
Neural Control of Movement - NCM Society
48
Society for the Neural Control of Movement
-
Upload
khangminh22 -
Category
Documents
-
view
1 -
download
0
Transcript of Neural Control of Movement - NCM Society

Society for the
Neural Control of Movement
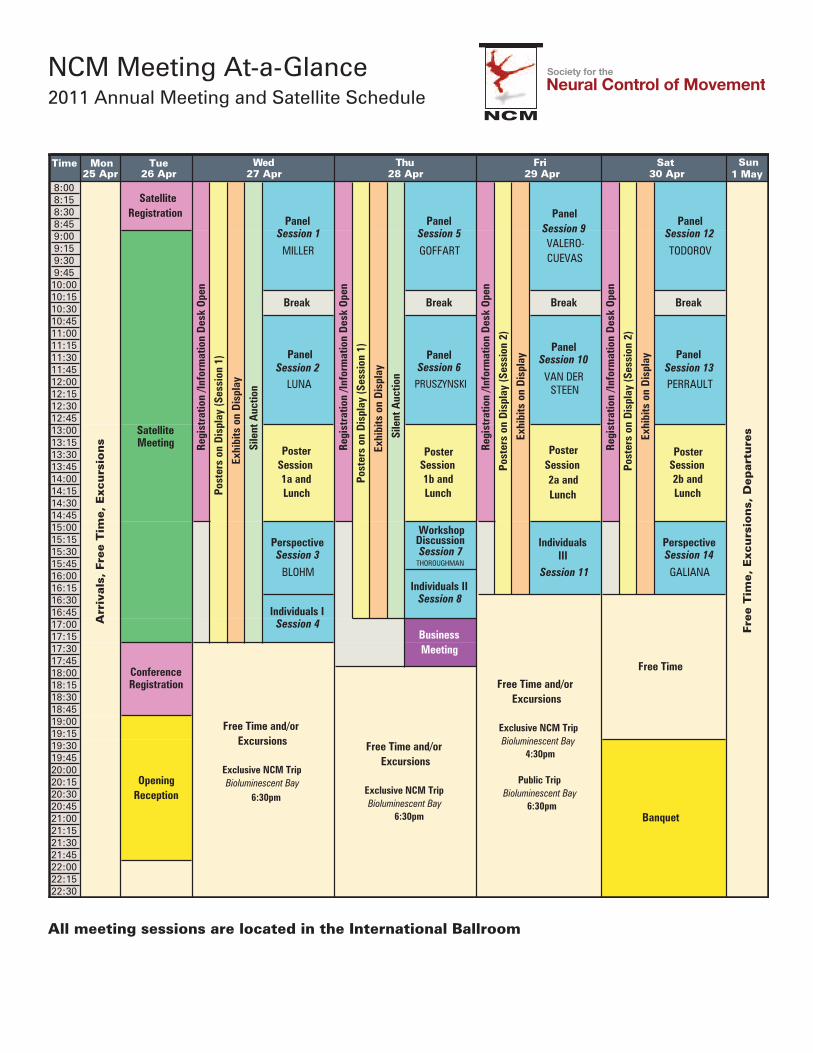
Time Mon Tue Sun25 Apr 26 Apr 1 May
8:008:158:308:459:009:159:309:45
10:0010:1510:3010:4511:0011:1511:3011:4512:0012:1512:3012:4513:0013:1513:3013:4514:0014:1514:3014:4515:0015:1515:3015:4516:0016:1516:3016:4517:0017:1517:3017:4518:0018:1518:3018:4519:0019:1519:3019:4520:0020:1520:3020:4521:0021:1521:3021:4522:0022:1522:30
Fre
eTim
e,
Excu
rsio
ns,
Dep
art
ure
s
Post
ers
onD
ispl
ay(S
essi
on1)
Post
ers
onD
ispl
ay(S
essi
on1)
Post
ers
onD
ispl
ay(S
essi
on2)
Post
ers
onD
ispl
ay(S
essi
on2)
Free Time and/or Excursions
Exclusive NCM TripBioluminescent Bay
4:30pm
Public TripBioluminescent Bay
6:30pm
Exhi
bits
onD
ispl
ay
IndividualsIII
Session 11
Exhi
bits
onD
ispl
ay
Free Time
Arr
ivals
,Fre
eTim
e,
Excu
rsio
ns Re
gist
ratio
n/In
form
atio
nD
esk
Ope
n
SatelliteMeeting
PanelSession 2
LUNA
PosterSession1a andLunch
PanelSession 13PERRAULT
Break
Individuals IISession 8
BusinessMeeting
Exhi
bits
onD
ispl
ay
PerspectiveSession 14
GALIANA
PosterSession2b and Lunch
Regi
stra
tion
/Info
rmat
ion
Des
kO
pen
PanelSession 6
PRUSZYNSKI
Free Time and/or Excursions
Exclusive NCM TripBioluminescent Bay
6:30pm
OpeningReception
ConferenceRegistration
Regi
stra
tion
/Info
rmat
ion
Des
kO
pen
PerspectiveSession 3
BLOHM
Individuals ISession 4
PanelSession 1
MILLER
Break
SatelliteRegistration
SatirFuhTdeW27 Apr
Sile
ntA
uctio
n
WorkshopDiscussionSession 7
THOROUGHMAN
30 Apr28 Apr 29 Apr
PosterSession1b and Lunch
PosterSession2a and Lunch
PanelSession 5
GOFFART
PanelSession 12
TODOROV
Banquet
Free Time and/or Excursions
Exclusive NCM TripBioluminescent Bay
6:30pm
PanelSession 9VALERO-CUEVAS
Break
PanelSession 10
VAN DERSTEEN
Regi
stra
tion
/Info
rmat
ion
Des
kO
pen
Sile
ntA
uctio
n
Break
Exhi
bits
onD
ispl
ay
NCM Meeting At-a-Glance2011 Annual Meeting and Satellite Schedule
All meeting sessions are located in the International Ballroom
Society for the
Neural Control of Movement

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 1
1 About NCM
2 Welcome Letter from the President
2 NCM Leadership
3 Membership Information
4 General Information
5 Special Meetings & Events
6 Future Meetings
6 NCM History
7 Satellite Meeting Program
8-9 Detailed Daily Program
11-27 Abstracts
29 Scholarship Winners
29 NCM Silent Auction
30-31 Poster Sessions by Author
33-41 Poster Sessions – Authors, Titlesand Affiliations
42-43 NCM Sponsors and Exhibitors
Poster Session Floor PlansInside back cover
About NCM
The Society for the Neural Control of Movement(NCM) is an international community ofscientists, clinician-investigators and students,all engaged in research whose common goal isto understand how the brain controlsmovement.
NCM was conceived in 1990 by Barry Peterson.With an initial leadership team that also includedPeter Strick and Marjorie Anderson, NCM wasformally established to bring together scientistsseeking to understand the neural mechanismsthat guide meaningful activities of daily life,primarily through the brain’s control of the eyes,head, trunk, and limbs. Early membersconsisted largely of systems neurophysiologists,behavioral, computational and theoreticalneurobiologists, and clinician-investigatorsinterested in disorders of motor function.
From the outset the goal of NCM was to providea useful gathering of investigators in an informaland casual setting to present and discuss wherewe are in a diverse and complex field, where weshould be going and how we might bestproceed as a community with multipleperspectives and approaches. The meeting wasto be unique in style, such that sessions wereformulated and proposed by small groups ofmembers themselves and geared to inform thelarger attending community through focusedpresentations integrated into themes. Sessionswould change in content with each yearlymeeting.
The inaugural NCM Meeting took place in April,1991 on Marcos Island, Florida, with roughly 140attendees. The success of the initial yearspromoted longevity and expansion of NCM andits meeting, both in attendance (now over 250,with membership over 400) and the breadth ofscientific content. Sessions cover all levels ofinquiry – from perception to genetic expression,and from whole organism to intracellularfunction, while also including computational andtheoretical approaches. This highly regardedmeeting continues to meet in desirable, family-friendly locations in late April or early Mayevery year.
Program Contents

2 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM
Elected members govern the Society for the Neural Control ofMovement. These members comprise the Board of Directorswho in turn elects Officers that comprise the ExecutiveCommittee. The Society’s Bylaws govern how the Boardmanages the Society.
Officers and Board members are elected for three-year termsand may be re-elected to one additional contiguous term. Thecurrent Board comprises the following Officers and Directors:
Officers (Executive Committee)President & Conference ChairGary D. Paige ([email protected])
Vice President & Scientific ChairDoug Munoz ([email protected])
Treasurer & SecretaryHenrietta L. Galiana ([email protected])
Development OfficerTerry Sanger ([email protected])
Welcome to our 21st Annual Meeting in San Juan,Puerto Rico. At this time it is fitting to provide somecomments about NCM, the meeting, and its venue.First, the Society is both healthy and growing. Wehave in recent years witnessed a progressivelyclimbing membership and meeting attendance (lastyear’s surprise volcano not withstanding). Indeedour Puerto Rico meeting attendance is among thehighest in our 21 year history. NCM continues toattract new and established investigators across ourunique blend of disciplines that in turn infuses anunusual range of presentation formats, all matchedby a mix of vibrancy, informality and collegiality. Ithank all of you, for all of that.
Second, NCM will continue to succeed only in light of novelty,which ultimately means new members and contributors thatstem from young investigators. Roughly a quarter of meetingattendance now constitutes students. Further, our scholarshipprogram has grown substantially, thanks to the remarkableefforts of Terry Sanger, our Development Officer. This yearmarks a truly exciting record of 24 awards scattered acrossNorth America and abroad. To continue to support thescholarship program we have added a Silent Auction to theprogram this year. Drop by and bid on the items we havebeen able to secure for your benefit. All proceeds will gotowards the Scholarship fund.
Third, we continue to hone our annual meetings throughexperiments. These include identifying new meeting sites,now among my primary roles. Puerto Rico has the potential tobecome the ‘new Florida’ for us for reasons about to beapparent to all of you. The costs, ecology, and conveniences(scientific and social) all add to substantial benefits for us. Ournext experiment will, as they say, ‘knock your socks off,’ as we
prepare to hold the 2012 meeting in Italy – Venice tobe exact. I look forward to presenting more on thisincredible opportunity at the Thursday afternoonbusiness meeting – please plan to attend this year.
Fourth, experiments in meeting structure continue.Doug Munoz is now our Vice President, responsiblefor the scientific content of the meeting, and hasorganized a magnificent program for San Juan(thanks, Doug – I am enjoying the reprieve…). Oneimportant structural change is that our social mealhas moved from a collective breakfast to a grazinglunch – grazing because this period will include daily
poster viewing as well as time to visit our sponsors’ exhibits.This also means that the meeting will engage attendeesthrough the day, ending earlier than in previous years to allowtime with family, friends and colleagues into the evening. I andthe Board (now identifiable by our badges) solicit yourfeedback on all of the above, as well as the meeting’s content.
Fifth, we are pleased with the growing collaboration we havewith De Armond Management Ltd. The guidance and supportthey are providing for our Society’s affairs and for theplanning and management of our meeting allow us toenhance the offerings and service to all members. Roz, Laurieand Marischal remain available to you throughout the meetingto help with any questions or support you need to ensure youhave a quality experience during the meeting.
Finally, I, my fellow Officers, and the NCM Board welcome allto a truly outstanding meeting here in San Juan.
Sincerely,
Gary PaigePresident
WelcomeTo the Society for the Neural Control of Movement21st Annual Meeting
NCM Leadership
Gary Paige, President
Doug Munoz Henrietta L. Galiana Terry Sanger

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 3
NCM AdministrationAssociation Secretariat& Conference Management([email protected])
De Armond Management Ltd.
Roz BeddallLaurie De ArmondMarischal De Armond
Administrative Assistant
Kathy Martin-Smith, Universityof Rochester
NCM membership is open to all scientists, principalinvestigators and students from around the world, pursuingresearch whose goal is to understand how the brain controlsmovement. Memberships are valid September 1 throughAugust 31 each year.
To become an NCM Member please visit us at the registrationdesk today.
NCM membership includes the following benefits:
• Opportunity to submit proposals and abstracts for sessionsat the Annual Meeting
• Opportunity to submit proposals for satellite meetings
• Opportunity to register for Annual NCM Meetings atreduced registration rates
• Access to the member resource database and othermembers’ web services
• Professional development and networking
• Ability to post job opportunities
• Access and ability to respond directly to job opportunitypostings
• Access to online NCM resources and Annual Meetingproceedings
• Access to scholarships (Grad Students and Post Docs)
• Opportunity to vote in Annual Elections of NCM Boardmembers
• Opportunity to stand for election to, and serve on, the NCMBoard of Directors
• Regular email updates and notices
Board Members
Name Institution Country Term
Andrea d’Avella1 Fondazione Santa Lucia Italy 2009 - 2012
Chris Miall1 University of Birmingham UK 2009 - 2012
Philip Sabes1 University of California USA 2009 - 2012San Francisco
James Lackner1 Brandeis University USA 2009 - 2012
Tim Ebner1 University of Minnesota USA 2010 - 2013
Lee Miller1 Northwestern University USA 2010 - 2013
John Krakauer1 Columbia University USA 2010 - 2013
Kathy Cullen1 McGill University Canada 2010 - 2013
Amy Bastian2 Kennedy Krieger Institute USA 2011 - 2014
Randy Flanagan2 Queens University Canada 2011 - 2014
Paul Cisek2 University of Montreal Canada 2011 - 2014
Jeroen Smeets1 VU University Amsterdam Netherlands 2011 - 2014
1 serving first 3 year term 2 serving second 3 year term
Membership Information
Board ServiceNominations for NCM Board service openin September. Nominations must comefrom members in good standing, and onlymembers are invited to stand for election.To learn more about Board service or ifyou are interested in serving on the NCMBoard, please discuss your interest withone of NCM’s Board members or Officers,or email [email protected].

4 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM
Meeting Venue
El San Juan Hotel & CasinoA Waldorf Astoria Hotel6063 Isla Verde AvenueSan Juan (Carolina), Puerto Rico
The meeting venue is the El San Juan Hotel & Casino. Allmeeting sessions will take place in this location.
Satellite MeetingSatellite Meeting registration fees include a complimentarydrink during a drop in gathering on April 25 and access to thefull day meeting with refreshment breaks and a buffet lunch.
Annual MeetingAnnual Meeting registration fees include access to allsessions including panel, perspective, workshop, and postersessions. Registration also includes daily refreshment breaks,grazing lunches, the Opening Reception and the ClosingBanquet.
Additional TicketsTickets can be purchased separately for your guests and/orchildren for the Opening Reception and Closing Banquet.Breakfast vouchers for the El San Juan breakfast buffet canbe purchased separately for all registrants and their guests ata discounted rate. These additional tickets can only bepurchased from the staff at NCM’s Registration Desk.
Name BadgesYour name badge is your admission ticket to the meetingsessions, coffee breaks, meals, reception and banquet. Pleasewear it at all times. At the end of the meeting we ask that yourecycle your name badge in one of the name badge recyclingstations that will be set out, or leave it at the RegistrationDesk.
To help identify and mentor our future investigators, studentdelegates have red edged badges. All other delegates haveclear badges. NCM Officers and Board Members, Exhibitorsand Staff will be identified by appropriate ribbons.
Dress CodeDress is casual for all NCM meetings and social events.
Registration and Information Desk HoursThe NCM Registration and Information Desk, located in thefoyer, will be open during the following dates and times:
Tuesday April 26 8:00 AM – 9:00 AM5:30 PM – 7:00 PM
Wednesday April 27 8:00 AM – 3:00 PM
Thursday April 28 8:00 AM – 3:00 PM
Friday April 29 8:00 AM – 3:00 PM
Saturday April 30 8:00 AM – 3:00 PM
If you need assistance during the meeting, please visit theRegistration Desk.
Message BoardFor your convenience, a Message Board will be located nearthe Registration Desk. Feel free to leave messages of interestto other meeting participants.
Poster InformationSet-Up / RemovalThere are two Poster Sessions during the Meeting andposters have been allocated to either one of the sessionsbased on poster themes. Poster presenters must set-up andremove their posters during the following times.
Session 1Set-up: Wednesday, April 27, between 7:00AM and 10:00AM
Remove: Thursday, April 28, between 5:00PM and 6:00PM
Session 2Set-up: Friday, April 29, between 7:00AM and 10:00AM
Remove: Saturday, April 30, between 4:30PM and 5:30PM
Information on Poster Authors (Lead), Poster Numbers andPoster Titles begins on page 27. For a complete copy of allthe poster abstracts, a limited supply of printed abstracts isavailable for purchase at the Registration Desk. Digital copiescan be downloaded from the Member Only section of theNCM Website.
Easy reference Poster Floor Plans for each session can befound on the inside back cover of this program.
StaffNCM staff from De Armond Management can be identified byribbons on their name badges. Feel free to ask any one of ourstaff for assistance.
Conference ExcursionsA limited number of spaces remain for the Bio-LuminescentBay kayak trips. If you are interested in joining one of thesetrips, please inquire at the Registration Desk.
Arrangements have also been made for NCM delegates andguests to reserve places on existing trips and excursionsorganized by GSI Puerto Rico, the El San Juan Hotel’s in-house group travel partner. Visit their travel desk, located justinside the exit to the pool and beach area, for moreinformation on the many trips they offer.
Daycare ServicesThe conference hotel provides a Kids Klub at a cost of$50/day for children ages 3 -15 years. This program runsfrom 10:00AM – 4:00PM daily. Lunch, snacks and drinks areprovided. This can be booked through the Pool Concierge,located just outside the exit to the pool and beach area.Advance reservations are encouraged.
If you need access to child care outside of these times, aprivate babysitting service is available at $20/hour. If you areinterested in this service, please visit the Hotel Conciergelocated next to the Hotel Front Desk.
Internet ServicesBedrooms booked through NCM’s group room block includecomplimentary wireless internet. Depending on signalstrength, this service will allow connection throughout theresort’s public spaces, meeting pre-function area and thepool area. Connection information is available through theHotel Front Desk.
No Smoking PolicyThe El San Juan Hotel is a completely non-smoking facility.Smoking areas are located outside the front entrance to thehotel or along the beach.
General Information

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 5
Special Meetings & EventsMonday, April 25 5:00 PM – 6:00 PM
Drop in Reception (Satellite Meeting Registrants)
Location: The Gold Bar, Hotel Lobby
Tuesday, April 26 7:00 PM – 10:00 PM
Opening Reception (cash bar, heavy hors d’oeuvres)
Location: Las Terraza
Wednesday, April 27 9:00 AM – 10:15 AM
Spouses, Guests and Family Meet, Mingle & Plan(light refreshments)
Location: International Ballroom Foyer
Thursday, April 28 9:00 AM – 10:15 AM
Spouses, Guests and Family Meet, Mingle & Plan(light refreshments)
Location: International Ballroom Foyer
Thursday, April 28
1:00 PM – 2:00 PMNCM Annual Luncheon for Trainee ScholarshipRecipients (by invitation only)
Location: The Banyan Tree
5:00 PM – 6:00 PMNCM Business Meeting
Location: Ballroom A
Saturday, April 30 7:30 PM – 8:00 PM
Reception (cash bar)
Location: Outside at the Encanto Bar & Grill
Banquet & Dance 8:00 PM – midnight
Location: Outside at the Encanto Bar & Grill
Floor Plan

6 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM
Meeting Dates City Country Hotel20th Annual Meeting* April 20 – 25, 2010 Naples, Florida USA Naples Beach Hotel & Golf Club19th Annual Meeting* April 28 – May 3, 2009 Waikoloa, Hawaii USA Waikoloa Beach Marriott Resort & Spa18th Annual Meeting April 29 – May 4, 2008 Naples, FLA USA Naples Beach Hotel & Golf Club17th Annual Meeting* March 25 – April 1, 2007 Seville Spain Melia Sevilla16th Annual Meeting* April 30 – May 7, 2006 Key Biscayne, FLA USA Sonesta Beach Resort15th Annual Meeting April 12 – 17, 2005 Key Biscayne, FLA USA Sonesta Beach Resort14th Annual Meeting* March 25 – April 3, 2004 Sitges Spain Melia Sitges13th Annual Meeting April 22 – 27, 2003 Santa Barbara, CA USA Fess Parker’s Doubletree Resort12th Annual Meeting* April 14 – 21, 2002 Naples, FLA USA Naples Beach Hotel & Golf Club11th Annual Meeting March 25 – 30, 2001 Seville Spain Melia Sevilla10th Annual Meeting April 9 – 17, 2000 Key West, FLA USA Wyndham Casa Marina Resort9th Annual Meeting* April 11 – 19, 1999 Kauai, Hawaii USA Princeville Resort8th Annual Meeting April 14 – 22, 1998 Key West, FLA USA Marriott Casa Marina Resort7th Annual Meeting* April 8 – 16, 1997 Cozumel Mexico Presidente Intercontinental6th Annual Meeting Apri 16 – 21, 1996 Marco Island, FLA USA Radisson Suite Beach Resort5th Annual Meeting April 18 – 25, 1995 Key West, FLA USA Marriott Casa Marina Resort4th Annual Meeting* April 13 – 22, 1994 Maui, Hawaii USA Maui Marriott Resort (Lahaina)3rd Annual Meeting April 13 – 18, 1993 Marco Island, FLA USA Radisson Suite Beach Resort2nd Annual Meeting April 21 – 26, 1992 Marco Island, FLA USA Radisson Suite Beach Resort1st Annual Meeting April 6 – 11, 1991 Marco Island, FLA USA Radisson Suite Beach Resort
* indicates a Satellite Meeting was held in conjunction with the Annual Meeting
NCM History
Future Meetings
2012 Annual Meeting and SatelliteWe are pleased to announce the 2012 Annual Meetingand Satellite will take place in one of the mostbeautiful cities in the world. Please plan to attend the22nd Annual Meeting in Venice, Italy.
A Satellite Meeting organized by Pierre-Paul Vidal, Ph.D.,CESeM (Centre d’Etude de la Sensorimotricité), UniversitéParis Descartes - CNRS - UMR 8194 and Andrea d’Avella,Ph.D., Laboratory of Neuromotor Physiology, IRCCSFondazione Santa Lucia will take place April 21 – 23, 2012.
The theme of the Satellite is: Sensorimotor Plasticity andLearning: from bench to bedside and it will have twomain goals:
• Describing the adaptation process, its complexities, and itsneural basis, while also understanding that what we aredealing with is a set of elaborate explicit and internalrepresentations of the environment and our relation to it ina continually changing manner.
• Honoring Professor Jose Maria Delgado Garcia, who is apioneer in the field of adaptation and learning, a dedicatedmentor to students, and an admired colleague and friend tomany.
The Annual Meeting will take place April 23 – 28. Bothmeetings will take place at the Hilton Molino Stucky Hotel,
located less than 10 minutes by complimentary hotel shuttleboat from historic Piazza San Marco. This is one meeting youwill not want to miss. Plan now to attend. Information aboutthe meeting and the location and the hotel (includingreservation information) is now available on the NCMWebsite.
Satellite Meetings
NCM’s Board welcomes suggestions for one or two-daySatellite Meetings in conjunction with future AnnualMeetings. Please discuss your ideas with NCM BoardMembers to formulate an early plan/proposal, and bring thisto the NCM President for further consideration (email:[email protected]).
Keynote Speakers
NCM provides the opportunity for members to suggestprominent colleagues in the field of neuroscience who wouldbe suitable candidates to provide a Keynote Address duringan Annual Meeting. The Keynote is an invited lecturedelivered by a prominent colleague whose contributions toneuroscience are widely acknowledged. Individuals andtopics outside the normal NCM community are encouraged.
If you wish to recommend a colleague as a future keynotepresenter please discuss with an NCM Board Member orOfficer or send an email to [email protected].
Since 1991 NCM’s annual meetings have provided a forumfor leading edge research, scholarly debate, the interchangeof ideas, and a platform for many exceptional established andemerging researchers in the field of Neural Science. We are
proud that this has all been accomplished in some of thenicest destinations in the world. Our history is strong and ourfuture is bright.

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 7
Motor learning: behavior, computation, and pathology
Organizers and Hosts:Amy Bastian, Randy Flanagan & Daniel Wolpert
April 26, 2011 • 9:00AM – 5:30PM
The ability to learn and refine motor actions is an essential part of human behavior. In recentyears there have been dramatic developments in our understanding of motor learning. The satellitemeeting will bring together a set of speakers with a broad range of expertise who will each beasked to focus on a single recent advance in neural and behavioral processes that contribute tomotor learning.
9:00AM-10:30AM Session 1 Neural basis of motor learning
Chair: Randy Flanagan Queens University
Krishna Shenoy Stanford University Toward a single trial view of motor preparationand learning
Pablo Celnik Johns Hopkins University Using non invasive brain stimulation to understandhuman motor learning
Joern Diedrichsen, University College London The neural representation of skilled finger movements
11:00AM-1:00PM Session 2 Normal motor learning
Chair: John Krakauer Johns Hopkins University
James Lackner Brandeis University Motor compensations for Impending Coriolis forces
David Ostry McGill University Motor learning results in changes to somatosensoryfunction and somatosensory areas of the brain
Paul Gribble University of Western Ontario Spatially selective enhancement of proprioceptive acuityfollowing motor learning
Rich Ivry University of California, Strategic and implicit processes in motor adaptationBerkeley
1:00PM-2:00PM Lunch
2:00PM-3:30PM Session 3 Normal motor learning and pathology
Chair: Amy Bastian Johns Hopkins University
Paul DiZio Brandeis University Development of context-specific motor adaptation
Rachael Seidler University of Michigan Cognitive contributions to skill learning
Robert Sainburg Penn State University Lateralization of motor learning, as revealed by studiesin unilateral stroke patients
4:00PM-5:30PM Session 4 Computational approaches to motor learning
Chair: Daniel Wolpert University of Cambridge
Terry Sanger University Southern California Learning a stochastic feedback controller withpopulations of spiking neurons
Maurice Smith Harvard University New insights into the generalization of visuomotoradaptation
Emo Todorov University of Washington Hierarchical learning: Combining the advantagesof realism and wishful thinking
Satellite Meeting Program San Juan, Puerto Rico

8 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM
DAY 1 Tuesday, April 268:00AM – 9:00AM Satellite Registration9:00AM – 5:30PM Satellite Meeting5:30PM – 7:00PM Meeting Registration7:00PM – 10:00PM Opening Reception
DAY 2 Wednesday, April 278:00AM – 10:15AM PANEL Session 1
Achieving a sense of movement: The intersection of somatosensationand motor controlOrganizer: L. Miller Participants: R. Sainburg, S. Helms Tillery, D. Weber
10:15AM – 10:45AM BREAK
10:45AM – 1:00PM PANEL Session 2
Normal and abnormal development of saccade controland its underlying neural circuitryOrganizer: B. Luna Participants: J. Sweeney, N. Alahyane, B. Clementz
1:00PM – 3:00PM POSTER Session 1aLUNCH
3:00PM – 4:30PM PERSPECTIVE Session 3
Neural network models in sensory-motor controlOrganizer: G. Blohm Participants: T. Sanger, P. Sabes
4:30PM – 5:30PM INDIVIDUAL PRESENTATIONS I Session 4
Mechanics and neural control of paw shake responseParticipants: B. Prilutsky, A. Klishko, D. Cofer, G. Cymbalyuk, R. Gregor, D. Edwards
Flexible modulation of perturbation-related activity in primary motor cortexby behavioural contextParticipants: M. Omrani, A. Pruszynski, S. Scott
Representation of ipsi- and bi-lateral finger movementsin primary motor cortexParticipants: J. Diedrichsen, N. Ward, T. Wiestler
Comparison of monkey and human responses to electrical stimulationwith a vestibular prosthesisParticipants: J. Phillips, S. Bierer, L. Ling, C. Kaneko, A. Fuchs, K. Nie, A. Nowack,S. Newlands, J. Rubinstein
DAY 3 Thursday, April 288:00AM – 10:15AM PANEL Session 5
Intercepting a moving objectOrganizer:L. Goffart Participants: V. Ferrera, J. Soechting, J. Smeets
10:15AM – 10:45AM BREAK
10:45AM – 1:00PM PANEL Session 6
Examining the sophistication of fast feedback responses: Reflexes asa window into voluntary controlOrganizer: A. Pruszynski Participants: F. Crevecoeur, C. Honeycutt, L. Selen
1:00PM – 3:00PM POSTER Session 1bLUNCH
3:00PM – 4:00PM WORKSHOP DISCUSSION Session 7
Human see, human do, but how?Organizer: K. Thoroughman Participants: L. Cohen, P. Gribble, P. Wanda, J. Bonaiuto
4:00PM – 5:00PM INDIVIDUAL PRESENTATIONS II Session 8
Effect of task constraints on limb stabization during human locomotionParticipants: A. Auyang, Y. Chang
Learning to be efficient: Split-belt walking adaptation reduces metabolic cost
Participants: J. Finley, A. Bastian, J. Gottschall
Detailed Daily ProgramAll sessions will be held in Ballroom A. Posters will be in Ballrooms B and C.

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 9
L-Dopa impairs ventral striatal recruitment and early phase sequence learningin patients with parkinson’s diseaseParticipants: R. Seidler, Y. Kwak, M. Muller, N. Bohnen, P. Dayalu
Distribution of locomotor interneurons in the lumbar ventral spinal cordof the newborn ratParticipants: J. Cazalets, M. Antri, N. Mellen
5:00PM – 6:00PM BUSINESS MEETING
DAY 4 Friday, April 298:00AM – 10:15AM PANEL Session 9
Myths and monsters in motor controlOrganizer: F. Valero-Cuevas Participants: J. Kutch, G. Loeb, E. Theodorou
10:15AM – 10:45AM BREAK
10:45AM – 1:00PM PANEL Session 10
Timing matters in eye hand coordinationOrganizer: J. van der Steen Participants: P. Lefèvre, F. Maij, C. Miall
1:00PM – 3:00PM POSTER Session 2aLUNCH
3:00PM – 4:30PM INDIVIDUAL PRESENTATIONS III Session 11
Reversing the reversal: Virtual tool use decreases reaction time and restoresrapid online corrections during conditions of visuomotor reversalParticipants: L. Baugh, J. Flanagan
Multisensory recalibration in motor controlParticipants: D. Henriques, E. Cressman, D. Salomonczyk
The nature of the multiple processes underlying motor learning in adaptationto velocity-dependent curl fieldsParticipants: J. Kennedy, C. Miall
Necessity is the mother of invention: Reconstructing missing sensory informationfor concurrent comparisons in multiple reference frames for eye-hand coordinationParticipants: M. Tagliabue, J. McIntyre
Measuring the generalization of visual perturbations of wrist movementsacross movement directions using mobile phonesParticipants: H. Fernandes , M. Albert, K. Kording
Is there a natural speed of reaching movements?Participants: B. Shabbott, P. Mazzoni
DAY 5 Saturday, April 308:00AM – 10:15AM PANEL Session 12
Thinking without the boxes: Towards an integrated view of sensorimotor integrationOrganizer: E. Todorov Participants: M. Churchland, J. Kalaska, W. Kristan
10:15AM – 10:45AM BREAK
10:45AM – 1:00PM PANEL Session 13
The role of biomechanics in the neural control of multijoint posture and movement:What remains for the neural controller?Organizer: E. Perreault Participants: M. Tresch, D. Franklin, L. Ting
1:00PM – 3:00PM POSTER Session 2bLUNCH
3:00PM – 4:30PM PERSPECTIVE Session 14
Sharing of sensory and motor signals in eye-head coordinationOrganizer: H. Galiana Participants: J. Van Opstal, D. Guitton
4:30PM – 7:30PM BREAK
7:30PM – 8:00PM RECEPTION
8:00PM – 11:00PM BANQUET

10 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 11
Abstracts Panel, Perspective, Workshop & Individual Sessions
PANEL Session 1Wednesday, April 278:00AM – 10:15AMAchieving a sense of movement: the intersection ofsomatosensation and motor controlLee Miller1, Stephen Helms Tillery2, Douglas Weber3, RobertSainburg4
1Northwestern University Feinberg School of Medicine, 2Arizona StateUniversity, 3University of Pittsburgh, 4Pennsylvania State University
The dorsal and ventral streams In the visual system may reflect adivision between information processing for action and perception. Thedegree to which somatosensory processing comprises a similardistinction is only speculative. Proprioception is often considered the“hidden” sense: While critical for movement, we are scarcely consciousof it. The sense of touch, on the other hand, is very much a part ofconscious sensation, while playing a less obvious role in movementcontrol. Both senses interact with vision, which complements, butcannot replace the role of either sense in the control of movement.These observations raise important basic scientific questions, but theyalso bring to light fundamental questions about the limits of recentattempts to replace lost somatosensory function through electricalstimulation of the nervous system. In this workshop we will examine theinteraction of these sensory modalities: how they combine to formperceptions of limb state and to guide movement planning andexecution. Sainburg will provide a background for understanding howvision and proprioception might be integrated for control of goaldirected movements. His work using a virtual environment to dissociatevision and proprioception provides evidence that while both modalitiescontribute to a single percept of limb position, discrepantrepresentations from each modality are independently incorporated intodifferent stages of motor planning, but interact to correct movementsduring execution. Helms Tillery will describe properties of cells recordedin cortical areas 1, 2, and 3b, while a monkey grasps objects. He uses avirtual environment to dissociate cell activity related to contact from thatrelated to movement. Many neurons encode both, making it difficult todisentangle their respective contributions. He is now working to decodeindividual finger and palmar-pad contact, and to understand how thebrain uses those two signals to create a coherent spatial tactile image.An ultimate goal is to use intracortical electrical microstimulation (ICMS)to create a realistic perception of object contact. Miller’s work parallelsthat of Helms Tillery, but is focused on the representation of kinematiclimb state and the effect of motor planning by mixed modality neuronsin area 2. He will describe the differential representation of passive andactive limb movement by populations of simultaneously recordedneurons. Using knowledge of this mapping from limb state to neuralstate, Miller has begun to investigate the monkey’s ability to perceiveand respond to “virtual” limb perturbations mediated by ICMS. Finally,Weber will discuss experiments designed to understand the encoding oflimb-state by populations of cutaneous and muscle afferents, with thegoal of delivering somatosensory feedback via peripheral stimulation.Weber records primary afferent activity in the dorsal root ganglia (DRG),and neural activity in cortical areas 3a and 3b during passive limbmovement. He then microstimulates through the DRG electrode arraywith pulse trains that mimic the discharge recorded during the passivemovements. Comparison of the natural and stimulus-evoked activity inS1 provides a convenient approach for assessing how closely thestimulation recreates the S1 neural response to passive movement. Weintend to reserve ample time for a discussion of both S1 codingproperties and the use of electrical stimulation to mimic afferent input.
PANEL Session 2Wednesday, April 2710:45AM – 1:00PMNormal and abnormal development of saccade controland its underlying neural circuitry.Beatriz Luna1, John Sweeney2, Nadia Alahyane3, Brett Clementz4
1University of Pittsburgh, 2University of Illinois in Chicago, 3QueensUniversity, 4University of Georgia
Saccadic eye movements are essential to explore the visualenvironment and guide other motor behaviors. Given the various stimulisurrounding us, we must select and generate the most appropriatesaccade among the multiple competing response options, relying uponthe environment cues and our internal goals. The saccade system allowsthus to study not only basic sensorimotor processes but also higher-order processes such as response inhibition, response planning, anddecision-making that support cognitive control and hence voluntarycontrol over behavior. Simple tasks such as prosaccade (look at aperipheral stimulus) and antisaccade (look away from the stimulus) tasksare extensively used to examine sensorimotor and cognitive control ofbehavior. The extended neural network that controls saccade generation(e.g., frontal, parietal and visual cortical areas, striatum, brainstem,cerebellum) overlaps with the brain circuits that are impaired in variousneurological and neurodevelopmental disorders. Moreover, its structurein terms of gray matter and white matter follows a precisedevelopmental time course during childhood and adolescence. Thebrain areas underlying sensorimotor control mature in early childhood,but synaptic pruning and myelination continue into adolescence, alongwith cortico-subcortical connectivity supporting top-down regulation ofbehavior. In sum, the saccade system provides a unique translationaltarget for investigating behavioral deficits and their neural substrateacross the lifespan. Such work can lead ultimately to refine the etiologyand endophenotypes of brain disorders. The goal of the session is todiscuss the latest behavioral and brain imaging findings on the ability tocontrol saccade initiation in typically-developing children andadolescents, and how this ability is compromised in two neuro-developmental disorders: schizophrenia and autism. Nadia Alahyane willshow that normal maturational changes in saccade performance dependcritically on maturational changes in both frontoparietal andfrontostriatal network activity before stimulus appearance, supportingsuccessful saccade initiation and inhibition. Beatriz Luna willdemonstrate that it is also the ability to retain an inhibitory set oversuccessive trials that is still maturing in adolescence along with theability to process errors. She will illustrate that this age-relatedimprovement relies on frontostriatal white matter tracts thatprogressively strengthen with increasing age, supporting effectiveconnectivity required for frontal top-down control of behavior. BrettClementz will present data indicating that antisaccade abnormalities inschizophrenia are not simply due to a failure of top-down control fromthe frontal cortex. He will show that the difficulty to inhibit theinappropriate response to the stimulus in the antisaccade task inschizophrenia is related to neural activity before stimulus appearance invisual cortex, which indexes the effect of top-down control needed toprepare response inhibition. John Sweeney will review alterations ofsaccade control in autism and their developmental trajectory, and reviewstudies showing that similar alterations can be seen in unaffectedrelatives of individuals with autism.

12 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM
PERSPECTIVE Session 3Wednesday, April 273:00PM – 4:30PMNeural network models in sensory-motor controlGunnar Blohm1, Terence Sanger2, Philip Sabes3
1Queen’s University, 2USC, 3UCSF
The NCM community has always been multi-disciplinary, exploringsensory-motor control through mathematical modeling techniques,behavioral observations, brain imaging and electrophysiology. However,linking findings across these sub-disciplines is often difficult and even inthe best of cases, only correlative. Here we show, with three examples,how artificial neural networks can overcome this challenge, providingboth conceptual and practical links between theory, neurophysiologyand behavior. Historically, modeling work in our community has beendominated by normative algebraic models (state-space model, opticalcontrol theory, Bayesian statistics, etc). While these models aresuccessful at predicting behavior and explaining ‘why the system is theway it is’, they do not address the ‘how’ questions. We believe thatneural network models are the first step toward identifying neuralmechanisms. Therefore, introducing artificial neural networks into ourresearch toolbox constitutes a unique opportunity to causally identifythe brain mechanisms underlying behavior in a number of ways. First,they can be utilized to make specific predictions as to what to look for inelectrophysiological experiments and thus maximize the efficiency andvalue of experiments in advancing knowledge. Second, they arepowerful tools for investigating potential neuronal mechanismsunderlying specific brain functions. The analysis of artificial neuralnetworks is not subject to experimental constraints, allowing us to gowell beyond what is possible with neurophysiological experiments.Third, neural network models can be artificially damaged to investigatepotential mechanisms for brain dysfunction. In this session, we will firstprovide a brief overview of modern artificial neural network approaches,including how they relate to each other and what kind of questions theyenable us to answer. We will then present three different examples, fromthe most biologically realistic networks to the most abstract. TerrySanger will describe the different types of phenomena that arise whennetworks compute using stochastically-driven spikes. Terry will showthat changes in computational bandwidth due to cellular injury or deathin spike-based neural population models may be able to predict some ofthe large-scale functional consequences of injury for patients. Next,mean field theory provides a transition between spiking and rate-basednetwork dynamics which results in a more abstract model of neuronalactivity in a network that is based on continuous-valued real numbers asopposed to discrete spikes. Philip Sabes will show how such rate-basednetworks can approximate normative statistical models, specifically foradaptive sensorimotor integration. He will then discuss the generalprinciples that can be derived from these network models, and showhow these principles are tested physiologically. Finally, a further step ofabstraction removes the temporal dynamics from the equations leadingto static neural networks called perceptrons. Gunnar Blohm willdemonstrate how such multi-layer perceptrons can provide crucialinsight into the brain mechanisms underlying sensory-motortransformations and make new testable predictions about neuronalproperties that one should find at the tip of the electrodes in areasinvolved in this process. We will conclude with a brief discussion ofcurrent limitations and challenges in the field.
INDIVIDUAL PRESENTATIONS ISession 4Wednesday, April 274:30PM – 5:30PMMechanics and neural control of paw shake responseBoris Prilutsky1, Donald Edwards2, David Cofer2, GennadyCymbalyuk2, Robert Gregor1, Alexander Klishko1
1Georgia Institute of Technology, 2Georgia State University
The paw shake response in the cat can be elicited by stimulations ofpaw skin afferents. The apparent goal of this reflex response is toremove the irritating stimulus by imparting extremely high peakaccelerations to the paw (~10 x acceleration of gravity). The mechanicsand muscle activity patterns during the paw shake have unique featuresthat have not been completely explained. For example, during the pawshake the gastrocnemius muscle (GA) is highly active and developingforce while shortening at velocities exceeding the reported maximalshortening velocity of this muscle. The soleus muscle (SO), a closefunctional synergist of the GA, is mostly silent during this behavior.During the paw shake, major hindlimb muscles are apparently organizedin the atypical anterior-posterior synergies rather than in the flexor-extensor synergies seen during locomotion. The goal of thispresentation is to summarize results of our experimental andcomputational studies aimed at explaining the mechanics and neuralcontrol during the paw shake. Major hindlimb muscles of several catswere surgically instrumented with EMG electrodes. The SO and GA inselected cats were also instrumented with sonomicrometry crystals tomeasure fascicle length changes. Paw shake was elicited by attachingadhesive tape to the paw. Hindlimb kinematics were measured using ahigh-speed motion capture system with simultaneous recordings ofEMG or sonomicrometry signals. The kinematics and a hindlimbgeometric model were used to calculate the origin-to-insertion (MTU)lengths of GA and SO, as well as hindlimb joint velocities, moments andpowers. The paw shake experiments were repeated after SO and/or GAmuscles were reinnervated. This procedure allowed for transectedaxons to reinnervate the muscles and eliminated autogenic length-dependent feedback. To interpret the experimental results and gainadditional insight into neural control of the paw shake, we developed acomputational model of the cat hindlimb and its neural control usingAnimatLab, software for modeling and simulations of neuromechanicalsystems. The model consisted of 5 hindlimb segments driven by 11 Hill-type muscle actuators. The muscles were controlled by a half-centeroscillator (CPG), stretch reflex circuits, and spinal pathways mediatingreciprocal inhibition and heterogenic force-dependent interactionsbetween antagonists and agonists. The experimental and computationalresults suggest that muscle and tendon elastic properties contributesubstantially to paw shake behavior. Stretch reflexes appear critical forreshaping muscle activity during paw shake — the anterior-posteriormuscle synergies could be achieved by modulating gains of stretchreflexes without reprogramming the CPG. On the other hand, removal ofautogenic stretch reflexes from the SO and/or GA did not influencemuscle activity substantially, suggesting that these muscles afterreinnervation receive heterogenic length-dependent feedback.
Flexible modulation of perturbation-related activity inprimary motor cortex by behavioural contextMohsen Omrani1, J. Andrew Pruszynski1, Stephen Scott1
1Queen’s University
Our response to someone bumping our arm varies dramatically from aminimal response if our arm is just resting on the table to a rapid precisecorrection if we are holding a cup of coffee. Our hypothesis is that thisselective use of sensory feedback is provided through a transcorticalpathway involving primary motor cortex (MI). Previous work has shownthat MI receives sensory feedback from the limb, but little is known onhow this feedback can be modified based on the behavioural goal. To

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 13
Six-AxisForce/Torque Sensors
Standard FeaturesSix Axes of Force/Torque Sensing (Fx Fy Fz Tx Ty Tz)
High Overload Protect terfaces for Ethernet, PC ,USB, EtherNet/ P, CAN, and mor Sizes from
17 mm – 250 mm diameter Custom sensors available
ApplicationsProduct Test Biomedical Resear r Force Resear Rehabilitation Resear Robotics
www.ati-ia.com919.772.0115

14 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 15
address this issue, we evaluated how MI neurons and upper limbmuscles respond to identical mechanical perturbations under twoconditions. In the first condition (Active task), monkeys performed apostural control task in which they maintained their hands at a centralspatial target and the limb was randomly perturbed. The monkey wasrewarded to return their hand to the spatial target in 750 ms. In thesecond condition (Passive task), monkeys were not required to performany arm motor task when they were perturbed. Instead monkeyswatched a movie without viewing their limb and the hand was servo-controlled to the same spatial location prior to each perturbation. Themonkey was rewarded on all trials regardless of their motor response.Although the two conditions were dramatically different, hand motionaway from the target was identical across the two tasks immediatelyfollowing the perturbation. However, hand motion in the active posturaltask quickly changed direction back towards the spatial goal at ~150 mspost-perturbation. In contrast, there was minimal hand motion back tothe target in the passive task. We evaluated the response of proximal-arm muscles during the two tasks. Brisk motor responses wereobserved within 50 ms following the perturbations in the active posturaltask. In contrast, EMG activity showed a marked reduction in responseto the perturbation in the passive task (on average a 56% decrease). Weevaluated the activity of 127 MI neurons in the active and passive tasks.Most neurons displayed perturbation-related activity 20 to 80 ms afterthe perturbation (70/85 in monkey P & 31/42 in monkey X). Initialanalysis of the data highlighted that perturbation-related activity wassimilar prior to 40 ms across the two tasks. After 40 ms, ~50% ofneurons showed a significant decrease in their response to theperturbation in the passive task, with some showing only a modestreduction and others displaying almost a complete cessation of activity.Across the cell population, the mean drop in the perturbation-relatedactivity was ~40% from the active to passive tasks. These results re-emphasize the robust presence of sensory feedback received in MI andthat this feedback is modulated rapidly in M1 based on the ongoingmotor behaviour. The fact that the initial phasic response was constantacross our tasks suggests that there is a relatively fixed default sensoryinput to M1 that must be shaped to provide selective feedback control.
Representation of ipsi- and bi-lateral fingermovements in primary motor cortexJoern Diedrichsen1, Nick Ward1, Tobias Wiestler1
1University College London
Finger movements lead usually to a suppression of the blood-oxygenation-level-dependent (BOLD) signal during functional magneticresonance imaging (fMRI) in the ipsilateral motor cortex. Based onneurophysiological studies, however, there is evidence that motor-cortical neurons also encode features of ipsilateral or bilateral hand andarm movements. Here we use a simple motor task to study theencoding of movements of single digits in ipsilateral motor cortex usingfMRI. Participants made visually paced key-presses on a piano-styleresponse board with one digit at a time, switching randomly betweendigits every 8.1s. For each voxel and run, we therefore obtained ameasure of how much the activity increased or decreased compared torest for movements of each finger. We then used multi-voxel patternanalysis to find regions that carried information about the moved digit,i.e. regions in which the local pattern of neural responses was differentdepending on which finger moved. As expected, we found large signalincreases and a clear digit representations in the primary motor cortexcontra-lateral to the movement. In ipsi-lateral motor cortex the BOLDsignal decreased compared to rest, but the region also showed above-chance accuracy in classification for the finger that moved. Furtheranalysis shows that the activity patterns associated with ipsi-lateralfinger movements matched the activity patterns associated with contra-lateral finger movements in a finger specific fashion, i.e. voxels in theright motor cortex that increased activity especially during left indexfinger movements, also showed the highest activity (i.e., were leastsuppressed) during right index finger movements. This result suggeststwo processes: Global suppression of the ipsi-lateral motor cortex and afinger-specific ipsi-lateral activation, possibly caused by mirror-symmetric spill-over from the other hemisphere. To test this idea, we
conducted a second experiment, in which we replicated the firstexperiment, but also measured activity during each possiblecombination of bimanual finger movements (one finger per hand). Wereasoned that, if the activity patterns in ipsilateral M1 were due to a spill-over effect, these patterns should be overridden in the bimanualcontext. Indeed, during bimanual movements, the activity patterns in thehand area of primary motor cortex only encoded the contralateral, butno longer the ipsilateral finger movement. However, we also found aneural representation that related to the specific bimanual movement,encoding the unique combination of contra- and ipsilateral fingers. Thisrepresentation was located dorsally and rostrally relative to bestcontralateral finger representation. In sum, our results argue that a)finger movements lead to a decrease of pre-synaptic activity inipsilateral motor cortex, that b) underneath this global suppression, afinger-specific activation can be found, c) that this finger-specificpatterns is overridden during bimanual hand movements, and that d) atrue representation of bimanual hand movements can be found in thetransitional area between primary and pre-motor cortex.
Comparison of monkey and human responses toelectrical stimulation with a vestibular prosthesis.James Phillips1, Steven Bierer1, Leo Ling1, Jay Rubinstein1, ShawnNewlands2, Amy Nowack1, Kaibao Nie1, Albert Fuchs1, Chris Kaneko1
1University of Washington, 2University of Rochester
Introduction: We have developed a vestibular prosthesis designed torestore function following damage to the vestibular end organ. It isconstructed of an internally implanted receiver stimulator and anexternal processor and rotation sensor. The internal device is beingimplanted in rhesus monkeys, and in patients to treat intermittentvertigo due to Meniere’s disease. This offers a unique opportunity tocompare the responses of monkeys and humans to comparableelectrical stimulation with a vestibular prosthesis.Methods: Weimplanted a prosthesis and inserted 2 or 3 leads with three stimulationsites each into the perilymphatic space of individual semicircular canalsin 7 monkeys. In a separate study, three leads were inserted in 3semicircular canals unilaterally in a human patient who received thedevice. Animals were stimulated with biphasic pulse trains of varyingfrequency and current. During mapping of the device in the humansubject, comparable electrical stimuli were used. Eye and headmovements were monitored with scleral and head coils and remote IRvideo in monkeys, and with IR video-oculography in the human subject.In addition, the human subject described the subjective sensations thatresulted from stimulation. Results: Monkeys displayed eye movementsin the plane of the implanted canal with electrical stimulation. Slowphase eye velocity increased with increasing stimulation current orfrequency. For low current stimulation for long durations, headunrestrained monkeys displayed eye and head movement in the planeof the stimulated canal. For short duration stimulation, eye movementswere primarily observed and gaze shifts were reliably perturbed. At highcurrents, roll tilt of the head was observed for lateral canal stimulation,indicating current spread to the utricle. The human patient alsodisplayed eye movements in the plane of each stimulated canal. The eyemovement velocity increased with increasing stimulation current andfrequency. The human subject reported no pain, taste, nausea, orauditory sensation with the stimulation, but did report a consistent rolltilt in addition to a whole body yaw rotation during lateral canalstimulation. The sensation of rotation scaled with increasing current andfrequency in each canal. Prolonged stimulation produced sustainednystagmus and a sensation of continuous whole body rotation.Conclusions: Electrical stimulation of the vestibular end organ with avestibular prosthesis produces comparable results in monkeys and inone human subject. The similar response in both species is promisingfor the primary application of this technology in treating bilateralvestibular loss in patients. This work is supported by NIH HHS-N-260-2006-00005-C, WH Coulter Foundation, and Cochlear Corp.

16 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM
PANEL Session 5Thursday, April 288:00AM – 10:15AMIntercepting a moving objectLaurent Goffart1, Vincent Ferrera2, John Soechting3, Jeroen Smeets4
1CNRS, 2University of Columbia, 3University of Minnesota, 4VrijeUniversiteit
The observation that animals can saccadically foveate and quickly reachwith the hand (or with an instrument) an object that moves in theperipheral visual field indicates that the nervous system is able to copewith the neuromuscular delays unfolding between the detection of anobject and its interception. This ability to intercept a moving objectopens the question of the neurobiological “representations” of aspatiotemporal event and of their functional organization. The aim of thispanel is to provide an overview of this topic by the presentation ofbehavioral and neurophysiological studies performed in non-human andhuman primates, and from the perspective of the oculomotor and limbmotor systems. After a general introduction showing how thisfundamental sensorimotor problem is handled by various animal forms(vertebrate and invertebrate), Laurent Goffart will present resultsobtained in the monkey that illustrate the brain’s flexibility incompensating for unexpected perturbations (induced by brief electricalstimulation in the deep superior colliculus) in the trajectory of saccadesaimed at a moving visual target. Then, Vincent Ferrera will open thequestion of the neural encoding of error signals, in the monkeyprefrontal cortex, that drive the oculomotor system for interceptinginvisible moving targets. The different parameters that enter into theprediction of the future target location for the eye tracking and handreaching movements in humans will afterwards be described by JohnSoechting. More particularly, it will be proposed that the extrapolation oftarget motion is based on target speed and distance for the hand motorsystem, while an extrapolation of target speed, based on a low-passfiltered version of target acceleration, drive the pursuit system. Finally,Jeroen Smeets will describe results of experiments designed to test howeye movements and visual information during the interception are usedto guide the hand. He will show that when human subjects had to hit acompletely predictable moving target through a static aperture in a wall,visual information during the last part of the movement enhancesperformance. In this task, subjects had to be precise both relative to themoving target and relative to the aperture at the moment of interception.The trade-off between these two precision-measures depended on theeye movements. If subjects fixated the aperture instead of pursuing thetarget, the variability relative to the moving target increased, and thatrelative to the aperture was reduced. After a short summary ofpresented results, a discussion will start with the audience to addressthe question whether the various forms of interception can be fused intoa common general framework and whether different “granularities” canbe detected in the brain’s representation of a simple spatiotemporalevent.
PANEL Session 6Thursday, April 2810:45AM – 1:00PMExamining the sophistication of fast feedbackresponses: Reflexes as a window into voluntarycontrolAndrew Pruszynski1, Claire Honeycutt2, Luc Selen3, FredericCrevecoeur4
1Umea University, 2Rehabilitation Institute of Chicago, 3RadboudUniversity Nijmegen, 4Queen’s University
Reflexes have traditionally been viewed as distant and less-sophisticatedcousins of voluntary control. However, there is a growing appreciation
that such fast feedback responses are flexibly tuned to various task-constraints such as movement amplitude, surface texture and intendedvigor. In this session, we will present recent experimental evidence fromhuman, clinical and monkey studies showing that: 1) fast feedbackresponses to various sensory stimuli express a degree of sophisticationtypically reserved for voluntary control; 2) these functional similaritiesmay reflect overlapping neural circuits; and 3) harnessing the linkbetween reflexive and voluntary control could provide a useful tool forstroke rehabilitation. Taken together, these studies emphasize thesimilarities between reflexive and voluntary control and suggest thatthey are more closely related than generally thought. Luc Selen willdiscuss how noisy task-information affects rapid feedback responseselicited by a mechanical perturbation. He will demonstrate that themagnitude of such responses reflects the accumulated evidencesupporting the validity of a particular movement. These results suggestthat sensory evidence from an ongoing decision processes is directlyprovided to the motor system to prepare for an upcoming action.Frederic Crevecoeur will demonstrate that the sophistication of rapidfeedback responses is present even when applied perturbationsapproach the level of natural variability. These findings suggest that thesame feedback control process that occasionally responds to counterlarge perturbations is continuously active during the normal,unperturbed movements that dominate our daily lives. AndrewPruszynski will discuss how fast feedback responses in primary motorcortex reflect an internal model of limb dynamics. He will show thatsingle neurons in primary motor cortex quickly respond to a mechanicalperturbation and that this response appropriately transforms shoulderand elbow motion information into a pattern of motor commands whichappropriately counter the underlying torque perturbation. ClaireHoneycutt will describe the effect of stroke on movement planning andinitiation. She will demonstrate that stroke subjects, who are impaired atvoluntarily initiating a planned reach relative to controls, are statistically-indistinguishable from controls when the movement is initiatedreflexively by a startling auditory stimulus. These results indicate thatstroke subjects have the capacity to plan and deliver movement asquickly as unimpaired individuals.
WORKSHOP DISCUSSIONSession 7Thursday, April 283:00PM – 4:00PMHuman see, human do, but how?Kurt Thoroughman1, James Bonaiuto2, Paul Wanda3, Paul Gribble4,Leonardo Cohen5
1Washington University in St Louis, 2California Institute of Technology,3Washington University in St Louis, 4University of Western Ontario,5NINDS, NIH
People routinely acquire new motor behaviors through observation. Animproved golf swing is more likely to be gained through coaching orstudy than through self-guided random exploration. Physical andoccupational therapists can teach patients to control movements morestably and safely through demonstration. Although coaching andtherapeutic demonstrations clearly and effectively influence humanbehavior, the facility of the human brain to transform visual capture intomovement is computationally and physiologically remarkable. Visualinformation is captured in a retinal space and needs to be transformedto usefully influence muscular output. Observation provides just asubset of the temporal and spatial features of movement that need to becontrolled. Neurophysiology and imaging have characterized qualitativefeatures of mirror neurons and mirror systems, but a rigorousidentification of how observation influences behavior is just nowemerging. After an introduction by K. Thoroughman, L. Cohen will detailhis foundational studies of behavior and transcranial magneticstimulation to show how action observation influences content andneural bases of motor memories in health and in disease. P. Gribble will

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 17
Quick Facts
www.bkintechnologies.comBKIN Technologies, Suite 1625, Biosciences Complex, 116 Barrie Street, Kingston, Ontario K7L 3N6 CanadaTel: +1(888) 533-4393 Email: [email protected]
The KINARM™ Family
Robotics: Probing brain function and dysfunctionBKIN Technologies is a leading developer of robotic technologies for probing brain function and dysfunction. Our patented KINARM™ product line enables basic and clinical researchers to explore sensory, motor and cognitive performance in both humans and NHPs. Our robotic and software tools create complex mechanical and visual worlds. BKIN‘s products are paradigm shifting and hold the promise of being the first quantitative and objective system to assess sensory, motor and cognitive function - critical instrumentation for the effective management of brain disease and injury.
Our Approach
The 2-dimensional ParadigmFundamental to BKIN’s approach is our 2-dimensional paradigm in which the workspace is restricted to a single plane. This 2-dimensional paradigm represents the optimal balance between behavioural complexity and measurement simplicity.
Integrated Virtual/Augmented RealityOur virtual/augmented reality systems allow visual information to be presented in the plane of the workspace. Aligning the visual and mechanical environments in this manner takes advantage of the natural coupling that exists between the visual and proprioceptive systems.
Our Goal
BKIN Technologies can help you choose the right product to fit your needs. Our multidisciplinary team is experienced in neuroscience, engineering, physiology, electronics, machining and software development. We work closely with basic and clinical researchers to integrate our technologies into their unique research programs to accelerate their research and achieve their goals.
• Multiple robotic platforms to fit with,and enable growth of, your research program
• 2-dimensional paradigm balancing complexity/simplicity
• Integrated VR for natural visual and proprioceptive coupling
• Sophisticatedengineering designed byneuroscientists for neuroscientists
“KINARM is a broad-based, versatile research facility that allows users to develop and design their own behavioural tasks to study sensory,
motor and/or cognitive deficits.”
– Dr. Stephen Scott, Professor, Centre of Neuroscience Studies, Queen’s University and Inventor of KINARM

18 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM
Precision Instrumentationfor the Sciences
MICROMANIPULATIONRock-solid precision and sub-micron accuracy. Whether your application requires the MPC-200 for single-handed access to multiple manipulators, the programmable capabilities of the MP-285, the simplicity of the MP-225, or one of our traditional, manual style instruments, we have a manipulator to meet your needs.
OPTICAL PRODUCTSWe’ve expanded our line of optical products to include the SmartShutter™, a full-featured controllable shutter, and the Lambda 10-3 controller, capable of commanding up to three filter wheels and two shutters. Other optical instruments include the DG-4 high-speed wavelength switcher, Lambda LS xenon light source, and a full range of filter wheels. Robust, reliable, and versatile, our imaging hardware can be driven by mostimaging software and adapted to any microscope.
MICROPIPETTE FABRICATIONSutter Instrument, the recognized leader in micropipette fabrication technology, offers leading edge technology in the P-1000 micropipette puller with an intuitive, full-featured interface. An extensive library of built-in programs is available through the color touch-screen display, taking the guesswork out of pipette pulling.
MICROINJECTIONThe XenoWorks™ microinjection system has been designed to meet the needs of a wide variety of applications that require the manipulation of cells and embryonic tissues including ICSI, ES Cell Microinjection, and Adherent Cell Microinjection.Highly-responsive movement and excellent ergonomicsintuitively link the user with the micropipette, improvingyield – saving time and resources.
ONE D I G I T A L D R I V E , N O V A T O , CA. 94949 PHONE: 415.883.0128 | FAX: 415.883.0572 EMAIL: [email protected] | WWW.SUTTER.COM

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 19
then describe a series of studies in which people capture knowledgeabout force-feedback (haptic) environments solely through observation,and what this capture reveals about neural representation of dynamics.P. Wanda will explore how experience of movement and observation,interleaved on a trial-by-trial basis, reveals the quantitative effect of eachon motor memory. J. Bonaiuto will present mathematical models he hasbuilt of the mirror system and hypothesize how elements of simpleimitation can evolve into the basis of mimicry of high-dimensional,complex behavior. We will conclude with discussion of how behavioral,computational, and physiological investigations can further identify howwe as humans can adapt motor behaviors not solely solitarily, butinterpersonally and interactively.
INDIVIDUAL PRESENTATIONS IISession 8Thursday, April 284:00PM – 5:00PMEffect of task constraints on limb stabization duringhuman locomotionArick Auyang1, Young-Hui Chang1
1Georgia Institute of Technology
Our goal was to investigate the use of motor redundancy to stabilize thetask level variables leg length and leg orientation changed whenpresented with locomotor task constraints. We previously showed thatleg length and leg orientation were stabilized through the structuring ofsegment angle variance during human hopping in place (Exp Brain Res192:253-264, 2009). We determined the amount of task variablestabilization and the structure of segment angle variance by quantifyingthe Goal-Equivalent Variance (GEV) and Non-Goal Equivalent Variance(NGEV) components of leg segment variance using an UncontrolledManifold approach (Exp Brain Res 126:289-306, 1999). Subjects hoppingat non-preferred frequencies showed increased leg length stabilizationthrough structuring segment angle variance. This work furtherdemonstrates that increasing leg segment variance structure is a robustcompensation strategy in response to task constraints and increasedtask difficulty during locomotion. Subjects hopped in place into a squaretarget placed on the ground. Three target sizes were used to vary taskdifficulty according to Fitts’ Law (0.25m2, 0.063m2, and 0.01m2). Wereasoned that decreasing target size should increase the need for legorientation stabilization. Leg orientation is typically stabilized in midaerial phase during hopping and we hypothesize that decreasing thetarget size should increase leg orientation stabilization through thestructuring of segment angle variance. In contrast, changing target sizeshould have no effect on the ability to stabilize leg length. Our resultsshow that decreasing the target size resulted in a significant increase inthe structure of segment angle variance to stabilize leg orientation.Surprisingly, we also observed a significant decrease in variancestructure for leg length stabilization. This suggests a trade off effectwhereby increases in the stabilization of an explicit task variable comesat the expense of stabilizing another implicit task variable. The results ofthis work suggest that the use of redundant combinations of segmentangles may be a robust strategy by which human locomotion isachieved when presented with new conditions and complexenvironments.
Learning to be efficient: Split-belt walking adaptationreduces metabolic costJames Finley1, Amy Bastian1, Jinger Gottschall2
1Johns Hopkins University, 2Pennsylvania State University
Human locomotion adapts for novel demands in natural environments,such as ice, snow or sand. This learning process is thought to be usedto improve walking economy (i.e. metabolic cost) and stability (i.e.
imbalance), though this has never been experimentally tested. Here weasked whether walking adaptation on a split-belt treadmill leads to amore economical walking pattern. In this experimental paradigm, whenthe treadmill belts are ‘split’ and moved at different speeds, subjectsinitially walk with a spatiotemporal asymmetry, but adapt back tosymmetry. Why do individuals select a symmetric gait pattern in anasymmetric environment? One hypothesis is that individuals typicallyselect a step length that minimizes energy consumption. Therefore, it ispossible that a similar optimization process drives the restoration ofsymmetry commonly observed during split-belt adaptation. We tested ifsplit-belt walking resulted in a transient increase in metabolic cost and ifthis cost decayed as step symmetry was restored. Eleven individualswere recruited for this study and each person walked in 4 conditions:three ‘tied’ belt conditions (belts at same speed at 0.5 m/s, 1.0 m/s, and1.5 m/s) and one split-belt condition (slow belt: 0.5 m/s, fast belt: 1.5m/s). Subjects walked in each of the tied conditions for 5 minutes andsubsequently adapted to the split-belt perturbation for 16 minutes. Atthe conclusion of adaptation, storage of a new pattern (i.e. an after-effect) was assessed during a 5 minute period when subjects walked atthe slow speed. Adaptation was characterized by a measure of stepsymmetry which is defined as the normalized difference in step lengthbetween the legs on the fast and slow belts. Metabolic power wascalculated using a standard equation based on oxygen consumption andcarbon dioxide production. Results showed that metabolic powerpeaked during early adaptation, and the amplitude of this peakcorrelated with the magnitude of the initial step symmetry perturbation(r2 = 0.58, p = 0.02). Metabolic power was then gradually reduced assubjects’ step lengths became more symmetric. During post-adaptation,when the belts were tied, a significant after-effect was present and wasassociated with a second rise in metabolic cost. Here again, stepsymmetry and metabolic power decayed until symmetry was restoredpost-adaptation. In sum, adaptation to a split-belt perturbation (i.e. goingfrom an asymmetric to symmetric gait) is associated with a minimizationof energy expenditure. This finding is important as it suggests thatimproving symmetry in patients with gait impairments using split-belttreadmill training may lead to more efficient walking. Supported by NIHHD048741, HD007414
L-DOPA impairs ventral striatal recruitment andearly phase sequence learning in patients withParkinson’s diseaseRachael Seidler1, Youngbin Kwak1, Martijn Muller1, Nicolaas Bohnen1,Praveen Dayalu1
1University of Michigan
The early phase of motor sequence learning relies on the ventral andanterior striatal circuitry, whereas the late phase relies on the dorsal andposterior striatal circuitry (Lehericy et al. 2005 PNAS). Early Parkinson’sdisease (PD) is mainly characterized by dopaminergic denervation of thedorsal and posterior striatum while sparing anterior and ventral regions.Dopaminergic medication improves dorsal and posterior striatumfunction by compensating for the loss of dopamine. However, previouswork has shown that dopaminergic medication interferes with theventral and anterior striatum function by overdosing this relatively intactstructure in early-state PD (Cools 2006 Neurosci Biobehav Rev). Here wetest whether these effects are also observed over the time course ofmotor sequence learning. In Study 1 (Kwak et al. 2010 J Neurophys)fourteen PD patients ON and OFF dopaminergic medications and 11healthy age-matched control participants performed an explicit motorsequence learning task. We observed a significant sequence learningimpairment associated with medication that was specific for the earlyphase of learning. In Study 2 (Kwak et al. under review), we investigatedthe neural substrate of this deleterious effect of L-DOPA on motorsequence learning. A new cohort of patients participated in Study 2.Seventeen PD patients ON and OFF L-DOPA and 21 control participantsperformed an explicit motor sequence learning during MRI scanning. Weagain observed an L-DOPA associated sequence learning impairment forearly learning. In addition, we found sequence learning-specificactivation during the early phase in the ventral putamen for controls andPD OFF but not for PD ON L-DOPA. The degree of L-DOPA associated

20 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM
Contact us for a demonstration
in your lab.
ripple LLC,© 2011 patents pending
connect to
toll free: (866) 324-5197 local: (801) 413-0139
fax: (801) [email protected]:www.rppl.comweb:
®
starting under
$60,000for 128 channelrecording systems
front ends for electrodes
32-channel animal microelectrode Nano
Micro
32-channel single reference - EEG, ECog, ENG
16-channel di�erential pairs - EMG
32-channel single reference - spikes and LFP
Surf D
Surf S
Digital I/O
Analog I/O
front ends for digital & analog
amplify, �lter, and digitize biopotential signals
connect to other instruments and audio devices
40 channels LVTTL i/o
56 channels, ±5 V analog i/o with audio in/out
neural interface system
neuroscience research
neuroprosthetic development
up to 512 electrodes
Open Source
Windows, Mac OS X, Linux
emsstding syorecror 128 channelffor 128 channel
$60,000ting undertars
sl
0r
Windows, Mac OS X, Linux
europrosthetic development
euroscience research
eural interface system
ows, Mac OS X, Linux
oprosthetic development
oscience research
al interface system connect to
x
ment
o
pen Source
p to 512 electrodes
front ends for electrodesamplify, �lter, and digitize biopotential signals
16-chrf D
Source
512 electrodes
or electrodesand digitize biopotential signals
EMGannel di�erential pairs -
Nano 32-channel animal microelectrode
32-ch
32-chrf S
cro
annel animal microelectrode
spikes and LFPannel single reference -
EEG, ECogannel single reference -
d LFP
g, ENG
front ends for digital & analog
56 channels, ±5
40 channels LVTT
connect to other instruments aront ends for digital & analog
Analog I/O
Digital I/O
V analog i/o with audio in/out
L i/o
nd audio devicesanalog
patents pending© 2011 LLC,ripple
web:email:
fax:local:
toll free:
eb: www.rppl.comail: .comsales@rppl
(801) 413-2874ax:(801) 413-0139al:
5197(866) 324-ee:

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 21
activation decrease was positively correlated with the degree ofsequence learning impairment in the early phase of learning. Thesefindings provide evidence for the selective effects of L-DOPA on theventral cortico-striatal loops involved in motor sequence learning inParkinson’s disease. In Study 3 (Kwak et al. in preparation) weinvestigated whether this L-DOPA associated impairment in earlysequence learning was larger for patients with less anterior and ventralstriatal dopaminergic denervation. A new cohort of 17 PD patientsunderwent 11C-dihydrotetrabenazine (DTBZ) PET scans to measure theirlevel of striatal denervation, calculated as the percentage decrease inDTBZ binding potential (BP) from age-matched controls in 6 differentstriatal subregions. We found a negative linear relationship between thelevel of L-DOPA associated impairment in early sequence learning andthe percentage BP decrease in the dorsal anterior and posteriorputamen of the more affected hemisphere. In sum, these experimentsdemonstrate that dopamine ‘over-dose’ of the ventral and anteriorstriatum results in impairments in the early phase of motor sequencelearning in PD patients. Supported by the Gustavus and Louise Pfeifferfoundation.
Distribution of locomotor interneurons in the lumbarventral spinal cord of the newborn ratJean rene Cazalets1, Nicholas Mellen, Myriam Antri1CNRS
Although the mammalian locomotor CPG has been localized to lumbarspinal cord, the functional-anatomical organization of flexor andextensor interneurons has not been characterized. Here, we tested thehypothesis that flexor and extensor interneuronal networks for walkingare physically segregated in the lumbar spinal cord as postulated earlierin the mudpuppy. For this purpose, we performed optical recordingsand lesion experiments from a horizontally sectioned hemispinal cordisolated from neonate rats. This hemi-spinal ventral cord preparationexposes locomotor networks at its dorsal surface, and produces well-organized fictive locomotion when superfused with 5-HT/NMDA. Spinalcord network activity accompanying fictive locomotion was visualizedusing the Ca2 indicator fluo-4 AM, while simultaneously monitoringmotor output at ventral roots L2 and L5. Interneurons that maintained astable phase relationship with motor output were retained for furtheranalysis. Rat locomotor interneurons were found along the segmentsrecorded from, but were concentrated at L1. This finding wascorroborated by the observation that lesions localized at segment L1induced more severe changes in motor activity than those in thesegment T13-L2. Whatever the lesion level we did not observe selectivedisruption of either flexor or extensor motor output. These resultssuggest that a physiological segregation of flexor and extensorinterneurons does not exist in the rat.
PANEL Session 9Friday, April 298:00AM – 10:15AMMyths and monsters in motor controlFrancisco Valero-Cuevas1, Jason Kutch1, Gerald Loeb1, EvangelosTheodorou1
1University of Southern California
Progress in the field of Sensorimotor Neuroscience hinges on a clearand up-to-date understanding of its fundamental tenets. While thevalidity of these tenets is routinely discussed and debated in theliterature, it is also necessary to present them to the community in amanner that challenges all of us to affirm, update or revise them. This isparticularly true when challenges or revisions to these tenets hinge onthe details of specialized computational approaches, a nuancedinterpretation of multiple experimental findings, a thorough knowledgeof a vast literature, integration of recent findings, and/or careful semanticdistinctions. We have assembled a team of four investigators withrelevant expertise to critically review four fundamental tenets in light of
recent work: Muscle redundancy; Valero-Cuevas Optimal Control;Theodorou Henneman’s Size Principle; Kutch Spinal Circuitry for MotorBehavior; Loeb The Panel will progress systematically from theperiphery inwards to update and revise our understanding of these fourcritical aspect of Sensorimotor Neuroscience. We begin by discussinghow the physics of tendon-driven systems defines the problem to besolved by the nervous system’and go on to show that we, in fact, havebarely enough muscles for versatile behavior. While these concepts donot change the fact that there exist multiple solutions for ‘simple’ tasks(i.e., with few constraints) and in the close vicinity of a valid coordinationpattern for a ‘complex’ task (i.e., with multiple constraints), they dostrongly set the scope of feasible solutions, necessitate ‘co-contraction’and ‘synergistic’ patterns independently of neural considerations andmake the system vulnerable to loss or weakness of even single muscles.The second part discusses the limitations of the optimal controlframework and its implications to neuromuscular systems. Optimalcontrol requires accurate knowledge of the dynamics and cost functions.Even though these requirements may be satisfied in tasks such as smallreaching movements, they are strongly violated in learning scenarios orcontrol tasks in which the dynamics and rewards are not known or vary(i.e., the majority of motor behaviors). We will provide an overview ofother control theoretic and machine learning approaches seldomapplied in our field motivated by these limitations and well suited toaddress them. These include reinforcement learning and modelpredictive, robust, and adaptive control. Next we transition to areassessment of the physiological machinery that drives the recruitmentof motor units and challenge the notion that recruitment system is asautonomous and robust as is generally believed. A noninvasiveinnovative approach called EMG-weighted averaging can reveal theprofile the motor unit activation pattern across the voluntary range (0-100% MVC) of an isometric contraction. By linking this experimentalapproach with computational models of motor unit activation we findthat motor unit recruitment is itself tuned in accordance with theirdirection of mechanical force output. Lastly, a computational assessmentof the most thorough compendium of spinal circuitry to date sheds lighton the high-level central drive necessary and sufficient to producecomplex voluntary movement and suggest the system can robustlymeet these demands in any given individual with minimal tuning of theparameters of the circuitry.
PANEL Session 10Friday, April 2910:45AM – 1:00PMTiming matters in eye hand coordinationJohannes van der Steen1, Philippe Lefèvre2, Femke Maij3, Chris Miall4
1Erasmus Medical Center, 2Université catholique de Louvain,3VU university Amsterdam, 4University of Birmingham
Orienting behavior in our environment, searching and reaching forobjects such as grasping a cup of coffee, pointing and attending tointeresting spots or drawing are very normal tasks in daily life. Suchtasks involving the eye and hand require a complex interaction betweencortical networks specialized for processing visual and proprioceptiveinformation and those specialized for planning and executing motortasks (Corbetta et al., 2008). Different subsets of neuronal populations inthe frontal and parietal cortex are responsible for processing distinctfeatures of eye hand coordination (see Wise et al. 1997 and Caminiti1998 for reviews). Temporal and spatial factors play a crucial role in theexecution of coordinated eye hand movements, and so we will presentand discuss the importance of these factors in sensorial processing andexecuting coordinated eye hand tasks. Femke Maij will present resultson the perception of briefly sensed stimuli during eye- and handmovements. Both the tactile and visual localization show systematicerrors. These errors have a temporal origin, as they can be manipulatedby adding irrelevant temporal information. She will present a model thatdescribes the results quantitatively as being caused by temporaluncertainty about the time of the stimulus, for the eye combined with aspatial prior of the tendency to believe that the stimulus occurred near

22 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM
the fovea. Chris Miall will present recent results on timing and spatialinformation from eye and hand tracking in artists drawing from livemodels, as well as more controlled drawing tasks and fMRI experiments.He suggests that in the process of using vision to guide the drawingaction, the spatial information is retained as a motor action, rather than avisual image. An important issue these studies highlight is the extent towhich visual control of the hand is achieved using non-foveal vision.Philippe Lefèvre will present data on eye-hand coordination whereparticipants performed a targeted tapping task with a hand-held objecttoward up and down targets. His presentation will focus on theimportance OF PREDICTION ON the timing of eye saccadesASSOCIATED TO THE HAND MOVEMENT and the role of prediction ingrip force modulation in high impact loads (collisions). By using catchtrials and a virtual environment, he demonstrates that this peak of gripforce is pre-programmed. Hans van der Steen will present data onrelative timing of coordinated eye-hand movements in tasks withvarying cognitive complexity. Healthy human subjects were instructed toperform various tapping tasks involving pro-tapping anti-tapping andspatial memory. The timing of healthy subjects in these tasks will bediscussed and contrasted against patients in the early onset phase ofAlzheimer dementia or in Parkinson’s disease. We will entice theaudience to participate in the discussion and propose that JeroenSmeets act as moderator in this session.
INDIVIDUAL PRESENTATIONS IIISession 11Friday, April 293:00PM – 4:30PMReversing the reversal: Virtual tool use decreasesreaction time and restores rapid online correctionsduring conditions of visuomotor reversal.Lee Baugh1, John Flanagan1
1Queen’s University
Despite our ability to adapt to conditions of visuomotor reversal, there isa reaction time cost in performing these movements when compared toconditions in which no reversal is present. Further, research has shownthat even after complete adaptation, fast corrections to a sudden jump intarget location are delayed and/or performed in the incorrect direction(as if no reversal was present). Motivated by the remarkable ability forhumans to adapt their movement plans to incorporate tool use, thepresent study assessed whether additional visual information similar towhat would be provided while using a tool would improve performance.Subjects were required to hit virtual targets with a cursor while vision ofthe arm was prevented and hand position was represented in themovement plane. During the reversal trials, tool-like visual informationconsisted of a stick pivoting around a center point connecting handposition and cursor position; therefore, leftward hand movements wererequired to hit targets appearing on the right. During the non-reversaltrials, the relationship between hand and cursor was represented by astick connecting hand position and cursor that translated with handmovement. In the No-Vision condition, task dynamics remained identicalto the Vision condition, but the virtual tool was not made visible toparticipants. As expected, presentation of the additional visualinformation significantly reduced reaction times during visuomotorreversal trials. A second experiment was conducted to determinewhether this gain in performance was maintained during ‘fast’corrections to a target jumping location upon movement onset on 20%of trials. It was found that the presentation of tool-like visual informationdecreased the time needed to correct the movement trajectory andincreased the accuracy of those corrections when the targetunexpectedly jumped location. Taken together, these results suggestthat visual information alone may be adequate to engage versatile tool-use neuronal mechanisms to increase task performance in a wide rangeof settings.
Multisensory recalibration in motor controlDenise Henriques1, Erin K Cressman2, Danielle Salomonczyk1
1York University, 2University of Ottawa
In this talk, I will discuss recent studies from my lab investigating howthe brain uses information from one sense to recalibrate other senses inorder to produce consistent representations for motor control. Morespecifically, this research asks whether recalibrating the arm motorsystem based on information from one sense, like vision, alsorecalibrates other senses such as proprioception. From previousresearch, we know that when subjects reach to visual targets withaltered visual feedback of the hand, subjects learn to adjust their handmovements in order to get the hand (or cursor) onto the target. We haverecently shown that this visuomotor adaptation also affects subjects’perceptual judgments regarding the location of their unseen handposition, even when estimating hand position after its actively versuspassively moved. That is, learning to reach with altered visual feedbackof the hand not only leads to motor recalibration, but also partialproprioceptive recalibration of the hand in the direction of thevisuomotor distortion. We have found that proprioception is recalibratedto a similar extent across the life-span and for patients with Parkinsondiseases. The extent of this recalibration is independent of the degree ofproprioceptive sensitivity. Moreover, we have found that mere exposureto a sensory discrepancy between the seen and felt hand position whenthe hand is passively moved also leads to motor and proprioceptiverecalibration of the hand. Taken together, our results suggest thatsensorimotor learning affects not only motor output and the predictedsensory consequences but also affects non-visual estimates of statichand position. In this talk, I will show evidence that suggests that thesemotor and sensory recalibrations (or plasticity) are separate processes,which rely on different error signals, including sensory discrepanciesbetween visual and proprioceptive estimates of hand position.Specifically, our results indicate that the patterns of motor and sensoryrecalibration tend to differ for bimanual transfer and generalizationacross the workspace. Even when these changes in movement andproprioceptive estimates consistently change, as in the case when wevary the magnitude of the visuomotor distortion, the motor and sensoryoutput do not correlate. These studies expand our understanding of theinteraction of vision and body-position sense in shaping the brain’smotor-control systems, as well as provide insight into the type of errorsignals and mechanisms that may underlie learning.
The nature of the multiple processes underlyingmotor learning in adaptation to velocity-dependentcurl fieldsJon Kennedy1, Chris Miall1
1University of Birmingham
What types of learning underlie adaptive behaviour in velocity-dependent curl fields? Smith, Ghazizadeh and Shadmehr (2006: PLOSBiology) have shown (using an adaptation, deadaptation, and error-clamp procedure) that adaptive performance during a reaching to targettask in a curl field is better explained by a model with two separatelearning modules with different timescales, than by a model with asingle learning module. However, Smith et al.’s (2006) model is silent onthe nature of the two learning modules (what is learnt by each of themodules). In adaptation to rotation of the visual field (prism adaptation),a combination of perceptual and non-perceptual adaptation underliesthe adaptive behavioural response, and the two types of adaptationhave different timescales (see Redding, Rossetti, & Wallace, 2005:Neuroscience and Biobehavioral Reviews, for a review). Ostry, Darainy,Mattar, Wong, and Gribble (2010: Journal of Neuroscience) found thatperception of the direction of near-to-far hand movements was shiftedclockwise following active adaptation to a clockwise curl field. Wesought to investigate whether this perceptual response in force fieldadaptation was associated with either the fast or the slow learningmodule identified by Smith et al. (2006). Thus, we combined Smith etal.’s (2006) and Ostry et al.’s (2010) procedures in order to correlateacross participants the (modelled) quantity of learning in the fast andslow learning modules with the size of the perceptual aftereffects. In our

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 23
data, Smith et al.’s (2006) results replicated: a dual processes modelbetter explained the data than a single process model. However, Ostry etal.’s (2010) perceptual aftereffects were not detected; nor were theresignificant correlations between the perceptual measures and thelearning in the fast and slow learning modules. Moreover, a series ofexperiments designed, first, to directly replicate Ostry et al.’s (2010)procedure alone, and, then, to modify that procedure to enhance theputative perceptual effect, did not replicate that effect. Hence, wetentatively conclude that both of the processes identified by Smith etal.’s (2006) model are behavioural in nature. In previous studies ofadaptation to curl fields (e.g. Smith et al.’s, 2006), behaviouraladaptation has been modelled under the assumption that participantscorrectly identified the nature of the disturbance (a velocity-dependentcurl field) and compensated accordingly (qualitatively optimally, but tovarying degrees). On the contrary, our data indicate that participantsemploy a range of sub-optimal strategies in response to the curl field.Thus, finally, we present an analysis of the types of strategic responsesadopted by participants, using system identification and computationalmodelling techniques.
Necessity is the mother of invention: Reconstructingmissing sensory information for concurrentcomparisons in multiple reference frames for eye-hand coordinationMichele Tagliabue1, Joseph McIntyre2
1Université Paris Descartes, 2CNRS
When aligning the hand to grasp an object, the CNS can combinemultiple sensory inputs encoded in multiple reference frames. Previousstudies have proposed how sensory information is selected, combinedand reconstructed. Distributed representations of spatial information indifferent reference frames and for different modalities would be createdin neural networks in the parietal cortex, whose recurrent structurewould ensure coherency between redundant internal representationsand allow for the reconstruction of sensory inputs that may be lacking.The same neural networks could also be involved in the optimalweighting of different sources of information to minimize perceptualuncertainty. Despite the remarkable capacities of these neural structures,recent experimental results in the literature suggest that when a directcomparison of target and hand is possible via a single sensory modality,the CNS avoids performing unnecessary coordinate transformations thatadd noise. In natural conditions, however, a direct comparison betweenstimulus and response is not always possible: for instance, when targetand hand do not share a common sensory modality (e.g., aligning theunseen hand to a visual target) or when body movements occurbetween target acquisition and manual response. Since sensorimotorintegration in these conditions has not been extensively investigated inthe literature, we studied the behavior of subjects who aligned the handto an external (visual or kinesthetic) target, when the feedback about thehand was visual, kinesthetic, or both; and we asked them to perform, ornot, head movements between the target memorization and responsephases. Moreover, rather than introducing conflict between the sensorysignals describing the target or hand themselves, as in previous studies,we instead introduced conflict between the reference frames used toencode their orientations in memory. With this technique we were ableto study situations in which it is impossible to perform the task usingonly a single sensory modality. By comparing experimental results toanalytical models based on principles of maximum likelihood, weshowed that brain does not compare optimal estimates of the target andhand orientation, as previously hypothesized, but instead optimallycombines the results of multiples comparisons performed in parallel indifferent modalities. Moreover, we demonstrated that CNS favors directcomparisons of egocentric sensory information whenever possible, inaccord with principles of maximum likelihood, but does not avoidreconstructing missing information at all costs. On the contrary, itappears that when at least one transformation of orientation informationis required, the reconstruction of other missing sensory information isbetter tolerated or even encouraged, perhaps because the additionalreconstruction is no more costly in terms of noise. Thus, for cross-modal tasks, or when head movements are involved, the CNS createsand uses both kinesthetic and visual representations, although themisalignment of the body with respect to vertical disrupts the
transformation of information between visual and kinesthetic sensation.We conclude that the necessity of producing at least one coordinatetransformation activates multiple, concurrent internal representations.
Measuring the generalization of visual perturbationsof wrist movements across movement directionsusing mobile phonesHugo Fernandes1, Mark Albert1, Konrad Kording11Northwestern University
In our lives we essentially never encounter the same situation twice. Dueto changes in the environment, our own body and our knowledge theproblems that we are solving are always different. Therefore,generalization is central to any behavior and measuring generalization isa promising behavioral window into neural representations and learningmechanisms. Recent studies in motor control have shown thatvisuomotor rotations for reaching have narrow generalization functions[1-3]: what we learn while moving in one direction only affectssubsequent movements into close directions. Here we wanted tomeasure the generalization functions for rotations of the wrist. To do sowe had 7 subjects hold a T-mobile G1 mobile phone in their hand. Thebuilt in acceleration sensor proved to be a convenient device tomeasure wrist movements during the behavioral protocol and itsprecision was assessed using a 3D Investigator Position Sensor(Optotrak). Subjects moved a cursor on the screen of the mobile phoneby tilting the wrist. Movements on the screen towards the training targetwere rotated and we measured how learning of this rotation in thetraining direction affected subsequent movements in other directions.We found that generalization is local and similar to generalizationpatterns of visuomotor rotation for reaching. Furthermore, the precisionof movements before the introduction of the perturbation showsanisotropy across directions. Movements to targets close to the medialand lateral directions were biased toward the distal direction byapproximately 10o. This bias appears to be relatively large incomparison to the typical standard deviation of movement and it mightreflect biomechanical effects or biases in perception. Most importantlywe established the use of mobile phones to run movement experimentsin motor control. Our application adds to the growing literature of usingmobile phones in medical contexts [4-6]. Cheaper and versatile ways ofcollecting data like in this study can make the recording and applicationof movement data much more ubiquitous. From clinical population, topopulations in hard to reach areas of the world, mobile phones providea useful tool for studying and using movement data in the future. 1. Paz,R., et al., Preparatory activity in motor cortex reflects learning of localvisuomotor skills. Nature neuroscience, 2003. 6(8): p. 882-890. 2.Krakauer, J., et al., Learning of visuomotor transformations for vectorialplanning of reaching trajectories. Journal of Neuroscience, 2000. 20(23):p. 8916. 3. Mattar, A. and D. Ostry, Modifiability of generalization indynamics learning. Journal of neurophysiology, 2007. 98(6): p. 3321. 4.Brezmes, T., J. Gorricho, and J. Cotrina, Activity recognition fromaccelerometer data on a mobile phone. Distributed Computing, ArtificialIntelligence, Bioinformatics, Soft Computing, and Ambient AssistedLiving, 2009: p. 796-799. 5. Mathews, A. and R. Butler. A vision for theuse of proactive mobile computing tools to empower people withchronic conditions. 2005: IEEE. 6. Rohs, M. and B. Gfeller, Using camera-equipped mobile phones for interacting with real-world objects.Advances in Pervasive Computing, 2004: p. 265-271.
Is there a natural speed of reaching movements?Britne Shabbott1, Pietro Mazzoni1
1Columbia University
The speed of our movements is a prominent aspect of our motorbehavior. While we are not much aware of movement speed when it isnormal, we easily notice movements that are unusually slow or fast. Thissuggests that movement speed might be a precisely controlled aspect ofmovement. Typically, the accuracy requirements of a task impose anupper boundary to how fast we can move while still being successful.However, natural reaching behaviors are regularly performed withmovement speeds that are well below the boundary for task accuracy,

24 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM
and yet they are not usually performed as slowly as possible. Such apreference for moving at certain speeds suggests the existence of anatural speed of movement. We hypothesized that natural movementspeed is carefully controlled by the nervous system and we developed aseries of experiments to determine 1) the existence and constancy ofnatural movement speeds, and 2) possible principles underlying naturalspeed selection. We first assessed natural reaching speed to a fixedtarget in the naive state. Within subject, the naive natural speed hadsmall variability compared to the range of speeds that could lead to tasksuccess. We then tested what speed subjects freely selected afterexperiencing other speeds. Subjects completed alternating blocks inwhich they reached either at their preferred speed or at an imposedspeed, chosen from a wide range. We found that preferred speed wasinfluenced, but not dictated by recent speed: the slope of therelationship between recent and preferred speed was less than 1, whichindicates that subjects still preferred a particular speed in spite of theinfluence of recent experience. We inferred this speed from the linear fitas the point where the preferred speed is the same as the recentlyimposed speed. This calculated natural speed reflects subjects’preference in the context of experiencing different speeds. Remarkably,the calculated natural speed was comparable to the naive natural speedfor each subject. These results demonstrate the existence of a naturalspeed of reaching movements that is reasonably consistent for eachsubject. We next considered whether optimization of movementkinematics might mediate speed selection. We found that velocityprofiles during natural reaching conditions were symmetrical, with peakvelocity occurring at the midpoint of each reach. Conversely, position atpeak velocity occurred prior to and following midpoint for slower andfaster movements, respectively. In addition, velocity profiles had moresubmovements, after the initial peak, for faster and slower movementsthan for natural-speed movements. These findings reveal a specificvelocity profile associated with natural speed, and suggest that naturalspeed may be selected so as to minimize online corrections. By varyingtarget distance and size, we found a Fitts-like linear relationship betweenmovement time and log(2 x target amplitude/width) for naturalmovements. Unlike Fitts’ law, in which speed reflects performance limits,this relationship reflects speed selection processes. These results areevidence for a “main sequence” governing reaching movements, which,unlike the saccade main sequence, is preferred and not obligatory.
PANEL Session 12Saturday, April 308:00AM – 10:15AMThinking without the boxes: Towards an integratedview of sensorimotor integrationEmanuel Todorov1, William Kristan2, Mark Churchland, John Kalaska
1University of Washington, 2University of California San Diego
“Understanding” often means breaking a system into pieces anddrawing labeled boxes around them. However the sensorimotor systemmay be too tightly integrated to be understood in this way, even whenstudied in artificial settings that promote boxology. Here we will discussrecent work pointing to an alternative view. Traditional distinctions suchas perception-cognition-action and decision-planning-execution are stilluseful, but no longer taken literally to mean separate computationsperformed in series by distinct brain areas. Instead we see them asdifferent aspects of the distributed neural dynamics that shape the flowof sensorimotor interactions with the environment. While this view isbroadly consistent with ecological and dynamical systems accounts, weseek to avoid the sense of mysticism sometimes present in suchaccounts, and instead be more specific about the neural mechanismsand the underlying computational principles. John Kalaska will critiquethe information-processing framework for interpreting neural function.This framework was originally inspired by cognitive psychology studiesof abstract problem solving, however the brain did not evolve originallyto play chess. Its ultimate purpose for millions of years has been topermit an individual to engage in real-time interaction with its
environment, enhancing survivability. He will review diverseneurophysiological data that are difficult to reconcile with the traditionalperception-cognition-action cascade of information processing, andappear to be more compatible with an alternative ethologically-inspiredview of interactive behavior as simultaneous processes that specifypotential motor actions and select between them. Bill Kristan willdiscuss the evidence that neurons responsible for producing one motorbehavior in the leech also contribute to the production of otherbehaviors. Simultaneous recordings from over 100 of these neuronsduring multiple behaviors show that (1) a substantial fraction of theneurons are active during more than one behavior; (2) the same neuroncan have more than one function during a single behavior; and (3) agroup of neurons can make the choice between two behaviors on thebasis of subtle variations in their joint activity patterns. None of theseobservations fit neatly into functional boxes, and instead point to acomplex and integrated dynamical system which is yet to be fullyunderstood. Mark Churchland will present results that challengetraditional thinking regarding the neural bases of planning andexecution. He will argue for an emerging paradigm that viewspreparatory activity not as a code for desired movement parameters, butrather as the initial state of a dynamical system. It will be argued that thedynamical system in question is reasonably simple, and involves thegeneration of one or more oscillatory rhythms, the sum of whichproduces a motor command. Emo Todorov will describe new optimalcontrol models where execution is the by-product of continuous re-planning. The movement plan is re-optimized every few milliseconds, soas to take into account perturbations and changes in the dynamics ortask goals. In this way the system can generate intelligent responses innovel circumstances, giving rise to apparent motor creativity. The newmodels produce complex dynamic behaviors: bouncing multiple ping-pong balls (on real robots), swimming to randomly-changing targets,walking and running and hopping.
PANEL Session 13Saturday, April 3010:45AM – 1:00PMThe role of biomechanics in the neural control ofmultijoint posture and movement: What remains forthe neural controller?Eric Perreault1, Lena Ting2, Matthew Tresch1, David Franklin3
1Northwestern University, 2Emory University and Georgia Tech,3Cambridge University
Generating coordinated movements and postures requires interactionbetween the nervous and musculoskeletal systems. This session willexplore how properties of the musculoskeletal system influencebehaviors that can be generated through neural control, and how thenervous system might exploit such properties. Examples will beprovided in which musculoskeletal biomechanics can either simplifyneural control, or limit its possible actions. A unifying theme will be thatactions of the nervous system depend strongly on body configurationand that postures can be selected to either increase or decreasedependence on the neural controller. The session will have fourpresentations, covering feedforward and feedback control in the upperand lower limbs to demonstrate the generality of the presentedconcepts. Matt Tresch will present work characterizing the configurationdependent actions of muscles (active and passive) in the rat hindlimband the relationship between these actions and muscle recruitment incomplex behaviors. Eric Perreault will present work demonstrating howthe intrinsic stiffness of individual muscles contributes to the multijointmechanics of the human arm. Intrinsic muscle properties can be tunedthrough changes in feedforward control of muscle activation (e.g.selective co-contraction). Hence, these results clarify the extent to whichfeedforward strategies can regulate arm mechanics at a given posture.Selecting arm posture allows further tuning of arm mechanics. DavidFranklin will examine how limb posture and neural control can be

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 25

26 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 27
adapted to improve energy efficiency. Arm postures are selected toprovide efficiency in the presence of force variability. When posturecontrol is limited, the CNS adapts arm mechanics to the environmentalinstability, in part through the tuning of feedback gains. Lena Ting willshow how changes in body posture during standing constrain thefeedback gains that can be used to stabilize the body in the face ofperturbations. Interactions between the dynamics of different bodyconfigurations and physiological delayed feedback control impose non-intuitive constraints and requirements on the actions of the nervoussystem, resulting in predictions about variability, modulation, selectionof neural feedback gains and strategy-switching (e.g. stepping) acrossconfigurations. Together these presentations will illustrate the need tounderstand and characterize the motor system’s biomechanicalcapabilities before meaningful conclusions about neural signals andneural control mechanisms can be made. We will leave time fordiscussion to address the common principles for motor control that arerevealed in these studies. Motivation and goals While it has long beenknown that the nervous and musculoskeletal systems interact toproduce movement, this session is timely in that is brings quantifiablepredictions to the study of motor control. Moreover, it is becoming clearthat ideas generated from upper and lower limb studies are convergingtowards common principles of neuromechanical control and theirrelationship to important topics such as variability, efficiency, andoptimality. These investigators have not presented an NCM paneltogether; Tresch and Ting were last on the same panel in 2003. We allmet at SfN to organize the proposed panel, and hope our selection willstimulate more widespread awareness and incorporation ofneuromechanical concepts in the NCM community.
PERSPECTIVE Session 14Saturday, April 303:00PM – 4:30PMSharing of sensory and motor signals in eye-headcoordinationHenrietta Galiana1, John Van Opstal2, Daniel Guitton3
1McGill University, 2Radboud University, 3MNI & McGill University
Years ago Dan Guitton organized a session for NCM where he posed thequestion ‘Is the head an eye or a limb’. Of course he was referring to thecontrol strategies then being proposed for limbs (requiring optimaltrajectory planning), and the simpler ballistic ones for eye saccades.
Today, such questions still remain without resolution in the communityand the number of theories and theoretical models continues to grow inboth limb and gaze control. In this session, we will re-examine severalaspects of eye-head coordination during gaze reorientation, in the lightof recent experimental observations. The resulting inferences arecompatible with evolving motor control models for posture andperipheral limbs, but do not require at any time detailed a prioritrajectory plans in the absence of obstacles. Three main areas are nowmature enough to reach a scientific consensus. 1. Does the SuperiorColliculus (SC) error map control gaze in open loop or closed loop?John Van Opstal will describe how the SC encodes multi-sensoryevoked eye-head coordination. He will present data on auditory- andaudiovisual evoked gaze shifts that argue for a dynamically updatedworld-centered goal representation, from which appropriate eye andhead control signals are derived. He will use a simple computationalmodel to replicate population encoding of normal and slow gaze shiftsfrom single-cell recordings, and explores whether the results canresolve the arguments over SC loop control. 2. Can the eyes fullycompensate for head perturbations during orienting gaze shifts? DanielGuitton will describe the coordination and mutual information exchangebetween the saccadic, head and vestibular systems during orientinghuman head-unrestrained gaze shifts, in which the head is subjected tounpredictable long-duration torques. He will show that the saccadic andvestibular systems coordinate to compensate for perturbations in orderto maintain the gaze trajectory invariant relative to control. Theobservations imply that vestibular signals continue to play acompensatory role even during gaze saccades. 3. How can brainstennetworks support compensation for perturbations during orienting gazeshifts? Mimi Galiana will present an updated model of gaze control forhead-free gaze shifts, which tightly inter-couples all sensory and motorsignals. To replicate recent data from Dan Guitton’s lab, it is necessary toallow a compensatory role for all sensory afferents (e.g. vestibular)throughout the saccadic and fixation intervals of a gaze shift. At least insome head perturbation directions (assisting torques), gaze trajectoriesand accuracy can remain very robust in humans - hence one can nolonger assume the VOR is off during saccades and that no othervestibular signal is available. We propose that the vestibular-only (VO)cells in vestibular nuclei described by Roy & Cullen (J. Neurosci. 2004)play an important role in maintaining gaze trajectory invariance. Hencetrajectory invariance need not require pre-planning. The final discussionwill highlight those points of agreement shared by the speakers, withcomments from the audience. The intent is to realize a schematicframework on which all future models can be built, tested or extended,rather than continue to elaborate divergent points of view. Links to limbcontrol and the role of spinal and vestibular reflexes in trajectoryevolution should also become clear.
Notes

28 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM
Mind-Fi is the closest thing tomental telepathy you’ve ever seen.
Introducing the
Mind-Fi Wireless 63 ChannelActive EEG
System
Mind-Fi is a wireless 63 channel active EEG head-cap that enables participants to
project their thoughts in real-time anywhere in a coverage area of 20x20 feet (6x6 meters).
Active electronics on the cap eliminate the need for scalp abrasion resulting in cap setup
time of only 5-10 minutes. �e cap weighs only a few ounces, so it’s comfortable to wear
and it will stay put as subjects move through the environment. To wash, just remove the
tiny transmitter and place it on the charger.
�ere’s also a small wireless access point to instantly receive the projected thoughts, flexible
data acquisition hardware and powerful, easy-to-install data acquisition software for any
Windows platform. Mind-Fi is suited to EEG, ERP and even BCI studies.
Features and Benefits:
• Quick setup and unexpected signal quality: Active head-stage on the cap, battery power and electrical isolation
• Compact: 63 channels on the cap and receiver
• Easy to install on any computer:USB 2.0 interface with Windows, MAC and Linux drivers
www.cortechsolutions.com • [email protected] • 910.362.1143

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 29
Scholarship Winners
New investigators and faculty are essential for the future ofany field of scientific inquiry. NCM has historicallyencouraged meeting participation by graduate students andpost-doctoral fellows. The scholarship program is designed toprovide partial support for them to participate in the meetingand is open to student and post-doc members in good
standing. Our scholarship program is funded through thesupport of our sponsors. Also as a result of residuals from anNIH R13 grant to last year’s Satellite Meeting organizer, RajGandhi, University of Pittsburgh, our scholarship programwas able to fund four additional students this year.
Scholarship Winner Mentor
Nadia Alahyane Centre for Neuroscience Studies, Queen’s University Douglas Munoz
James Bonaiuto Andersen Laboratory, California Institute of Technology Richard Andersen
Jordan Brayanov Neuromotor Contol Lab, Harvard University Maurice Smith
Jessica Brooks Department of Physiology, McGill University Kathleen Cullen
Gavin Buckingham Centre for Brain and Mind, University of Western Ontario Melvyn Goodale
Gabriela Cantarero Johns Hopkins University, Human Brain Physiology Pablo Celnikand Stimulation Laboratory
Claudia Casellato, Neuroengineering and medical robotics Laboratory, Politecnico di Milano Giancarlo Ferrigno
Tanuj Gulati Drexel University College of Medicine Simon Giszter
Helen Huang Neuromechanics Lab, University of Colorado at Boulder Alaa A. Ahmed
Matthew Kaufman Neurosciences Graduate Program, Stanford University Krishna Shenoy
Trisha Kesar Physical Therapy Department, University of Delaware Darcy Reisman/Stuart Binder-Macleod
Femke Maij Faculty of Human Movement Science, VU University Amsterdam Jeroen Smeets
Laura Malone Biomedical Engineering Department, Johns Hopkins University Amy Bastien
Alejandro Melendez-Calderon Imperial College of Science Technology and Medicine Etienne Burdet
Jean-Jacques Orban de Xivry Biomedical Engineering and Neuroscience, Reza ShadmehrJohns Hopkins School of Medicine
Muriel Panouilleres Centre de Recherche en Neurosciences de Lyon Denis Pelisson
Myrthe Plaisier VU University in Amsterdam Jeroen Smeets
Jinsook Roh Rehabilitation Institute of Chicago and Northwestern University William Rymer, Randall Beer,Eric Perreault
Britne Shabbott Motor Performance Laboratory, Columbia University Medical Center Pietro Mazzoni
Natela Shanidze Neuroscience Program, University of Michigan Michael King
Michele Tagliabue Centre d’Etude de la Sensorimotricité, Université Paris Descartes Joseph McIntyre
Jordan Taylor University of California, Berkeley Richard Ivry
Evangelos Theodorou Brain and Body Dynamics Lab, University of Southern California Francisco Valero-Cuevas
Paul Wanda Biomedical Engineering, Washington University in St. Louis Kurt Thoroughman
NCM has many generous supporters who haveprovided a variety of items for our inaugural silentauction. Check it out, you’ll find gift certificates forlocal restaurants and attractions and other items ofinterest to the NCM community. If you have anitem you wish to donate to the auction, it’s not toolate to do so – please bring it to the RegistrationDesk and we’ll add it to the auction.
Bidding is open Wednesday from 8:00 AM –
3:00 PM and on Thursday between 8:00 and 6:00.Bid once! Bid many times! Bid before the auctioncloses following the NCM Business Meeting andthen enjoy your winnings for the rest of themeeting.
The Silent Auction table is located next tothe Registration Desk. All proceeds go towardssupporting the NCM’s Scholarship Fund. Yoursupport helps support our future.
NCM SilentAuction
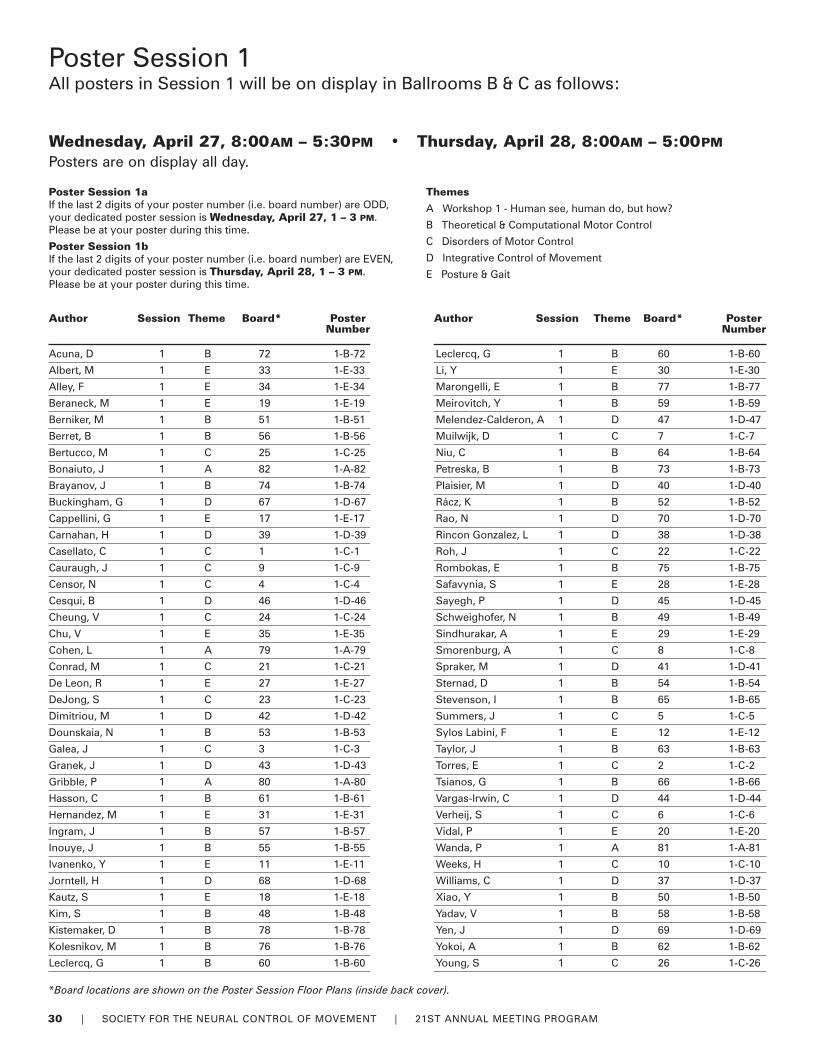
30 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM
Poster Session 1All posters in Session 1 will be on display in Ballrooms B & C as follows:
Author Session Theme Board* Poster Author Session Theme Board* PosterNumber Number
Acuna, D 1 B 72 1-B-72 Leclercq, G 1 B 60 1-B-60
Albert, M 1 E 33 1-E-33 Li, Y 1 E 30 1-E-30
Alley, F 1 E 34 1-E-34 Marongelli, E 1 B 77 1-B-77
Beraneck, M 1 E 19 1-E-19 Meirovitch, Y 1 B 59 1-B-59
Berniker, M 1 B 51 1-B-51 Melendez-Calderon, A 1 D 47 1-D-47
Berret, B 1 B 56 1-B-56 Muilwijk, D 1 C 7 1-C-7
Bertucco, M 1 C 25 1-C-25 Niu, C 1 B 64 1-B-64
Bonaiuto, J 1 A 82 1-A-82 Petreska, B 1 B 73 1-B-73
Brayanov, J 1 B 74 1-B-74 Plaisier, M 1 D 40 1-D-40
Buckingham, G 1 D 67 1-D-67 Rácz, K 1 B 52 1-B-52
Cappellini, G 1 E 17 1-E-17 Rao, N 1 D 70 1-D-70
Carnahan, H 1 D 39 1-D-39 Rincon Gonzalez, L 1 D 38 1-D-38
Casellato, C 1 C 1 1-C-1 Roh, J 1 C 22 1-C-22
Cauraugh, J 1 C 9 1-C-9 Rombokas, E 1 B 75 1-B-75
Censor, N 1 C 4 1-C-4 Safavynia, S 1 E 28 1-E-28
Cesqui, B 1 D 46 1-D-46 Sayegh, P 1 D 45 1-D-45
Cheung, V 1 C 24 1-C-24 Schweighofer, N 1 B 49 1-B-49
Chu, V 1 E 35 1-E-35 Sindhurakar, A 1 E 29 1-E-29
Cohen, L 1 A 79 1-A-79 Smorenburg, A 1 C 8 1-C-8
Conrad, M 1 C 21 1-C-21 Spraker, M 1 D 41 1-D-41
De Leon, R 1 E 27 1-E-27 Sternad, D 1 B 54 1-B-54
DeJong, S 1 C 23 1-C-23 Stevenson, I 1 B 65 1-B-65
Dimitriou, M 1 D 42 1-D-42 Summers, J 1 C 5 1-C-5
Dounskaia, N 1 B 53 1-B-53 Sylos Labini, F 1 E 12 1-E-12
Galea, J 1 C 3 1-C-3 Taylor, J 1 B 63 1-B-63
Granek, J 1 D 43 1-D-43 Torres, E 1 C 2 1-C-2
Gribble, P 1 A 80 1-A-80 Tsianos, G 1 B 66 1-B-66
Hasson, C 1 B 61 1-B-61 Vargas-Irwin, C 1 D 44 1-D-44
Hernandez, M 1 E 31 1-E-31 Verheij, S 1 C 6 1-C-6
Ingram, J 1 B 57 1-B-57 Vidal, P 1 E 20 1-E-20
Inouye, J 1 B 55 1-B-55 Wanda, P 1 A 81 1-A-81
Ivanenko, Y 1 E 11 1-E-11 Weeks, H 1 C 10 1-C-10
Jorntell, H 1 D 68 1-D-68 Williams, C 1 D 37 1-D-37
Kautz, S 1 E 18 1-E-18 Xiao, Y 1 B 50 1-B-50
Kim, S 1 B 48 1-B-48 Yadav, V 1 B 58 1-B-58
Kistemaker, D 1 B 78 1-B-78 Yen, J 1 D 69 1-D-69
Kolesnikov, M 1 B 76 1-B-76 Yokoi, A 1 B 62 1-B-62
Leclercq, G 1 B 60 1-B-60 Young, S 1 C 26 1-C-26
*Board locations are shown on the Poster Session Floor Plans (inside back cover).
Poster Session 1aIf the last 2 digits of your poster number (i.e. board number) are ODD,your dedicated poster session isWednesday, April 27, 1 – 3 PM.Please be at your poster during this time.
Poster Session 1bIf the last 2 digits of your poster number (i.e. board number) are EVEN,your dedicated poster session is Thursday, April 28, 1 – 3 PM.Please be at your poster during this time.
Themes
A Workshop 1 - Human see, human do, but how?
B Theoretical & Computational Motor Control
C Disorders of Motor Control
D Integrative Control of Movement
E Posture & Gait
Wednesday, April 27, 8:00AM – 5:30PM • Thursday, April 28, 8:00AM – 5:00PMPosters are on display all day.
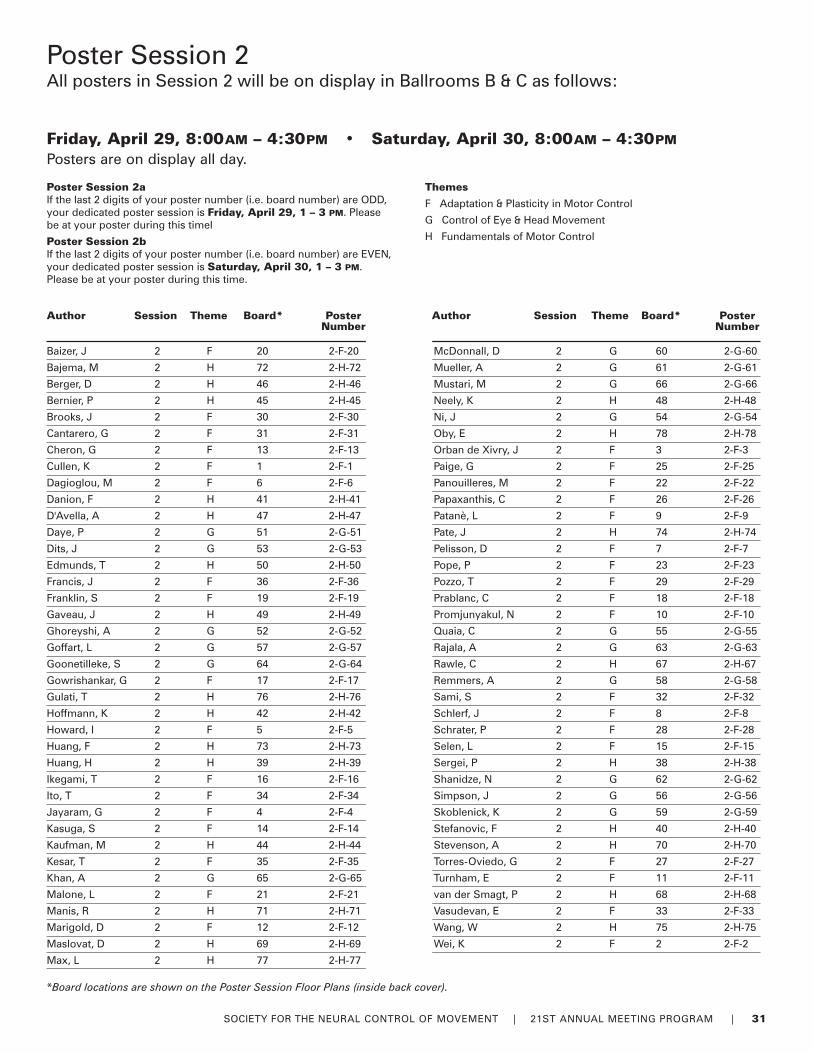
SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 31
Author Session Theme Board* Poster Author Session Theme Board* PosterNumber Number
Baizer, J 2 F 20 2-F-20 McDonnall, D 2 G 60 2-G-60
Bajema, M 2 H 72 2-H-72 Mueller, A 2 G 61 2-G-61
Berger, D 2 H 46 2-H-46 Mustari, M 2 G 66 2-G-66
Bernier, P 2 H 45 2-H-45 Neely, K 2 H 48 2-H-48
Brooks, J 2 F 30 2-F-30 Ni, J 2 G 54 2-G-54
Cantarero, G 2 F 31 2-F-31 Oby, E 2 H 78 2-H-78
Cheron, G 2 F 13 2-F-13 Orban de Xivry, J 2 F 3 2-F-3
Cullen, K 2 F 1 2-F-1 Paige, G 2 F 25 2-F-25
Dagioglou, M 2 F 6 2-F-6 Panouilleres, M 2 F 22 2-F-22
Danion, F 2 H 41 2-H-41 Papaxanthis, C 2 F 26 2-F-26
D'Avella, A 2 H 47 2-H-47 Patanè, L 2 F 9 2-F-9
Daye, P 2 G 51 2-G-51 Pate, J 2 H 74 2-H-74
Dits, J 2 G 53 2-G-53 Pelisson, D 2 F 7 2-F-7
Edmunds, T 2 H 50 2-H-50 Pope, P 2 F 23 2-F-23
Francis, J 2 F 36 2-F-36 Pozzo, T 2 F 29 2-F-29
Franklin, S 2 F 19 2-F-19 Prablanc, C 2 F 18 2-F-18
Gaveau, J 2 H 49 2-H-49 Promjunyakul, N 2 F 10 2-F-10
Ghoreyshi, A 2 G 52 2-G-52 Quaia, C 2 G 55 2-G-55
Goffart, L 2 G 57 2-G-57 Rajala, A 2 G 63 2-G-63
Goonetilleke, S 2 G 64 2-G-64 Rawle, C 2 H 67 2-H-67
Gowrishankar, G 2 F 17 2-F-17 Remmers, A 2 G 58 2-G-58
Gulati, T 2 H 76 2-H-76 Sami, S 2 F 32 2-F-32
Hoffmann, K 2 H 42 2-H-42 Schlerf, J 2 F 8 2-F-8
Howard, I 2 F 5 2-F-5 Schrater, P 2 F 28 2-F-28
Huang, F 2 H 73 2-H-73 Selen, L 2 F 15 2-F-15
Huang, H 2 H 39 2-H-39 Sergei, P 2 H 38 2-H-38
Ikegami, T 2 F 16 2-F-16 Shanidze, N 2 G 62 2-G-62
Ito, T 2 F 34 2-F-34 Simpson, J 2 G 56 2-G-56
Jayaram, G 2 F 4 2-F-4 Skoblenick, K 2 G 59 2-G-59
Kasuga, S 2 F 14 2-F-14 Stefanovic, F 2 H 40 2-H-40
Kaufman, M 2 H 44 2-H-44 Stevenson, A 2 H 70 2-H-70
Kesar, T 2 F 35 2-F-35 Torres-Oviedo, G 2 F 27 2-F-27
Khan, A 2 G 65 2-G-65 Turnham, E 2 F 11 2-F-11
Malone, L 2 F 21 2-F-21 van der Smagt, P 2 H 68 2-H-68
Manis, R 2 H 71 2-H-71 Vasudevan, E 2 F 33 2-F-33
Marigold, D 2 F 12 2-F-12 Wang, W 2 H 75 2-H-75
Maslovat, D 2 H 69 2-H-69 Wei, K 2 F 2 2-F-2
Max, L 2 H 77 2-H-77
*Board locations are shown on the Poster Session Floor Plans (inside back cover).
Poster Session 2All posters in Session 2 will be on display in Ballrooms B & C as follows:
Poster Session 2aIf the last 2 digits of your poster number (i.e. board number) are ODD,your dedicated poster session is Friday, April 29, 1 – 3 PM. Pleasebe at your poster during this timel
Poster Session 2bIf the last 2 digits of your poster number (i.e. board number) are EVEN,your dedicated poster session is Saturday, April 30, 1 – 3 PM.Please be at your poster during this time.
Themes
F Adaptation & Plasticity in Motor Control
G Control of Eye & Head Movement
H Fundamentals of Motor Control
Friday, April 29, 8:00AM – 4:30PM • Saturday, April 30, 8:00AM – 4:30PMPosters are on display all day.

32 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 33
Session 1 Posters are listed by theme.
A - WorkshopHuman see, human do, but how?Team Leader: Kurt Thoroughman
Poster Presentations:
1-A-79 Leonardo G Cohen
1-A-80 Paul L Gribble
1-A-81 Paul A Wanda
1-A-82 James Bonaiuto
B - Theoretical & ComputationalMotor Control
1-B-48 Learning a new motor skill with a highdimension motor system: Preliminary resultsSung Shin Kim1, Nicolas Schweighofer1, Stefan Schaal1, RobertScheidt2
1University of Southern California, 2Marquette University
1-B-49 Arm choice in fast pointing movementsdepends on balance between inertial property of thearm and neural noiseNicolas Schweighofer1, Rieko Osu2, T. Yoshioka 2, Yupeng Xiao1
1University of Southern California, 2ATR
1-B-50 Multi-source execution noise predicts optimalmovement duration in reaching movements withoutvisual feedbackYupeng Xiao1, James Gordon1, Nicolas Schweighofer1
1University of Southern California
1-B-51 Optimality criteria generalize from lineardeterministic to stochastic and nonlinear settingsMax Berniker1, Megan O'Brien2, Konrad Kording1, Alaa Ahmed2
1Northwestern University, 2University of Colorado
1-B-52 A computational approach to quantifyingmuscle redundancy, and a sample application to guidethe detection of strategies to mitigate fatigueKornelius Rácz1, Jason Kutch1, Francisco Valero-Cuevas1
1University of Southern California
1-B-53 Preferred patterns of joint control duringunconstrained arm movementsNatalia Dounskaia1, Wanyue Wang1
1Arizona State University
1-B-54 State space analysis of intrinsic timing inthrowingDagmar Sternad1, Rajal Cohen2
1Northeastern University, 2Oregon Health and Science University
1-B-55 A comprehensive computational frameworkto evaluate grasp quality of tendon-driven hands witharbitrary topologyJoshua Inouye1, Jason Kutch1, Francisco Valero-Cuevas1
1University of Southern California
1-B-56 Reaching to a bar: How do we handle targetredundancy?Bastien Berret1, Enrico Chiovetto2, Francesco Nori1, Thierry Pozzo1
1Italian Institute of Technology, 2University Clinic Tübingen
1-B-57 A single-rate context-dependent processunderlies rapid adaptation to familiar object dynamicsJames Ingram1, Ian Howard1, J Randall Flanagan2, Daniel Wolpert1
1University of Cambridge, 2Queen’s University
1-B-58 A mathematical model of motor lateralizationVivek Yadav1, Robert Sainburg1
1Pennsylvania State University
1-B-59 Kinematic analyses of Israeli SignLanguage (ISL)Yaron Meirovitch1, Tamar Flash1
1Weizmann Institute of Science
1-B-60 Decoding the 3D visuomotor transformationof velocity signals for manual tracking using neuralnetworksGuillaume Leclercq1, Philippe Lefèvre1, Gunnar Blohm2
1Université catholique de Louvain, 2Queen's University
1-B-61 Learning to control dynamic objects withtime-varying energy constraintsChristopher Hasson1, Tian Shen1, Dagmar Sternad1
1Northeastern University
1-B-62 Flexible bimanual action coordinated bygain-field encoding of both arms' kinematics ininternal modelAtsushi Yokoi1, Masaya Hirashima1, Daichi Nozaki1
1Graduate School of Education, The University of Tokyo
1-B-63 Strategic generalization duringvisuomotor adapationJordan Taylor1, Richard Ivry1, Maurice Smith2, Jordan Brayanov2,Laura Hieber1
1University of California, Berkeley, 2Harvard University
1-B-64 Multi-scale highspeed motoneuron simulationusing FPGAChuanxin Niu1, Terrence Sanger1, Sirish Nandyala1
1University of Southern California
1-B-65 Statistical assessment of the stability ofsingle neuron movement representation in primarymotor cortexIan Stevenson1, Anil Cherian1, Brian London1, Jacob Reimer2, MarcSlutzky1, Konrad Kording1, Lee Miller1, Nicholas Hatsopoulos2, EricLindberg1, Nicholas Sachs1
1Northwestern University, 2University of Chicago
1-B-66 Biologically inspired learning algorithm forcontrolling biologically plausible neuro-musculoskeletal systemsGeorge Tsianos1, Gerald Loeb1, Jared Goodner1, Yao Li 1
1University of Southern California
Poster Sessions Authors, Titles and Affiliations

34 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM
1-B-72 The structure of visual scenes affectsperceived depths in a motor taskDaniel Acuna1, Konrad Koerding2, Ian Stevenson2
1Research Institute of Chicago, 2Rehabilitation Institute of Chicago
1-B-73 Movement curvature related tobiomechanical properties of the armBiljana Petreska1, Aude Billard2
1Harvard University, 2EPFL (École Polytechnique Fédéralede Lausanne)
1-B-74 Generalization patterns reveal thatvisuomotor rotation adaptation is composed of twodistinct components: Local shift learning and globalrotation learningJordan Brayanov1, Biljana Petreska1, Maurice Smith1
1Harvard University
1-B-75 Task-specific dynamics and motor babblingfor robotic hand controlEric Rombokas1
1University of Washington
1-B-76 Stiffness modulation in forceregulation tasksMax Kolesnikov1, Ferdinando Mussa-Ivaldi1, Kevin Lynch2
1Rehabilitation Institute of Chicago, 2Northwestern University
1-B-77 The advantage of flexible neuronal tunings inneural network models for motor learningEllisha Marongelli1, Ranjan Khan1, Kurt Thoroughman1
1Washington University in St. Louis
1-B-78 Testing predictions of different costfunctions for optimal control in arm movementsDinant Kistemaker1, Jeremy Wong1, Paul Gribble1
1University of Western Ontario
C - Disorders of Motor Control
1-C-1 Primary dystonia in children: How are motorcontrol and motor learning affected? Upper limbprotocols: Kinematics and EMGs to characterize thedisorder and force field paradigm to investigate thesensory-driven learning rateClaudia Casellato1, Giovanna Zorzi2, Alessandra Pedrocchi1, GiancarloFerrigno1, Nardo Nardocci2
1 Politecnico di Milano, 2 Fondazione IRCCS Istituto Neurologico CarloBesta
1-C-2 Larger impairment of endogenously-evokedautomated than exogenously-evoked intendedreaching modes in patients with Parkinson’s diseaseElizabeth Torres1, Kenneth Heilman2, Howard Poizner3
1Rutgers University, 2University of Florida, 3University of California,San Diego
1-C-3 Dopamine-dependent response to surprise inParkinson's diseaseJoseph Galea1, Sven Bestmann1, Mazda Beigi1, Marjan Jahanshahi1,John Rothwell1
1University College London
1-C-4 Impaired motor memory reconsolidation inpatients with chronic strokeNitzan Censor1, Leonardo Cohen1
1NIH
1-C-5 Age-related differences in attentional cost ofinterlimb coordinationJeffery Summers1, Hakuei Fujiyama1, Mike Garry1, Mark Hinder1
1University of Tasmania
1-C-6 Effects of early stage dementia onvisuomotor coordinationSimone Verheij1, Johan Pel1, Danya Muilwijk1, Hans van der Steen1
1Erasmus Medical Center
1-C-7 Visuomotor coordination in Parkinson'sdiseaseDanya Muilwijk1, Johan Pel1, Simone Verheij1, Hans van der Steen1
1Erasmus Medical Center
1-C-8 Visual feedback of the non-moving limbimproves active joint-position sense of the impairedlimb in Spastic Hemiparetic Cerebral PalsyAna Smorenburg1, Geert Savelsbergh2, Annick Ledebt3, FrederikDeconinck1
1Manchester Metropolitan University, 2Manchester MetropolitanUniversity / VU University, 3VU University
1-C-9 Total bimanual force productiondifferentiates movement variability and regularity inchronic stroke and age-matched controlsJames Cauraugh1, Neha Lodha1
1University of Florida
1-C-10 Effect of cerebellar damage on passiveversus active proprioception across Cartesian spaceHeidi Weeks1, Amy Bastian1
1Johns Hopkins University
1-C-21 Effects of movement velocity on handstrength after strokeMegan Conrad1, Derek Kamper2
1Rehabilitation Institute of Chicago / Northwestern University,2Rehabilitation Institute of Chicago / Illinois Institute of Technology
1-C-22 Alterations in upper limb synergy structureand activation in chronic stroke survivorsJinsook Roh1, Eric Perreault1, William Rymer1, Randall Beer1
1Rehabilitation Institute of Chicago / Northwestern University
1-C-23 The effect of movement speed on movementquality during reach-grasp-lift after stroke
Stacey DeJong1, Sydney Schaefer1, Catherine Lang1
1Washington University in St. Louis School of Medicine
Poster Sessions Authors, Titles and Affiliations

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 35
1-C-24 Is there a relationship between motorrecovery in stroke survivors and selectivereinforcement or attenuation of muscle synergyactivations?Vincent Chi Kwan Cheung1, Paolo Bonato2, Emilio Bizzi1, SabrinaPaganoni2, Patrick Kasi2, Andrea Valsecchi2
1MIT, 2Spaulding Rehabilitation Hospital
1-C-25 The cost function effects on the speed-accuracy trade-off in childhood dystonia using anovel assisted communication deviceMatteo Bertucco1, Terence Sanger1
1University of Southern California
1-C-26 The effects of transcranial stimulation onsymptoms of dystonia in childrenScott Young1, Terence Sanger1
1University of Southern California
D - Integrative Control of Movement
1-D-37 Colour-induced relationship between moodand reaching kinematics during a goal-directedaiming taskCamille Williams1, Heather Carnahan1, Lawrence Grierson2
1University of Toronto, 2McMaster University
1-D-38 The proprioceptive map which underliesmovement and tactile sensation is constructedindividuallyLiliana Rincon Gonzalez1, Stephen Helms Tillery1
1Arizona State University
1-D-39 Thunberg wetness illusion and the control ofgrip force at the fingers when grasping objects ofvarious temperaturesHeather Carnahan1, Camille Williams1, Adam Dubrowski2
1University of Toronto, 2Sick Children's Hospital
1-D-40 The size-mass illusionMyrthe Plaisier1, Jeroen Smeets1
1VU University
1-D-41 Cerebellar activity is related to forceamplitude but not rate of forceMatthew Spraker1, Daniel Corcos1, Stephan Swinnen2, DavidVaillancourt1, Janey Prodoehl1
1University of Illinois at Chicago, 2K.U.Leuven
1-D-42 Task-dependent cross limb reflexesMichael Dimitriou1, David Franklin1, Daniel Wolpert1
1University of Cambridge
1-D-43 The role of the superior parietal lobule inexplicit versus implicit control of decoupled visually-guided reachingJoshua Granek1, Laure Pisella2, Yves Rossetti2, Lauren Sergio1
1York University, 2INSERM U.864
1-D-44 Separation of visual object features andgrasp strategy in primate ventral premotor cortexCarlos Vargas-Irwin1, John Donoghue1, Lachlan Franquemont1,Michael Black2
1Brown University Neuroscience Department, 2Brown UniversityDepartment of Computer Science
1-D-45 LFP activity within PMd and SPL duringdecoupled visually-guided reachingPatricia Sayegh1, Kara Hawkins 1, Lauren Sergio1
1York University
1-D-46 Inter-individual variability in catchingkinematics: Evidence of multiple and non-stereotypedcontrol strategies?Benedetta Cesqui1, Francesco Lacquaniti1, Alessandro Portone1,Andrea d'Avella1
1IRCSS Santa Lucia Foundation
1-D-47 Mechanisms for disturbance attenuation inhuman-human collaborative tasksAlejandro Melendez-Calderon1, Victoria Komisar1, GowrishankarGanesh2, Etienne Burdet1
1Imperial College London, 2ATR Computational NeuroscienceLaboratories
1-D-67 Learning from mistakes: Improving initialfingertip force scaling by observing lifting errorsGavin Buckingham1, Paul Gribble1, Minnie Tang1, Melvyn Goodale1
1University of Western Ontario
1-D-68 Synaptic noise in motor output cells of thecerebellumHenrik Jorntell1, Fredrik Bengtsson1
1Lund University, Section for Neurophysiology
1-D-69 Sparse control of human locomotion:Evidence from hopscotchJasper Yen1, Young-Hui Chang1
1Georgia Institute of Technology
1-D-70 Visual and proprioceptive integration inmotor cortexNaveen Rao1, John Donoghue1
1Brown University
E - Posture & Gait
1-E-11 The development of spinal motor output forwalking in humansYuri Ivanenko1, Germana Cappellini1, Nadia Dominici1, RichardPoppele2, Francesco Lacquaniti3
1Santa Lucia Foundation, 2University of Minnesota, 3University ofRome Tor Vergata
1-E-12 Gait transitions in simulated reduced gravityFrancesca Sylos Labini1, Yuri Ivanenko1, Germana Cappellini1, VelioMacellari2, Joe McIntyre3, Francesco Lacquaniti4
1 Santa Lucia Foundation, 2Istituto Superiore di Sanità, 3UniversitéParis 5, 4University of Rome Tor Vergata

36 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM
KEY DATESCALL FOR ABSTRACTS
(opening dates)
Team (Multi-Author) October 28, 2011Individual (Oral & Poster) December 16, 2011
Registration will open in the fall of 2011.
www.ncm-society.org
DISCOVER THE MAGIC OF
VENICE, ITALY22ND ANNUAL NCM MEETING
Hilton Molino StuckyVenice, Italy
MOLINO STUCKYSPECIAL NCM GROUP RATES
Double Room (single occupancy) ! 209.00*Double Room (double occupancy) ! 239.00**Rate includes full buffet breakfast
Hotel reservations now open!Visit the website for reservation details.
Satellite MeetingApril 22 - 23, 2012
Annual MeetingApril 23 – 28, 2012

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 37
1-E-17 Functional topography of motor pools in thehuman spinal cord and the biomechanics oflocomotionGermana Cappellini1, Francesco Lacquaniti2, Yuri Ivanenko3, NadiaDominici3, Richard Poppele4
1Fondazione Santa Lucia, 2Santa Lucia Foundation and University ofRome Tor Vergata, 3Santa Lucia Foundation, 4University of Minnesota
1-E-18 Angular momentum based assessment ofdynamic balance performance in hemiparetic walkingSteven Kautz1, Richard Neptune2, Cameron Nott1
1Medical University of South Carolina, 2The University of Texas
1-E-19 Postural control and spatial orientation ofvestibular-deficient mice in the absence of gravityMathieu Beraneck1, Lionel Reveret2, Mathieu Rodriguez2, EstelleDuveau2, Anne Le Séach1, Pierre-Paul Vidal1
1CNRS - University Paris Descartes, 2INRIA Rhône-Alpes
1-E-20 The first five hundred milliseconds of a fallPierre-Paul Vidal1
1CNRS
1-E-27 Effects of robotic-assisted locomotor trainingand functional electrical stimulation in spinallycontused ratsRay De Leon1, Deborah Won1, Lauren Conn1, Sina Askari1, TeKangChao1, Connie Lee1
1Cal State LA
1-E-28 A common feedback model can explainmuscle activity in postural responses to discrete andcontinuous perturbationsSeyed Safavynia1, David Jilk2, Lena Ting3
1Emory University, 2Georgia Institute of Technology, 3Georgia Instituteof Technology / Emory University
1-E-29 Prenatal motor development for differentincubation periods affects postural control but notearly gaitAnil Sindhurakar1, Kornelius Racz1, Francisco Valero-Cuevas1, NinaBradley1
1University of Southern California
1-E-30 Human postural sway during quiet stance: Anonlinear optimal control perspectiveYao Li1, William Levine2, Gerald Loeb1
1University of Southern California, 2University of Maryland
1-E-31 Forward dynamic model of the momentumarrest phase of whole-body downward reachingmovements: Effects of age and functional impairmentManuel Hernandez1, James Ashton-Miller1, Neil Alexander1
1University of Michigan
1-E-33 Automated activity recognition andidentification using mobile phone accelerometry
Mark Albert1, Konrad Kording1
1Northwestern University
1-E-34 Contralateral contributions to ankle stiffnessduring human quiet stanceFerryl Alley1, P. J. Stapley1, R. E. Kearney1
1McGill University
1-E-35 Body weight supported treadmill walking maydecrease stabilityVirginia Way Tong Chu1, Brian Schmit2, T. George Hornby3
1Rehabilitation Institute of Chicago, 2Marquette University, 3Universityof Illinois at Chicago
Notes

38 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM
Session 2 Posters are listed by theme.
F - Adaptation & Plasticityin Motor Control
2-F-1 Internal models of self motion: Computationsthat suppress reafference in early vestibularprocessingKathleen Cullen1
1McGill University
2-F-2 Carrying water: The generalization functionof perturbations of the hand's weightKunlin Wei1, Konrad Koerding2, Ian Stevenson2, Xiang Yan1
1Peking University, 2Northwestern University
2-F-3 Corticospinal reorganization during force-field adaptationJean-Jacques Orban de Xivry1, Reza Shadmehr2, Michelle Harran2,Yousef Salimpour2, Mohammad Ali Ahmadi Pajouh2
1Université catholique de Louvain, 2Johns Hopkins University
2-F-4 Modulating locomotor adaptation withcerebellar stimulationGowri Jayaram1, Byron Tang1, Pablo Celnik1, Amy Bastian1
1Johns Hopkins University
2-F-5 Motor actions have a temporally-decayingaugmenting effect on state-dependent motor learningIan Howard1, David Franklin1, James Ingram1, Daniel Wolpert1
1University of Cambridge
2-F-6 Cerebellar modulation by TDCS during amotor taskMaria Dagioglou1, Chris Miall1
1University of Birmingham
2-F-7 A functional imaging study of the adaptationof saccadic eye movementsDenis Pelisson1, Pierre Fonlupt2, Aline Miquée2, Christian Urquizar2,Peggy Gérardin2
1INSERM, CNRS and Univ. of Lyon, 2Lyon Neuroscience ResearchCenter, INSERM U1028, CNRS UMR5292, Univ. Lyon1
2-F-8 Cerebellar mechanisms underlying differentforms of visuomotor adaptationJohn Schlerf1, Pablo Celnik2, Joseph Galea3
1Johns Hopkins University, 2Johns Hopkins School of Medicine,3University College London
2-F-9 The role of modularity in learning novelkinematic internal modelsLaura Patanè1, Giulio Sandini1, Alessandra Sciutti1, Bastien Berret1,Francesco Nori1
1Fondazione Istituto Italiano di Tecnologia
2-F-10 Pedaling-related brain activity in chronicstroke survivors: An fMRI studyNuttaon Promjunyakul1
1Marquette University
2-F-11 Structural learning induced by both randomand gradual motor task variationEdward Turnham1, Daniel Braun1, Daniel Wolpert1
1University of Cambridge
2-F-12 Rightward visual distortion from prismsproduces adaptation and unilateral, symmetricgeneralization during precision stepping and obstacleavoidance walking tasksDaniel Marigold1, Scott Alexander1, Brent Flodin1
1Simon Fraser University
2-F-13 Whisker stimulation at 8Hz induces LTD incerebellar Purkinje cell layer in alert miceGuy Cheron1, Javier Marquez-Ruiz2
1Univeristé Libre de Bruxelles, 2Universidad Pablo de Olavide
2-F-14 Automatic processing of multiple errorinformation in visuomotor learningShoko Kasuga1, Daichi Nozaki1, Masaya Hirashima1
1The University of Tokyo
2-F-15 Retention of motor memory during wholebody accelerationLuc Selen1, Adjmal Sarwary1, Pieter Medendorp1
1Donders Institute for Brain, Cognition and Behaviour
2-F-16 Endpoint error affects trajectory adaptationduring force field learningTsuyoshi Ikegami1, Ganesh Gowrishankar1, Tricia Gibo2, MitsuoKawato3, Rieko Osu1
1National Institute of Information and Communications Technology(NICT) / ATR, 2Johns Hopkins University, 3ATR
2-F-17 Modifications in free trajectory afteradaptation to a force field inducing target-errorGanesh Gowrishankar1, Rieko Osu2, Etienne Burdet3, TsuyoshiIkegami1
1National Institute of Information and Communications Technology(NICT), 2CNS-ATR, National Institute of Information andCommunications Technology (NICT), 3Imperial College London
2-F-18 Visuomotor adaptation to prism-alteredapparent distanceClaude Prablanc1, Anne-Emmanuelle Priot2, Corinne Roumes2, JustinPlantier2, Olivier Sillan3
1Inserm-Lyon Neuroscience Research Center, 2Army, 3Inserm-LyonNeuroscience Research Center
2-F-19 Visuo-motor reflex sensitivity is transientlyincreased during dynamic learningSae Franklin1, David Franklin1, Daniel Wolpert1
1University of Cambridge
Poster Sessions Authors, Titles and Affiliations

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 39
2-F-20 Expression of double cortin in unipolar brushcells of the ventral paraflocculus, flocculus and dorsalcochlear nucleus of the adult rat: Evidence for adultneurogenesis?Joan Baizer1, Senthilvelan Manohar 1, Nicholas Paolone1, SarahHayes1, Richard Salvi1
1University at Buffalo
2-F-21 Walking adaptation--When you get older,forget itLaura Malone1, Amy Bastian1
1Johns Hopkins University
2-F-22 Role of the posterior intra-parietal sulcus inmaintaining eye movement accuracy: An fMRI-guidedTMS studyMuriel Panouilleres1, Peggy Gerardin2, Laure Hudry2, RomeoSalemme2, Christian Urquizar2, Alessandro Farne2, Denis Pelisson2
1INSERM U1028, 2INSERM U1028, CNRS UMR5292,University Lyon 1
2-F-23 What is the impact of cerebellar stroke onresting-state functional connectivity?Paul Pope1, Dagmar Timmann2, Roxana Burciu2, Nina Theysohn2,Chris Miall1
1University of Birmingham, 2University of Duisburg-Essen
2-F-25 Adaptation of sound localization by eyeposition is linearly related to ocular eccentricityGary Paige1, Kathryn Cooper2, William O'Neill1
1University of Rochester, 2University of Rochester, College of Arts,Science & Engineering
2-F-26 Inter-hemispheric inhibition during mentalactions: An ipsilateral silent period studyCharalambos Papaxanthis1, Nicolas Gueugneau1, Marco Bove2,Thierry Pozzo1
1INSERM U887: Motricté et Plasticité, 2Departement of experimentalMedecine
2-F-27 Motor learning of space and time to controlsymmetry in split-belt walkingGelsy Torres-Oviedo1, Amy Bastian1, James Finley1
1Johns Hopkins University / Kennedy Krieger Institute
2-F-28 Task requirements predict transition tospecificity in visuomotor learningPaul Schrater1, Jacqueline Fulvio1
1University of Minnesota
2-F-29 Short term hand non-use: Behavioural andcortical effectsThierry Pozzo1, Luciano Fadiga2, Marco Bove, Michela Bassolino2
1Université de Bourgogne, 2Istituto Italiano di Tecnologia
2-F-30 Altering head plant dynamics induces rapidlearning in the neural circuit that underlies thecancellation of vestibular reafferenceJessica Brooks1, Kathleen Cullen1
1McGill University
2-F-31 Neurophysiology of interference in skilllearningGabriela Cantarero1, Pablo Celnik1, Rachel Salas1, Rebecca O'Malley1,Byron Tang1
1Johns Hopkins Medical Institution
2-F-32 Sequence learning modulates the restingbrainSaber Sami1, Ali Chowdhury1, Chris Miall1
1Birmingham University
2-F-33 Structure learning in a locomotor adaptationtaskErin Vasudevan1, Ting Feng2, Amy Bastian3
1Moss Rehabilitation Research Institute, 2Johns Hopkins School ofMedicine, 3Johns Hopkins School of Medicine and Kennedy KriegerInstitute
2-F-34 Somatosensory-auditory training changesspeech articulatory motionTakayuki Ito1
1Haskins Laboratories
2-F-35 Training characteristics influence themagnitude of motor learning during post-stroke gaitrehabilitationTrisha Kesar1, Darcy Reisman1, Stuart Binder-Macleod1
1University of Delaware
2-F-36 Neural network reorganization aftermicrostimulation in VPL and somatosensory cortexJoseph Francis1, Weiguo Song1
1SUNY Downstate Medical School
G - Control of Eye & Head Movement
2-G-51 Head-unrestrained tracking of targets in 2-Dreveals head tracking strategies and head-unrestrained catch-up saccades triggeringmechanismsPierre Daye1, Gunnar Blohm2, Philippe Lefèvre3
1National Eye Institute, NIH, 2Queen's University, 3Universitécatholique de Louvain
2-G-52 Do visual-vestibular interactions affectoculomotor system's tactics?Atiyeh Ghoreyshi1, Henrietta Galiana1
1McGill University
2-G-53 Vergence and version interactions duringspatial memory (updating)Joyce Dits1, Johan Pel1, Hans van der Steen1
1Erasmus Medical Center

40 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM
2-G-54 Eye position affects perception of wholebody direction in linear motionJianguang Ni1, Dominik Straumann1, Itsaso Olasagasti1
1University Hospital Zurich
2-G-55 Spatial summation properties of humanocular following responseChristian Quaia1, Boris Sheliga1, Lance Optican1
1National Eye Institute
2-G-56 Simultaneous recording from Purkinje andbasket / stellate cells in the rabbit flocculusJohn Simpson1, Tom Ruigrok2, Robert Hensbroek1
1NYU Medical School, 2Erasmus Medical Center
2-G-57 Robustness in the spatial localization of amoving targetLaurent Goffart1, Jérome Fleuriet1
1CNRS
2-G-58 Effects of a disturbed visual percept onvisuospatial memoryAngelique Remmers1, Joyce Dits1, Johan Pel1, Hans Van der Steen1
1Erasmus Medical Center
2-G-59 Effects of the NMDA antagonist ketamine onrule memory and response suppression in the anti-saccade taskKevin Skoblenick1, Brian Soper1, Stefan Everling1
1University of Western Ontario
2-G-60 Toward the development of a blink prosthesisfor facial paralysis PatientsDaniel McDonnall1, Christopher Smith1, Robert Askin1, M. DouglasGossman2, Shane Guillory1
1Ripple, 2University of Louisville
2-G-61 Local GABA-ergic networks suppress Purkinjecell firing in the oculomotor vermisAdrienne Mueller1, Farrel Ric Robinson1
1University of Washington
2-G-62 Galvanic vestibular stimulation suppressesvestibulo-ocular (VOR) and vestibulo-colic (VCR)reflexes in head-unrestrained guinea pigNatela Shanidze1, W. Michael King1
1University of Michigan
2-G-63 Methylphenidate affects oculomotor taskswitching and prefrontal cortex activityAbigail Rajala1, Luis Populin1
1University of Wisconsin - Madison
2-G-64 Neuromuscular evidence for a subcorticalvisuo-motor reflex in a hemidecorticate patientSamanthi Goonetilleke1, Brian Corneil1, Kate Rath-Wilson2, TimothyDoherty1, Daniel Guitton2
1University of Western Ontario, 2McGill University
2-G-65 Deficits in saccade production in parietaldamaged patientsAarlenne Khan1, Annabelle Blangero2, Douglas Munoz1, Laure Pisella3
1Queen's University, 2City College of New York, 3Inserm 864
2-G-66 Smooth pursuit and vestibular ocularbehavior in strabismic monkeysMichael Mustari1, Seiji Ono1
1University of Washington
H - Fundamentals of Motor Control
2-H-38 Continuous reflexive guidance of armmovement in space by the visual signal from thetarget. New hypothesis.Perfiliev Sergei1, Johan Wessberg1
1Institute of Neuroscience and Physiology, University of Gothenburg
2-H-39 Reduction of metabolic cost when learningnovel arm reaching dynamicsHelen Huang1, Bianca Bzdel1, Rodger Kram1, Alaa Ahmed1
1University of Colorado, Boulder
2-H-40 A scalable single-model based spinal-like controller for muscular manipulation of atwo-joint armFilip Stefanovic1, Henrietta Galiana1
1McGill University
2-H-41 Superposition between voluntary andautomatic grip force control during objectmanipulationFrederic Danion1
1Universite de la Mediterranee
2-H-42 Connections between PMd and superioprcolliculusKlaus-Peter Hoffmann1, Claudia Distler1, Vicente Reyes-Puerta1
1Ruhr-Universität Bochum
2-H-44 Cortical preparatory activity avoids causingmovement by remaining in a muscle-neutral spaceMatthew Kaufman1, Mark Churchland1, Stephen Ryu1, KrishnaShenoy1
1Stanford University
2-H-45 Effector selective recruitment of humandorso-medial PPC during reachingPierre-Michel Bernier1, Scott Grafton1
1University of California Santa Barbara
2-H-46 Adaptation to novel visuomotortransformation depends on compatibility with musclesynergiesDenise Berger1, Andrea d'Avella2, Dinesh Pai3, Timothy Edmunds3,Reinhard Gentner2
1IRCCS Fondazione Santa Lucia, 2Santa Lucia Foundation, 3Universityof British Columbia
Poster Sessions Authors, Titles and Affiliations

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 41
2-H-47 Robustness of muscle synergies duringvisuomotor adaptationAndrea D'Avella1, Timothy Edmunds2, Dinesh Pai2, Reinhard Gentner1
1Santa Lucia Foundation, 2University of British Columbia
2-H-48 Lateralized brain circuits mediate static anddynamic precision grip force productionKristina Neely1, Stephen Coombes1, Peggy Planetta1, DavidVaillancourt1
1University of Illinois at Chicago
2-H-49 Kinematic and electromyographic patterns ofvertical pointing movements performed with theshoulder, the elbow, and the wrist jointJeremie Gaveau1, Laurent Demougeot1, Bastien Berret2, LucianoFadiga3, Thierry Pozzo4, Charalambos Papaxanthis1
1INSERM U887 Motricite-Plasticite, University of Burgundy, 2ItalianInstitute of Technology, 3Dipartimento di Scienze Biomediche eTerapie Avanzate-Sezione di Fisiologia Umana, Università degli,4INSERM U887 Motricite-Plasticite, University of Burgundy, InstitutUniversitaire de France
2-H-50 A virtual environment for investigating thecontrol architecture in tool manipulationTimothy Edmunds1, Reinhard Gentner2, Dinesh Pai1, Andrea d'Avella2
1University of British Columbia, 2Santa Lucia Foundation
2-H-67 Set-size effects demonstrate a role of parietaland frontal theta oscillations in movement targetselectionChristian Rawle1, R. Chris Miall1, Peter Praamstra2
1University of Birmingham, 2Radboud University Nijmegen
2-H-68 Calibrating grasp stiffness during a gradedforce taskPatrick van der Smagt1, Hannes Hoeppner1, Claudio Castellini1
1DLR / RM
2-H-69 Motor preparation of spatially and temporallydefined movements: Evidence from startleDana Maslovat1, Nicola Hodges1, Romeo Chua1, Ian Franks1
1University of British Columbia
2-H-70 Transcranial magnetic stimulation to theprimary motor cortex can influence the early releaseof a preplanned movement by a startling acousticstimulusAndrew Stevenson1, Dana Maslovat1, Romeo Chua1, Ian Franks1
1University of British Columbia
2-H-71 Characteristics of a three-fingered graspduring a pouring task requiring dynamic stabilityRyan Manis1, Veronica Santos1
1Arizona State University
2-H-72 Preparatory mode shifts in response to dwelltime requirements in a reciprocal aiming taskMichael Bajema1, Anthony Carlsen2, Sam Perlmutter3, ColumMacKinnon3
1Kettering University, Northwestern University, 2University of Ottawa,3Northwestern University
2-H-73 Inference of control scheme in rhythmicmanual controlFelix Huang1, James Patton2, Ferdinando Mussa-Ivaldi1
1Northwestern University, 2University of Illinois Chicago
2-H-74 Optimizing redundant wrist and forearmrotationsJenny Pate1, Steven Charles1
1Brigham Young University
2-H-75 Directional preferences during dominant andnondominant arm movementsWanyue Wang1, Natalia Dounskaia1
1Arizona State University
2-H-76 Modular spinal primitives and their pulseadjustments in frog prey strike behaviorTanuj Gulati1, Corey Hart1, Simon Giszter1
1Drexel University College of Medicine
2-H-77 Relation between somatosensory detectionaccuracy and efferent control in the human jawLudo Max1, Ayoub Daliri1
1University of Washington
2-H-78 Muscle-like neurons in primary motor cortexEmily Oby1, Lee Miller1, Rebecca Friesen1
1Northwestern University

42 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM
NCM Sponsors and Exhibitors
NCM receives support from a number ofcompanies providing services to ourcommunity. This sponsor support fundsour Scholarship Program and allows usto support the attendance at the meetingfor many students.
The 2011 Annual Meeting is beingsupported by the following sponsoringcompanies, some of whom haveexhibits at the meeting. Please showyour appreciation for their support bylearning about their products andservices, and for those with exhibits,make time to visit with them while youare at the meeting.
Arrington Research, Inc.www.ArringtonResearch.com
220 Hz ViewPoint EyeTracker® systemsfrom Arrington Research are nowshipping. Systems include a SoftwareDevelopers Kit (SDK), real-time Ethernetcommunication, built-in stimuluspresentation, post-hoc data analysistools, a MATLAB toolbox, many other3rd Party product interfaces andexamples. Great for both human andnon-human primates and can beprovided with Analog and TTLcommunication to ensure seamlesscommunication with your existingequipment. ViewPoint EyeTracker®systems are the easiest and best valueavailable and include light-weight headmounted, HMD and head fixed systems.Arrington Research has been providingreliable affordable eye trackers for theresearch market worldwide for almost15 years.
ATI Industrial Automationwww.ati-ia.com
ATI Industrial Automation is a world-leading developer of Multi-AxisForce/Torque Sensing Systems,Automatic Tool Changers, RoboticCollision Sensors and Robotic DeburringTools. Our engineering-centric stafffocuses on providing customer solutionsto robotic, automation, and sensingapplications. Our Multi-AxisForce/Torque Sensors measure allcomponents of force and torque (Fx, Fy,Fz, Tx, Ty, and Tz) and are used in awide variety of applications including:robotic surgery, haptics, rehabilitation,and neurology. Key features include:High overload protection, high-speed
output, span temperature compensation,and high signal-to-noise ratio. OurSensors are available in a wide range ofsizes from 17 mm to 330 mm andinclude environmental protection (IP) onmost models. Visit our website atwww.ati-ia.com for more information.
Visit our exhibit at booth F.
BKIN Technologieswww.bkintechnologies.com
BKIN Technologies is a leadingdeveloper of robotic technologies forprobing brain function and dysfunction.Our patented KINARM™ product lineenables basic and clinical researchers toexplore sensory, motor and cognitiveperformance in both humans and NHPs.Our robotic and software tools createcomplex mechanical and visual worlds.BKIN’s products are paradigm shiftingand hold the promise of being the firstquantitative and objective system toassess sensory, motor and cognitivefunction - critical instrumentation for theeffective management of brain diseaseand injury.
Blackrock Microsystemswww.blackrockmicro.com
Founded in May 2008 near the BlackrockDesert in Utah, Blackrock Microsystemsdraws on a legacy of high techinnovation that began with BionicTechnologies, a spin-off from theUniversity of Utah in 1997. The companyprovides enabling tools for theneuroscience, neural engineering andneuroprosthetics research and clinicalcommunity worldwide. Blackrock hascreated a commercial vision to lead themarket in implantable microsystems.Our team caters to the immediate andfuture needs of the research markets.From miniature electrodes andintelligent microsystems, to complexdata acquisition systems and ground-breaking algorithms, we drive successfulcommercialization of cutting edgetechnology.
Visit our exhibit at booth A.
Cortech Solutions, Incwww.cortechsolutions.com
See us in the exhibition hall to learnabout Mind-FiTM, our wireless 63/126channel EEG system as well as the
Biosemi ActiveTwo and g.tec BCIsystems. We specialize in systems forEEG, sEMG and fNIRS in advancedneuroscience applications, and ERP,ERD, BCI and new combinations of thesetechniques with motion capture andother technologies are among ourcapabilities. Our clients include theworld’s leading motor control andrehabilitation research centers. Jointhem and let us provide solutions toyour research needs!
Visit our exhibit at booth B.
Motion Lab Sytems, Incwww.motion-labs.com
With a range of five MA300 EMGSystem packages, Motion Lab Systemsoffers the best value for a completeEMG System at an affordable price. Ourrange of EMG Systems and accessoriescontinue to feature the unrivaled dataintegrity and high quality performanceour customers have come to expect andour competitive prices make excellenceaffordable to everyone. We offersystems from the introductory MA300-XII system though to our 16 channel,high performance MA300-28. Allsystems are available as high reliabilitycabled systems or upgraded to highperformance Radio Telemetry systems.
Visit our exhibit at booth D.
Northern Digital Inc.www.ndigital.com
For 30 years, Northern Digital Inc.motion capture systems have earned thereputation as the gold standard formotion capture among researchscientists. With unparalleled accuracy,speed and resolution, the OptotrakCertus and 3D Investigator Systems willhelp you achieve your research goals.Both systems easily integrate with otherresearch devices, ensuring true-synchronous collection of data thatmeets all your needs. The OptotrakCertus and 3D Investigator’s “active”technology means you will never wastetime sorting data points. Furthermore,time consuming calibrations are notrequired. With a wide array of options,Northern Digital Inc. can meet therequirements of your most challengingresearch projects.
Visit our exhibit at booth E.

SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM | 43
Psychology Press Journals /Routledge Journalsc/o Taylor and Francis Group
www.taylorandfrancis.com
For two centuries, Taylor & Francis hasbeen fully committed to the publicationof scholarly information. Under ourPsychology Press and Routledgeimprints, we publish a wide variety ofjournals in the psychology field. Visit usto view our titles and to pick up FREEsample copies of our journals!
Visit our exhibit at booth H.
Ripplewww.rppl.com
Ripple provides neurophysiology dataacquisition systems for research. Oursystems are compact, portable, andheavily optimized for real-time, closed-loop control applications with hundreds
of channels of EMG, EEG andmicroelectrode data. Many of oursoftware components are available asOpen Source with support for Windows,Mac OS X, and Linux.
Visit our exhibit at booth C.
Sutter Instrumentswww.sutter.com
For over 35 years, Sutter Instrument hasbeen an international leader in themanufacture of precisioninstrumentation for the neurologicalsciences. With a proven dedication tocustomer satisfaction and wide range oftechnical expertise, we are able to offersuperior products that allow researchersto push the boundaries of science. Ourfocus is micropipette fabrication,micromanipulation, microinjection andoptical imaging. New products includethe touch screen P-1000 micropipette
puller, DG-4 PLUS fast wavelengthswitcher with 30% more output and0.5msec switching time, MOM MoveableObjective microscope, and MP-78/MPC-78 large moving platform stage for patchslice or in vivo experiments.
Tucker-Davis Technologieswww.tdt.com
Tucker-Davis Technologies is a leadingmanufacturer of modular DSP-baseddata acquisition and stimulus generationsystems, offering products ranging fromelectrodes to complete workstations forneurophysiology and evoked potentials.Stop by our exhibit for information onthe latest additions to the System 3platform, including our next generationmulti I/O processors, real-time datastreamer, and video tracker.
Visit our exhibit at booth G.
CONFERENCE, EVENT &ASSOCIATION SPECIALISTS
[email protected] • www.dearmondmanagement.com
ConferenceManagement
Conference planningand consulting servicesto support your eventfrom conception todelivery
ConferenceRegistration
Provide your attendeeswith a simple,convenient, online wayto register and pay foryour event
Conference WebDevelopment
Support yourconference with awebsite that speaks toyour audience anddrives participation andregistration
AbstractManagement
Collect, review andadminister yourAbstracts online withour integrated AbstractManagement system
AssociationManagement
Freedom to focus onyour long-termstrategies, while wemanage your dailyoperations and memberservices
E-Communications
Connect with yourcommunity, delegatesand members witheffective messagingusing our E-Communicationssystem
De Armond Management Ltd is a full service Conference and Association Managementcompany, specializing in supporting Scientific, Academic and Research organizations and groups withtheir Conference, Congress, and Member Management requirements. Our services include:
To learn more about us or to secure ourservices for your conference or organization,please call 1 800 472-7644 or e-mail us.

44 | SOCIETY FOR THE NEURAL CONTROL OF MOVEMENT | 21ST ANNUAL MEETING PROGRAM

ToFo
yer
Exhibits
Exhibits
58
57
56
55
54
53
52
51
50
49
48
47
46
4544 43 42 41
59
60
61
62
63
64
65
66
67
68
78
77
76
75
74
73
72
71
70
69
79 80 81 82
16 15 14 13
27
28
29
30
31
32
33
34
35
36
26
25
24
23
22
21
20
19
18
17
1
2
3
4
5
6
7
8
9
10
11
12
H A
BG
F C
DE
40 39 38 37
Food
Food
Food
Food
Wed, April 27 & Thurs April 28Ballroom B & C
A Workshop (Thoroughman)
B Theoretical & Computational Motor Control
C Disorders of Motor Control
D Integrative Control of Movement
E Posture & Gait
Blank Poster Spots
ToFo
yer
Exhibits
Exhibits
Food
Food
Food
Food
58
57
56
55
54
53
52
51
50
49
48
47
46
4544 43 4142 40 39 38 37
59
60
61
62
63
64
65
66
67
68
78
77
76
75
74
73
72
71
70
69
79 80 81 82
27
28
29
30
31
32
33
34
35
36
26
25
24
23
22
21
20
19
18
17
1
2
3
4
5
6
7
8
9
10
11
12
16 15 14 13
H A
BG
F C
DE
F Adaptation & Plasticity in Motor Control
G Control of Eye & Head
H Fundamentals of Motor Control
Blank Poster Spots
Fri, April 29 & Sat, April 30Ballroom B & C
Poster Layout
Session 1
Poster Layout
Session 2
Poster Session Floor Plans Ballroom B & C
ExhibitorsA Blackrock Microsystems
B Cortech Solutions, Inc
C Ripple
D Motion Lab Sytems, Inc
E Northern Digital Inc
F ATI Industrial Automation
G Tucker-Davis Technologies
H Psychology Press Journals / RoutledgeJournals c/o Taylor and Francis Group
* For information on the exhibitors, pleasevisit the Sponsor/Exhibitor profilesbeginning on page 42.
ExhibitorsA Blackrock Microsystems
B Cortech Solutions, Inc
C Ripple
D Motion Lab Sytems, Inc
E Northern Digital Inc
F ATI Industrial Automation
G Tucker-Davis Technologies
H Psychology Press Journals / RoutledgeJournals c/o Taylor and Francis Group
* For information on the exhibitors, pleasevisit the Sponsor/Exhibitor profilesbeginning on page 42.

Society for the
Neural Control of Movement
The 21st Annual NCM Meetingthanks our sponsors, supporters and donors




















