
Ultraviolet visual sensitivity in three avian lineages: paleognaths, parrots, and passerines

Biological Journal of the Linnean Society (2000), 70: 121–146. With 6 figures
doi: 10.1006/bijl.1999.0396, available online at http://www.idealibrary.com on
Mouthpart evolution in adults of the basal,‘symphytan’, hymenopteran lineages
MARK JERVIS∗
Cardiff School of Biosciences, Cardiff University, Cardiff CF1 3TL
LARS VILHELMSEN
Department of Systematic Zoology, EBC, Uppsala University, Norbyvagen 18D,75236 Uppsala, Sweden
Received 4 January 1999; accepted for publication 7 July 1999
We review feeding biology and mouthpart structure generally among adults of the basalhymenopteran, or ‘symphytan’, lineages (sawflies, woodwasps, horntails and their relatives).These insects feed on a wide range of materials: floral and extrafloral nectar, pollen, plant(floral and leaf ) tissues, plant (angiosperm) sap, the juice of ripe fruit, the spermatial fluid ofrust fungi, sternorrhynchan bug honeydew, and insect tissues. Adults show feeding-relatedmouthpart specialization either for consuming pollen (the Xyelidae only) or for consuming‘concealed’ floral nectar (several families). Seven functional types of elongated proboscis or‘concealed-nectar extraction apparatus (CNEA)’ have previously been recognized amongHymenoptera. We identify an additional type, which appears to be unique among Hy-menoptera and has probably evolved directly from unspecialized mouthparts (labiomaxillarycomplex). In total, three types of CNEA are known to occur in ‘Symphyta’. Type 1 occursin Pamphiliidae, Megalodontesidae, Argidae, Pergidae, Tenthredinidae, Cimbicidae andCephidae. Type 5 occurs in Pergidae (in two unidentified species of Euryinae). Type 8 occursin Tenthredinidae (in the genus Nipponorhynchus Takeuchi). CNEA of some type or other hasarisen at least twice within the family Tenthredinidae and at least twice within the pergidsubfamily Euryinae. Evolutionary parallelism in CNEA structure has occurred between thebasal, ‘symphytan’, hymenopteran lineages and the Apocrita, a phenomenon hitherto notmentioned in the literature. Within the ‘Symphyta’, possession of Type 1 CNEA appears tobe a ground plan feature of each of the following taxa: the pergid genus Eurys Newman, themegalodontesid genus Megalodontes Latreille (the only extant representative of the Me-galodontesidae) and the tenthredinid genus Cuneala Zirngiebl, while possession of Type 8appears to be a ground plan feature of the tenthredinid genus Nipponorhynchus Takeuchi.However, in general among ‘Symphyta’, possession of CNEA is characteristic of only smalland taxonomically subordinate groups, suggesting that CNEA has evolved independentlymany times within the basal hymenopteran lineages rather than being inherited from acommon ancestor early in the evolutionary history of the Hymenoptera. In other words,ecological expediency rather than phylogenetic history mainly accounts for its distributionpattern within the basal lineages. The results of a morphological survey of ‘Symphyta’indicate that the habit of exploiting ‘concealed’ nectar, by means of CNEA, is fairly
∗Corresponding author. E-mail: [email protected]
1210024–4066/00/050121+26 $35.00/0 2000 The Linnean Society of London

M. JERVIS AND L. VILHELMSEN122
widespread (CNEA is present in at least half of the extant families), contradicting the viewheld in the literature on flower visiting that it is altogether absent. We discuss our findingsin relation to the occurrence and evolution of ‘suctorial’ mouthparts in other Hymenopteraand in non-hymenopteran insects: Mecopterida, Coleoptera and Hemiptera. 2000 The Linnean Society of London
ADDITIONAL KEY WORDS:—sawflies – parasitoids – predators – palynivores – nec-tarivores – bees – mouthparts – comparative morphology – flower visiting – evolution –phylogeny – evolutionary parallelism.
CONTENTS
Introduction . . . . . . . . . . . . . . . . . . . . . . . 122Foods and mouthparts of ‘Symphyta’ . . . . . . . . . . . . . . . 122
Foods . . . . . . . . . . . . . . . . . . . . . . . 122Mouthparts . . . . . . . . . . . . . . . . . . . . . 126
Discussion . . . . . . . . . . . . . . . . . . . . . . . 137Acknowledgements . . . . . . . . . . . . . . . . . . . . 142References . . . . . . . . . . . . . . . . . . . . . . . 142
INTRODUCTION
Jervis (1998) recognized seven functional types of mouthpart specialization (an elong-ated proboscis formed from the labiomaxillary complex) in Hymenoptera, relating tothe extraction of ‘concealed’ floral nectar. Such morphology is termed ‘concealed-nectar extraction apparatus (CNEA)’; nectar in this case is considered to be concealedinsofar as it is inaccessible to wasps of otherwise similar size/morphology that haveunspecialized mouthparts. The wasps usually either cannot enter the corolla bodily orplace their heads inside (this also applies to other forms of nectar reservoir, i.e. thoseformed by the calyx or by overlapping bracts), as either the floral aperture is too narrowor floral structures such as the stamens and the stigma form a barrier.
Broad-scale patterns in CNEA occurrence that were identified by Jervis (1998)within the Hymenoptera included a strong degree of evolutionary parallelism inCNEA structure between the Ichneumonoidea and its putative sister-group, theAculeata, and also the presence of CNEA in several ‘symphytan’ lineages as well aswithin the Apocrita. Jervis’s (1998) treatment of ‘Symphyta’ was, however, cursory.Based on examination both of the literature and of the ‘Symphyta’ collections ofThe Natural History Museum, London, and the Zoological Museum, Copenhagen,this paper sheds further light on the occurrence and evolution of CNEA within the‘Symphyta’, and brings together the hitherto highly scattered literature on thefeeding biology of adult ‘Symphyta’ generally. The ‘Symphyta’ comprises fourteenfamilies in six superfamilies (Gauld & Bolton, 1988). They are characterized onlyby the absence of the ‘wasp-waist’ observed in most Apocrita. It has long beenrecognized that the ‘Symphyta’ is not a natural group, but is simply the collectivename for the basalmost lineages within the Hymenoptera (Vilhelmsen, 1997).
FOODS AND MOUTHPARTS OF ‘SYMPHYTA’
Foods
Adult ‘Symphyta’, like ‘Parasitica’ ( Jervis, 1998), consume (floral and extrafloral)nectar, pollen, plant (floral and leaf ) tissues, plant sap, the juice of ripe fruit, the

MOUTHPART EVOLUTION IN HYMENOPTERAN LINEAGES 123
spermatial fluid of rust fungi, sternorrhynchan bug honeydew, and insect tissues.The nectar taken is of four types:
(1) the fully exposed type such as that of Apiaceae (umbellifers) and Rosaceae. Thisoccurs as either a liquid or a solid. Solidification occurs as a result of waterevaporation; unlike nectar contained within a tubular corolla or other ‘concealing’structure, it rapidly equilibrates with ambient humidity, becoming highly viscousand eventually solid. It is not known whether ‘Symphyta’ can exploit solidifiednectar.
Willis & Burkill (1895, 1903, 1908) observed feeding by the following: Tenthredoarcuata Forster (a CNEA-bearing species) consuming the nectar of Rubus sp. and ofa range of umbellifer species, Pristiphora carinata (Hartig) (a CNEA-bearing species)consuming the nectar of Saxifraga hypnoides L., and Dolerus aeneus Hartig (a CNEA-less species) consuming the nectar of Alchemilla alpina L.
(2) that situated under a flap of petal tissue as in Ranunculus repens L. (Ranunculaceae);this can be exploited by CNEA-less as well as CNEA-bearing wasps ( Jervis et al.,1993; N.D. Springate, pers. comm.), and is considered here to be unconcealed;
(3) that occurring in short, broad, tubular corollas, as in some Asteraceae (com-posites); this can be exploited by CNEA-less wasps (the nectar can be reached usingtheir unspecialized labiomaxillary complex; the glossa can be everted to some extent[Jervis et al., 1993; Jervis, 1998], see below), so it is considered here to be unconcealed.
Willis & Burkill (1903) observed the CNEA-bearing species, Tenthredo arcuata,consuming the nectar of Achillea millefolium L.;
(4) that occurring in long, narrow, tubular corollas, as in some Asteraceae andMyoporaceae. This nectar is exploited only by means of CNEA. Field observationsof concealed-nectar feeding are mentioned below (see ‘Mouthparts’).
Consumption of extrafloral nectar has been observed in Tenthredinidae: Tenthredoinstabilis (Klug), T. scalaris (Klug) (both feeding from Prunus extrafloral nectaries;mouthparts not examined in either wasp species), and Aglaostigma tesselata (Klug)(CNEA-bearing, feeding from Prunus and Vicia sativa L.) (Rathay, 1883).
Pollen feeding has been observed in several families, including the Xyelidae, theArgidae and the Tenthredinidae (especially Nematinae) (Benson, 1950; Liston, 1980;Smith, 1993). One of us (LV) has observed pollen grains in the CNEA and alimentarycanal of Megalodontes Latreille (Megalodontesidae), and pollen grains have been foundin the guts of several Cretaceous fossil Xyelidae: from the angiosperm Afropollis sp.(Winteraceae) (Caldas, Martins-Neto & Lima-Filho, 1989) and from pinaceous andother gymnospermous trees (Krassilov & Rasnitsyn, 1983). Pollen taken by extantspecies includes that of anemophilous trees such as Salix and Pinus sylvestris L., andthat of entomophilous plants such as Apiaceae, Rosaceae and Ranunculaceae(Cameron, 1882; Hobby, 1932b; Benson, 1950). Xyela Dalman adults apparentlyfeed on a wide range of gymnospermous and non-gymnospermous pollen sources(e.g. see Smith, 1993). It is also likely that pollen grains which become trapped andso contaminate nectar and other liquids (Todd & Vansell, 1942; Willmer, 1980) arealso consumed by some floral nectar-feeders and moisture drinkers (‘Symphyta’ areknown to imbibe free water such as dew and cuckoo spit, see below), as an incidentalconsequence of imbibing the liquid (see Jervis [1998] for discussion of other wasps).
Plant floral tissues eaten include petals and stamens (Willis & Burkill, 1908;

M. JERVIS AND L. VILHELMSEN124
Rohwer, 1913; Benson, 1950). Willis & Burkill (1908) observed Tenthredo arcuata (aCNEA-bearing species) eating those of Valeriana officinalis L.. The same species wasalso recorded “devouring the flower” of Ranunculus acris L.. Benson (1950) mentionsthat sawflies of the genus Corynis Thunberg (Cimbicidae) (species not indicated;several bear CNEA) consumes petals, particularly those of Ranunculus (the nectariesin Ranunculus occur at the bases of the petals [Percival, 1965], so the wasps maytherefore consume nectar as well as floral tissues). Stamen filaments may, in somecases, impede exploitation of exposed floral nectar (see ‘Parasitica’ [Patt et al., 1997]),and so may be eaten partly in order to gain access. For example, Rohwer (1913)describes how a female of Tenthredo arcuata (a CNEA-bearing species) completelydevoured four stamens of an umbellifer prior to feeding on the nectar. Macroxyelaferruginea (Cresson) (Xyelidae) eats the inner parts of flower buds (Garlick, 1923).
‘Symphyta’ are also known to eat non-floral tissues (Chawner, 1921; Garlick,1923; Ross, 1933; Woolatt, 1946). The tenthredinids Macrophya punctumalbum (L.)(CNEA-less) and Sciapteryx consobrina (Klug) (CNEA-less) chew leaves, whereas thetenthredinid Nematus oligospilus Forster (CNEA-less) and the argid Sphacophilus apios(Ross) (mouthparts not examined) graze on the leaf pubescence. Macroxyela ferrugineaeats the inner parts of leaves, as well as of flower buds (see above).
Plant (angiosperm) sap feeding has been recorded in some species. Dolerus unicolor(Beauvois) (Tenthredinidae) (CNEA-less) consumes the sap exuding naturally frommaple buds (Goulet, 1986), and Cimbex americana Leach (Cimbicidae) (CNEA-less)makes transverse incisions with its mandibles into the bark of small branches andthen imbibes the sap (Stein, 1974).
Some nematine Tenthredinidae have been observed consuming the juice of ripefruit (Benson, 1950).
According to Benson (1950), there is a marked tendency for adults of ‘Symphyta’to exploit the larval host plants as food. The five records of non-floral plant tissuefeeding by adults, involving the tenthredinids Macrophya punctumalbum, Sciapteryxconsobrina and Nematus oligospilus, the argid Sphacophilus apios, and the xyelid Macroxyelaferruginea (Chawner, 1921; Woollatt, 1946; Ross, 1933; Garlick, 1923), involvedfeeding on the larval host plant species. Macroxyela ferruginea adults were also recordedfeeding on the tissues of plant species not used by larvae, but nevertheless showeda marked preference for the larval food plant (Garlick, 1923). According to Liston(1980), adults of certain species of Amauronematus Konow (Tenthredinidae) (a mostlyCNEA-bearing genus) feed exclusively on the catkins of the larval food plant (Salix).He also mentions that adults of Abia candens Konow (Cimbicidae) (a CNEA-bearingspecies) prefer to feed at the inflorescences of the larval food plant, Succisa pratensisMoensch (Dipsacaceae), although they will also visit those of umbellifers. Linsley &MacSwain (1959) mention that both adults and larvae of Tenthredo pectoralis Norton(CNEA-less) feed on the floral tissues (not specified) of Ranunculus californicus Bentham.Notable exceptions to Benson’s (1950) aforementioned generalization include: (a)Dolerinae (Tenthredinidae), whose larvae feed on grasses, sedges, rushes or horsetailsbut whose adults visit other plants for feeding e.g. Dolerus aeneus sometimes eats thepollen of Pinus sylvestris; (b) Xyela julii (Brebisson) (CNEA-less) whose larvae feed onthe staminate ‘flowers’ of P. sylvestris but whose adults prefer to eat the pollen of Betulaor Cytisus (Felton, 1956); and (apparently) (c) the tenthredinid genus NipponorhynchusTakeuchi (CNEA-bearing), whose whose larvae are thought to feed on ferns, butwhose adults feed mainly, if not exclusively, on angiosperm concealed-nectar (aninference based on the possession of CNEA in that genus, see below). [With regard

MOUTHPART EVOLUTION IN HYMENOPTERAN LINEAGES 125
to floral preferences, Liston (1980) suggests that in small, weakly flying species, floralspecificity may be particularly advantageous, as it increases the chances of findinga mate. There are no established cases of sawflies having a mating system in whichmales search for females specifically at feeding sites, as opposed to other sites (for aputative example among Hymenoptera ‘Parasitica’, see Belokobylskij & Jervis [1998]).However, Liston mentions that male sawflies (apparently meaning Tenthredinidae)tend to fly from flower to flower, whereas females are more sedentary; this behaviouris consistent with a ‘males search for females at feeding sites’ strategy but the systemis considered most likely to occur in species where females feed predictably at welldefined feeding sites [Godfray, 1994].]
Feeding on sternorrhynchan bug honeydew is practised by several ‘Symphyta’.Rathay (1883) recorded Dolerus ?pratensis (L.) and Macrophya crassula (Klug) (CNEA-less) feeding on the honeydew of Aphis euonymi (Fab.). Zoebelein (1955) observedseveral other Tenthredinidae feeding on honeydew from the following sources: aphid(Aphis sambuci L. on Sambucus nigra L.), coccid (Eulcanium pulchrum King. on Quercusrobur L.), Physokermes piceae Schrk. on Picea excelsa [Lamb.] Link) or psyllid (Psyllacrataegi Schrk. on Crataegus laevigata (Poiret) DC.); all are tree-associated Sternor-rhyncha. Diprion pini (L.) (Diprionidae) (CNEA-less) was recorded feeding on thehoneydew of the pine aphid Cinara pini L. (Zoebelein, 1955). It is unclear as towhether the honeydew consumed in these cases was liquid, solid or occurred inboth states (like exposed nectar, honeydew rapidly solidifies).
The sugar-containing spermatial fluid of rust fungi is known to be consumed bysome Tenthredinidae: Dolerus ?nigratus (Muller), Dolerus sp., Aglaostigma tessellata (Klug)(CNEA-bearing), ?Hoplocampa chrysorrhoea (Klug) and Macrophya punctumalbum (CNEA-less) (Rathay, 1883; Buller, 1950).
Members of several genera of Tenthredinidae and one genus each of Xiphydriidaeand Cimbicidae have been recorded preying on insects (Hobby, 1932a,b,c; Korolkov,1914; Rohwer, 1913; Matsumura, 1927; Iwata, in Clausen, 1940; Benson, 1950;Okutani, 1957; Togashi, 1959, 1978; Wolf, 1968; Liston, 1980; Preston-Mafham &Preston-Mafham, 1993) (reports of insectivory by Siricidae have not been cor-roborated [Chrystal, 1928]). Apparently, the habit is practised mainly by females(Hobby, 1932c), and often takes place on flowers such as umbellifers (e.g. see Rohwer[1913]). Prey include other sawflies (Tenthredinidae, including conspecific males),Diptera (Bibionidae, Tipulidae, Ceratopogonidae, Empididae, Syrphidae, Strati-omyidae, Scathophagidae, Calliphoridae, Fanniidae, Tachinidae), Coleoptera (Can-tharidae, Chrysomelidae, Mordellidae, Nitidulidae), Hymenoptera (Braconidae,Ichneumonidae, Sphecidae, Apidae), Plecoptera (Perlidae), Odonata (Pla-tycnemididae) Hemiptera (Aphididae, Psyllidae and Aphrophoridae) and Lepidoptera(Geometridae, Bombycidae and Lymantriidae). The aphrophorid prey eaten werelarvae enclosed by their cuckoo-spit which the sawfly also consumed (Benson, 1950).The lymantriid prey record is by L.O. Howard, of Cimbex americana captured flyingwith a gypsy moth caterpillar held in its mandibles (Anonymous, 1896). Matsumura(1927) similarly reported on Xiphydria kawakami Matsumura (mouthparts not ex-amined) carrying, in flight, its caterpillar prey. Presumably the prey in these caseswas being transported to a site where feeding could take place undisturbed, e.g. bypredators.
Murray (1927) observed an adult of Tenthredo arcuata (CNEA-bearing) feeding onan adult of Empis sp. that was itself in the process of consuming a predator (Dolichopussp.); given the probable generalist habits of adults of the three predators in the

M. JERVIS AND L. VILHELMSEN126
interaction, this is possibly an example of intraguild predation (i.e. the predatorscould belong to a guild exploiting the same prey species, see Rosenheim et al.[1995]). Tenthredo nigropicta (Smith) (mouthparts not examined) consumed up to threefull-grown larvae of its chrysomelid beetle prey (Melasoma, Plagiodera) per day (Iwata,in Clausen, 1940). Adults of Rhogogaster Konow (whose members examined so farare CNEA-less) are known to be particularly voracious predators (Liston, 1980).Whether any ‘Symphyta’ are exclusively predacious as adults is not known (althoughTenthredo nigropicta is a prime candidate); some tenthredinids are known to feed bothon prey and on floral foods (Cameron, 1882).
Some of the prey of ‘Symphyta’ are actually or potentially ‘beneficial’ insects (seeabove). For example, the prey of Xiphydria kawakami include silkworms; Matsumura(1927) regarded the woodwasp as a pest, but it is not known if this is its currentstatus.
Consumption of more than one type of food material is likely to be commonamong ‘Symphyta’ having unspecialized mouthparts, but there is a paucity ofinformation on this subject. Tenthredo arcuata consumes insect and plant tissues aswell as floral nectar (see above), as might many other CNEA-bearing species (seebelow).
Wolf (1968) observed a female of Tenthredo livida L. (CNEA-less) masticating aseries of faecal pellets from caterpillars of another tenthredinid, Nematus salicis (L.).Each pellet was masticated for up to two minutes, but in no case was a pelletingested. This behaviour is likely to be an opportunistic means of obtaining moistureand/or nutrients, and could be common among groups of ‘Symphyta’ whose femalessearch for oviposition sites in leaves.
Mouthparts
The mouthparts sensu lato of the ‘typical’ ‘symphytan’ (e.g. as seen in the membersof the superfamily Tenthredinoidea) comprise a symmetrical labrum (not strictly amouthpart), a pair of at most slightly asymmetrical mandibles, paired maxillae, anunpaired labium, and a hypopharynx.
In predatory ‘Symphyta’ the mandibles are used to puncture the integument ofthe prey and then cut the tissues into fragments; active movements of othermouthparts (maxillae, labium, hypopharynx, see below) are presumably involved iningestion (for a review of the mechanics of feeding in chewing predators, see Evans[1964], Evans & Forsythe [1985] and Chapman [1995]). The same would apply tofeeding on plant tissues (e.g. see Rohwer [1913] on ‘symphytans’).
It is possible that honeydew feeding symphytans use their mandibles to scrapedried honeydew from surfaces (e.g. leaves), as in some parasitoid wasps (see Jervis[1998]); it is unclear how the honeydew fragments are then processed prior toingestion.
Specialization of the mandibles, the labrum, the hypopharynx and the maxillaefor pollen feeding occur in Xyelidae but apparently not in other families containingpollen feeders (Argidae, Tenthredinidae, Megalodontesidae). The mandibles ofxyelids are highly asymmetrical, and their molar (grinding) surfaces operate in themanner of a pestle and mortar (Fig. 1A,B). The labrum bears a group of closelyjuxtaposed spines and hairs on its epipharyngeal (inner) surface (Vilhelmsen, 1996);this ‘epipharyngeal brush’, which lies to one side of the labral mid line, is used to

MOUTHPART EVOLUTION IN HYMENOPTERAN LINEAGES 127
Figure 1. Left (A) and right (B) mandibles (posterior view) of Macroxyela ferruginea (Say), and maxillarypalp (C) of Xyela julii (Brebisson). Scale bars=100 lm.
clean the mandibles of pollen. The hypopharynx has a distinct concavity lined withtransverse rows of cuticular teeth (the ‘infrabuccal pouch’ or ‘triturating basket’) onits anterior surface, opposite the mandibles (Vilhelmsen, 1996). It serves to processfurther the pollen after the grains have been crushed by the mandibles. [Note,however, that pollen grains do not need to be ground by insects in order for nutrientsto be extracted from them (Gilbert, 1981; Haslett, 1983).] The role of the maxillaein gathering pollen is discussed below.
A similar system, including the presence of asymmetrical mandibles, severalepipharyngeal brushes, an infrabuccal pouch, and elongate maxillary palps, is alsoobserved in the pollen feeding moth Micropteryx calthella (L.) (Hannemann, 1956) and

M. JERVIS AND L. VILHELMSEN128
other basal Lepidoptera (see Vilhelmsen, 1996); its presence has been suggested byKristensen (1984) to be a ground plan feature of the Lepidoptera. An epipharyngealbrush + asymmetrical mandibles is found in the psocodean Amphigeronia bifasciataLatreille (Vilhelmsen, unpublished), and an epipharyngeal brush + infrabuccalpouch are present in the basal coleopteran Priacma serrata Leconte. Pollen feedinghas not been demonstrated for these two taxa, however. It is difficult to decidewhether these features have evolved independently in the different taxa possessingthem, or whether the similarities reflect common descent; in the latter case, this‘triturating basket’-complex must have been lost within Hymenoptera, Lepidopteraand Coleoptera.
Feeding on liquids (e.g. nectar, sap, honeydew and fungal spermatial fluid)involves exclusively the labium and/or maxillae (for the condition in CNEA-lessHymenoptera, see Jervis’s [1998] figure 1, and also various figures in Saini & Dhillon[1979a,b] and Vilhelmsen [1996]). These structures are firmly connected by amembrane (“oral field” of Duncan [1939]) to form the labiomaxillary complex whichis operated as a unit, independently of the mandibles. Each maxilla comprises abasal sclerite or cardo, at the distal end of which a stipes is articulated. Each stipesbears three structures: a maxillary palpus laterally, a galea (usually hairy) and a mediallacinia. The labium comprises the postmentum basally and the prementum distally. Theprementum bears laterally a pair of labial palpi and distally a glossa and a pair ofparaglossae. The glossa and paraglossae in a ‘typical’ ‘symphytan’ are of about equallength, having transverse rows of minute cuticular hairs that are flattened to somedegree, covering their anterior surfaces (Vilhelmsen, 1996). The variation in theconfiguration of the distal part of the labium exhibited by the Xyelidae confoundsinferences as to whether or not this is the ground plan state for the Hymenoptera(Vilhelmsen, 1996).
The hypopharynx also requires mentioning here: this mouthpart structure, whichis not part of the labiomaxillary complex per se, is a mostly membranous lobeseparated from the prementum by the salivary outlet; it lies posteroventrally to themouth and forms the posterior wall of the cibarium (which is the anterodorsal partof the preoral cavity). Situated in the hypopharynx, and forming the floor of thecibarium, is the sitophore (‘pharyngeal plate’ of some authors) (Vilhelmsen, 1996).
In Hymenoptera CNEA has been formed through elongation and other modi-fication of various parts of the labiomaxillary complex, including in one case thehypopharynx (see Type 8 CNEA, below). The seven functional types of CNEApreviously recognized within the order are as follows (see Jervis, 1998) (Table 1):
Type 1. Slightly to moderately elongated glossa, galeae and, in some taxa, paraglossae;glossa not greatly elongated relative to the galeae, and either rounded distally or atmost only shallowly forked; prementum elongated in some species; nectar extraction(=‘nectar loading’, see below) performed by the glossa (paraglossae also involvedin some ‘Symphyta’ and Aculeata).
Type 2. Elongated glossa and galeae; glossa greatly elongated relative to the galeaeand very deeply forked; prementum elongated in some species; nectar extractionperformed by glossa.
Type 3. Resembling Type 1 but with maxillary palps markedly elongate; nectarextraction performed by glossa and (if deployed) maxillary palps (=auxiliary nectarextraction mechanism is some circumstances).

MOUTHPART EVOLUTION IN HYMENOPTERAN LINEAGES 129
T 1. CNEA recorded among Hymenoptera. (∗=pressent in parasitoid members)
Type of CNEA
Taxon 1 2 3 4 5 6 7 8
SPamphiloideaPamphiliidae +Megalodontesidae +TenthredinoideaPergidae + +Argidae +Cimbicidae +Tenthredinidae + +CephoideaCephidae +A‘P’
EvaniodeaGasteruptiidae +IchneumonoideaIchneumonidae∗ + + + +?Braconidae∗ + + + + + + +ChalcidoideaLeucospidae∗ +AChrysidoideaChrysididae∗ +VespoideaPompilidae∗ +Vespidae + +ApoideaSphecidae +Apidae + + + + +
Type 4. Elongated glossa, galeae and labial palps; glossa exposed for much of itslength in dried specimens where the CNEA is everted; labial palps closely alignedwith glossa; nectar extraction performed by glossa and labial palps.
Type 5. Elongated maxillary palps used for nectar extraction.
Type 6. Elongated glossa and labial palps; labial palps used for nectar extraction.
Type 7. Elongated labial palps enclosed within elongated maxillary palps, all fourstructures used for nectar extraction.
Jervis (1998) pointed to the possible existence of an additional type of CNEA(Type 8), occurring in Scoliidae; while CNEA has indeed been found to exist in thisgroup (Saini & Dhillon, 1979a,b), it appears to be of Type 1 (see figure 15 in Saini& Dhillon [1979b]).
The following Types of CNEA occur in ‘Symphyta’:
Type 1. This was recorded in a diverse range of taxa (genus names follow Abe &Smith [1991]). Note that in some (e.g. see discussion of Megalodontes), elongation of

M. JERVIS AND L. VILHELMSEN130
Figure 2. Type 1 CNEA of Megalodontes olivieri (Megalodontesidae) showing the maxillary palps, thelabial palps, the left galea, the left paraglossa and the glossa. Taken using a low-vacuum SEM andreproduced by permission of The Natural History Museum and its Trustees.
the glossa and paraglossae is negligible (not evident even), only the prementum andgaleae being ‘produced’ as such (e.g. merceti Konow).
Pamphiliidae: (Pamphiliinae) Onycholyda sertata (Konow), Pamphilius inanitus Villersand P. semicinctus (Norton).
Megalodontesidae: Megalodontes escalerai Konow, M. flabellicornis (Germar), M.interruptus Gussakovskij, M. kuznetsovi Dovnar-Zapolskij, M. merceti, M. nitens (Frey-muth), M. olivieri (Brulle) (Fig. 2), M. skorniakowii (Freymuth), M. xanthosomusZhelochovtsev.
CNEA occurs in all species of Megalodontes, but varies somewhat (Springate, 1995),although it corresponds to Type 1 in all cases (but see below). Only the two extremeforms of CNEA are mentioned here: (i) the shortest, with the glossa barely longerthan the galeae, distinctly flared and strongly downwardly curved, and the galeaeswollen apically (judaicus Lepeletier, kuznetsovi, merceti); (ii) the longest, with the glossamuch longer than the galeae, parallel-sided and slightly downwardly curved, andthe galeae elongate (most species within the genus).
Argidae: (Arginae) Arge katangana Forsius, Arge sauteri (Enslin), Cibdela janthina (Klug),(Sterictiphorinae) Didymia albipes (Konow) and Trochophora lobata (Erichson).
Pergidae: (Pterygophorinae), Pterygophorus cinctus Klug, P. distinctus (Rohwer) (Fig.3), P. insignis Kirby, Lophyrotoma analis (Costa) L. interrupta (Klug), L. opima (Konow),L. zonalis (Rohwer), (Euryinae) Clarissa atrata Turner, C. divergens Kirby, C. hebeBenson, C. tasbates Naumann (see Naumann, 1997), Eurys aeratus Newman, E. aglaiaBenson, E. bellus Rohwer, E. calliphenges Benson, E. chloe Benson, E. laetus (Westwood)(Fig. 4A,B), E. niditus Kirby, E. nitens (Kirby), E. pulcher Benson and E. rutilans Morice(Fig. 4C).
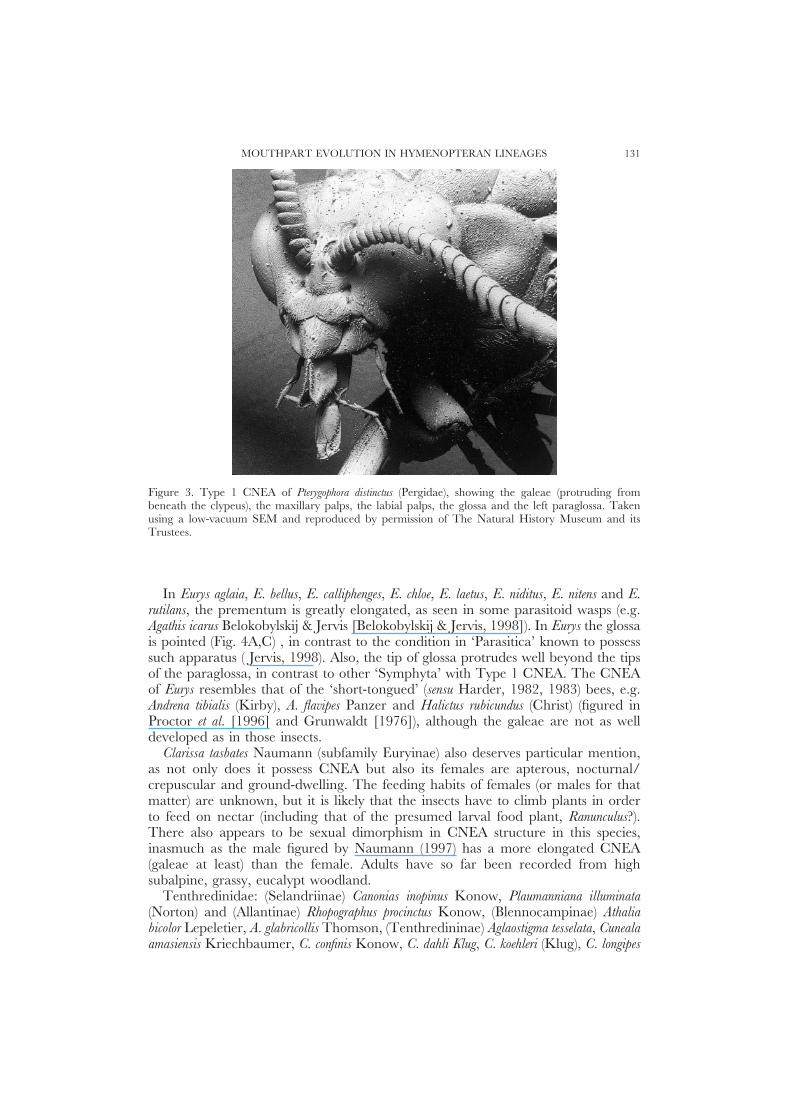
MOUTHPART EVOLUTION IN HYMENOPTERAN LINEAGES 131
Figure 3. Type 1 CNEA of Pterygophora distinctus (Pergidae), showing the galeae (protruding frombeneath the clypeus), the maxillary palps, the labial palps, the glossa and the left paraglossa. Takenusing a low-vacuum SEM and reproduced by permission of The Natural History Museum and itsTrustees.
In Eurys aglaia, E. bellus, E. calliphenges, E. chloe, E. laetus, E. niditus, E. nitens and E.rutilans, the prementum is greatly elongated, as seen in some parasitoid wasps (e.g.Agathis icarus Belokobylskij & Jervis [Belokobylskij & Jervis, 1998]). In Eurys the glossais pointed (Fig. 4A,C) , in contrast to the condition in ‘Parasitica’ known to possesssuch apparatus ( Jervis, 1998). Also, the tip of glossa protrudes well beyond the tipsof the paraglossa, in contrast to other ‘Symphyta’ with Type 1 CNEA. The CNEAof Eurys resembles that of the ‘short-tongued’ (sensu Harder, 1982, 1983) bees, e.g.Andrena tibialis (Kirby), A. flavipes Panzer and Halictus rubicundus (Christ) (figured inProctor et al. [1996] and Grunwaldt [1976]), although the galeae are not as welldeveloped as in those insects.
Clarissa tasbates Naumann (subfamily Euryinae) also deserves particular mention,as not only does it possess CNEA but also its females are apterous, nocturnal/crepuscular and ground-dwelling. The feeding habits of females (or males for thatmatter) are unknown, but it is likely that the insects have to climb plants in orderto feed on nectar (including that of the presumed larval food plant, Ranunculus?).There also appears to be sexual dimorphism in CNEA structure in this species,inasmuch as the male figured by Naumann (1997) has a more elongated CNEA(galeae at least) than the female. Adults have so far been recorded from highsubalpine, grassy, eucalypt woodland.
Tenthredinidae: (Selandriinae) Canonias inopinus Konow, Plaumanniana illuminata(Norton) and (Allantinae) Rhopographus procinctus Konow, (Blennocampinae) Athaliabicolor Lepeletier, A. glabricollis Thomson, (Tenthredininae) Aglaostigma tesselata, Cunealaamasiensis Kriechbaumer, C. confinis Konow, C. dahli Klug, C. koehleri (Klug), C. longipes

M. JERVIS AND L. VILHELMSEN132
Figure 4. Type 1 CNEA of: (A&B) Eurys laetus, and (C) E. rutilans (Pergidae). It is fully everted in (A)and (C) and fully retracted in (B). Abbreviations: ga, galea; gl, glossa; lp, labial palp; mp, maxillarypalp; pg, paraglossa. Reproduced from Benson (1934) by permission of the Royal EntomologicalSociety of London.
Konow, Tenthredo arcuata Forster, T. acerrima Benson, T. algoviensis Enslin, T. largiflavaEnslin, T. schaefferi Klug, T. schaefferi form perkinsi Morice, T. umbrica Benson, Zaschizonyxpluricincta (Norton), (Nematinae) Pristiphora carinata, Amauronematus amentorum Forster,A. amplus Konow, A. articola Dalla Torre, A. dahlbomi Thomson, A. enslini Lindqvist,A. fallax Lepeletier, A. groenlandicus Malaise, A. hebes Konow, A. helleni Lindqvist, A.hulteni Malaise, A. hyperboreus Thomson, A. latiserra Malaise, A. microphyes Forster, A.stordalensis Strand, A. taeniatus (Lepeletier), A. tillbergi Malaise.
Type 1 CNEA appears to be very widespread within Tenthredo L. and AmauronematusKonow and may be a ground plan feature for each genus. It is very likely such afeature for the genus Cuneala Zirngiebl. In the genus Athalia Leach Type 1 occursin some members of each of the bicolor Lepeletier (e.g. bicolor itself ), fumipennis Konowand glabricollis Thomson species-groups (e.g. glabricollis itself ) (see Benson, 1962), andhas obviously evolved independently at least three times within the genus.
In Cuneala the glossa and paraglossae (the tips of the latter protrude beyond thetip of the glossa) are exposed (i.e. not concealed by the galeae) for most of theirlength (c. 0.7 of glossa, c. 0.75 of paraglossae in C. koehleri), which is unusual forType 1 CNEA (being more characteristic of Type 2). In many dried specimens the

MOUTHPART EVOLUTION IN HYMENOPTERAN LINEAGES 133
glossa and paraglossa have become separated, giving the proboscis a distinctly trifidappearance. However, overall the CNEA most closely corresponds to Type 1.
Cimbicidae: (Abiinae) Abia candens, A. sachalinensis Takeuchi, Orientabia japonica(Cameron), (Coryninae) Corynis concinna (Stein), C. crassicornis (Rossi), C. frontina(Konow), C. lateralis (Brulle), C. orientalis (Konow), C. sp. nr sanguinea (Vollenhoven),Zaraea fasciata (L.) and Z. triangularis Takeuchi.
Cephidae: (Cephinae) Hartigia linearis (Schrank), Janus kashivorus Yano & Sato,Calameuta filiformis (Eversmann), C. gaullei (Konow), Cephus cultratus Eversmann, C.nigrinus Thomson, C. pygmeus L., Cephus japonicus (Forsius) (?), and Characopygus moriceiKonow.
As with an unspecialized labiomaxillary complex, removal of nectar by Type 1CNEA is attributable in the main to repeated eversion and retraction of theprementum and thus of the glossa and paraglossae (see Jervis, 1998). The glossaand paraglossae, which are densely covered with presumably hydrophilic hairs (seebelow) and together form the functional ‘tongue’, are slightly rolled at their edgesin many CNEA-bearing ‘symphytans’ possessing a non-pointed glossa. In Cunealaand Eurys the pointed glossa appears to be semi-tubular with a ventral groove. Asin the unspecialized labiomaxillary complex, saliva issues from the salivary outletthat opens onto the anterior surface of the glossa (see Vilhelmsen’s [1996] figure1A), but the route it then takes is likely to vary according to glossal morphology. Inspecies with a non-pointed glossa the saliva would be dispersed over the superiorsurface of the glossa and of both paraglossae, but in the case of a pointed glossa(i.e. Cuneala, Eurys) it would be directed onto the inferior surface by the paraglossae(i.e. into the ventral groove, thereby keeping the saliva separate from the nectar ornectar/saliva mixture that has been loaded onto the superior surface; see Snodgrass’s[1935] discussion of Type 4 CNEA in bees).
The nectar or nectar/saliva mixture would be drawn onto and into the hairysurfaces of the glossa and paraglossae (‘nectar loading’ sensu Kingsolver & Daniel[1995]) by a combination of attraction (owing to the hydrophily of the hairs) andcapillarity (owing to the close juxtaposition of the hairs). Nectar is also likely to bedrawn up between the narrow spaces between the glossa and paraglossa, bycapillarity. Ingestion of the nectar/saliva mixture that is eventually ‘unloaded’ fromthe glossa/paraglossae when the ‘tongue’ is retracted (see Jervis [1998] for discussionof the mechanics of unloading) would be achieved through pumping movements ofthe cibarial and pharyngeal musculature.
Based on the disposition of the labiomaxillary complex in some of the driedspecimens examined, and in Benson’s (1934) figure of E. laetus (his and our fig. 4B),it appears that Eurys species with a greatly elongated prementum can retract theCNEA to such an extent that, except for the glossa, it is concealed in anterior viewby the galeae. Whether this occurs only during nectar unloading, occurs only whenthe CNEA is inactive, or occurs in both circumstances, is not known.
Type 5. This CNEA occurs in two unidentified species of Pergidae of the subfamilyEuryinae (Houston, 1983). The adults studied by Houston (1983) were collectedfrom the flowers of Eremophila sp. (Myoporaceae), which has its nectar well concealedat the base of the corolla (see discussion by Houston [1983]). Nectar would bedrawn up between the palps by capillarity, and then be ingested by means of alapping motion of the glossa coupled with cibarial and pharyngeal pumping (Houston,1983; Jervis, 1998).

M. JERVIS AND L. VILHELMSEN134
Type 5 CNEA is shared with Braconidae and Apidae (Houston, 1983; Jervis,1998). Whether it is a flexible structure or is always held rigid (see Houston’s Fig.9), is not known. The pergids Acordulecera longicornis Enderlein and Aulacomerus retusicornis(Konow) resemble some cryptine ichneumonids and some chelonine braconids inhaving elongated maxillary palps but, as in those wasps, the palps in museumspecimens are not closely aligned (which would be necessary for nectar to be drawnup by capillarity), so they cannot be firmly placed in this category of CNEA untilobservations are made of feeding insects.
Adults of some Xyelidae: Xyelinae (Xyela and Pleroneura Ashmead) and Ma-croxyelinae (Xyelecia Ross, but not Macroxyela Kirby and Megaxyela Ashmead), alsohave modified maxillary palps; in this case the second and third segments are greatlyelongated and expanded but remain cylindrical (Fig. 1C). This modification is mostprobably associated not with nectar feeding but with pollen feeding. The palps havea few elongate hairs situated close to their apices but lack the dense covering ofcurved hairs used for pollen collection as seen in the aculeate families Mutillidaeand Tiphiidae ( Jervis, 1998). Nevertheless, they could be used in the gathering ofpollen grains as in the moth Micropteryx calthella (see Hannemann, 1956). [The absenceof elongate maxillary palps in Macroxyela and Megaxyela is in accordance with theobservations of Macroxyela ferruginea feeding on buds (Garlick [1923], see ‘Foods’above) rather than pollen. However, these two genera retain the other peculiarmouthpart features (highly asymmetrical mandibles (Fig. 1A, B), epipharyngealbrush, pronounced infrabuccal pouch (Vilhelmsen, 1996) associated with pollen-feeding in other xyelids. We consider it most likely that pollen-feeding is ancestralto the family, the feeding habit of Macroxyela being secondarily derived.]
Type 8. This occurs in the two known species (not examined by us) in the tenthredinidgenus Nipponorhynchus Takeuchi (Takeuchi & Tokunaga, 1941; Naito, 1973, 1975)(Fig. 5). Takeuchi & Tokanuga were correct in remarking that this CNEA is unusualamong Hymenoptera generally. Uniquely, the prementum and stipites are greatlyelongated, the latter closely aligned laterally with the former. The lateral surfacesof the prementum are concave, as is the internal (i.e. inwards facing) surface of eachstipes. Together, the prementum and stipites form a structure with two foodconduction channels. The galeae, the maxillary palps, the glossa/paraglossae andthe labial palps are not obviously specialized. However, in contrast to otherHymenoptera, the hypopharynx extends over the glossa. Its tip, like that of theglossa, is densely covered in hairs, suggesting involvement in nectar loading (seebelow).
Saliva would be released from the tip of the CNEA and the resulting nectar/saliva mixture would be loaded onto the glossa and paraglossae, probably initiatedby contact with the tip of the hypopharynx. Once a quantity of fluid has beenloaded onto the glossa/paraglossae (which present a relatively small surface area inthis CNEA), the meal would then be drawn up the two nectar conduction channelsby capillarity. According to Takeuchi & Tokunaga (1941) the prementum andstipites together form a rigid structure; whether this rigidity is confined to feedingor also applies to the CNEA in repose, needs to be determined.
Nipponorhynchus, which is confined to Japan, forms a clade in the Selandriinae withthe CNEA-less genus Adelesta Ross, which is confined to eastern North America(Naito, 1975). Like Adelesta, Nipponorhynchus appears to be associated as larvae withferns that form part of a relict Tertiary flora in each continent (Naito, 1975); it is
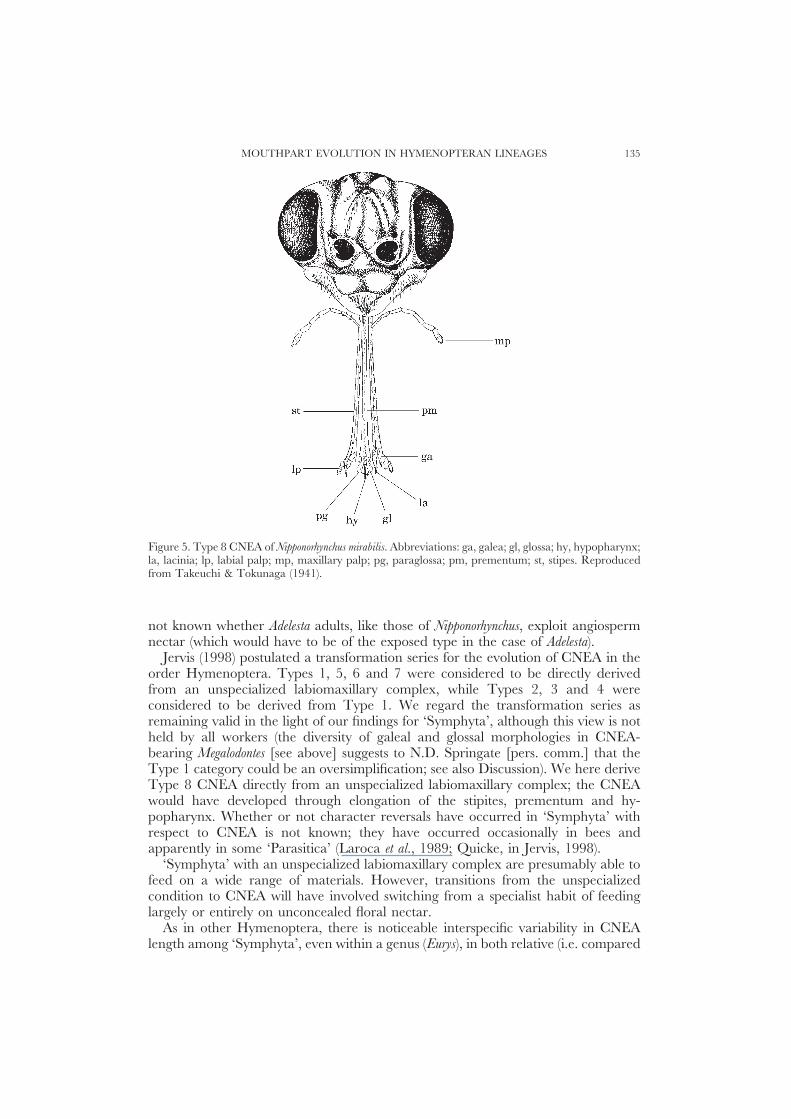
MOUTHPART EVOLUTION IN HYMENOPTERAN LINEAGES 135
Figure 5. Type 8 CNEA of Nipponorhynchus mirabilis. Abbreviations: ga, galea; gl, glossa; hy, hypopharynx;la, lacinia; lp, labial palp; mp, maxillary palp; pg, paraglossa; pm, prementum; st, stipes. Reproducedfrom Takeuchi & Tokunaga (1941).
not known whether Adelesta adults, like those of Nipponorhynchus, exploit angiospermnectar (which would have to be of the exposed type in the case of Adelesta).
Jervis (1998) postulated a transformation series for the evolution of CNEA in theorder Hymenoptera. Types 1, 5, 6 and 7 were considered to be directly derivedfrom an unspecialized labiomaxillary complex, while Types 2, 3 and 4 wereconsidered to be derived from Type 1. We regard the transformation series asremaining valid in the light of our findings for ‘Symphyta’, although this view is notheld by all workers (the diversity of galeal and glossal morphologies in CNEA-bearing Megalodontes [see above] suggests to N.D. Springate [pers. comm.] that theType 1 category could be an oversimplification; see also Discussion). We here deriveType 8 CNEA directly from an unspecialized labiomaxillary complex; the CNEAwould have developed through elongation of the stipites, prementum and hy-popharynx. Whether or not character reversals have occurred in ‘Symphyta’ withrespect to CNEA is not known; they have occurred occasionally in bees andapparently in some ‘Parasitica’ (Laroca et al., 1989; Quicke, in Jervis, 1998).
‘Symphyta’ with an unspecialized labiomaxillary complex are presumably able tofeed on a wide range of materials. However, transitions from the unspecializedcondition to CNEA will have involved switching from a specialist habit of feedinglargely or entirely on unconcealed floral nectar.
As in other Hymenoptera, there is noticeable interspecific variability in CNEAlength among ‘Symphyta’, even within a genus (Eurys), in both relative (i.e. compared

M. JERVIS AND L. VILHELMSEN136
to head length) and absolute terms. Species with the shortest apparatus (hardlyprotruding below mandibles in repose) include Tenthredo species (Tenthredinidae),Clarissa hebe, Lophyrotoma analis (Pergidae), Onycholyda sertata (Pamphiliidae), Didymiaalbipes, Trochophora lobata (Argidae) and Janus kashivorus (Cephidae), while those withthe longest include several Eurys species (Pergidae) (Fig. 4A–C) and Nipponorhynchusmirabilis (Tenthredinidae) (Fig. 5). In the specimen of E. chloe examined, the tip ofthe CNEA reaches the mid coxae. Megalodontes olivieri (Fig. 2) and Pterygophorus distinctus(Pergidae) (Fig. 3) and Cuneala species (Tenthredinidae) have moderately long CNEA.
The observed variability in CNEA length undoubtedly reflects in large part anadaptive response to variability in floral morphology, i.e. the exploitation of corollasof different lengths. However, studies on bumblebees have shown that CNEA lengthis not necessarily an accurate predictor of the flower species exploited, for tworeasons: (i) measured length and functional length are not necessarily synonymous(Harder, 1982); (ii) whereas CNEA length is the morphological feature most closelyassociated with flower choice, its importance in determining the range of flowerspecies exploited can vary significantly both with other insect morphological (bodysize) and with flowering plant ecological factors (abundance, species richness) (Harder,1985). Furthermore, it is clear both from our study of ‘Symphyta’ and from researchon ‘Parasitica’ ( Jervis, 1998) that exposed floral nectar sources can be included inthe resource range of CNEA-bearing species. Unfortunately, there is a major paucityof information available on the floral nectar sources exploited by CNEA-bearing‘Symphyta’ that would help elucidate to what extent CNEA length is a predictorof flower species range: as with ‘Parasitica’ ( Jervis et al., 1993), the majority ofpublished and data label records of flower visiting by ‘Symphyta’ neither indicatethe food resource (pollen, nectar) consumed nor rule out the possibility that thewasps were visiting for purposes other than feeding on plant materials (i.e. predation,mating). Liston (1980) observed Athalia species (with Type 1 CNEA) visiting theflowers of Labiatae, and Cuneala species (Type 1) those of Geranium; both of theseplant species have ‘concealed’ nectar . Riek (1970) remarks that in Australia, Euryinae(whether with Type 1 or 5 CNEA is not known) are common in early summer onblossoms, especially the small, tubular, blue flowers of Wahlenbergia (Campanulaceae)but are also attracted to the flowers of eucalypts and other plants. N.D. Springate(pers. comm.) observed Megalodontes cephalotes (Type 1) consuming both the (concealed?) nectar of Tripleurospermum inodorum (L.) Schultz (Asteraceae) and the unconcealednectar of Ranunculus repens.
As far as is known, no ‘Symphyta’ exhibit the downwards elongation of the headthat would facilitate exploitation of more deeply concealed nectar than absoluteCNEA length would indicate (cf. some Apocrita, some Agathidinae, some Leuco-spidae, some bees [Jervis, 1998]).
The structure and distribution of glossal and paraglossal hairs in CNEA-bearing‘Symphyta’ was not examined extensively. The hairs are scale-like and are arrangedin transverse rows in the two CNEA-bearing species studied: Pterygophorus distinctusand Eurys bella. The hairs are either scale-like or capitate in CNEA-less ‘Symphyta’(Michener & Brooks, 1984; Vilhelmsen, 1996) (see CNEA-bearing ‘Parasitica’ inJervis [1998]). Flattening of part or all of each hair presents a larger surface areafor nectar loading.
CNEA was not observed in either: Anaxyelidae (Syntexis Rohwer examined);Xyelidae (Xyela Dalman, Xyelecia Ross, Macroxyela Kirby and Pleroneura Ashmeadexamined); Diprionidae (Gilpinia Benson, Monoctenus Dahlbom, Neodiprion Rohwer

MOUTHPART EVOLUTION IN HYMENOPTERAN LINEAGES 137
examined; Blasticotomidae (Blasticotoma filiceti Klug, B. atra Zhelokhovtsev, B. nipponicaTakeuchi, Paremphytus flavipes [Takeuchi] and Runaria reducta [Malaise] examined);Siricidae (Sirex L., Urocerus Geoffroy, Xeris Costa, Xoanon Semenov-Tian-Shanskij,Eriotremex Benson, Tremex Jurine examined); Xiphydriidae (Xiphydria Latreille ex-amined); Orussidae (Orussus Latreille examined); the cephid genus Athetocephus Benson;or the following genera of pamphiliids: (Cephalciinae) Acantholyda A. Costa, CaenolydaKonow, Cephalcia Panzer and (Pamphiliinae) Neurotoma Konow; of argids: (Arginae)Scobina Lepeletier; of pergids: (Perginae) Acanthoperga Shipp, Cerealces Kirby, PergaLeach, Pseudoperga Guerin, Xyloperga Shipp, (Euryinae) Ancyloneura Cameron, (Sy-zygoniinae) Incalia Cameron, Lagideus Konow and Syzygonia Klug, (Parasyzygoniiae)Parasyzygonia Rohwer, (Acordulecerinae) Giladeus Brethes, (Loboceratinae) AulacomerusSpinola, (Philomastiginae) Philomastix Froggat; of cimbicids: (Cimbicinae) CimbexOlivier, (Pachylostictinae) Pachylosticta Klug and Pseudopachylosticta Mallach, and ofthe following species of tenthredinids: (Selandriinae) Dolerus aeneus, D. niger (L.),D. unicolor, (Tenthredininae) Macrophya punctumalbum, Zaschizonyx montana (Cresson),(Nematinae) Nematus oligospilus, (Blennocampinae) Athalia circularis (Klug), A. cordataLepeletier, A. liberta (Klug) and A. rosae (L.). Those families known only from fossilmaterial (see Abe & Smith [1991]) were not examined.
Even among those families lacking CNEA, the mouthparts exhibit considerablevariation. The configuration of the mouthparts in Xyelidae has already beenmentioned. In the woodwasps (Anaxyelidae, Siricidae, Xiphydriidae) and in Orus-sidae, several transformation series in mouthpart structure were revealed byVilhelmsen (1996): the labrum is displaced posteriorly to the tips of the mandibles(as in Pamphiloidea and Cephidae) and reduced in size, and the glossa andparaglossae are shortened, the glossa having a characteristic inverted U-shape inAnaxyelidae, Orussidae and Apocrita. All of these changes are most pronounced inthe latter two taxa, supporting the sister-group relationship between them(Vilhelmsen, 1997). These changes are probably not related to feeding, but ratherserve to free the mandibles for chewing an escape tunnel following eclosion of theadult wasp; the woodwasps, the Orussidae, and the basalmost apocritan familiespupate in wood, which must consitute a formidable barrier. Indeed, the mandiblesin most Hymenoptera are reinforced by deposits of zinc and/or manganese, whichmakes them more resistant to abrasion, a trait inferred to have evolved in thecommon ancestor of Cephidae (whose members also pupate within plant tissue),‘Siricoidea’, Orussidae and Apocrita (Quicke et al., 1998).
The functional mouth in Siricidae is typically very small; the mouthparts otherthan the mandibles are very reduced (the maxillary components are reduced, andthe labium is transformed into a tubular ‘totaglossa’) (Vilhelmsen, 1996) (seeDiscussion)
DISCUSSION
The conventional wisdom in the literature concerned with flower visiting (Kevan& Baker, 1983; Proctor et al., 1996) is that ‘Symphyta’ cannot exploit ‘concealed’floral nectar. As can be seen from this study, not only can they exploit such nectar,but also the possession of CNEA for the purpose appears to be fairly widespreadwithin the ‘Symphyta’, being present in at least half of the 14 extant families. CNEA

M. JERVIS AND L. VILHELMSEN138
Xyelidae
Blasticotomidae
Tenthredo (1)
Nematus
Diprionidae
Cimbicidae (1)
Argidae (1)
Pergidae (1, 5)
Pamphiliidae (1)
Megalodontesidae 1
Cephidae (1)
Anaxyelidae
Dolerus
Siricidae
Orussidae
'Tenthredinidae' (1, 8)
APOCRITA (1–7)
Xiphydriidae
Athalia (1)
Figure 6. Cladogram of the extant basal hymenopteran lineages, with the occurrence of CNEA mappedonto it. CNEA is indicated, out of parentheses when possessed by all members of the taxon, and inparentheses when present in some members only. Information for CNEA of the Apocrita was takenfrom Jervis (1998). The cladogram was derived by analysis in PAUP 3.1.1 (Swofford, 1993; somecharacters treated as ordered, TBR-branch swapping, random additions sequence, 20 replications,successive weighting using the consistency index applied until the length and topology of the resultingtree no longer changed) of a comprehensive data set, scored for six outgroup and 38 hymenopterantaxa (Vilhelmsen, 1999) Mouthpart characters were excluded from the analysis.
of some kind or other has independently evolved several times within the ‘Symphyta’,both among and within families (Fig. 6), and in some cases within genera (e.g.Athalia). Also, two types of CNEA have arisen independently within the pergidsubfamily Euryinae (Types 1 and 5) and within the tenthredinid subfamily Se-landriinae (Types 1 and 8).
The occurrence of different types of CNEA in the ‘symphytan’ families issummarized in Table 1 and Figure 6. CNEA was arguably present in the commonancestor of one family only, the Megalodontesidae. It is also possible that CNEA isa ground plan feature of the Cephidae, being absent only from Athetocephus amongthe taxa examined here, from which it might have been lost secondarily. This cannotbe confirmed at present, however, since there is no phylogeny available for thisfamily. Both the Megalodontesidae and Cephidae are small taxa, comprising aroundthirty and around a hundred species, respectively (Megalodontesidae: Springate,1995; Cephidae: Goulet, 1993). These two cases apart, the presence of CNEA in‘Symphyta’ is characteristic of taxa of genus rank (e.g. Eurys, Nipponorhynchus, Cuneala)or lower. Jervis (1998) revealed a similar pattern in the Apocrita: in that group

MOUTHPART EVOLUTION IN HYMENOPTERAN LINEAGES 139
CNEA is a ground plan feature of only one subfamily, the Parnopinae (Chrysididae),and two families, the Leucospidae and (a long-standing observation) the Apidae. Thepattern of independent acquisition of CNEA observed within the order Hymenopteracontrasts significantly with the incidence of ‘suctorial’ mouthparts observed in mostof the other large insect orders, particularly those comprising the Mecopterida (=the orders Mecoptera, Siphonaptera, Diptera, Trichoptera and Lepidoptera), wheresuch mouthparts are diagnostic of entire orders or large parts of them (Kristensen1991; see below). Our study has also revealed a degree of parallel evolution betweenthe basal hymenopteran lineages and the Apocrita, the mouthpart transformations:unspecialized labiomaxillary complex to Type 1, and unspecialized labiomaxillarycomplex to Type 5 being common to the two groups; such parallelism has hithertonot been remarked upon (although Houston [1983] remarks on the sharing of thesame type of haustellate mouthparts [Type 5] between some Pergidae and someApoidea). Jervis (1998) also observed parallelism in evolution of CNEA between theIchneumonoidea and its putative sister-group, the Aculeata.
CNEA has not been included as a character in recent cladistic treatments ofthe Hymenoptera (Vilhelmsen, 1997; Ronquist et al., 1999), although mouthpartcharacters were included. Indeed, it would be inappropriate to include CNEA,because of the non-homology of the constituent parts (this study and that of Jervis[1998]). Even among taxa assigned the same type of CNEA, there might bedifferences. For example, both Megalodontesidae and most Cephidae have Type 1,yet in the former family the labrum is elongate and is incorporated into the probosciswhile in the latter it is short and excluded (Vilhelmsen, 1996) (in the former, thelabrum forms a roof to the CNEA, but is unlikely to be directly involved intransporting nectar to the mouth). This further corroborates the view that the CNEAare not homologous in these families.
The most recent review of the palaeontological literature on insect mouthpartsmaintains the view that CNEA among Hymenoptera is restricted to known fossilsof Aculeata (Labandeira, 1997), but this may simply reflect the fact that entomologiststend not to be aware that CNEA occurs in non-aculeate Hymenoptera. It wouldbe useful to determine whether or not CNEA is present in fossil ‘symphytans’, as‘Symphyta’ may have pre-dated, or at least coincided with, the Aculeata in evolvingmorphological specializations for dealing with the nectar-concealing floral structuresof the early composite angiosperms (so far, the earliest recorded fossil Hymenopteraknown to bear CNEA have been bees, recorded in late Cretaceous [Turonian toCampanian] amber [Michener & Grimaldi, 1988]; many fossil ‘Symphyta’ pre-datethis period, the oldest known hymenopteran fossil from the Triassic being assignedto the Xyelidae (Rasnitsyn 1969, 1988).
We likewise cannot assume CNEA to be absent in those extant families in whichwe have not recorded such apparatus (i.e. Xyelidae, Diprionidae, Blasticotomidae,Xiphydriidae, Orussidae; the Siricidae being an exception, see below). CNEA canbe expected to occur in at least some of these (and some of the extinct) groups fortwo reasons. Firstly, a sitophore (see Mouthparts, above) is present in all ‘Symphyta’(as in apparently all other Hymenoptera), so all have the anatomical potential todevelop CNEA. Vilhelmsen (1996) suggested that the sitophore is a probablesynapomorphy for the Hymenoptera + Mecopterida, and inferred that this isindicative of it being a ‘preadaptation’ for the development of ‘suctorial’ mouthparts(i.e. a proboscis) (we use the term ‘suctorial’ broadly, rather than in the strictsense used by Kingsolver & Daniel [1995]) (as noted above, in ‘Foods’; in those

M. JERVIS AND L. VILHELMSEN140
Hymenoptera having an unspecialized labiomaxillary complex the glossa can beeverted to some extent and thus be used as a probing device).
Suctorial mouthparts probably evolved independently in the common ancestorsof the Trichoptera, Siphonaptera and Diptera (Kristensen, 1991), and they haveapparently evolved only once within the Lepidoptera, being diagnostic of theglossatan moths which comprise the majority of the order (Kristensen, 1984).Furthermore, a sitophore is present in the largest hemimetabolan order, the Hem-iptera, which is characterized primarily by the possession of ‘suctorial’ mouthparts(Chaudonneret, 1990). This further strengthens the view that a sitophore is anecessary prequisite for the evolution of CNEA.
The distribution of both a sitophore and ‘suctorial’ mouthparts in the Coleopteracould provide an additional test of our hypothesis. These structures are absent fromPriacma serrata Leconte (a representative of the basalmost extant lineage of theColeoptera [Beutel, 1997]), and presumably from the ground plan of the order, sowe can reasonably infer that lack of a sitophore has constrained the occurrence ofCNEA or other ‘suctorial’ apparatus in adult beetles. It is indeed the case thatCNEA is very rare among Coleoptera despite many members being consumers ofnectar and other floral resources (e.g. see Evans [1973] and Grinfel’d [1975] forexamples of taxa). The only CNEA-bearing beetles we are aware of are among theMeloidae (e.g. the genus Nemognatha Illiger: adults have elongated and hairy [ontheir internal surfaces] galeae; Leptopalpus Guerin der Meneville: adults have elongatedand hairy [on their internal surfaces] maxillary palps [Grinfel’d, 1975]). A narrowsclerite traversing the floor of the cibarium has been observed in many beetles, butit is probably not homologous with the sitophore of Hymenoptera and Mecopterida(Kristensen, 1999). It would be useful to examine whether a sitophore is present inCNEA-bearing beetles such as Nemognatha.
[Another noteworthy pattern is in the fate of the mandibles in the adults ofCNEA-bearing Hymenoptera and proboscis-bearing Mecopterida. In ‘Symphyta’,as in other Hymenoptera ( Jervis, 1998), CNEA has not developed at the expenseof the mandibles. Fully developed mandibles are required for adult eclosion,emergence and consumption of solid foods (see ‘Foods’, above). In proboscis-bearingMecopterida, however, the mandibles are more or less reduced (Chaudonneret,1990).]
The second reason for expecting CNEA to be present in several of the ‘symphytan’families in which we have so far not recorded such apparatus is that some of thesegroups contain large-bodied insects. Thus, not only will adults have a high absolutemetabolic requirement (e.g. see Gilbert [1985]) and concomitant strong need toexploit energy-rich food sources (discussed below) but also they will be unable togain access to nectar in long tubular corollas or other ‘concealing’ structures, otherthan by means of CNEA.
We therefore consider there to be strong grounds for assuming that, providedthere exist both the necessary ecological opportunities (albeit infrequent generally,as ‘concealed’ nectar sources are rare in most habitats [Gilbert & Jervis, 1998]) andthe necessary selection pressures (discussed below), CNEA can evolve unconstrainedamong ‘Symphyta’.
Siricids are, however, unlikely to possess CNEA. Their labiomaxillary complexis typically reduced (see ‘Mouthparts’), and adults characteristically do not feed asadults (egg development and maintenance are fuelled by stored fats [Zondag &

MOUTHPART EVOLUTION IN HYMENOPTERAN LINEAGES 141
Nuttall, 1977]). They also lack rectal papillae, which would not be required if theadults are non-feeding.
Floral nectar can be used to fuel several physiological processes in ‘Symphyta’:egg production, somatic maintenance and locomotion (in winged species mainlyflight, but in wingless species only walking, e.g. see Naumann [1984, 1997] andShinohara [1986]). It appears that most ‘Symphyta’ are synovigenic (e.g. see Okutani[1957], Tait [1962], Heron [1966], Maeta et al. [1981], Hatakeyama et al. [1990]),i.e. females eclose with only part of their lifetime egg complement mature and readyfor laying, and so need to feed in order to mature the remaining oocytes. Comparedwith other potential foods, floral nectar is generally low both in proteinaceousmaterials and in lipids, but is high in carbohydrates. The large lipid demand foregg yolk formation (symphytan eggs are yolk-rich) would be satisfied by the conversion,via the tricarboxylic cycle, of nectar sugars (early on in life, however, the fat bodyreserves carried over from the larval stage could supply much or all of the demand).Through elongation of different labiomaxillary components (galeae, prementum,glossa and paraglossae in Type 1 CNEA, maxillary palps only in Type 5, prementumand stipites in Type 8), and also elongation of the hypopharynx in Type 8, a non-trivial proportion of ‘Symphyta’ have switched from feeding on unconcealed floralnectar sources to feeding on concealed ones. This begs the question: what are themain fitness advantages to possessing CNEA, as compared with an unspecializedlabiomaxillary complex? Gilbert & Jervis (1998) identified three:
(i) Compared with exposed nectar, nectar concealed either in long narrow, tubularcorollas or in other deep tubular or cup-shaped structures that act as a nectarreservoir, remains relatively dilute. It is therefore of low viscosity and does notrequire dilution with saliva prior to ingestion, and so extraction can be rapid (i.e.absolute handling time is low), provided the CNEA is not too long (beyond a certainlength relative to body size, CNEA can be cumbersome, see Plowright & Plowright[1997]).
(ii) Compared with exposed nectar sources, sources of concealed nectar oftenoffer a much greater volume of nectar and so are usually more sugar-rich in absoluteterms. Thus, potentially a greater amount of nectar sugar can be extracted, and agreater net energy return from feeding obtained (this assumes that anatomical/physiological specializations can be developed, in conjunction with CNEA: broadly,those for the ingestion of large volumes of nectar and those for the excretion ofexcess water [but see iii, below]).
(iii) Sources of concealed nectar are often the only readily available source ofwater required for maintaining an appropriate hygrothermal physiological balance.As with parasitoid wasps ( Jervis, 1998), many ‘Symphyta’ possessing CNEA occurin, and may be restricted to, arid or semi-arid habitats (e.g. most Megalodontes species,many of the CNEA-bearing Pergidae).
One disadvantage of possessing CNEA is that it can preclude consumption ofadditional, in some cases richer, sources of egg production materials. CNEA Type5 perhaps cannot be used, for mechanical reasons (the glossa is too short, seeHouston [1983]) to consume unconcealed liquid foods, occurring as relatively thin‘films’, such as exposed floral nectar, liquid honeydew, plant sap, fruit juice andfungal spermatial fluid. However the same probably does not apply to CNEA Types1 and 8 (cf. Gilbert & Jervis, 1998). For example, Megalodontes cephalotes (Type 1CNEA) consumes the unconcealed nectar (as defined here) of Ranunculus repens (N.D.Springate, pers. comm.). Also, Roy (1993, 1994, 1996) recorded feeding by andrenid

M. JERVIS AND L. VILHELMSEN142
and halictid bees (Type 1 CNEA) on fungal spermatial fluid (note that these beeshave a pointed glossa (Michener & Brooks, 1984) as in the CNEA-bearing ‘sym-phytans’ Eurys and Cuneala). Having CNEA probably does tend to preclude ex-ploitation of contaminant pollen (i.e. in nectar), as the latter would block the nectartransport system (Gilbert & Jervis, 1998; Megalodontes appears to be an exception,see ‘Foods’), but probably does not preclude the direct consumption of prey andplant tissues and pollen from anthers, as mainly the mandibles are used (see Tenthredoarcuata, and also vespid wasps).
This paper, we hope, will serve to draw attention not only to the diversity ofmouthpart structure but also the diversity of feeding strategies among adult ‘Sym-phyta’. To date, studies on symphytan feeding ecology have been confined to thelarval stages (e.g. see Heitland & Pschorn-Walcher [1993]). As in other Hymenoptera,and also Diptera and Lepidoptera, the type of adult feeding strategy, combined withits reproductive and other physiological correlates, can have major populationdynamic consequences (see Jervis & Kidd, 1999; Gilbert & Jervis, 1998; Owen &Gilbert, 1989; Miller, 1996). Given the group’s economic and in some instancesecological importance, it is thus imperative that more be discovered about adultfeeding ecology.
ACKNOWLEDGEMENTS
We thank the following: Mike Fitton, Suzanne Lewis and Christine Taylor forenabling MJ to examine the ‘Symphyta’ collection of the Natural History Museum,London; the trustees of the aforementioned institution for permitting MJ to examinethe collection; Suzanne Lewis for also assisting MJ with SEM work; John Deeming,Dianne Edwards, Ian Gauld, Conrad Labandeira, Brian Levey, Peter Price, AlexRasnitsyn, David Sheppard and Neil Springate for valuable advice and discussion;Neil Springate for reading and providing valuable comments on a draft; twoanonymous reviewers for valuable comments; George Else for providing insightsinto bee mouthparts; Heicke Rothfritz for translating German; Jane Sparks forhelping with inter-library loans; Howard Ensign Evans whose writings inspired MJto research into Hymenoptera; Julia, William and George Jervis for moral supportto MJ.
REFERENCES
Abe M, Smith DR. 1991. The genus-group names of Symphyta (Hymenoptera) and their typespecies. Esakia 31: 1–115.
Anonymous. 1896. Minutes of meeting on March 5, 1896. Proceedings of the Entomological Society ofWashington 4: 31.
Belokobylskij S, Jervis MA. 1998. Two new species of Agathis (Hymenoptera, Braconidae, Aga-thidinae) from Spain, with a record of mating by one species on flowers. Journal of Natural History32: 1217–1225.
Benson RB. 1934. A classification of the sawflies of the family Pterygophoridae, with a revision ofthe Australian members of the subfamily Euryinae (Hymenoptera, Symphyta). Transactions of theRoyal Entomological Society of London 82: 461–478.
Benson RB. 1950. An introduction to the natural history of British sawflies. Transactions of the Societyfor British Entomology 10: 45–138.

MOUTHPART EVOLUTION IN HYMENOPTERAN LINEAGES 143
Benson RB. 1962. A revision of the Athaliini (Hymenoptera: Tenthredinidae). Bulletin of the BritishMuseum (Natural History) 11: 333–382.
Beutel R 1997. Uber Phylogenmese und Evolution der Coleoptera (Insecta). Abhandlungen des Natur-wissenschaftliches Vereins in Hamburg 31: 1–164.
Buller AHR. 1950. Researches on fungi Vol. VII: the sexual process in the Uredinales. Toronto: University ofToronto Press.
Caldas MB, Martins-Neto RG, Lima-Filho FP. 1989. Afropollis sp. (polem) no trato intestinal devespa (Hymenoptera: Apocrita: Xyelidae) no Cretaceo da Bacia do Araripe. Actas Simp. Geol. Nordeste13: 195–196.
Cameron P. 1882. A monograph of the British phytophagous Hymenoptera. (Tenthredo, Sirex and CynipsLinne) Volume I. London: The Ray Society.
Chapman RF. 1995. Mechanics of food handling by chewing insects. In: Chapman RF, de Boer G,eds. Regulatory mechanisms in insect feeding. London: Chapman & Hall, 3–31.
Chaudonneret J. 1990. Les pieces buccales des insectes. Edition Hors Serie du Bulletin Scientifique deBourgogne 1–256.
Chawner ET. 1921. The egg-laying of sawflies. Entomologist’s Monthly Magazine 57: 19.Chrystal N. 1928. The Sirex wood-wasps and their importance in Forestry. Bulletin of Entomological
Research 189: 219–247.Clausen CP. 1940. Entomophagous insects. New York: McGraw-Hill.Duncan DC. 1939. A contribution to the biology of North American vespine wasps. Stanford University
Publications, Biological Sciences 8: 1–272.Evans MEG. 1964. A comparative account of the feeding methods of the beetles Nebria brevicollis (F.)
(Carabidae) and Philonthus decorus (Grav.) (Staphylinidae). Transactions of the Royal Society of Edinburgh66: 91–109.
Evans MEG. 1973. The life of beetles. London: George Allen & Unwin.Evans MEG, Forsythe TG. 1985. Feeding mechanisms, and their variation in form, of some adult
ground-beetles (Coleoptera: Carabidae). Journal of Zoology, London 206: 113–143.Felton JC. 1956. Some sawflies (Hym., Symphyta) collected in Berkshire. Entomologist’s Monthly Magazine
92: 220.Garlick WG. 1923. Notes on the feeding habits of an adult sawfly. Canadian Entomologist 55: 256–257.Gauld ID, Bolton B. 1988. The Hymenoptera. London/Oxford: The Natural History Museum/Oxford
University Press.Gilbert FS. 1981. Foraging ecology of hoverflies: morphology of the mouthparts in relation to feeding
on nectar and pollen in some common urban species. Ecological Entomology 10: 385–392.Gilbert FS. 1985. Diurnal activity patterns in hoverflies (Diptera, Syrphidae). Ecological Entomology 10:
385–392.Gilbert FS, Jervis MA. 1998. Functional, evolutionary and ecological aspects of feeding-related
mouthpart specializations in parasitoid flies. Biological Journal of the Linnean Society 61: 495–535.Godfray HCJ. 1994. Parasitoids: Behavioural and Evolutionary Ecology. Princeton NJ: Princeton University
Press.Goulet H. 1986. The genera and species of the Nearctic Dolerini (Symphyta: Tenthredinidae:
Selandriinae): classification and phylogeny. Memoirs of the Entomological Society of Canada 135: 1–208.Goulet H. 1993. Superfamilies Cephoidea, Megalodontoidea, Orussoidea, Siricoidea, Tenth-
redinoidea, and Xyeloidea. In: Goulet H, Huber JT, eds. Hymenoptera of the World: an IdentificationGuide to Families. Ottawa: Agriculture Canada, 101–129.
Grinfel’d EK. 1975. Anthophily in beetles (Coleoptera) and a critical evaluation of the cantharophiloushypothesis. Entomological Review 4: 18–22.
Grunwaldt W. 1976. Andrena grossella, eine Insekten-Art mit 9-gliedrigen Maxillar- and Labialpalpen.Nachrichttenblatt der Bayerischen Entomologen 25: 65–70.
Hannemann HJ. 1956. Die Kopfmuskulatur von Micropteryx calthella (L.). Zoologische Jarbucher derAnatomie und Ontogenie der Tiere 75: 177–206.
Harder LD. 1982. Measurement and estimation of functional proboscis length in bumblebees(Hymenoptera: Apidae). Canadian Journal of Zoology 60: 1073–1079.
Harder LD. 1983. Functional differences of the proboscides of short- and long-tongued bees(Hymenoptera: Apoidea). Canadian Journal of Zoology 61: 1580–1586.
Harder LD. 1985. Morphology as a predictor of flower choice by bumblebees. Ecology 66: 198–210.Haslett J. 1983. A photographic account of pollen digestion by adult hoverflies. Physiological Entomology
8: 167–171.

M. JERVIS AND L. VILHELMSEN144
Hatkeyama M, Sawa M, Oishi K. 1990. Ovarian development and vitellogenesis in the sawfly,Athalia rosae ruficornis Jakovlev (Hymenoptera, Tenthredinidae). Invertebrate Reproduction and Development17: 237–245.
Heitland W, Pschorn-Walcher H. 1993. Feeding strategies of sawflies. In: Wagner MR, Raffa KF,eds, Sawfly life history adaptations to woody plants. San Diego: Academic Press, 93–118.
Heron RJ. 1966. The reproductive capacity of the larch sawfly and some factors of concern in itsmeasurement. Canadian Entomologist 98: 561–578.
Hobby BM. 1932a. The prey of sawflies (Hym., Tenthredinidae). Proceedings of the Royal EntomologicalSociety of London A 7: 14–15.
Hobby BM. 1932b. On the predacious habit in Tenthredo marginella F., and on possible mimeticassociations in sawflies. Proceedings of the Royal Entomological Society of London A 7: 15–116.
Hobby BM. 1932c. Further records of sawflies and their prey (Hym., Tenthredinidae). Proceedings ofthe Royal Entomological Society of London A 7: 35–36.
Houston TF. 1983. An extraordinary new bee and adaptation of palpi for nectar-feeding in someAustralian Colletidae and Pergidae. Journal of the Australian Entomological Society 22: 263–270.
Jervis MA. 1998. Functional and evolutionary aspects of mouthpart structure in parasitoid wasps.Biological Journal of the Linnean Society 63: 461–493.
Jervis MA, Kidd NAC. 1999. Parasitoid adult feeding behaviour and biological control. In: HawkinsBA, Cornell HV, eds. Theoretical approaches to biological control. Cambridge: Cambridge UniversityPress, 131–151.
Jervis MA, Kidd NAC, Fitton MG, Huddleston T, Dawah HA. 1993. Flower-visiting byhymenopteran parasitoids. Journal of Natural History 27: 67–105.
Kevan PG, Baker HG. 1983. Insects as flower visitors and pollinators. Annual Review of Entomology28: 407–453.
Kingsolver JG, Daniel TL. 1995. Mechanics of food handling by fluid-feeding insects. In: ChapmanRF, de Boer G, eds. Regulatory mechanisms in insect feeding. London: Chapman & Hall, 32–73.
Korolkov DM. 1914. Pests of orchards. Materials for the Study of the Injurious Insects of the Government ofMoscow 5: 1–93.
Krassilov VA, Rasnitsyn AP. 1983. A unique find: pollen in the intestine of Early Cretaceoussawflies. Palaeontological Journal 1982: 80–95.
Kristensen NP. 1984. Studies on the morphology and systematics of primitive Lepidoptera (Insecta).Steensrupia 10: 141–191.
Kristensen NP. 1991. Phylogeny of extant hexapods. In: The Insects of Australia, Second Edition. Carlton:Melbourne University Press, 125–140.
Kristensen NP. 1999. Phylogeny of the endopterygote insects, the most successful lineage of livingorganisms. European Journal of Entomology.
Labandeira CC. 1997. Insect mouthparts: Ascertaining the palaeobiology of insect feeding strategies.Annual Review of Ecology and Systematics 28: 153–193.
Laroca S, Michener CD, Hofmeister RM. 1989. Long mouthparts among ‘short-tongued’ beesand the fine structure of the labium in Niltonia (Hymenoptera: Colletidae). Journal of the KansasEntomological Society 62: 400–410.
Linsley EG, MacSwain JW. 1959. Ethology of some Ranunculus insects with emphasis on competitionfor pollen. University of California Publications in Entomology 16: 1–33.
Liston AD. 1980. Why sawflies of the Tenthredo arcuata-schaefferi complex (Hymenoptera, Ten-threclinidine) visit flowers. Annales Entomologici Fennici 46: 85–88.
Maeta Y, Kurihara M, Hara K. 1981. Biological and histological studies on the ovarian developmentof the European pine sawfly Neodiprion sertifer Geoffory (Hymenoptera, Diprionidae). Kontyu 49:577–596.
Matsumura S. 1927. Summary of the Japanese Xiphydria species. Insecta Matsumura, Sapporo 1: 202–206.Michener CD, Brooks RW. 1984. Comparative study of the glossae of bees (Apoidea). Contributions
of the American Entomological Institute 22: 1–73.Michener CD, Grimaldi DA. 1988. The oldest fossil bee: Apoid history, evolutionary stasis, and
antiquity of social behaviour. Proceedings of the National Academy of Sciences 85: 6424–6426.Miller WE. 1996. Population behaviour and adult feeding capacity in Lepidoptera. Environmental
Entomology 25: 213–226.Murray J. 1927. A carnivorous sawfly. Naturalist 1927: 139.Naito T. 1973. A new species of Nipponorhynchus Takeuchi from Japan (Hymenoptera,: Tenthredinidae).
Kontyu 41: 95–96.

MOUTHPART EVOLUTION IN HYMENOPTERAN LINEAGES 145
Naito T. 1975. Phylogeny and distribution of five related genera, Eriocampidea, Hemitaxonus, Pseudo-hemitaxonus, Nipponorhynchus and Adelesta, referred to the Selandriinae (Hymenoptera, Tenthredinidae).Kontyu 43: 330–342.
Naumann ID. 1984. An apterous sawfly (Hymenoptera: Symphyta) from Papua New Guinea.Systematic Entomology 9: 339–349.
Naumann ID. 1997. A remarkable, new Australian sawfly with brachypterous, nocturnal or crepuscularfemales (Hymenoptera: Symphyta: Pergidae). Journal of Natural History 31: 1335–1346.
Okutani T. 1957. Some remarks on the biology of the Japanese Symphyta (Studies on SymphytaIX). Insect Ecology 6: 139–150.
Owen J, Gilbert F. 1989. On the abundance of hoverflies. Oikos 55: 183–193.Patt JM, Hamilton GC, Lashomb JH. 1997. Foraging success of parasitoid wasps on flowers:
interplay of insect morphology, floral architecture and searching behaviour. Entomologia Experimentaliset Applicata 83: 21–30.
Percival MS. 1965. Floral biology. London: Pergamon.Plowright CMS, Plowright RC. 1997. The advantage of short tongues in bumblebees (Bombus) –
analyses of species distributions according to flower corolla depth, and of working speeds on whiteclover. Canadian Entomologist 129: 51–59.
Preston-Mafham R, Preston-Mafham K. 1993. The Encyclopedia of land invertebrate behaviour. London:Blandford Press.
Proctor M, Yeo PF, Lack A. 1996. The natural history of pollination. London: Collins New Naturalists.Quicke DLJ, Wyeth P, Fawke JD, Basibuyuk HH, Vincent JFV. 1998. Manganese and zinc in
the ovipositors and mandibles of hymenopterous insects. Zoological Journal of the Linnean Society 124:387–396.
Rasnitsyn AP. 1969. Origin and evolution of lower Hymenoptera. Transactions of the Institute ofPaleontology 123: 1–196 (In Russian; translated in 1979 by Amerind Publishing Co., New Delhi).
Rasnitsyn AP. 1988. An outline of the evolution of the hymenopterous Insects (Order Vespida).Oriental Insects 22: 115–145.
Rathay E. 1883. Untersuchungen uber die Spermogonien der Rostpilze. [Studies on the spermogoniaof the rust fungi]. Denkschriften der Kaiserlichen Akademie der Wissenschaften zu Wien 46: 1–51.
Riek EF. 1970. Hymenoptera. In: The insects of Australia. Carlton: Melbourne University Press.Rohwer SA. 1913. Notes on the feeding habits of two adult sawflies. Proceedings of the Entomological
Society of Washington 15: 148–149.Ronquist F, Rasnitsyn A, Roy A, Erikson K, Lindgren M. 1999. Phylogeny of the Hymenoptera:
A cladistic reanalysis of Rasnitsyn’s (1988) data. Zoological Scripta 28: 13–50.Rosenheim JA, Kaya HK, Ehler LE, Marois JJ, Jaffee BA. 1995. Intraguild predation among
biological control agents – theory and evidence. Biological Control 5: 303–335.Ross HH. 1933. The description and life history of a new sawfly Sterictiphora apios (Argidae: Hymen.).
Proceedings of the Entomological Society of Washington 35: 13–19.Roy B. 1993. Floral mimicry by a plant pathogen. Nature 362: 56–58.Roy B. 1994. The effects of pathogen-induced pseudoflowers and buttercups on each other’s insect
visitation. Ecology 75: 352–358.Roy B. 1996. A plant pathogen influences pollinator behaviour and may influence reproduction of
non-hosts. Ecology 77: 2445–2457.Saini MS, Dhillon SS. 1979a. Comparative morphology of galea and lacinia in Hymenoptera.
Entomon 4: 149–155.Saini MS, Dhillon SS. 1979b. Glossal and paraglossal transformation in order Hymenoptera. Entomon
4: 355–360.Shinohara A. 1986. A new apterous sawfly from Sulawesi, Indonesia (Hymenoptera: Pergidae:
Pereyiinae), and the pleural origin of the ventral region of the sawfly mesothorax. Systematic Entomology11: 247–253.
Smith DR. 1993. Systematics, life history, and distribution of sawflies. In: Wagner MR, Raffa KF,eds, Sawfly life history adaptations to woody plants. San Diego: Academic Press, 3–32.
Snodgrass RE. 1935. Principles of insect morphology. New York & London: McGraw-Hill.Springate ND. 1995. Megalodontidae (Insecta: Hymenoptera): a systematic revision and cladistic
analysis. Ph.D. thesis, University de Neuchatel.Stein JD. 1974. Elm sawfly. United States Department of Agriculture Forest Service, Forest Pest Leaflet 142: 1–6.Swofford DL. 1993. PAUP. Phylogenetic Analaysis Using Parsimony, Version 3.1.1. Champaign: Illinois State
Natural History Survey.

M. JERVIS AND L. VILHELMSEN146
Tait NN. 1962. The anatomy of the sawfly Perga affinis Kirby (Hymenoptera: Pergidae). AustralianJournal of Zoology 10: 652–684.
Takeuchi K, Tokunaga M. 1941. Morphological study of the head and mouthparts of a remarkablesaw-fly, Nipponorhynchus mirabilis Takeuchi. Tenthredo 3: 275–291.
Todd FE, Vansell GH. 1942. Pollen grains in nectar and honey. Journal of Economic Entomology 35:728–731.
Togashi I. 1959. Food habits of some sawflies. Kontyu 27: 256.Togashi I. 1978. Insects of Mt. Hakusan. Kanazawa: Hoppo Shuppan-sha.Vilhelmsen L. 1996. The preoral cavity of lower Hymenoptera (Insecta): comparative morphology
and phylogenetic significance. Zoologica Scripta 25: 143–170.Vilhelmsen L. 1997. The phylogeny of the lower Hymenoptera (Insecta), with a summary of the
early evolutionary history of the order. Journal of Zoological and Systematic Evolutionary Research 35:49–70.
Vilhelmsen L. 1999. The comparative anatomy of the basal hymenopteran lineages, and itsphylogenetic implications. Unpublished Ph.D. thesis, Zoological Museum, University of Copen-hagen.
Willis JC, Burkill IH. 1895. Flowers and insects in Great Britain I. Annals of Botany 9: 227–273.Willis JC, Burkill IH. 1903. Flowers and insects in Great Britain II. Annals of Botany 17: 313–349.Willis JC, Burkill IH. 1908. Flowers and insects in Great Britain IV. Annals of Botany 22: 603–649.Willmer PG. 1980. The effect of insect visitors on nectar constituents in temperate plants. Oecologia
47: 270.Wolf F. 1968. Sur le regime alimentaire des tenthredininae adultes (Hymenoptera Symphyta). Bulletin
Annuel de la Societe Royale de Belgique 104: 275–276.Woollatt LH. 1946. Some biological notes on Sciapteryx consobrina Klug (Hym. Symphyta: Tenthre-
dinidae). Entomologist’s Monthly Magazine 82: 18–19.Zoebelein G. 1955. Der honigtau als nahrung der Insekten I [Honeydew as food of insects]. Zeitschrift
fur Angewandte Entomologie 38: 369–416.Zondag R, Nuttall MJ. 1977. Sirex noctilio Fabricius (Hymenoptera: Siricidae). Forest and Timber Insects
in New Zealand 20: 1–8.
Copyright © 2022 FDOKUMEN




















