
Myofiber adaptational response to exercise in a mouse model of nemaline myopathy
11
ABSTRACT: In some muscle diseases, such as muscular dystrophy, ex- ercise can increase muscle damage and alter myofiber adaptation. We determined whether this is also true for the congenital muscle disease nemaline myopathy using our mouse model of this disease. Nemaline mice expressing a mutant -tropomyosin slow protein [-Tm slow (Met9Arg)] in skel- etal muscle underwent 4 weeks of treadmill exercise. Exercise increased slow/oxidative myofibers, but different fibers were involved in these trans- formations in nemaline mice. Despite similar expression of the mutant -Tm slow protein in muscles of the nemaline mouse, muscles responded in a unique manner that did not reflect fiber-type composition. For example, the particular fibers involved in fast-to-slow transformation were specific for each muscle examined. In contrast to the muscular dystrophies, exercise did not result in muscle damage nor did it cause an increase in rod-containing fibers; however, the fiber-type distribution of rod-containing fibers was altered in a muscle-specific fashion. That exercise did not exacerbate the pathology (i.e., nemaline rod formation) supports its use in nemaline myopathy pa- tients. This study shows that fibers of a similar type respond to increased activity differently in different muscles and suggests that fibers of similar type may be functionally distinct in different muscles. Muscle Nerve 30: 470 – 480, 2004 MYOFIBER ADAPTATIONAL RESPONSE TO EXERCISE IN A MOUSE MODEL OF NEMALINE MYOPATHY VISALINI NAIR-SHALLIKER, MSc, 1 ANTHONY J. KEE, PhD, 1 JOSEPHINE E. JOYA, DVM, 1 CHRISTINA A. LUCAS, PhD, 2 JOSEPH F. HOH, PhD, 2 and EDNA C. HARDEMAN, PhD 1 1 Muscle Development Unit, Children’s Medical Research Institute, Westmead, NSW 2145, Australia 2 Department of Physiology and Institute for Biomedical Research, Building F13, University of Sydney, Sydney, NSW 2006, Australia Accepted 18 June 2004 Nemaline myopathy is the most common congenital disease of skeletal muscle, presenting at birth or in early childhood with hypotonia and muscle weak- ness. A defining feature of this condition is the pres- ence of electron dense structures, called rods, in the sarcomeres. Nemaline rods are thought to be accu- mulations of Z-line proteins, principally -actinin, formed through an unchecked expansion of the Z line. 13 Mutations in five genes encoding proteins found in or associated with the thin filaments of the sarcomere have been held responsible for some forms of nemaline myopathy: -tropomyosin, -tro- pomyosin slow (-Tm slow ), nebulin, -skeletal actin, and troponin T slow. 29 The first nemaline-associated mutation to be identified was a single base change (Met to Arg) in the gene encoding -Tm slow (TPM3). 11 Patients with this mutation were diag- nosed with childhood-onset nemaline myopathy. We have generated a transgenic mouse model of nemaline myopathy by expressing this dominant negative -Tm slow (Met9Arg) mutant in skeletal mus- cle. 6 This model of nemaline myopathy has all of the features of the human disease including the pres- ence of nemaline rods in skeletal muscle and an increase in slow/oxidative fibers. As has been ob- served in muscles of a patient with nemaline myop- athy, 22 the number of rod-containing fibers within muscles varied significantly. In the mouse model, we were able to determine that this variation was not dependent on the amount of mutant protein Abbreviations: ANOVA, analysis of variance; CSA, cross-sectional area; Tm, tropomyosin; ECU, extensor carpi ulnaris; EDL, extensor digitorum lon- gus; MyHC, myosin heavy chain; ND, not detected; PBS, phosphate-buffered saline; SDS–PAGE, sodium dodecyl sulphate–polyacrylamide gel electro- phoresis; SOL, soleus; WT, wildtype Key words: congenital muscle diseases; endurance exercise; myofiber transformation; transgenic mouse; tropomyosin Correspondence to: E. Hardeman, e-mail: [email protected] © 2004 Wiley Periodicals, Inc. Published online 18 August 2004 in Wiley InterScience (www.interscience. wiley.com). DOI 10.1002/mus.20138 470 Exercise in Mice with Nemaline Myopathy MUSCLE & NERVE October 2004
-
Upload
independent -
Category
Documents
-
view
1 -
download
0
Transcript of Myofiber adaptational response to exercise in a mouse model of nemaline myopathy

ABSTRACT: In some muscle diseases, such as muscular dystrophy, ex-ercise can increase muscle damage and alter myofiber adaptation. Wedetermined whether this is also true for the congenital muscle diseasenemaline myopathy using our mouse model of this disease. Nemaline miceexpressing a mutant �-tropomyosinslow protein [�-Tmslow(Met9Arg)] in skel-etal muscle underwent 4 weeks of treadmill exercise. Exercise increasedslow/oxidative myofibers, but different fibers were involved in these trans-formations in nemaline mice. Despite similar expression of the mutant�-Tmslow protein in muscles of the nemaline mouse, muscles responded ina unique manner that did not reflect fiber-type composition. For example, theparticular fibers involved in fast-to-slow transformation were specific for eachmuscle examined. In contrast to the muscular dystrophies, exercise did notresult in muscle damage nor did it cause an increase in rod-containing fibers;however, the fiber-type distribution of rod-containing fibers was altered in amuscle-specific fashion. That exercise did not exacerbate the pathology(i.e., nemaline rod formation) supports its use in nemaline myopathy pa-tients. This study shows that fibers of a similar type respond to increasedactivity differently in different muscles and suggests that fibers of similar typemay be functionally distinct in different muscles.
Muscle Nerve 30: 470–480, 2004
MYOFIBER ADAPTATIONAL RESPONSETO EXERCISE IN A MOUSE MODEL OFNEMALINE MYOPATHY
VISALINI NAIR-SHALLIKER, MSc,1 ANTHONY J. KEE, PhD,1
JOSEPHINE E. JOYA, DVM,1 CHRISTINA A. LUCAS, PhD,2
JOSEPH F. HOH, PhD,2 and EDNA C. HARDEMAN, PhD1
1Muscle Development Unit, Children’s Medical Research Institute, Westmead,NSW 2145, Australia2Department of Physiology and Institute for Biomedical Research, Building F13,University of Sydney, Sydney, NSW 2006, Australia
Accepted 18 June 2004
Nemaline myopathy is the most common congenitaldisease of skeletal muscle, presenting at birth or inearly childhood with hypotonia and muscle weak-ness. A defining feature of this condition is the pres-ence of electron dense structures, called rods, in thesarcomeres. Nemaline rods are thought to be accu-mulations of Z-line proteins, principally �-actinin,formed through an unchecked expansion of the Zline.13 Mutations in five genes encoding proteinsfound in or associated with the thin filaments of the
sarcomere have been held responsible for someforms of nemaline myopathy: �-tropomyosin, �-tro-pomyosin slow (�-Tmslow), nebulin, �-skeletal actin,and troponin T slow.29 The first nemaline-associatedmutation to be identified was a single base change(Met to Arg) in the gene encoding �-Tmslow
(TPM3).11 Patients with this mutation were diag-nosed with childhood-onset nemaline myopathy.
We have generated a transgenic mouse model ofnemaline myopathy by expressing this dominantnegative �-Tmslow(Met9Arg) mutant in skeletal mus-cle.6 This model of nemaline myopathy has all of thefeatures of the human disease including the pres-ence of nemaline rods in skeletal muscle and anincrease in slow/oxidative fibers. As has been ob-served in muscles of a patient with nemaline myop-athy,22 the number of rod-containing fibers withinmuscles varied significantly. In the mouse model, wewere able to determine that this variation was notdependent on the amount of mutant protein
Abbreviations: ANOVA, analysis of variance; CSA, cross-sectional area;Tm, tropomyosin; ECU, extensor carpi ulnaris; EDL, extensor digitorum lon-gus; MyHC, myosin heavy chain; ND, not detected; PBS, phosphate-bufferedsaline; SDS–PAGE, sodium dodecyl sulphate–polyacrylamide gel electro-phoresis; SOL, soleus; WT, wildtypeKey words: congenital muscle diseases; endurance exercise; myofibertransformation; transgenic mouse; tropomyosinCorrespondence to: E. Hardeman, e-mail: [email protected]
© 2004 Wiley Periodicals, Inc.Published online 18 August 2004 in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/mus.20138
470 Exercise in Mice with Nemaline Myopathy MUSCLE & NERVE October 2004

present in the muscle because all muscles expressedsimilar levels of �-Tmslow(Met9Arg) protein. Thenumber of rod-containing fibers in individualmuscles was consistent within litters of�-Tmslow(Met9Arg) mice and through generations.The increase in number of slow/oxidative fibers waspresent at 1 month of age and was maintainedthrough adulthood, indicating disruption of theearly postnatal maturation of the different fibertypes.
Skeletal muscle is composed of a number of dif-ferent fibers that have been classified into four maintypes: a single slow and three fast (IIA, IIX or IID,and IIB) fiber-types. Fibers of a particular type ex-press a specific myosin heavy chain (MyHC) isoform:MyHC slow/I�, IIA, IIX, and IIB. Thus the expres-sion of MyHC isoforms is commonly used as a basisof fiber-type classification. Altered muscle activitypromotes conversion between the different fibertypes. With exercise training, there is an increase inoxidative enzyme capacity and a shift in fiber-typeand MyHC expression from slow to fast isoforms(reviewed elsewhere15). In rodent fast-twitch muscles[extensor digitorum longus (EDL), tibialis anterior,gastrocnemius] there is a decrease in fast MyHC IIBisoform expression and an increase in MyHC IIXand IIA content.2,3,8 In the slow-twitch muscles of therat (e.g., soleus), a shift occurs from MyHC IIX andIIA to the slow MyHC I(�) isoform.3,8 The slowercontractile phenotype of the muscle is thought to bedue largely to a shift from fast to slow MyHC isoformexpression.15 With exercise regimens of high inten-sity or increased duration, the shift to slower MyHCisoforms is often associated with hypertrophy of theslower fibers8,9 leading to increased muscle strengthand decreased fatigability.9
In the present study we have used our mousemodel of nemaline myopathy to examine whetherexercise exacerbates the muscle pathology andwhether disruption of the sarcomeric thin filamentby the mutant �-Tmslow protein alters myofiber ad-aptational response to exercise. Another importantissue was whether exercise training leads to an in-crease in the severity of sarcomere disruption asso-ciated with nemaline myopathy (i.e., nemalinerods). As pathology varies widely between muscles inhumans and the Tmslow(Met9Arg) mouse, we hy-pothesized that myofiber adaptation is muscle spe-cific.
MATERIALS AND METHODS
This study was approved by our local Animal Careand Ethics Committee.
Transgenic Mice. Generation and initial character-ization of the �Tmslow(Met9Arg) transgenic mice(FVB/NJ background strain) has been describedpreviously.6 The �Tmslow(Met9Arg) transgene(driven by the human skeletal actin promoter) ex-presses in all fibers at differing levels.6 A mouse line(Line 4) that has high expression of the mutant Tmtranscript and a significant amount of sarcomeredisruption in fast fibers (e.g., nemaline rods andcytoplasmic bodies)6 was used in these studies. Non-transgenic mice arising from the matings of hemizy-gous transgenic mice were used as control wildtype(WT) animals. Mice were genotyped by Southernhybridization as described by Corbett et al.6
Endurance Exercise. Seven male transgenic and 7male WT mice were fed food and water ad libitumand, 2 weeks prior to exercise, were housed in aroom with reversed light/dark cycle (light off: 7:00;light on: 19:00). Mice were exercised in a light-filledroom during the period of maximum activity (9:00to 14:00). At 11 weeks of age, the mice were primedto exercise on a motorized treadmill (ColumbusExer-4/8, Columbus, OH) for 1 week. The primingregimen increased gradually in intensity and in du-ration from 7–10 m.min�1 for 20 min on the first dayof priming to 15 m.min�1 for 70 min on the final dayof priming.
Endurance exercise training began at age 12weeks. Prior to the commencement of the exerciseregime, total body weight of each mouse was deter-mined. Mice were exercised for 2 h daily and theintensity of the endurance exercise was increasedprogressively. In week 1, the treadmill was set at anincline of 6% and speed of 15–17 m.min�1. In week2, the incline and the speed were increased to 8%and 20 m.min�1, respectively. In weeks 3 and 4, theincline was increased to 10% with a further increasein speed to 25 m.min�1.
At the end of the exercise regime (day 29), themice were sacrificed by cervical dislocation and mus-cles were collected that were predominately fast(EDL; extensor carpi ulnaris, ECU) or slow (soleus,SOL) and had a range of rod-containing fibers(ECU, 60%; EDL, 35%; and SOL, � 1%). Musclesfrom the right limbs were weighed and stored at�80°C for MyHC gel analysis. Muscles from the leftlimbs were coated in tissue freezing medium (Pro-SciTech, Brisbane, Australia), frozen in melting iso-pentane prechilled in liquid nitrogen, and stored inliquid nitrogen for immunohistochemical analysis.
Immunohistochemistry. Quantification of fiber typeand morphometry were performed as described by
Exercise in Mice with Nemaline Myopathy MUSCLE & NERVE October 2004 471

Corbett and colleagues,6 with minor modifications.Muscles were equilibrated to �24°C prior to section-ing. Transverse sections (8-�m thickness) were cutfrom the midsection of the muscles. Each section wasfixed in 2% paraformaldehyde in phosphate-buff-ered saline (PBS) for 2 min, followed by threewashes in PBS. After the final PBS wash, nonspecificsites were blocked in 10% normal goat serum for 1 hat room temperature. The goat serum was removedand primary monoclonal antibody was applied toeach section and left overnight at 4°C. Sections weretreated with monoclonal antibodies obtained fromsupernatants of hybridoma cultures to detect thefollowing MyHC isoforms: slow/type I� (undilutedBA-F8),4 type IIB (undiluted BF-F3),21 and type IIA(undiluted SC 71).21 All hybridomas were purchasedfrom the German Collection of Microorganisms andCell Cultures. The sections were washed and incu-bated in 0.3% H2O2 in PBS for 15 min at roomtemperature to quench endogenous peroxidase ac-tivity. The sections were washed thoroughly and in-cubated in secondary antibody (peroxidase-conju-gated goat/antimouse immunoglobulins; Dako,Glostrup, Denmark; 1:100 dilution) for 20 min atroom temperature. Detection was carried out usingthe DAB peroxidase substrate kit (Vector Laborato-ries, Burlingame, CA). All washes were carried out inPBS at room temperature.
To determine whether the number of rod-con-taining fibers was altered with exercise, muscle sec-tions were stained with antibodies to �-actinin, themajor constituent of nemaline rods. Sections weremade and prepared for immunohistochemistry asdescribed above. The primary antibody mixturecomprised the monoclonal myosin heavy chains ofinterest and the rabbit-polyclonal �-actinin-2 and -3(1:200; gift from Alan Beggs). A monoclonal anti-body directed against MyHCIIX was obtained fromsupernatants of hybridoma cultures as described pre-viously.12 The sections were washed thoroughly andincubated in (1:1000 dilution) secondary antibody(Alexa Fluor 488 goat anti-mouse immunoglobulinsand Alexa Fluor 594 goat anti-rabbit immunoglobu-lins; Molecular Probes OR, Sydney, Australia) for 1 hat room temperature. The number of rods in specificfiber types was determined on sections stained withthe �-actinin antibody and co-stained with a MyHC-specific antibody. The total number of rods in allfiber types was also determined to provide an esti-mate of the number of rod-containing fibers foreach fiber type as a proportion of all rod-containingfibers in a muscle. At least 300 fibers from eachmuscle were counted.
NADH-TR Assay. Oxidative fibers were detected bystaining for NADH-tetrazolium reductase (NADH-TR) as described by Allen and colleagues.2 Muscleswere cut, fixed, and washed as described above. Sec-tions were pre-incubated in 0.2 mol/L Tris-HCl (pH7.4) for 30 min at room temperature and then incu-bated for up to 15 min at 37°C in a reaction mixturecontaining 1.5 mM NADH and 1.5 mM nitrotetrozo-lium blue in 0.2 mol/L Tris (pH 7.4). Followingincubation, the sections were removed from the re-action mixture and washed thoroughly in distilledwater.
Fiber Morphometry. Images from the muscle sec-tions were captured using a digital camera. Fiberdiameters were obtained from the stained sectionsusing the Image-Pro software (Media Cybernetics,Silver Spring, MD). Fiber diameter was measured asthe shortest axis for each fiber within each section.This allows for fiber distortion due to imperfecttransverse sectioning and fiber compression.23 Cross-sectional area (CSA) was estimated (CSAest) for eachfiber in each muscle assuming each fiber is repre-sented as a cylinder. Accordingly, the CSAest of eachfiber was calculated as �r,2 where r is the radius ofthe fiber. The CSAest of each specific fiber type wasexpressed as a percentage of the CSAest of the entiresection to correct for the effect of angle of section.
Myosin Heavy Chain (MyHC) Isoform Composition.
Frozen tissues were crushed and suspended in 10volumes of extraction buffer containing 0.3 M NaCl,0.1 M NaH2PO4, 50 mM Na2HPO4, 10 mM sodiumpyrophosphate, 1 mM MgCl2, 10 mM EDTA, and 1.4mM mercaptoethanol.5 The suspension was allowedto sit on ice for 1 h, and then centrifuged at 23,000g for 5 min at 4°C. To the resulting supernatant wasadded an equal volume of 100% glycerol and theextract was stored at �20°C.
Myosin heavy chain isoforms were separated onsodium dodecyl–sulphate polyacrylamide electro-phoresis (SDS–PAGE) as described by others.1,26
Briefly, the MyHC extract was resuspended in sam-ple buffer containing Laemmli Sample Buffer (Bio-rad, Sydney, Australia) and 10% mercaptoethanol.Samples were loaded onto a 0.75 mm, 8% separatingand 3% stacking vertical acrylamide mini gel (Mini-protean Protean III, Biorad, Sydney, Australia). Thegels were electrophoresed at 70 V for 30 h at 4°C.The upper chamber buffer contained 0.1 M Tris, 150mM glycine, 0.1% SDS, and 0.07% mercaptoethanoland the lower chamber buffer contained 50 mMTris, 75 mM glycine, and 0.05% SDS.
472 Exercise in Mice with Nemaline Myopathy MUSCLE & NERVE October 2004
At the end of the run, the gels were stained usinga silver nitrate kit (Biorad, Sydney, Australia) andscanned using a Biorad GS800 densitometer. Therelative amount of protein in each band was quanti-fied using the Quantity One software (Biorad, Syd-ney, Australia).
Data and Statistical Analysis. Results are presentedas means and standard errors of the mean (SE). TheLevene’s test was performed to assure homogeneityof variance together with the Kolmogorov–Smirnov(Lilliefors) test to check for normality of the data.24
All data presented satisfied the above tests and soparametric analysis was employed.
Initially, global differences between groups weredetermined using two-way analysis of variance(ANOVA), with the factors being exercise and thepresence of the mutant transgene.24 If statisticallysignificant differences were detected by ANOVA, in-dividual comparisons were made between groupsusing the post-hoc Fisher’s LSD test (� level � 0.05).
In order to assess changes in the distribution offiber diameters between groups, the diameter ofindividual fibers of each fiber type (I, IIA, IIB orNADH-positive oxidative fibers) were grouped intothree size classes: (1) average, (2) larger than aver-age, and (3) smaller than average. The diametercut-off for grouping was based on the mean andstandard deviation (SD) of the fiber diameters forthe control mice (unexercised WT). The average-sized fibers were defined as fibers with diameterswithin �1 SD of the mean diameter of the controlmice. The larger-than-average fibers were defined asfibers larger than 1 SD above the mean diameter ofthe control mice, and the smaller-than-average fiberswere smaller than 1 SD of the mean diameter of thecontrol mice. A significant decrease in fiber size(atrophy) was defined as a statistically significantincrease in the percentage of smaller-than-averagefibers or a decrease in the percentage of larger-than-average fibers. Conversely, a significant increase infiber size (hypertrophy) was defined as a statistical
increase in the percentage of larger-than-averagefibers or a decrease in the percentage of smaller-than-average fibers.
RESULTS
Both WT and nemaline mice successfully completedthe exercise training regimen. However, the nema-line mice ran on the treadmill with an abnormal gaitcompared to the WT mice: they ran with the hind-limbs close to the body, tail dragging on the tread-mill and toes splayed laterally resulting in an archedback.
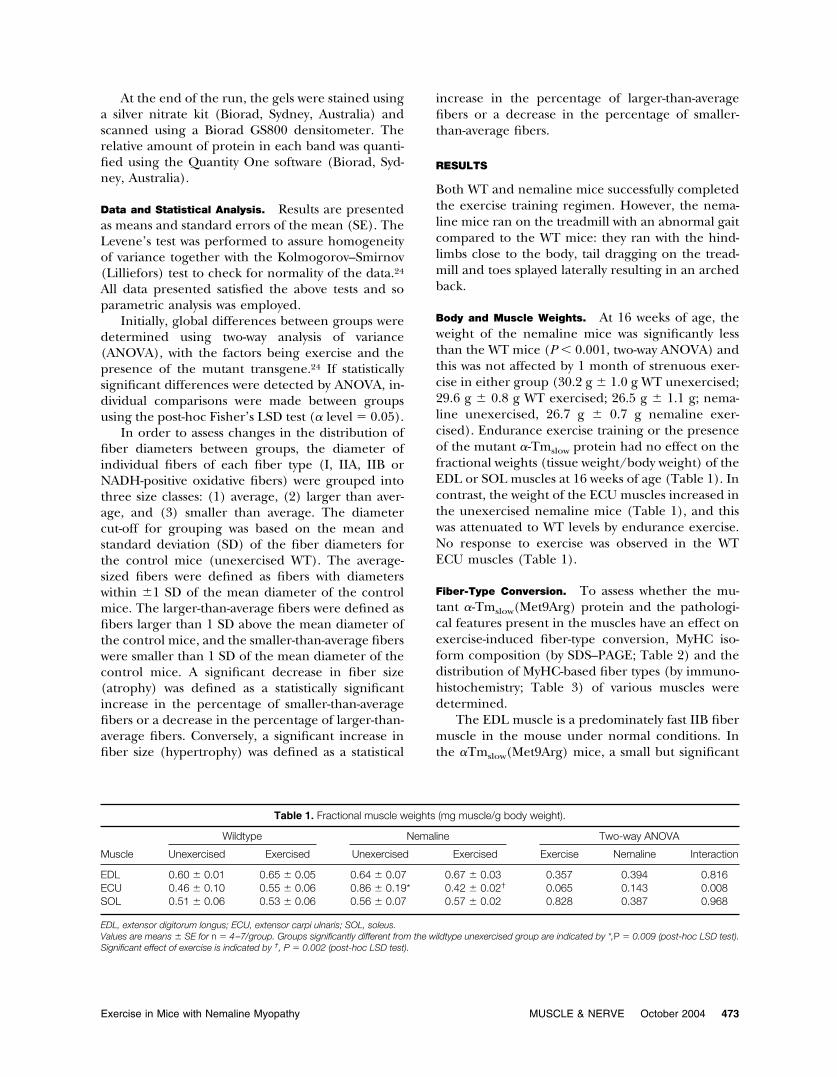
Body and Muscle Weights. At 16 weeks of age, theweight of the nemaline mice was significantly lessthan the WT mice (P � 0.001, two-way ANOVA) andthis was not affected by 1 month of strenuous exer-cise in either group (30.2 g � 1.0 g WT unexercised;29.6 g � 0.8 g WT exercised; 26.5 g � 1.1 g; nema-line unexercised, 26.7 g � 0.7 g nemaline exer-cised). Endurance exercise training or the presenceof the mutant �-Tmslow protein had no effect on thefractional weights (tissue weight/body weight) of theEDL or SOL muscles at 16 weeks of age (Table 1). Incontrast, the weight of the ECU muscles increased inthe unexercised nemaline mice (Table 1), and thiswas attenuated to WT levels by endurance exercise.No response to exercise was observed in the WTECU muscles (Table 1).
Fiber-Type Conversion. To assess whether the mu-tant �-Tmslow(Met9Arg) protein and the pathologi-cal features present in the muscles have an effect onexercise-induced fiber-type conversion, MyHC iso-form composition (by SDS–PAGE; Table 2) and thedistribution of MyHC-based fiber types (by immuno-histochemistry; Table 3) of various muscles weredetermined.
The EDL muscle is a predominately fast IIB fibermuscle in the mouse under normal conditions. Inthe �Tmslow(Met9Arg) mice, a small but significant
Table 1. Fractional muscle weights (mg muscle/g body weight).
Muscle
Wildtype Nemaline Two-way ANOVA
Unexercised Exercised Unexercised Exercised Exercise Nemaline Interaction
EDL 0.60 � 0.01 0.65 � 0.05 0.64 � 0.07 0.67 � 0.03 0.357 0.394 0.816ECU 0.46 � 0.10 0.55 � 0.06 0.86 � 0.19* 0.42 � 0.02† 0.065 0.143 0.008SOL 0.51 � 0.06 0.53 � 0.06 0.56 � 0.07 0.57 � 0.02 0.828 0.387 0.968
EDL, extensor digitorum longus; ECU, extensor carpi ulnaris; SOL, soleus.Values are means � SE for n � 4–7/group. Groups significantly different from the wildtype unexercised group are indicated by *,P � 0.009 (post-hoc LSD test).Significant effect of exercise is indicated by †, P � 0.002 (post-hoc LSD test).
Exercise in Mice with Nemaline Myopathy MUSCLE & NERVE October 2004 473
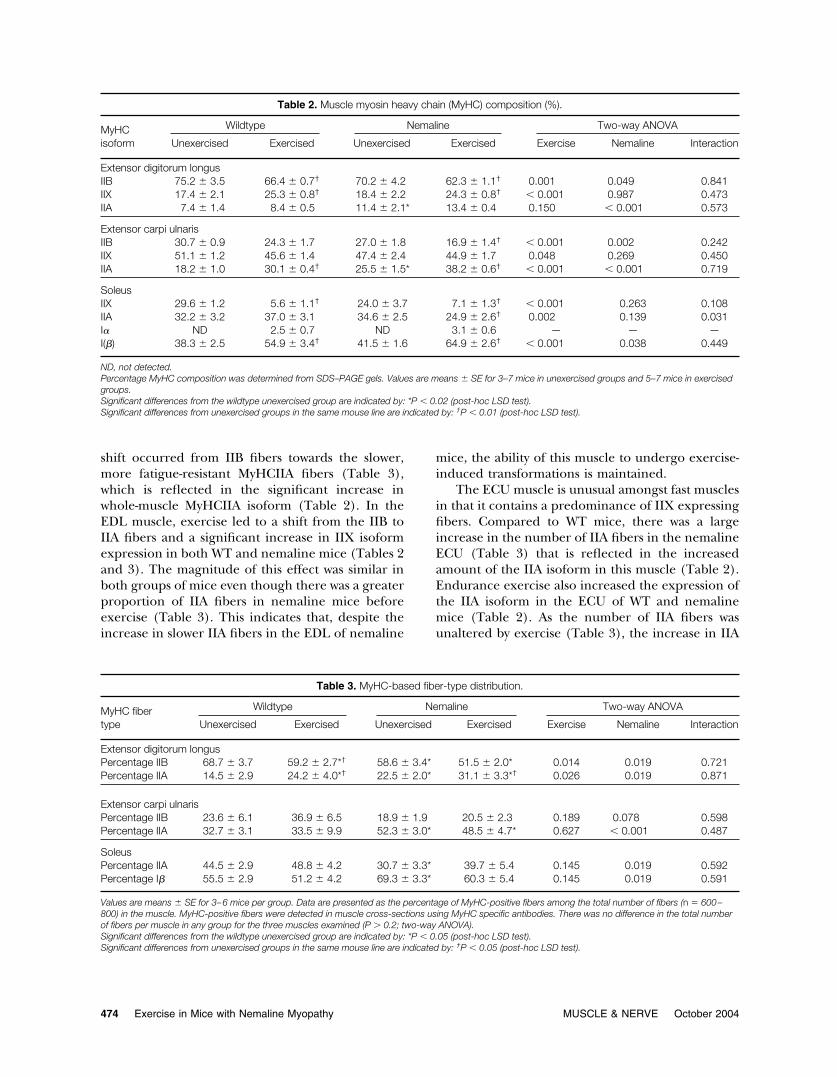
shift occurred from IIB fibers towards the slower,more fatigue-resistant MyHCIIA fibers (Table 3),which is reflected in the significant increase inwhole-muscle MyHCIIA isoform (Table 2). In theEDL muscle, exercise led to a shift from the IIB toIIA fibers and a significant increase in IIX isoformexpression in both WT and nemaline mice (Tables 2and 3). The magnitude of this effect was similar inboth groups of mice even though there was a greaterproportion of IIA fibers in nemaline mice beforeexercise (Table 3). This indicates that, despite theincrease in slower IIA fibers in the EDL of nemaline
mice, the ability of this muscle to undergo exercise-induced transformations is maintained.
The ECU muscle is unusual amongst fast musclesin that it contains a predominance of IIX expressingfibers. Compared to WT mice, there was a largeincrease in the number of IIA fibers in the nemalineECU (Table 3) that is reflected in the increasedamount of the IIA isoform in this muscle (Table 2).Endurance exercise also increased the expression ofthe IIA isoform in the ECU of WT and nemalinemice (Table 2). As the number of IIA fibers wasunaltered by exercise (Table 3), the increase in IIA
Table 2. Muscle myosin heavy chain (MyHC) composition (%).
MyHCisoform
Wildtype Nemaline Two-way ANOVA
Unexercised Exercised Unexercised Exercised Exercise Nemaline Interaction
Extensor digitorum longusIIB 75.2 � 3.5 66.4 � 0.7† 70.2 � 4.2 62.3 � 1.1† 0.001 0.049 0.841IIX 17.4 � 2.1 25.3 � 0.8† 18.4 � 2.2 24.3 � 0.8† � 0.001 0.987 0.473IIA 7.4 � 1.4 8.4 � 0.5 11.4 � 2.1* 13.4 � 0.4 0.150 � 0.001 0.573
Extensor carpi ulnarisIIB 30.7 � 0.9 24.3 � 1.7 27.0 � 1.8 16.9 � 1.4† � 0.001 0.002 0.242IIX 51.1 � 1.2 45.6 � 1.4 47.4 � 2.4 44.9 � 1.7 0.048 0.269 0.450IIA 18.2 � 1.0 30.1 � 0.4† 25.5 � 1.5* 38.2 � 0.6† � 0.001 � 0.001 0.719
SoleusIIX 29.6 � 1.2 5.6 � 1.1† 24.0 � 3.7 7.1 � 1.3† � 0.001 0.263 0.108IIA 32.2 � 3.2 37.0 � 3.1 34.6 � 2.5 24.9 � 2.6† 0.002 0.139 0.031I� ND 2.5 � 0.7 ND 3.1 � 0.6 — — —I(�) 38.3 � 2.5 54.9 � 3.4† 41.5 � 1.6 64.9 � 2.6† � 0.001 0.038 0.449
ND, not detected.Percentage MyHC composition was determined from SDS–PAGE gels. Values are means � SE for 3–7 mice in unexercised groups and 5–7 mice in exercisedgroups.Significant differences from the wildtype unexercised group are indicated by: *P � 0.02 (post-hoc LSD test).Significant differences from unexercised groups in the same mouse line are indicated by: †P � 0.01 (post-hoc LSD test).
Table 3. MyHC-based fiber-type distribution.
MyHC fibertype
Wildtype Nemaline Two-way ANOVA
Unexercised Exercised Unexercised Exercised Exercise Nemaline Interaction
Extensor digitorum longusPercentage IIB 68.7 � 3.7 59.2 � 2.7*† 58.6 � 3.4* 51.5 � 2.0* 0.014 0.019 0.721Percentage IIA 14.5 � 2.9 24.2 � 4.0*† 22.5 � 2.0* 31.1 � 3.3*† 0.026 0.019 0.871
Extensor carpi ulnarisPercentage IIB 23.6 � 6.1 36.9 � 6.5 18.9 � 1.9 20.5 � 2.3 0.189 0.078 0.598Percentage IIA 32.7 � 3.1 33.5 � 9.9 52.3 � 3.0* 48.5 � 4.7* 0.627 � 0.001 0.487
SoleusPercentage IIA 44.5 � 2.9 48.8 � 4.2 30.7 � 3.3* 39.7 � 5.4 0.145 0.019 0.592Percentage I� 55.5 � 2.9 51.2 � 4.2 69.3 � 3.3* 60.3 � 5.4 0.145 0.019 0.591
Values are means � SE for 3–6 mice per group. Data are presented as the percentage of MyHC-positive fibers among the total number of fibers (n � 600–800) in the muscle. MyHC-positive fibers were detected in muscle cross-sections using MyHC specific antibodies. There was no difference in the total numberof fibers per muscle in any group for the three muscles examined (P � 0.2; two-way ANOVA).Significant differences from the wildtype unexercised group are indicated by: *P � 0.05 (post-hoc LSD test).Significant differences from unexercised groups in the same mouse line are indicated by: †P � 0.05 (post-hoc LSD test).
474 Exercise in Mice with Nemaline Myopathy MUSCLE & NERVE October 2004
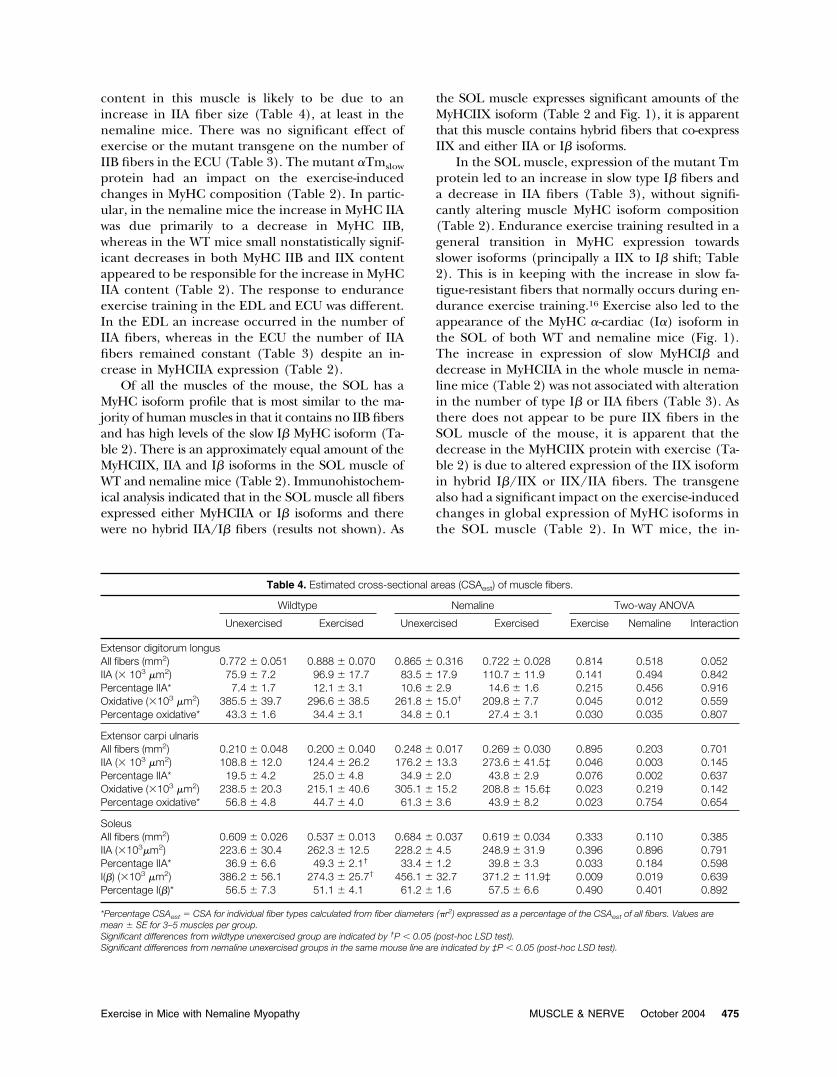
content in this muscle is likely to be due to anincrease in IIA fiber size (Table 4), at least in thenemaline mice. There was no significant effect ofexercise or the mutant transgene on the number ofIIB fibers in the ECU (Table 3). The mutant �Tmslow
protein had an impact on the exercise-inducedchanges in MyHC composition (Table 2). In partic-ular, in the nemaline mice the increase in MyHC IIAwas due primarily to a decrease in MyHC IIB,whereas in the WT mice small nonstatistically signif-icant decreases in both MyHC IIB and IIX contentappeared to be responsible for the increase in MyHCIIA content (Table 2). The response to enduranceexercise training in the EDL and ECU was different.In the EDL an increase occurred in the number ofIIA fibers, whereas in the ECU the number of IIAfibers remained constant (Table 3) despite an in-crease in MyHCIIA expression (Table 2).
Of all the muscles of the mouse, the SOL has aMyHC isoform profile that is most similar to the ma-jority of human muscles in that it contains no IIB fibersand has high levels of the slow I� MyHC isoform (Ta-ble 2). There is an approximately equal amount of theMyHCIIX, IIA and I� isoforms in the SOL muscle ofWT and nemaline mice (Table 2). Immunohistochem-ical analysis indicated that in the SOL muscle all fibersexpressed either MyHCIIA or I� isoforms and therewere no hybrid IIA/I� fibers (results not shown). As
the SOL muscle expresses significant amounts of theMyHCIIX isoform (Table 2 and Fig. 1), it is apparentthat this muscle contains hybrid fibers that co-expressIIX and either IIA or I� isoforms.
In the SOL muscle, expression of the mutant Tmprotein led to an increase in slow type I� fibers anda decrease in IIA fibers (Table 3), without signifi-cantly altering muscle MyHC isoform composition(Table 2). Endurance exercise training resulted in ageneral transition in MyHC expression towardsslower isoforms (principally a IIX to I� shift; Table2). This is in keeping with the increase in slow fa-tigue-resistant fibers that normally occurs during en-durance exercise training.16 Exercise also led to theappearance of the MyHC �-cardiac (I�) isoform inthe SOL of both WT and nemaline mice (Fig. 1).The increase in expression of slow MyHCI� anddecrease in MyHCIIA in the whole muscle in nema-line mice (Table 2) was not associated with alterationin the number of type I� or IIA fibers (Table 3). Asthere does not appear to be pure IIX fibers in theSOL muscle of the mouse, it is apparent that thedecrease in the MyHCIIX protein with exercise (Ta-ble 2) is due to altered expression of the IIX isoformin hybrid I�/IIX or IIX/IIA fibers. The transgenealso had a significant impact on the exercise-inducedchanges in global expression of MyHC isoforms inthe SOL muscle (Table 2). In WT mice, the in-
Table 4. Estimated cross-sectional areas (CSAest) of muscle fibers.
Wildtype Nemaline Two-way ANOVA
Unexercised Exercised Unexercised Exercised Exercise Nemaline Interaction
Extensor digitorum longusAll fibers (mm2) 0.772 � 0.051 0.888 � 0.070 0.865 � 0.316 0.722 � 0.028 0.814 0.518 0.052IIA (� 103 �m2) 75.9 � 7.2 96.9 � 17.7 83.5 � 17.9 110.7 � 11.9 0.141 0.494 0.842Percentage IIA* 7.4 � 1.7 12.1 � 3.1 10.6 � 2.9 14.6 � 1.6 0.215 0.456 0.916Oxidative (�103 �m2) 385.5 � 39.7 296.6 � 38.5 261.8 � 15.0† 209.8 � 7.7 0.045 0.012 0.559Percentage oxidative* 43.3 � 1.6 34.4 � 3.1 34.8 � 0.1 27.4 � 3.1 0.030 0.035 0.807
Extensor carpi ulnarisAll fibers (mm2) 0.210 � 0.048 0.200 � 0.040 0.248 � 0.017 0.269 � 0.030 0.895 0.203 0.701IIA (� 103 �m2) 108.8 � 12.0 124.4 � 26.2 176.2 � 13.3 273.6 � 41.5‡ 0.046 0.003 0.145Percentage IIA* 19.5 � 4.2 25.0 � 4.8 34.9 � 2.0 43.8 � 2.9 0.076 0.002 0.637Oxidative (�103 �m2) 238.5 � 20.3 215.1 � 40.6 305.1 � 15.2 208.8 � 15.6‡ 0.023 0.219 0.142Percentage oxidative* 56.8 � 4.8 44.7 � 4.0 61.3 � 3.6 43.9 � 8.2 0.023 0.754 0.654
SoleusAll fibers (mm2) 0.609 � 0.026 0.537 � 0.013 0.684 � 0.037 0.619 � 0.034 0.333 0.110 0.385IIA (�103�m2) 223.6 � 30.4 262.3 � 12.5 228.2 � 4.5 248.9 � 31.9 0.396 0.896 0.791Percentage IIA* 36.9 � 6.6 49.3 � 2.1† 33.4 � 1.2 39.8 � 3.3 0.033 0.184 0.598I(�) (�103 �m2) 386.2 � 56.1 274.3 � 25.7† 456.1 � 32.7 371.2 � 11.9‡ 0.009 0.019 0.639Percentage I(�)* 56.5 � 7.3 51.1 � 4.1 61.2 � 1.6 57.5 � 6.6 0.490 0.401 0.892
*Percentage CSAest � CSA for individual fiber types calculated from fiber diameters (�r2) expressed as a percentage of the CSAest of all fibers. Values aremean � SE for 3–5 muscles per group.Significant differences from wildtype unexercised group are indicated by †P � 0.05 (post-hoc LSD test).Significant differences from nemaline unexercised groups in the same mouse line are indicated by ‡P � 0.05 (post-hoc LSD test).
Exercise in Mice with Nemaline Myopathy MUSCLE & NERVE October 2004 475

crease in MyHCI� was due primarily to a decrease inMyHCIIX abundance, whereas in the nemaline micethere was a decrease in abundance of both the IIAand IIX isoforms.
Fiber Size. A change in fiber size is an importantadaptive response to endurance exercise that can leadto an increase in muscle strength. In this study wemeasured fiber diameter and fiber CSA to assess theeffect of the mutant �Tmslow protein on the trophicresponse of fibers to endurance exercise training.
In EDL muscles, there was no statistical effect ofthe mutant �Tmslow protein or endurance exerciseon the modal diameter of IIB fibers (Fig. 2A). Inmice, the oxidative fibers of the EDL muscle expresspredominantely IIA or IIX MyHC isoforms. Thepresence of the mutant �Tmslow protein led to adecrease in oxidative fiber size indicated by a de-crease in fiber diameter (Fig. 2A) and CSA (Table4). An additional decrease in oxidative fiber diame-ter occurred when the nemaline mice were exer-cised, a decrease that was similar to exercised WTmice (Fig. 2A). In nemaline mice there was a signif-icant decrease in IIA fiber diameter and an increasein diameter of these fibers with exercise (post-hocLSD test; Fig. 2A).
In the ECU, there was a significant increase in CSAof IIA fibers in nemaline mice before exercise (Table4) and an additional increase in IIA fiber diameter andCSA with exercise (Fig. 2B and Table 4, respectively).However, oxidative fiber diameter and CSA decreasedin these mice (Fig. 2B and Table 4, respectively). As IIXand IIA fibers are the only oxidative fibers in ECUmuscle, these findings indicate that the decrease inoxidative fiber size in nemaline mice with exercise wasdue to a decrease in IIX fiber size. In the ECU fromWT mice, exercise had no effect on IIA or oxidativefiber size (Fig. 2B and Table 4).
In the SOL muscle of nemaline mice there wasan overall increase (combined unexercised and ex-ercised data; P � 0.03, two-way ANOVA) in CSA of I�fibers compared to WT mice (Table 4) but no effectof the transgene on IIA fiber diameter (Fig. 2C) orCSA (Table 4). Endurance exercise decreased typeI� fiber diameters and CSA in WT and nemalinemice (Fig. 2C and Table 4, respectively), but de-creased oxidative fiber diameters in the WT but notnemaline mice (Fig. 2C).
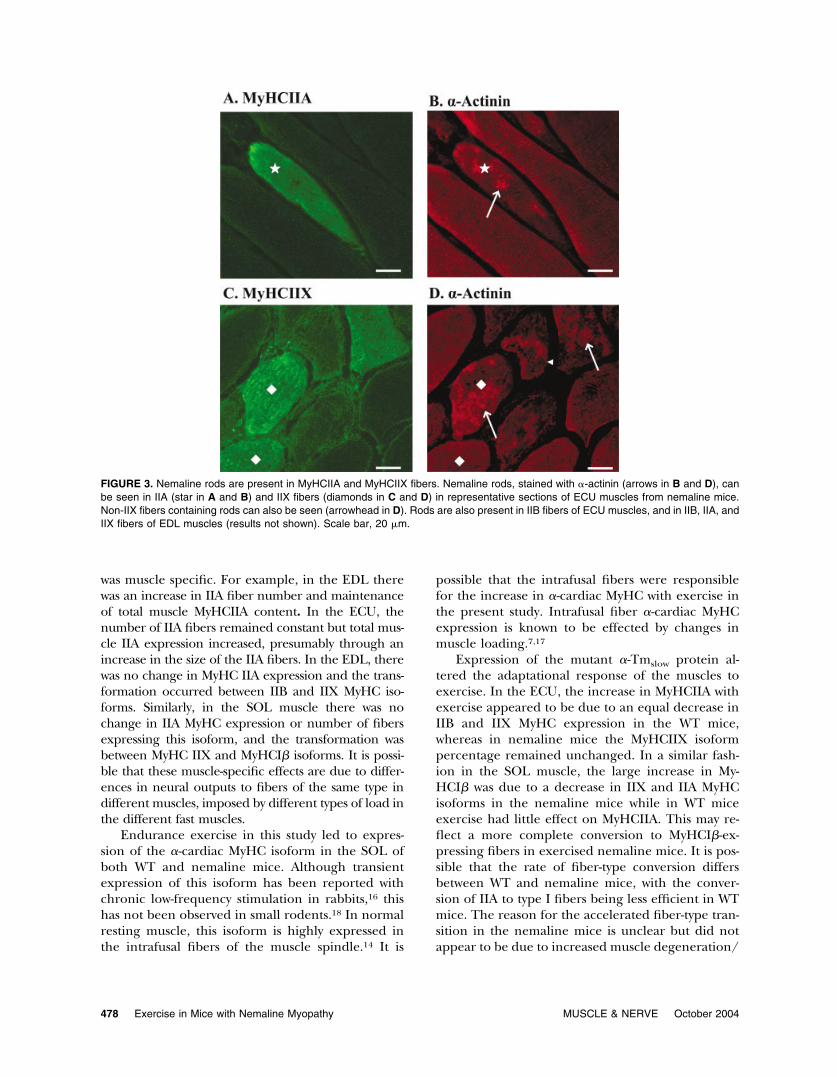
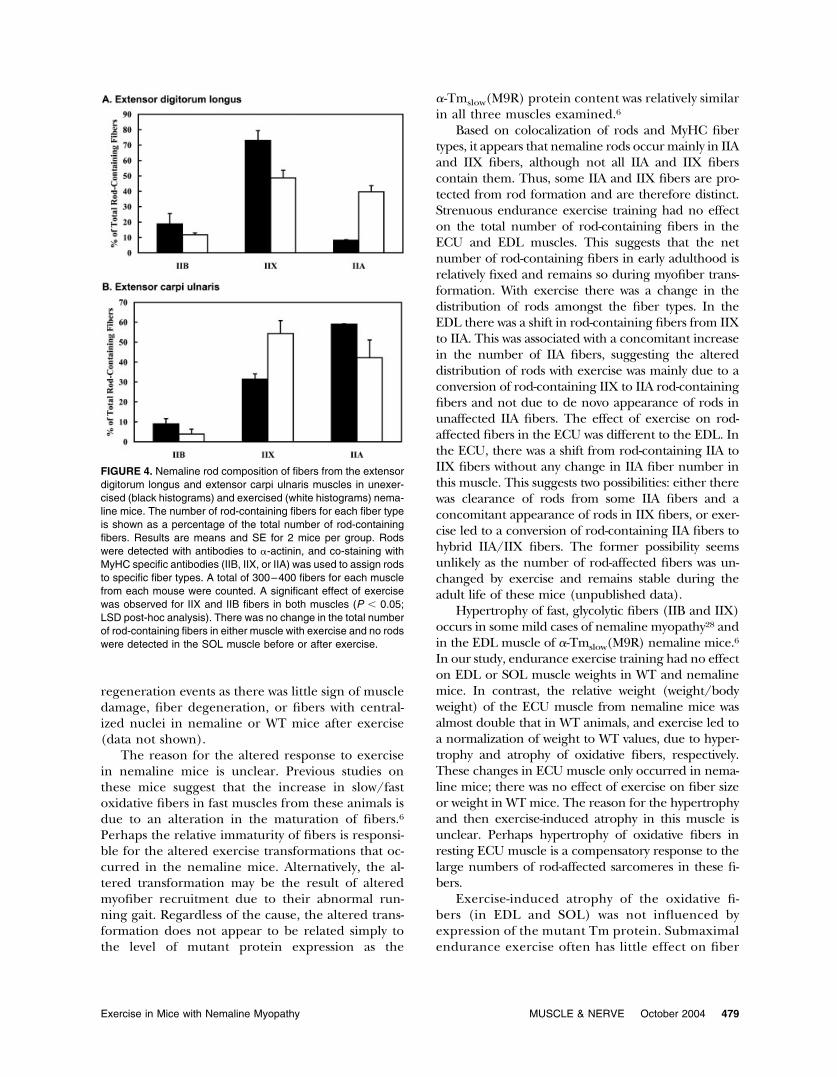
Colocalization of Rods and MyHC Fiber-Types. As re-ported previously, the majority of fibers in the ECUhad rods (�60%; Fig. 3D), whereas in the EDL rodswere present in only one third of fibers.6 Co-immu-nostaining of rods (�-actinin) and MyHC isoforms(Fig. 3) indicate that rods are mainly in IIX fibers inEDL muscles (Fig. 4A) and in IIA and IIX fibers inECU muscles (Fig. 4B) of unexercised nemalinemice. The higher proportion of rod-containing fi-bers in the ECU than EDL muscle is due to thegreater number of IIA fibers in the ECU (Table 3).There was no increase in the total number of rod-containing fibers in EDL or ECU, but the distribu-tion of rod-containing fibers between fiber types wasaltered. In the EDL muscle, exercise produced ashift in rod-containing fibers from IIX to IIA fibers(Fig. 4A), associated with an increase in the numberof MyHCIIA fibers (Table 3). Conversely, in the ECUexercise produced a shift in rod-containing fibersfrom IIA to IIX fibers (Fig. 4B).
DISCUSSION
We have used our transgenic model of nemaline my-opathy to examine whether the presence of the mutant�-tropomyosinslow protein [�-Tmslow(Met9Arg)] inmuscle alters exercise-induced myofiber transforma-tions. Endurance exercise resulted in a general shiftto slower, more fatigue-resistant fibers, but the nor-mal fast-to-slow fiber-type progression appeared tobe altered in nemaline mice. The altered myofibertransformation was not correlated with the level ofmutant �-Tmslow protein or the number of sarco-mere disruptions. The specific fibers transformed byexercise in nemaline mice varied in different mus-cles despite similar expression of the mutant�-Tmslow protein. Nemaline rods were presentmainly in IIX and IIA fibers and the total number ofrod-containing fibers remained unchanged with ex-ercise. The exercise-induced fiber transformationswere associated with a muscle-specific change in thedistribution of rods between IIX and IIA fibers.
FIGURE 1. Sodium dodecyl sulphate–polyacrylamide electro-phoresis (SDS–PAGE) gels showing myosin heavy chain(MyHC) expression in response to exercise in the SOL muscle ofWT (�) and nemaline () mice. An example of WT heart ventriclehas been included (lane 1) to show the location of the I� cardiacMyHC isoform. Note that SOL muscle does not express MyHCIIB and that the slow MyHC isoform � I.
476 Exercise in Mice with Nemaline Myopathy MUSCLE & NERVE October 2004
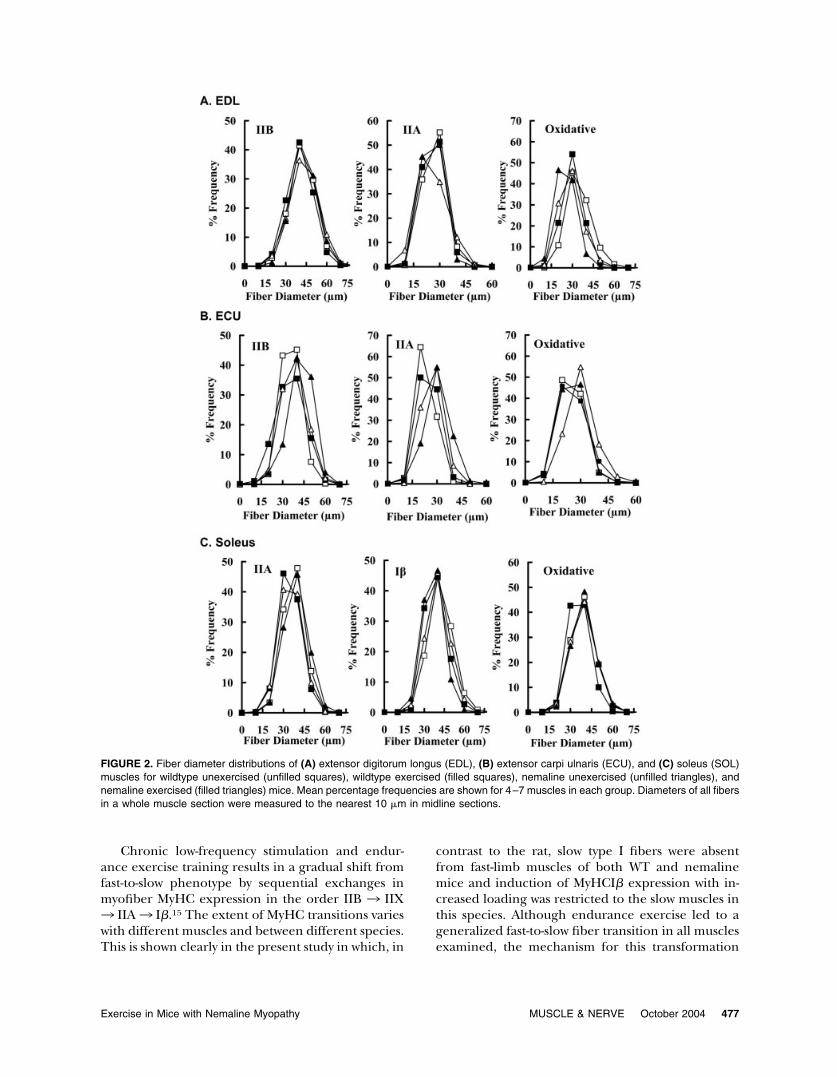
Chronic low-frequency stimulation and endur-ance exercise training results in a gradual shift fromfast-to-slow phenotype by sequential exchanges inmyofiber MyHC expression in the order IIB 3 IIX3 IIA3 I�.15 The extent of MyHC transitions varieswith different muscles and between different species.This is shown clearly in the present study in which, in
contrast to the rat, slow type I fibers were absentfrom fast-limb muscles of both WT and nemalinemice and induction of MyHCI� expression with in-creased loading was restricted to the slow muscles inthis species. Although endurance exercise led to ageneralized fast-to-slow fiber transition in all musclesexamined, the mechanism for this transformation
FIGURE 2. Fiber diameter distributions of (A) extensor digitorum longus (EDL), (B) extensor carpi ulnaris (ECU), and (C) soleus (SOL)muscles for wildtype unexercised (unfilled squares), wildtype exercised (filled squares), nemaline unexercised (unfilled triangles), andnemaline exercised (filled triangles) mice. Mean percentage frequencies are shown for 4–7 muscles in each group. Diameters of all fibersin a whole muscle section were measured to the nearest 10 �m in midline sections.
Exercise in Mice with Nemaline Myopathy MUSCLE & NERVE October 2004 477
was muscle specific. For example, in the EDL therewas an increase in IIA fiber number and maintenanceof total muscle MyHCIIA content. In the ECU, thenumber of IIA fibers remained constant but total mus-cle IIA expression increased, presumably through anincrease in the size of the IIA fibers. In the EDL, therewas no change in MyHC IIA expression and the trans-formation occurred between IIB and IIX MyHC iso-forms. Similarly, in the SOL muscle there was nochange in IIA MyHC expression or number of fibersexpressing this isoform, and the transformation wasbetween MyHC IIX and MyHCI� isoforms. It is possi-ble that these muscle-specific effects are due to differ-ences in neural outputs to fibers of the same type indifferent muscles, imposed by different types of load inthe different fast muscles.
Endurance exercise in this study led to expres-sion of the �-cardiac MyHC isoform in the SOL ofboth WT and nemaline mice. Although transientexpression of this isoform has been reported withchronic low-frequency stimulation in rabbits,16 thishas not been observed in small rodents.18 In normalresting muscle, this isoform is highly expressed inthe intrafusal fibers of the muscle spindle.14 It is
possible that the intrafusal fibers were responsiblefor the increase in �-cardiac MyHC with exercise inthe present study. Intrafusal fiber �-cardiac MyHCexpression is known to be effected by changes inmuscle loading.7,17
Expression of the mutant �-Tmslow protein al-tered the adaptational response of the muscles toexercise. In the ECU, the increase in MyHCIIA withexercise appeared to be due to an equal decrease inIIB and IIX MyHC expression in the WT mice,whereas in nemaline mice the MyHCIIX isoformpercentage remained unchanged. In a similar fash-ion in the SOL muscle, the large increase in My-HCI� was due to a decrease in IIX and IIA MyHCisoforms in the nemaline mice while in WT miceexercise had little effect on MyHCIIA. This may re-flect a more complete conversion to MyHCI�-ex-pressing fibers in exercised nemaline mice. It is pos-sible that the rate of fiber-type conversion differsbetween WT and nemaline mice, with the conver-sion of IIA to type I fibers being less efficient in WTmice. The reason for the accelerated fiber-type tran-sition in the nemaline mice is unclear but did notappear to be due to increased muscle degeneration/
FIGURE 3. Nemaline rods are present in MyHCIIA and MyHCIIX fibers. Nemaline rods, stained with �-actinin (arrows in B and D), canbe seen in IIA (star in A and B) and IIX fibers (diamonds in C and D) in representative sections of ECU muscles from nemaline mice.Non-IIX fibers containing rods can also be seen (arrowhead in D). Rods are also present in IIB fibers of ECU muscles, and in IIB, IIA, andIIX fibers of EDL muscles (results not shown). Scale bar, 20 �m.
478 Exercise in Mice with Nemaline Myopathy MUSCLE & NERVE October 2004
regeneration events as there was little sign of muscledamage, fiber degeneration, or fibers with central-ized nuclei in nemaline or WT mice after exercise(data not shown).
The reason for the altered response to exercisein nemaline mice is unclear. Previous studies onthese mice suggest that the increase in slow/fastoxidative fibers in fast muscles from these animals isdue to an alteration in the maturation of fibers.6Perhaps the relative immaturity of fibers is responsi-ble for the altered exercise transformations that oc-curred in the nemaline mice. Alternatively, the al-tered transformation may be the result of alteredmyofiber recruitment due to their abnormal run-ning gait. Regardless of the cause, the altered trans-formation does not appear to be related simply tothe level of mutant protein expression as the
�-Tmslow(M9R) protein content was relatively similarin all three muscles examined.6
Based on colocalization of rods and MyHC fibertypes, it appears that nemaline rods occur mainly in IIAand IIX fibers, although not all IIA and IIX fiberscontain them. Thus, some IIA and IIX fibers are pro-tected from rod formation and are therefore distinct.Strenuous endurance exercise training had no effecton the total number of rod-containing fibers in theECU and EDL muscles. This suggests that the netnumber of rod-containing fibers in early adulthood isrelatively fixed and remains so during myofiber trans-formation. With exercise there was a change in thedistribution of rods amongst the fiber types. In theEDL there was a shift in rod-containing fibers from IIXto IIA. This was associated with a concomitant increasein the number of IIA fibers, suggesting the altereddistribution of rods with exercise was mainly due to aconversion of rod-containing IIX to IIA rod-containingfibers and not due to de novo appearance of rods inunaffected IIA fibers. The effect of exercise on rod-affected fibers in the ECU was different to the EDL. Inthe ECU, there was a shift from rod-containing IIA toIIX fibers without any change in IIA fiber number inthis muscle. This suggests two possibilities: either therewas clearance of rods from some IIA fibers and aconcomitant appearance of rods in IIX fibers, or exer-cise led to a conversion of rod-containing IIA fibers tohybrid IIA/IIX fibers. The former possibility seemsunlikely as the number of rod-affected fibers was un-changed by exercise and remains stable during theadult life of these mice (unpublished data).
Hypertrophy of fast, glycolytic fibers (IIB and IIX)occurs in some mild cases of nemaline myopathy28 andin the EDL muscle of �-Tmslow(M9R) nemaline mice.6In our study, endurance exercise training had no effecton EDL or SOL muscle weights in WT and nemalinemice. In contrast, the relative weight (weight/bodyweight) of the ECU muscle from nemaline mice wasalmost double that in WT animals, and exercise led toa normalization of weight to WT values, due to hyper-trophy and atrophy of oxidative fibers, respectively.These changes in ECU muscle only occurred in nema-line mice; there was no effect of exercise on fiber sizeor weight in WT mice. The reason for the hypertrophyand then exercise-induced atrophy in this muscle isunclear. Perhaps hypertrophy of oxidative fibers inresting ECU muscle is a compensatory response to thelarge numbers of rod-affected sarcomeres in these fi-bers.
Exercise-induced atrophy of the oxidative fi-bers (in EDL and SOL) was not influenced byexpression of the mutant Tm protein. Submaximalendurance exercise often has little effect on fiber
FIGURE 4. Nemaline rod composition of fibers from the extensordigitorum longus and extensor carpi ulnaris muscles in unexer-cised (black histograms) and exercised (white histograms) nema-line mice. The number of rod-containing fibers for each fiber typeis shown as a percentage of the total number of rod-containingfibers. Results are means and SE for 2 mice per group. Rodswere detected with antibodies to �-actinin, and co-staining withMyHC specific antibodies (IIB, IIX, or IIA) was used to assign rodsto specific fiber types. A total of 300–400 fibers for each musclefrom each mouse were counted. A significant effect of exercisewas observed for IIX and IIB fibers in both muscles (P � 0.05;LSD post-hoc analysis). There was no change in the total numberof rod-containing fibers in either muscle with exercise and no rodswere detected in the SOL muscle before or after exercise.
Exercise in Mice with Nemaline Myopathy MUSCLE & NERVE October 2004 479

size,19 whereas oxidative fiber atrophy has beenobserved following prolonged high-intensity en-durance exercise25,27 and in rats selectively bredfor endurance running.10 The oxidative fiber atro-phy in our study may reflect the extensive natureof the exercise program. It has been suggestedthat atrophy of oxidative fibers is a normal re-sponse to extensive endurance exercise, improv-ing the efficiency of contraction and the diffusionof metabolites to the sarcomere and waste prod-ucts out of the muscle fiber.10,20
In conclusion, expression of a nemaline myopathy–producing mutation results in altered exercise-inducedtransformations that are muscle- and fiber-type specific.The total number of rod-affected fibers in muscles fromnemaline mice appears to be remarkably stable even dur-ing exercise-induced fiber transformations. This indicatesthat exercise does not exacerbate the muscle pathology inthis disease model. However, the distribution of rodsamongst fiber types is altered with exercise presumablythrough fiber-type conversion and not formation of rodsin unaffected fibers. The presence of rod-affected fibersin the muscles of nemaline mice may have altered musclefiber size and influenced the trophic response to exercise.The altered myosin heavy chain expression in nemalinemice is likely to be a consequence of the disruption of thesarcomeric thin filament by the mutant �Tmslow protein.
We gratefully acknowledge Professor Alan Beggs for providing the�-actinin antibodies. We also thank Gill Butler-Browne, Eva Chin,Mark Corbett, Peter Gunning, and Kathy North for their helpfulcomments about aspects of this work. We are indebted to WendyThornton and the other members of the BioServices Unit of CMRIfor their help in managing the animals. This investigation was sup-ported by National Medical Research Council Grant 185206.
REFERENCES
1. Agbulut O, Li Z, Mouly V, Butler-Browne GS. Analysis ofskeletal and cardiac muscle from desmin knock-out and nor-mal mice by high resolution separation of myosin heavy-chainisoforms. Biol Cell 1996;88:131–135.
2. Allen DL, Harrison BC, Maass A, Bell ML, Byrnes WC, Lein-wand LA. Cardiac and skeletal muscle adaptations to volun-tary wheel running in the mouse. J Appl Physiol 2001;90:1900–1908.
3. Baldwin KM, Haddad F. Effects of different activity and inac-tivity paradigms on myosin heavy chain gene expression instriated muscle. J Appl Physiol 2001;90:345–357.
4. Borrione AC, Zanellato AM, Saggin L, Mazzoli M, Azzarello G,Sartore S. Neonatal myosin heavy chains are not expressed inNi-induced rat rhabdomyosarcoma. Differentiation 1988;38:49–59.
5. Butler-Browne GS, Herlicoviez D, Whalen RG. Effects of hy-pothyroidism on myosin isozyme transitions in developing ratmuscle. FEBS Lett 1984;166:71–75.
6. Corbett MA, Robinson CS, Dunglison GF, Yang N, Joya JE,Stewart AW, Schnell C, Gunning PW, North KN, HardemanEC. A mutation in alpha-tropomyosin(slow) affects musclestrength, maturation and hypertrophy in a mouse model fornemaline myopathy. Hum Mol Genet 2001;10:317–328.
7. De Doncker L, Picquet F, Butler-Browne G, Falempin M.Expression of myosin heavy chain isoforms along intrafusalfibers of rat soleus muscle spindles after 14 days of hindlimbunloading. J Histochem Cytochem 2002;50:1543–1554.
8. Demirel HA, Powers SK, Naito H, Hughes M, Coombes JS.Exercise-induced alterations in skeletal muscle myosin heavychain phenotype: dose-response relationship. J Appl Physiol1999;86:1002–1008.
9. Fitts, RH, Widrick JJ. Muscle mechanics: adaptations withexercise-training. Exerc Sport Sci Rev 1996;24:427–473.
10. Howlett RA, Gonzalez NC, Wagner HE, Fu Z, Britton SL,Koch LG, Wagner PD. Selected contribution: skeletal musclecapillarity and enzyme activity in rats selectively bred forrunning endurance. J Appl Physiol 2003;94:1682–1688.
11. Laing NG, Wilton SD, Akkari PA, Dorosz S, Boundy K, Knee-bone C, Blumbergs P, White S, Watkins H, Love DR, Haan E.A mutation in the alpha tropomyosin gene TPM3 associatedwith autosomal dominant nemaline myopathy NEM1. NatGenet 1995;10:249.
12. Lucas CA, Kang LH, Hoh JF. Monospecific antibodies againstthe three mammalian fast limb myosin heavy chains. BiochemBiophys Res Commun 2000;272:303–308.
13. Morris EP, Nneji G, Squire JM. The three-dimensional structureof the nemaline rod Z-band. J Cell Biol 1990;111:2961–2978.
14. Pedrosa F, Soukup T, Thornell LE. Expression of an alphacardiac-like myosin heavy chain in muscle spindle fibres. His-tochemistry 1990;95:105–113.
15. Pette D. Training effects on the contractile apparatus. ActaPhysiol Scand 1998;162:367–376.
16. Peuker H, Conjard A, Pette D. Alpha-cardiac-like myosinheavy chain as an intermediate between MHCIIa and MHCIbeta in transforming rabbit muscle. Am J Physiol 1998;274:C595–C602.
17. Picquet F, De Doncker L, Falempin M. Expression of myosinheavy chain isoforms in rat soleus muscle spindles after 19 daysof hypergravity. J Histochem Cytochem 2003;51:1479–1489.
18. Putman, CT, Conjard A, Peuker H, Pette D. Alpha-cardiac-likemyosin heavy chain MHCI alpha is not upregulated in trans-forming rat muscle. J Muscle Res Cell Motil 1999;20:155–162.
19. Rennie, MJ, Tipton KD. Protein and amino acid metabolismduring and after exercise and the effects of nutrition. AnnuRev Nutr 2000;20:457–483.
20. Rivero, JL, Talmadge RJ, Edgerton VR. Fibre size and meta-bolic properties of myosin heavy chain-based fibre types in ratskeletal muscle. J Muscle Res Cell Motil 1998;19:733–742.
21. Schiaffino S, Gorza L, Sartore S, Saggin L, Ausoni S, VianelloM, Gundersen K, Lomo T. Three myosin heavy chain isoformsin type 2 skeletal muscle fibres. J Muscle Res Cell Motil1989;10:197–205.
22. Shafiq SA, Dubowitz V, Peterson HC, Milhorat AT. Nemalinemyopathy: report of a fatal case, with histochemical and elec-tron microscopic studies. Brain 1967;90:817–828.
23. Song, SK, Shimada N, Andrienko T. Orthogonal diameters inthe analysis of muscle fiber size and form. Nature 1963;200:1220–1221.
24. SPSS Inc. SPSS Advanced statistics manual. Chicago: SPSSInc.; 1997.
25. Swaddle JP, Biewener AA. Exercise and reduced muscle massin starlings. Nature 2000;406:585–586.
26. Talmadge RJ, Roy RR. Electrophoretic separation of rat skel-etal muscle myosin heavy-chain isoforms. J Appl Physiol 1993;75:2337–2340.
27. Tamaki N. Effect of endurance training on muscle fiber typecomposition and capillary supply in rat diaphragm. Eur J ApplPhysiol Occup Physiol 1987;56:127–131.
28. Wallgren-Pettersson C. Congenital nemaline myopathy. A clini-cal follow-up of twelve patients. J Neurol Sci 1989;89:1–14.
29. Wallgren-Pettersson C, Laing NG. Report of the 83rd ENMCInternational Workshop: 4th Workshop on Nemaline Myop-athy. Neuromuscul Disord 2001;11:589–595.
480 Exercise in Mice with Nemaline Myopathy MUSCLE & NERVE October 2004




















