
Nectar: an energy drink used by European songbirds during spring migration
11
1 23 Journal of Ornithology ISSN 0021-8375 Volume 152 Number 4 J Ornithol (2011) 152:923-931 DOI 10.1007/s10336-011-0675-4 Nectar: an energy drink used by European songbirds during spring migration Jacopo G. Cecere, Fernando Spina, Susanne Jenni-Eiermann & Luigi Boitani
-
Upload
independent -
Category
Documents
-
view
2 -
download
0
Transcript of Nectar: an energy drink used by European songbirds during spring migration

1 23
Journal of Ornithology ISSN 0021-8375Volume 152Number 4 J Ornithol (2011) 152:923-931DOI 10.1007/s10336-011-0675-4
Nectar: an energy drink used by Europeansongbirds during spring migration
Jacopo G. Cecere, Fernando Spina,Susanne Jenni-Eiermann & Luigi Boitani

1 23
Your article is protected by copyright
and all rights are held exclusively by Dt.
Ornithologen-Gesellschaft e.V.. This e-offprint
is for personal use only and shall not be self-
archived in electronic repositories. If you
wish to self-archive your work, please use the
accepted author’s version for posting to your
own website or your institution’s repository.
You may further deposit the accepted author’s
version on a funder’s repository at a funder’s
request, provided it is not made publicly
available until 12 months after publication.

ORIGINAL ARTICLE
Nectar: an energy drink used by European songbirdsduring spring migration
Jacopo G. Cecere • Fernando Spina •
Susanne Jenni-Eiermann • Luigi Boitani
Received: 27 September 2010 / Revised: 15 January 2011 / Accepted: 14 February 2011 / Published online: 4 March 2011
� Dt. Ornithologen-Gesellschaft e.V. 2011
Abstract Nectar exploitation by European birds mainly
refers to passerines feeding on exotic plants, although some
recent studies described nectar-feeding by trans-Saharan
passerines on local plants. We examined which birds and
plants are involved in nectar consumption and investigated
the consequences of nectar use on plasma blood glucose
concentrations during spring migration at a stopover site in
the central Mediterranean. We recorded 12 opportunistic
nectar-feeding species, with Sylvia warblers in particular
showing a marked nectarivory, and 6 plants used by birds,
no one of which shows typical ‘‘bird flowers’’ features.
Moreover, we demonstrated that nectar uptake led to an
increase in plasma glucose concentrations and that nectar
was drunk not only by birds in poor physical condition. The
nectar consumption is fully in accordance with time-min-
imising migration models, allowing birds to obtain water
and energy at stopover sites in a short time: nectar is easy
to find and quick to digest.
Keywords Migrant � Pollen transport � Plasma glucose
concentration � Sylvia � Stop-over site
Zusammenfassung Nektarkonsum durch europaische
Singvogel wurde bisher vornehmlich an exotischen Pflanzen
beobachtet. Jungere Studien beschreiben aber auch Nektar-
konsum durch trans-Sahar Zugvogel an einheimischen
Pflanzen. In dieser Studie untersuchten wir wahrend des
Fruhlingszuges an einem Rastplatz im zentralen Mittelmeer-
raum, welche Vogelarten welche Pflanzen fur den Nektar-
konsum nutzen, und welche Auswirkungen die
Nektaraufnahme auf die Plasmakonzentration der Glukose
hat. 12 Arten, insbesondere Grasmucken, tranken Nektar an
insgesamt sechs Pflanzenarten, von denen keine die typischen
Merkmale der Ornithophilie zeigte. Die Nektaraufnahme
fuhrte zu einem Anstieg der Plasmaglukosekonzentration und
wurde vor allem von Vogeln in schlechter korperlicher Ver-
fassung genutzt. Nektar ist eine Energiequelle mit vielen
Vorteilen: sie ist schnell zu finden, rasch aufzunehmen und
leicht zu verdauen. Dieser Vorteil, Wasser und Energie
in kurzer Zeit aufzunehmen, reduziert die Zeit, die Zugvogel
an einem Rastplatz verbringen mussen und unterstutzt das
Zeit-Minimierungsmodell.
Introduction
Plants pollinated by birds are generally typified by tubular,
red and scentless flowers with a particular nectar compo-
sition and quantity (Johnson and Steiner 2000; Nicolson
2002; Nicolson and Fleming 2003; Rodrıguez-Girones and
Communicated by C. G. Guglielmo.
J. G. Cecere (&) � L. Boitani
Department of Biology and Biotechnology Charles Darwin,
University of Rome ‘‘La Sapienza’’, Viale dell’Universita 32,
00185 Rome, Italy
e-mail: [email protected]
L. Boitani
e-mail: [email protected]
F. Spina
Istituto Superiore per la Protezione e la Ricerca Ambientale,
Via Ca Forcanetta 9, 40064 Ozzano Emilia, BO, Italy
e-mail: [email protected]
S. Jenni-Eiermann
Swiss Ornithological Institute, Seerose 1,
6204 Sempach, Switzerland
e-mail: [email protected]
123
J Ornithol (2011) 152:923–931
DOI 10.1007/s10336-011-0675-4
Author's personal copy

Santamarıa 2004), although not all bird-flowers fit this
description (Ollerton 1998). Specialised nectarivores birds,
which regularly visit and pollinate plants, belong to only
seven families (Trochilidae, Drepaninidae, Nectariniidae,
Promeropidae, Meliphagidae, Psittacidae and Fringilli-
dae). But many other species, belonging to several families
(see, for example, Gryj et al. 1990; Symes et al. 2008;
Brown et. 2010a), are occasional nectarivores (Johnson and
Nicolson 2008); this is true of Sylvia warblers in Europe
(Stiles 1981; Ford 1985; Laursen et al. 1997; Pellmyr 2002)
which have representatives that opportunistically visit
flowers and feed on nectar, but do not necessarily pollinate
them.
Nectar exploitation by European birds mainly refers to
passerines feeding on exotic, introduced plants whose
flowers often show features typically used to attract birds
as pollinators (Holm and Laursen 1982; Prinzinger 1988;
Proctor et al. 1996; Harrup 1998; Merino and Nogueras
2003), but it has also been recorded for European plants,
such as Rhamnus alaternus, Ferula communis, Acer, Cra-
taegus, Salix, Ribes (reviewed by Ford 1985; Kay 1985;
Calvario et al. 1989; Schwilch et al. 2001). In 1989, Bur-
quez described the effective pollination of an exotic plant,
Fritillaria imperialis, by Parus caeruleus in Great Britain
(Burquez 1989), and in 2005, there was the first confir-
mation of a native bird-pollinated plant, Anagyris foetida,
in Europe (Ortega-Olivencia et al. 2005). However, the
ecological interactions between flowering plants and
European birds are still poorly studied.
In Europe, nectar consumption has also been observed in
migratory birds landing at stopover sites on Mediterranean
islands (Schwilch et al. 2001; Cecere et al. 2010); nectar-
feeding by trans-Saharan passerines has also recently been
described at oases in the Sahara desert (Salewski et al.
2006). During spring migration, trans-Saharan passerines
must cross two large ecological barriers—the Sahara desert
and the Mediterranean Sea—in order to reach their
breeding areas. Hence, they have to fly for approximately
2,000 km over inhospitable land or water. Some of them,
however, may have the chance to land on suitable areas
such as Mediterranean islands, after non-stop flights of up
to 14–16 h across the sea (Pilastro et al. 1995).
During such endurance flights, migrants have to deal
with a particular physiological situation, since they have to
fast, even up to several days, despite very high energy-
demanding conditions (Jenni and Jenni-Eiermann 1998). It
has been shown that the energy used for flight is mainly
derived from fat stores and to a lesser extent from proteins
(Jenni and Jenni-Eiermann 1998). Proteins not having a
storage form, they are obtained from the breast muscles
and the digestive organs (Schwilch et al. 2002). In a pre-
vious study on Ventotene Island (Italy), it was suggested
that nectar is an ideal first food for birds with a reduced
digestive capacity, because (1) monosaccharides do not
have to be digested as they are absorbed directly, and (2)
protein catabolism is stopped (Schwilch et al. 2001). It has
not yet been established whether nectar consumption
increases the plasma glucose concentration and is therefore
useful for reversing the process of lipolysis and protein
breakdown, whether it would only be a short-term aid for
burst flights or could rather be useful to continue the
migration journey in the longer term.
The aims of this study were to: (1) examine which bird
and plant species are involved in nectar consumption on
Ventotene Island; and (2) test the hypothesis that songbirds
gain physiological benefit in term of plasma glucose con-
centration from nectar consumption. Nectar is normally
characterised by high glucose concentration, and we pre-
dict that its exploitation leads to an increase in bird’s
plasma blood glucose level and that this benefit might
persist after few hours from the last feeding, at the post-
absorptive state. Moreover, we expect that nectar is more
frequently used by Sylvia warblers in poor physical con-
dition after endurance flights, while birds in good condition
target on more usual and nourishing food.
Methods
The field work was carried out between 1 April and 30 May
2006 and 2007 at Ventotene Island, 50 km off the Tyrrhe-
nian coast of Italy. Ventotene is a small inhabited island
(approx. 1.3 km2) covered by several small fields which
were formerly all cultivated. Many fields are now aban-
doned and are characterised by pioneer vegetation. Natural
Mediterranean maquis is confined to cliffs and small sur-
rounding areas. Several species of spring migrants arrive at
Ventotene Island from North Africa, after crossing at least
500 km of open sea by non-stop flights (Pilastro et al.
1995). Since 1998, the Istituto Superiore per la Protezione e
la Ricerca Ambientale, the Italian Ringing Centre, has co-
ordinated the ‘‘Progetto Piccole Isole’’ (Small Islands Pro-
ject) in order to investigate strategies adopted by spring
migrants when crossing the central Mediterranean (Spina
et al. 1993). During 2006 and 2007, 282 m of mist-net were
checked every hour and every day between 1 h before
sunrise to 1 h after sunset, except in bad weather conditions
(strong wind or driving rain), between 1 March and 31 May.
Nectar-feeding birds and plants census
A 1-km transect across the mosaic of fields in Ventotene
was carried out four times per week between 0900 and
1200 hours in order to collect qualitative data on nectar-
feeding bird species and the plants used by birds. The
transect was walked 48 times during 2006 and 40 times
924 J Ornithol (2011) 152:923–931
123
Author's personal copy

during 2007 for 30 min on average (total approximately
44 h). When an unrecorded bird species was found feeding
on a flower or an already recorded bird species was found
taking nectar from a new plant species, we checked for the
presence of insects or other invertebrates as a potential food
resource on each flower of the individual plant visited. The
absence of invertebrates on the plant and a second visit of
the bird feeding on the checked flowers within 10 min were
regarded as nectar exploitation. We excluded the possibility
that pollen is the target of the birds’ visits to flowers, since
Schwilch et al. (2001) clearly showed by faecal analysis that
birds do not feed on pollen at this site. According to the
authors, in fact, the anthers of Brassica are too large and the
smaller Ferula anthers are ingested just accidentally.
A census of nectar-feeding birds was also carried out
during the ringing activity on Ventotene during 2006 and
2007. For each captured bird, the presence or absence of
pollen on the bill, forehead and breast feathers was asses-
sed and scored. The presence of pollen grains on the
plumage of European migrating songbirds is considered
proof of nectar consumption by different authors (Ash et al.
1959, 1961; Laursen et al. 1997; Schwilch et al. 2001;
Salewski et al. 2006). In particular, birds feeding on nectar
from Brassica sp. usually had yellow and granular pollen
around their bill and on their forehead (Fig. 1), while birds
feeding on nectar from Ferula sp. usually had yellow and
gluey pollen around their bill and on their breast feathers
(Schwilch et al. 2001). We calculated the percentage of
captured individuals carrying pollen per species between
mid-April and mid-May for both study years.
Physical condition
The fat and muscle condition of each bird was also scored
during ringing activity on Ventotene; according to Kaiser
(1993), fat scores were assigned by estimating the visible
amount of subcutaneous fat deposition inside the furcula
and on the abdomen (9 score levels). Muscle scores were
assigned by estimating the thickness of the breast muscles
(4 score levels) (Bairlein 1995 based on Gosler 1991) as an
estimate of breast muscle protein mass (Jenni et al. 2000).
Plasma blood glucose concentration
High glucose concentrations have been found in the nectar
of Brassica sp. and Ferula sp. (Schwilch et al. 2001). To
assess the possible effect of nectar consumption on the
plasma glucose level of the birds we measured plasma
blood glucose concentration in Garden Warblers (Sylvia
borin): (1) which had been feeding on nectar, hence
showing fresh pollen on bill and feathers (n = 55; from
now onwards: ‘‘GW.P.’’); and (2) which probably did not
feed on nectar, showing no signs of pollen (n = 53; from
now onwards ‘‘GW.NO-P.’’). For this part of the study, we
considered only birds with fresh pollen on plumage which
recently fed on nectar, contrary to birds with dried and
agglutinated pollen, which most likely fed on nectar at
stopover sites visited prior to reaching the trapping site
(Cecere et al. 2010). Plasma blood glucose concentration
was measured by a handheld plasma glucose monitor
(Bayer Glucometer Ascensia Elite�) normally used on
humans, but also animals. The reliability of this method to
accurately measure plasma glucose level in birds has been
demonstrated by Lieske et al. (2002), and the same glu-
cometer was also used in recent studies on diel variation in
plasma glucose concentration in birds (Downs et al. 2010;
Lobban et al. 2010). It works with disposable test strips and
requires just one drop of blood. Blood was obtained by
puncturing the brachial vein.
For this part of the study, a sample of 108 Garden
Warblers with muscle score 1 and fat score between 0 and
2 were tested on Ventotene Island during the spring of
2007. The mist-nets were observed continuously from a
hide, and when a bird flew into the net, the exact trapping
time was recorded. Glucose is assimilated very quickly;
therefore, to avoid the possible influence of the last food
intake on glucose concentration, we analysed the blood in
the post-absorptive state, exactly 90 min after capture,
when birds were considered to be in a post-absorptive,
fasting state (Jenni-Eiermann and Jenni 1996). All tests
were carried out between 0900 and 1800 hours.
Statistical analyses
After testing for normal distribution (Shapiro–Wilk test),
we used parametric tests. General Linear Model (GLM)
was used to assess the possible influence of nectar-feeding,
fat reserves, test hour and the possible interactions of theseFig. 1 Common Whitethroat (Sylvia communis) with fresh pollen of
Brassica sp. on beak and forehead. Photo: J.G. Cecere
J Ornithol (2011) 152:923–931 925
123
Author's personal copy

variables with plasma glucose concentration; all variables
were entered both forwards and backwards, and selected by
SBC (Schwarz Bayesian information criterion), in order to
choose the model that best balances goodness of fit against
model complexity. In order to test whether nectar is more
frequently used by birds in weaker physical condition we
applied Logistic Regression to analyse the possible influ-
ence of fat and muscle on presence/absence of pollen on
plumage, as an indication of nectar consumption. All
analyses were performed with the statistical program SAS
ver. 9.2 (SAS Institute, Cary, NC, USA).
Results
The birds and plants involved in nectar consumption
Birds
During ringing activity on Ventotene, 68 passerine species
(23,614 individuals) were trapped between 1 April and 30
May 2006, and 72 passerine species (26,864 individuals)
were trapped in the same period in 2007. Pollen was
present on the bill, forehead or breast of birds belonging to
12 species trapped during the 2 years (Table 1). Individu-
als belonging to the other 74 passerine specie were trapped
with no indication of nectar feeding, including Icterine
Warbler (Hippolais icterina) that was recorded with pollen
on plumage, with just one individual, in 1997 (Table 1).
Sylvia species had high frequencies of birds with pollen,
while Phylloscopus species, collected only during spring
2006, had lower percentages (Table 1). Among migrating
Sylvia warblers, the species with the highest degree of
nectarivory was the Blackcap (S. atricapilla), 64.4% of
birds with pollen on plumage (2006 and 2007 data analysed
together), followed by Subalpine Warbler (S. cantillans),
Garden Warbler and Common Whitethroat (S. communis),
with 54.8, 38.7, and 34.3%, respectively. Lesser White-
throat (S. curruca), Spectacled Warbler (S. conspicillata)
and Ruppell’s Warbler (S. rueppelli) were trapped in too
small numbers to calculate percentages, while 50% of the
trapped resident Sardinian Warblers (S. melanocephala)
showed indications of nectar consumption (2006 and 2007
data analysed together). Among Phylloscopus, the species
with the highest frequency of birds with pollen on plumage
was Chiffchaff (P. collybita) with 28.6%, followed by
Willow Warbler (P. trochilus), 6.7% and Wood Warbler
(P. sylatrix), 0.2% (2006 data). The Italian Sparrow (Pas-
ser italiae) was the only species not belonging to Sylvidae
trapped with pollen on plumage. The percentages of birds
with pollen on Ventotene, per species for each year of the
study, are reported in Table 1.
During the qualitative transect census, the same species
were observed feeding on nectar, except for Wood Warbler
and Ruppell’s Warbler (Table 2). In May 2007, some
flocks (5–20 individuals) of Italian Sparrows were
observed several times feeding on nectar from Ferula sp.
just in front of the ringing station and two individuals with
Table 1 Sample size of species recorded with pollen on foreheads, bill or breast during the 2 years of study (2006–2007) and in 1997 (Schwilch
et al. 2001)
Species 1997 (Schwilch et al. 2001) 2006 2007
With
pollen
Birds
checked
% With
pollen
Birds
checked
% With
pollen
Birds
checked
%
Subalpine Warbler (S. cantillans) 250 2,244 11.1 171 309 55.3 159 293 54.3
Garden Warbler (S. borin) 169 852 19.8 856 2,620 32.7 546 998 54.7
Common Whitethroat (S. communis) 70 794 8.8 283 855 33.1 176 483 36.4
Blackcap (S. atricapilla) 31 271 11.4 14 23 60.9 15 22 68.2
Sardinian Warblera (S. melanocephala) 1 74 1.4 1 6 16.7 4 4 100.0
Willow Warbler (P. trochilus) 3 1,485 0.2 12 179 6.7
Chiffchaff (P. collybita) 4 595 0.7 4 14 28.6
Wood Warbler (P. sibilatrix) 1 308 0.3 1 429 0.2
Icterine Warbler (H. icterina) 1 588 0.2 0 1,631 0.0 0 1,205 0.0
Italian Sparrowa (P. Italiae) 0 29 0.0 2 40 5.0
Lesser Whitethroat (S. curruca) 1 2
Spectacled Warblerb (S. conspicillata) 1 2
Ruppell’s Warbler (S. rueppelli) 1 2
Total 530 7,211 7.35 1,344 6,100 22.03 903 3,047 29.64
a Breeding species in Ventoteneb Occasionally breeding species in Ventotene
926 J Ornithol (2011) 152:923–931
123
Author's personal copy

pollen on the forehead were also mist-netted. Although this
species has been commonly breeding in several colonies on
the island during the 20 years of the ringing project, it had
never been observed feeding on nectar before.
Plants
Six plant species were used by birds in Ventotene
(Table 2), of which four were local (Brassica montana,
Ferula communis, Fig. 2; Lavatera arborea and probably
Malva sylvestris), and two were introduced species (Pru-
nus avium and Pittosphorum tobira). None of these plants
show any bird-flower characteristics. Their flowers differ
in size and structure: medium size for B. montana, L. arbo-
rea, M. sylvestris and P. avium, and small for P. tobira and
F. communis, the flowers of the last being organised in an
inflorescence, while those of the other plants are single.
F. communis was the plant visited by all 10 bird species
observed during trasects, followed by B. montana (7 spe-
cies; Table 2). Only the Subalpine Warbler, an early pas-
sage long-distance migrant, was observed feeding on the
nectar of Prunus avium that blooms between late March
Ta
ble
2P
lan
tsp
ecie
sv
isit
edb
yso
ng
bir
ds
feed
ing
on
nec
tar
on
Ven
tote
ne
Isla
nd
insp
rin
g
Pla
nt
Flo
wer
Syl
via
atr
ica
pil
laS
.ca
nti
lla
ns
S.
bo
rin
S.
com
mu
nis
S.
curr
uca
S.
con
spic
illa
taS
.m
ela
no
cep
ha
laP
hyl
losc
op
us
tro
chil
us
P.
coll
ybit
aP
ass
erit
ali
ae
Fam
ily
Sp
ecie
sS
tru
ctu
re,
size
Bra
ssic
ace
ae
Bra
ssic
ain
can
aS
ing
le,
med
ium
44
44
44
4
Ap
iace
ae
Fer
ula
com
mu
nis
Infl
ore
scen
ceo
f
smal
lfl
ore
ts
44
44
44
44
44
Ma
lva
cea
eL
ava
tera
arb
ore
aS
ing
le,
med
ium
/
larg
e
44
44
Ma
lva
sylv
estr
isS
ing
le,
med
ium
4
Ro
sace
ae
Pru
nu
sa
viu
mS
ing
le,
med
ium
4
Pit
tosp
ora
cea
eP
itto
sph
oru
mto
bir
aS
ing
le,
smal
l4
4
Fig. 2 Above Blackcap (Sylvia atricapilla) feeding nectar on
B. montana. Photo: courtesy Paolo Fumagalli. Below Subalpine
Warbler (S. cantillans) feeding nectar on Ferula communis. Photo:
courtesy Marzia Mirabile
J Ornithol (2011) 152:923–931 927
123
Author's personal copy
and early April, when most trans-Saharan migrants have
not yet reached the island. Only Garden Warblers and
Common Whitethroats were observed feeding on nectar of
Pittosphorum tobira which is a plant used for hedges in
several local gardens and blooms in May, when the two
warblers are the most common species on Ventotene. Just
one Garden Warbler was observed nectar feeding on Malva
sylvestris.
Effects of nectar consumption on plasma blood
glucose concentration
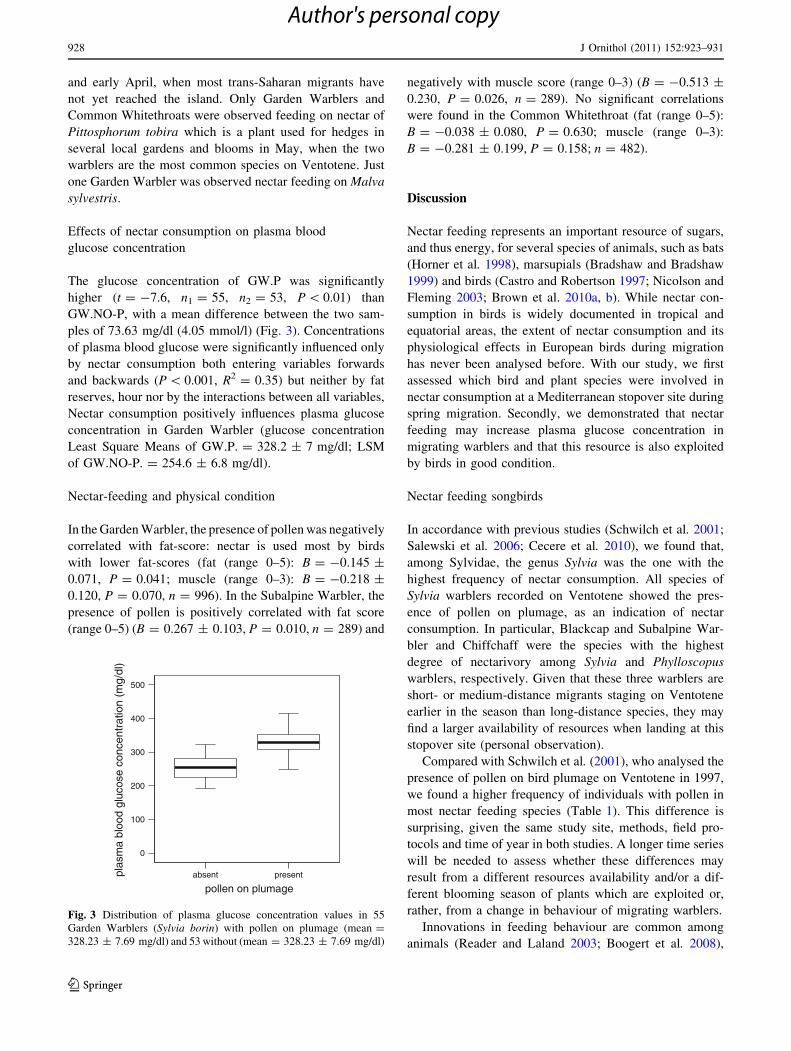
The glucose concentration of GW.P was significantly
higher (t = -7.6, n1 = 55, n2 = 53, P \ 0.01) than
GW.NO-P, with a mean difference between the two sam-
ples of 73.63 mg/dl (4.05 mmol/l) (Fig. 3). Concentrations
of plasma blood glucose were significantly influenced only
by nectar consumption both entering variables forwards
and backwards (P \ 0.001, R2 = 0.35) but neither by fat
reserves, hour nor by the interactions between all variables,
Nectar consumption positively influences plasma glucose
concentration in Garden Warbler (glucose concentration
Least Square Means of GW.P. = 328.2 ± 7 mg/dl; LSM
of GW.NO-P. = 254.6 ± 6.8 mg/dl).
Nectar-feeding and physical condition
In the Garden Warbler, the presence of pollen was negatively
correlated with fat-score: nectar is used most by birds
with lower fat-scores (fat (range 0–5): B = -0.145 ±
0.071, P = 0.041; muscle (range 0–3): B = -0.218 ±
0.120, P = 0.070, n = 996). In the Subalpine Warbler, the
presence of pollen is positively correlated with fat score
(range 0–5) (B = 0.267 ± 0.103, P = 0.010, n = 289) and
negatively with muscle score (range 0–3) (B = -0.513 ±
0.230, P = 0.026, n = 289). No significant correlations
were found in the Common Whitethroat (fat (range 0–5):
B = -0.038 ± 0.080, P = 0.630; muscle (range 0–3):
B = -0.281 ± 0.199, P = 0.158; n = 482).
Discussion
Nectar feeding represents an important resource of sugars,
and thus energy, for several species of animals, such as bats
(Horner et al. 1998), marsupials (Bradshaw and Bradshaw
1999) and birds (Castro and Robertson 1997; Nicolson and
Fleming 2003; Brown et al. 2010a, b). While nectar con-
sumption in birds is widely documented in tropical and
equatorial areas, the extent of nectar consumption and its
physiological effects in European birds during migration
has never been analysed before. With our study, we first
assessed which bird and plant species were involved in
nectar consumption at a Mediterranean stopover site during
spring migration. Secondly, we demonstrated that nectar
feeding may increase plasma glucose concentration in
migrating warblers and that this resource is also exploited
by birds in good condition.
Nectar feeding songbirds
In accordance with previous studies (Schwilch et al. 2001;
Salewski et al. 2006; Cecere et al. 2010), we found that,
among Sylvidae, the genus Sylvia was the one with the
highest frequency of nectar consumption. All species of
Sylvia warblers recorded on Ventotene showed the pres-
ence of pollen on plumage, as an indication of nectar
consumption. In particular, Blackcap and Subalpine War-
bler and Chiffchaff were the species with the highest
degree of nectarivory among Sylvia and Phylloscopus
warblers, respectively. Given that these three warblers are
short- or medium-distance migrants staging on Ventotene
earlier in the season than long-distance species, they may
find a larger availability of resources when landing at this
stopover site (personal observation).
Compared with Schwilch et al. (2001), who analysed the
presence of pollen on bird plumage on Ventotene in 1997,
we found a higher frequency of individuals with pollen in
most nectar feeding species (Table 1). This difference is
surprising, given the same study site, methods, field pro-
tocols and time of year in both studies. A longer time series
will be needed to assess whether these differences may
result from a different resources availability and/or a dif-
ferent blooming season of plants which are exploited or,
rather, from a change in behaviour of migrating warblers.
Innovations in feeding behaviour are common among
animals (Reader and Laland 2003; Boogert et al. 2008),
absent present
pollen on plumage
0
100
200
300
400
500
)ld/gm(
noitartnecnocesoculg
doolba
msalp
Fig. 3 Distribution of plasma glucose concentration values in 55
Garden Warblers (Sylvia borin) with pollen on plumage (mean =
328.23 ± 7.69 mg/dl) and 53 without (mean = 328.23 ± 7.69 mg/dl)
928 J Ornithol (2011) 152:923–931
123
Author's personal copy

where a new behaviour adopted by few individuals can
rapidly spread through social learning (Lefebvre and Pal-
ameta 1988; Reader 2004). This might be the case for the
Italian Sparrows that consumed nectar in our study, despite
it not having been documented previously on Ventotene.
However, nectar feeding is well described for the conge-
nerous House Sparrows (P. domesticus) (Leveau 2008).
Although social learning is easier to hypothesise for a
resident species such as the Italian Sparrow, which can
theoretically observe several individuals of migratory
songbirds feeding on nectar, social learning has also been
demonstrated for migrants at stopover sites during social
foraging activity (Nemeth and Moore 2007).
Plants involved in nectar exploitation
Six plant species, belonging to five different families, were
used by birds for nectar feeding on Ventotene island. None
of them have clear bird-flower features, such as tubular, red
and scentless flowers (Johnson and Steiner 2000; Rodrı-
guez-Girones and Santamarıa 2004). F. communis and
B. montana were the plants used by the highest number of
bird species (Table 2), which is consistent with the
importance of these plant species for nectar-feeding song-
birds on Ventotene, as shown by previously studies
(Schwilch et al. 2001; Cecere et al. 2010). The fact that six
plant species used by birds are characterised by different
floral structure and size suggests a strong opportunism by
warblers. The use of Ferula and Brassica was already
described in Schwilch et al. (2001), although B. montana,
with leaves lacking a petiole, was incorrectly identified as
B. fruticulosa, which has leaves with a petiole (Pignatti
1982). To our knowledge, the use of nectar of the other
four plants (Lavatera arborea, Prunus avium, Pittospho-
rum tobira and probably Malva sylvestris) by birds has
never been previously described in Europe. The plasticity
of some bird species to nectar feeding, also supported by
their opportunistic behaviour, can be an important selective
pressure driving plants to adapt towards the use of birds as
pollinators. Johnson and Nicolson (2008) showed that, both
in Africa and in America, plants pollinated by occasional
nectarivores birds are characterised by specific nectar
properties which differ from those of plants pollinated by
specialised nectarivores birds. However, this model still
has to be properly investigated in the Mediterranean region.
Consequences of nectar consumption
In our study, Garden Warblers with pollen on their plum-
age had higher levels of plasma glucose concentration
compared with birds with no pollen. All birds were tested
in a short-term fasting state, characterised by a significant
decrease of triglycerides and a significant increase of
ketone bodies in plasma, indicating fat catabolism (Jenni-
Eiermann and Jenni 1996). It is known that, during this
state, glucose is regulated within narrow limits in Garden
Warblers: no significant change in plasma glucose con-
centration was observed between the resorptive state, a
short-term fasting state (90–120 min without food) and a
long-term fasting state (10–12 h) (Jenni-Eiermann and
Jenni 1991). In our study, the difference in plasma glucose
levels between birds with and without pollen on plumage is
remarkable. Considering the normally narrow range of
variation (see above), this difference could be explained
only by the food ingested, hence by nectar feeding (Fig. 2).
Since only birds with fresh pollen on plumage were ana-
lysed, we can maintain that these birds fed on nectar in situ,
as suggested by Cecere et al. (2010). During spring
migration, birds with fresh pollen on plumage are present
on Ventotene Island, but they are virtually absent from
other two nearby islands, Ponza and Zannone, located in
the same archipelago and, therefore, within the same
migratory flyway (Cecere et al. 2010). This observation
indicates that fresh pollen came directly from Ventotene as
also confirmed by the time budget analysis reported in the
same study.
The increase in plasma glucose, as suggested by
Schwilch et al. (2001), may stop protein catabolism and
play a significant role in helping birds with a reduced
digestive capacity. It is possible that nectar, as a source of
energy, could be an important aid for migrating birds after
endurance flights across challenging ecological barriers.
Nectar feeding and physical condition
Nectar was fed by birds disposing of sufficient fat reserves
(Common Whitethroat, Subalpine Warbler) as well as by
birds disposing of small fat reserves (Garden Warbler).
Muscle score had a significant effect only in Subalpine
Warblers and not in the other species. These results suggest
that all birds, independent of their physical condition, used
nectar as a quickly absorbed energy drink. Moreover,
nectar may also be a valued water source (Symes et al.
2011) on dry Mediterranean islands. It is therefore rea-
sonable to think that its use could reduce the time spent by
migrants at the stopover site. Future studies should confirm
this hypothesis and clarify whether there is a real long
term-advantage, for nectar feeding birds, in the ensuing
completion of migration and the settlement at breeding
territories, verses migrants which only ate proteins en
route.
Acknowledgments We are extremely grateful to Andrea Ferri and
all other ringers who participated in the field activities at Ventotene
ringing station. We thank Carlo Catoni and Diego Rubolini for
important and useful suggestions and Simona Imperio for her
enthusiastic and constructive discussions during the preparation of the
J Ornithol (2011) 152:923–931 929
123
Author's personal copy

manuscript. We also thank Fabrizio Bartolucci and Laura Cornara for
the identification of Brassica montana and Anna McCann for the first
proofreading. Result from Progetto Piccole Isole (ISPRA) paper no. 45.
References
Ash JS (1959) Pollen contamination by birds. Br Birds 52:421–426
Ash JS, Jones PH, Melville R (1961) The contamination of birds with
pollen and other substances. Br Birds 54:93–100
Bairlein F (1995) European-African songbird migration network:
manual of field methods. Institut fur Vogelforschung ‘Vogelw-
arte Helgoland’, Wilhelmshaven
Boogert NJ, Reader SM, Hoppitt H, Laland KN (2008) The origin and
spread of innovations in starlings. Anim Behav 75:1509–1518
Bradshaw SD, Bradshaw FJ (1999) Field energetics and the
estimation of pollen and nectar intake in the marsupial honey
possum, Tarsipes rostratus, in heathland habitats of South-
Western Australia. J Comp Physiol B 169:569–580
Brown M, Downs CT, Johnson SD (2010a) Sugar preferences and
digestive efficiency in an opportunistic avian nectarivore,
the dark-capped bulbul Pycnonotus tricolour. J Ornithol
151:637–643
Brown M, Downs CT, Johnson SD (2010b) Sugar preferences of a
generalist nonpasserine flower visitor, the African speckled
mousebird (Colius striatus). Auk 127:781–786
Burquez A (1989) Blue tits, Parus caeruleus, as pollinators of the
crown imperial, Fritillaria imperialis, in Britain. Oikos
55:335–340
Calvario E, Fraticelli F, Gustin M, Sarrocco S, Sorace A (1989) The
blackcap Sylvia atricapilla and the garden warbler Sylvia borinas pollinator of Rhamnus alaternus (Rhamnaceae). Avocetta
13:53–55
Castro I, Robertson AW (1997) Honeyeaters and the New Zealand
forest flora: the utilisation and profitability of small flowers. NZ J
Ecol 21:169–179
Cecere JG, Matricardi C, Frank B, Imperio S, Spina F, Gargallo G,
Barboutis C, Boitani L (2010) Nectar exploitation by songbirds
at Mediterranean stopover sites. Ardeola 57:143–157
Downs CT, Wellmann AE, Brown M (2010) Diel variation in plasma
glucose concentrations of malachite sunbirds Nectarina famosa.
J Ornithol 151:235–239
Ford HA (1985) Nectarivory and pollination by birds in southern
Australia and Europe. Oikos 44:127–131
Gosler AG (1991) On the use of greater covert moult and pectoral
muscle as measures of condition in passerines with data for the
great tit Parus major. Bird Study 38:1–9
Gryj E, Martinez del Rio C, Baker I (1990) Avian pollination and
nectar use in Combretum fruticosum (Loefl.). Biotropica
22:266–271
Harrup B (1998) Wintering blackcaps taking nectar from, and
probably pollinating, Mahonia. Br Birds 91:201
Holm E, Laursen K (1982) Observation and experiments on the oral
apparatus and its function in some warblers species (Sylviidae).
Zool Anz Jena 209:224–246
Horner MA, Fleming TH, Sahley CT (1998) Foraging behaviour and
energetics of a nectar-feeding bat, Leptonycteris curasoae(Chiroptera: Phyllostomidae). J Zool Lond 244:575–586
Jenni L, Jenni-Eiermann S (1998) Fuel supply and metabolic
constraints in migrating birds. J Avian Biol 29:521–528
Jenni L, Jenni-Eiermann S, Spina F, Schwabl H (2000) Regulation of
protein breakdown and adrenocortical response to stress in birds
during migratory flight. Am J Physiol Regul Integr Comp
Physiol 278:1182–1189
Jenni-Eiermann S, Jenni L (1991) Metabolic responses to flight and
fasting in night-migrating passerines. J Comp Physiol B
161:465–474
Jenni-Eiermann S, Jenni L (1996) Metabolic differences between the
post-breeding, moulting and migratory periods in feeding and
fasting passerine birds. Funct Ecol 10:62–72
Johnson SD, Nicolson SW (2008) Evolutionary associations between
nectar properties and specificity in bird pollination system. Biol
Lett 4:49–52
Johnson SD, Steiner KE (2000) Generalization versus specialization
in plant pollination systems. Trends Ecol Evol 15:140–143
Kaiser A (1993) A new multi-category classification of subcutaneous
deposits of songbird. J Field Ornithol 64:246–255
Kay QON (1985) Nectar from willow catkins as a food source for
blue tits. Bird Study 32:40–44
Laursen K, Holm E, Sørensen I (1997) Pollen as a marker in
migratory warblers, Sylviidae. Ardea 85:223–231
Lefebvre L, Palameta B (1988) Mechanisms, ecology and population
diffusion of socially learned, food-finding behaviour in feral
pigeons. In: Zentall T, Galef BG Jr (eds) Social learning:
psychological and biological perspectives. Erlbaum, Hillsdale,
pp 141–160
Leveau LM (2008) Dynamics of nectarivory in the house sparrow in
an urban environment. Ornitol Neotrop 19:275–281
Lieske CL, Ziccardi MH, Mazet JAK, Newman SH, Gardner IA
(2002) Evaluation of 4 handheld blood glucose monitors for use
in seabird rehabilitation. J Avian Med Surg 16:277–285
Lobban K, Downs C, Brown M (2010) Diel variation in plasma
glucose concentration in some South African avian frugivore.
Emu 110:66–70
Merino S, Nogueras I (2003) Loquat Eriobotrya japonica as a winter
nectar source for birds in central Spain. Ardeola 50:265–267
Nemeth Z, Moore FR (2007) Unfamiliar stopover sites and the value
of social information during migration. J Ornithol 148:369–376
Nicolson SW (2002) Pollination by passerine birds: why are the
nectars so dilute? Comp Biochem Physiol B 131:645–652
Nicolson SW, Fleming PA (2003) Nectar as food for birds: the
physiological consequences of drinking dilute sugar solutions.
Plant Syst Evol 238:139–153
Ollerton J (1998) Sunbird surprise for syndromes. Nature
394:726–727
Ortega-Olivencia A, Rodrıguez-Riano T, Valtuena FJ, Lopez J,
Devesa JA (2005) First confirmation of a native bird-pollinated
plant in Europe. Oikos 110:578–590
Pellmyr O (2002) Pollination by animals. In: Herrera CM, Pellmyr O
(eds) Plant-animal intersection. An evolutionary approach.
Blackwell, Oxford, pp 157–184
Pignatti S (1982) La Flora d’Italia. Ed agricole, Bologna
Pilastro A, Baccetti N, Massi A, Montemaggiori A, Roselli A, Spina F
(1995) Direction of migration and fat consumption rate estimates
in spring migrating garden warblers (Sylvia borin). Ric Biol
Selvag 12:435–445
Prinzinger R (1988) Nektar als regelmaßige Zugnahrung bei
Grasmucken? J Ornithol 129:475–478
Proctor M, Yeo P, Lack A (1996) The natural history of pollination.
Timber Press, Portland, OR
Reader SM (2004) Distinguishing social and asocial learning using
diffusion dynamics. Learn Behav 32:90–104
Reader SM, Laland KN (2003) Animal innovation: an introduction.
In: Reader SM, Laland KN (eds) Animal innovation. Oxford
University Press, Oxford, pp 3–35
Rodrıguez-Girones MA, Santamarıa L (2004) Why are so many bird
flowers red? PLoS Biol 2:e350
Salewski V, Almasi B, Schlageter A (2006) Nectarivory of Palearctic
migrants at a stopover site in the Sahara. Br Bird 99:299–305
930 J Ornithol (2011) 152:923–931
123
Author's personal copy

Schwilch R, Mantovani R, Spina F, Jenni L (2001) Nectar consump-
tion of warblers after long-distance flights during spring
migration. Ibis 143:24–32
Schwilch R, Grattarola A, Spina F, Jenni L (2002) Protein loss during
long-distance migratory flight in passerine birds: adaptation and
constraint. J Exp Biol 205:687–695
Spina F, Massi A, Montemaggiori A, Baccetti N (1993) Spring
migration across central Mediterranean: general results from
‘‘Progetto Piccole Isole’’. Vogelwarte 37:1–94
Stiles FG (1981) Geographical aspects of bird-flower coevolution,
with particular reference to Central America. Ann Miss Bot Gard
68:323–351
Symes TC, Nicolson SW, McKechnie AE (2008) Response of avian
nectarivore to the flowering of Aloe marlothii: a nectar oasis
during dry South African winters. J Ornithol 149:12–22
Symes TC, McKechnie AE, Nicolson SW, Woodborne SM (2011)
The nutritional significance of a winter-flowering succulent for
opportunistic avian nectarivores. Ibis 153:110–121
J Ornithol (2011) 152:923–931 931
123
Author's personal copy




















