brain astrocytes. Dev. Brain Dysfunct.1997; 10: 350–8.
73 Laping NJ, Nichols NR, Day JR, Jhonson SA, Finch CE. Trans-criptional control of hippocampal glial fibrillary acidic proteinand glutamine synthetase in vivo: opposite responses to to corticosterone. Endocrinology 1995; 135: 1928–33.
Neuroendocrine–immune circuitry 415

74 Nichols N, Osterburg HH, Masters JN, Millar SL, Finch CE.Messenger RNA for glia fibrillary acidic protein is decreased in the rat brain following acute and and chronic corticosteronetreatment. Mol. Brain Res. 1990; 7: 1–7.
75 Besedowski HO, Sorkin E. Thymus involvement in sexual maturation. Nature 1974; 249: 356–9.
76 Grossman CJ. Regulation of the immune system by sexsteroids. Endocr. Rev. 1984; 5: 435–43.
77 Farookhi R, Wesolowski E, Trasler JM, Robairg B. Modulationby neonatal thymectomy of the reproductive axis in male andfemale rats during development. Biol. Reprod. 1988; 27: 1267.
78 Grossman CJ. Are there underlying immune–neuroendocrineinteractions responsible for immunological sexual dimor-phism? Prog. Neuroendocrinimmunol. 1990; 3: 75–80.
79 Marchetti B, Guarcello V, Triolo G, Morale MC, Farinella Z,Scapagnini U. Luteinizing hormone-releasing hormone(LHRH) as natural messenger in neuro-immune-endocrinecommunications. In: Hadden JW, Masek K, Nisticò G (eds).Interactions Among CNS, Neuroendocrine and ImmuneSystems. Rome-Milan: Pythagora Press, 1989; 127–46.
80 Gallo F, Morale MC, Sambataro D, Farinella Z, Marchetti B.The immune response during carcinogen-induced tumorigene-sis. Breast Cancer Res. Treat. 1993; 125: 332–45.
81 Calzolari A. Recherches experimentales sur un rapport prob-able entre la fonction du thymus et celle des testicules. Arch.Ital Biol. (Tor) 1979; 307: 71–6.
82 Marchetti B, Guarcello V, Scapagnini U. Luteinizing hormone-releasing hormone agonist (LHRH-A) binds to lymphocytes and modulates the immune response. In: Castagnetta L, NenciI (eds). Biology and Biochemistry of Normal and Cancer CellGrowth. London: Harwood Accademic Press, 1988; 149–52.
83 Sundaram K, Didolkar A, Thau R, Chandhuri M, Schmidt F.Antagonists of luteinizing hormone releasing hormone bind torat mast cells and induce histamine release. Agents Actions1988; 25: 307.
84 Marchetti B, Guarcello V, Morale MC et al. Luteinizinghormone-releasing hormone (LHRH) binding sites in the ratthymus: Characteristic and biological function. Endocrinology1989; 125: 1025–36.
85 Standaert FE, Chew BP, De Avila D, Reeves JJ. Presence ofluteinizing hormone-releasing hormone binding sites in cul-tured porcine lymphocytes. Biol. Reprod. 1992; 46: 997–1002.
86 Maier CC, Marchetti B, LeBoeuf RD, Blalock JE. Thymocytesexpress a mRNA that is identical to hypothalamic luteinizinghormone-releasing hormone mRNA. Cell. Mol. Neurobiol.1992; 12: 447–54.
87 Azad N, Emanuele NV, Halloran MM, Tentler J, Kelley MR.Presence of luteinizing hormone-releasing hormone (LHRH)mRNA in rat spleen lymphocytes. Endocrinology 1991; 128:679–84.
88 Azad N, La Paglia N, Abel K et al. Immunoactivation enhancesthe concentration of luteinizing hormone-releasing hormonepeptide and its gene expression in human peripheral T-lymphocytes. Endocrinology 1993; 133: 215–22.
89 Wilson TM, Yu-Lee L, Kelly MR. Coordinate gene expression of luteinizing hormone-releasing hormone (LHRH) and theLHRH-receptor following prolactin stimulation in the rat Nb2 T cell line: Implications for a role in immunomodulation andcell-cycle gene expression. Mol. Endocrinol. 1995; 9: 44–53.
90 Marchetti B, Labrie F. Prolactin inhibits pituitary LHRH recep-tors in the rat. Endocrinology 1982; 111: 1209–18.
91 Radivick S, Ticknor CM, Nakayama Y et al. Evidence for directestrogen regulation of the human gonadotropin-releasinghormone gene. J. Clin. Invest. 1991; 88: 1649.
92 Marchetti B, Morale MC, Pelletier G. Sympathetic nervoussystem control of thymus gland maturation: autoradiographiccharacterization and localization of the beta
2-adrenergic
receptor in the rat thymus gland and presence of a sexualdimorphism during ontogenic development. Prog. Neuro-endocrin. Immunol. 1990; 3: 103–11.
93 Marchetti B, Morale MC, Pelletier G. The thymus gland as amajor target for the central nervous system and the neuro-endocrine system: neuroendocrine modulation of thymic beta
2-
adrenergic receptor distribution as revealed by in vitroautoradiography. Mol. Cell. Neurosci. 1990; 1: 10–21.
94 Morale MC, Gallo F, Batticane N, Marchetti B. The immuneresponse evokes up- and down-modulation of a b
2-adrenergic
receptor messenger RNA concentration in the male rat thymus.Mol. Endocrinol. 1992; 6: 1513–22.
95 Marchetti B, Morale MC, Paradis P, Bouvier M. Characteriza-tion, expression and hormonal control of a β
2-adrenergic recep-
tor. Am. J. Physiol. 1994; 267: E718–E31.96 Marchetti B, Fortier M, Poyet P, Folléa N, Pelletier G, Labrie F.
β2-adrenergic receptor in the rat mammary gland during
pregnancy and lactation: characterization, distribution, andcoupling to adenylate cyclase. Endocrinology 1990; 126:565–76.
97 Batticane N, Morale MC, Gallo F, Farinella Z, Marchetti B.Luteinizing hormone-releasing hormone signaling at the lymphocyte involves stimulation of interleukin-2 receptorexpression. Endocrinology 1991; 128: 277–86.
98 De Kloet ER, Reul JMHM. Feedback action and tonic influenceof corticosteroids on brain function: a concept arising from theheterogeneity of brain receptor systems. Psychoneuro-endocrinology 1987; 12: 83–105.
99 De Kloet ER, Vreugdenhil E, Oitzl MS, Joels M. Brain corticosteroid receptor balance in health and disease. Endocr.Rev. 1998; 19: 83–105.
100 McEwen BS, Biron CA, Brunson KW et al. The role of adreno-corticoids as modulators of immune function in health anddisease: neural, endocrine and immune interactions. Brain Res.Rev. 1997; 23: 79–133.
101 Troiano L, Monti D, Cossarizza A et al. Involvement of CD45in dexamethasone and heat-shock induced apoptosis of rat thy-mocytes. Biochem. Biophys. Res. Commun. 1995; 214: 941–8.
102 Peiffer A, Morale MC, Barden N, Marchetti B. Modulation ofglucocorticoid receptor gene expression in the thymus by thesex steroid hormone milieu and correlation with sexual dimorphism of immune response. Endocrine J. 1994; 2:181–91.
103 Tabibzadeh SS, Santhanam U, Sehgal PB, May LT. Cytokine-induced production of IFN-beta
2/IL-6 by freshly explanted
human endometrial stromal cells. J. Immunol. 1989; 42:3134–9.
104 Zanker B, Wal G, Wieder KJ, Strom TB. Evidence that gluco-corticosteroids block the expression of the human interleukin-6 gene by accessory cells. Transplantation 1990; 49: 183–9.
105 Spangelo BL, Judd AM, MacLeod RM, Goodman DW, IsaksonPC. Endotoxin-induced release of interleukin-6 from rat medialbasal hypothalami. Endocrinology 1990; 127: 1779–85.
106 Critchlow V, Liebelt RA, Bar-Sela M, Mountcastle W, Lipscomb HS. Sex differences in resting pituitary adrenal function in the rat. Am. J. Physiol. 1963; 205: 807–12.
107 Gala RR, Westphal U. Corticosteroid-binding globulin in therat: studies on the sex differences. Endocrinology 1965; 77:841–4.
108 Turner BB, Weaver DA. Sexual dimorphism of glucocorticoidin rat brain. Brain Res. 1985; 343: 16–19.
MC Morale et al.416

Neuroendocrine–immune circuitry 417
109 Chisari ANB, Peron MJ, Giovanbattista A, Spinedi E. Gender-dependent charcateristics of the hypothalamo-corticotrope axisfunction in glucocorticoid-repleted and glucocorticoid-depletedrats. J. Endocrinol. Invest. 1998; 21: 737–43.
110 Greenstein BD, Fitzpatrick FTA, Kendall MD, Wheeler MJ.Regeneration of the thymus in old male rats treated with astable analogue of LHRH. J. Endocrinol. 1987; 112: 345–50.
111 Chiodi H. Thymus hypertrophy induced by castration in old ratsand mice. Fed. Proc. 1977; 35: 277A.
112 Marchetti B, Guarcello V, Morale MC et al. Luteinizinghormone-releasing hormone (LHRH) agonist restoration of ageassociated decline of thymus weight, thymic LHRH receptors,and thymocyte proliferative capacity. Endocrinology 1989;125: 1037–47.
113 Ataya KM, Sakr W, Blacker CM, Mutchnick MA, Latif ZA.Effect of GnRH agonists on the thymus in female rats, ActaEndocrinol. 1989; 121: 833–8.
114 Blacker CM, Ataya KM, Savoy-Moore RT, Subramanian MG,Mutchnick MA, Dunbar J. The gonadotropin-releasinghormone agonist leuprolide affects the thymus and other non-reproductive systems of female rats. Acta Endocrinol.1991; 125: 581–6.
115 Rao LV, Cleveland RP, Ataya KM. Sequential changes in func-tional lymphocyte subpopulations during long term adminis-tration of GnRH agonist in postpubertal female mice.Endocrine J. 1993; 1: 451–7.
116 Marchetti BV, Guarcello MC, Morale G et al. A physiologicalrole for the neuropeptide luteinizing hormone-releasinghormone (LHRH) during the maturation of thymus gland function. Int. J. Neurosci. 1990; 51: 287–9.
117 Marchetti B, Morale MC, Batticane N, Gallo F, Farinella Z,Cioni M. Aging of the reproductive-neuroimmune axis. Acrucial role for the hypothalamic neuropeptide luteinizinghormone-releasing hormone. Ann. NY Acad. Sci. 1991; 621:159–72.
118 Jacobson JD, Nisula BC, Steinberg AD. Modulation of expression of murine lupus by gonadotropin-releasing-hormone analogs. Endocrinology 1994; 134: 2516–23.
119 Jacobson JD, Ansari MA, Mansfield ME, McArthur CP,Clement LT. Gonadotropin-releasing hormone increases CD4+T-lymphocyte numbers in an animal model of immuno-deficiency. J. Allergy Clin. Immunol. 1999; 104: 653–8.
120 Shanks N, Windle RJ, Harbuz MS, Jessop DS, Ingram C,Lightman SM. Early-life exposure to endotoxin alters hypo-thalamic-pituitary-adrenal function and predisposition toinflammation. Proc. Natl Acad. Sci. 2000; 97: 5645–50.
121 McFarlin DEM, McFarland HF. Multiple sclerosis. N. Engl. J.Med. 1982; 307: 1183–8.
122 Dowdell KC, Gienapp IE, Stukman S, Wardrop RM, WhitacreC. Neuroendocrine modulation of chronic relapsing experi-mental autoimmune encephalomyelitis: a critical role for thehypothalamic-pituitary-adrenal axis. J. Neuroimmunol. 1999;100: 243–51.
123 Voskhul RR, Pitchekian-Halabi H, Mackenzie-Graham A.Gender differences in autoimmune demyelination in the mouse:implications for multiple sclerosis. Ann. Neurol. 1996; 39:724–33.
124 Barnes PJ, Adcock I. Anti-inflammatory actions of steroids: molecular mechanisms. TIPS 1993; 14: 436–41.
125 Wilckens T. Glucocorticoids and immune function: physio-logical relevance and pathogenic potential of hormonal dys-function. TIPS 1995; 16: 193–7.
126 Pépin MC, Pothier F, Barden N. Impaired type II gluco-corticoid-receptor function in mice bearing antisense RNAtransgene. Nature 1992; 355: 725–8.
127 Stec I, Barden N, Reul JMHM, Holsboer F. Dexamethazonenon-suppression in transgenic mice expressing antisense RNAto the glucocorticoid receptor. J. Psychiatr. Res. 1993; 28: 1–5.
128 Barden N, Stec ISM, Motkowski A, Holsboer F, Reul JMHM.Endocrine profile and neuroendocrine challenge tests in transgenic mice expressing antisense-RNA against the gluco-corticoid receptor. Neuroendocrinol. 1997; 3: 212–20.
129 Karanth S, Linthorst ACE, Stalla GK, Barden N, Holsboer F,Reul JMHM. Hypothalamic-pituitary-adrenocortical axischanges in a transgenic mouse with impaired glucocorticoidreceptor function. Endocrinology 1997; 138: 3476–85.
130 Sacedon R, Vicente A, Varas A et al. Partial blockade of T-celldifferentiation and marked alteration of the thymic micro-environment in transgenic mice with a dysfunctional gluco-corticoid receptor function. J. Neuroimmunol. 1999; 98:157–67.




















